
Continuity and Change among the Kuhl Irrigation Systems of ...
Upload
cadic-conicetCategory
view
1download
0
ORIGINAL PAPER
Assessing genetic variation and population structureof invasive North American beaver (Castor CanadensisKuhl, 1820) in Tierra Del Fuego (Argentina)
Marta Susana Lizarralde Æ Graciela Bailliet ÆSebastian Poljak Æ Mariana Fasanella ÆCecilia Giulivi
Received: 8 August 2007 / Accepted: 10 August 2007
� Springer Science+Business Media B.V. 2007
Abstract The North American beaver (Castor
Canadensis) was introduced into Isla Grande de
Tierra del Fuego, Argentina in 1946 as a potential
source of wild fur. The species showed high growth
potential, reaching close to 100,000 individuals from
an original founding stock of 25 females and 25
males. Beavers adapted rapidly to their new environ-
ment and became invasive, providing an excellent
model of successful adaptation of introduced popu-
lations to a new habitat. In this study, we used
polymorphic mitochondrial (mt) DNA to evaluate
genetic variation in the introduced beaver population
from Tierra del Fuego. Nucleotide variation in partial
sequences of Cytochrome b (500 bp) and 12S rRNA
(421 bp) genes and the main non-coding D-loop
region (521 bp) were analyzed. Our study allowed to
identify 10 different mtDNA lineages in the invasive
population, none of them shared among the source
populations. The pattern observed is a consequence
of cessation of gene flow following expansion of the
founding beaver population since the time of its
introduction. This approach contributes to the under-
standing of effects of genetic changes on survival
ability and reproductive success of invasive species.
It also has important management implications to
invasive species.
Keywords Castor Canadensis � Control region �Cytochrome b � D-loop � Genetic variation �Haplotype � Invasive population �Mitochondrial marker � 12S rRNA �Tierra del Fuego
Introduction
The management of biological invasions requires
establishing general principles to predict the perfor-
mance of a species in a new locality. Consideration of
population genetics, with explicit analyses of the
genetic structure of invasive species, may allow more
effective management of invasive species. Examples
of potential applications of population genetics in
studies of invasive species include predicting inva-
siveness to reduce occurrence of new invasions,
M. S. Lizarralde (&) � S. Poljak � M. Fasanella
Centro Regional de Estudios Genomicos, Universidad
Nacional de La Plata, Av. Calchaquı km 23.5 Piso 4,
CP 1888 Florencio Varela, Buenos Aires, Argentina
e-mail: [email protected]
M. S. Lizarralde � S. Poljak
Centro Austral de Investigaciones Cientıficas, CADIC-
CONICET, Ushuaia 9410 Tierra del Fuego, Argentina
G. Bailliet
Instituto Multidisciplinario de Biologıa Celular,
IMBICE, La Plata 1900, Argentina
C. Giulivi
Department of Molecular Biosciences, School
of Veterinary Medicine, University of California,
Davis, CA 95616, USA
123
Biol Invasions
DOI 10.1007/s10530-007-9161-6

predicting the efficacy of alternative control efforts,
and improving management of invasive species
within native communities (Sakai et al. 2001).
In 1946, 25 breeding pairs of North American
beaver, Castor canadensis Kuhl, 1820, were released
in the central region of Isla Grande, the largest island
of Tierra del Fuego Archipelago (Fig. 1). Since this
introduction, the beaver population has rapidly grown
and spread from Isla Grande to other islands of the
archipelago (Lizarralde 1993), thus becoming invad-
ers (Moorman 2007; Anderson et al. 2006a;
Lizarralde et al. 2004; Jasick et al. 2002).
Castor canadensis is native to North America
where it is widely distributed, and it has been
introduced in several Eurasian localities where it
competes with the European species C. fiber (Jenkins
and Busher 1979). The introduction of beavers into
Tierra del Fuego Archipelago dramatically affected
this sub-Antarctic ecosystem. Given that the combi-
nation of insular conditions and Antarctic influence
determines an extreme climate in the archipelago that
provides only marginal possibilities for the develop-
ment of many native species, any alteration—such as
introduction of exotic fauna—represents a significant
modification (Lizarralde and Escobar 2000). The
establishment of beavers induced irreversible changes
to the structure and function of biological communi-
ties (Lizarralde et al. 1996, 2004).
Castor canadensis has adapted successfully to sub-
Antarctic ecosystems as a geographically isolated
invasive species (Lizarralde 2006; Lizarralde et al.
2004). During its expansion, the dispersal rate of the
species has ranged from 2 to 6 linear kilometers per
year along different watercourses. As a result,
beavers occur at present throughout most of Isla
Grande de Tierra del Fuego in both the Chilean and
Argentinean sectors. The latest census of Isla Grande
and other islands in the Beagle Channel indicate that
the beaver population is near the maximum carrying
capacity of the area (Lizarralde et al. 2004). Esti-
mated population density is 0.7 beaver colonies per
km2, with more than 100,000 animals in a geograph-
ical range of approximately 70,000 km2. Physical
changes caused by the beavers have become an
integral part of the southern landscape. At present,
beavers have colonized all watersheds and streams of
Isla Grande and other islands of the Archipelago,
causing several modifications of the forest ecosystem,
such as (a) expansion of wetlands, (b) higher water
table levels and (c) alteration of the biochemical
composition of the water, sediments and soils of
riparian areas by accumulation of organic phospho-
rus, carbon and nitrogen (Lizarralde et al.1996;
Coronato et al. 2003). These changes have had
important consequences for the flora and fauna of
the region (Anderson et al. 2006b; Martinez Pastur
et al. 2006; Moorman et al. 2006).
