Agonistic sounds signal male quality in the Lusitanian toadfish
8
Dear Author, Please, note that changes made to the HTML content will be added to the article before publication, but are not reflected in this PDF. Note also that this file should not be used for submitting corrections.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Agonistic sounds signal male quality in the Lusitanian toadfish

Dear Author, Please, note that changes made to the HTML content will be added to the article before publication, but are not reflected in this PDF. Note also that this file should not be used for submitting corrections.

UNCO
RRECTED P
RO
OF
1Q1 Agonistic sounds signal male quality in the Lusitanian toadfish
2Q2 M. Clara P. Amorim a,⁎, Carlotta Conti a, Teresa Modesto b, Amparo Gonçalves c, Paulo J. Fonseca d
3 a MARE — Marine and Environmental Sciences Centre, ISPA— Instituto Universitário, Lisbon, Portugal4 b Centro de Ciências do Mar, Universidade do Algarve, Campus de Gambelas, 8000-810 Faro, Portugal5 c Division of Aquaculture and Upgrading, Portuguese Institute for the Sea and Atmosphere, I.P. (IPMA, I.P.), Lisbon, Portugal6 d Departamento de Biologia Animal and cE3c — Centre for Ecology, Evolution and Environmental Changes, Faculdade de Ciências, Universidade de Lisboa, Lisbon, Portugal
7
9 H I G H L I G H T S
10
11 • Acoustic communication during agonis-12 tic behaviour is widespread in fishes.13 • Breeding Lusitanian toadfish males de-14 fend nests from intruders with sounds.15 • Low fundamental frequency (high16 mean pulse period) reflected high an-17 drogen levels.18 • The dominant harmonic frequency de-19 creased with sonic muscle lipid content.20 • Spectral content of fish sounds signal21 male traits that are key to fight out-22 come.23
2 4 G R A P H I C A L A B S T R A C T
25
26
2728
30303132
3334
3 5
a b s t r a c t3 6 a r t i c l e i n f o
37 Article history:38 Received 26 February 201539 Received in revised form 27 May 201540 Accepted 1 June 201541 Available online xxxx
42 Keywords:43 Acoustic communication44 Fish45 Temporal cues46 Spectral cues47 Steroid hormones48 Territorial defence
49Acoustic communication during agonistic behaviour is widespread in fishes. Yet, compared to other taxa, little is50knownon the information content of fish agonistic calls and their effect on territorial defence. Lusitanian toadfish51males (Halobatrachus didactylus) are highly territorial during the breeding season and use sounds (boatwhistles,52BW) to defend nests from intruders. BW present most energy in either the fundamental frequency, set by the53contraction rate of the sonic muscles attached to the swimbladder, or in the harmonics, which are multiples of54the fundamental frequency. Here we investigated if temporal and spectral features of BW produced during terri-55torial defence reflect aspects of male quality that may be important in resolving disputes. We found that higher56mean pulse period (i.e. lower fundamental frequency) reflected higher levels of 11-ketotestosterone (11KT), the57main teleost androgen which, in turn, was significantly relatedwithmale condition (relative bodymass and gly-58cogen content). BW dominant harmonic mean and variability decreased with sonic muscle lipid content. We59found no association between BWduration andmale quality. Taken together, these results suggest that the spec-60tral content of fish agonistic sounds may signal male features that are key in fight outcome.
61 © 2015 Published by Elsevier Inc.
6263
64
65
661. Introduction
67Animals' interactions, such as conflicts over limited resources, are68mediated by the exchange of signals that provide information for adap-69tive decisionmaking [1]. Territorial defence is often a key component of70an individual's fitness and includes both signalling territorial ownership
Physiology & Behavior xxx (2015) xxx–xxx
⁎ Corresponding author.E-mail addresses: [email protected] (M.C.P. Amorim), [email protected]
(C. Conti), [email protected] (T. Modesto), [email protected] (A. Gonçalves),[email protected] (P.J. Fonseca).
PHB-10895; No of Pages 7
http://dx.doi.org/10.1016/j.physbeh.2015.06.0020031-9384/© 2015 Published by Elsevier Inc.
Contents lists available at ScienceDirect
Physiology & Behavior
j ourna l homepage: www.e lsev ie r .com/ locate /phb
Please cite this article as: M.C.P. Amorim, et al., Agonistic sounds signal male quality in the Lusitanian toadfish, Physiol Behav (2015), http://dx.doi.org/10.1016/j.physbeh.2015.06.002

UNCO
RRECTED P
RO
OF
71 and chasing away intruders. In this context, signals should provide72 species/sex-specific information to inform potential intruders of the73 owner's presence on site, and also provide information on fighting abil-74 ity and motivation for territorial confrontations [1]. Hence, information75 content of territorial signals is expected to depend on whether a signal76 serves only for territorial ownership advertisement, to reveal the77 sender's quality and motivation, or both [1].78 Multiple studies show that vocalizations can influence the resolution79 of contests as they reflect the sender's body size or condition (e.g.80 [2–4]). For example, low frequency vocalizations may give a reliable81 indication of a large body size and deter potential attackers [2] or, in the82 case of humans, influence the perception of leadership capacity [5]. In83 fish, the largest group of vertebrates exhibiting widespread evolution84 of sound production [6], acoustic signals are often produced during con-85 tests, including territorial defence [7]. In this taxon there is evidence for86 salient information content in the temporal pattern of sound pulses [6].87 In addition, there is a strong selection pressure favouring complexity in88 the spectro-temporal content of vocal signals [8]. Surprisingly, little is89 known on whether temporal and spectral features of agonistic calls re-90 flect the sender's quality and affect territorial defence (e.g. [7,9,10]). Im-91 portantly, while it has been shown that in somefishes larger individuals92 produce longer, louder and lower frequency agonistic sounds [11–14],93 few studies have provided an integrative view on how agonistic sounds94 reflect multiple aspects of male quality such as physical and physiologic95 features.96 Fishes from the Batrachoididae family have become a key97 neuroethological model for studying acoustic communication in verte-98 brates because mate attraction and territorial defence in theses fishes99 rely heavily on acoustic signalling [15–17]. While mating advertisement100 calls (boatwhistles and hums) produced by batrachoidid nest-holders101 have been also implied in signalling territorial ownership and in spacing102 out individuals [9,16,18], grunts are considered themain agonistic call [8,103 15]. Within this family, Lusitanian toadfish (Halobatrachus didactylus)104 males mainly defend their nests with agonistic boatwhistles (BW) that105 are similar to mating advertisement BW except for presenting lower106 dominant frequencies and weaker amplitude modulation [16]. As BW107 have shown to be rather complex signals [8] they render the opportunity108 to assess the salience of relevant sound features for territorial defence in109 teleosts. Typically, BWare relatively longmultiharmonic signalswherein110 the fundamental frequency is determined by the firing rate of the vocal–111 acoustic neural network that drives the contraction rate of the paired112 sonic muscles attached to the swimbladder [10].113 Here we relate spectral and temporal features of the agonistic BW114 with Lusitanian toadfish male quality, including biometric, condition,115 and physiological features.We predict that BWacoustic features convey116 information onmale quality including energetic reserves and hormonal117 status which influence the outcome of contest behaviour [19].
