
Bases Ecológicas para o Desenvolvimento Sustentável - Aula 1 - A Atmosfera e o Homem
Upload
independentCategory
view
1download
0
Bacterias en la atmósfera
Irma Rosas, Eva Salinas, Leticia Martínez, Carlos Eslava y Alejandro Cravioto
1. Introducción
Una gran variedad de organismos que cambian su localización geográfica durante su ciclo de vida, lo hacen a través de la atmósfera. Por tal motivo, las partículas biológicas están siempre presentes en la atmósfera, aunque su número y viabilidad cambien con las horas del día, las condiciones del tiempo, las estaciones del año y la ubicación geográfica. El tamaño de la biota que fluye en la atmósfera varía desde micrómetros, como en el caso de virus, bacterias, esporas y polen, hasta milímetros, como las semillas y los insectos sin alas. Se pueden encontrar cerca del suelo o a grandes alturas, pero su presencia mas alláde la tropósfera no se ha determinado con precisión.
La aerobiología es una disciplina relativamente nueva, surgida alrededor de 1930 y que se encarga de estudiar el aerotransporte pasivo de los microorganismos, su identificación, comportamiento, movimientosy supervivencia, conjuntando los conocimientos de la microbiología,la meteorología, la física de los aerosoles y la química atmosférica.
La atmósfera en general no se considera un hábitat de los microorganismos, ya que sólo algunos de ellos son capaces de reproducirseallí.1 Durante su transporte bajan su tasa metabólica y se recuperan hasta que se impactan sobre un organismo o un medio con las condiciones óptimas para crecer o infectar. Sin embargo, su presencia en la atmósferatiene gran relevancia desde el punto de vista ecológico, por el grado de dispersión que pueden adquirir y que difícilmente lograrían, siendo su hábitat primario terrestre o acuático.2 Considerando la física atmosférica, las aerobacterias se asocian con los núcleos de condensación, los núcleos de congelación y con su enriquecimiento por efecto de la niebla.3,4
Los estudios aerobiológicos se iniciaron como resultado de un interés epidemiológico, para tratar un patógeno de animales, plantas o del hombre. Recientemente este tipo de investigaciones se han incrementado enel área agrícola, incluyendo plaguicidas microbianos, actividades de

composteo y daño por heladas, entre otros. Asimismo, en zonas urbanas se ha registrado la introducción de microorganismos a la atmósfera asociada a la turbulencia vehicular y a la gran densidad poblacional, lo que ha dado lugar a un campo de estudio específico. Actualmente, en respuesta a la problemática del terrorismo, se ha desarrollado infraestructura para la detección y dispersión de armas biológicas.
La mayoría de las bacterias que entran a la atmósfera provienen de fuentes naturales como la vegetación, el suelo y los cuerpos de agua, y en menor proporción de las actividades antropogénicas; su supervivencia ydistribución están moduladas por factores biológicos, meteorológicos (como el viento, la radiación solar, la temperatura, la humedad relativa)y por la química atmosférica.
Su presencia en la atmósfera ha sido demostrada por su crecimiento en medios de cultivo (denominándose cultivables); sin embargo, se considera que esto representa sólo una pequeña fracción de la población que llega ala atmósfera, de forma tal que la mayoría podría estar muerta o encontrarse en forma viable no cultivable.
2. La atmósfera como parte del hábitat
El continuo intercambio entre agua, aire y suelo nos obliga a incluir a la atmósfera como parte del hábitat de los organismos y a descubrir en ella diversos factores que determinan su distribución, en donde la dispersión juega un papel muy importante. Es un hecho que sólo para ciertos organismos la atmósfera será eficiente, y para otros resultará unmedio hostil en el cual sólo algunos podrán soportar la presión, por lo que se le considera un medio selectivo.
Por las dimensiones de este fluido de gases y partículas que rodea a la superficie terrestre, es necesario describir a la atmósfera considerando sus diferentes capas cuya altura esta relacionada con la temperatura, la presión barométrica y la densidad del aire.
A la tropósfera o atmósfera baja (que comprende los primeros diez kilómetros), por su cercanía a los ecosistemas terrestres y acuáticos, llegarán las diversas partículas biológicas en sus formas esporuladas o vegetativas, por mecanismos activos o pasivos; se distribuirán vertical yhorizontalmente, dependiendo de la energía disponible (viento, corrientesde convección, remolinos locales, etc.) lo que les proporcionará flotación y movimiento.
En esta atmósfera baja encontraremos diversas capas asociadas a las condiciones del tiempo, como noches claras, días nublados o soleados. Así

la capa límite laminar podrá tener una altura que va de pocos centímetroshasta alcanzar varios metros; una vez que las partículas biológicas entran a la capa de los remolinos locales y a la capa turbulenta, podrán llegar a zonas cuyas barreras geográficas no lo permiten.
En las noches la condición isoterma o la inversión térmica determinarán la estabilidad de la atmósfera, de tal forma que las partículas introducidas a la troposfera permanecerán en algunas ocasiones cerca del suelo (figura 1).
Figura 1. Las capas de la atmósfera y la dispersión de las partículasbiológicas
Fuente: modificado Gregory 1961.5
Es importante considerar que las dimensiones y la dinámica en cada capa atmosférica dependerán en parte de la orografía, del tipo de suelo y

dosel del lugar; es por ello que se habla de lo contrastante entre el comportamiento de una atmósfera urbana respecto de una rural (figura 2). Para el caso de las concentraciones de aerobacterias registradas durante el día se reporta: urbana> bosques> rural> zona costera.
Este enriquecimiento de partículas biológicas en la zona urbana está asociado a diversas fuentes antropogénicas, pero principalmente a la contaminación del suelo aunado a la turbulencia, que facilita la introducción de partículas a la atmósfera a diferentes alturas.
FIGURA 2. Comparación de las diversas capas atmosféricas en una zonaurbana y una rural
CLP ( capa límite etaria), CLU (capa límite urbana), CLR (capa limite rural), CDU (capa del doselurbano).
3. Las fuentes de las aerobacterias

A la atmósfera se pueden introducir una gran variedad de partículas de origen biológico, como granos de polen, esporas fúngicas, bacterias, algas, protozoarios, insectos y, ocasionalmente, virus. En general, las partículas predominan en las partes bajas de la atmósfera cerca de las fuentes locales de generación. Sin embargo, algunas esporas de hongos y bacterias pigmentadas se han recuperado a 48-77 Km. de altura.6
Las bacterias constituyen uno de los grupos más abundantes en el ambiente. En condiciones naturales se les encuentra en el suelo, el agua,y las plantas, principalmente como organismos saprobios. Debido a que carecen de mecanismos activos de liberación, son introducidas a la atmósfera por procesos mecánicos, directamente por la acción del viento yla lluvia sobre el suelo, los cuerpos de agua y la superficie de las hojas, e indirectamente por la acción de las olas y la formación de burbujas sobre los sistemas acuáticos.
Las actividades antropogénicas, como el tráfico vehicular, las plantas detratamiento de aguas residuales, los centros de manejo de desechos sólidos, el movimiento de los animales en suelos expuestos, las prácticasagrícolas y la manipulación de composta7, 8, 9 entre otros, liberan una gran cantidad de bacterias a la atmósfera, produciendo la contaminación de las áreas circundantes (cuadro1).
Cuadro1. Fuentes naturales y antropogénicas que contribuyen a incrementarla concentración de bacterias a la atmósfera
UFC: unidades formadoras de colonias; ND: no detectable
Fuente Concentración (UFC m-3)Naturales
Costa ND - 560Bosques 385 - 1.2 x 103Pastizales 127 - 587Matorral desértico 2 - 283
AntropogénicasZona urbana 539 - 7.2 x103Calles transitadas 100 - 13 x 103Parques 100 - 2.5 x 103Estación de transferencia de basura 350 - 14 x 103Planta recicladora de basura 1.1 x 103 - 2.8 x 107Planta de composteo 1 x 103 - 11 x 106