Likewise, beaver specimens have been repeat-
edly recorded in the continental sector of Chile’s
Brunswick Peninsula since 1994, a fact that con-
firms the entrance of the species into that sector of
continental Patagonia (Skewes et al. 2006; Ander-
son et al. 2006b). Consequently, detailed and
systematized information about beaver invasion
processes at regional level is extremely relevant.
Data such as behavior of the species in other areas
of the world, ecological characteristics of their
original geographical distribution, age of the pop-
ulation in the new environment, extent of the
invasion, and knowledge of genetic aspects may
represent key information when it comes to plan-
ning management strategies.
Fig. 1 Isla Grande de
Tierra del Fuego. Location
of the study area within
Tierra del Fuego National
Park according to Martinez
Pastur et al. (2006)
M. S. Lizarralde et al.
123

The analysis of the genetic attributes of invasive
species suggests that their adaptive success involves
genetic traits that are determinant enough to ensure
rapid expansion in a new environment (Nei 1975;
Roughgarden 1979; Sakai et al. 2001). Of particular
interest is the concept that biological invasions allow
species to alter their own genetic pattern (Wright
1955; Carson 1987) and exhibit an extremely wide
range of evolutionary patterns (Brown and Marshall
1986). In addition, recent awareness of the loss of
genetic variability due to inbreeding and founder
effects in invaders (Sakai et al. 2001) and their
significance for the preservation of these species
warrants an extensive study of the Tierra del Fuego
beaver population.
Relatively low genetic variability has been
observed in several natural populations, such as
muskrat Ondatra zibethicus introduced in Finland, or
beaver C. fiber in Europe (Ellegreen et al. 1993;
Durka et al. 2005) and it has been attributed to
extensive population bottleneck phenomena. In the
case of C. canadensis introduced in Tierra del Fuego,
the impressive growth of this population suggests that
its viability and fertility were not drastically affected
by genetic founder effect. But, until now, genetic
structure of the introduced beaver population was
unknown.
The goals of this work were to evaluate genetic
diversity within this invasive beaver population and
test the current population structure, based on the
following considerations: (i) Tierra del Fuego bea-
vers are geographically isolated from other
populations of C. canadensis; (ii) they represent an
interesting model case to study the consequences of
population founder events initiated by the introduc-
tion of a small number of animals, and (iii) the
North American beaver established in Isla Grande
de Tierra del Fuego is also an informative case for
studying invasion processes. To address these goals,
we analyzed a hypervariable partial sequence of the
mitochondrial DNA (mtDNA) D-loop (or control
region), which has already proved to be suitable for
investigations at the population level in several
mammalian species (Ducroz et al. 2005; Durka
et al. 2005; Reyes et al. 2003; Matson and Baker
2001; Matsuhashi et al. 1999, Douzery and Randi
1997; Brown et al. 1986). We also analyzed
segments of cytochrome b and 12S rRNA as sources
of complementary information about the genetic
structure of the population. This study was per-
formed on a population from the southern area of
Isla Grande de Tierra del Fuego located within
Tierra del Fuego National Park (Fig. 1), and its
results are particularly important for the planifica-
tion of future management programs in this and
other areas with similar problems.
Finally, the genetic information obtained from
this work is especially significant to monitor the
present advance of this population onto newly
invaded areas of the archipelago, such as Brunswick
Peninsula in Chile, and to be used in future control
programs, both to assess the integrity of genetically
different units and to ensure successful control
actions that take advantage of the negative impacts
of inbreeding.
Materials and methods
Tissues and specimens analyzed
A total of 30 C. canadensis specimens from water-
courses (Cruz, Negro, Los Castores, Lapataia and
Ensenada creeks) located within 75 km2 Tierra del
Fuego National Park study area (54� 50’ 32.4’’ SL, 68�32’ 11.5’’ WL), 15 km west of Ushuaia city (Fig. 1),
were analyzed. DNA was extracted from samples of
fresh tissue (liver, spleen, muscle) of specimens
collected by park guards (Table 1). In addition, samples
of fixed tissues (liver and muscle) of 5 specimens from a
source population (Admiralty Island, Alaska, USA)
were included as outgroup for the analysis. Three sets of
mt-DNA (cytochrome b, D-loop and 12S rRNA)
C. canadensis and C. fiber sequences obtained from
GenBank database (accession numbers AF293348,
AF155878, AF155879, CF1389529, AY623644-6,
AY623642-3, DQ088702-3, AY012111 and U67297)
were also used for the comparisons.
Skulls and mandibles of the specimens analyzed
were deposited in the mammal collection of Museo de
Ciencias Naturales de La Plata, Argentina. Tissues and
other data associated with each specimen are refer-
enced directly to each voucher specimen and stored
along with a field catalog number in the following
laboratory collections: Centro Regional de Estudios
Genomicos, Florencio Varela, Argentina (Pcc) and US
Forest Service, Juneau, Alaska, USA (JB02).
Genetic structure of invasive beaver population in Tierra del Fuego
123

A detailed list of specimens sequenced per sam-
pling site is given below.
