![Exposure to benzo[a]pyrene of Hepatic Cytochrome P450 Reductase Null (HRN) and P450 Reductase Conditional Null (RCN) mice: Detection of benzo[a]pyrene diol epoxide-DNA adducts by immunohistochemistry](https://static.fdokumen.com/doc/165x107/63259f17c9c7f5721c022d3b/exposure-to-benzoapyrene-of-hepatic-cytochrome-p450-reductase-null-hrn-and-p450.jpg)
Biotransformacion del Benzo a pireno
88
INSTUTUTO POLITECNICO NACIONAL SECRETARIA DE INVESIGACIÓN Y POSGRADO ESCUELA NACIONAL DE CIENCIAS BIOLOGICAS SECCIÓN DE ESTUDIOS DE POSGRADO E INVESTIGACIÓN “BIOACTIVACIÓN DE COMPUESTOS ORGÁNICOS HALOGENADOS POR LA ACTIVIDAD DE LA GLUTATIÓN S-TRANSFERASA TETHA Y DEL CYP 2E1 EN EL CHARAL BLANCO Chirostoma riojai” TESIS QUE PARA OBTENER EL GRADO DE MAESTRO EN CIENCIAS QUIMICOBIOLÓGICAS PRESENTA: HIDROBIÓL. ROBERTO DE JESÚS GONZÁLEZ ORTÍZ DIRECTOR de tesis: DR. ARMANDO VEGA LÓPEZ Codirectora de tesis: DRA. ALICIA RAMÍREZ RAMÍREZ México D. F., Enero 2010
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Biotransformacion del Benzo a pireno

INSTUTUTO POLITECNICO NACIONAL SECRETARIA DE INVESIGACIÓN Y POSGRADO
ESCUELA NACIONAL DE CIENCIAS BIOLOGICAS
SECCIÓN DE ESTUDIOS DE POSGRADO E INVESTIGACIÓN
“BIOACTIVACIÓN DE COMPUESTOS ORGÁNICOS HALOGENADOS POR LA ACTIVIDAD DE LA GLUTATIÓN S-TRANSFERASA TETHA Y DEL CYP 2E1 EN EL
CHARAL BLANCO Chirostoma riojai”
TESIS QUE PARA OBTENER EL GRADO DE
MAESTRO EN CIENCIAS QUIMICOBIOLÓGICAS
PRESENTA:
HIDROBIÓL. ROBERTO DE JESÚS GONZÁLEZ ORTÍZ
DIRECTOR de tesis: DR. ARMANDO VEGA LÓPEZ Codirectora de tesis: DRA. ALICIA RAMÍREZ RAMÍREZ
México D. F., Enero 2010



Índice
Relación de figuras…………………………………………..……………………………………………………….i Relación de tablas…………………………………………………………………………………………………..iii Abreviaturas……………………………………………………………………………………………………..…..iv Símbolos……………………………………………………………………………………………………………...iv RESUMEN…………………………………………………………………………………………………………….v ABSTRACT…………………………………………………………………………………………………………vii
1. INTRODUCCIÓN………………………………………………………………………………………………...1 2. ANTECEDENTES…………………………………………………………………………………………………1
2.1 BIFENILOS POLICLORADOS……………………………………………………………………………...1 2.1.1 Toxocinética ambiental………………………………………………………………………………...3 2.1.2. Efectos tóxicos…………………………………………………………………………………………..4 2.1.3 Biotransformación………………………………………………………………………………………5 2.1.4 Normatividad…………………………………………………………………………………………....5
2.2 HIDROCARBUROS AROMÁTICOS POLICÍCLICOS (HAP´s)..............................................................6 2.2.1 Toxocinética……………………………………………………………………………………………...8 2.2.2 Efectos tóxicos…………………………………………………………………………………………....8
2.2.2.1 Naftaleno…………………………………………………………………………………………8 2.2.3 Biotransformación de los HAP’s………………………………………………………………………8
2.2.3.1 Naftaleno…………………………………………………………………………………………9 2.2.3.2 Benzo[a]pireno…………………………………………………………………………………..9
2.2.4 Normatividad…………………………………………………………………………………………..11 2.3 Biomarcadores…………………………………………………………………………………………………11
2.3.1 Uso de biomarcadores en estudios ecotoxicológicos………………………………………………11 2.3.1.1 Radicales libres y especies reactivas de oxígeno………………………………………….12 2.3.1.2 Estrés oxidativo………………………………………………………………………………...12 2.3.1.3 Lipoperoxidación celular……………………………………………………………………..13 2.3.1.4 Generación de hidroperóxidos lipídicos…………………………………………………...13 2.3.1.5 Oxidación de proteínas……………………………………………………………………….16 2.3.1.6 Generación de formaldehido………………………………………………………………...16
2.4 Biotransformación de los xenobióticos…………………………………………………………………….18 2.4.1 Sistema de oxidasas de función mixta (MFO)………………………………………………………18
2.4.1.1 Citocromo 2E1 (CYP 2E1)………………………………………………………………………19 2.4.2 Glutatión S-transferasas……………………………………………………………………………….19
2.5 ZONA DE ESTUDIO…………………………………………………………………………………………21 2.6 Generalidades sobre el charal de Santiago (Chirostoma riojai)………………………………………...23 2.7 Bioactivación y conjugación de PCB´s y HAP’s en organismos acuáticos……………………………..23
3. JUSTIFICACIÓN………………………………………………………………………………………………...25 4. HIPÓTESIS…………….…...…………………………………………………………………………………….25 5. OBJETIVOS……….………..…………………………………………………………………………………….26
5.1 OBJETIVO GENERAL………………………………………………………………………………………26 5.2 OBJETIVOS ESPECÍFICOS………………………………………………………………………………...26
6. METODOLOGÍA………………………………………………………………………………………………...27 6.1 Determinaciones realizadas en el laboratorio……………………………………………………………28
6.1.1 Proteínas totales………………………………………………………………………………………...28 6.1.2 Biomarcadores de bioactivación……………………………………………………………………..28
6.1.2.1 Actividad del CYP 2E1………………………………………………………………………..28 6.1.2.2 Actividad de la glutatión S-transferasa isoforma tetha…………………………………..28 6.1.3 Biomarcadores de estrés oxidativo…………………………………………………………….28
6.1.3.1 Anión superóxido………………………………………………………………………..28 6.1.3.2 Peróxido de hidrógeno…………………………………………………………………..28

6.1.3.3 Formaldehído……………………………………………………………………………..29 6.1.4 Biomarcadores de daño…………………………………………………………………………29
6.1.4.1 Substancias reactivas al ácido tiobarbitúrico (TBAR´s)……………………………29 6.1.4.2 Hidroperóxidos lipídicos………………………………………………………………29 6.1.4.3 Oxidación de proteínas (formación de carbonilos)………………………………....29
6.1.5 Determinaciones ambientales………………………………………………………………....29 6.1.5.1 Materia orgánica total en sedimentos y agua………………………………………..29
6.1.5.1.1 Materia orgánica total en sedimentos………………………………………29 6.1.5.1.2 Materia orgánica total en aguas………………………………………………30 6.1.5.1.3 Materia orgánica hidrosoluble……………………………………………….30
6.1.5.3.1 Materia orgánica hidrosoluble en sedimentos…………..……….30 6.1.5.3.2 Materia orgánica hidrosoluble en aguas………………….………30
6.1.5.2 Clorofilas…………………………………...……………………………………………..30 6.1.5.3 HAP´s en agua, sedimento y tejidos…………………………………………………..31
6.1.5.3.1 HAP´s en sedimentos…………………………………………………………31 6.1.5.3.2 HAP´s en aguas por extracción líquido-líquido…………………………...31 6.1.5.3.3 HAP´s en tejidos……………………………………………………………….31 6.1.5.4 PCB´s en sedimentos, agua y tejidos…………………………………………..31
6.1.5.4.1 PCB´s en sedimentos y aguas…...…………………………………….31 6.1.5.4.2 PCB´s en tejidos……………………………………………………...…32
6.2 Determinaciones ambientales (realizadas en campo)…………………………………………………...32 7. RESULTADOS…………………………………………………………………………………………………...33
7.1 PRIMER MUESTREO (sequìa fría)…………………………………………………………………….....33 7.1.1 Contenido de HAP´s en agua y sedimentos………………………………………………………...33 7.1.2. Contenido de HAP´s en hígado y vísceras de peces………………………………………............34 7.1.3. Factor de bioconcentración (FBC)…………………………………………………………………...36 7.1.4. Biomarcadores de estrés oxidativo, radicales libres………………………………………………36 7.1.5. Biomarcadores de daño……………………………………………………………………………….38 7.1.6. Biomarcadores de activación……...………………………………………………………………….40 7.1.7. Análisis de componentes principales……………………………………………………………….42
7.1.7.1 Biomarcadores vs. HAP´s. Hígado…………………………………………………………...42 7.1.7.2 Biomarcadores vs. HAP´s. Víscera…………………………………………………………...43 7.1.7.3 Materia orgánica, clorofilas y feofitina vs. HAP´s. Agua…………………………………45 7.1.7. 4 Materia orgánica vs. HAP´s. Sedimento……………………………………………………45
7.2 SEGUNDO MUESTREO (lluvias)…………………………………………………………………………47 7.2.1. Contenido de HAP´s en agua y sedimentos………………………………………………………..47 7.2.2 Contenido de HAP´s en hígado y vísceras de peces……………………………………………….48 7.2.3 Factor de bioconcentración……………………………………………………………………………50 7.2.4 Biomarcadores de estrés oxidativo, radicales libres……………………………………………….50 7.2.5 Biomarcadores de daño………………………………………………………………………………..52 7.2.6 Biomarcadores de activación………………………………………………………………………….54
7.2.7 Análisis de componentes principales……………………………………………………………….56 7.2.7.1Biomarcadores vs. HAP´s. Hígado……………………………………………………………56 7.2.7.2 Biomarcadores vs. HAP´s. Víscera…………………………………………………………...56 7.2.7.3 Materia orgánica, clorofilas y feofitina vs. HAP´s. Agua…………………………………58 7.2.7.4 Materia orgánica, clorofilas y feofitina vs. HAP´s. Sedimentos…………………………58
8. DISCUSIÓN………………………………………………………………………………………………………61 8.1 Niveles de HAP´s en el agua, sedimento y órganos (hígado y vísceras)………...…………………...61 8.2 Factor de bioconcentración (FBC)………………………………………………………………………….63 8.3 Biomarcadores…………………...…………………………………………………………………………...64
9. CONCLUSIONES………………………………………………………………………………………………..66 10. LITERATURA CITADA……………………………………………………………………………………….67

i
RELACIÓN DE FIGURAS Fig. No.
Página
Fig. 1. Estructura básica de los PCB´s. 1
Fig. 2. Estructura química del naftaleno, pireno y benzo[a]pireno. 6
Fig. 3. Biotransformación del benzo[a]pireno. 3
Fig. 4. Peroxidación de lípidos. 14 Fig. 5. Generación de hidroperóxidos lipídicos. 15
Fig. 6. Formación del sulfóxido de metionina y de la sulfona de metionina. 16
Fig. 7. Formación de alcoholes como resultado de la peroxidación de lípidos. 17
Fig. 8. Ciclo catalítico del citocromo P450. 18
Fig. 9. Laguna de Zumpango. 21
Fig. 10. Zona lacustre del Valle de México, indicando la ubicación actual de las zonas que circundaban a este cuerpo de agua.
22
Fig. 11. Ejemplares de Chirostoma riojai. 23
Fig. 12. Diagrama de flujo de las determinaciones realizadas en laboratorio y campo.
27
Fig. 13.
PRIMER MUESTREO (sequía fría)
Contenido de HAP´s en el agua colectada en las estaciones a diferentes profundidades.
33
Fig. 14. Contenido de HAP´s evaluado en los sedimentos. 34
Fig. 15.
Determinaciones en peces (hígado y vísceras)
Contenido de pireno.
35
Fig. 16. Contenido de benzo[a]pireno. 35
Fig. 17. Factor de bioconcentración de los HAP´s. 36
Fig. 18. Niveles de peróxido de hidrógeno (H2O2). 37
Fig. 19. Niveles de superóxido (O2-). 37
Fig. 20. Niveles de formaldehído. 38
Fig. 21. Substancias reactivas al ácido tiobarbitúrico (TBAR´s). 39
Fig. 22. Hidroperóxidos lipídicos . 39
Fig. 23. Oxidación de proteínas. 40
Fig. 24. Actividad del CYP2E1. 41
Fig. 25. Actividad de la GSTT. 41
Fig. 26. Gráfico bidimensional de puntuaciones sobre los componentes principales en hígado. 42
Fig. 27. Gráfico bidimensional de puntuaciones sobre los componentes principales en víscera.
44
Fig. 28.
Otras determinaciones
Gráfico bidimensional de puntuaciones sobre los componentes principales en agua.
45
Fig 29. Gráfico bidimensional de puntuaciones sobre los componentes principales en sedimento.
46

ii
Fig. No. Página
Fig. 30.
SEGUNDO MUESTREO (lluvias)
Contenido de HAP´s en el agua colectada en las estaciones a diferentes profundidades.
47
Fig. 31. Contenido de HAP´s evaluado en los sedimentos. 48
Fig. 32.
Determinaciones en peces (hígado y vísceras)
Contenido de pireno.
49
Fig. 33. Contenido de benzo[a]pireno. 49
Fig. 34. Factor de bioconcentración de los HAP´s. 50
Fig. 35 Niveles de peróxido de hidrógeno (H2O2). 51
Fig. 36. Niveles de superóxido (O2-). 51
Fig. 37. Niveles de formaldehído. 52
Fig. 38. Substancias reactivas al ácido tiobarbitúrico (TBAR´s). 53
Fig. 39. Hidroperóxidos lipídicos. 53
Fig. 40. Oxidación de proteínas. 54
Fig. 41. Actividad del CYP2E1. 55
Fig. 42. Actividad de la GSTT. 55
Fig. 43. Gráfico bidimensional de puntuaciones sobre los componentes principales en hígado. 56
Fig. 44. Gráfico bidimensional de puntuaciones sobre los componentes principales en víscera. 58
Fig. 45.
Otras determinaciones Gráfico bidimensional de puntuaciones sobre los componentes principales en agua.
59
Fig. 46. Gráfico bidimensional de puntuaciones sobre los componentes principales en sedimento.
60

iii
RELACIÓN DE TABLAS
Tabla No. Página
Tabla 1.
Propiedades fisicoquímicas de PCB´s con 5, 6 y 7 sustituciones de cloro en su estructura.
2
Tabla 2. Toxicidad aguda de algunas formulaciones de PCB´s. 4
Tabla 3. Propiedades físicas del naftaleno, pireno y benzo[a]pireno. 7
Tabla 4. Propiedades físicoquímicas del naftaleno, pireno y benzo[a]pireno. 7
Tabla 5.
Destino de los contaminantes orgánicos en los sedimentos con base en su Koc y su solubilidad en el agua.
7
Tabla 6. Clasificación de la peligrosidad del benzo[a]pireno. 9
Tabla 7.
Longitudes de onda (λ) para determinar los HAP´s considerados en este estudio.
31
Tabla 8.
PRIMER MUESTREO (sequía fría)
Matriz de correlaciones entre biomarcadores e HAP´s, evaluados en hígado.
43
Tabla 9. Matriz de correlaciones entre biomarcadores e HAP´s, evaluados en víscera. 44
Tabla 10. Matriz de correlaciones entre materia orgánica, pigmentos fotosintéticos e HAP´s, evaluados en agua.
45
Tabla 11. Matriz de correlaciones entre materia orgánica e HAP´s, evaluados en sedimentos.
46
Tabla 12.
SEGUNDO MUESTREO (lluvias)
Matriz de correlaciones entre biomarcadores e HAP´s, evaluados en hígado.
57
Tabla 13. Matriz de correlaciones entre biomarcadores e HAP´s, evaluados en víscera. 57
Tabla 14. Matriz de correlaciones entre materia orgánica, pigmentos fotosintéticos e HAP´s, evaluados en agua.
59
Tabla 15. Matriz de correlaciones entre materia orgánica e HAP´s, evaluados en sedimentos.
60

iv
ABREVIATURAS
ACP´s análisis de componentes principales CYP 2E1 citocromo P450 isoforma E1 DL50 concentración letal media GSH glutatión reducido HAP´s hidrocarburos aromáticos policícliclos H2O2 peróxido de hidrógeno HOOo hidroperóxido lipídico Koc coeficiente de distribución en el suelo
Kow coeficiente de partición octanol-agua FBC factor de bioconcentración GSTT, GSTθ glutatión S-transferasa isoforma tetha LMP límite máximo permisible MAD malondialdehído m. o. materia orgánica MOD materia orgánica disuelta MOP materia orgánica particulada MOT materia orgánica total O2
- anión superóxido PCB´s bifenilos policlorinados TBAR´s substancias reactivas al ácido tiobarbitúrico USEPA, EPA Agencia de protección ambiental de los E. U. SÍMBOLOS mPa milipascales (unidad de presión) t ½ tiempo de vida media

v
RESUMEN
Como resultado de las actividades humanas en los ambientes dulceacuícolas pueden llegar a depositarse contaminantes con la capacidad de generar estrés oxidativo en los organismos acuáticos, tales como los bifenilos policlorados (PCB´s) y los hidrocarburos aromáticos policíclicos (HAP´s). Además, dichas substancias son biotransformados por las oxidasas de función mixta (tales como el CYP 2E1) y las enzimas dependientes del glutatión (como la GSTT) hacia compuestos más polares que sean más fácilmente eliminados por el organismo (detoxificación). Sin embargo, también pueden generarse especies químicas con mayor toxicidad a la de los compuestos originales (bioactivación). Por otra parte, ambos procesos pueden ser modificados por la presencia de otros contaminantes ambientales. En conjunto, la respuesta tóxica sobre los organismos acuáticos puede variar dependiendo de los xenobióticos en cuestión, la especie de que se trate, la etapa del ciclo de vida en que se encuentre, sus hábitos alimenticios y la época del año, entre otros. En particular, el charal de Santiago (Chirostoma riojai) es una especie dulceacuícola endémica de la zona central de nuestro país. Fue descrita hace poco más de 50 años, pero debido a la destrucción de su hábitat, la contaminación, el uso de agua para riego, actividades agrícolas, industriales y domésticas, el número de localidades donde habita se ha reducido drásticamente, por lo que actualmente se encuentra en peligro de extinción. Por lo anterior, para el presente trabajo se decidió evaluar la respuesta pro-oxidante del charal de Santiago expuesto a mezclas ambientales complejas de PCB´s e HAP´s en la Laguna de Zumpango, Estado de México. En la Laguna de Zumpango se colectó agua (en la superficie, en el nivel de compensación y en el fondo) además de sedimentos, en tres estaciones representativas del cuerpo de agua para el análisis de compuestos orgánicos halogenados (COH) [bifenilos policlorados (PCB´s): mono-ortho-substituidos, di-ortho-substituidos y no-ortho-substituidos e hidrocarburos aromáticos policíclicos (HAP´s): naftaleno, pireno y benzo[a]pireno (BaP)], además de parámetros fisicoquímicos evaluados in situ y en laboratorio. Los charales fueron divididos en tres grupos de acuerdo a su relación peso-talla. En hígado y vísceras se analizaron biomarcadores de la respuesta pro-oxidante (concentración de peróxido de hidrógeno, hidroperóxidos lipídicos, lipoperoxidación, oxidación de proteínas) y de la bioactivación (actividad del CYP2E1 y de la GSTT). También se evaluó el contenido de HAP´s en estos órganos. Los resultados se analizaron por medio de ANOVA seguido de la prueba de Dunnett. Las relaciones entre los biomarcadores (variables dependientes) y la concentración de HAP (variables independientes) se probó por regresión lineal y exponencial.