Planta de tratamiento de aguas residuales 1 x 102 - 2 x 105
Zona rural 202 - 3.4 x 103Campo agrícola 46 - 6.5 x 103Empacadora de algodón 3.3 x 106 - 19 x 106
Las bacterias suspendidas en la atmósfera generalmente se encuentran asociadas a partículas, por lo que su concentración aumenta durante la época de secas, debido al transporte convectivo de las partículas provenientes de las superficies secas. Durante la época de lluvias su número disminuye significativamente debido al lavado de la atmósfera.10
Las plantas, al ser un hábitat natural de muchos microorganismos (saprobios o patógenos), entre los que se incluyen las bacterias, contribuyen de manera importante a incrementar el número de éstas suspendidas en el aire, por la acción del viento y de la lluvia, así comopor el roce entre las mismas hojas.11 Durante la época húmeda del año la vegetación puede liberar una mayor carga bacteriana a la atmósfera que elsuelo.
La superficie de los vegetales se considera un sistema abierto, en continuo intercambio con la atmósfera. Este hábitat aéreo, cercano a la superficie de las plantas y con el que se mantiene cierta relación, se conoce con el nombre de filosfera, y sus habitantes son llamados epífitos.12 En gran parte el interés por el estudio de la filosfera se deriva de la necesidad de conocer el comportamiento (dispersión, colonización, sobrevivencia y patogenicidad), así como el control de los fitopatógenos, que son abundantes en este ambiente.
La comunidad bacteriana que compone la filosfera es muy amplia y en realidad se conoce muy poco acerca del número y especies que la integran,debido a que muchos de sus miembros se encuentran dentro del grupo de organismos no cultivables. Recientemente se han desarrollado técnicas moleculares para tratar de conocer la identidad de los miembros que integran a la comunidad de bacterias epífitas. Entre éstas se presenta una alta tasa de transferencia de plásmidos, a la vez que la abundancia de fagos encontrados sobre las plantas sugiere que la transducción también puede prevalecer. Por otra parte, la superficie de las hojas se considera como un medio de colonización hostil para las bacterias, debidoa los frecuentes cambios en la disponibilidad de agua, incidencia de la radiación y baja disponibilidad de nutrimentos, por lo que dichas cepas pueden servir como fuente de genes que codifiquen para tolerar condiciones de estrés. Esto sugiere que la superficie de las hojas puede

considerarse un área importante para la diseminación horizontal de genes y por lo tanto un sustento para la diversificación microbiana.12
Por otra parte, los animales y el hombre constituyen una fuente importante de bacterias patógenas. Las bacterias contenidas en la saliva se liberan a la atmósfera al hablar, toser y estornudar; la descamación de la piel y el cabello es una fuente constante de generación de virus, bacterias y hongos; las heces de animales y humanos pueden contaminar el suelo con microorganismos potencialmente patógenos, y existe la posibilidad de que sean suspendidos posteriormente en la atmósfera. En diversas muestras de polvo urbano (datos no publicados) y casero de la Ciudad de México se ha aislado la bacteria Escherichia coli, indicadora de contaminación fecal, y que constituye el 40% del total de bacterias coliformes aisladas en el polvo,13 lo que indica un riesgo potencial de contaminación por ésta y otras bacterias patógenas, así como por virus o parásitos.
Se reconoce que las bacterias están presentes en la atmósfera de ambientes extramuros, y que su inhalación representa un riesgo para la salud, ya sea en su forma vegetativa o parte de sus compuestos estructurales denominados “compuestos biogénicos”, como son los lipopolisacáridos de la membrana externa de las bacterias Gram negativas y los ácidos teicoicos de las Gram positivas.
5. Muestreadores
Existen diferentes tipos de muestreadores para colectar las partículas suspendidas en la atmósfera así como para determinar su distribución por tamaño. Algunos se han diseñados para el muestreo de polvo o partículas no viables, mientras que otros se usan exclusivamente para la colecta de bioaerosoles o microorganismos. A continuación se describirán algunos de los muestreadores más ampliamente usados en el área de la aerobiología para el aislamiento de bacterias.
Impactadores
El principio de colecta por impactación se basa en la tendencia de una partícula a desviarse del flujo de aire debido a la inercia, cuando la corriente de aire se curva al pasar por una superficie sólida o semisólida. Las partículas se separan de la corriente de aire y se impactan sobre la superficie.

Impactador de cascada
Dentro de esta clase de muestreadores el más usado en los estudios aerobiológicos es el impactador para partículas viables Andersen (Graseby, Atlanta, GA.). Este equipo está constituido por una serie de seis placas de aluminio, cada una con 400 orificios cuyo diámetro disminuye sucesivamente, por lo que la velocidad del aire se incrementa de una etapa a la siguiente. Succiona un flujo de aire de 28.3 L min.-1 (1 pie3) por medio de una bomba de vacío. Las partículas que son acarreadas en la corriente de aire, con un diámetro aerodinámico entre >15 a 1 ?m, son separadas por su tamaño en seis fracciones al pasar por las placas perforadas. Las partículas de una masa mayor son depositadas en la etapa superior, mientras que las partículas más pequeñas, capaces de mantenerse en el flujo de aire a baja velocidad, son transportadas sucesivamente a mayor velocidad y se impactan sobre la superficie de colecta de las siguientes etapas.14 Bajo cada placa se coloca una caja Petri con agar, en cuya superficie se desarrollarán las partículas viables (figura 3).
Existe una versión del impactador de cascada Andersen de dos etapas, cadauna con 200 orificios. Al igual que el muestreador anterior, succiona un flujo de aire de 28.3 L min-1 y las partículas son separadas en las fracciones respirable y no-respirable. La etapa superior corresponde a las placas 1 y 2 del muestreador de seis etapas y la inferior a las etapas 3 a 6 (figura 3).
Figura 3. Muestreadores de cascada Andersen de dos y seis etapas

Muestreadores de una etapa.
Existen diferentes modelos de muestreadores de una etapa. El muestreador portátil Burkard (Rickmansworth, England) colecta las partículas suspendidas con un flujo de aire de 10 L min.-1 a través de una placa perforada con 100 orificios. Se recomienda el uso de este equipo para la colecta de partículas < 5 ?m de diámetro, con una eficiencia > 95%. El muestreador N6-Andersen, el cual es una adaptación del equipo de seis etapas, sólo usa la sexta etapa del muestreador. El uso de los impactadores de una etapa es más económico en términos del número de placas de agar requeridas y del tiempo empleado en el procesamiento de las muestras; sin embargo, presenta la desventaja de no fraccionar la muestra por tamaños.
Impactadores en líquido (Impingers)
En este equipo de muestreo el aire succionado con una bomba de vacío se colecta directamente sobre un medio líquido. La mayoría de los impingers están hechos de vidrio Pyrex, con una sola cámara de colecta y un conducto para la succión del aire, el cual cuenta con un orificio críticoque determina la velocidad del flujo de aire.
Uno de los modelos es el AGI-30 (all-glass impinger), en el cual el flujode aire llega a 30 mm de la base del muestreador. Esto incrementa la eficiencia del muestreo de partículas viables, ya que reduce la velocidada la que son impactadas y disminuye el daño causado por el contacto con

la base del muestreador. Este equipo funciona con un flujo de aire de 12.5 L min-1 y generalmente se usan 20 ml de medio de colecta. La ventajade este muestreador es que se puede realizar una serie de diluciones del líquido de colecta cuando la concentración de microorganismos es muy alta.
El empleo de este tipo de equipo no se ha reducido únicamente a la colecta de partículas fúngicas y bacterias suspendidas en el aire, se ha empleado exitosamente en la colecta de algas, amibas de vida libre,15 virus y recientemente se ha utilizado el líquido de colecta en la detección de diversos microorganismos por medio de la técnica de la reacción en cadena de la polimerasa (PCR).16 El método de PCR es rápido ysensible, por lo que puede ser usado como una alternativa para la evaluación de la calidad del aire.
Impingers con fraccionamiento de tamaño
En 1960 May diseñó un muestreador que combina las ventajas de colectar las partículas suspendidas dentro de un medio líquido, con la de fraccionar las partículas por su tamaño.17 Este muestreador es conocido como MSLI (multistage all glass liquid impinger) y se presenta en tres tamaños que colectan 55, 20 y 10 L min-1 por medio de una bomba de vacío.Las partículas suspendidas en la corriente de aire se separan en tres fracciones, que corresponden por su tamaño, a la depositación en la región extra-torácica, bronquial y alveolar del tracto respiratorio (figura 4).
Una alternativa al uso del MSLI, es el impactador en líquido Burkard, el cual al igual que el muestreador anterior separa las partículas en tres fracciones (>10 ?m; 10-4 ?m; <4 ?m) con base en su diámetro aerodinámico (DA) . La ventaja de este equipo es que está fabricado en aluminio, por lo que su diseño es más exacto, y existe un riesgo menor durante los muestreos ya que es menos frágil (figura 4).
Figura 4. Muestreadores en fase líquida con fraccionamiento de tamaño

Muestreadores de centrífuga
La colecta de microorganismos por centrifugación permite la creación de un torbellino que produce que las partículas suspendidas en el aire se impacten sobre la superficie de colecta. El muestreador más común de estetipo es el Biotest RCS (Reuter Centrifugal Air Sampler; Alemania). El aire es succionado por el rotor del muestreador, que al girar crea una fuerza centrífuga y ocasiona la impactación de las partículas. Sobre las paredes de la cámara se coloca una tira plástica con agar en la que se desarrollarán las colonias de microorganismos, después de ser retirada del equipo e incubada a la temperatura adecuada. El motor funciona por medio de baterías y succiona un flujo de aire de 40 L min.-1 (figura 5). Es un equipo pequeño y de fácil manejo, por lo que su uso se ha popularizado especialmente en la evaluación de la calidad microbiológica de ambientes hospitalarios.18 Sin embargo, no es un equipo recomendado para el muestreo de ambientes ocupacionales ya que la superficie de las tiras de agar se saturan fácilmente.