Specimens examined
Castor canadensis 1 (Pcc240) male (M) Los Castores
creek, Tierra del Fuego National Park, TFNP, Argen-
tina; 2 (Pcc229) M Los Castores creek, TDFNP; 3
(Pcc226) female (F) Negro creek TDFNP; 4 (Pcc239)
M Cruz creek TDFNP; 5 (Pcc227) M Negro creek
TDFNP; 6 (Pcc228) F Los Castores creek TDFNP; 7
(Pcc210) F Cruz creek TDFNP; 8 (Pcc235) F Cruz
creek; 9 (Pcc 224) F Cruz creek TDFNP; 10 (Pcc 218) F
Ensenada TDFNP; 11 (Pcc99) F Lapataia TDFNP; 12
(Pcc204) F Ensenada TDFNP; 13 (Pcc225) F Los
Castores creek TDFNP; 14 (Pcc87) F Lapataia
TDFNP, 15 (Pcc89) F Lapataia TDFNP, 16 (Pcc90)
F Lapataia TDFNP, 17 (Pcc91) M Lapataia TDFNP, 18
(Pcc100) F Los Castores creek TDFNP, 19 (Pcc 123) F
Los Castores creek TDFNP, 20 (Pcc165) M Cruz creek
TDFNP, 21 (Pcc200) M Negro creek TDFNP, 22
(Pcc201) M Negro creek TDFNP, 23 (Pcc203) F
Ensenada TDFNP, 24 (Pcc204) F Ensenada TDFNP,
25 (Pcc209) M Cruz creek TDFNP, 26 (Pcc211) F
Negro creek TDFNP, 27 (Pcc217) M Ensenada
TDFNP, 28 (Pcc213) M Cruz creek TDFNP, 29
(Pcc214) M Los Castors creek TDFNP, 30 (Pcc215)
F Los Castores Creek TDFNP, 31 (JB02-01) M Mile
2.8 Greens Cr. Rd Admiralty Island (GCRAI), Alaska
USA; 32 (JB02-02) F Mile 4.8 GCRAI; 33 (JB02-04) F
Mile 3.8 GCRAI; 34 (JB02-05) M Mile 3.8 GCRAI; 35
(JB02-06) F Mile 2.6 GCRAI.
DNA extraction, amplification and sequence
analyses
DNA was extracted from fresh and fixed liver, kidney
and muscle tissue following the sodium duodecyl
sulfate-proteinase K/phenol/RNase method (Sam-
brock et al. 1989). Samples were concentrated by
ethanol precipitation. Amplified sequences were
cytochrome b gene (approximately 600 bp), 12
SrRNA (approx. 400 bp) gene and D-loop or control
region (approx. 500 bp).
Primer sequences
The letters L and H in the following sequences denote
light and heavy strands, respectively; the number
indicates position of the 30 base of the primer in the
complete mouse mtDNA sequence (Bibb et al. 1981).
Cytochrome b gene was amplified using a combina-
tion of primers MVZ 05 - L14115 (50-CGAAGCTTG
ATATGAAAAACCATCGTTG-30) and MVZ14 -
H15825 (50-GGTCTTCATCTYHGGYTTACAAGA
C-30) (Smith and Patton 1993), 12S rRNA gene with
L1091 (50- AAAAAGCTTCAAACTGGGATTAGA
Table 1 Castor canadensis examined for the sequence variation of mitochondrial DNA fragments of Cytochrome b, D-loop and 12
rRNA (12S)
Haplotype Frequency Base pairs Accesion number Sampling site locations
Cytb A 0.38 500 AY793641 East (Cruz) and West (Lapataia, Los Castores )
Cytb B 0.19 500 AY793642 East (Ensenada) and West (Los Castores, Negro)
Cytb C 0.12 500 AY793643 East (Cruz) and West (Negro, Lapataia)
Cytb D 0.31 500 AY793644 Admiralty Island (Alaska, USA)
DLoop A 0.18 503 AY787822 East (Cruz, Ensenada)
DLoop B 0.12 503 AY787823 West (Negro, Los Castores)
DLoop C 0.06 503 AY787824 West (Negro, Lapataia)
DLoop D 0.12 503 AY787825 East (Cruz, Ensenada)
DLoop E 0.12 503 AY787826 East (Cruz, Ensenada)
DLoop F 0.06 503 AY787827 West (Lapataia, Negro, Los Castores)
DLoop G 0.31 521 AY968083 Admiralty Island (Alaska, USA)
12SrRNA 1.00 421 AY787828 West and East TDFNP sites, Admiralty Island (USA)
Individuals are arranged according to their, type of haplotype, deposited number and geographical location origin: Tierra del Fuego
National Park (Western and East), Argentina and Admiralty Island (AIUSA), Alaska, USA
M. S. Lizarralde et al.
123

TACCCCACTAT-30) and H1478 (50- TGACTGCA-
GAGGGTGACGGGCGGTGTGT- 30) described by
Kocher et al. (1989). Hypervariable domain I (HVI)
(Saccone et al. 1987) of the control region (D-loop)
was amplified using the following universal primers:
Thr-L15926 (50-CAATTCCCCGGTCTTGTAAACC-
30) located in the neighboring tRNA-pro gene and
DL-H16340 (50-CCTGAAGTAGGAACCAGATG-
30) (Vila et al. 1999; Ducroz et al. 2005; Durka et al.
2005).
Amplification of the double-stranded product was
performed in 25 ll or 50 ll total reaction volume
with two polymerase chain reaction (PCR) thermal
profiles using Thermus aquaticus DNA-polymerase
in a MJ thermocycler. About 25 ll of PCR mix
contained 1.25 U of Platinum Taq DNA polymerase
(InvitrogenTM), 2.5 ll of 10 · Tag polymerase buffer
with (NH4) SO4, 1.5 mM of MgCl2, 200 lM of
DNTP’s and 5 pmol of each primer. Thermal profiles
of cytochrome b and D-Loop consisted of denatur-
ation for 1 min at 94�C, annealing for 1.5 min at
55�C and extension for 2–6 min at 72�C; the cycle
was repeated 40 times. Thermal profile of 12 S rRNA
consisted of denaturation for 2 min at 94�C, anneal-
ing for 1 min at 65�C and extension for 1–3 min at
73�C; the cycle was repeated 30 times.