vi
No detectaron PCB´s en la columna de agua, mientras que la concentración de HAP´s presentó un patrón diferencial en los compartimentos ambientales. En el agua sólo se detectó naftaleno, pero en los sedimentos el compuesto más abundante fue pireno, seguido del benzo[a]pireno y en la estación más contaminada (embarcadero) se encontraron los tres HAP’s bajo estudio. Estos hallazgos denotan que la actividad antropogénica es determinante en la contaminación por HAP´s de este cuerpo de agua. En hígado y vísceras se cuantificaron elevados factores de bioconcentración (FBC) de pireno y bajos para el BaP. El hígado tuvo un FBC de pireno >2,000 y en vísceras fue mayor a 2,500. Sin embargo, para el BaP el FBC fue menor a 10. Al respecto, es posible sugerir interacciones complejas entre los componentes bióticos y abióticos. Respecto a la generación de radicales libres todos ellos fueron detectados y además hubo diferencia entre los tejidos. En ningún caso se halló lipoperoxidación significativa, pero los hidroperoxidos lipídicos estuvieron incrementados. Esto demuestra que el estrés oxidativo se detiene en fases tempranas. También se encontró oxidación de proteínas. En hígado y víscera, los hidroperoxidos lipídicos y la lipoperoxidación, además de la oxidación de proteínas tuvieron una relación significativa con los niveles de pireno (p<0.001). Estos resultados sugieren que el daño oxidativo está asociado con el metabolismo realizado por las enzimas de la función oxidasa, específicamente las isoenzimas del citocromo P450, por lo que es posible concluir que la relación entre los HAP’s y las fuerzas endógenas pro-oxidantes generan estrés oxidativo y en el caso de los fosfolípidos, los daños se detienen en etapas tempranas pero en las proteínas, ocurre una oxidación significativa.

vii
ABSTRACT
As a result of the human activities, pollutants in the freshwater ecosystems able to induce oxidative stress in the aquatic organisms can be loaded, such as polychlorinated biphenyls (PCB´s) and polyaromatic aromatic hydrocarbons (PAH´s). Also, this substances are biotransformed by mixed oxidase function enzymes (such as the CYP 2E1) and by the glutathione dependent enzymes (as the GSTT) to a more polar compounds that could be eliminated by the organism more easily (detoxification). However, as a consequence of this process, chemical species with higher toxicity regarding to original compounds can also be generated (bioactivation). On the other hand, both processes can be modified by the occurrence of other environmental pollutants. Overall, the toxic response on the aquatic organisms can vary depending of the type of xenobiotics, of the species, of the stage of lifespan, of its feeding behavior, and the season, among others. In particular, the Toluca silverside (Chirostoma riojai) is a freshwater species endemic of the Central Plateau of Mexico. It was described more than 50 years ago, but due to its habitat destruction, by the contamination, by the use of water for watering, by agricultural, industrial and domestic activities, the number of its localities has decreased drastically. As a consequence, this day its status is endangered. For the above-mentioned, in the present study we decided to evaluate the pro-oxidant response of the Toluca silverside exposed to environmental mixtures of PCB´s and PAH´s in Zumpango Lake, State of Mexico. In Zumpango Lake, water samples in the surface, in the compensation level and in the bottom waster, also sediments were collected in three representative stations of the water body for the analysis of halogenated organic compounds (HOC) [polychlorinated biphenyls (PCB’s): mono-ortho-substituted, di-ortho-substituted and non-ortho-substituted, and also polycyclic aromatic hydrocarbons (PAH´s): naphthalene, pyrene and benzo[a]pyrene (BaP)] besides physicochemical parameters evaluated in situ and in laboratory. The specimens were divided in three groups according to its weight-size relationship. In liver and viscera, biomarkers of pro-oxidant response (concentration of peroxide of hydrogen, lipid hydroperoxides, lipid peroxidation and carbonyl proteins) were assayed, also biomarkers of bioactivation (activity of the CYP2E1, and of the GSTT) as well as the content of PAH´s in the organs were analyzed. The results were analyzed by means of ANOVA followed by Dunnett test. The relationships among the biomarkers (dependent variables) and the concentration of HOC (independent variables) it was proven by lineal and exponential regression. PCB’s in the water column were not detected, while the concentration of PAH’s have a differential pattern in the environmental compartments. In the water column only naphthalene was detected, but in sediments the most copious hydrocarbon was the pyrene followed by the benzo[a]pyrene. In the most polluted station (harbor) three of

viii
the PAH’s under study were founded. These findings denote that the antropogenic activity is crucial in the contamination of this water body with PAH’s. In liver and viscera high bioconcentración factors (FBC) of pyrene followed by BaP were quantified. The liver had a BCF of pireno >2,000 and in viscera was higher at 2,500. However, for the BaP, the FBC was smaller at 10. In this respect it is possible to suggest complex interactions between the biotic and abiotic compartments. Regarding to the generation of free radicals, all of these were detected, as well as differences among the studied tissues were noted. In any case lipid peroxidation was observed; however, the lipid hydroperoxides content were detected. This finding denotes that the oxidative stress stops in early phases of the chain reaction. Also, carbonyl proteins were noted in both tissues. In liver and viscera, the lipid hydroperoxides and the lipid peroxidation as well as the carbonyl proteins had a significant relationship with the levels of pyrene (p <0.001). These results show that the oxidative damage is associated with the metabolism carried out by the enzymes of the mixed oxidase function, specifically the cytochrome P450 isoenzymes. It is possible to conclude that the relationship between the PAH’s with the pro-oxidizers endogenous forces generates oxidative stress, in the case of the fosfolipids of membranes the damages stops in early stages, but in case of the proteins, it occurs in a significant magnitude.

1
1. INTRODUCCIÓN El charal de Santiago (Chirostoma riojai ) es una especie dulceacuícola que se encuentra en la lista roja de especies en peligro de extinción. Se considera la degradación de su hábitat como la principal causa de la disminución de las poblaciones naturales de esta especie (Figueroa-Lucero y col., 2003). Por otro lado, en los ambientes dulceacuícolas se pueden acumular contaminantes ambientales con el potencial de causar estrés oxidativo en los organismos acuáticos, tales como los hidrocarburos aromáticos policíclicos (HAP´s) y los bifenilos policlorinados (PCB´s) (Di Giulio y col., 1989). Dichos contaminantes son biotransformados y bioactivados por las enzimas de la función mixta oxidasa, como el CYP2E1 y por enzimas de la fase II, como las pertenecientes a la familia de la glutatión-S-transferasa (GST), tal es el caso de la GST isoforma tetha (GSTT). 2. ANTECEDENTES 2.1 BIFENILOS POLICLORADOS Los bifenilos policlorados (PCB´s por sus siglas en inglés) son una serie de aceites de uso industrial con alta resistencia al calor, formados por la unión de dos anillos aromáticos (Fig. 1) los cuales pueden tener hasta 10 substituciones de átomos de cloro (Watts, 1998; Eljarrat y Barceló, 2003).
Fig 1. Estructura básica de los PCB´s (Merck, 1996).
Estos compuestos se utilizaron por más de ocho décadas en sistemas de refrigeración, la industria eléctrica (transformadores y condensadores) y en menor grado como fluidos hidráulicos, lubricantes, plastificantes, impermeabilizantes, retardantes de flama, pinturas y pigmentos (PNUMA y FAO, 1992; Moltó y col., 1998). Se comenzaron a producir en 1929 por la compañía Monsanto (Estados Unidos) con el nombre de Aroclor, debido a la necesidad de contar con un aceite enfriador para transformadores eléctricos que tuviera propiedades dieléctricas y fuera más estable y menos inflamable que el aceite mineral utilizado hasta entonces y de esta manera evitar que los arcos voltaicos producidos en las plantas eléctricas pudieran provocar la ignición del mismo (Watts, 1998; Fernández-Bremauntz y col., 2004).

2
Ejemplos de otros nombres comerciales para los PCB´s son Chlorpen (Alemania) y Kanechlor (Japón) (Moltó y col., 1998), Chloerxtol, Clophen, Dykanol, Fenclor, Inerteen, Noflamol, Phenoclor, Pyralene, Pyranol, Santotherm, Sovol y Therminol (PNUMA y FAO, 1992). Al nivel mundial, los principales productores son Monsanto (E.U.A), Caffaro (Italia), Kanegafuchi (Japón), Rhône-Poulenc (Francia), Atochem (Francia), Bayer (Alemania) y Chemko (Checoslovaquia) (PNUMA y FAO, 1992). Los bifenilos policlorados son producidos por medio de la cloración de un esqueleto bifenilo en presencia de un catalizador (Moltó y col., 1998). Teóricamente se pueden formar hasta 209 productos distintos ó congéneres (denominados así debido a que se generan de manera simultánea en el proceso de cloración) (Moltó y col., 1998; Eljarrat y Barceló, 2003).Ya que no es posible controlar el número de substituciones de Cl ó la posición en que éstas ocurren (Watts, 1998), las formulaciones comerciales de PCB´s no contienen un solo congénere, sino mezclas complejas de hasta 132 de éstos compuestos (Bommanna y col., 1994). Estas características dificultan la determinación de estos compuestos en las diferentes matrices ambientales (EPA, 1996a; EPA, 1996b; Eljarrat y Barceló, 2003), a la vez que son de relevancia toxicológica, ya que la respuesta tóxica producida por la mezcla puede ser diferente a la causada por sus componentes de manera individual. Por ejemplo, si uno de ellos estimula ó inhibe la biotransformación de otro componente de la mezcla puede modificar su toxicidad y la forma en que es distribuido y acumulado en los tejidos (Kleinow y col., 1987)). Otros factores que pueden modificar la respuesta tóxica son el patrón de sustitución de los átomos de cloro sobre el anillo bifenilo y el porcentaje de cloración (Ifremer, 2005), ya que al aumentar el número de sustituciones de cloro se incrementa también su toxicidad (Repetto,1997; Eljarrat y Barceló, 2003). En la tabla 1 se muestran las propiedades fisicoquímicas de algunos PCB´s: Tabla 1. Propiedades fisicoquímicas de PCB´s con 5, 6 y 7 sustituciones de cloro en su estructura (Ifremer, 2005).
Compuesto Peso molecular Solubilidad en agua a 25 oC
(μg/L)
Presión de vapor a 25 oC
(mPa)
Log Kow
PCB 105 (5) 326.4 4-72 0.304-30 6.53-6.93
PCB 156 (6) 360.9 0.40-38 0.2-12 6.7-7.3
PCB 180 (7) 395.3 0.31-6.56 0.086-0.506 7.14-7.36 Nota: Los números entre paréntesis se refieren al número de átomos de Cl- en la molécula.
Por su elevada persistencia y toxicidad, en 1979 se prohibió la producción y se limitó la distribución de PCB´s en E.U.A. Actualmente son considerados carcinógenos por la Agencia de Protección Ambiental de Estados Unidos (EPA) y la Agencia Internacional

3
para la Investigación en Cáncer (IARC, por sus siglas en inglés) (Moltó y col., 1998). Además, están incluidos en la Lista de Contaminantes Orgánicos Persistentes (COP´s), adoptada en el Convenio de Estocolmo, Suecia, en 1991 (Eljarrat y Barceló, 2003; Fernández-Bremauntz et al., 2004) y en la Lista de Contaminantes Prioritarios de la Unión Europea (Bro-Rasmussen, 1994). Sin embargo, en México y otros países dichos compuestos aún se encuentran presentes en los transformadores y otros dispositivos eléctricos (Watts, 1998). En nuestro país aún permanecen unas 5,000 ton de PCB´s sin destruir (Fernández-Bremauntz et al., 2004). 2.1.1 Toxocinética ambiental Los PCB´s con 5 ó más átomos de cloro son muy resistentes a la biodegradación (PNUMA y FAO, 1992) ya que al aumentar el porcentaje de cloración disminuye su solubilidad en agua al igual que su capacidad de evaporación y volatilización. Sin embargo, también se incrementa su afinidad por los tejidos adiposos y su adsorción en los sedimentos. Estos fenómenos les permiten biacumularse y bioconcentrarse en los organismos (Ifremer, 2005). Los principales congéneres de PCB´s que pueden bioacumularse poseen 5 a 7 átomos de cloro (penta, hexa y heptacloro bifenilos), ya que los compuestos con mayor número de sustituciones de este halógeno se unen firmemente a los sedimentos y de esta manera se reduce la biodisponibilidad para los organismos. Aquellos con menor proporción de cloración son rápidamente biotransformados y eliminados por lo que son biacumulados de manera reducida (Safe y col., 1982). En peces y crustáceos se han encontrado factores de bioconcentración (FBC) de hasta 270,000 (PNUMA y FAO, 1992). Una vez en los sedimentos, los PCB´s se unen fuertemente al carbón orgánico (Manahan, 2005), por lo que la vida media de estos compuestos (t ½) puede alcanzar alrededor de 5 años (PNUMA y FAO, 1992). Bajo estas condiciones ciertas bacterias anaerobias de los sedimentos pueden modificar la biodisponibilidad de los PCB´s por su elevado tiempo de residencia reduciendo la proporción de congéneres con mayor número de átomos de cloro y aumentando de manera simultánea, el porcentaje de compuestos menos clorados. Ya que los PCB´s con mayor grado de cloración actúan como aceptores de electrones durante la respiración microbiana anaerobia a este proceso se le denomina deshalorespiración (Manahan, 2005). Por otro lado, en compuestos con más de cinco átomos de cloro también puede ocurrir en cierta medida la degradación por fotólisis (PNUMA y FAO, 1992).

4
Watts (1998) sugiere que las formulaciones de PCB´s con más de cinco átomos de Cl sean consideradas de riesgo elevado debido a su alta persistencia y toxicidad. 2.1.2. Efectos tóxicos Los PCB´s y sus congéneres pueden ser absorbidos a través de la piel, pulmones y tracto gastrointestinal. A nivel ocupacional, las principales vías de exposición son la pulmonar y dérmica. Por vía oral, la principal fuente de exposición es a través de la ingestión de pescado contaminado (PNUMA y FAO, 1992; Ford y col., 2001). Algunos de los efectos provocados por los PCB´s son inhibición de la acción de la dopamina y otros neurotrasmisores (Fielder y Martin, 1994), inmunosupresión, promoción de tumores e interferencia con la utilización del calcio (Dean y col., 1994). A nivel agudo, pueden causar náuseas, letargo, pigmentación café en la piel y uñas, edema subcutáneo en el rostro, aspecto peculiar en los folículos capilares, lagrimeo excesivo, dificultad en la visión y problemas gastrointestinales (Merck, 1996). En organismos acuáticos (peces), se han reportado valores de DL50 entre 3 y 3000 ug/L (PNUMA y FAO, 1992). A continuación se presentan los valores de DL50 reportados para algunos productos comerciales (Tabla 2): Tabla 2. Toxicidad aguda de algunas formulaciones de PCB´s
(PNUMA y FAO, 1992)
Organismo Formulación DL50 (mg/kg)
Rata (vía oral) Aroclor 1221(*) 4,000
Rata (vía oral) Aroclor 1268(*) 10,000
Conejo Aroclor 1242(*) 1,000
Conejo Aroclor 1268(*) 3,000 (*): Los últimos dos dígitos del nombre del producto indican el porcentaje de cloración
en peso en la formulación (p. ej., el Aroclor 1221 posee un 21% de átomos de cloro)
A nivel crónico en humanos, el signo más característico de intoxicación es la aparición de cloracné en regiones específicas del cuerpo (por debajo y al lado de los ojos, detrás de las orejas), pero también se puede presentar lesiones cutáneas sobre el cuello, hombros, abdomen, escroto y pene en el caso de los varones (Ford y col., 2001). Daño hepático, reducción en la capacidad pulmonar, irritación ocular y disminución en el peso del producto en madres expuestas (Merck, 1996) y alteración del metabolismo del Ca++ por vía hormonal (PNUMA y FAO, 1992).

5
También se ha documentado el potencial de los PCB´s y sus metabolitos para alterar los procesos reproductivos en el hombre y los peces, al interferir con la función endócrina (Ifremer, 2005). Sin embargo, tal vez el efecto más importante a nivel crónico sea su carcinogenicidad, la cual se ha observado en animales de laboratorio y es probable que también ocurra en el hombre (PNUMA y FAO, 1992). Esto sucede a través de mecanismos epigénicos tales como la inducción de enzimas, el incremento en la multiplicación celular y alteraciones en la señalización (Ford y col., 2001). Según los datos epidemiológicos los principales órganos afectados son el hígado, los conductos biliares y la vesícula biliar (PNUMA y FAO, 1992). A consecuencia del elevado número de congéneres de PCB´s que pueden contener las formulaciones comerciales y para facilitar los estudios relacionados con estos compuestos se han agrupado en 5 categorías de daño (1A, 1B, 2, 3 y 4). Además, se reducido su número de 209 a 36 congéneres basándose en su potencial para causar toxicidad, la capacidad para inducir las enzimas involucradas en su bioactivación, ocurrencia en el ambiente y abundancia relativa en los tejidos animales (McFarland y Clarke, 1989). En el estudio de la toxicidad producida por los PCB´s se ha utilizado el concepto de Factor Equivalente de Toxicidad (TFE, por sus siglas en inglés), el cual está basado en los siguientes criterios: a) el compuesto debe poseer similitud estructural con la 2,3,7,8-tetraclorodibenzo-p-dioxina (TCDD) considerada el contaminante ambiental más potente conocido (Safe, 1987), b) la respuesta tóxica del compuesto debe estar mediada por su unión al receptor aril hidrocarburo (AhR), y 3) el compuesto debe ser persistente y bioacumularse a lo largo de la cadena alimenticia (Eljarrat y Barceló, 2003). 2.1.3 Biotransformación Su biotransformación ocurre en el hígado a través de las isoenzimas del citocromo P450 (CYP 450) y de las glucuronil transferasas (Fielder y Martin, 1994). Posteriormente, pueden ser eliminados a través de la bilis y orina. En el hombre, el tiempo de vida media varía directamente con el grado de cloración del compuesto, de 6 meses en los menos sustituidos a más de 2 años en los altamente clorados (Ford y col., 2001). 2.1.4 Normatividad Se ha establecido un límite máximo permisible (LMP) para la protección de la vida acuática para los PCB´s de 30 ug/L (ppb) (PNUMA y FAO, 1992).