Figura 5. Muestreador de centrífuga Biotest
5. Distribución temporal y espacial de las aerobacterias
Las bacterias son menos numerosas en la atmósfera que los hongos, aunque algunas veces pueden encontrarse en la atmósfera como células independientes, generalmente lo hacen asociadas a partículas que varían en tamaño de <0.65 a >7.0 ?m DA (diámetro aerodinámico), excepto en zonascosteras donde los aerosoles bacterianos son de menor tamaño. En esta área más del 84% de las partículas transportadoras de bacteria tienen un tamaño promedio de 3.6 ?m DA. La razón de este tamaño se debe a que las bacterias se asocian a materiales amorfos de plantas, a escamaciones dérmicas, suelos, hongos, aerosoles marino, núcleos de condensación o bien pueden ser introducidas a la atmósfera en conglomerados bacterianos,protegidos por solutos o sustancias mucosas.19 La figura 6 muestra la distribución de las aerobacterias por tamaño, en muestreos realizados en la Ciudad de México a las 10:00 h. durante la época de secas.10
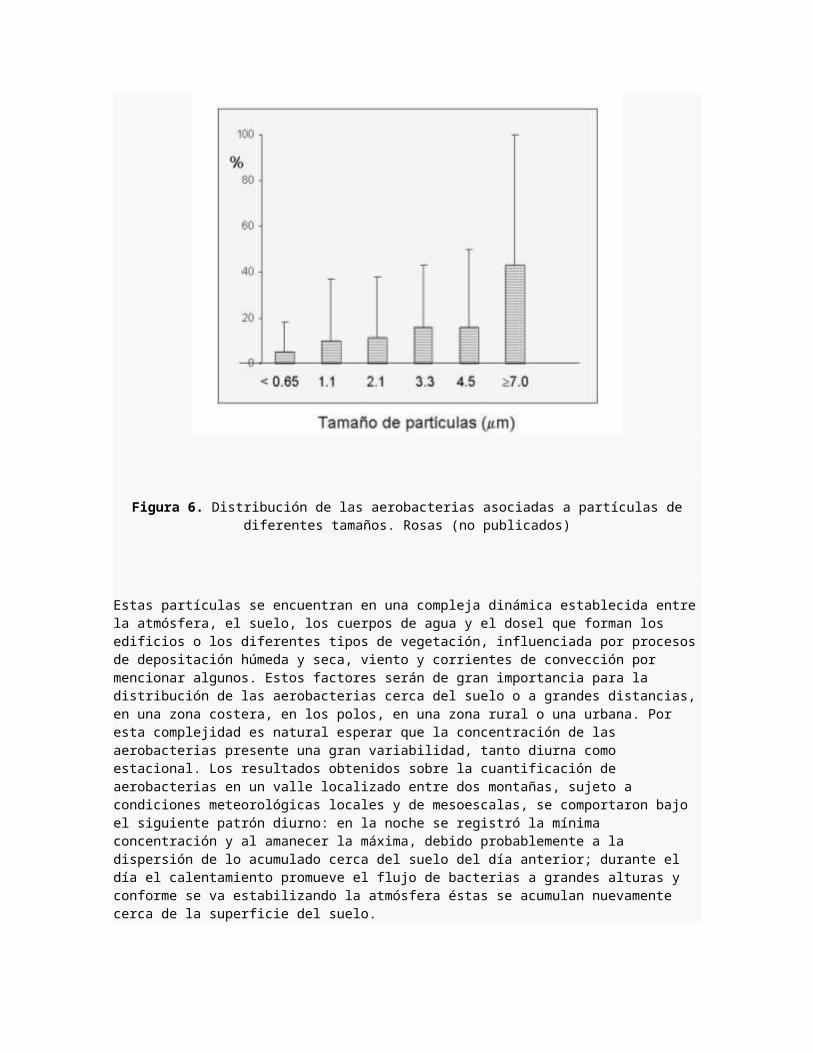
Figura 6. Distribución de las aerobacterias asociadas a partículas dediferentes tamaños. Rosas (no publicados)
Estas partículas se encuentran en una compleja dinámica establecida entrela atmósfera, el suelo, los cuerpos de agua y el dosel que forman los edificios o los diferentes tipos de vegetación, influenciada por procesosde depositación húmeda y seca, viento y corrientes de convección por mencionar algunos. Estos factores serán de gran importancia para la distribución de las aerobacterias cerca del suelo o a grandes distancias,en una zona costera, en los polos, en una zona rural o una urbana. Por esta complejidad es natural esperar que la concentración de las aerobacterias presente una gran variabilidad, tanto diurna como estacional. Los resultados obtenidos sobre la cuantificación de aerobacterias en un valle localizado entre dos montañas, sujeto a condiciones meteorológicas locales y de mesoescalas, se comportaron bajo el siguiente patrón diurno: en la noche se registró la mínima concentración y al amanecer la máxima, debido probablemente a la dispersión de lo acumulado cerca del suelo del día anterior; durante el día el calentamiento promueve el flujo de bacterias a grandes alturas y conforme se va estabilizando la atmósfera éstas se acumulan nuevamente cerca de la superficie del suelo.

Durante la tarde la entrada de aire y la brisa limpian el valle, y finalmente en la noche, el aire que baja por la pendiente de las montañastermina por desplazar lo acumulado durante el día.19 Se considera que el proceso de las primeras horas de la mañana (dilución) y el de la tarde (acumulación) se da en diversas localidades (bosque, urbano y rural), mientras que los otros procesos dependen de la cercanía a océanos o montañas (figura 7).
Figura 7. Distribución horaria de las aerobacterias durante la época desecas
Fuente: Lighthart,1997.19
Se cuenta con poca información acerca de la variación estacional de las aerobacterias; sin embargo, en algunos lugares de Europa, Estados Unidos y Canadá se han encontrado altas concentraciones de éstas en verano y bajas en invierno, asociadas con períodos polvosos de sequía durante la primera temporada, mientras que en primavera e invierno la lluvia y la nieve permiten a la disminución de concentración de aerobacterias.
Se tiene información de que el tipo de aerobacterias varía con las horas del día. En la noche las Gram positivas esporuladas presentan su

concentración mínima (17%) y las Gram negativas su máxima (22%); durante el día se invierte el proceso con 35% y 12%, respectivamente. También se ha evaluado la proporción de bacterias pigmentadas: durante el día disminuye la proporción de bacterias no pigmentadas debido a su sensibilidad a la radiación solar, por lo que se registra una mayor proporción de aerobacterias pigmentadas.20
La distribución espacial de las aerobacterias es dependiente de los flujos y la modulación meteorológica. El flujo es definido como el númerode bacterias que pasan a través de una unidad de área por una unidad de tiempo (UFC m-2 s-1) y esto a su vez, está determinado por la dinámica atmosférica. Sin embargo, se tienen pocos datos al respecto. En un matorral desértico,21, 22 se han encontrado 4.7 UFC m-2 s-1, 75 UFC m-2 s-1por encima de campos de trigo, y 543 UFC m-2 s-1 en campos de alfalfa.23, 24 Diferentes observaciones señalan que estos flujos varían durante el día a pesar de que se considera que la tasa de crecimiento de las bacterias epífitas se mantiene constante. En general puede mencionarse que:
Las aerobacterias decrecen con la altura (la capa de inversión representa una barrera para la dispersión de las bacterias).
La concentración de aerobacterias cambia dependiendo de las características de la superficie por la que atraviesa la masa de aire.
Los sistemas climáticos frontales con vientos en ráfaga pueden incrementar la concentración, mientras que disminuyen con la lluvia.
Las actividades en zonas urbanas y rurales pueden aumentar la entrada de bacterias a la atmósfera.
El efecto de la contaminación atmosférica puede afectar su viabilidad y por lo tanto su distribución espacial.
Algunas bacterias pueden viajar grandes distancias y otras pueden afectarsolamente a nivel local y su supervivencia es por corto tiempo.
Las aerobacterias son más numerosas en las ciudades (4000 m-3, promedio 850) que en áreas rurales (3400 m-3, promedio 99).25, 26, 27
Hasta este momento, utilizando el mismo muestreador y las mismas condiciones de cultivo, se obtuvieron los siguientes datos: las bacteriasGram positivas (73-90%) son el grupo predominante entre las aerobacterias, siendo abundantes las Micrococaceas y los Staphylococcus;

las formas esporuladas registran altas concentraciones en los bosques, entre intermedias y altas en zonas rurales, y bajas proporciones en zonascosteras.21 No ha sido posible identificar aproximadamente 21% de las Gram positivas cultivables.
Las Gram negativas representan una baja proporción de la población de lasaerobacterias, donde los grupos representativos son las Pseudomonas (5.5-10.7%), principalmente en zonas rurales, y las Xanthomonas (0-7.6%) en zonas costeras (cuadro 2).
En la atmósfera de zonas marinas se han registrado concentraciones de aerobacterias de 1-32 UFC m-3, incluyendo Micrococcus, Sarcina, Bacillus,Corynebacterium, bacilos pleomórficos, Gram positivos y formas esporuladas. En la estratosfera (18–27 km), no se ha determinado cuál es la mejor técnica para hacer un muestreo, sin embargo, se han logrado aislar micrococos, bacilos Gram positivos, negativos y bacterias difteroides.28
Cuadro 2. Bacterias cultivables aisladas de la atmósfera de ambientesextramuros
Microorganismo Ubicación Porcentaje deltotal
Bacillus
Oregon, EEUU (bosque, costa, urbano, rural)
16.92Arthrobacter 1.22Clavibacter 1.92Curtobacteruim 8.06Rhodococcus 0.84Micrococcus 1.46Pseudomonas 2.46Xanthomonas 2.24Staphylococcus Beijing, China 30.5Micrococcus 7.15Corynebacterium 12.85Brevibacterium 10.9Listeria 30.3Bacillius 51.3Escherichia coli
Ciudad de México (zona urbana) 4.9
Serratia 2.1