118 2. Methods
119 2.1. Subject males and maintenance
120 The Lusitanian toadfish, like other batrachoidids, has two fixedmale121 reproductive phenotypes that differ in a suite ofmorphological andneu-122 roendocrine traits [10,20]. Type I males nest under rocks or in crevices,123 are territorial, vocalize to attract mates and provide parental care to the124 eggs in the nest. In contrast, Type II males are not territorial and sneak125 fertilizations [20].126 We captured territorial males that spontaneously occupied artificial127 concrete shelters placed in the lower intertidal of the Tagus River es-128 tuary (Military Air Force Base 6, Montijo, Portugal; 38° 42′N; 8° 58′W).129 Fish use these shelters as nests during the breeding season that spans130 from May to July in Portugal (e.g. [17]). We used plastic round131 swimming-pools (2.5 m diameter, 0.5 m water depth) as stock and ex-132 perimental tanks. Tanks were placed on the sand just above the high133 tide shoreline, near the collection area and under a shadow net cover
134held 170 cm high to prevent excessive solar radiation and water135heating. Water temperature varied in all tanks from 18 to 26 °C136(mean = 21.4 °C), within the range of the estuary water temperature137variation during the same period. Tank water was renewed every1382–3 days, by pumping directly from the estuary. A natural light cycle139was maintained as tanks were outdoors.
1402.2. Territorial experiments
141We carried out territorial intrusion experiments following142Vasconcelos et al Q3. [10]. Briefly, 24 h before trials two subject males143were placed in an experimental tank provided with two roof tiles (in-144ternal dimensions 44 cm × 18 cm × 10 cm) placed 50 cm apart and14520 cm away from the tank's border. In each 1 h trial, two intruder146males were placed sequentially in the experimental tank, one at the147start of the trial and the second 30 min after, both remaining in the148tank until the end of the trial (Supplementary material, Video 1). This149experimental design resembles the natural chorusing aggregations,150where territorial males nest close together [21] and may sequentially151attract competitor males [17]. Visual and acoustic behaviours were152monitored during trials and noted in detail on paper. Fish weremarked153with small cuts between the fin rays to ensure their identity. Marking154did not cause any measurable change in the fish behaviour. Fish were155never used in more than one trial.156We placed one hydrophone (High Tech 94 SSQ, High Tech Inc.,157Gulfport, MS, USA; frequency response: 30 Hz to 6 kHz ± 1 dB; voltage158sensitivity: −165 dB re. 1 V/μPa) in front of each nest, at about 10 cm159from its entrance and from the tank bottom, attached to a wooden rod160kept over the tank. Simultaneous two channel recordings were made161with a USB audio capture device (Edirol UA-25, Roland, Osaka, Japan;16216 bit, 44.1 kHz acquisition rate per channel) connected to a laptop163and down-sampled to 6 kHz by Adobe Audition 3.0 (Adobe Systems,164San José, CA, USA). Recorded sounds could be attributed to a particular165territorial male (henceforth named nest-holder) due to the close prox-166imity of each hydrophone to one nest. Also, in the course of similar ex-167periments (e.g. [16]) we have observed that only nest-holders produce168sounds. In one exception (unpublished data), the intruder entered the169nest and also made BW but these could clearly be distinguished from170the nest-holder's sounds due to spectral differences.171At the end of the 19 trials, residents (n = 38) and intruders (n =17234) that engaged in social interactions were measured to the nearest173mm for total length (TL) and to the nearest g for total bodymass (M).174Note that in four trials one of the intruders did not interact with nest-175holders. Residents measured mean ± sd (range) 42.0 ± 3.0 (37.5–49.5)176cm in TL and intruders 36.9 ± 2.7 (30.0–42.0) cm in TL. Residents pro-177duced on average 4.7 ± 5.9 (range: 0–22, mode = 1) BW during trials.178For this studywe only considered residentmales thatmade at least 4179BW (n = 14; note that one male only had 2 BW suitable for analysis).180These nest-holders made an average of 10.4 ± 6.4 (4–22) BW during181a 1 h trial. All resident males were not used for sound analysis and the182intruders were returned to the estuary after trials. Territorial males183that made fewer than 4 BW did not differ in TL from more vocal males184(t test, p N 0.05). They likely made fewer sounds because they experi-185enced fewer interactions with intruders than the males that were186more vocal (t = 3.34, d.f. = 36, p b 0.01, mean no. of interactions =1874.9 vs. 2.8 for more vocal vs. less vocal fish).188The 14 males used to relate sounds with male quality averaged18941.9±1.8 (39.3–46.0) cm in TL and 1221±199 (970–1600) g inM. Im-190mediately after the end of trials, thesemales were sacrificedwith an ex-191cessive dosage of MS222 (tricaine methane sulphonate; Pharmaq,192Norway) buffered (1:1) with sodium bicarbonate. Blood samples were193collected from the caudal vein in heparinized syringes within 4 min194from first handling of the fish. Plasma samples were subsequently sep-195arated by centrifugation (6000 rpm for 5 min) on site and stored on196ice until taken to the lab, where theywere stored at−20 °C until steroid197quantification. Following blood sampling, males were immediately
2 M.C.P. Amorim et al. / Physiology & Behavior xxx (2015) xxx–xxx
Please cite this article as: M.C.P. Amorim, et al., Agonistic sounds signal male quality in the Lusitanian toadfish, Physiol Behav (2015), http://dx.doi.org/10.1016/j.physbeh.2015.06.002

UNCO
RRECTED P
RO
OF
198 dissected and the mass of the gonads (MG), the liver (ML), and of the199 sonic muscles (MSM) was weighed to the nearest mg, and eviscerated200 body mass to the nearest g (ME). A sample of body muscle (epaxial201 muscle fibres) was taken. Samples of body and sonic muscles were im-202 mediately placed in ice and in dry ice until stored in the lab at −20 °C203 and−80 °C for subsequent quantification of the lipid and glycogen con-204 tent, respectively. Lipids and glycogen are important sources of energy205 in fish and are often used as a direct measure of body condition [22].206 They are also important metabolic substrates of sonic muscle activity,207 i.e. sound production (e.g. [23]).