6
Sin embargo, no existen valores de referencia similares en la legislación ambiental mexicana para estas substancias y únicamente se hallan considerados en la NOM-133-ECOL-2000, la cual establece las especificaciones de protección ambiental para el manejo de PCB´s, materiales contaminados y equipos que los contengan (Fernández-Bremauntz, 2004). 2.2 HIDROCARBUROS AROMÁTICOS POLICÍCLICOS (HAP´s) Los hidrocarburos polinucleares (PNA´s, por sus siglas en inglés) son una serie de compuestos aromáticos heterocíclicos formados por la fusión de dos o más anillos bencénicos (Furton y Pentzke, 1998), los cuales pueden tener substituciones de átomos de oxígeno, nitrógeno ó azufre. Entre éstos, se hallan los hidrocarburos aromáticos policíclicos (PAH´s, por sus siglas en inglés) formados únicamente por carbono e hidrógeno (Watts, 1998). Los HAP´s son moléculas moderadamente estables; sus fuentes de emisión principales provienen de la combustión incompleta de combustibles fósiles (tales como carbón, petróleo y gasolina) y materia orgánica, pero también pueden encontrarse en las fracciones más pesadas de los derivados del petróleo (p. ej. aceites lubricantes y asfalto) y en el humo del cigarrillo (Watts, 1998; Cram y col., 2004). El más sencillo de los HAP´s es el naftaleno, el cual contiene únicamente dos anillos bencénicos unidos entre sí (Fig. 2). Los otros HAP´s considerados en el presente trabajo son el naftaleno, pireno y benzo[a]pireno, los cuales poseen 2, 4 y 5 anillos fusionados, respectivamente (Fig. 2).
Fig. 2. Estructura química del naftaleno, pireno y benzo[a]pireno (Merck, 1996).
Sin embargo, en al ambiente se pueden encontrar mezclas extremadamente complejas de HAP´s, incluyendo isómeros y formas alquiladas (Furton y Pentzke, 1998). Las propiedades fisicoquímicas de estos compuestos están influidas por el número de anillos bencénicos y en consecuencia, por su peso molecular (Ifremer, 2005). A continuación, se presentan las propiedades físicas de los compuestos considerados en este estudio (Tablas 3 y 4):
Naftaleno Pireno Benzo[a]pireno

7
Tabla 3. Propiedades físicas del naftaleno, pireno y benzo[a]pireno (Merck, 1996)
Compuesto No. registro CAS
Peso molecular
Densidad Punto fusión Punto ebullición
naftaleno 91-20-3 128.17 1.162 80.2 oC 217.9 oC
pireno 129-00-0 202.26 1.271 156 oC 404 oC
benzo[a]pireno 50-32-8 252.32 179.3 oC 310 oC
Tabla 4. Propiedades físicoquímicas del naftaleno, pireno y benzo[a]pireno (Cram y col., 2004)
Compuesto
Solubilidad en agua (mg/L)
Presión de vapor (KPa)
Log Kow
Log Koc
(L kg-1)
naftaleno 31.0 0.012 3.37 -
pireno 0.14 3.33 x 10-7 5.18 4.22-5.65
benzo[a]pireno 3.8 x 10-3 7.45 x 10-10 6.04 5.48-6.74 Nota: Kow se refiere al coeficiente de partición octanol-agua y Koc, al coeficiente de distribución del compuesto orgánico entre el suelo y el agua de poro.
En particular, Ney (1990) ha propuesto un esquema para predecir el comportamiento de los contaminantes orgánicos en los sedimentos utilizando su coeficiente de distribución en el suelo (Koc) y su solubilidad en agua (Tabla 5):
Tabla 5. Destino de los contaminantes orgánicos en los sedimentos con base en su Koc y su solubilidad en el agua (SA) (modificado de Ney, 1990).
Log Koc < 3 SA> 1,000 ppm
Log Koc >4 SA< 10 ppm
Adsorción - X
Solubilidad X -
Movilidad X -
Biodegradación X -
Dispersión X -
Acumulación - X
Bioacumulación - X
Persistencia - X

8
2.2.1 Toxocinética En el hombre la exposición a estos compuestos puede darse a través de alimentos contaminados, de manera ocupacional ó por el asfalto aplicado a las carreteras (Furton y Pentzke, 1998). Al incrementarse el número de anillos de los HAP´s, su solubilidad en agua disminuye y al igual que en el caso de los PCB´s, su adsorción en los sedimentos y la acumulación en los tejidos adiposos de los organismos aumenta (Ifremer, 2005). En los sedimentos, se unen principalmente a la fracción constituida por carbón orgánico (Manahan, 2005). En el ambiente, los HAP´s pueden sufrir foto-oxidación por la acción de la luz ultravioleta, al absorber y transferir la energía de la radiación UV hacia el oxígeno molecular generando de esta manera especies reactivas, las cuales pueden dañar los tejidos de manera directa ó al promover reacciones de óxido-reducción y provocan la muerte celular (Ifremer, 2005). Se ha observado este efecto fototóxico inducido por la luz UV sobre los HAP´s en células humanas, plantas, peces, anfibios y zooplancton (Cram y col., 2004). 2.2.2 Efectos tóxicos 2.2.2.1 Naftaleno A nivel agudo, se ha reportado que puede causar envenenamiento sin importar la vía de absorción (oral, inhalatoria ó dérmica), el cual se manifiesta principalmente por la presencia de anemia hemolítica (USEPA, 2003). Sin embargo, también pueden ocurrir náuseas, vómito, dolor de cabeza, diaforesis, hematuria, fiebre, necrosis hepática, ictericia, agrandamiento del bazo, convulsiones y coma (Merck, 1996). 2.2.3 Biotransformación de los HAP’s Una vez que han ingresado al organismo, los HAP´s pueden ser biotransformados en compuestos menos tóxicos y luego eliminados (detoxificación) ó bioactivados hacia metabolitos reactivos con mayor toxicidad que el compuesto original, con potencial mutagénico ó carcinogénico (Ifremer, 2005). En particular, el naftaleno y pireno son bioactivados por el CYP 1A1 y el CYP 2E1 (Yang y col., 1999).

9
2.2.3.1 Naftaleno En el hígado, este compuesto es oxidado hacia metabolitos reactivos (USEPA, 2003) como naftol y naftoquinona, pero también puede ser conjugado directamente con el glutatión para formar tioéteres (Leikin y Paloucek, 2008). Dichos metabolitos reaccionan posteriormente con enzimas y macromoléculas de membrana, ejerciendo su efecto tóxico sobre la sangre, el ojo y en estudios animales, afectando también el pulmón (USEPA, 2003). Un dato sobresaliente es el hecho que los individuos con deficiencia de la enzima glucosa 6-fosfato deshidrogenasa son más susceptibles a desarrollar anemia hemolítica luego de la exposición a este compuesto. Esto se asocia también con niveles de glutatión reducido (GSH) bajos, el cual se encarga de proteger a los eritrocitos contra daño oxidativo (USEPA, 2003). Una situación parecida puede darse en los organismos juveniles, ya que las vías encargadas de la detoxificación de los metabolitos reactivos aún no se encuentran bien desarrolladas (USEPA, 2003). 2.2.3.1 Benzo[a]pireno Después de la oxidación inicial del benzo[a]pireno por el CYP 450 (Fig. 3) se forma un 7,8 epóxido, el cual es transformado por la acción de la enzima epóxido hidrolasa en un 7,8 diol. Este último compuesto es oxidado nuevamente por las oxidasas de función mixta hacia el antiisómero (+)7,8-diol-9,10-epóxido (Manahan, 2003), el cual ha demostrado propiedades mutagénicas y carcinogénicas en animales de laboratorio (Rubin, 2001). Por lo anterior, este compuesto ha sido considerado como potencialmente carcinógeno para el hombre por diversas instancias en los E.U.A.(Tabla 6). Tabla 6. Clasificación de la peligrosidad del benzo[a]pireno (James y Saranko, 2000).
Organismo Clase de peligro
ACGIH A2: sospechoso de ser carcinógeno en el hombre
NIOSH Substancias ocupacionales potencialmente carcinógenas
NTP Agentes potencialmente carcinógenos en el hombre
IARC 2A: Posiblemente carcinógeno para el hombre (de manera individual)
ACGIH: American Conference of Governmental Industrial Hygienists; NIOSH: National Institute for Occupational Safety and Health; NTP: National Toxicology Program; IARC: International Agency for Research on Cancers.

10
Además, el Programa de las Naciones Unidas para el Medio Ambiente (PNUMA) ha considerado ampliar la lista de contaminantes orgánicos persistentes (COP´s) para incluir a los HAP´s (Fernández-Bremauntz y col., 2004).
Fig. 3. Biotransformación del benzo[a]pireno (modificado de Manahan, 2003).
CYP 450
Benzo[a]pireno (+)Benzo[a]pireno7,8 epóxido
epóxido hidrolasa
H2O
(+)Benzo[a]pireno7,8 dihidrodiol
(+)Benzo[a] pireno7,8 diol-9, 10-epóxido-2 (carcinogénico)
CYP 450

11
2.2.4 Normatividad En Estados Unidos, se ha establecido un Límite de Referencia para la Salud (HRL, por sus siglas en inglés) para naftaleno de 140 ug/L, equivalentes a 0.14 ppm (USEPA, 2003). En nuestro país, las Normas Oficiales Mexicanas no consideran Límites Máximos Permisibles (LMP´s) para la protección de la vida acuática para los HAP´s (Scram y col., 2004). 2.3 Biomarcadores Un biomarcador es una variación ya sea en los componentes ultraestructurales, al nivel de organelos, en los procesos bioquímicos o fisiológicos y que son medibles en un sistema biológico ó en muestras y además puede ser inducido por los xenobióticos (NRC, 1989). Tales variaciones pueden indicar la magnitud de la respuesta del organismo a los contaminantes, así como establecer una relación causal entre la presencia de una substancia química y su efecto (Fosi, 1994). Los biomarcadores pueden clasificarse en tres tipos (NRC, 1997):
a) De exposición. Se refieren a la cuantificación de un compuesto ó su metabolito, ó a una respuesta única atribuible a un compuesto ó grupo de ellos.
b) De efecto. Son respuestas en el organismo que puedan estar directamente
relacionadas con la exposición.
c) De susceptibilidad. Es la medición de las capacidades innatas ó inducidas del organismo para responder a un tóxico ambiental.
2.3.1 Uso de biomarcadores en estudios ecotoxicológicos En este tipo de estudios, se puede evaluar el estado de salud del ecosistema utilizando especies centinela como bioindicadores ó de modo más específico, investigar el estado de salud de una población ó de una especie en riesgo expuesta a contaminantes ambientales (Fosi, 1994).

12
2.3.1.1 Radicales libres y especies reactivas de oxígeno Los radicales libres son especies químicas con uno ó más electrones desapareados (Halliwell y Cross, 1994). Hace más de medio siglo, se propuso que la generación de radicales libres podría ser la responsable de los efectos tóxicos provocados por el oxígeno molecular (O2) (Gerschman y col., 1954). Esta hipótesis se popularizó y fue convertida en la teoría de la toxicidad del O2 causada por el superóxido (O2.), luego del descubrimiento de la enzima superóxido dismutasa (SOD) en 1969 por McCord y Fridovich (Fridovich, 1989). Este radical, junto con el radical hidroxilo (OH.) y el peróxido de hidrógeno (H2O2), se denominan en conjunto especies reactivas de oxígeno (ROS, por sus siglas en inglés) (Halliwell y Cross, 1994). En los organismos aerobios, las especies reactivas de oxígeno pueden formarse de manera normal durante procesos que involucran la transferencia de electrones, tales como la cadena respiratoria mitocondrial (Könisberg, 1992), la biotransformación de xenobióticos por el CYP 450 (Fridovich, 1989), la auto-oxidación de flavinas, catecolaminas e hidroquinonas catalizada por metales de transición (Fe y Cu) presentes intracelularmente (Halliwell y Gutteridge, 1989) ó la incidencia de luz ultravioleta sobre las moléculas (Reppeto, 1997). En particular, se ha reportado la participación del O2. y H2O2 en funciones celulares de importancia, p. ej. como segundos mensajeros en cascadas de señalización (Valko y col., 2006; Gómez-Quiroz y Cuevas-Baena, 2008) y su producción por los neutrófilos y macrófagos durante los procesos inflamatorios como respuesta a agentes infecciosos (Valko y col., 2006). Sin embargo, el O2. y H2O2 son precursores del radical hidroxilo, el cual es el agente oxidante más potente encontrado en los sistemas biológicos (Halliwell y Gutteridge, 1989; Kruk, 1998). Debido a su elevada reactividad, el OH. puede reaccionar prácticamente con cualquier molécula biológica con que entre en contacto (Halliwell y Cross, 1994), por lo que puede generar estrés oxidativo y dañar importantes componentes celulares, tales como membranas, proteínas y ácidos nucleicos (Poli y col., 2004). 2.3.1.2 Estrés oxidativo Es un desbalance entre las fuerzas pro-oxidantes y antioxidantes lo cual puede desembocar en una serie de daños en la célula ó en los procesos adaptativos frente a dicho desbalance (Halliwell y Gutteridge, 1999).

13
En los organismos acuáticos, este desequilibrio puede ser producido por variaciones en la disponibilidad de oxígeno en los tejidos de los organismos (Storey, 1996) ó por la acción de los contaminantes sobre los organismos. 2.3.1.3 Lipoperoxidación celular La lipoperoxidación es uno de los principales efectos celulares causados por la generación de radicales libres (Fig. 4). Es un proceso degenerativo que actúa sobre los ácidos grasos poliinsaturados de las membranas celulares y cuya consecuencia más importante es la destrucción paulatina de éstas. Dicha ruptura genera una variedad de compuestos (alcoholes, cetonas, aldehídos y éteres) entre los que se encuentra el malondialdehído (MAD) (Buege y Aust, 1978). Existen defensas antioxidantes inespecíficas como las vitaminas A, C y E, ácido úrico y ceruloplasmina y específicas o enzimáticas como la superóxido-dismutasa (SOD), catalasa (CAT) y las enzimas dependientes del glutatión glutatión peroxidasa (GPx) y glutatión reductasa (GR)) para prevenir este proceso (Comporti, 1993). 2.3.1.4 Generación de hidroperóxidos lipídicos Los ácidos grasos poliinsaturados de las membranas biológicas son particularmente susceptibles de ser atacados por radicales libres. En la fase de iniciación del proceso de lipoperoxidación, la sustracción de un átomo de hidrógeno por el radical hidroxilo genera un radical lípidico con un carbono centrado (dieno conjugado); éste reacciona con el oxígeno molecular (O2) para formar un radical peroxilo (radical lípido peróxido), dando lugar a hidroperóxidos lipídicos (Fig 5) (Halliwell, 1992). Los radicales peroxilo pueden sustraer un átomo de hidrógeno de un ácido graso adyacente, comenzando una serie de reacciones autocatalíticas en cadena (fase de propagación) que convierten a muchos lípidos de membrana en hidroperóxidos lipídicos. La presencia de estos daños altera su función al modificar su fluidez y permitiendo el paso de iones como el Ca++ a través de la misma, lo cual activa a las fosfolipasas y eventualmente provoca su ruptura. Además, los hidroperóxidos lipídicos pueden atacar y dañar proteínas de membrana (Halliwell, 1992). Finalmente, los hidroperóxidos lipídicos pueden ser convertidos en una serie de productos de bajo peso molecular (alcanos, alquenos, derivados hidroxi ó epoxi, cetonas y polihidroxiperóxidos), los cuales por sí mismos pueden ser tóxicos para la célula (Fuller y col., 1988). A este proceso se le denomina lipoperoxidación.

14
Fig. 4. Peroxidación de lípidos (Buege y Aust, 1973).

15
Fig 5. Generación de hidroperóxidos lipídicos (basado en Halliwell y Gutteridge, 1989)

16
2.3.1.5 Oxidación de proteínas Las proteínas realizan una gran cantidad de funciones celulares, tales como la recepción y trasmisión de señales, el transporte de iones, las respuestas a condiciones de estrés, el metabolismo energético, la duplicación y reparación del DNA, la traducción y la transcripción. Cuando el O2- y el H2O2 se unen a residuos de metionina ó cisteína, pueden activarlas ó inactivarlas (Hansberg-Torres, 2002). El O2- también puede reaccionar con los aminoácidos triptofano, tirosina, histidina y lisina (Davies y Dean, 1997). El grado de oxidación en las proteínas se puede medir cuantificando el número de carbonilos, los cuales se forman por la oxidación de de los aminoácidos prolina y arginina, rupturas en la cadena peptídica (Hansberg-Torres, 2002) ó la reacción de los aldehídos (producto de la peroxidación de lípidos y la oxidación de azúcares) con las proteínas (Hermes-Lima, 2006). Cuando esta modificación es irreversible (Fig. 6), un cambio en la estructura de la proteína puede llevar a un daño funcional, alterando los procesos en los cuales interviene (Repetto, 1997). La mayoría de los daños producidos por el O2- son de este tipo y en términos generales marcan a las proteínas para su degradación proteolítica (Hansberg-Torres, 2000).
Fig. 6. Formación del sulfóxido de metionina (reversible) y de la sulfona de metionina(irreversible) (Hansberg-Torres, 2002). 2.3.1.6 Generación de formaldehído Como se ha mencionado, el radical hidroxilo (OH.) es el radical del oxígeno más reactivo que se conoce (Halliwell y Gutteridge, 1989; Kruk, 1998) y que debido a su reactividad, puede sustraer hidrógenos, p. ej. de los dobles enlaces de los ácidos grasos insaturados de las membranas biológicas (Halliwell y Gutteridge, 1989). La lipoperoxidación genera una variedad de compuestos, entre los que se puede encontrar alcoholes (Buege y Aust, 1978) (Fig. 7). Estos alcoholes pueden ser oxidados a sus correspondientes aldehídos (Dorfman y Adams, 1973) por las enzimas contenidas en los microsomas hepáticos (Cederbaum y Cohen, 1980) ó por las catalasas, en el caso de alcoholes de bajo peso molecular, tales como metanol y etanol (Halliwell y Gutteridge, 1989). Si el alcohol

17
oxidado es el metanol (CH3OH), el aldehído producido es el metanal (HCHO) ó formaldehído.
Fig. 7. Formación de alcoholes como resultado de la peroxidación de lípidos (modificado de Repetto, 1997).

18
2.4 Biotransformación de los xenobióticos Una vez que han ingresado al organismo, las substancias sufren una serie de reacciones químicas que cambian sus propiedades químicas y sus efectos toxicológicos. Dichas reacciones, modifican la polaridad del compuesto, afectando así la forma en que es distribuido y facilitando su eliminación del organismo (detoxificación). Sin embargo, también pueden producirse metabolitos con mayor toxicidad que la del compuesto original, a este proceso se le denomina bioactivación (Alvares y Pratt, 1990). Las reacciones de biotransformación pueden dividirse en dos clases: las de fase I y las de fase II. En las primeras, la finalidad es incrementar la polaridad del compuesto al oxidarlo, reducirlo ó romper enlaces éster ó amida. En las reacciones de fase II ó de conjugación, el compuesto original ó uno de sus metabolitos es acoplado a substratos endógenos tales como el ácido glucurónico, ácido acético ó ácido sulfúrico (Alvares y Pratt, 1990). 2.4.1 Sistema de oxidasas de función mixta (MFO) Comprende una superfamilia de enzimas mono-oxigenasas que catalizan la incorporación de oxígeno al sustrato. Se denominan oxidasas de función mixta (MFO, por sus siglas en inglés) debido a que del oxígeno molecular se reduce un átomo de oxígeno hacia agua, como lo hacen las oxidasas, mientras el otro es adicionado al sustrato (Turpein, 2006) dando como resultado alcoholes y epóxidos (Repetto, 1997). Estas enzimas contienen un átomo de Fe y para realizar su función requieren de oxígeno, un agente reductor derivado del NADPH y un transporte facilitado a través del citocromo P450 (CYP) (Martin y col., 1985) (Fig. 8).
Fig 8. Ciclo catalítico del citocromo P450 (Turpein, 2006).

19
La superfamilia de los CYP´s con base en su homología ha sido dividida en familias (enzimas con más de 40% de la secuencia de aminoácidos idéntica) y éstas a su vez, en subfamilias (si más del 55% de la secuencia de animoácidos se repite). En el hombre, actualmente se han descrito hay 18 familias de CYP´s y 42 subfamilias, las cuales abarcan 57 genes que codifican para dichos CYP´s y 58 pseudogenes. Las enzimas de las familias 1-3 realizan la mayoría de la biotransformación de los xenobióticos, mientras el resto está involucrado en funciones fisiológicas como las biosíntesis de ácidos grasos, hormonas esteroides y ácidos biliares (Turpein, 2006). De todos los sistemas enzimáticos involucrados en las reacciones de biotransformación, este trabajo estará enfocado al CYP 2E1 y la GSTT por su importancia en la biotransformación y/o biactivación de los PCB’s y de los HAP’s. 2.4.1.1 Citocromo 2E1 (CYP 2E1) Esta subfamilia está codificada por un solo gen. Se encuentra en cantidades relativamente altas en el hígado (2-4%). Los sustratos para este CYP usualmente son compuestos hidrofóbicos ó de bajo peso molecular y para su estudio se han empleado clorzoxazona, acetominofén, enflurano y halotano (Turpein, 2006). El CYP 2E1 puede ser inducido por varios de sus sustratos (tales como el etanol, acetona y pirazol); por el contrario, puede ser inhibido por la piridina y el disulfiram (Turpein, 2006). 2.4.2 Glutatión S-transferasas Consisten en una familia de isoenzimas encargadas de la conjugación de los xenobióticos y los aldehídos resultado de la peroxidación de lípidos con el glutatión (GSH) (Hermes-Lima, 2004). Este proceso puede ocurrir con un gran número de xenobióticos y compuestos endógenos electrofílicos (Jakoby, 1990) y constituye el primer paso en la vía de la formación del ácido mercaptúrico (Commandeur y col., 1995). Las GST´s están ampliamente distribuidas en todos los organismos incluyendo bacterias, hongos, plantas y animales, ya sea en el citoplasma ó el núcleo (Hermes-Lima, 2004). La conjugación de dichos compuestos electrofílicos con el glutatión, previene que estos reaccionen con los centros nucleofílicos de moléculas importantes para la célula, tales como las proteínas y el DNA (Commandeur y col., 1995). Además, las GST´s son importantes para la síntesis de prostaglandinas y leucotrienos (Hermes-Lima, 2004).