Enterobacter 46.4Aeromonas 5.5Klebsiella 1No fermentadoras 32
6. Viabilidad de las aerobacterias
Cuando los organismos son estresados por aerosolización, frío, calor, congelación, radiación, contaminación química o choque osmótico, pueden morir ó ser dañados, aunque otros pueden permanecer aparentemente sin resentir sus efectos. Las bacterias dañadas son importantes ya que no soncapaces de crecer en medios de cultivo (viables no cultivables) como lo harían las especies no dañadas; sin embargo, éstas se podrían reproducir al encontrar un ambiente adecuado, obteniendo suplementos metabólicos queles permitirían reparar el daño sufrido y así poderse reproducir.
De tal forma que la supervivencia de una bacteria al ser estresada, dependerá de su capacidad de reparar sus funciones biológicas afectadas ypodría mencionarse de manera general que lo realizan a través de dos tipos de procesos:
1) Físico-químicos2) Enzimático (dependiente de energía)
Debe mencionarse que las diferentes estructuras y compuestos de las bacterias no presentan la misma estabilidad termodinámica, siendo menos estables las membranas que los ácidos nucleicos; consecuentemente la molécula dañada dependerá de la energía del factor estresante. Tal es el caso de la desecación que cuenta con menos energía que la radiación UV, por lo que la primera afectará más fácilmente a las membranas.29 En el cuadro 3 se presentan las moléculas blanco asociadas con los factores estresantes, lo que resulta importante porque nos da idea del tipo de proceso que se deberá llevar a cabo durante la reparación.
La reparación de estructuras superficiales es necesaria ante el estrés por deshidratación o rehidratación, los cuales producen daño a estas estructuras y a las membranas; una forma en que este daño se expresa es através de la afectación de enzimas hidrolíticas; metales como el Mg, Fe, o Zn son importantes en los mecanismos de reparación, pero se ha observado que una completa reparación tiene lugar sólo en presencia de fuentes de carbón. La congelación y la descongelación producen este mismotipo de daño y se ha registrado una recuperación favorable con péptidos.

La reparación del transporte activo es un proceso que llevan a cabo algunas bacterias afectadas por deshidratación. El mecanismo de reparación se debe en parte por un incremento del ?-metil glucósido, aunque el mecanismo completo de reparación aún no se conoce.
El oxígeno, el ozono, los diferentes contaminantes ambientales y la radiación exacerban la desecación de las bacterias debido a la formación de radicales libres, los cuales oxidarán principalmente a los lípidos, afectando la fluidez de la membrana.
La reparación del daño por radiación ultravioleta induce en el ADN un dímero ciclobutano pirimidina, cercano a las uniones citosina o timina, formando un anillo ciclobutano. Muchas bacterias poseen una enzima muy específica capaz de separar este dímero, restaurando así la secuencia de bases; esta enzima tiene requerimientos estrictos de luz. La reparación también puede ser por corte del dímero de la pirimidina, completándose elespacio por la acción de la ADN-polimerasa usando la banda complementariade ADN como molde.
Se conoce que el gen rulAB, presente en plásmidos de bacterias que habitan en la filosfera, y el gen umuDC, presente en cromosoma de bacterias entéricas, están involucrados en la reparación del ADN después de la exposición a radiación ultravioleta.30
De lo anteriormente descrito, se desprende que los microorganismos que cuentan con grandes reservas de ATP y trehalosa, entre otros, serán capaces de reparar los daños causados por el estrés de la aerosolización.
Cuadro 3. Moléculas blanco afectadas por factores ambientales estresantes
Factor estresante Molécula blancoHumead relativa y temperatura Fosfolípidos de membrana, proteínas
Oxígeno Fosfolípidos, proteínasOzono Fosfolípidos, proteínasFactor aire libre (ozono+olefins)
Fosfolípidos, proteínas y ácidos nucléicos
Rayos g, rayos X y UV Fosfolípidos, proteínas y ácidos nucléicos
Fuente: Cox 1995.29

7. Relación de las aerobacterias con las fases de transición en la atmósfera
Las partículas solubles e insolubles, orgánicas e inorgánicas, naturales o antropogénicas, participan en las fases de transición vapor-liquido, vapor-sólido (hielo) y líquido-sólido.
Está bien documentado el hecho de que las aeropartículas actúan como núcleos de condensación en la formación de nubes. Sin embargo, la mayoríade ellas requieren de temperaturas de aproximadamente –10oC. La peculiaridad de algunas partículas biológicas, principalmente bacterias, es que tienen la capacidad de inducir congelación a temperaturas de –2oC,aunque la mayoría lo hacen a – 4oC. Esto significa que si el agua que lasrodea está fría, el proceso de formación de la nube puede ser inducido por estas partículas biológicas como núcleos activos de congelación.
Algunos organismos han desarrollado mecanismos mediante los cuales minimizan el daño debido a la congelación, mediante la formación de hieloextracelular. Específicamente las bacterias sintetizan proteínas con diferentes funciones:31
Proteínas núcleo de hielo, actúan como soporte para formar hielo. Proteínas anti-núcleo, inhiben la formación del núcleo de hielo,
por medio de una partícula extraña en la gota de agua. Proteínas anticongelantes, disminuyen las temperaturas de
congelación, modifican o inhiben el crecimiento de los cristales dehielo y su recristalización, y protegen a la membrana celular de los daños inducidos por el frío.
Los genes involucrados en la formación de núcleos de congelación han sidosecuenciados para Erwinia ananas (inaA) y para varias especies de Pseudomonas (inaW, inaZ); la alta homología y su posible ancestro común aún es tema de discusión.32
Se conoce poco del papel de las partículas biológicas en la formación de núcleos de condensación de nubes, aunque se especula que ciertas partículas con gran afinidad por el agua podrían tener un papel activo. Datos experimentales señalan que los granos de polen presentan propiedades higroscópicas, ya que adquieren agua de la atmósfera a una humedad relativa menor al 100%.33

Por otro lado, es importante mencionar que se considera que la niebla actúa como una fuente generadora de bioaerosoles secundarios, ya que algunas aerobacterias se duplican durante los eventos con niebla.4
8. Aerobacterias patógenas
Fitopatógenos
Para los microorganismos la dispersión es una oportunidad para que muchosde sus propágulos se distribuyan amplia y azarosamente entre áreas y hospederos, principalmente inhóspitos. Uno de los mecanismos utilizados en la dispersión consiste en la formación de propágulos resistentes y la liberación de cantidades considerables de esporas, situación que incrementa notablemente la colonización de un hábitat. Tal es el caso de numerosos hongos fitopatógenos transportados por el aire. Puccinia graminis es capaz de liberar millones de esporas por m2 en un campo de trigo, su liberación puede realizarse en períodos cortos o hasta por seismeses y no requiere de ningún otro vehículo o vector para su diseminación.34 En el caso de las bacterias, muchas de éstas provienen dela filosfera35 y su dispersión se ve favorecida durante las primeras horas de la mañana de días soleados, cuando las hojas están secas y la velocidad del viento supera 1 m/s, lo que permite que se efectúe el transporte de alrededor de 100 UFC/m2/s; sin embargo, esta concentración puede ser muy variable.23 La inmigración y emigración de fitopatógenos también puede ocurrir durante la lluvia, la irrigación de los cultivos y su cosecha, a través de aerosoles generados y transportados por el vientoo cuando las gotas de lluvia golpean la superficie de hojas infectadas.36Sin embargo, este mecanismo también puede reducir rápidamente las poblaciones de bacterias, debido al efecto de lavado que ejerce sobre ellas. Finalmente los patógenos son depositados en el suelo, el cual constituye una fuente de microorganismos que pueden ser nuevamente suspendidos en la atmósfera, siempre que la velocidad del viento sea mayor a 0.2 m/s, y el suelo se encuentre lo suficientemente seco.23, 37 Otro mecanismo de dispersión se presenta a través de vectores, como algunos insectos y aves, los cuales transportan hongos y bacterias epífitas en sus extremidades.38, 39
El transporte aéreo es extremadamente deletéreo, ya que las células son expuestas a radiación ultravioleta (UV), desecación y altas temperaturas;no obstante, algunas bacterias pueden tolerar dichas condiciones. La dispersión de los fitopatógenos puede ser local, si se realiza dentro delmismo cultivo, o de forma global, como ocurre durante las tormentas de