208 2.3. Lipid and glycogen quantification
209 Lipid content was measured following Olsson et al. [24]. Muscle210 samples were desiccated at 60 °C for 24 h and were weighed on a211 scale (Sartorius RC210D, Göttingen, Germany) to the nearest 0.01 mg.212 Lipids were extracted in 100 ml of petroleum ether (Sigma-Aldrich, St.213 Louis, MO, USA) for 8 h. Relative body lipid content of each male was214 measured as the difference in dry weight before and after lipid extrac-215 tion expressed as g/100 g tissue dry weight.216 Glycogen content was determined according to the method de-217 scribed by Viles and Silverman [25]. Dry samples (25 mg) were218 boiled with 15 ml of 33% potassium hydroxide for 15 min. After219 cooling, 0.5mlwas taken and 50 μl of a saturated sodium sulphate solu-220 tion and 2ml of ethanolwere added. Sampleswere placed in an ice bath221 for 30 min for glycogen precipitation. Following centrifugation the pre-222 cipitate was dissolved in 0.5 ml of distilled water again precipitated223 with 1 ml of ethanol (for 30 min) and centrifuged. The precipitate was224 re-dissolved in 0.4 ml of distilled water and 3 ml of anthrone-reagent225 was added (prepared with concentrated sulphuric acid, water and226 anthrone — Merck 101468). This mixture was heated at 90 °C for227 20 min. The absorbance was then measured at 620 nm. A calibration228 curve was prepared using glycogen type II from oyster (Sigma G8751)229 as a standard. The results were expressed as μg/mg tissue dry weight.
230 2.4. Hormone quantification
231 Plasma levels of testosterone (T), 11 keto-testosterone (11KT) and232 cortisol (CORT) were measured by radioimmunoassay (RIA). 11KT and233 CORT are respectively the predominant androgen and corticosteroid in234 teleost fish, including the Lusitanian toadfish [20]. Previously to the as-235 says, 100 μl plasma samples were extracted for free and conjugated ste-236 roids using the techniques described in Scott and Canário [26] and237 Damasceno-Oliveira et al. [27]. RIA methodology and cross reactions238 of antisera for T, 11KT and CORT were described in Vasconcelos et al.239 [17]. The results of the different fractions (free and conjugated) were240 summed. Intra-assay and inter-assay precision (coefficient of variation)241 was6.3% and 5.2% for T, 3.9% and 5.5% for 11KT, 7.8% and 10.3% for CORT,242 respectively.
243 2.5. Behavioural and sound analyses
244 We tallied the number of agonistic resident–intruder interactions,245 which started with the intruder's approach (when the intruder reached246 within a body length from the nest) and partial or total intrusions in the247 nest (hereafter named intrusions). For resident fish we tallied the fre-248 quency of agonistic escalated behaviours (escalated fight, EF) such as249 chasing, bite attempts, bites and mouth–mouth fight. Non-escalated vi-250 sual behaviour such as approach or mouth opening with the extension251 of fins was not observed. We also considered the number of emitted252 BW by the resident.253 Acoustic analysis was performed using Adobe Audition 3.0 and254 Raven 1.2 forWindows (Bioacoustics Research Program, Cornell Lab-255 oratory of Ornithology, Ithaca, NY, USA). We measured BW duration256 (Dur, ms), from the start of the first pulse to the end of the last pulse,257 and pulse period (PP) of the BW tonal phase [cf. 28] (ms, average
258peak-to-peak interval of six consecutive pulses in the middle of this259segment— Fig. 1a). We calculated themean and coefficient of variation260(CV, CV=SD /mean) for the BWduration and pulse period per individ-261ual. Because the dominant frequency (frequency with the highest ener-262gy component in the BW tonal phase) can be in either the fundamental263frequency (H0) or one of its harmonics (H1 to H3) [28], we further cal-264culated the most common dominant harmonic for each fish (i.e. the265mode).We then averaged the observed frequency values for the sounds266that presented that harmonic as the dominant frequency, hereafter267named dominant harmonic (DH,Hz; Fig. 1a). As ameasure of its variabil-268ity we used the ratio between the number of sounds that presented the269DHby the total number of sounds. This ratio indicates if amale produces270BW with the dominant frequency always in one particular harmonic271(DH = 1) or in different harmonics (DH b 1). Temporal patterns were272measured from the oscillogram and frequency measures from the273power spectrum (sampling frequency 8 kHz, Hamming window, filter274bandwidth 10 Hz). We analysed an average ± sd (range) of 7.3 ± 2.5275(4–10) BW per male.
2762.6. Statistical analyses
277We investigated whether the number of BW or the number of EF278made bynest-holderswas correlatedwith the number of intruder inter-279actions (only intrusions in the case of EF) and with the nest-holder's280quality (see below) with Pearson correlation. We tested if BW acoustic281parameter (Dur mean, Dur CV, PP mean, PP CV, DH, DH variability)282reflected nest-holder's features with linear regression analyses, exclud-283ing the male for which only 2 BW were analysed. Dependent variable,284i.e. acoustic parameters were not correlated (Pearson correlation,285p N 0.05). We log10-transformed the dominant harmonic mean and var-286iability to meet the assumption of normality.287Weconsidered the following descriptors for resident quality: steroid288levels (T, 11KT, CORT), lipid content of somatic and sonic muscles289(LipidM and LipidSM, respectively), glycogen content of somatic and290sonic muscles (GlycM and GlycSM, respectively), and total length291(log10TL). We also used the residuals of the simple linear regression of292sonic muscle, gonads and liver mass on eviscerated body mass (RMSM,293RMG, RML, respectively) as metrics of these parameters controlled for294the influence of body size. For example a high positive residual of295RMSM indicates a heavier than average sonic mass for a given body296size. In addition, we used the residuals of ME on TL (COND) as a metric297of body condition. 11KT, GlycM and COND were positively correlated298with each other (Pearson correlation, r=0.75–0.77, p b 0.01). LipidSM299was also related with RML (r= 0.57, p b 0.05). The remaining male de-300scriptors were unrelated. To avoid multicollinearity and to reduce the301number of explanatory variables we excluded GlycM, COND and RML
302from analyses. We further excluded T as it is physiologically related303with 11KT [20].304Our final regressionmodels complied with all assumptions of multi-305ple linear regression. All model residuals were normally distributed.306Further residual analyses were performed using Durbin–Watson statis-307tics, residual plots as well as multicollinearity tests (variance inflation308factors, VIF). All statistical analyseswere performed using SPSS forWin-309dows (20.0, SPSS Inc., Chicago, IL, USA).