20
Por otra parte, se ha visto que las glutatión S-transferasas poseen actividad tipo glutatión peroxidasa independiente de selenio (denotada como GST-Px), por lo que pueden catalizar peróxidos orgánicos, tales como los hidroperóxidos lipídicos, pero no los peróxidos inorgánicos (p. ej. el H2O2). Esta capacidad está distribuida de manera diferencial entre las diferentes clases de GST´s, siendo más alta para la GSTθ (ó GST T), moderada para la GSTα y baja para las otras isoformas, y se ha visto que juega un papel importante en la detoxificación de hidroperóxidos orgánicos en mamíferos (Hermes-Lima, 2006). En los vertebrados, se han identificado 6 clases de GST´s, 4 en la fracción citosólica [alpha (α), mu (µ), pi (π) y tetha (θ)] (Mannervik y col., 1992) y 2 en la fracción microsomal (sigma y kappa). Las diferencias de las isoformas están basadas en las secuencias del amino terminal, especificidad del sustrato, sensibilidad a los inhibidores, punto isoeléctrico y análisis inmunológicos (Hermes-Lima, 2006). Dentro de la fracción citoplasmática se han encontrado dos tipos de la isoforma tetha (θ), la GST T1-1 (Meyer y col., 1991) y la GST T2-2 (Hussey y Hayes, 1992). Con esta nomenclatura, GST T1-1 se refiere a una isoenzima GST de la clase tetha, formada por dos subunidades (es decir, un dímero) del tipo T1.

21
2.5 ZONA DE ESTUDIO La Laguna de Zumpango (Fig. 9) se encuentra situada al noreste del Estado de México, en el Municipio de Zumpango de Ocampo (INEGI, 2000).
Fig. 9. Laguna de Zumpango, Estado de México. URL4 (2008) Google Earth, Image© Digital Globe.
Forma parte de los antiguos lagos de la Cuenca del Valle de México (Texcoco, Chalco, Zumpango y Xochimilco) (Fig. 10) y por sus dimensiones (1,865 Ha de superficie y 18 km de longitud en su bordo), se considera uno de los cuerpos de agua más importantes del Estado de México (INEGI, 2001) y el más grande de la ex zona lacustre del Valle de México. Hasta 1789 fue una cuenca cerrada y de gran extensión, posteriormente el espejo de agua se redujo tanto por la utilización del agua de sus tributarios y de la tierra para uso agrícola. A principios del siglo XX se comenzaron a drenar su aguas hacia el Río Tula a través del Tajo de Nochistongo para tratar de evitar las inundaciones que afectaban a la Ciudad de México, lo cual redujo considerablemente su superficie (INEGI, 2001).

22
Fig. 10. Zona lacustre del Valle de México, indicando la ubicación actual de las zonas que circundaban a este cuerpo de agua (INEGI, 2001).

23
Actualmente, este embalse se halla incluido dentro de las Regiones Hidrológicas Prioritarias de la CONABIO (Comisión Nacional para Conocimiento y Uso de la Biodiversidad) (Arriaga-Cabrera y col., 1998), es una de las Reservas Naturales Protegidas bajo el control del Estado de México y fue declarado “Santuario del Agua” en 2003 (INEGI, 2008). 2.6 Generalidades sobre el charal de Santiago (Chirostoma riojai) Chirostoma riojai (Fig. 11) es una especie de pez zooplanctófaga endémica del Antiplano Mexicano, de importancia económica para las poblaciones que habitan las riberas de los lagos del curso alto del sistema Lerma-Santiago (Figueroa-Lucero y col., 2003).
Fig. 11. Ejemplares de Chirostoma riojai (Laboratorio de Toxicología Ambiental, ENCB-IPN).
Esta zona se caracteriza por ser un gran centro ictiológico de endemismos del país, con 58% de especies endémicas (Miller y Smith, 1986). Sin embargo, desde hace tiempo se han desarrollado importantes núcleos humanos en la región. A lo largo de su cauce, actualmente se presenta un alto grado de urbanización e industrialización afectando la calidad del aire, agua, suelo y comunidades naturales que ahí habitan (Figueroa-Lucero y col., 2003). El charal de Santiago Chirostoma riojai se describió por primera vez en 1965. Sin embargo, en 1997 sólo se registró su presencia en 9 localidades del Alto Lerma y en la actualidad sólo se le reporta en dos localidades: la Laguna de Santiago Tilapa que fue donde se describió el holotipo y el embalse Ignacio Ramírez (Figueroa-Lucero y col., 2003), por lo que actualmente se halla incluida en las especies en peligro de extinción (NOM-059-SEMARNAT-2001). 2.7 Bioactivación y conjugación de PCB´s y HAP’s en organismos acuáticos Existen trabajos previos realizados en organismos acuáticos y animales de laboratorio donde se ha estudiado el papel de la GSTT y el CYP 2E1 en la biotransformación y bioactivación de varios compuestos orgánicos.

24
Por ejemplo, en animales de laboratorio, se ha investigado el papel del CYP 2E1 en la biotransformación del etanol, la biactivación de varios solventes industriales y como generador de radicales libres (Turpein, 2006) y la conjugación de epóxidos e hidroperóxidos a través de la GSTT (Blocki y col., 1994). En relación con los organismos acuáticos, Wall y Crivello (1998) reportaron por primera vez la actividad del CYP 2E1 en peces teleósteos y Geter y col. (2003) observaron la inducción de este CYP en peces Oryzias latipes expuestos a compuestos clorados y compuestos generadores de radicales libres (etanol y acetona). Por otro lado, en referencia al estrés oxidativo en organismos acuáticos, se pueden citar los siguientes trabajos: Thomas y Wofford (1993), expusieron al pez Micropogonias undulatus a Aroclor 1254, observando un incremento en los niveles de malondialdehído en el hígado (in vivo e in vitro) y ovario (in vitro) de los peces. En otra investigación, en la cual se expuso al bagre de canal Ictalurus punctatus a sedimentos contaminados con HAP´s, se encontró que a todos los periodos de tiempo evaluados, los niveles hepáticos de malondialdehído fueron significativamente mayores que en los controles (Di Giulio y col., 1993). En relación con la bioacumulación de compuestos orgánicos halogenados (PCB´s), Otto y Moon (1996) evaluaron la acumulación de estos compuestos en el músculo de peces Ameriurus nebulosus colectados en un cuerpo de agua contaminado con estas substancias, hallando un incremento en los niveles de PCB´s de 22 veces con respecto a los detectados en peces provenientes de un sitio no contaminado utilizado como referencia.

25
3. JUSTIFICACIÓN La presencia en el ambiente de compuestos aromáticos halogenados (PCB´s) e hidrocarburos aromáticos policíclicos (HAP´s) puede representar un riesgo potencial para la salud y los ecosistemas acuáticos. En el caso del charal de Santiago (Chirostoma riojai) además de ser una especie endémica está en peligro de extinción. Aunque no existen reportes que argumenten las causas de su desaparición, la contaminación de los cuerpos de agua podría ser un factor importante limitante para su distribución, además de acelerar la reducción de sus poblaciones naturales. Se sabe que los HAP´s y PCB´s, ambos grupos de contaminantes ubicuos en los ecosistemas acuáticos, requieren de la bioactivación para llevar a cabo sus efectos tóxicos y además pueden acumularse en los organismos generando radicales libres y causando una serie de daños celulares como la peroxidación de lípidos y la oxidación de proteínas. Por lo anterior, son necesarios numerosos estudios donde sobresale evaluar la biactivación de compuestos aromáticos halogenados (PCB´s y HAP´s) a través de las enzimas CYP 2E1 y GSTT en el charal de Santiago C. riojai, especie endémica de nuestro país. Los resultados obtenidos permitirán por un lado diagnosticar el estado de salud de esta población del charal de Santiago habitante de la Laguna de Zumpango y por otro lado, proporcionarán información sobre los niveles de contaminación de este cuerpo de agua para el establecimiento de medidas preventivas y/o correctivas para la protección de esta especie en peligro de extinción.
4. HIPÓTESIS
Si la Laguna de Zumpango, Estado de México está contaminada con compuestos orgánicos halogenados como los PCB´s y HAP´s, entonces sus efectos tóxicos podrán ser evaluados a través de biomarcadores de biactivación y de estrés oxidativo en el charal de Santiago (C. riojai) habitante de este cuerpo de agua y será posible encontrar relaciones entre la respuesta biológica y los niveles de estos contaminantes presentes en los diferentes compartimentos ambientales (agua y sedimento).

26
5. OBJETIVOS
5.1 OBJETIVO GENERAL Evaluar la biactivación de compuestos orgánicos halogenados (PCB´s y HAP´s) en el charal de Santiago Chirostoma riojai, colectado en la Laguna de Zumpango, Estado de México, durante dos periodos bien diferenciados del año: sequía fría (diciembre 2008) y lluvias (julio 2009).
5.2 OBJETIVOS ESPECÍFICOS
1) Evaluar los niveles de hidrocarburos aromáticos policíclicos (HAP´s) y de los bifenilos policlorados (PCB´s) presentes en el agua, sedimento y peces de la Laguna de Zumpango, Estado de México.
2) Determinar el factor de bioconcentración (FBC) de los compuestos mencionados
en el hígado y víscera de los peces.
3) Medir la actividad de los fuerzas endógenas pro-oxidantes como el CYP 2E1 y la glutatión S-transferasa isoforma tetha en el charal de Santiago (Chirostoma riojai) expuesto mezclas ambientales complejas y sus relaciones con el estrés oxidativo.
4) Cuantificar la concentración de especies reactivas del oxigeno (peróxido de hidrógeno y anión superóxido) producidos de manera endógena y sus relaciones con el estrés oxidativo.
5) Valorar el estrés oxidativo en el hígado y vísceras de los peces, utilizando como biomarcadores los niveles de lipoperoxidación, hidroperóxidos lipídicos y oxidación de proteínas.

27
6. METODOLOGÍA
Fig. 12. Diagrama de flujo de las determinaciones realizadas en laboratorio y campo.

28
6.1 Determinaciones realizadas en el laboratorio 6.1.1 Proteínas totales A pH ácido (3.9), el verde de bromocresol se une a las proteínas, dando como resultado un cambio de coloración de verde hacia azul. De esta manera, la cantidad de proteínas presentes en la muestra es proporcional a la absorbancia, la cual es evaluada a 625 nm y determinada por interpolación en una curva patrón de albúmina.
6.1.2 Biomarcadores de bioactivación 6.1.2.1 Actividad del CYP 2E1 (Cannady y col., 2003) La hidroxilación del p-nitrofenol por el CYP 2E1 es evaluada a pH débilmente ácido (6.8) en presencia de NADH. La actividad se evalúa mediante la formación de p-nitrocatecol a 520 nm y se expresa en relación a la cantidad de proteínas totales contenidas en la muestra, como mM/ min/mg proteína. El coeficiente de extinción molar para el p-nitrocatecol es de 91 mM cm-1. 6.1.2.2 Actividad de la glutatión S-transferasa isoforma tetha (Warholm y col., 1994) En esta determinación se cuantifica la producción de formaldehído en presencia de GSH, usando como sustrato al diclorometano con reactivo de Nash (86 uL de ácido acético, 62 uL de acetil acetona y 4.6 g de acetato de amonio/ 10 mL). La actividad de la enzima se evalúa a 414 nm, a pH de 7.4 y se expresa como ug formaldehído/ min/mg proteína usando una curva estándar de formaldehído.
6.1.3 Biomarcadores de estrés oxidativo 6.1.3.1 Anión superóxido (Schlezinger y col., 1999) El dihidroetidio es oxidado por el anión superóxido en presencia de NADH. El producto oxidado se evalúa espectrofluorométricamente a 530 nm de emisión y 624 nm de excitación. La concentración de anión superóxido se estima mediante un coeficiente de extinción molar de 7,000 M cm-1. 6.1.3.2 Peróxido de hidrógeno (Schlezinger y col., 1999) Esta técnica se basa en la oxidación del diacetato de diclorodihidrofluoresceína que realiza el H2O2. El producto oxidado emite fluorescencia a 485 nm de excitación y 525 nm de emisión en presencia de NADH. Para el cálculo se usa una curva patrón de H2O2.

29
6.1.3.3 Formaldehído (basado en Warholm y col., 1994) La cuantificación de este compuesto se realiza a 414 nm, por medio de la adición del reactivo de Nash como se detalló previamente.
6.1.4 Biomarcadores de daño
6.1.4.1 Substancias reactivas al ácido tiobarbitúrico (TBAR´s) (Buege y Aust, 1978) El malondialdehído (MDA) es uno de los productos del proceso de lipoperoxidación. En esta técnica, se hace reaccionar este compuesto con el ácido tiobarbitúrico (TBA), dando como resultado un complejo color rosa-anaranjado, el cual es evaluado a 535 nm usando un coeficiente de extinción molar de 1.56 x 105 M cm-1 y se expresan como sustancias reactivas al ácido tiobarbitúrico. 6.1.4.2 Hidroperóxidos lipídicos (Furtado-Filho y col., 2007) En esta reacción, los hidroperóxidos lipídicos son oxidados a pH ácido por un complejo ferroso-xilenol naranja. El complejo resultante Fe3+-xilenol naranja es evaluado a 530 nm mediante una curva de calibración de hidroperoxido cumeno (Sigma Aldrich) de 0.1 a 0.5 mM. 6.1.4.3 Oxidación de proteínas (formación de carbonilos) (Ansaldo y col., 2007) Los productos más importantes del ataque de los radicales libres sobre las proteínas son los derivados carbonilo, los cuales son evaluados por medio de su derivatización con 2,4-dinitrofenil-hidrazina (DNPH), seguida de precipitación ácida y su posterior lectura a 366 nm. La concentración de derivados carbonilo se estima mediante el coeficiente de extinción molar de 0.022 mM cm-1
6.1.5 Determinaciones ambientales 6.1.5.1 Materia orgánica total en sedimentos y agua (APHA, 1992) 6.1.5.1.1 Materia orgánica total en sedimentos En esta técnica, la materia orgánica (m. o.) presente en las muestras es oxidada con K2Cr2O7 y H2SO4. A continuación, se titulan con sulfato ferroso en presencia de un indicador (ferroína), hasta lograr un vire de color de esmeralda a canela-rojizo.

30
Finalmente, la m. o. es cuantificada al comparar los volúmenes de titulación de las muestras con la del testigo y normalizada utilizando el peso seco del sedimento. 6.1.5.1.2 Materia orgánica total en aguas Para cuantificar la m. o., se hace pasar un volumen determinado de agua a través de un filtro de fibra de vidrio de 0.45 micras, el cual es colocado en un matraz Erlenmeyer. El resto de la determinación es igual al empleado para las muestras de sedimento. 6.1.5.1.3 Materia orgánica hidrosoluble (APHA, 1992) 6.1.5.3.1 Materia orgánica hidrosoluble en sedimentos En esta técnica, se agrega un volumen de agua a las muestras de sedimento, se agita en Vórtex y se centrifuga. A continuación, el sobrenadante es filtrado, acidificado con HCl y leído a 275 nm. El carbono orgánico hidrosoluble es cuantificado por interpolación en una curva patrón preparada con biftalato de potasio y normalizada respecto al peso seco del sedimento. 6.1.5.3.2 Materia orgánica hidrosoluble en aguas Se toma un volumen determinado de muestra, la cual es filtrada. En seguida, sobre el filtrado se repite el mismo procedimiento que en inciso anterior. 6.1.5.2 Clorofilas (Axler y Owen, 1994) La cuantificación de los pigmentos fotosintéticos se realiza espectrofotométricamente, evaluando a distintas longitudes de onda (430 nm para clorofila “a”, 453 nm para clorofila “b” y 665 nm para feofitinas) la cantidad de pigmentos contenidos en un volumen determinado de muestra, los cuales han sido extraídos previamente bajo condiciones alcalinas (acetona al 90% a pH 10.0), e interpolando posteriormente las absorbancias leídas en ecuaciones preestablecidas con estándares de clorofila “a” y “b” (Sigma Aldrich).

31
6.1.5.3 HAP´s en agua, sedimento y tejidos (Livingstone y col., 1993) 6.1.5.3.1 HAP´s en sedimentos Las muestras de sedimento son pesadas y se realiza una extracción con diclorometano agitando en vortex por 5 y centrifugadas a 3,500 rpm/15 min. En seguida, el sobrenadante obtenido es filtrado 3 veces, utilizando lana de fibra de vidrio y colectado en frascos color ámbar, los cuales son etiquetados y mantenidos en congelación hasta el momento de las determinaciones. La evaluación de los HAP´s se hace por espectroscopía de fluorescencia, utilizando las siguientes longitudes de onda (Tabla 7) e interpolando en una curva patrón. Tabla 7. Longitudes de onda (λ) para determinar los HAP´s considerados en este estudio (Livingstone y col., 1993)
Compuesto λ Excitación (nm) λ Emisión (nm)
naftaleno 273 360
pireno 341 380
benzo[a]pireno 380 430
6.1.5.3.2 HAP´s en aguas por extracción líquido-líquido Utilizando un embudo de separación, se toma un volumen de agua y se adiciona diclorometano. A continuación, se agita y se deja reposar hasta que se separen las fases. Enseguida, la fase conteniendo el disolvente se filtra tres veces en lana de fibra de vidrio y colectada en frascos color ámbar. El resto del procedimiento es igual que en el inciso anterior. 6.1.5.3.3 HAP´s en tejidos Los charales son pesados, homogeneizados y adicionados con diclorometano. Luego de una agitación en vortex y centrifugación, la fase conteniendo al disolvente es separada, filtrada tres veces con lana de fibra de vidrio y colectada en frascos color ámbar, repitiendo los mismos pasos que en los dos incisos anteriores. 6.1.5.4 PCB´s en sedimentos, agua y tejidos (Vega-López y col., 2008) 6.1.5.4.1 PCB´s en sedimentos y aguas La extracción se realiza de manera similar que en el caso de los HAP´s, con la diferencia de que se utiliza metanol grado HPLC como disolvente. Posteriormente, la fase

32
conteniendo metanol es separada y filtrada 3 veces en lana de vidrio y colectada en frascos color ámbar. En seguida, se realiza una extracción en fase sólida, haciendo pasar el filtrado a través de un cartucho C-18 previamente activado. Los compuestos orgánicos adsorbidos en la fase estacionaria son eluídos con metanol HPLC y colectados en frascos color ámbar. Finalmente, los PCB´s son determinados por CG, utilizando un detector de captura de electrones, blancos de reactivos y estándares con las concentraciones apropiadas. PCB´s en aguas, extracción líquido-líquido Nuevamente, el procedimiento de extracción es muy similar a la efectuado con los HAP´s, sólo que el disolvente utilizado es metanol grado HPLC. Con el filtrado obtenido, se repiten los mismos pasos que en el inciso anterior. 6.1.5.4.2 PCB´s en tejidos Al igual que en los dos incisos previos, para la extracción se procede como en el caso de los HAP´s, pero utilizando metanol grado HPLC. Posteriormente, las proteínas contenidas en el filtrado, son separadas por precipitación ácida, por la adición de H2SO4 al 0.2% y agitación en el vortex, seguida de centrifugación (modificado de Flanagan y col., 2006) (Apéndice 1). Una vez hecho lo anterior, el filtrado es analizado por CG bajo las mismas condiciones que en el inciso anterior.
6.2 Determinaciones ambientales
(realizadas en campo) En las muestras de agua y sedimento: pH (NMX-AA-008-SCFI-2000), temperatura (NMX-AA-007-SCFI-2000) y potencial de óxidoreducción (NMX-AA-008-SCFI-2000). En las estaciones de muestreo: transparencia por el disco de Secchi y profundidad.

33
7. RESULTADOS PRIMER MUESTREO (sequía fría) 7.1.1 Contenido de HAP´s en agua y sedimentos El naftaleno fue el más abundante en agua y tuvo una distribución localizada en el embarcadero al nivel superficial y al fondo de la isla, estación cercana al afluente de la laguna (Fig. 13).
Fig. 13. Contenido de HAP´s en el agua colectada en las estaciones a diferentes profundidades durante la sequía fría ( I-S: isla-superficie; I-Nc: isla-nivel de compensación; I-F: isla-fondo; C-S: centro-superficie; C-Nc: centro-nivel de compensación; C-F: centro-fondo; E-S: embarcadero-superficie; E-Nc: embarcadero-nivel de compensación; E-F: embarcadero-fondo). En los sedimentos, el embarcadero fue el sitio más contaminado (Fig. 14). Respecto a los compuestos el más abundante también fue el naftaleno; sin embargo, el benzo[a]pireno se detectó en todos los sitios a diferencia de los otros HAP’s.