Double-stranded PCR products were purified with
Qiaquik PCR Purification Kit (QiagenTM), eluted in
water following the manufacturer’s indications, and
directly sequenced in both directions using the same
primers for amplifications. Sequencing was con-
ducted using ABI Prism Automated 377 and 3100
Genetic Analyzer Sequencers (Applied Biosys-
temsTM), at the Division of Sequencing Services of
Universidad Nacional de San Martın (Buenos Aires,
Argentina) and the Advanced Genetic Analysis
Center of University of Minnesota (Minnesota,
USA) respectively. Sequences for both strands were
determined.
The sequences were visually edited, managed and
aligned using Biology WorkBench 3.2 (http://work
bench.sdsc.edu) or Chromas 2.3 (Technelysium Pty.
Ltd. 1998–2004 http://www.technelysium.com.au)
and the computer software CLUSTAL W (Thompson
et al. 1994). Alignments were then optimized manu-
ally. To evaluate sequence divergence analyses
between groups, we clustered populations from study
area of 75 km2 Tierra del Fuego National Park of
using Phylip 3.57 (Felsenstein 2004) into two groups:
West (Los Castores, Negro and Lapataia creeks) and
East (Ensenada and Cruz creeks) (Table 1).
Sequences were deposited in GenBank under
accession numbers AY793641 to AY793644,
AY787822 to AY787827, AY968083, and AY787828.
A median-joining network (MJ) (Bandell et al.
1999) was constructed using Network 4 (http://www.
fluxus-engineering.com) to visualize relationships
among haplotypes. Haplotype diversity (h) and pair-
wise divergence (Nei 1987; Tajima 1993) were
computed using ARLEQUIN version 3.01 (http://
cmpg.unibe.ch/software/arlequin3) (Excoffier et al.
2006). Analysis of genetic diversity was performed as
described in Nei (1987). Pairwise differences among
haplotypes were used as measurement of molecular
distance (Nei 1987, Tajima 1993, Excoffier 2004).
Mismatch distribution analysis followed Schenider
and Ecoffier (1999). Correlation of genetic and geo-
graphical distances was tested by a Mantel test
between the pairwise Fst values and mean net number
of pairwise differences (Tajima 1993; Fu 1997). All
computation analysis were performed as implemented
in ARLEQUIN (1,000 permutations).
Results
Sequence analysis of a total number of 35 specimens
analyzed in this study revealed 12 new haplotypes.
These new Castor canadensis mt-DNA haplotypes
are available from GenBank under numbers
AY793641-44 (cytochrome b), AY 787822 to AY
787827 and AY968083 (D-loop), and AY787828
(12S rRNA). The sequences are not complete and
have the standard gene order of mammals. The 12
new haplotypes reported here represent a data set
with 18 mt-DNA genomes (GenBank database
http://www.ncbi.nlm.nih.gov/Genbank) from distinct
C. canadensis populations in the Northern and
Southern Hemispheres.
Cytochrome b sequences
Sequence divergence
Cytochrome b sequences of 500 base pairs were
available. Sequence data for 3 new haplotypes (CytbA,
CytbB and CytbC) were detected and also deposited in
Genetic structure of invasive beaver population in Tierra del Fuego
123

GenBank (Accession numbers AY793641-44).
Sequence analysis of the 5 C. canadensis samples
from the source population revealed another new
different haplotype (CytbD) also deposited in Gen-
Bank (Accession number AY793644). Gene diversity
obtained was 0.7583 ± 0.0603 (mean ± SD).
Nucleotide sequence variation within the 500 bp
of cytochrome b consisted of 3 variable sites located
at positions 208, 327 and 330. All substitutions were
A/T transitions. Table 2 shows the distribution of
variable sites among C. canadensis haplotypes in the
500 bp alignment analysis. Heteroplasmy was not
detected in the nucleotide sequence of the fragment.
Haplotype CytbA was dominant and shared among
the sites studied. Frequency of haplotypes ranged
from 0.12 to 0.38 (Table 1). Nucleotide diversity (p)
was 0.49 ± 0.35.
The new 4 C canadensis sequences differed from
previously reported sequences (Kuehn et al. 2000;
Montgelard et al. 2002; Ducroz et al. 2005). Haplo-
type CytbD from the source population also differed
by 3 point mutations from that of the invasive TFNP
population. Sequence analysis comparisons with
C. fiber as outgroup revealed 98 variable sites, 77
of them phylogenetically informative. Transitions of
sites 144 (A/G) and 147 (T/C) discriminated between
beaver species, as well as inversions at sites 258–259
and 265–267 occurring in the first 306 bp (GenBank
accession numbers C. canadensis AY793641-46,
AY793373-75-76, AF293348, AF155878 vs. C. fiber
AF155879, AJ389529).