3103. Results
311Nest-holders experienced one to nine interactions with intruders312(mean ± SD = 4.6 ± 2.2), from which 46.2% were approaches, 4.6%313were approaches followed by intrusions, and 49.2% were direct partial314or total intrusions. None experienced a nest takeover. Nest-holders315only responded to intruders' approaches with BW (Fig. 1b). During in-316trusions nest-holders reacted with BW or with BW followed by EF317(when the intruder would not flee upon a BW). In a few occasions, sub-318ject males did not apparently react, i.e. did not show a visible change in319visual or acoustic behaviour, or immediately escalated their behaviour
3M.C.P. Amorim et al. / Physiology & Behavior xxx (2015) xxx–xxx
Please cite this article as: M.C.P. Amorim, et al., Agonistic sounds signal male quality in the Lusitanian toadfish, Physiol Behav (2015), http://dx.doi.org/10.1016/j.physbeh.2015.06.002
UNCO
RRECTED P
RO
OF
320 (Fig. 1b). The number of BWwas correlatedwith the number of interac-321 tions (r= 0.61, p b 0.05) but not with male features. The number of EF322 was in turn only correlated with own body glycogen levels (r=−0.56,323 p b 0.05).324 Agonistic BW Dur averaged 640 ms (range 437–933 ms) and pre-325 sented a relatively high coefficient of variation (mean = 0.3, range326 0.1–0.5). PP averaged 23ms (range 19.7–26.9ms) andwasmore stereo-327 typed (mean CV = 0.05, range 0.01–0.07). DH was most commonly328 found on the first harmonic (DH mean = 89 Hz, range 41–146 Hz),329 and on average males maintained a particular dominant frequency330 band in 70% of BW, i.e. DH variability averaged 0.7 (range 0.5–1.0).
331Only three out of 13 males produced BW always with the same domi-332nant frequency, i.e. with DH variability = 1.333We asked whether these acoustic parameters reflect male quality.334Both Dur mean and Dur CV were not significantly related with any335male quality variable (regression analyses, p N 0.05). However, several336male quality variables were significantly associated with pulse period337and spectral characteristics of BW. PP mean increased with 11KT338(R2 = 0.63) and decreased with relative gonad mass (RMG: R2 = 0.22,339Table 1, Fig. 2a,b). PP CV was negatively affected by CORT (R2 = 0.37;340Fig. 2c). DH mean was negatively associated with LipidSM (R2 = 0.35,341Table 1, Fig. 3a). DH variability decreased with LipidSM (R2 = 0.52)
Rel
.am
plitu
de (
dB)
0.4 0.60.20
Frequency (kHz)
-40
-20
0
DHSound duration
6 PP
100 ms
(a)
72.7
27.3
Approach
45.7
42.9
5.7 5.7
IntrusionBW
BW+EF
EF
NR
(b)
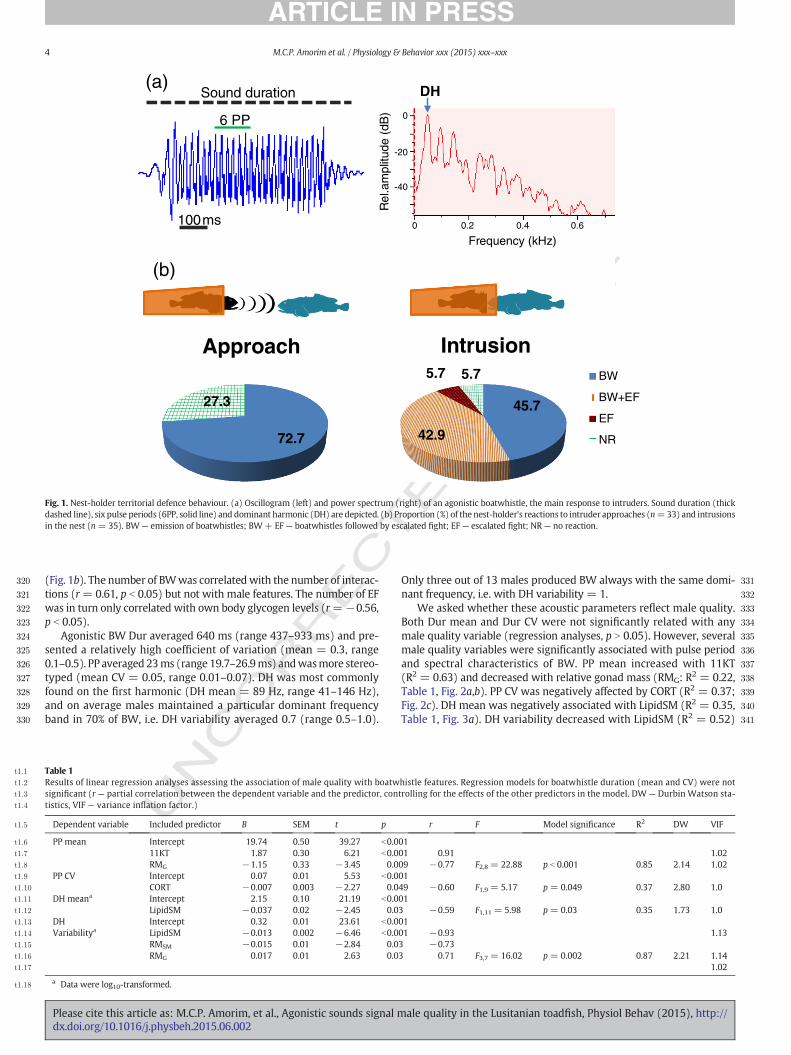
Fig. 1. Nest-holder territorial defence behaviour. (a) Oscillogram (left) and power spectrum (right) of an agonistic boatwhistle, the main response to intruders. Sound duration (thickdashed line), six pulse periods (6PP, solid line) and dominant harmonic (DH) are depicted. (b) Proportion (%) of the nest-holder's reactions to intruder approaches (n=33) and intrusionsin the nest (n = 35). BW— emission of boatwhistles; BW+ EF — boatwhistles followed by escalated fight; EF — escalated fight; NR— no reaction.