34
Fig. 14. Contenido de HAP´s evaluado en los sedimentos durante la sequía fría.
7.1.2 Contenido de HAP´s en hígado y vísceras de peces En contraste con los compartimentos ambientales, en los peces se detectó pireno, pero no naftaleno. La concentración de pireno tuvo una relación inversa con la talla de los organismos, más que entre tejidos (Fig. 15). También se detectó benzo[a]pireno pero en este caso si se observaron diferencias entre tejidos. Su concentración fue mayor en vísceras y además presentó relación inversa con la talla (Fig. 16).
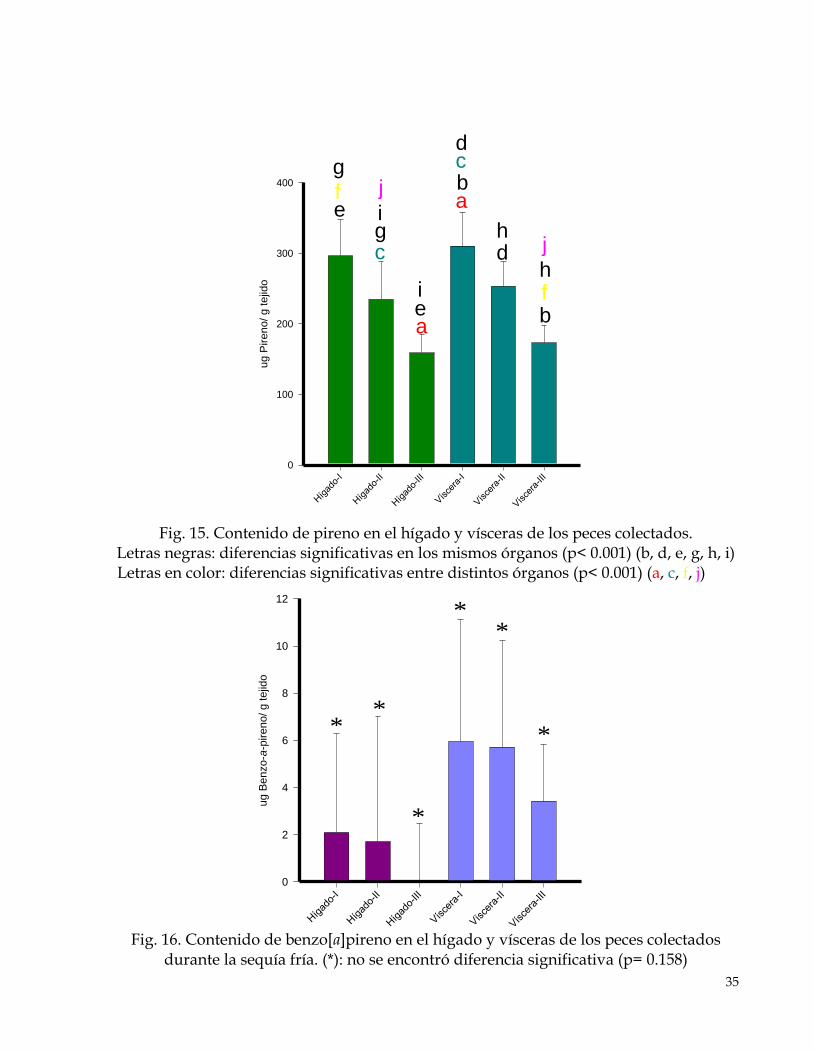
35
Híg
ado-
I
Híg
ado-
II
Híg
ado-
III
Vísce
ra-I
Vísce
ra-II
Vísce
ra-II
I
ug
Pire
no
/ g
te
jido
0
100
200
300
400
Híg
ado-
I
Híg
ado-
II
Híg
ado-
III
Vísce
ra-I
Vísce
ra-II
Vísce
ra-II
I
ug B
enzo-a
-pireno/ g tejid
o
0
2
4
6
8
10
12
Fig. 15. Contenido de pireno en el hígado y vísceras de los peces colectados. Letras negras: diferencias significativas en los mismos órganos (p< 0.001) (b, d, e, g, h, i) Letras en color: diferencias significativas entre distintos órganos (p< 0.001) (a, c, f, j)
Fig. 16. Contenido de benzo[a]pireno en el hígado y vísceras de los peces colectados
durante la sequía fría. (*): no se encontró diferencia significativa (p= 0.158)
a
c
a b c d
e
e f g
g
i
i
j
b f
j h
d h
* *
*
* *
*

36
7.1.3 Factor de bioconcentración (FBC) De los HAP’s en estudio, el pireno mostró un elevado factor de bioconcetración tanto en vísceras como en hígado (Fig. 17).
Fig. 17. Factor de bioconcentración de los HAP´s en hígado y vísceras de los charales
colectados durante la sequía fría.
7.1.4 Biomarcadores de estrés oxidativo, radicales libres El contenido de radicales libres presentó una diferencia entre los diferentes tejidos. El peróxido de hidrógeno se detectó en todas las tallas, tanto en hígado como de vísceras (Fig. 18). En el caso anión superoxido, su cantidad fue mayor en hígado y en vísceras fue mínima (Fig. 19). La concentración de formaldehido fue mayor en hígado que en vísceras (Fig. 20). También se observó una relación inversa con la talla en el caso del órgano hepático.

37
Fig. 18. Niveles de peróxido de hidrógeno (H2O2) en el hígado y víscera de los charales colectados durante la sequía fría. (*) no se encontró diferencia significativa (p= 0.326).
Híg
ado-
I
Híg
ado-
II
Híg
ado-
III
Vísce
ra-I
Vísce
ra-II
Vísce
ra-II
I
mM
/ g
tejid
o
0
200
400
600
800
1000
1200
1400
1600
1800
Fig. 19. Niveles de superóxido (O2-) en el hígado y víscera de los charales colectados
durante la sequía fría. (*): no se encontró diferencia significativa (p= 0.012).
Híg
ado-I
Híg
ado-II
Híg
ado-II
I
Vísce
ra-I
Vísce
ra-II
Vísce
ra-II
I
mM
/ g
te
jid
o
0
50
100
150
200
250
* * *
*
*
*
*
*
* * * *

38
Fig. 20. Niveles de formaldehído en el hígado y víscera de los charales colectados
durante la sequía fría. (a, b): diferencia significativa (p< 0.001). 7.1.5 Biomarcadores de daño No se observaron diferencias significativas en la lipoperoxidación de ninguno de los órganos bajo estudio (Fig. 21). La cantidad de hidroperóxidos lipídicos, que es un estadio intermedio del daño, fue mayor en el hígado que en vísceras y además se halló una relación inversa con la talla (Fig. 22). La oxidación de las proteínas estuvo más elevada en el hígado que en vísceras (Fig. 23). En el caso del hígado hubo relación inversa con la talla de los organismos.
Híg
ado-
I
Híg
ado-
II
Híg
ado-
III
Vísce
ra-I
Vísce
ra-II
Vísce
ra-II
I
mM
form
ald
ehíd
o/ g tejid
o
0
100
200
300
400
a
a
b
b

39
Fig. 21. Substancias reactivas al ácido tiobarbitúrico (TBAR´s) en el hígado y víscera de los charales colectados durante la sequía fría. (*) no se encontró diferencia significativa
(p= 0.472).
Fig. 22. Hidroperóxidos lipídicos en el hígado y víscera de los charales colectados
durante la sequía fría. (a,b): diferencia significativa (p= 0.002).
Híg
ado-I
Híg
ado-II
Híg
ado-II
I
Vís
cera
-I
Vís
cera
-II
Vís
cera
-III
mM
/ g
tejid
o
0
20
40
60
80
100
* *
*
* * *
Híg
ado-I
Híg
ado-II
Híg
ado-II
I
Vísce
ra-I
Vísce
ra-II
Vísce
ra-II
I
mM
/ g
te
jid
o
0
20
40
60
80
a
a
b
b

40
Fig. 23. Oxidación de proteínas en el hígado y víscera de los charales colectados durante la sequía fría. (a,b,c) diferencia significativa(p< 0.001).
7.1.6 Biomarcadores de activación No se observaron diferencias significativas entre las tallas ni tejidos en la actividad del CYP2E1, pero en todos los casos hubo catálisis (Fig. 24). En contraste con este citocromo, la actividad de la GSTT fue estadísticamente significativa y mayor en el hígado que en vísceras (Fig. 25). También en el hígado hubo una relación inversa con la talla.
Híg
ado-I
Híg
ado-II
Híg
ado-II
I
Vísce
ra-I
Vísce
ra-II
Vísce
ra-II
I
mM
/ m
g p
rote
ína
/ g
te
jid
o
0
10
20
30
40
a
a
b
b c
c

41
Híg
ado-I
Híg
ado-II
Híg
ado-II
I
Vísce
ra-I
Vísce
ra-II
Vísce
ra-II
I
mM
/ g
pro
teín
a/ g
tejid
o
0
2
4
6
8
Fig. 24. Actividad del CYP2E1 en el hígado y víscera de los charales colectados durante la sequía fría. (*) no se encontró diferencia significativa (p= 0.214).
Fig. 25. Actividad de la GSTT en el hígado y víscera de los charales colectados durante la sequía fría. (a,b,c,d,e,f,g): diferencia significativa (p< 0.001).
* *
* *
*
*
Híg
ado-I
Híg
ado-II
Híg
ado-II
I
Vísce
ra-I
Vísce
ra-II
Vísce
ra-II
I
ug
fo
rma
lde
híd
o/
min
/ m
g p
rote
ína
/ g
te
jid
o
0
20
40
60
80
c a
a
b
b
c
d d
g
e e f
f
g

42
7.1.7. Análisis de componentes principales (ACP´s). 7.1.7.1 Biomarcadores vs. HAP´s. Hígado La actividad de la GSTT y la oxidación de proteínas estuvieron relacionadas con el benzo[a]pireno y con el pireno de manera directa. La lipoperoxidación y los hidroperóxidos lipídicos y el contenido de formaldehido tuvieron relación inversa con esos hidrocarburos (Fig. 26 y tabla 8).
Fig. 26. Gráfico bidimensional de puntuaciones sobre los componentes principales
en el hígado del charal colectado durante la sequía fría. GSTT: glutatión S-transferasa tetha; H2O2: peróxido de hidrógeno. HOOo: hidroperóxidos lipídicos; O2-: anión
superóxido.
-0.20
-0.10
0.00
0.10
0.20
0.30
0.40
0.50
0.02 0.04 0.06 0.08 0.10 0.12 0.14 0.16 0.18 0.20
GSTT
Benzo[a]pireno
Pireno
Oxidación de
proteínas H2O2
HOOo
Formaldehído CYP 2E1 O2
- Lipoperoxidación

43
Tabla 8. Matriz de correlaciones entre biomarcadores e HAP´s, evaluados en hígado.
H
2O
2
O2
-
Fo
rmal
de
hid
o
CY
P 2
E1
GS
TT
lip
op
ero
xid
ació
n
HO
Oo
oxid
aci
ón
d
e
pro
teín
as
pir
en
o
ben
zo
[a]p
ire
no
H2O2 1
O2- 0.150 1
formaldehído 0.381 0.921 1
CYP 2E1 0.343 0.447 0.543 1
GSTT 0.121 -0.099 -0.057 0.143 1
lipoperoxidación 0.332 0.919 0.927 0.487 -0.029 1
HOOo 0.389 0.917 0.952 0.588 0.003 0.940 1
oxidación de proteínas
0.156 0.715 0.761 0.319 0.163 0.751 0.752 1
Pireno 0.208 0.132 0.272 0.268 0.278 0.114 0.354 0.395 1
benzo[a]pireno 0.031 0.213 0.252 -0.082 0.299 0.159 0.323 0.485 0.742 1
Nota: las variables con correlación más alta entre aparecen resaltadas.
7.1.7.2 Biomarcadores vs. HAP´s. Víscera En víscera la concentración de benzo[a]pireno y pireno inducen la actividad del CYP2E1 y la producción del anión superóxido y de formaldehido, pero abaten la actividad de la GSTT. Mientras mayor fue la cantidad de estos hidrocarburos, se redujo la oxidación de proteínas, la lipoperoxidación y la concentración celular de peróxido de hidrógeno (Fig. 27 y tabla 9).

44
Fig. 27. Gráfico bidimensional de puntuaciones sobre los componentes principales
en víscera de charal colectado durante la sequía fría. H2O2: peróxido de hidrógeno; O2-: anión superóxido. GSTT: glutatión S-transferasa tetha.
Tabla 9. Matriz de correlaciones entre biomarcadores e HAP´s, evaluados en víscera.
H2O
2
O2
-
Fo
rmal
deh
ido
CY
P 2
E1
GS
TT
lip
op
ero
xid
ació
n
HO
Oo
oxid
aci
ón
d
e
pro
teín
as
pir
en
o
ben
zo
[a]p
iren
o
H2O2 1
O2- -0.075 1
formaldehído 0.036 -0.147 1
CYP 2E1 0.085 0.143 0.317 1
GSTT 0.055 -0.249 -0.326 -0.242 1
lipoperoxidación 0.717 0.078 0.339 0.052 -0.370 1
HOOo -0.017 -0.102 -0.043 -0.277 -0.226 0.090 1
oxidación de proteínas
-0.017 -0.102 -0.043 -0.277 -0.226 0.090 1.000 1
pireno -0.102 0.452 -0.019 -0.059 0.018 0.040 -0.251 -0.251 1
benzo[a]pireno -0.174 0.673 0.066 0.020 -0.131 0.112 0.082 0.082 0.633 1
Nota: las variables con correlación más alta entre aparecen resaltadas.
-0.40
-0.30
-0.20
-0.10
0.10
0.20
0.30
0.40
-0.40 -0.30 -0.20 -0.10 0 0.10 0.20 0.30 0.40
Oxidación de
proteínas
H2O2
Lipoperoxidación
Formaldehído
Benzo[a]pireno
O2-
Pireno
CYP 2E1
GSTT