Population relationships
The median-joining (MJ) network showed haplotype
CytbB as a central cluster connected mainly by short
branches to haplotypes CytbA and CytbC, while
haplotype CytbD (source population) formed a cluster
connected by relatively long branches to the rest
(Fig. 2a). Mean number of pairwise differences was
1.48 ± 0.94. Fu’s Fs test of selective neutrality
(original number of alleles k = 4) was Fs = 0.65;
Mean Theta (Pi) = 0.99 ± 0.47. Expected number of
alleles was 4.17. Pairwise divergence (p distance)
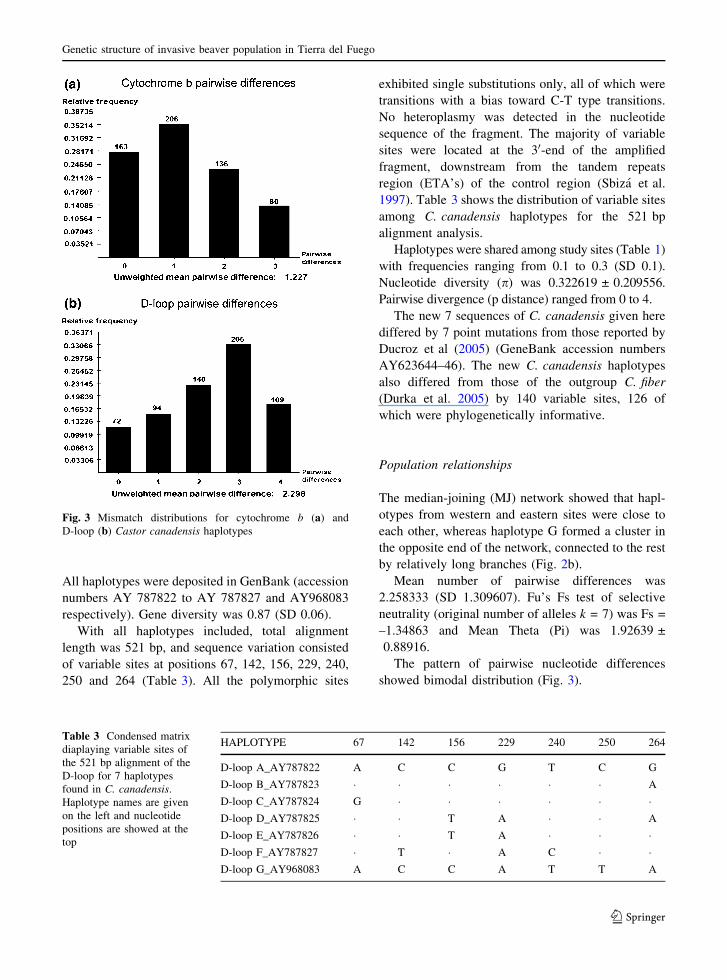
ranged from zero to 3 (Fig. 3). The pattern of
pairwise nucleotide differences showed a trimodal
distribution.
D-loop sequences
Sequence divergence
Six haplotypes were detected within D-loop
sequences (D-LoopA, D-LoopB, D-LoopC, DLoopD,
D-LoopE and D-LoopF) (Table 1). The amplified
fragment was 503 bp long. After alignment, the
haplotypes differed by 6 point mutations. The 5
C. canadensis sequences from the source population
differed by 7 point mutations and showed another
different haplotype (D-LoopG) that was markedly
longer (521 bp) than those from invasive population.
Table 2 Condensed matrix diaplaying variable sites of the
500 bp alignment of the Cytochrome b for 4 haplotypes found
in C. canadensis. Haplotype names are given on the left and
nucleotide positions are showed at the top
Haplotype 208 327 330
Cytb A_ AY793641 G G G
Cytb B_ AY793642 A � �Cytb C_ AY793643 A � A
Cytb D_ AY793644 A A A
Fig. 2 Median joining (MJ) network for C. canadensis (a)
Cytochrome b and (b) DLoop haplotypes (Table 1). Circle
areas are proportional to haplotype frequencies
M. S. Lizarralde et al.
123
All haplotypes were deposited in GenBank (accession
numbers AY 787822 to AY 787827 and AY968083
respectively). Gene diversity was 0.87 (SD 0.06).
With all haplotypes included, total alignment
length was 521 bp, and sequence variation consisted
of variable sites at positions 67, 142, 156, 229, 240,
250 and 264 (Table 3). All the polymorphic sites
exhibited single substitutions only, all of which were
transitions with a bias toward C-T type transitions.
No heteroplasmy was detected in the nucleotide
sequence of the fragment. The majority of variable
sites were located at the 30-end of the amplified
fragment, downstream from the tandem repeats
region (ETA’s) of the control region (Sbiza et al.
1997). Table 3 shows the distribution of variable sites
among C. canadensis haplotypes for the 521 bp
alignment analysis.
Haplotypes were shared among study sites (Table 1)
with frequencies ranging from 0.1 to 0.3 (SD 0.1).
Nucleotide diversity (p) was 0.322619 ± 0.209556.
Pairwise divergence (p distance) ranged from 0 to 4.
The new 7 sequences of C. canadensis given here
differed by 7 point mutations from those reported by
Ducroz et al (2005) (GeneBank accession numbers
AY623644–46). The new C. canadensis haplotypes
also differed from those of the outgroup C. fiber
(Durka et al. 2005) by 140 variable sites, 126 of
which were phylogenetically informative.
Population relationships
The median-joining (MJ) network showed that hapl-
otypes from western and eastern sites were close to
each other, whereas haplotype G formed a cluster in
the opposite end of the network, connected to the rest
by relatively long branches (Fig. 2b).
Mean number of pairwise differences was
2.258333 (SD 1.309607). Fu’s Fs test of selective
neutrality (original number of alleles k = 7) was Fs =
–1.34863 and Mean Theta (Pi) was 1.92639 ±
0.88916.
The pattern of pairwise nucleotide differences
showed bimodal distribution (Fig. 3).
Fig. 3 Mismatch distributions for cytochrome b (a) and
D-loop (b) Castor canadensis haplotypes
Table 3 Condensed matrix
diaplaying variable sites of
the 521 bp alignment of the
D-loop for 7 haplotypes
found in C. canadensis.
Haplotype names are given
on the left and nucleotide
positions are showed at the
top
HAPLOTYPE 67 142 156 229 240 250 264
D-loop A_AY787822 A C C G T C G
D-loop B_AY787823 � � � � � � A
D-loop C_AY787824 G � � � � � �D-loop D_AY787825 � � T A � � A
D-loop E_AY787826 � � T A � � �D-loop F_AY787827 � T � A C � �D-loop G_AY968083 A C C A T T A
Genetic structure of invasive beaver population in Tierra del Fuego
123

12 S rRNA Sequences
Sequence divergence
Mitochondrial 12S rRNA sequence data for 421 base
pairs were also available. A unique haplotype was
detected in all sampled specimens and deposited in
GeneBank (accession number AY787828) (Table 1).