polvo. Un ejemplo de transporte a grandes distancias lo representa Puccinia graminis, el cual fue transportado del norte de México y el sur de Texas al norte de los Estados Unidos de América y Canadá durante la primavera, haciendo un recorrido similar en sentido contrario durante el verano y otoño. Se han registrado transportes a través de los océanos de fitopatógenos que van de Australia a Sudáfrica, de Angola a Brasil y de África al Caribe y América.40, 41, 42 Se requiere del diseño de modelos donde los eventos de liberación, dispersión y depositación en el hospedero estén interconectados con los fenómenos de turbulencia atmosférica y de esta forma poder predecir cómo, cuándo y dónde serán transportados los fitopatógenos.43
Efecto de los microorganismos en las plantas.
Las plantas superiores sirven como un ambiente donde pueden desarrollarsepoblaciones y comunidades microbianas, que incluyen bacterias, levaduras y hongos. Las bacterias constituyen uno de los colonizadores más numerosos, registrando concentraciones de 107 bacterias/cm2.12, 44 Las actividades de las plantas, la nutrición y los componentes celulares proveen una fuente de nutrimentos para dichos microorganismos, estos a suvez, son la causa de respuestas morfológicas, bioquímicas o patológicas en el individuo con el cuál están asociados. Esta relación entre macro y microorganismos es altamente interdependiente, debido a la proximidad física tan estrecha que presentan.34 Las relaciones microbianas pueden ocurrir en la rizósfera, en la filosfera o en el suelo, a una distancia cercana de la planta. El interés por dichas relaciones, se debe al impacto económico de los fitopatógenos en la producción agrícola, con unaclara necesidad de entender su comportamiento y control.
Estudios sobre la diversidad de bacterias realizados con las secuencias del ARNr del 16S revelan comunidades más complejas a las reportadas con anterioridad.45 En el cuadro 4 se enlistan algunas de las especies fitopatógenas frecuentemente aisladas en hospederos con importancia económica. Las bacterias pueden afectar el desarrollo y la productividad de una planta por diferentes mecanismos: disminuyendo la eficiencia del proceso fotosintético, como patógenos, actuando como núcleos de congelación y por la producción de fitohormonas. Sin embargo, poco se conoce sobre el papel de colonizadores no patógenos; Pseudomonas fluorescens y Pantoea agglomerans son dos especies epífitas que inhiben la colonización de fitopatógenos y actualmente son explotadas comercialmente.12, 46, 47 Los factores que participan en la relación planta-bacteria son la distribución geográfica y clima, la fenología y edad de la planta, y la genética de las bacterias.

Dentro de las características de los fitopatógenos se encuentra la abundancia de formas pigmentadas, como Erwinia herbicola, que confieren ventajas selectivas para colonizar ambientes expuestos a radicación solarintensa.48 Otro de los habitantes comunes en las hojas es Pseudomonas syringae, cuyo éxito en la colonización se debe a su capacidad para utilizar como fuente de energía y carbono compuestos como el metanol y metilaminas, los cuales se encuentran disponibles en la superficie de lashojas y constituyen una de las principales fuentes de emisión de metanol a la atmósfera.49 Esta característica le permite a la bacteria parasitar más de 80 especies de plantas, causando lesiones necróticas. Otro mecanismo de daño es su participación como núcleo de congelación.50
Barreras para la colonización
La filosfera puede ejercer una acción selectiva sobre los fitopatógenos. Las condiciones químicas, físicas y biológicas determinan cuáles células sobrevivirán, proliferarán o morirán, y aunque existen diversas especies con distribución global, algunas pueden ser excluidas debido a una o más propiedades del hábitat potencial. La condición que evita la presencia deotras especies es considerada una barrera, aunque no necesariamente debe ser un obstáculo físico, ya que pueden ser sustancias u organismos los que pueden evitar la colonización microbiana. En el caso específico de las plantas, éstas poseen cubiertas que protegen los sitios potenciales de colonización y reducen la difusión de nutrimentos, limitando el crecimiento bacteriano. Tal es el caso de la cutícula externa de los frutos, la cera de las hojas, los exudados de resina, gomas y taninos.51 Existen también compuestos fenólicos y glucósidos fungistáticos, y la presencia de diferentes enzimas, como la peroxidasa, involucrada en la resistencia contra hongos patógenos.34 Estos microorganismos también están sujetos a condiciones extremas de desecación, radiación UV, cambiosde temperatura, humedad relativa, velocidad del viento, y lluvia ácida, características por las que se considera a la filosfera como un ambiente estrés.46
Genes asociados con la patogenicidad
Se han identificado numerosos genes que capacitan a una bacteria fitopatógena en el proceso de colonización. Durante el contacto entre unaplanta y un microorganismo patógeno se produce una cascada de eventos, dentro de los cuales se distinguen dos tipos de respuesta. La primera requiere de un receptor que interactúe con alguna proteína de la bacteria, desarrollando una reacción protectora inmediata de la planta;

en esta situación, la bacteria es denominada avirulenta para un genotipo específico.52, 53 En el otro tipo de respuesta existen proteínas virulentas que afectan a la planta produciendo radicales libres en el sitio de penetración del patógeno, generando la muerte de las células infectadas.
En ambos casos se requiere de la transcripción de genes que confieran características de resistencia a las bacterias. Tal es el caso del incremento de polisacáridos extracelulares que fortalecen las paredes, laproducción de pigmentos como la melanina y la expresión de genes rulAB, responsables de mecanismos de reparación de ADN, características que les permiten a las bacterias resistir condiciones de desecación y tolerar la radiación UV.
La utilización de diferentes compuestos orgánicos como sustratos, motilidad, osmotolerancia y la capacidad para adherirse a superficies, son también adaptaciones de fitopatógenos. Algunas de estas características las codifican genes comunes, y otras genes patógenos, como es el caso del gen gae que codifica un pili tipo IV que participa enla adherencia de las bacterias.54 El modelo mejor estudiado es hrp (reacción de hipersensibilidad y patogenicidad) de Pseudomonas syringae, presente en la mayoría de las bacterias fitopatógenas Gram negativas y responsable de la necrosis de células vegetales. Algunos genes similares a hrp han sido identificados de especies no patógenas, en donde funcionancomo promotores de crecimiento en la rizósfera. Dentro de las funciones de las proteínas codificadas por los genes hrp se encuentran la regulación de la transcripción, la síntesis de los componentes de una víade secreción y la síntesis de proteínas de secreción de esta vía, lo que conforma un sistema de secreción tipo III, similar al que utilizan los patógenos de animales como Yersinia, Shigella y Salmonella spp.55, 56, 57
En otros fitopatógenos se han caracterizado nueve genes que muestran gransimilitud con las secuencias de hrp altamente conservados, denominados hrc.
Otro grupo de genes requeridos en la formación de lesiones y producción de toxinas es gacS (activador global del sensor cinasa) y gacA. En este sistema las proteínas cinasas codificadas por gacS sirven como un sensor de estímulos ambientales, en donde la señal es transmitida por fosforilación de un regulador citoplásmico, que induce cambios en la transcripción del gen gacA responsable de la producción de numerosos metabolitos que contribuyen al biocontrol de hongos de la rizósfera; ambos genes también influyen en la producción de proteasas extracelulares.58 El gen ice presente en cepas de Pseudomonas syringae esel responsable del daño por congelación, ya que muestra una correlación positiva entre la temperatura de congelación y el tamaño de la población de cepas ice +. Actualmente una estrategia de control de daño es la

utilización de cepas recombinantes ice –.12 Los genes asociados a procesos fitopatógenos continúan siendo estudiados en otras especies, conla finalidad de esclarecer el comportamiento de la microbiota presente enla filosfera.
Figura 8. Comportamiento de un fitopatógeno
La figura 8 nos muestra cómo un fitopatógeno (Pseudomonas syringae) puedeser dispersado a partir de una semilla infectada y llegar hasta la superficie de la hoja; su desarrollo depende de la susceptibilidad y estado vegetativo de la planta, la intensidad de la lluvia y del establecimiento de poblaciones numerosas. En caso contrario, si las condiciones que prevalecen son de desecación y calor, aunado al arribo a una planta desfavorable, solamente sobrevivirá el fitopatógeno. Durante la colonización es necesaria la expresión tanto de genes ordinarios como de genes patógenos (gac, hrp, ice), los cuales favorecen el incremento de