45
7.1.7.3 Materia orgánica, clorofilas y feofitina vs. HAP´s. Agua En la sequía fría, la concentración de los hidrocarburos en agua se incrementó conforme los pigmentos fotosintéticos, las feofitinas y la materia orgánica de partículas. Y disminuyó con la cantidad de materia orgánica disuelta (Fig. 28 y tabla 10).
Fig. 28. Gráfico bidimensional de puntuaciones sobre los componentes principales en agua. MOD: materia orgánica disuelta; MOT: materia orgánica total; MOP: materia
orgánica particulada. Tabla 10. Matriz de correlaciones entre materia orgánica, pigmentos fotosintéticos e HAP´s, evaluados
en agua. MOT: materia orgánica total; MOD: materia orgánica disuelta; MOP: materia orgánica particulada.
MO
T
MO
D
MO
P
na
ftal
en
o
be
nz
o[a
]pir
en
o
clo
rofi
la a
clo
rofi
la b
feo
fiti
na
MOT 1
MOD 0.405 1
MOP -0.926 -0.028 1
naftaleno -0.500 -0.994 0.135 1
benzo[a]pireno 0.069 -0.884 -0.441 0.830 1
clorofila a -0.979 -0.212 0.983 0.315 -0.268 1
clorofila b -0.961 -0.135 0.994 0.240 -0.343 0.997 1
feofitina -0.989 -0.266 nafta 0.367 -0.214 0.998 0.991 1
Nota: las variables con correlación más alta entre aparecen resaltadas.
-1.00
-0.80
-0.60
-0.40
-0.20
0.20
0.40
0.60
0.80
1.00
-1.00 -0.80 -0.60 -0.40 -0.20 0 0.20 0.40 0.60 0.80 1.00
MOD
MOT
Benzo[a]pireno
Naftaleno
feofitina Clorofila a
Clorofila b MOP
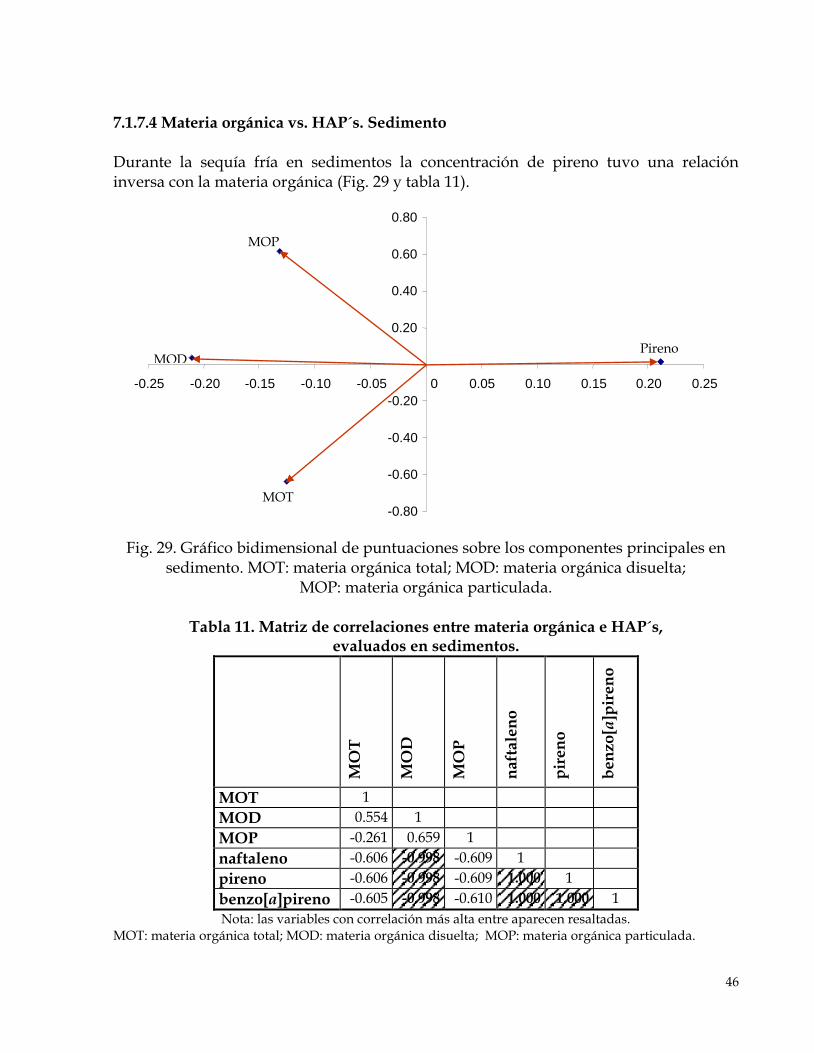
46
7.1.7.4 Materia orgánica vs. HAP´s. Sedimento Durante la sequía fría en sedimentos la concentración de pireno tuvo una relación inversa con la materia orgánica (Fig. 29 y tabla 11).
Fig. 29. Gráfico bidimensional de puntuaciones sobre los componentes principales en
sedimento. MOT: materia orgánica total; MOD: materia orgánica disuelta; MOP: materia orgánica particulada.
Tabla 11. Matriz de correlaciones entre materia orgánica e HAP´s,
evaluados en sedimentos.
MO
T
MO
D
MO
P
naft
ale
no
pir
en
o
ben
zo
[a]p
iren
o
MOT 1
MOD 0.554 1
MOP -0.261 0.659 1
naftaleno -0.606 -0.998 -0.609 1
pireno -0.606 -0.998 -0.609 1.000 1
benzo[a]pireno -0.605 -0.998 -0.610 1.000 1.000 1
Nota: las variables con correlación más alta entre aparecen resaltadas. MOT: materia orgánica total; MOD: materia orgánica disuelta; MOP: materia orgánica particulada.
-0.80
-0.60
-0.40
-0.20
0.20
0.40
0.60
0.80
-0.25 -0.20 -0.15 -0.10 -0.05 0 0.05 0.10 0.15 0.20 0.25
MOP
MOD
MOT
Pireno
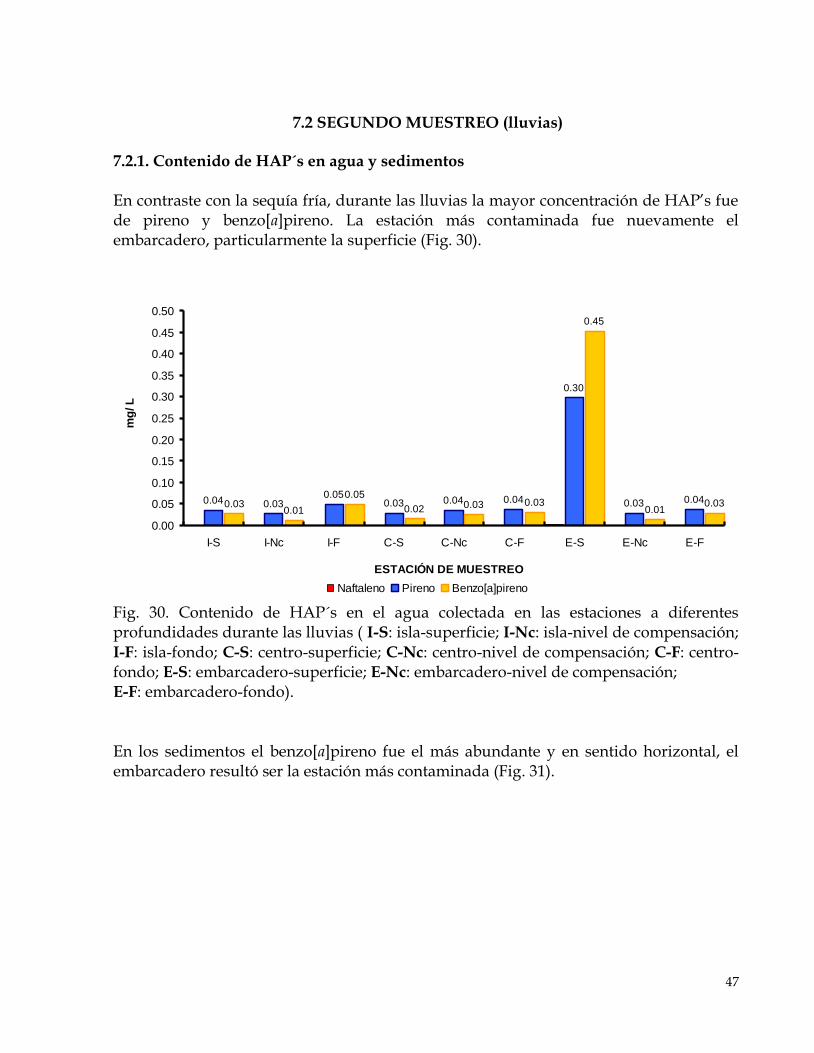
47
7.2 SEGUNDO MUESTREO (lluvias) 7.2.1. Contenido de HAP´s en agua y sedimentos En contraste con la sequía fría, durante las lluvias la mayor concentración de HAP’s fue de pireno y benzo[a]pireno. La estación más contaminada fue nuevamente el embarcadero, particularmente la superficie (Fig. 30).
0.04 0.030.05
0.03 0.04
0.30
0.03 0.040.030.01
0.05
0.02 0.03 0.03
0.45
0.010.030.04
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.40
0.45
0.50
I-S I-Nc I-F C-S C-Nc C-F E-S E-Nc E-F
ESTACIÓN DE MUESTREO
mg
/ L
Naftaleno Pireno Benzo[a]pireno
Fig. 30. Contenido de HAP´s en el agua colectada en las estaciones a diferentes profundidades durante las lluvias ( I-S: isla-superficie; I-Nc: isla-nivel de compensación; I-F: isla-fondo; C-S: centro-superficie; C-Nc: centro-nivel de compensación; C-F: centro-fondo; E-S: embarcadero-superficie; E-Nc: embarcadero-nivel de compensación; E-F: embarcadero-fondo). En los sedimentos el benzo[a]pireno fue el más abundante y en sentido horizontal, el embarcadero resultó ser la estación más contaminada (Fig. 31).
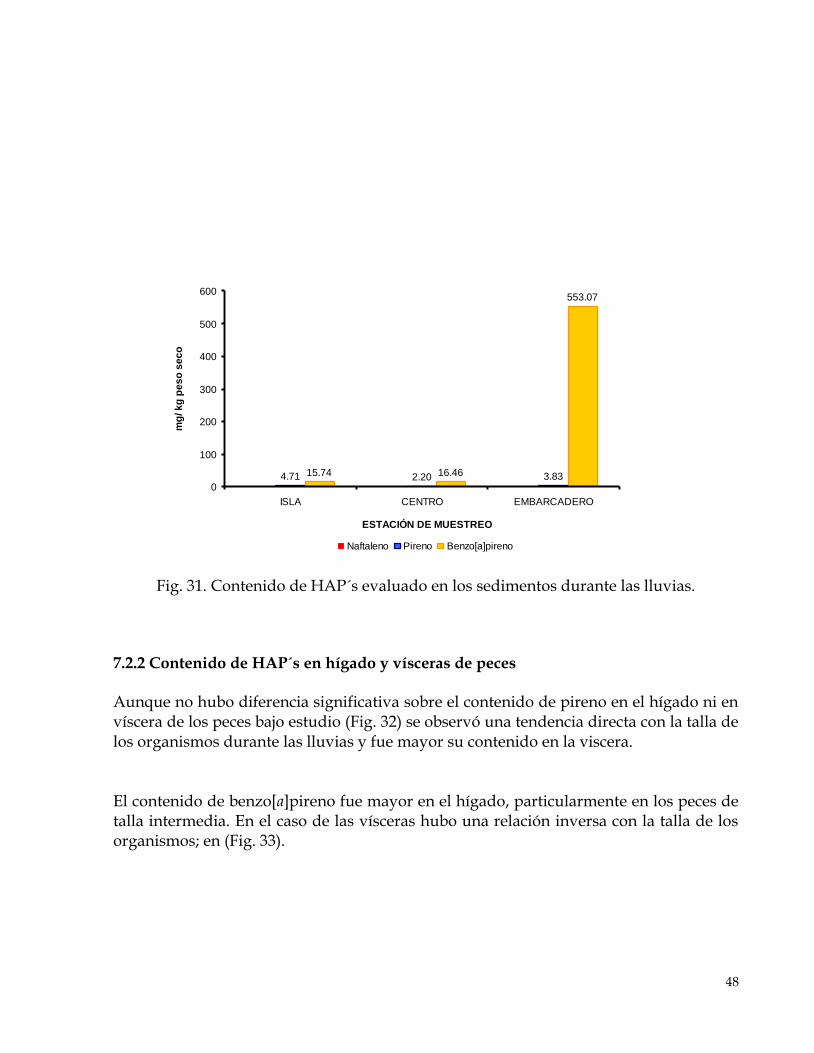
48
4.71 2.20 3.8315.74 16.46
553.07
0
100
200
300
400
500
600
ISLA CENTRO EMBARCADERO
ESTACIÓN DE MUESTREO
mg
/ k
g p
es
o s
ec
o
Naftaleno Pireno Benzo[a]pireno
Fig. 31. Contenido de HAP´s evaluado en los sedimentos durante las lluvias. 7.2.2 Contenido de HAP´s en hígado y vísceras de peces Aunque no hubo diferencia significativa sobre el contenido de pireno en el hígado ni en víscera de los peces bajo estudio (Fig. 32) se observó una tendencia directa con la talla de los organismos durante las lluvias y fue mayor su contenido en la viscera. El contenido de benzo[a]pireno fue mayor en el hígado, particularmente en los peces de talla intermedia. En el caso de las vísceras hubo una relación inversa con la talla de los organismos; en (Fig. 33).

49
Híg
ado-
I
Híg
ado-
II
Híg
ado-
III
Vísce
ra-I
Vísce
ra-II
Vísce
ra-II
I
ug
pir
en
o/ g
tejid
o
0
500
1000
1500
2000
2500
3000
3500
Fig. 32. Contenido de pireno en el hígado y vísceras de los peces colectados durante las
lluvias. (*): no se encontró diferencia significativa (p= 0.145)
Híg
ado-
I
Híg
ado-
II
Híg
ado-
III
Vísce
ra-I
Vísce
ra-II
Vísce
ra-II
I
ug
/ g
te
jido
0
2000
4000
6000
8000
10000
12000
14000
Fig. 33. Contenido de benzo[a]pireno en el hígado y vísceras de los peces colectados
durante las lluvias. (a): diferencia significativa (p= 0.035)
*
a
*
* *
*
*
a

50
7.2.3 Factor de bioconcentración (FBC) Al igual que en la sequía fría, durante las lluvias los mayores factores de bioconcentración se documentaron para el pireno seguido del benzo[a]pireno, pero la magnitud disminuyó drásticamente comparativamente con el primer muestreo. Tampoco en estas épocas se encontró naftaleno (Fig. 34).
0.000 0.0000.000
309.132
2.468 9.334
0
50
100
150
200
250
300
350
Hígado Vísceras
TEJIDO ANALIZADO
Facto
r d
e B
ioco
ncen
tració
n
(FB
C)
Naftaleno Pireno Benzo[a]pireno
Fig. 34. Factor de bioconcentración de los HAP´s en hígado y vísceras de los charales colectados.
7.2.4 Biomarcadores de estrés oxidativo, radicales libres En contraste con la sequía fría, la concentración celular de peróxido de hidrógeno fue substancialmente mayor en víscera que en hígado aunque no hubo relación aparente con la talla del organismo (Fig. 35). En el hígado se observó una relación inversa con el tamaño de los ejemplares. A diferencia del peróxido de hidrógeno, la concentración del anión superóxido fue mayor en hígado que en víscera (Fig. 36). En los peces de mayor talla su cantidad fue la más elevada. Otro de los radicales libres bajo estudio fue el formaldehido. Su concentración fue mayor en el hígado que en vísceras (Fig. 37) pero no hubo una relación con la talla de los peces. En contraste, en la víscera tuvo una relación inversa con el tamaño de los especímenes.

51
Híg
ado-
I
Híg
ado-
II
Híg
ado-
III
Vísce
ra-I
Vísce
ra-II
Vísce
ra-II
I
mM
/ g
te
jid
o
0
1000
2000
3000
4000
5000
Fig. 35. Niveles de peróxido de hidrógeno (H2O2) en el hígado y víscera de los charales
colectados durante las lluvias. (a,b,c,d,e): diferencia significativa (p< 0.001).
Híg
ado-
I
Híg
ado-
II
Híg
ado-
III
Vísce
ra-I
Vísce
ra-II
Vísce
ra-II
I
mM
/ g
te
jid
o
0
10000
20000
30000
40000
50000
Fig. 36. Niveles de superóxido (O2-) en el hígado y víscera de los charales colectados durante las lluvias. (a,b,c,d,e,f): diferencia significativa (p< 0.001).
a
a b
b c
c
e d
d e
a
a b
b
c
c
d
d
e
e
f
f

52
Híg
ado-
I
Híg
ado-
II
Híg
ado-
III
Vísce
ra-I
Vísce
ra-II
Vísce
ra-II
I
mM
form
ald
ehíd
o/ g tejid
o
0
20
40
60
80
100
120
140
160
180
Fig. 37. Niveles de formaldehído en el hígado y víscera de los charales colectados
durante las lluvias. (a, b,c): diferencia significativa (p< 0.001).
7.2.5 Biomarcadores de daño Como sucedió durante la sequía fría, en las lluvias no hubo diferencias significativas en los niveles de lipoperoxidación. Sin embargo, fue mayor en hígado que en víscera (Fig. 38). Los hidroperóxidos lipídicos fueron más elevados en el hígado de los peces más pequeños que en víscera (Fig. 39). En éstas, los daños tuvieron una relación inversa con la talla de los peces. La oxidación de proteínas fue más alta en víscera que en hígado y además tuvo una relación inversa con el tamaño de los especímenes. En hígado, el daño fue significativo sólo en los peces de talla mayor (Fig. 40).
a
a
b
b
c
c

53
Híg
ado-
I
Híg
ado-
II
Híg
ado-
III
Vísce
ra-I
Vísce
ra-II
Vísce
ra-II
I
mM
/ g
te
jido
0
1
2
3
4
5
6
7
8
Fig. 38. Substancias reactivas al ácido tiobarbitúrico (TBAR´s) en el hígado y víscera de
los charales colectados. (*) no se encontró diferencia significativa (p= 0.020).
Híg
ado-
I
Híg
ado-
II
Híg
ado-
III
Vísce
ra-I
Vísce
ra-II
Vísce
ra-II
I
mM
/ g t
ejid
o
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
1.8
Fig. 39. Hidroperóxidos lipídicos en el hígado y víscera de los charales colectados.
(a,b,c): diferencia significativa (p< 0.001).
*
*
*
* * *
a
a
b
b
c
c

54
Híg
ado-
I
Híg
ado-
II
Híg
ado-
III
Vísce
ra-I
Vísce
ra-II
Vísce
ra-II
I
mM
/ m
g p
rote
ína/
g te
jido
0
1
2
3
4
5
Fig. 40. Oxidación de proteínas en el hígado y víscera de los charales colectados.
(a,b) diferencia significativa(p< 0.001). 7.2.6 Biomarcadores de activación La actividad del CYP 2E1 evaluada en la víscera de los peces superó por mucho a la hallada en el hígado, donde fue cercana a cero (Fig. 41) No se encontraron diferencias significativas de la actividad de la GSTT en los órganos de los peces (Fig. 42).
b a
a
b

55
Híg
ado-
I
Híg
ado-
II
Híg
ado-
III
Vísce
ra-I
Vísce
ra-II
Vísce
ra-II
I
mM
/ g p
rote
ína/ g tejid
o
0
10
20
30
40
50
60
Fig. 41. Actividad del CYP2E1 en el hígado y víscera de los charales colectados durante las lluvias. (a,b,c,d,e,f,g): diferencia significativa (p< 0.001).
Híg
ado-
I
Híg
ado-
II
Híg
ado-
III
Vísce
ra-I
Vísce
ra-II
Vísce
ra-II
I
g fo
rmald
ehíd
o/ m
in/ m
g p
rote
ína
/ g t
ejid
o
0.0
0.5
1.0
1.5
2.0
2.5
Fig. 42. Actividad de la GSTT en el hígado y víscera de los charales colectados durante las lluvias. (*): no se encontró diferencia significativa (p= 0.008).
a
a
b
b c
c
d
d e
e f
g
g
*
*
*
*
*
*

56
7.2.7 Análisis de componentes principales (ACP´s).
7.2.7.1Biomarcadores vs. HAP´s. Hígado En el hígado, se encontraron elevados niveles de correlación (mayores a 0.9) entre la concentración de piero y benzo[a] y los biomarcadores de estrés oxidativo (anión superóxido y formol), entre los biomarcadores de daño (lipoperoxidación e hidroperóxidos lipídicos) y entre estos dos grupos (producción del anión superóxido y formaldehído con los niveles de lipoperoxidación e hidroperóxidos lipídicos) (Fig. 26 y Tabla 8).
Fig. 43. Gráfico bidimensional de puntuaciones sobre los componentes principales en hígado del charal colectado durante las lluvias. H2O2: peróxido de hidrógeno; O2-: anión superóxido; 1: lipoperoxidación. 2: benzo[a]pireno; 3: hidroperóxidos lipídicos; 4: CYP
2E1; 5: GSTT
7.2.7.2 Biomarcadores vs. HAP´s. Víscera En las vísceras, en cambio, sólo se encontró una correlación importante (1.0) entre los niveles de hidroperóxidos lipídicos y la oxidación de proteínas (Fig. y Tabla 9).
-0.60
-0.40
-0.20
0.20
0.40
0.60
-0.10 -0.05 0 0.05 0.10 0.20 0.25 5 2
Oxidación de
proteínas
H2O2
O2-
Pireno
Formaldehído 1
3 4

57
Tabla 12. Matriz de correlaciones entre biomarcadores e HAP´s, evaluados en hígado.
H2O
2
O2
-
Fo
rmal
de
hid
o
CY
P 2
E1
GS
TT
lip
op
ero
xid
ació
n
HO
Oo
ox
ida
ció
n
de
p
rote
ína
s
pir
en
o
be
nz
o[a
]pir
en
o
H2O2 1
O2- -0.374 1
formaldehído -0.332 -0.086 1
CYP 2E1 0.104 0.024 0.637 1
GSTT -0.205 -0.202 0.621 0.342 1
lipoperoxidación -0.303 -0.050 0.742 0.450 0.583 1
HOOo -0.043 -0.281 0.631 0.431 0.196 0.451 1
oxidación de proteínas
-0.154 -0.306 0.581 0.498 0.432 0.429 0.403 1
pireno -0.290 0.267 0.504 0.245 0.095 0.112 0.465 0.203 1
benzo[a]pireno -0.364 -0.235 0.901 0.489 0.771 0.669 0.604 0.661 0.399 1
Nota: las variables con correlación más alta entre aparecen resaltadas.
Tabla 13. Matriz de correlaciones entre biomarcadores e HAP´s, evaluados en víscera.
H2O
2
form
ald
eh
ído
CY
P 2
E1
GS
TT
lip
op
ero
xid
ació
n
HO
Oo
oxid
aci
ón
d
e
pro
teín
as
pir
en
o
ben
zo
[a]p
iren
o
H2O2 1
formaldehído -0.162 1
CYP 2E1 0.081 -0.182 1
GSTT 0.314 -0.352 0.602 1
lipoperoxidación 0.409 -0.067 0.390 0.459 1
HOOo 0.140 0.085 0.247 0.138 -0.045 1
oxidación de proteínas
0.109 -0.176 0.823 0.655 0.264 0.049 1
pireno 0.300 -0.387 0.456 0.364 0.027 0.212 0.400 1
benzo[a]pireno 0.027 -0.195 0.961 0.488 0.341 0.302 0.677 0.537 1
Nota: las variables con correlación más alta entre aparecen resaltadas.