t1:1 Table 1t1:2 Results of linear regression analyses assessing the association of male quality with boatwhistle features. Regression models for boatwhistle duration (mean and CV) were nott1:3 significant (r — partial correlation between the dependent variable and the predictor, controlling for the effects of the other predictors in the model. DW — Durbin Watson sta-t1:4 tistics, VIF — variance inflation factor.)
t1:5 Dependent variable Included predictor B SEM t p r F Model significance R2 DW VIF
t1:6 PP mean Intercept 19.74 0.50 39.27 b0.001t1:7 11KT 1.87 0.30 6.21 b0.001 0.91 1.02t1:8 RMG −1.15 0.33 −3.45 0.009 −0.77 F2,8 = 22.88 p b 0.001 0.85 2.14 1.02t1:9 PP CV Intercept 0.07 0.01 5.53 b0.001t1:10 CORT −0.007 0.003 −2.27 0.049 −0.60 F1,9 = 5.17 p = 0.049 0.37 2.80 1.0t1:11 DH meana Intercept 2.15 0.10 21.19 b0.001t1:12 LipidSM −0.037 0.02 −2.45 0.03 −0.59 F1,11 = 5.98 p = 0.03 0.35 1.73 1.0t1:13 DH Intercept 0.32 0.01 23.61 b0.001t1:14 Variabilitya LipidSM −0.013 0.002 −6.46 b0.001 −0.93 1.13t1:15 RMSM −0.015 0.01 −2.84 0.03 −0.73t1:16 RMG 0.017 0.01 2.63 0.03 0.71 F3,7 = 16.02 p = 0.002 0.87 2.21 1.14t1:17 1.02
t1:18 a Data were log10-transformed.
4 M.C.P. Amorim et al. / Physiology & Behavior xxx (2015) xxx–xxx
Please cite this article as: M.C.P. Amorim, et al., Agonistic sounds signal male quality in the Lusitanian toadfish, Physiol Behav (2015), http://dx.doi.org/10.1016/j.physbeh.2015.06.002

UNCO
RRECTED P
RO
OF
342 andwith RMSM (R2=0.18), but increasedwith RMG (R2=0.13, Table 1,343 Fig. 3b–d).
344 4. Discussion
345 The Lusitanian toadfish used BW as the primary response to in-346 truders, both during approaches and while facing actual intrusions in347 the nest (Fig. 1). Nest-holders typically only resorted to escalated fight348 when opponents did not flee after first receiving BW, and only after349 the intruder entered the nest. These results are consistent with BW sig-350 nalling territorial ownership and serving as a warning of a possible at-351 tack. Congruently, BW have been suggested as deterrent signals in352 territorial intrusions [16]. In addition, BWmay also function in resource353 holding potential assessment, if acoustic features relate to fighting abil-354 ity [19].355 We found that circulating steroid hormones were associated with356 pulse period mean and CV. Males with elevated levels of 11KT pro-357 duced BWwith higher mean pulse period, i.e. with lower fundamen-358 tal frequency [4]. In turn, 11KT was positively correlated with male359 condition (relative body mass and glycogen content). Males with360 higher CORT levels produced BWwith less variable fundamental fre-361 quencies (lower PP CV). Our results are consistent with the well-362 known role of sex and stress steroids as regulators of acoustic signals363 used during social behaviour [10,29]. Steroids not only shape vocal364 control circuits in fish but also modulate vocal motor patterning365 through rapid non-genomic effects on the activity of the hindbrain366 vocal central pattern generator [30]. Such effects have been exten-367 sively shown in other batrachoidids. In the plainfin midshipman368 fish, Porichthys notatus, intramuscular injections of 11KT and CORT369 rapidly increase the durations of fictive calls but do not affect funda-370 mental frequency [31]. 11KT and CORT administered exogenously
371via implanted food items, also increase calling rate of mating calls372in territorial Gulf toadfish, Opsanus beta [32]. While our study points373to different effects of 11KT and CORT on BW features they are consis-374tent with steroids modulating the activity of the central mechanisms375of sound production. Indeed, the vocal central pattern generator of376batrachoidids [30,33] expresses abundant androgenmRNA receptors377[29] and plays an essential role in setting the duration and pulse rep-378etition rate of calls [34].379Despite the trade-off with, e.g. the immune system, elevated levels380of steroids are also socially adaptive since they regulate fight or flight re-381sponses and the allocation of energy resources [35,36]. 11KT levels are382usually elevated in territorial batrachoidid males in the pre-nesting pe-383riod and in the first half of the breeding season, encompassing territorial384establishment, courtship and spawning (e.g. [20,37]) In these periods,385social challengesmodulate the release of sex or stress steroid hormones,386while the release of these hormones in turn facilitates adequate re-387sponses to changes in social environment. This reciprocal interaction388between steroids and behaviour is known as the challenge hypothesis389[38]. Consequently, having fluctuations of circulating steroid levels390encoded in the fundamental frequency of the toadfish agonistic391boatwhistle is likely relevant in opponent assessment, as the intruders392may obtain valuable information on the nest-holder's agonistic motiva-393tion, energetic availability and reproductive status.394Our present results further suggest that dominant harmonic395mean and variability signal sonic muscle condition (relative mass396and mainly lipid content). We found that males with higher lipid397levels and relative mass made BW with lower dominant harmonics.398In addition, males with greater lipid content in their sonic muscles399presented a lower variability in the BW dominant harmonic. That400is, they tended to present the maximum spectral energy in the401same harmonic. Sonic muscle features are androgen dependent.
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0
11-ketotestosterone (ng/ml)
18
20
22
24
26
28
Pul
se p
erio
d m
ean
(ms)
-3 -2 -1 0 1 2
RMG
20
22
24
26
28
Pul
se p
erio
d m
ean
(ms)
0 1 2 3 4 5 6 7
Cortisol (ng/ml)
0.00
0.02
0.04
0.06
0.08
Pul
se p
erio
d C
V
(a) (b)
(c)
R2 = 0.63 R2 = 0.22
R2 = 0.37
Fig. 2. Relation between pulse periodmeanwith (a) circulating 11-ketotestosterone and (b) relative gonadmass (RMG). Relation between pulse period variability (coefficient of variation,CV) with circulating cortisol (c). Lines of univariate regressions and 95% confidence interval bands are shown.