la población. En esta etapa las bacterias pueden producir lesiones, que constituyen espacios donde el organismo patógeno puede sobrevivir durantecondiciones climáticas desfavorables. Después de este evento el patógeno está preparado para la emigración y dispersión de la enfermedad. Si la planta llega a un estado maduro y logra producir semillas, el patógeno puede permanecer en ellas hasta que sean sembradas y así iniciar un nuevociclo.46
Cuadro 4. Bacterias aisladas de la filosfera de especies de importanciaeconómica
Hospedero: Centeno Hospedero: OlivoPseudomonas fluorescent Pseudomonas syringaeXanthomonas campestris Xanthomonas campestrisFlexibacter spp. Erwinia herbicolaListeria spp. Acinetobacter acetiStaphylococcus saprophyticus Gluconobacter oxydansKlebsiella spp. Pseudomonas fluorescensAcinetobacter spp. Bacillus megateriumErwinia herbicola Leuconostoc mesenteroidesPseudomonas spp. Lactobacillus plantarumStaphylococcus spp. Curtobacterium plantarumBacillus spp. Micrococcus luteusMicrococcus luteus Arthobacter globiformisAislados no identificados Klebsiella planticolaHospedero: Pera y Manzano Streptococcus faeciumErwinia amylovora Clavibacter sp.Pseudomonas fluorescens Micrococcus sp.Pantoea agglomerans Serratia marcescensHospedero: Olmo Bacillus subtilisBacillus megaterium Cellulomonas flavigenaHospedero: Frijol Erwinia spCurtobacterium citreum Zymomonas mobilisHospedero: Arroz Bacillus sp.Curtobacterium albidum Alcaligenes faecalisHospedero: Diversas Plantas Erwinia carotovoraSphingomonas pruni Pseudomona aeruginosa

La presencia de las bacterias en la atmósfera ha sido investigada principalmente por su potencial patógeno en plantas y animales, incluyendo al hombre, ya que tanto las estructuras aéreas vegetales como el tracto respiratorio se consideran sistemas abiertos, en continuo intercambio con la atmósfera.
Patógenos de humanos
En general puede mencionarse que la concentración de bioaerosoles en extramuros es mayor que la existente en intramuros; a pesar de ello la posibilidad de infección para la población general en los ambientes externos es menor. Sin embargo, la entrada de bioaerosoles a los ambientes intramuros59 representa un peligro importante principalmente enlos hospitales, en los que los pacientes con problemas inmunes pueden serafectados tanto por los microorganismos patógenos como por los oportunistas presentes en estos.
Recientemente se ha investigado la diseminación de patógenos a través de los océanos, analizando la dispersión de nubes de polvo de los desiertos africanos. Dependiendo de los vientos y la época del año, las nubes de polvo pueden llegar al norte de Europa, América del Norte, Sudamérica, América Central y el Caribe. Aunque durante mucho tiempo se consideró quela transmisión de microorganismos patógenos por esta vía era poco factible, principalmente por el tiempo que tienen que permanecer expuestos a la luz ultravioleta durante el viaje (5-7 días para cruzar elAtlántico y 7-9 días para el Pacífico), se ha mostrado que diferentes microorganismos, incluidos algunos patógenos para humanos, pueden realizar este recorrido y sobrevivir.42
Se cuenta con información42 que señala que existen billones de microorganismos por tonelada de polvo; sin embargo, el mismo estudio refiere que con los métodos de cultivo tradicionales se puede aislar únicamente el 1% de los microorganismos asociados a muestras de polvo. Por otro lado, dicho trabajo considera que la concentración de patógenos es muy baja para causar infección en humanos, aunque es importante tomar en cuenta que un individuo con respuesta inmune deficiente puede infectarse inclusive con dosis bajas y constituirse en una posible fuentede transmisión en ambientes cerrados.
Las principales fuentes de bacterias en el aire son originadas por el hombre, siendo las más importantes las aguas negras y los desechos de origen animal. La degradación y digestión de los desechos produce aerosoles que contienen bacterias, algunas de las cuales pueden ser patógenas como es el caso de los estreptococos y las coliformes fecales. El viento y las corrientes turbulentas de aire tienen enorme influencia

sobre la distancia que recorren las partículas después de ser liberadas. Un estudio realizado en la ciudad de Marsella60 mostró que el número de bacterias se incrementaba con la temperatura y la velocidad del viento. La identificación de las bacterias mostró que la localización geográfica tenía influencia cualitativa y cuantitativa sobre la biota del aire, observándose un incremento global de los microorganismos, en particular de las bacterias Gram negativas, sobre todo en el área urbana.
Las bacterias pueden producir endosporas que le confieren resistencia contra los cambios ambientales, la temperatura y la congelación. Aunque la mayoría de las bacterias esporuladas son anaerobios estrictos (Clostridium), las hay facultativas como en el caso de Bacillus. Las esporas pueden ser transportadas a grandes distancias y dispersadas por el viento, por lo que en los últimos años han adquirido gran relevancia ante la inminencia de atentados bioterroristas.
Los virus, al igual que las bacterias, normalmente son introducidos a la atmósfera a través de desechos de origen humano y animal. Sin embargo, supresencia como partículas individuales en el aire es rara. Su detección eidentificación en muestras de aire es complicada, por lo que la evidenciade su presencia en bioaerosoles se ha establecido mediante estudios epidemiológicos en veterinaria. No obstante, existen reportes de la presencia del virus de la rabia en grutas habitadas por murciélagos, y degrupos de enterovirus como: echovirus, poliovirus y coxsackievirus provenientes de muestras de aire obtenidas en sitios de riego con aguas negras.
El efecto más importante en la población humana (desde el punto de vista económico y por el número de personas afectadas), atribuido a los aerosoles de origen extramuros, son los problemas de hipersensibilidad, en particular la rinitis alérgica, el asma, así como algunas infecciones.Un ejemplo al respecto lo constituye la legionelosis, padecimiento cuyo agente etiológico es Legionella pneumophila. Se ha descrito que el principal mecanismo de infección en este caso es la transmisión directa del medio ambiente por la inhalación de gotas de agua aerosolizadas o partículas que contienen Legionella viable; la bacteria se deposita posteriormente en los alvéolos de los pulmones ocasionando una enfermedadrespiratoria severa.
Componentes de las bacterias implicados en el daño a humanos
La presencia en los bioaerosoles de componentes de la pared celular de bacterias, como es el caso de la endotoxina de las Gram negativas y los ácidos lipoteicoicos de las Gram positivas, representa un problema de

salud. La inhalación de estos compuestos causa reacciones febriles y una respuesta inflamatoria intensa en los individuos expuestos.61
Uno de los componentes principales, responsable de dicho efecto son las endotoxinas, término empleado para describir el lipopolisacárido (LPS) dela membrana externa de las bacterias Gram negativas, el cual produce un efecto tóxico. El lípido A del LPS es químicamente distinto de otros lípidos de las membranas biológicas y es el responsable de la actividad tóxica de la molécula.62
Las endotoxinas son ubicuas, dada la naturaleza cosmopolita de las bacterias Gram negativas. En ambientes extramuros se ha encontrado un patrón estacional en la concentración de endotoxinas presentes en el aire, siendo superior durante el verano.63 Sin embargo, las concentraciones más altas en la atmósfera (2-7 ?g m-3) se han obtenido enambientes ocupacionales, como son las fábricas donde se procesan fibras vegetales (como el algodón), plantas de tratamiento de aguas residuales ybioterios, entre otros.64, 65, 66
La respuesta inflamatoria inducida por la endotoxina provoca la liberación de citocinas a través de un receptor de membrana denominado CD14 y una proteína de unión al LPS (LBP) presente en el suero. Aunque secreía que CD14 era un receptor específico para la endotoxina, ahora se sabe que también los peptidoglucanos y los ácidos lipoteicoicos de la pared celular de las bacterias Gram positivas son capaces de estimular macrófagos alveolares, a través de un mecanismo dependiente de CD14.67 Sin embargo, CD14 no es un receptor que funcione como transductor de señales, y requiere de otras proteínas receptoras, como TLR (toll like receptor), que controlan la señalización. El resultado es la activación del factor nuclear NF-?? responsable de la transcripción de genes encargados de la producción de citocinas. La endotoxina durante la señalización utiliza una proteína TLR4, mientras que los peptidoglucanos y los ácidos lipoteicoicos requieren de TLR2; ambas proteínas, de manera independiente o combinada, están involucradas en la respuesta inflamatoria.
Las partículas actúan como transportadores de compuestos biogénicos así como de microorganismos y alergenos. Diversos estudios epidemiológicos han demostrado que el incremento de aeropartículas menores a 10 ?m (PM10), consideradas como partículas inhalables, afecta la salud de niñosy adultos. Esta situación se ve reflejada en un aumento en las ausencias escolares por infecciones respiratorias como la bronquitis, la exacerbación asmática, etc. Tal situación incrementa las visitas a hospitales e incluso la tasa de mortalidad.68, 69, 70
Las evidencias epidemiológicas antes mencionadas sólo comprometen el tamaño y la concentración de las partículas; sin embargo, la composición