This sequence was compared with GenBank
sequences (Accession numbers AY012111 and
U67297). Nucleotide sequence revealed 1 point
mutation (transition A/G) within the 421 bp.
Discussion
The exotic North American beaver C. canadensis had
adapted successfully to the sub-Antarctic ecosystems
as a geographically isolated invasive species. After
the initial introduction, beavers spread throughout the
entire Tierra del Fuego Archipelago and colonized all
the islands located between the Atlantic and Pacific
oceans; recently, they have also invaded new areas in
the Chilean sector of continental Patagonia. Their
rapid population growth, which has led to a current
population size of about 100,000 individuals from 25
founding pairs, indicates that the initial population
collapse did not have any drastic effects on the
viability or fertility of the individuals. Thus, this
exotic beaver population represents an informative
model for the study of genetic consequences of
invasion processes, including possible founder effects
caused by introduction of a small number of animals.
Although partial sequences of different mtDNA
markers have been previously reported for C. canad-
ensis in separate works (Nedhal et al. 1996; Murphy
et al. 2001; Ducroz et al. 2005), the present investi-
gation is the first combination of cytochrome b, 12S
rRNA and D-loop sequence analysis. The C. canad-
ensis populations examined in this study display high
genetic divergence at intrapopulation level.
Our data revealed the existence of several lineages
in the invasive beaver population. As a preliminary
result, the new mitochondrial DNA sequences sug-
gest clear differences between South and North
American beaver populations; in particular, the
cytochrome b transition at site 327 (Table 2) allowed
discrimination between invasive and source popula-
tions of C. canadensis. Transversions in the D-loop
or control region were not detected in our samples,
possibly because these are rare events at the popu-
lation level, although they may occur at a few
consecutive sites (Wood and Phua 1996; Douzery and
Randi 1997).
One of the most remarkable results of our study is
the genetic structuring detected within the invasive
beaver population, which could also have been
present in the source population. It may be hypoth-
esized that, after the expansion of the introduced
beavers and establishment of their current range, the
invasive population evolved according to the isola-
tion-by-distance model, as in the case of C. fiber
populations (Durka et al. 2005). This study revealed
new haplotypes different from previously reported
ones. These elements, together with the close simi-
larity of the new haplotypes within sites, suggest that
the pattern observed is a consequence of cessation of
gene flow following expansion of the founder 50-
individual population (founding type) at some point
from the time of introduction. Thus, disappearance of
geographically intermediate populations by isolation
would have produced the genetic structuring pattern
showed by C. canadensis in Tierra del Fuego.
Alternatively, this pattern could be the result of
genetic structuring that was already present in the
founder population.
Beavers disperse along the rivers and watershed
network of Isla Grande, mainly by using larger fast-
flowing rivers, and gene flow between local beaver
populations could be restricted by features of these
fluvial systems. For example, in the case of European
beavers the watersheds of Eurasian rivers could have
functioned as effective barriers to gene flow (Durka
et al. 2005).
Low DNA variation has been repeatedly reported
in taxa that have undergone severe bottlenecks or
founder effects. The European beaver C. fiber seems
to be one of the mammals with lowest variability
levels, according to studies of mitochondrial DNA
and major histocompatibility complex (MHC)
(Ducroz et al. 2005, Milishnikov and Saveljev
2001; Ellegren et al. 1993). Whereas Scandinavian
beavers exhibit extremely low variation of multilocus
minisatellite fingerprinting and complete monomor-
phism at MHC class I and II loci, the populations
from the European sector of Russia show substantial
polymorphism at minisatellite but not at MHC loci
(Ellegren et al. 1993).
M. S. Lizarralde et al.
123

Even though the depth of the split between the
South (Tierra del Fuego Archipelago) and North
American beaver populations could not be clearly
determined from these preliminary data, the inter-
ruption of gene flow between both populations clearly
suggests possible fixation of the divergent new
haplotypes after severe drift in the archipelago
population. The extreme genetic variation showed
by cytochrome b and especially D-loop haplotypes is
probably a consequence of migration and reinvasion
processes between Chilean and Argentinean popula-
tions. Similar genetic variability has been observed
between several European and Eurasian beaver
populations thought to have undergone extensive
bottlenecks or founder effects (Ellegren et al. 1993,
1994; Ducroz et al. 2005; Durka et al. 2005).
These results give rise to new questions. What is
the magnitude of the genetic drift that occurred in the
invasive beaver population of 25 mating pairs from
North America that expanded into a population of
probably more than 100,000 animals? Will they
successfully invade new areas of continental southern
Patagonia or will they remain restricted to Tierra del
Fuego Archipelago? Have invasive beavers under-
gone extensive physiological changes in the process
of adaptation to the Southern Cone? These and many
other questions remain unanswered, and further
comparative analyses of mtDNA variation across
the range of this species in the Argentinean and
Chilean sectors of Tierra del Fuego Archipelago will
be required to address them. Comparisons with North
American populations will be helpful to distinguish
founder effects generated at the time of introduction
from preexisting genetic structuring. DNA typing of
beavers from different source areas by means of PCR
analysis could be another useful approach to assess
the status of polymorphisms prior to the founder
effect. Furthermore, studies using nuclear markers,
such as microsatellites and genes with likely adaptive
significance, will help to assess the extent of genetic
variation and provide a complement to this dataset.
Acknowledgements This study was supported by a grant
from Consejo Nacional de Investigaciones Cientıficas y
Tecnicas, CONICET, Argentina (PIP 4306/96) to ML and a
Diversity Initiative Grant (2002–2003) from the University of
Minnesota to CG. Special thanks to Mrs. Virginia Haynes
(UMD) for technical assistance in the sequencing process. We
thank Julio Escobar and Guillermo Deferrari (CADIC-
CONICET) for participation and assistance in the collection
of material used in this study; all the staff at Tierra del Fuego
National Park (TFNP), and especially Dr. Thomas Hanley (US
Forest Service, Juneau, Alaska, USA) for providing tissue
samples of North American specimens and for his special
collaboration. We also thank Dr. Claudio Bravi (IMBICE) for
his advice and comments.