5M.C.P. Amorim et al. / Physiology & Behavior xxx (2015) xxx–xxx
Please cite this article as: M.C.P. Amorim, et al., Agonistic sounds signal male quality in the Lusitanian toadfish, Physiol Behav (2015), http://dx.doi.org/10.1016/j.physbeh.2015.06.002

UNCO
RRECTED P
RO
OF
402 Sonic muscle mass, morphological and physiological traits adapted403 for rapid contraction are driven seasonally by androgens in404 batrachoidids and other vocal fish, paralleling the increased vocal ac-405 tivity in the breeding season (e.g. [39–42]). For example, androgens406 drive an increase of mitochondria-filled sarcoplasm in sonic muscles407 necessary for sustained aerobic activity of muscle contraction [39]408 that is key to reproductive success [17]. The effect of steroids on409 sonic muscle function also depends on the expression of hormone410 receptors and enzymes involved in steroid signalling pathways,411 which in the plainfin midshipman diverge between calling and412 non-calling type I males [43]. Lipids, likely reflecting previous forag-413 ing success and health status, are one of the major metabolic sub-414 strates of sonic muscles during prolonged aerobic contraction and415 can noticeably decrease during the period of maximal vocal activity416 [40]. In this context, low and stable BW dominant harmonics are in-417 formative of androgen driven traits, such as sonic muscle mass, and418 likely of the individuals foraging competitive ability. Other hypothe-419 ses should also be taken into consideration. For example, harmonic420 variability could be associated with the lipid levels in sonic muscles.421 As steroids are lipophilic hormones, high lipid levels in muscle fibres422 could provide a ‘sink’ for 11KT [44] and enhance the aforementioned423 morphological and physiological traits for sustained rapid contrac-424 tion. Alternatively, harmonic variability could be an epiphenomenon425 related to intrinsic muscle contraction variability unrelated with426 male quality.427 In summary, agonistic BW not only function to fend off potential428 nest intruders but also potentially convey relevant information for429 male assessment. Agonistic BW with low and stable fundamental430 and dominant frequencies signal elevated steroid levels that mediate431 social interactions and trigger high sonic muscle condition required
432for sustained sound production. Low sound frequency typically sig-433nals male quality even when not related with body size [5]. An inter-434esting result in the present study is the information associated with435BW frequency stability. Signal stability may increase redundancy in436information and therefore communication efficacy [1]. The present437work also suggests high physiological condition associated with438low frequency stereotyped signals as theseweremade by individuals439with better body and sonic muscle condition and high steroid levels.440Recent studies have pointed to the existence of non-linear acous-441tic components in batrachoidid BW [8,45]. For example, similar to442the Lusitanian toadfish, the oyster toadfish BW start with a broad-443frequency non-harmonic grunt-like component [28,45,46]. One of444the proposed mechanisms underlying this chaotic initial grunt445phase of the BW is the arrhythmic (but in-phase) recruitment of446the bilateral sonic muscles encoded by the vocal central neural447network [45]. This raises the hypothesis that the stability of the fun-448damental and dominant frequencies of BW is also advertising preci-449sion in the neural circuitry underlying the vocal pattern generator,450which may be dependent on development issues. Future studies451should address the role of vocal stability vs. plasticity in vocal inter-452actions and whether such fine variability in spectral parameters of453fish calls can be perceived by fish.454Supplementary data to this article can be found online at http://dx.455doi.org/10.1016/j.physbeh.2015.06.002.
456Acknowledgements
457We thank the Air Force Base No. 6 of Montijo (Portugal) for allowing458this study in their military establishment. We are grateful to André459Alves and Daniel Alves for the help with the field work. Fish drawings
0 2 4 6 8 10 12 14
LipidSM (g)
1.6
1.8
2.0
2.2
Log1
0-D
H (
Hz)
0 2 4 6 8 10 12 14
LipidSM (g)
0.15
0.20
0.25
0.30
0.35
Log 1
0-D
H v
aria
bilit
y
-3 -2 -1 0 1 2
RMSM
0.15
0.20
0.25
0.30
0.35
Log 1
0-D
H v
aria
bilit
y
-3 -2 -1 0 1 2
RMG
0.15
0.20
0.25
0.30
0.35
Log 1
0-D
H v
aria
bilit
y
(a) (b)
(c) (d)
R2 = 0.35 R2 = 0.52
R2 = 0.18 R2 = 0.13
Fig. 3. Relation between the dominant frequency mean (DH) with sonic muscle lipid content (LipidSM). (a) Relation between the dominant frequency variability with (b) sonic musclelipid content (LipidSM), (c) relative sonic muscle mass (RMSM) and (d) relative gonad mass (RMG). Lines of univariate regressions and 95% confidence interval bands are shown.
6 M.C.P. Amorim et al. / Physiology & Behavior xxx (2015) xxx–xxx
Please cite this article as: M.C.P. Amorim, et al., Agonistic sounds signal male quality in the Lusitanian toadfish, Physiol Behav (2015), http://dx.doi.org/10.1016/j.physbeh.2015.06.002

UNCO
RRECTED P
RO
OF
460 in Fig. 1 weremade byManuel Vieira. This study was funded by Science461 and Technology Foundation, Portugal (project PTDC/MAR/118767/462 2010, pluriannual programme UI&D 331/94 and UI&D 329, and grant463 SFRH/BPD/41489/2007 to M.C.P.A).