58
Fig. 44. Gráfico bidimensional de puntuaciones sobre los componentes principales en víscera. H2O2: peróxido de hidrógeno; GSTT: glutatión S-transferasa tetha
HOOo: hidroperóxidos lipídicos.
7.2.7.3 Materia orgánica, clorofilas y feofitina vs. HAP´s. Agua En las muestras de agua, se encontraron altas correlaciones (mayores a 0.9) entre la cantidad de pigmentos fotosintéticos (clorofilas a y b, feofitina) y HAP´s con la materia orgánica total, y entre los HAP´s (pireno y benzo[a]pireno) (Fig. 45 y Tabla 14). 7.2.7.4 Materia orgánica, clorofilas y feofitina vs. HAP´s. Sedimentos En los sedimentos, se hallaron correlaciones importantes (mayores a 0.9) entre la cantidad de HAP´s (pireno y benzo[a]pireno) y la cantidad de materia orgánica disuelta (Fig. 46 y Tabla 15).
-0.40
-0.30
-0.20
-0.10
0.10
0.20
0.30
0.40
0.50
0.60
-0.15 -0.10 -0.05 0 0.05 0.10 0.15 0.20 0.25 0.30
Formaldehído
H2O2
Lipoperoxidación
GSTT
Pireno
CYP 2E1
Benzo[a]pireno HOOo
Oxidación de proteínas

59
Fig. 45. Gráfico bidimensional de puntuaciones sobre los componentes principales en agua durante las lluvias. MOD: materia orgánica disuelta; MOP: materia orgánica
particulada. MOT: materia orgánica total.
Tabla 14. Matriz de correlaciones entre materia orgánica, pigmentos fotosintéticos e HAP´s, evaluados en agua.
MO
T
MO
D
MO
P
clo
rofi
la a
clo
rofi
la b
feo
fiti
na
pir
en
o
ben
zo
[a]p
iren
o
MOT 1
MOD -0.414 1
MOP 0.729 0.321 1
clorofila a 0.963 -0.154 0.886 1
clorofila b 0.911 -0.001 0.947 0.988 1
feofitina 0.957 -0.133 0.896 1.000 0.991 1
pireno 0.995 -0.325 0.791 0.985 0.946 0.981 1
benzo[a]pireno 0.995 -0.325 0.791 0.985 0.946 0.980 1.000 1
Nota: las variables con correlación más alta entre aparecen resaltadas.
-0.30
-0.20
-0.10
0.10
0.20
0.30
0.40
0.50
0.60
0.70
0.80
-0.04 -0.02 0 0.02 0.04 0.06 0.08 0.10 0.12 0.14 0.16
MOD
MOP
pireno
MOT
feofitina Clorofila b
Clorofila a

60
Fig. 46. Gráfico bidimensional de componentes principales en sedimento. MOP: materia orgánica particulada; MOD: materia orgánica disuelta
MOT: materia orgánica total.
Tabla 15. Matriz de correlaciones entre materia orgánica e HAP´s, evaluados en sedimentos.
MO
T
MO
D
MO
P
pir
en
o
ben
zo
[a]p
iren
o
MOT 1
MOD 0.864 1
MOP -0.950 -0.663 1
pireno 0.782 0.989 -0.547 1
benzo[a]pireno 0.707 0.967 -0.450 0.994 1
Nota: las variables con correlación más alta entre aparecen resaltadas. MOT: materia orgánica total; MOD: materia orgánica disuelta;
MOP: materia orgánica particulada.
-0.60
-0.40
-0.20
0.20
0.40
0.60
0.80
1.00
-0.20 -0.15 -0.10 -0.05 0 0.05 0.10 0.15 0.20 0.25
MOD
Benzo[a]pireno
Pireno
MOP
MOT

61
8. DISCUSIÓN Los estudios toxicológicos relacionados con el estrés oxidativo y la biactivación de compuestos orgánicos en peces son importantes por varias razones. Por ejemplo, algunos trabajos indican que los sistemas enzimáticos de detoxificación de los organismos terrestres, evolucionaron partir de los existentes en los organismos acuáticos como una adaptación a las condiciones de una atmósfera rica en oxígeno (Alvares y Pratt, 1990). Además, pueden proporcionar información valiosa sobre diferencias en la biotransformación de xenobióticos entre organismos acuáticos y especies roedoras, las cuales comparten un mayor número de características con el hombre. Por ejemplo, de manera general, se acepta que la capacidad de detoxificación de los peces es menor comparada con la de los mamíferos. Sin embargo, en estudios realizados in vitro en microsomas hepáticos del lenguado y rata con benzo[a]pireno, se encontró que la cantidad del metabolito benzo[a]pireno-7,8-diol producido fue mayor en los peces que en la rata (Varanasi y col., 1986). Aunque se ha visto que los peces pueden realizar algunas reacciones de biotransformación observadas en los mamíferos, existen diferencias en la velocidad a cual se llevan a cabo, la importancia de las vías utilizadas y los productos formados (Kleinow y col., 1987). Por otra parte, los resultados sobre la capacidad de biotransformación de peces obtenidos en especies individuales, pueden ser comparados para tratar de hallar tendencias generales. Por ejemplo, al exponer microsomas de varias especies de peces a HAP´s (benzo[a]pireno), se encontró que en los teleósteos (peces óseos) se producía mayor número de metabolitos con anillos con grupos dioles (Stegeman y Woodin, 1980) mientras en otros, como en la raya (Leucornia erinacea) se generaban más compuestos fenólicos (Bend y col., 1979). 8.1 Niveles de HAP´s en el agua, sedimento y órganos (hígado y vísceras) En los muestreos realizados en agua en las dos épocas del año (sequía fría y lluvias) la estación más contaminada con HAP´s fue el embarcadero (nivel de superficie). En la temporada de sequía fría en esta estación predominó el naftaleno, seguido del benzo[a]pireno y pireno. En cambio, durante la época de lluvias, los mayores niveles de HAP´s correspondieron al benzo[a]pireno y pireno, siendo detectados en todas las estaciones (embarcadero, centro e isla) en los tres niveles de profundidad (superficie, nivel de compensación y fondo), aunque con una diferencia de hasta tres órdenes de magnitud en comparación con la temporada anterior.

62
Las diferencias en los niveles de HAP´s en la columna de agua, pueden ser causados por diferencias en el tamaño de la molécula, a mayor peso molecular, menor solubilidad en el agua, por su coeficiente de partición octanol-agua, por su grado de adsorción a la materia orgánica, por la degradación microbiana, por la luz ó la temperatura (USEPA, 2003). El hecho de que la mayor cantidad de estos compuestos fueran encontrados en el embarcadero, demuestra que el impacto de las actividades antropogénicas, tales como el derrame de combustible y el uso de lubricantes utilizados por los motores de las embarcaciones, es determinante en las concentraciones de HAP´s de este cuerpo de agua. Por otra parte, la disminución en los niveles de los HAP´s más abundantes en la época de lluvias en comparación con la sequía fría y su presencia en todas las estaciones de muestreo, podría explicarse por el incremento en los aportes de agua del exterior a la laguna. Ya sea por las precipitaciones ó a través de los ríos que desembocan en ella, generando turbulencias, reduciendo de esta manera las concentraciones de HAP´s en el agua y distribuyéndolos más uniforme a través de la laguna. Otro factor que podría contribuir a la mezcla de estos compuestos, sería que al ser un ambiente acuático somero (su profundidad promedio es menor a 6 m), se dificultaría su estratificación, permitiendo la combinación de las masas de agua en la laguna; sin embargo, en el caso de la Laguna de Zumpango se detectó un gradiente de temperaturas de hasta 9º C en la sequía fría y de 5º C durante las lluvias entre las aguas superficiales y las profundas. Durante la temporada de sequía fría, los niveles promedio de naftaleno evaluados en las muestras de agua (59.62 mg/L) fueron muy similares a los obtenidos en muestreos previos por Ortíz-Cornejo (2008) para este cuerpo de agua con 62.8 mg/L. En el caso de los niveles de HAP´s en sedimentos, los resultados obtenidos mostraron similitudes con los observados en la columna de agua. Por ejemplo, las mayores concentraciones de estos compuestos fueron encontrados en el embarcadero en ambos periodos de muestreo. Durante la sequía fría el principal contaminante detectado fue el naftaleno, siendo reemplazado en magnitud por el benzo[a]pireno en la temporada de lluvias. Además, en ambas temporadas del año, el HAP más abundante fue el benzo[a]pireno que se halló presente en las tres estaciones de muestreo. Sin embargo, durante el periodo de lluvias, en todas las estaciones se detectó pireno en los sedimentos además del BaP. La distribución más uniforme de los HAP´s durante la temporada de lluvias en los sedimentos, podría ser explicada por un aumento en el acarreo de materiales con carga eléctrica, de los ríos hacia la laguna, tales como las arcillas, sobre las cuales podrían ser adsorbidos los HAP´s (Manahan, 2003) y transportados de esta manera por las corrientes ó las turbulencias generadas por las precipitaciones, asociándose en

63
conglomerados, aumentando así su peso y finalmente precipitándose hacia el fondo de este cuerpo de agua. Por otro lado, los altos niveles de HAP´s en agua y sedimento observados durante la sequía fría, podría estar relacionada con una disminución en su degradación por factores abióticos, tales como la luz ultravioleta (fotólisis) ó la temperatura (USEPA, 2003) ya que en esta época del año es menor irradiación solar. Por un lado, el transporte sucede por vía detritófaga ya que en la base de la pirámide alimenticia del charal, se encuentras organismos que están en contacto con los sedimentos (Figueroa-Lucero y col., 2003). Pero tampoco se puede descartar la influencia de los productores primarios y de los fitoplanctívoros como transportadores de HAP. En el caso de los órganos bajo estudio se observó un comportamiento diferenciado entre las dos épocas del año. En la sequía fría, los niveles de pireno fueron similares en ambos órganos, aunque tuvo relación inversa con el tamaño de los peces. Este fenómeno se debe posiblemente a que los sistemas enzimáticos de biotransformación se hallan menos desarrollados en los organismos más jóvenes como es considerado para la mayoría de los organismos (Repetto, 1997). 8.2 Factor de bioconcentración (FBC) En ambos periodos de muestreo, los PAH´s más abundantes en el hígado y vísceras fueron el pireno, seguido del benzo[a]pireno. Sin embargo, a diferencia de la temporada de sequía fría, en la época de secas únicamente se detectaron los dos compuestos en la víscera de los peces. Las diferencias en los niveles de ambos PAH´s en estos órganos pueden reflejar su composición diferencial (p. ej. contenido de lípidos) y por la capacidad de detoxificación. La primera de estas propiedades está relacionada con el grado de afinidad de los HAP´s por los tejidos. En el segundo caso, entre mayor sea la capacidad de detoxificación del compuesto, su eliminación del organismo será más rápida y su acumulación se reducirá. Por otro lado, durante la época de sequía fría se obtuvieron FBC´s para el pireno en hígado y víscera mayores a 2,000 y 2,500, respectivamente, por lo que este HAP cae dentro de la categoría de los contaminantes orgánicos persistentes propuesta por la EPA (substancias con un FBC mayor a 1,000). Además, ya que este hidrocarburo posee un log Kow de 5.18, también cumple con los criterio del PNUMA (log Kow> 4.0) para ser incluido en la Lista de Contaminantes Orgánicos Persistentes del Convenio de Estocolmo y por sus niveles de bioacumulación, puede ser altamente tóxico para el ambiente (ecotóxico) (Cram y col., 2004). No detectaron PCB en la columna de agua y estos resultados concuerdan con estudios previos donde tampoco se encontraron estos xenobióticos en la Laguna de Zumpango

64
(Vega-López y col., 2009). Estos hallazgos denotan que la inducción de radicales libres y el estrés oxidativo observados en ambos muestreos se deben a otros compuestos como los HAP’s entre otros. En el caso de mezclas de xenobióticos, la presencia de un componente puede influenciar la toxicidad causada por otro, al inducir ó inhibir su biotransformación. En particular, la actividad de las monooxigenasas hepáticas microsomales puede alterarse debido a contaminantes ambientales, tales como los PCB´s y HAP´s (Kleinow y col., 1987). 8.3 Biomarcadores En los dos periodos evaluados, los niveles de formaldehído e hidroperóxidos lipídicos observados en el hígado fueron mayores a los obtenidos en las vísceras, principalmente en los peces de talla pequeña. Respecto a la generación de radicales libres, los mayores niveles de H2O2 se encontraron en charales de talla intermedia particularmente en el intestino, lo que denota nuevamente la importancia de la vía trófica. En este caso también está implicada en la generación de estas especies reactivas del oxígeno. El desacoplamiento del flujo electrónico durante los procesos redox de las enzimas del citocromo P 450 y de los xenobióticos es responsable de la generación de ROS (Schlezinger y col., 1999). En el presente trabajo se observó la presencia de formaldehído en los peces, particularmente en el hígado de los peces durante la sequía fría y las lluvias se encontraron diferencias significativas entre órganos y entre tallas, presentándose los valores mayores en el hígado de los charales de talla media a pequeña. En mamíferos se ha reportado que el CYP2E1 produce metabolitos pro-carcinógenos (Schlezinger y col., 1999; Moreno Grau, 2003). La GSTT genera metabolitos intermedios como el S-clorometil glutatión, que es conjugado por la beta-liasa y cuyo resultado son tiocetonas y iones episulfonio, especies muy electrofílicas capaces de reaccionar con diferentes biomoléculas como el DNA, además de su metabolito final que es el formaldehido (Flanagan y col., 1990; Dekant y Vamvakas, 1993; Green, 1997; Landi y col., 1999). Los resultados del presente trabajo indican que existen inductores de estas enzimas en la Laguna de Zumpango como pueden ser los halometanos y los HAP’s. En la temporada de sequía fría, la relación entre los biomarcadores de estrés oxidativo (formaldehído y anión superóxido) y los de daño (lipoperoxidación e hidroperóxidos lipídicos) fue más evidente en el hígado de los peces. Ya que no se encontraron diferencias significativas entre los niveles del CYP 2E1 en los dos órganos evaluados, y se sabe que el desacoplamiento del flujo electrónico durante los procesos redox de las enzimas del CYP 450 es responsable de la generación de especies reactivas de oxígeno (Vega-López y col., 2009). Los niveles del anión superóxido observados podrían ser generados por otras isoformas del CYP 450 distintas al CYP 2E1 como el CYP 1A1 o el

65
CYP 1A2. En el caso del formaldehído, este compuesto es probable se forme a partir de los productos generados a consecuencia del proceso de peroxidación de lípidos (Buege y Aust, 1978), por la oxidación de compuestos por los microsomas hepáticos (Cederbaum y Cohen, 1980) ó durante la conjugación de compuestos clorados con el glutatión (Moreno-Grau, 2003). De manera similar, en la época de lluvias, no se observaron diferencias significativas entre la actividad de la GSTT en el hígado y víscera de los peces, por lo que en la conjugación de los metabolitos de los HAP´s podrían estar implicadas otras vías. En particular, en el pez sol (Solea solea) expuesto a fenantreno y el lenguado (Parophrys vetulus) se ha documentado la glucorinización como principal vía de conjugación (Nishimoto y col., 1992; Hillenweck y col., 2008). Sin embargo, en esta última especie se menciona la conjugación con GSH como la segunda vía más importante, y no ocurre la conjugación con sulfato. Algunos de los factores que pueden influir en la respuesta tóxica de los organismos acuáticos, son la especie bajo estudio, el tiempo de exposición y el tipo de xenobiótico. En particular, la lipoperoxidación en el hígado de los charales en las dos épocas del año, fue aproximadamente 18 veces menor que lo observado en el hígado de la carpa común (Cyprinus Carpio) colectada en este mismo cuerpo de agua (Ortíz-Cornejo, 2008) lo cual indicaría que Chirostoma riojai es una especie capaz de compensar el daño oxidativo a través de diferentes rutas. Por otra parte, en una exposición a corto plazo se observó en el mezclapique amarillo (Girardinichthys viviparus) expuesto a agua de la Laguna de Zumpango, que la lipoperoxidación presentó una respuesta dependiente del sexo de los animales. En machos se incrementó hasta 148 veces con respecto a los controles y en hembras, la respuesta fue irregular (Vega-López y col., 2008). Estos resultados indican la importancia de los fenómenos de adaptación y tolerancia desarrollados por el charal de Santiago, más no implica que estos fenómenos puedan mantenerse indefinidamente. En el intestino los hidroperoxidos lipídicos y la lipoperoxidación tuvieron relaciones significativas con los niveles de H2O2 (p<0.001 y p<0.05, respectivamente), pero no hubo diferencias entre tallas. La ausencia de diferencias entre las tallas indica que el grado de desarrollo de los peces no es un factor determinante en la susceptibilidad para que sucedan estos daños. Esta respuesta sugiere que el estrés oxidativo se detiene en etapas tempranas de las reacciones en cadena. Hermes-Lima (2004) reporta que las reacciones en cadena pueden detenerse a través de polimeración entre radicales libres como los hidroperóxidos o a través de captadores o defensas inespecíficas como varias vitaminas o el glutatión. Probablemente estos mecanismos fueron responsables de minimizar el daño sobre los fosfolípidos de membrana. Sin embargo, destaca la necesidad de otros estudios para demostrar su participación en el charal de Santiago. Respecto a la participación de los HAP’s en el estrés oxidativo, en este estudio se demostró que en hígado, los hidroperoxidos lipídicos y la lipoperoxidación además de

66
la oxidación de proteínas tuvieron una relación significativa con los niveles de pireno (p<0.001) y de BaP (p<0.05). Estos resultados muestran que el daño oxidativo hepático está asociado con el metabolismo realizado por las enzimas de la función oxidasa, específicamente las isoenzimas del citocromo P450 (Di Giulio y col., 1989). Basta mencionar que los patrones de respuesta del daño oxidativo entre los peces de la misma clase fueron similares a las enzimas que bioactivan a los COH (CYP2E1 y GSTT) aunque las relaciones no fueron significativas. Esto probablemente indica que la GSTT no desempeña un papel fundamental en la conjugación de los metabolitos de los COH. Probablemente los metabolitos de los COH en esta especie se conjugan por otras vías de conjugación. En el pez sol (Solea solea) expuesto a fenantreno y en el lenguado (Parophrys vetulus) se documento la glucorinación como vía principal de conjugación (van der Oost y col., 2003; Hillenweck y col., 2008). Sin embargo, en esta última especie se menciona la conjugación con GSH como la segunda vía más importante y no ocurre la sulfuconjugación.
No existe información disponible sobre la toxicidad de estos compuestos en Atheriniformes por lo que sobresale la necesidad de continuar estudiando al respecto.
9. CONCLUSIONES Gracias a los hallazgos del presente estudio es posible concluir que la relación entre los niveles de H2O2 producidos y el grado de daño oxidativo son producidos por la biotransformación de los HAP’s hacia metabolitos más tóxicos, particularmente a través del CYP2E1. Además en los charales de la Laguna de Zumpango prevalece un daño temprano previo a la destrucción de la membrana y que depende del órgano de los peces expuestos.
Hacen falta otros estudios como son los reproductivos o demográficos para determinar el riesgo del charal de Santiago (Chirostoma riojai) habitante de la Laguna de Zumpango.

67
10. LITERATURA CITADA APHA, AWWA y WPCF, (1992). Métodos Normalizados para el Análisis de Aguas Potables y Residuales, 17th ed., Ediciones Díaz de Santos, España. Alvares, A. P. y Pratt, W. B., (1990). Pathways of Drug Metabolism. In: Pratt, W. B. y Taylor, P. (eds.). Principles of Drug Action. The Basis of Pharmacology, 3rd ed., Churchill Livingstone, UK, 365-421. Ansaldo, M., Sacristán, H. y Wider, E., (2007). Does starvation influence the antioxidant status of the oxidative gland of Nacella concinna in experimental conditions?. Comp. Biochem. Physiol. Part C, 146(1-2): 118-123. Arriaga-Cabrera, L., Aguilar-Sierra, J., Alcocer-Durand, R., Jiménez-Rosenberg, E., Muñoz-López, E. y Vásquez-Domínguez (coords.), (1998). Regiones Hidrológicas Prioritarias. Escala de trabajo 1: 4 000 000. 2ª ed., Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México. Axler, R. P. y Owen, C. J., (1994). Measuring Chlorophyll and Phaeophytin: Whom Should You Believe?. Lake and Reserv. Manage., 8(2): 143-151. Bend, J. R., Ball, L. M., Elmamlouk, T. H., James, M. O. y Philpot, R. M., (1979). Micromosomal mixed-function oxidation in untreated and polycyclic aromatic hydrocarbon treated fish. In: (Khan, M. A. Q., Lech, J. J. y Menn, J. J. (eds.). Pesticide and Xenobiotic Metabolism in Aquatic Organism. ACS Symposium Series, American Chemical Society, Washington, DC. Bommanna, G., Loganathan, G. y Kannan, K., (1994). Global organochlorine contamination trends: an overview, AMBIO 23(3): 187. Bro-Rasmussen, F., (1994). EEC water quality objectives for chemical dangerous to aquatic environments (List 1), Rev. Environ. Contam. Toxicol., 137: 83. Buege, J. A. y Aust, S. D., (1978). Microsomal lipid peroxidation. Methods in Enzymology, 52: 302-310. Cannady, E. A., Dyer, C. A., Christian, P. J., Sipes, G. y Hoyer, P. B., (2003). Expression and activity of cytochromes P450 2E1, 2A, and “B” in the mouse ovary: the effects of 4-vinylcyclohexene and its diepoxide metabolite. Toxicol. Sci., 73: 423-430. Cederbaum, A. I. y Cohen, G., (1980). Biochem. Biophys. Res. Commun., 97: 730-736.