References
Anderson CB, Rozzi R, Torres-Mura JC, McGehee SM,
Sherriffs MF, Schuettler E, Rosemond AD (2006a) Exotic
vertebrate fauna in the remote and pristine sub-Antarctic
Cape Horn Archipelago region of Chile. Biodivers Con-
serv 10:3295–3313
Anderson C, Griffith C, Rosemond A, Rozzi R, Dollenz O
(2006b) The effects of Invasive North American beavers
on riparian plant communities in Cape Horn, Chile: do
exotic beavers engineer differently in sub-Antartic eco-
systems? Biol Conserv 128:467–474
Bandell HJ, Forster P, Rol A (1999) Median-Joining networks
for inferring intraspecific phylogenetics. Mol Ecol Evol
16:37–48
Bibb MJ, Van Etten RA, Wright CT, Walberg MW, Clayton
DA (1981) Sequence and gene organization of mouse
mitochondrial DNA. Cell 2:167–80
Brown G, Gadaleta G, Pepe C, Saccone C, Sbisa E (1986)
Structural conservation and variation in the D-loop con-
taining region of vertebrate mitochondrial DNA. J Mol
Biol 192:503–511
Brown AHD, Marshall DR (1986) Evolutionary changes
accompanying colonization in plants. In: Scudder GCE,
Reveal JL (eds) Evolution today. Carnegie-Mellon
University, Pittsburgh, PA, 351–363
Carson HL 1987 Colonization and speciation In: Gray AJ,
Crawley MJ, Edwards PJ (eds) Colonization, succession
and stability Oxford, Blackwell Scientific Publications
Coronato A, Escobar JM, Mallea C, Roig C, Lizarralde MS
(2003) Geomophorlogy characteristics of mountain rivers
colonized by Castor canadensis in Tierra del Fuego,
Argentina. Ecologıa Austral 13:15–26
Doyzery E, Randi E (1997) The mitochondrial control region
of cervidae: evolutionary patterns and phylogenetic con-
tent. Mol Biol Evol 14(11):1154–1166
Ducroz JF, Stubbe M, Saveljev AP, Heidecke D, Samjaa R,
Ulevicius A, Stubbe A, Durka W (2005) Genetic variation
and population structure of the Eurasian beaver Castorfiber in eastern Europe and Asia based on mtDNA
sequences. J Mammal 86:1059–1067
Durka W, Babik W, Ducroz JF, Heidecke D, Rosell F, Samjaa R,
Saveljev AP, Stubbe A, Ulevicius A, Stubbe M (2005)
Mitochondrial phylogeography of the Eurasian beaver
Castor fiber L. Mol Ecol 14:3843–3856
Ellegren H, Hartman G, Johansson M, Anderson L (1993)
Major histocompatibility complex monomorphism and
low levels of DNA fingerprinting variability in a reintro-
duced and rapidly expanding population of beavers. Proc
Natl Acad Sci USA 90:8150–8153
Ellegren H, Johansson M, Hartman G, Anderson L (1994)
DNA fingerprinting with the human 33.6 minisatellite
Genetic structure of invasive beaver population in Tierra del Fuego
123

probe identifies sex in beavers Castor fiber. Mol Ecol
3:273–274
Excoffier L, Laval G., Schneider S (2006) Arlequin ver. 3.01:
an integrated software package for population genetics
data analysis. Evol Bioinform Online 1:47–50
Excoffier L (2004) Patterns of DNA sequence diversity and
genetic structure after a range expansion: lessons from the
infinite-island model. Mole Ecol 13(4):853–864
Felsenstein J (2004) PHYLIP – phylogeny inference package
version 3.6. Department of Genetics, University of
Washington, Seattle
Fu Y-X (1997) Statistical tests of neutrality of mutations
against population growth, hitchhiking and backgroud
selection. Genetics 147:915–925
Jaksic FM, Iriarte JA, Jimenez JE, Martinez DR (2002)
Invaders without frontiers: cross-borders invasion of
exotic mammals. Biol Invasions 4:157–173
Jenkins SH, Busher PE (1979) Castor canadensis. Mamm
Species 120:1–8
Kotcher TD, Thomas WK, Meyer A, Edwards SV, Paabo S,
Villablanca FX, Wilson AC (1989) Dynamics of mito-
chondrial DNA evolution in animals: amplification and
sequencing with conserved primers. Proc Natl Acad Sci
USA 86:6196–6200
Kuehn R, Shwad G, Shroeder W, Rottmann O (2000) Differ-
entiation of Castor fiber and Castor Canadensis by
noninvasive molecular methods. Zoo Biol 19:511–515
Lizarralde MS (1993) Current status of the introduced beaver
population in Tierra del Fuego (Argentina). AMBIO
22(6):351-358
Lizarralde MS (2006) Control of invasive beaver in Archi-
pelago of Tierra del Fuego. Paper presented at the
International Workshop on the beaver problem in the
Southern Cone, Wildlife Conservation Society, Punta
Arenas, Chile, 6–8 December 2006
Lizarralde MS, Escobar JM (2000) Exotic mammals in Tierra
del Fuego, Argentina, Ciencia Hoy 10(56):52–63
Lizarralde MS, Deferrari GA, Alvarez SE, Escobar JM (1996)
Nutrient dynamic alterations induced by beaver (Castorcanadensis) on the southern beech forest (Nothofagus).