464 References
465 [1] J.W. Bradbury, S.L. Vehrencamp, Principles of Animal Communication, Sinauer Asso-466 ciates, Sunderland, MA, 1998.467 [2] N.B. Davies, T.R. Halliday, Deep croaks and fighting assessment in toads Bufo bufo,468 Nature 274 (1978) 683–685, http://dx.doi.org/10.1038/274683a0.469 [3] W.D. Brown, A.T. Smith, B. Moskalik, J. Gabriel, Aggressive contests in house crickets:470 size, motivation and the information content of aggressive songs, Anim. Behav. 72471 (2006) 225–233, http://dx.doi.org/10.1016/j.anbehav.2006.01.012.472 [4] M.C.P. Amorim, J.M. Simões, N. Mendonça, N.M. Bandarra, V.C. Almada, P.J. Fonseca,473 Lusitanian toadfish song reflects male quality, J. Exp. Biol. 213 (2010) 2297–3004,474 http://dx.doi.org/10.1242/jeb.044586.475 [5] C.A. Klofstad, R.C. Anderson, S. Peters, Sounds like a winner: voice pitch influences476 perception of leadership capacity in both men and women, Proc. R. Soc. B 279477 (2012) 2698–2704, http://dx.doi.org/10.1098/rspb.2012.0311478 [6] F. Ladich, Sound production and acoustic communication, in: G. von der Erude, J.479 Mogdans, B.G. Kapoor (Eds.), The Senses of Fish Adaptations for the Reception of480 Natural Stimuli, Kluwer, Dordrecht and Narosa, New Delhi 2004, pp. 210–230481 [7] F. Ladich, A.A. Myrberg Jr., Agonistic behavior and acoustic communication, in: F.482 Ladich, S.P. Collin, P. Moller, B.G. Kapoor (Eds.), Communication in Fishes, vol. 1, Sci-483 ence Publishers, Enfield, NH 2006, pp. 121–148484 [8] A.N. Rice, B.R. Land, A.H. Bass, Nonlinear acoustic complexity in a fish ‘two-voice’485 system, Proc. Biol. Sci. B 278 (2011) 3762–3768, http://dx.doi.org/10.1098/rspb.486 2011.0656.487 [9] L. Remage-Healey, A.H. Bass, Rapid elevations in both steroid hormones and vocal488 signaling during playback challenge: a field experiment in Gulf toadfish, Horm.489 Behav. 47 (2005) 297–305, http://dx.doi.org/10.1016/j.yhbeh.2004.11.017.490 [10] K.P. Maruska, J.A. Sisneros, Sex steroid-dependent modulation of acoustic com-491 munication systems in fishes, in: F. Ladich (Ed.), Sound Communication in Fishes,492 Springer-Verlag, Wien, 2015 (in pressQ4 ).493 [11] A.A. Myrberg Jr., S.J. Ha, H. Shamblott, The sounds of bicolor damselfish494 (Pomacentrus partitus): predictors of body size and a spectral basis for individual495 recognition and assessment, J. Acoust. Soc. Am. 94 (1993) 3067–3070, http://dx.496 doi.org/10.1121/1.407267.497 [12] F. Ladich, Sound characteristics and outcome of contests in male croaking gouramis498 (Teleostei), Ethology 104 (1998) 517–529, http://dx.doi.org/10.1111/j.1439-0310.499 1998.tb00087.x.500 [13] L.E. Wysocki, F. Ladich, The ontogenetic development of auditory sensitivity, vocal-501 ization and acoustic communication in the labyrinth fish Trichopsis vittata, J. Comp.502 Physiol. A. 187 (2001) 177–187.503 [14] M.C.P. Amorim, S.S. Pedroso, M. Bolgan, J.M. Jordão, M. Caiano, P.J. Fonseca, Painted504 gobies sing their quality out loud: acoustic rather than visual signals advertise male505 quality and contribute to mating success, Funct. Ecol. 27 (2013) 289–298, http://dx.506 doi.org/10.1111/1365-2435.12032.507 [15] A.H. Bass, J.R. McKibben, Neural mechanisms and behaviors for acoustic communi-508 cation in teleost fish, Prog. Neurobiol. 69 (2003) 1–26, http://dx.doi.org/10.1016/509 S0301-0082(03)00004-2.510 [16] R.O. Vasconcelos, J.M. Simões, V.C. Almada, P.J. Fonseca, M.C.P. Amorim, Vocal be-511 haviour during territorial intrusions in the Lusitanian toadfish: boatwhistles also512 function as territorial ‘keep-out’ signals, Ethology 116 (2010) 155–165, http://dx.513 doi.org/10.1111/j.1439-0310.2009.01722.x.514 [17] R.O. Vasconcelos, R. Carriço, A. Ramos, T. Modesto, P.J. Fonseca, M.C.P. Amorim, Vocal515 behavior predicts reproductive success in a teleost fish, Behav. Ecol. 23 (2012)516 375–383, http://dx.doi.org/10.1093/beheco/arr199.517 [18] H.E. Winn, Vocal facilitation and the biological significance of toadfish sounds, in:518 W.N. Tavolga (Ed.), Marine Bio-acoustics, vol. 2, Pergamon Press, Oxford, UK 1967,519 pp. 283–304520 [19] Y.Y. Hsu, R.L. Earley, L.L. Wolf, Modulation of aggressive behaviour by fighting521 experience: mechanisms and contest outcomes, Biol. Rev. 81 (2006) 33–74, http://522 dx.doi.org/10.1017/S146479310500686X.523 [20] T. Modesto, A.V.M. Canário, Morphometric changes and sex steroid levels dur-524 ing the annual reproductive cycle of the Lusitanian toadfish, Halobatrachus525 didactylus, Gen. Comp. Endocrinol. 131 (2003) 220–231, http://dx.doi.org/526 10.1016/S0016-6480(03)00027-3.527 [21] M.C.P. Amorim, J.M. Simões, P.J. Fonseca, V.C. Almada, Patterns of shelter usage and528 social aggregation by the vocal Lusitanian toadfish, Mar. Biol. 157 (2010) 495–503,529 http://dx.doi.org/10.1007/s00227-009-1335-6.530 [22] S. Chellappa, F.A. Huntingford, R.H.C. Strang, R.Y. Thomson, Condition factor and531 hepatosomatic index as estimates of energy status in male three-spine stickleback,532 J. Fish Biol. 47 (1995) 775–787, http://dx.doi.org/10.1111/j.1095-8649.1995.533 tb06002.x.