de las mismas tiene un papel muy importante en el tipo de respuesta desarrollada. Alfaro-Moreno et al.71 demuestran que las partículas ambientales de origen industrial con concentraciones elevadas de metales de transición, colectadas en la zona norte de la Ciudad de México, son capaces de inducir apoptosis y daño al ADN en macrófagos y fibroblastos. En cambio, partículas colectadas en la zona centro, un área donde se combina el tráfico vehicular y la contaminación biológica, son responsables de una respuesta inflamatoria más significativa.
Otros investigadores, como Soukup y Becker72 y Becker et al.73 han demostrado que bacterias ambientales, como Pseudomonas sp. y Staphylococcus lentus, asociadas a las aeropartículas, son las responsables de la producción de citocinas en macrófagos y células CHO transfectadas con CD14, ya que su producción puede ser reducida hasta en un 50% cuando se utilizan inhibidores de endotoxina como la polimixina B o bloqueando el receptor CD14. Este tipo de respuesta es tres veces más alto con bacterias Gram negativas; sin embargo, éstas se presentan en menor concentración en la atmósfera. Osornio-Vargas et al.74 reportaron resultados similares en donde la producción de citocinas se ve disminuidapor la proteína neutralizante de endotoxina (PNEr); sin embargo, no se observa una completa inhibición, lo que sugiere la participación de otroscomponentes presentes en las partículas durante la respuesta inflamatoria. El aire de ambientes urbanos registra concentraciones relativamente altas de bacterias y endotoxina asociadas a partículas, porlo que su inhalación constituye un riesgo para la salud.
Bibliografía
1.Dimmick, R., H. Wolochow y M. Chatigny. 1979. Evidences that bacteria can form new cells in airborne particles. Applied Enviromental Microbiology 37: 924-927. American Society of Microbiology, EE.UU.
2.Gage, S., S. Isard y M. Colunga. 1999. Ecological scaling of aerobiological dispersal process. Agricultural and Forest Meteorology 97: 249-261. Elsevier Science, Gran Bretaña.
3.Wurzler, S., A. Bott, S. Gruber, K. Diehl y S. Mathias-Maser. 1999.The influence of biological aerosol particles on cloud microphysics: numerical case studies using new experimental data. Journal of Aerosol Science 30: S811-S812. Elsevier Science, Gran Bretaña.

4.Fuzzi, S., P. Mandreoli y A. Perfetto. 1997. Fog droplets an atmospheric source of secondary biological aerosol particles. Atmospheric Environment 31: 267-290. Elsevier Science, Gran Bretaña.
5.Gregory, P. 1961. The microbiology of the atmosphere. Plunin, N. (ed). Interscience Publishers, New York, EE.UU.
6.Imshenetsky, A. A., S. V. Lysenko y G.A. Kazakov. 1978. Upper boundary of the biosphere. Applied Environmental Microbiology 35 (1): 1-5.
7.Hughes, K. A. 2003. Aerial dispersal and survival of sewage derivedfaecal coliforms in Antarctica. Atmospheric Environment 37(22): 3147-3155.
8.Laine, M. M., K. S. Jorgensen, H. S. Kiviranta, T. S. Vartiainen, J. K. Jokela, A. K. Adibi y M. K. Salkinoja-Solonen. 1999. Bioaerosols and particles release during composting of contaminatedsawmill soil. Bioremediation Journal 3(1): 47-58.
9.Rosas, I., C. Calderón, E. Salinas, y J. Lacey. 1996. Airborne microorganisms in a domestic waste transfer station. En: Muilenberg, M. y H. Burge (eds.) Aerobiology: Proceedings of the Pan-American Aerobiology Association. CRC, Lewis Publishers, EE.UU.pp. 89-98.
10. Rosas, I., A. Yela y C. Santos-Burgoa. 1994. Occurrence of airborne enteric bacteria in Mexico City. Aerobiologia 10 (1): 39-45. Kluwer Academic Publisher, Holanda.
11. Butterworth, J. y H. A. McCartney. 1991. The dispersal of bacteria from leaf surfaces by water splash. Journal of Applied Bacteriology 71: 484-496.
12. Lindow, S. E. y J. H. Leveau. 2002. Phyllosphere microbiology. Current Opinion in Biotechnology 13(3): 238-243
13. Rosas, I., E. Salinas, A. Yela, E. Calva, C. Eslava, y A. Cravioto. 1997. Escherichia coli in settled dust and air samples collected in residential environments in Mexico City. Applied Environmental Microbiology 63(10): 4093-4095.
14. Andersen, A. A. 1958. New sampler for the collection, sizing and enumeration of viable airborne particles. Journal of Bacteriology. 76: 471-484.
15. Bonilla, P., L. Urban, N. S. Vega, I. Rosas, R. Ortiz, E. Ramírez e I. Guerra. 2001. Free-living amebas isolated from air anddust samples from homes of asthmatic children in Mexico City. En: John Libbey (ed.). IXth International Meeting on the Biology and Pathogenicity of Free-Living Amoebae Proceedings. pp. 97-101.
16. Álvarez, A. J., M. B. Buttner y L. D. Stetzenbach. 1995. PCR for bioaerosols monitoring: sensitivity and environmental

interference. Applied Environmental. Microbiology 61(19): 3639-3644.
17. May, K. R. 1966. Multistage liquid impinger. Bacteriological Reviews 30: 559-570.
18. Casewell, M. W., N. Nesai y E. Lease. 1986. The use of the reuter centrifugal air sampler for the estimation of bacterial counts in different hospital locations. Journal of Hospital Infection 7: 250-260.
19. Lighthart, B. 1997. The ecology of bacteria in the alfresco atmosphere. FEMS Microbiology Ecology 23: 263-274. Elsevier, Gran Bretaña.
20. Tong, Y. y Lighthart, B. 1997. A study of the relationship between pigmented outdoor atmospheric bacteria and solar radiation.Photochem Photobiology 65: 103-106.
21. Lighthart, B. y B. Shaffer. 1995. Airbone bacteria in the atmospheric surface layer: temporal distribution above a grass seedfield. Applied Enviromental Microbiology 61: 1492-1496. American Society of Microbiology, EE.UU.
22. Shaffer, B. y B. Lighthart. 1997. Survey of Airborne bacteriaat four diverse locations in Oregon: Urban, rural forest and coastal. Microbial Ecology 34: 167-177. Springer-Verlag, Nueva York, EE.UU.
23. Lindemann, J. y C. Upper. 1985. Aerial dispersal of epiphiticbacteria over bean plants. Applied Enviromental Microbiology 50: 1229-1232. American Society of Microbiology, EE.UU.
24. Lindemann, J., H. Constantinidou, W. Barchet, y C. Upper. 1982. Plants and sources of airborne bacteria including ice nucleation active bacteria. Applied Environmental Microbiology 44: 1059-1063. American Society of Microbiology, EE.UU.
25. Bovallius, A., B. Bucht, R. Roffey, y P. Anas. 1978. Three-year investigation of the natural airborne bacterial flora at four localities in Sweden. Applied Environmental Microbiology 35: 847-852. American Society of Microbiology, EE.UU.
26. Jones, B. y J. Cookson. 1983. Natural artmospheric microbal conditions in a typical suburban area. Applied Environmental Microbiology 45: 919-934. American Society of Microbiology, EE.UU.
27. Lacey, J. y J. Benett. 1995. Outdoor air sampling techniques.En: Cox, S. y M. Wathes (eds.). Bioaerosols Handbook. Lewis Publishers; Londres, Gran Bretaña. pp. 407-471.
28. Bovallius, A., R. Roffey y E. Henningson. 1980. Long-range transmission of bacteria. Annals New York Academic of Science 353: 186-200. Nueva York, EE.UU.