Austral Ecol 6:101–105
Lizarralde MS, Escobar JM, Deferrari GA (2004) Invader
species of Argentina: a review about beaver (Castorcanadensis) population situation on Tierra del Fuego
ecosystem. Interciencia 29(7):352–356
Martinez Pastur G, Lencinas V, Escobar J, Quiroga P,
Malmierca L, Lizarralde MS (2006) Understory succes-
sion in areas of Nothofagus forests in Tierra del Fuego
(Argentina) affected by Castor canadensis. J Appl Vegetat
Sci 9:143–154
Matson CW, Baker RJ (2001) DNA sequence variation in the
mitochondrial control region of red-backed voles
(Clethrionomys). Mol Biol Evol 18:1494–1501
Matsuhashi TR, Masuda R, Mano T, Yoshida MC (1999)
Microevolution of the mitochondrial DNA control region
in the Japanese brown bear (Ursus arctos) populations.
Mol Biol Evol 16:676–684
Montgelard C, Bentz S, Tirard C, Verneau O, Catzeflis FM
(2002) Molecular systematics of Sciurognathi (Rodentia):
the mitochondrial cytochrome b and 12S rRNA genes
support the Anomaluroidea (Pedetidae and Anomaluri-
dae). Mol Phylogenet Evol 22:220–233
Milishnikov AN, Saveljev AP (2001) Genetic divergence and
similarity of introduced populations of European beaver
(Castor fiber L.1758) from Kirov and Novosibinsk oblasts
of Russia. Russ J Genet 37:108–111
Moorman MC (2007) The conservation implications of inva-
sive beaver and trout on native freshwater fish in the Cape
Horn Biosphere Reserve, Chile. M.S. Thesis. Department
of Earth, Atmosphere and Marine Sciences, North Caro-
lina State University. Major Advisor: Dr. David Eggleston
Moorman MC, Anderson CB, Gutiirrez AG, Charlin R, Rozzi
R (2006) Watershed conservation and aquatic benthic
macroinvertebrate diversity in the Alberto D’Agostini
National Park, Tierra del Fuego, Chile. Anales del Insti-
tuto de la Patagonia 34:41–58
Murphy WJ, Eizirik E, Johnson WE, Zhang YP, Ryder OA,
OBrien SG (2001) Molecular phylogenetics and the ori-
gins of placental mammals. Nature 409(6820):614–618
Nedbal MA, Honeycutt RL, Schlitter DA (1996) Higher level
systematics of rodents (Mammalia, Rodentia): evidence
from the mitochondrial 12SrRNA gene. J Mamm Evol
3(3):201–237
Nei M (1987) Molecular evolutionary genetics. Columbia
University Press, New York, NY, USA
Nei M (1975) Molecular population genetics and evolution.
Elsevier Ed. New York
Reyes A, Nevo E, Saccone C (2003) DNA sequence variation
in the mitochondrial control region of subterranean mole
rats, Spalax ehrenbergi superspecies, in Israel. Mol Biol
Evol 20:622–632
Roughgarden J (1979) Theory of population genetics and
evolutionary ecology. Macmillan, New York
Saccone C, Attimonelli M, Sbisa E (1987) Structural elements
highly preserved during the evolution of the D-loop
containing region in vertebrate mitochondrial DNA. J Mol
Evol 26:205–211
Sakai AK, Allendorf FW, Holt JS, Lodge DM, Molfsky J, With
KA, Baughman S, Cabin RJ, Cohen JE, Ellstrand NC,
McCauley DE, O’Neil P, Parker IM, Thompson JN,
Weller SG (2001) The population biology of invasive
species. Annu Rev Ecol Syst 32:305–332
Skewes O, Gonzalez F, Olave R, Avila A, Vargas V, Paulsen P,
Kvnig HE (2006) Abundance and distribution of Ameri-
can beaver, Castor canadensis (Kuhl 1820), in Tierra del
Fuego and Navarino islands, Chile. Eur J Wildl Res
52:292–296
Sambrock J, Fritsch EF, Maniatis T (1989) Molecular cloning,
a laboratory manual. Cold Spring Harbor Laboratory
Press, New Cork
Sbiza E, Tanzariello F, Reyes A, Pesole G, Saccone C (1997)
Mammalian mitochondrial D-loop region structural analysis:
identification of new conserved sequences and their func-
tional and evolutionary implications. Gene 205:125–140
Smith MF, Patton JL (1993) Variation in mitochondrial cyto-
chrome b sequence in natural populations of South
American akodontine rodents (Muridae Sigmodontinae).
Mol Biol Evol 8(1):85–103
M. S. Lizarralde et al.
123

Tajima F (1993) Measurement of DNA polymorphism In:
Takahata N, Clark AG (eds) Mechanisms of molecular
evolution. introduction to molecular paleopopulation
biology, Japan Scientific Societies Press, Sinauer Asso-
ciates Inc., Tokyo, Sunderland, MA, pp 37–59
Thompson JD, Higgins DG, Gibson TJ (1994) Clustal W:
improving the sensitivity of progressive multiple sequence
alignment through sequence weighting, positions-specific
gap penalties and weight matrix choice. Nucleic Acids
Res 22:4673–4680
Vila C, Amorin IR, Leonard JA (1999) Mitochondrial DNA
phylogeography and population history of the grey Wolf
Canis lupus. Mol Ecol 8:2089–2103
Wood NJ, Phua SH (1996) Variation in the control region
sequence of the sheep mitochondrial genome. Anim Genet
27:25–33
Wright S (1955) Classification of the factors of evolution. Cold
Spring Harb Symp Quant Biol 20:16–24
Genetic structure of invasive beaver population in Tierra del Fuego
123
Copyright © 2022 FDOKUMEN




