68
Comporti, M., (1993). Lipid peroxidation: Biopatological significance. Molec. Aspects. Med., 14: 199-207. Commandeur, J. N. M., Stijntjes, G. J. y Vermeulen, N. P. E., (1995). Enzymes and Transport Systems Involved in the Formation and Disposition of Glutathione S-Conjugates. Role in Bioactivation and Detoxication Mechanisms of Xenobiotics. Pharmacological Reviews, 47(2): 273-330. Cram, S., Ortíz, R. y Páez, R., (2004). Hidrocarburos aromáticos policíclicos. In: (Fernández-Bremauntz, A., Yarto-Ramírez, M. y Castro-Díaz, J. (Comp.). Las substancias tóxicas persistentes en México. SEMARNAT & INE, México, 173-199. Davies, M. J. y Dean, R. T., (1997). Radical-mediated protein oxidation. Oxford University Press, Oxford. Dean, J. H., Cornacoff, J. B., Rosenthal, G. J. y Luster, M. I., (1994). In: Hayes, A. W. (ed.). Principles and Methods of Toxicology, Raven Press, New York, 1065. Dekant, W. y Vamvakas, S., (1993). Glutathione-dependent bioactivation of xenobiotics. Xenobiotica, 23(8): 373-387. Di Giulio, R. T., Washburn, P. C., Wenning, R. J., Winston, G. W. y Jewel, C. S., (1989). Biochemical responses in aquatic animals: a review of determinants of oxidative stress. Environ. Toxicol. Chem., 8: 1103-1123. Di Giulio, R. T., Habig, C. y Gallagher, E. P., (1993). Effects of Black Rock Harbor sediments on indices of biotransformation, oxidative stress, and DNA integrity in channel catfish. Aquat. Toxicol., 26: 1-22. Dorfman, L. M. y Adams, G. E., (1973). Natl. Stand. Ref. Data Ser. (U. S. Natl. Bur. Stand.) NSRDS-NBS 46. EPA, (1996a). Method 8082. Polychlorinated biphenyls (PCB´s) by gas chromatography. Test Methods for Evaluating Solid Waste Physical/Chemical Methods (SW-846). Update III. U. S. Environmental Protection Agency, Office of Solid Waste & Economic, Methods and Risk Analysis Division. EPA, (1996b). Method 8072C. Semivolatile organic compounds by gas chromatography/mass spectrometry (GC/MS). Test Methods for Evaluating Solid Waste Physical/Chemical Methods (SW-846). Update III. U. S. Environmental Protection Agency, Office of Solid Waste & Economic, Methods and Risk Analysis Division.

69
Eljarrat, E. y Barceló, D., (2003). Priority lists for persistent organic pollutants and emerging contaminants base don their relative potency in environmental samples. Trends in Analytical Chemistry, 22(10): 655-665. Fernández-Bremauntz, A., Yarto-Ramírez, M. y Castro-Díaz, J., (Comp)., (2004). Las substancias tóxicas persistentes en México. SEMARNAT & INE, México. Fielder, R. J. y Martin, D., (1994). In: Ballantyne, B., Marrs, T. y Turner, P. (eds.), General and Applied Toxicology, Macmillan, Wimbledon, 1133. Figueroa-Lucero, G., Meza-González, O. R., Hernández-Rubio, Ma. C., Castro, T. B., Barriga-Sosa, I. de los A., Rodríguez-Canto, A. y Arredondo-Figueroa, J. L., (2003). Crecimiento y sobrevivencia de larvas del Charal del Alto Lerma Chirostoma riojai (Solórzano y López, 1965) (Atheriniformes: Atheronipsidae) en condiciones de laboratorio. II Congreso Iberoamericano Virtual de Acuicultura (CIVA), 927-935. Flanagan, R. J., Morgan, P. E., Spencer, E. P. y Whelpton, R., (2006). Micro-extraction techniques in analytical toxicology: short review. Biomed. Chromatogr., 20: 530-538. Ford, M. D., Delaney, K. A., Ling, L. J. y Erickson, T., (2001). Clinical Toxicology. W. B. Saunders Company, Philadelphia. Fosi, M. C., (1994). Nondestructive Biomarkers in Ecotoxicology. Environ. Health Perspect, 102(Suppl 12): 49-54. Fridovich, I., (1989). Superoxide dismutases. An adaptation to a paramagnetic gas. J. Biol. Chem., 264: 7761-7764. Fuller, B. J., Gower, J. D. y Gree, C. J., (1988). Free radical damage and organ preservation: fact or fiction. A review of the interrelationship between oxidative stress and physiological ion disbalance. Cryobiology, 25: 377-393. Furton, K. G. y Pentzke, (1998). Polycyclic Aromatic Hydrocarbons. In: Shibamoto, T. (ed.) Chromatographic Analysis of Environmental and Food Toxicants, Marcel Dekker, NY, 1-30. Furtado-Filho, O. V., Polcheira, C., Machado, D. F., Mourăo, G. y Hermes-Lima, (2007). Selective oxidative stress markers in a South American crocodilian species. Comp. Biochem. Physiol. Part C, 146(1-2): 241-254. Gerschman, K., Gilbert, D. L., Nye, S. W. Dwyer, S. y Fenn, W. O., (1954). Oxygen poisoning and X-irradiation: a mechanism in common. Science, 119: 623-626.

70
Gómez-Quiroz, L. E. y Cuevas-Bahena, D. B., (2008). ERO y señalización. In: Könisberg-Fainstein, M. (ed.). Radicales libres y estrés oxidativo. Aplicaciones médicas. Manual Moderno, México, 487-499. Green, T., (1997). Methylene chloride induced mouse liver and lung tumors: an overview of the role of mechanistic studies in human safety assessment. Human and Experimental Toxicology, 17(2): 84-87. Halliwell, B. y Gutteridge, J. M. C., (1989). Free radicals in Biology and Medicine, 2nd ed., Clarendon Press, Oxford. Halliwell, B., (1992). Reactive oxygen species and the central nervous system. Journal of Neurochemistry, 59: 1609-1623. Halliwell, B. y Cross, C. E., (1994). Oxygen-derived Species: Their Relation to Human Disease and Environmental Stress. Environ. Health Perspect., 102(Supp 10): 5-12 Hansberg-Torres, W., (2002). Biología de las especies de oxígeno reactivas. In: Cea Bonilla, A., del Arenal-Mena, I. P., Riveros-Rosas, H. y Vázquez-Contreras, E. (eds.). Mensaje bioquímico, Vol. XXVI, Fac. Medicina, UNAM, México, 19-54. Hermes-Lima, M., (2004). Oxygen in biology and biochemistry: role of free radicals. In: Storey, K. B. (ed.). Functional metabolism: Regulation and Adaptation, John Wiley & Sons, 319-368. Higson, S. P. J., (2007). Química analítica. McGraw-Hill Interamericana, México. Hillenweck, A., Canlet, C., Mauffret, A., Debrauwer, L., Claireaux, G. y Cravedi, J. P., (2008). Characterization of biliary metabolites of fluoranthene in the common sole (Solea solea). Environmental Toxicology and Chemistry, 27 (12): 2575-2581. Hussey, A. J. y Hayes, J. D., (1992). Characterization of a human class- θ-glutathione S-transferase with activity towards 1-menapthyl sulphate. Biochemistry Journal, 286: 929-935. Ifremer, A. J., (2005). Stream 4. D4.1.2. Report on the experimental results from literature of selected chemicals on the dose-response relationships. INERIS & European Commission, France. INEGI, (2000). XII Censo General de Población y Vivienda 2000. Resultados Preeliminares. Instituto Nacional de Estadística y Geografía (INEGI), México.

71
INEGI, (2001). Síntesis de Información Geográfica del Estado de México y Anexo Cartográfico. Instituto Nacional de Estadística y Geografía (INEGI), México. INEGI, (2008). Anuario Estadístico del Estado de México. Instituto Nacional de Estadística y Geografía (INEGI), México. Jacoby, W. B., (1990). Reactions of glutathione transferases: a pattern for the enzymes of detoxication. In: Hayes, J. D., Pickett, C. B. y Mantle, T. J. (eds.). Glutathione S-transferases and Drug Resistance, Taylor & Francis, London, 87-96. James, R. C. y Saranko, C. J., (2000). Chemical carcinogénesis. In: Williams, P. L., James, R. C. y Roberts, S. M., (eds. ). Principles of Toxicology. Environmental and Industrial Applications, 2nd ed., John Wiley & Sons, NY, 265-324. Kleinow, K. M., Melancon, M. J. y Lech, J. L., (1987). Biotransformation and induction: Implications for Toxicity, Bioaccumulation and Monitoring of Environmental Xenobiotics in Fish. Enviromental Health Perspectives, 71: 105-119. Könisberg, M., (1992). Bioenergética de la cadena respiratoria mitochondrial. Universidad Autónoma Metropolitana, Unidad Iztapalapa, México. Kruk, I., (1998). Environmental Toxicology and Chemistry of Oxygen Species. Springer-Verlag, Berlin. Landi, S., Hanley, N. M., Kligerman, A. D. y De Marini, D. M., (1999). Induction of sister chromatid exchanges in human peripheral blood lymphocytes by bromoform: investigation of the role of the GSTT-1 pltmorphism. Mutation Research, 429(2): 261-277. Livingstone, D. R., Lemaire, P., Matthews, A., Peters, L., Bucke, D. y Law, R. J., (1993). Pro-oxidant, Antioxidant and 7-Ethoxiresorufin O-Deethylase (EROD) Activity Responses in Liver of Dab (Limanda limanda) Exposed to Sediment Contaminated with Hydrocarbons and Other Chemicals. Mar. Pollut. Bull., 26(11): 602-606. Leikin, J. B. y Paloucek, F. P., (2008). Poisoning and Toxicology Handbook, 4th ed., CRC Press, Boca Raton. Manahan, S. E., (2003). Toxicological Chemistry and Biochemistry, 3rd ed, CRC Press, Boca Raton. Manahan, S. E., (2005). Environmental Chemistry, 8th ed., CRC Press, Boca Raton.

72
McFarland, V. A. y Clarke, J. U., (1989). Environmental Occurrence, Abundance and Potencial Toxicity of Polychlorinated Biphenyl Congeners: Considerations for a Congener-Specific Analysis. Environmental Health Perspectives, 81(225-239). Mannervik, B., Awasthi, Y., C., Board, B., Hayes, J. D., Di Ilio, C., Ketterer, B., Listowski, I., Morgenstern, R., Muramatsu, M., Pearson, W. R., Pickett, C. B., Sato, K., Widersten, M. y Wolf, C. R., (1992). Nomenclature for human glutathione transferases. Biochemistry Journal, 282: 305-308. Martin, D. W. Jr., Mayes, P. A., Rodwell, V. W. y Granner, D. K., (1985). Harper´s Review of Biochemistry, 20th ed., Lange Medical Publiclations, Los Altos, CA. Merck, (1996). The Merck Index. An Encyclopedia of Chemicals, Drugs and Biologicals, 20th ed., Merck & Co., Inc., Whitehouse Station, N. J., USA. Meyer, D. J., Coles, B., Pemble, S. E., Gilmore, K. S., Fraser, G. M. y Ketterer, B., (1991). θ: A new class of glutathione transferases purified from rat and man. Biochemistry Journal, 274: 409-414. Miller, R. R. y Smith, M. L., (1986). Origin and Geography of the Fishes of Central Mexico. In: Howtt, E. O., (ed.). Zoo-geography of North American Fishes, Willey Interscience, New York, 487-517. Moltó, J. C., Font, G., Picó, Y. y Mañes, J., (1998). Polyclorinated biphenyls In: Shibamoto, T., (ed.). Cromatographic Analysis of Environmental and Food Toxicants. Chromatographic Science Series No. 77, Marcel Dekker, NY, 31-76. Moreno-Grau, Ma. D., (2003). Toxicología Ambiental. Mc Graw-Hill/Interamericana de España, España. Ney, R., (1990). Fate and transport of organic chemicals in the environment. Government Institutes, Maryland, E. U. Nishimoto, M., Yanagida, G. K., Stein, J. E., Baird, W. M. y Varanasi, U., (1992). The metabolism of benzo(a)pyrene by English sole (Parophrys vetulus): Comparison between isolated hepatocytes in vitro and liver in vivo. Xenobiotica, 22(8): 949-961. NOM-059-SEMARNAT-2001. Protección Ambiental. Especies nativas de México de flora y fauna silvestres. Categorías de riesgo y especificaciones para su inclusión, exclusión ó cambio. Lista de especies en riesgo. Diario Oficial de la Federación del 6 de marzo de 2002.

73
NMX-AA-007-SCFI-2000. Análisis de agua. Determinación de la temperatura en aguas naturals, residuals y residuals tratadas. Método de prueba (cancela a la NMX-AA-007-1980). Diario Oficial de la Federación del 18 de diciembre de 2000. NMX-AA-008-SCFI-2000. Análisis de agua. Determinación el pH. Método de prueba (cancela a la NMX-AA-008-1980). Diario Oficial de la Federación del 18 de diciembre de 2000. NRC, (1997). Biological Markers in Environmental Health Research. Environ. Health Perspect., 74: 3-9 NRC, (1989). Biological Biomarkers in Reproductive Toxicology. National Academy Press, Washington. Ortíz-Cornejo, N. L., (2008). Inmunotoxicidad de hidrocarburos aromáticos policíclicos en la carpa común (Cyprinus carpio) crónicamente expuesta al agua de la Laguna de Zumpango. Tesis de Maestría en Ciencias Quimicobiológicas. IPN-ENCB, México. 109 p Otto, D. M. E. y Moon, T. W., (1996). Phase I and II enzymes and antioxidant responses in different tissues of brown bullheads from relatively polluted and non-polluted systems. Arch. Environ. Contam. Toxicol., 31: 141-147. Poli, G., Leonarduzzi, G., Biasi, F. y Chiarpotto, E., (2004). Oxidative stress and cell signaling. Curr. Med. Chem., 11: 1163-1882. PNUMA (Programa de las Naciones Unidas para el Medio Ambiente) y FAO (Organización de las Naciones Unidas para la Agricultura y la Alimentación), (1992). Aplicación del procedimiento de información y consentimiento previos a las Substancias Químicas Prohibidas ó Severamente Limitadas que circulan en el comercio internacional, FAO & PNUMA, Roma-Ginebra. Repetto, M., (1997). Toxicología fundamental, 3a ed., Díaz de Santos, Madrid. Richter, G., (1978). Plant Metabolism, physiology and biochemistry of primary metabolism. Trad. D. Williams. Georg Thieme Publishers, London, 25-53, 132-139. Rubin, H., (2001). Synergistic mechanism in carcinogenesis by polycyclic aromatic hydrocarbons and by tobacco smoke: a biohistorical perspective with updates. Carcinogenesis, 22: 1903-1930. Safe, S., Parkinson, A., Robertson, L., Cockerline, R., Safe, L., Bandiera, S. y Okey, A., (1982). PCBs as AHH inducers. In: Hutzinger, O., Frei, R. W., Merian, E. y Pocchiari, F.

74
(eds.) Chlorinated Dioxins and Related Compounds. Pergamon Press, New York, 383-392. Safe, S., (1987). Determination of 2, 3, 7, 8-TCCD toxic equivalent factors (TEFs): Support for the use of the in vitro AHH induction assay. Chemosphere, 16(4): 791-802. Schlezinger, J. J., White, R. D. y Stegeman, J. J., (1999). Oxidative inactivation of cytochrome P-450 1A (CYP1A) stimulated by 3,3’,4,4’-tetrachlorobiphenyl: production of reactive oxygen by vertebrate CYP1As. Mol. Pharmacol., 56: 588-597. Stegeman, J. J. y Woodin, B. R., (1980). Patterns of benzo[a]pirene metabolism in liver of the marine fish Stenotomus versicolor. Fed. Proc., 39, 1752 Storey, K. B., (1996). Oxidative stress: animal adaptations in nature. Brazilian Journal of Medical and Biological Research, 29: 1715-1733. Thomas, P. y Wofford, H. W., (1993). Effects of cadmium and Aroclor 1254 on lipid peroxidation, glutathione peroxidase activity, and selected antioxidants in Atlantic croaker tissues. Aquat. Toxicol., 27: 159-178. Turpein, M., (2006). Cytochrome P450 enzymes-in vitro, in vivo, and in silico studies. Acta Universitatis Oluluensis, D 895: 17-39. USEPA, (2003). Regulatory Determination Support Document for Naphtalene. U.S. EPA (Environmental Protection Agency). Office of Water. Standards and Risk Management Division. EPA 815-R-03-014, Washington, D. C. Valko, M., Rhodes, C. J., Moncol, J., Izakovic, M., y Mazur, M., (2006). Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chemico-Biological Interactions, 160: 1-40. van der Oost, R., Beyer, J. y Vermeulen, N. P. E., (2003). Fish bioaccumulation and biomarkers in environmental assessment: a review. Environmental Toxicology and Pharmacology, 13: 57-149. Varanasi, U., Nishimoto, M., Reichter, W. L., y LeEberhart, B. T., (1986). Comparative metabolism of benzo[a]pyrene and covalent binding to hepatic DNA in English sole, starry flounder and rat. Cancer Res., 46: 3817-3824. Vega-López, A., Jiménez-Orozco, F. A., García-Latorre, E y Domínguez-López, M. L., (2008). Oxidative stress response in an endangered goodeid fish (Girardinichtys viviparus) by exposure to water from its extant localities. Ecotoxicology and Environmental Safety 71: 94-103.

75
Vega-López, A., Jiménez-Orozco, F. A., Jiménez-Zamudio, L. A., García-Latorre, E. y Domínguez-López, M. L., (2009). Phase I Enzyme Induction in Girardinichtys viviparus, an Endangered Goodeid Fish, Exposed to Water from Native Localities Enriched with Polychlorinated Biphenyls. Arch. Environ. Contam. Toxicol., 57: 561-570. Wall, K. L. y Crivello, J., (1998). Chlorzoxazone metabolism by winter flounder liver microsomes: evidence for existence of a CYP 2E1-like isoform in teleosts. Toxicol. Appl. Pharmacol., 151: 98-104. Warholm, M., Alexandre, A. K., Högberg, J., Sigvardsson, K., y Rannug, A., (1994). Polymorphic distribution of glutathione transferase activity with methyl chloride in human blood. Pharmacogenetics, 4: 307-311. Watts, R. J., (1998). Hazardous Wastes. Sources, Pathways, Receptors. John Wiley & Sons, New York, 68-71, 76-81, 114-117. Yang, M., Koga, M., Katoh, T. y Kawamoto, T., (1999). A study for the proper application of urinary naphthols, new biomarkers for airbone polycyclic hydrocarbons. Arch. Environ. Contam. Toxicol., 36: 99-108. Yu, H., (2002). Environmental carcinogenic polycyclic aromatic hydrocarbons: photochemistry and phototoxicity. Journal of Environmental Science and Health, 20(2): 149-183.




![Effects of Fatty Acids on Benzo[a]pyrene Uptake and Metabolism in Human Lung Adenocarcinoma A549 Cells](https://static.fdokumen.com/doc/165x107/63334a02a290d455630a124b/effects-of-fatty-acids-on-benzoapyrene-uptake-and-metabolism-in-human-lung-adenocarcinoma.jpg)
![A comparative study of the reproductive effects of methadone and benzo [a] pyrene in the pregnant and pseudopregnant rat](https://static.fdokumen.com/doc/165x107/631dd0ecb5acdf8d60026ce4/a-comparative-study-of-the-reproductive-effects-of-methadone-and-benzo-a-pyrene.jpg)




![Photophysical properties and computational investigations of tricarbonylrhenium(I)[2-(4-methylpyridin-2-yl)benzo[ d]-X-azole]L and tricarbonylrhenium(I)[2-(benzo[ d]-X-azol-2-yl)-4-methylquinoline]L](https://static.fdokumen.com/doc/165x107/631cc3eea906b217b9071d28/photophysical-properties-and-computational-investigations-of-tricarbonylrheniumi2-4-methylpyridin-2-ylbenzo.jpg)
![The effects of benzo[a]pyrene on leucocyte distribution and antibody response in rainbow trout (Oncorhynchus mykiss)](https://static.fdokumen.com/doc/165x107/63254a034643260de90dad35/the-effects-of-benzoapyrene-on-leucocyte-distribution-and-antibody-response-in.jpg)
![Production of reactive oxygen species and 8-hydroxy-2′deoxyguanosine in KB cells co-exposed to benzo[a]pyrene and UV-A radiation](https://static.fdokumen.com/doc/165x107/6336f6781c5ab7fce2057d09/production-of-reactive-oxygen-species-and-8-hydroxy-2deoxyguanosine-in-kb-cells.jpg)


![Alterations to proteome and tissue recovery responses in fish liver caused by a short-term combination treatment with cadmium and benzo[a]pyrene](https://static.fdokumen.com/doc/165x107/6335a389b5f91cb18a0b7e03/alterations-to-proteome-and-tissue-recovery-responses-in-fish-liver-caused-by-a.jpg)
![Cadmium alters the formation of benzo[a]pyrene DNA adducts in the RPTEC/TERT1 human renal proximal tubule epithelial cell line](https://static.fdokumen.com/doc/165x107/634608ef38eecfb33a06d537/cadmium-alters-the-formation-of-benzoapyrene-dna-adducts-in-the-rptectert1-human.jpg)