29. Cox, C. 1995. The aerobiological pathway of microorganisms. John Wiley y Sons (eds.). Nueva York, EE.UU. pp. 293.
30. Kim, J. y G. Sundi. 2000. Regulation of the rulAB mutagenic DNA repair operon of Pseudomonas syringe by UV-b (290 to 320 nanometers) radiation and analysis of rulAB- mediated mutability invitro and in planta. Journal of Bacteriology 182: 6137-6144.
31. Kawahara, H. 2002. The structures and functions of ice crystal-controlling proteins from bacteria. Journal of Bioscience and Bioengineering. 94: 492-496. Elsevier Science, Gran Bretaña.
32. Abe, K., S. Watabe, Y. Emori, M. Watanabe y S. Arai. 1989. Anice nucleation active gene of Erwinia ananas . FEBS letter 258: 297-300.
33. Matthias-Maser, S., B. Bogs y R. Jaeniske. 2000. The size distribution of primary biological aerosol particles in cloud wateron the mountain Kleiner Felberg/Taunus (Frg.). Atmospheric Research54: 1-13. Elsevier Science, Gran Bretaña.
34. Alexander, M. 1971. Microbial Ecology. Wiley (eds.) Nueva York, EE.UU. pp.511.
35. Lindemann, J., D. C. Arny, S. S. Hirano y C. D. Upper. 1981. Dissemination of bacteria, including Pseudomonas syringae, in a bean plot. Phytopathology. 71:890
36. Walker, J. C., y P. N. Patel. 1964. Splash dispersal and windas factors in epidemiology of halo blight of bean. Phytopathology 54: 140-141.
37. Delany, A. C., y S. Zenchelsky. 1976. The organic component of wind erosion generated soil derived aerosol. Soil Science 121: 146-155.
38. Evans, R. N., y D. C. Prusso. 1969. Spore dispersal by birds.Mycologia. 61(4): 832.
39. Venette, J. R. 1982. How bacteria find their hosts. En: M. S.Mount y G.H. Lacy (eds.). Phytopathogenic Prokaryotes. Vol. 2. Academic Press, Nueva York, EE.UU. pp.3-30.
40. Pedgley, D. E. 1986. Long distance transport of spores. MacMillan Publishing Company (ed). Nueva York. EE.UU.
41. Nagarajan, S., y D. V. Singh. 1990. Long distance dispersion of rust phatogens. Annals Review Phytopathology 28: 139-153.
42. Griffin, D., V. Garrison, J. Herman y E. Shinn. 2001. Africandesert dust in the Caribbean atmosphere: Microbiology and public health. Aerobiologia 17: 203-207.
43. Aylor, D., y M. Irwin. 1999. Aerial dispersal of pests and pathogens: implications for integrated pest management. Agricultural and Forest Meteorology 97: 233-234.

44. Ruinen, J. 1956. Ocurrence of Beijerinckia species in the “phyllosphere”. Nature 177: 220-221.
45. Yang, C. H., D. E. Crowley, J. Borneman y N. T. Keen. 2001. Microbial phyllosphere populations are more complex than previouslyrealized. Proceedings of the National Academy of Science 98: 3889-3894.
46. Hirano, S. S., y C. D. Upper. 2000. Bacterial in the leaf ecosystem with emphasis on Pseudomonas syringae a pathogen, ice nucleus and epiphyte. Microbiology and Molecular Biology Review. 64(3): 624-653.
47. Johnson, K. B., V. O. Stockwell, T. L. Sawyer y D. Sugar. 2000. Assessment of environmental factors influencing growth and spread of Pantoea agglomerans on and among blossoms of pear and apple. Phytopathology 90: 1285-1294.
48. Sundin, G. W., y J. L. Jacobs. 1999. Ultraviolet radiation (UVR) sensitivity analysis and UVR survival strategies of a bacterial community from phyllosfphere of field-grown peanut (Arachis hypogeae L.). Microbial Ecology 38: 27-38.
49. Corpe, W. A., y S. Rheem. 1989. Ecology of the methylotrophicbacteria on living leaf surfaces. FEMS Microbiology Ecology 62: 243-250
50. Upper, C. D., y G. Vali. 1995. The discovery of bacterial icenucleation and its role in the injury of plants by frost. En: R. E.Lee, Jr., G. J. Warren, y L. V. Gusta (eds.). Biological Ice Nucleation and its Application. American Phytopathological Society,St Paul, Minn. , pp. 29-39.
51. Schönherr, J., y P. Baur. 1996. Cuticule permeability studies: a model for estimating leaching of plant metabolites to leaf surfaces. En: Morris, C.E., P. C. Nicot and C. Nguyen (eds.).The Aerial plant surface microbiology. Plenum Press, Nueva York, EE.UU. pp. 1-23.
52. Piffanelli, P., A. Devoto y P. Schulze-Lefert. 1999. Defence signalling in cereals. Current Opinion Plant Biology 2: 295-300.
53. Martin, G. B. 1999. Functional analysis of plant disease resistence genes and their downstream effectors. Current Opinion Plant Biology 2: 273-279.
54. Suoniemi, A., K. Björklöf, K. Haahtela, y M. Romantschuk. 1995. Pili of Pseudomonas syringae pathovar syringae enhance initiation of bacterial epiphytic colonization of bean. Microbiology 141: 497-503.
55. Gopalan, S., W. Wei, y S. Y. Hei. 1996. hrp gene-dependent induction of hin1: a plant gene activated rapidly by both harpins and the avrPto gene-mediated signal. Plant Journal 10: 591-600.

56. Hei, S. Y. 1998. Type III protein secretion systems in plant and animal pathogenic bacteria. Annual Review of Phytopatology 36: 363-392.
57. Hirano, S. S., A. O. Charkowski, A. Collmer, D. K. Willis y C. D. Upper. 1999. Role of the hrp type III protein secretion system in growth Pseudomonas syringae pv. syringae B728a on host plant in the field. Proceedings of the National Academy of Science 96: 9851-9856.
58. Laville, J., C. Voisard, C. Keel, M. Maurhofer, G. Défago, y D. Haas. 1992. Global control in Pseudomonas fluorescens mediating antibiotic synthesis and suppression of black root rot of tobacco. Proceedings of the National Academy of Science 89: 1562-1566.
59. Burge, H. A. 1994. Bioaerosols Investigation. En: H. A. Burge(ed.). Bioaerosols. Max Eisenberg Series Edition. pp.1-24.
60. Di Giorgio, C., Krempff A., Guiraud H., Binder P., Tiret C., y Dumenil G. 1996. Atmospheric pollution by airborne microorganismsin the city of Marseilles. Atmospheric Environment 30: 155-160.
61. Rosas, I., H. McCartney, R. Payne, C. Calderón, J. Lacey, R. Chapela y S. Ruíz-Velazco. 1998. Analysis of the relationships between environmental factors (aeroallergens, air pollution and weather) and asthma emergency admissions to a hospital in Mexico City. Allergy 53: 394-401.
62. Milton, D. K. 1995. Endotoxins. En: H. A. Burge (ed.). Bioaerosols: Indoor Air Research Series. Lewis Publishers, Boca Raton, Florida, EE.UU. pp. 77-86.
63. Park, J. H., D. L. Spiegelman, H. A. Burge, D. R. Gold, G. L.Chew y D. K. Milton. 2000. Longitudinal study of dust and airborne endotoxin in the home. Environmental Health Perspective 108: 1023-1028.
64. Breum, N. O., B. H. Nielse, E. M. Nielsen, U. Midtgaard y O. M. Poulsen. 1997. Dustiness of compostable waste: a methodological approach to quantify the potential of waste to generate airborne microorganisms and endotoxin. Waste Management and Research 15(2): 169-187.
65. Rosas, I., C. Calderón, E. Salinas, L. Martínez, E. Alfaro-Moreno, D. K. Milton y A. R. Osornio-Vargas. 2001. Animal and worker exposure to dust and biological particles in animal care houses. Aerobiologia 17: 49-59.
66. Rylander, R., y P. Morey. 1982. Airborne endotoxin in industries processing vegetable fibers. American Industrial HygieneAssociation Journal 43(11): 811.

67. Landmann, R., B. Muller y W. Zimmerli. 2000. CD14, new aspects of ligand and signal diversity. Microbes and Infection 2: 295-304.
68. Schwartz, J., D. Slater, T. V. Larson, W. E. Pierson, y J. Q.Koening. 1993. Particulate air pollution and hospital emergency room visits for asthma in Seattle. American Review Respiratory Disease 147: 826-831.
69. Borja-Aburto, V., D. Loomis, S. Bangdiwala, C. Shy, y R. Rascon-Pacheco. 1997. Ozone, suspended particulates and daily mortality in Mexico City. American Journal of Epidemiology 145(3): 258-268.
70. Samet, J. M., S. Zeger, F. Dimici, F. Curriero, I. Coursac, D. W. Dockery, J. Schwartz y A. Zanobetti. 2000. The national morbidity, mortality and air pollution study part II: Morbidity, mortality and air pollution in the United States. HEI Research Report Number 94, Part II.
71. Alfaro-Moreno, E., L. Martínez, C. García-Cuellar, J. Bonner,C. Murray, I. Rosas, S. Ponce de León y A. Osornio-Vargas. 2002. Biologic effects induced in vitro by PM10 from three different zones of Mexico City. Environmental Health Perspectives 110(7): 715-720.
72. Soukup, J. y S. Becker. 2001. Human alveolar macrophage responses to air pollution particles are associated with insoluble components of course material, including particle endotoxin. Toxicology Applied of Pharmacology 171: 20-26.
73. Becker, S., M. Fenton y J. Soukup. 2002. Involment of microbial components and toll-like receptors 2 and 4 in cytokine responses to air pollution particles. American Journal Respiratory Cell Molecular Biology 27: 611-618.
74. Osornio-Vargas, A., E. Alfaro-Moreno, L. Martínez, C. García-Cuellar, J. C. Bonner, J. Clifford Murray, S. Ponce de León e I. Rosas. 2003. Comparative cellular effects induced by PM2.5 and PM10from two zones of México City. Environment Health Perspective. 111(10): 1289-1293.
Copyright © 2022 FDOKUMEN




