534[23] S. Mitchell, J. Poland, M.L. Fine, Does muscle fatigue limit advertisement calling in535the oyster toadfish Opsanus tau? Anim. Behav. 76 (2008) 1011–1016, http://dx.536doi.org/10.1016/j.anbehav.2008.03.024.537[24] K.H. Olsson, C. Kvarnemo, O. Svensson, Relative costs of courtship behaviours in538nest-building sand gobies, Anim. Behav. 77 (2009) 541–546, http://dx.doi.org/10.5391016/j.anbehav.2008.10.021.540[25] P. Viles, J. Silverman, Determination of starch and cellulose with anthrone, Anal.541Chem. 21 (1949) 950–953, http://dx.doi.org/10.1021/ac60032a019.542[26] A.P. Scott, A.V. Canário, 17α,20β-Dihydroxi-4-pregnen-3-one 20-sulphate: a major543new metabolite of the teleost oocyte maturation-inducing steroid, Gen. Comp.544Endocrinol. 85 (1992) 91–100, http://dx.doi.org/10.1016/0016-6480(92)90176-K.545[27] A. Damasceno-Oliveira, B. Fernandez-Duran, J. Goncalves, E. Couto, A.V.M. Canario, J.546Coimbra, Plasma steroid hormone levels in female flounder Platichthys flesus and the547influence of fluctuating hydrostatic pressure, Comp. Biochem. Phys. A 163 (2012)548272–277, http://dx.doi.org/10.1016/j.cbpa.2012.08.001.549[28] M.C.P. Amorim, R.O. Vasconcelos, Variability in the mating calls of the Lusitanian550toadfish Halobatrachus didactylus: potential cues for individual recognition, J. Fish551Biol. 72 (2008) 1355–1368, http://dx.doi.org/10.1111/j.1095-8649.2008.01974.x.552[29] P.M. Forlano, M. Marchaterre, D.L. Deitcher, A.H. Bass, Distribution of androgen re-553ceptor mRNA expression in vocal, auditory and neuroendocrine circuits in a teleost554fish, J. Comp. Neurol. 518 (2010) 493–512, http://dx.doi.org/10.1002/cne.22233.555[30] A.H. Bass, L. Remage-Healey, Central pattern generators for social vocalization:556androgen-dependent neurophysiological mechanisms, Horm. Behav. 53 (2008)557659–672, http://dx.doi.org/10.1016/j.yhbeh.2007.12.010.558[31] L. Remage-Healey, A.H. Bass, Rapid, hierarchical modulation of vocal patterning by559steroid hormones, J. Neurosci. 24 (2004) 5892–5900, http://dx.doi.org/10.1523/560JNEUROSCI.1220-04.2004.561[32] L. Remage-Healey, A.H. Bass, A rapid neuromodulatory role for steroid hormones in562the control of reproductive behavior, Brain Res. 1126 (2006) 27–35, http://dx.doi.563org/10.1016/j.brainres.2006.06.049.564[33] M.L. Fine, F.A. Chen, D.A. Keefer, Autoradiographic localization of dihydrotestoster-565one and testosterone concentrating neurons in the brain of the oyster toadfish,566Brain Res. 709 (1996) 65–80, http://dx.doi.org/10.1016/0006-8993(95)01275-3567[34] B.P. Chagnaud, R. Baker, A.H. Bass, Vocalization frequency and duration are coded in568separate hindbrain nuclei, Nat. Commun. 2 (346) (2011) 1–11, http://dx.doi.org/10.5691038/ncomms1349.570[35] J.C. Wingfield, S. Lynn, K.K. Soma, Avoiding the ‘costs’ of testosterone: ecological571bases of hormone–behavior interactions, Brain Behav. Evol. 57 (2001) 239–251,572http://dx.doi.org/10.1159/000047243.573[36] A.F.H. Ros, K. Becker, A.V.M. Canário, R.F. Oliveira, Androgen levels and energy me-574tabolism in Oreochromis mossambicus, J. Fish Biol. 65 (2004) 895–905, http://dx.575doi.org/10.1111/j.1095-8649.2004.00484.x.576[37] M.L. Fine, M.S. Johnson, D.W. Matt, Seasonal variation in androgen levels in the oys-577ter toadfish, Copeia 2004 (2004) 235–244, http://dx.doi.org/10.1643/CP-03-073R1.578[38] J.C. Wingfield, R.E. Hegner, A.M. Dufty Jr., G.F. Ball, The “challenge hypothesis”: the-579oretical implications for patterns of testosterone secretion, mating systems and580breeding strategies, Am. Nat. 136 (1990) 829–846 (Stable URL: http://www.jstor.581org/stable/2462170).582[39] R.K. Brantley, J.C. Wingfield, A.H. Bass, Sex steroid levels in Porichthys notatus, a fish583with alternative reproductive tactics, and a review of the hormonal bases for male584dimorphism among teleost fishes, Horm. Behav. 27 (1993) 332–347, http://dx.doi.585org/10.1006/hbeh.1993.1025.586[40] M.A. Connaughton, M.L. Fine, M.H. Taylor, The effects of seasonal hypertrophy and587atrophy on fiber morphology, metabolic substrate concentration and sound charac-588teristics of the weakfish sonic muscle, J. Exp. Biol. 200 (1997) 2449–2457.589[41] T. Modesto, A.V.M. Canário, Hormonal control of swimbladder sonic muscle dimor-590phism in the Lusitanian toadfish Halobatrachus didactylus, J. Exp. Biol. 206 (2003)5913467–3477, http://dx.doi.org/10.1242/jeb.00581.592[42] M.L. Fine, K.R. Pennypacker, Hormonal basis for sexual dimorphism of the sound-593producing apparatus of the oyster toadfish, Exp. Neurol. 92 (1986) 289–298,594http://dx.doi.org/10.1016/0014-4886(86)90081-6595[43] R.M. Genova, M.A. Marchaterre, R. Knapp, D. Fergus, A.H. Bass, Glucocorticoid and596androgen signaling pathways diverge between advertisement calling and non-597calling fish, Horm. Behav. 62 (2012) 426–432, http://dx.doi.org/10.1016/j.yhbeh.5982012.07.010.599[44] G.T. Crossin, M. Poisbleau, L. Demongin, O. Chastel, T.D. Williams, M. Eens, P.600Quillfeldt, Migratory constraints on yolk precursors limit yolk androgen deposition601and underlie a brood reduction strategy in rockhopper penguins, Biol. Lett. 8 (2012)6021055–1058, http://dx.doi.org/10.1098/rsbl.2012.0476.603[45] C.P.H. Elemans, A.F. Mensinger, L.C. Rome, Vocal production complexity correlates604with neural instructions in the oyster toadfish (Opsanus tau), J. Exp. Biol. 217605(2014) 1887–1893, http://dx.doi.org/10.1242/jeb.097444.606[46] M.L. Fine, R.F. Thorson, Use of passive acoustics for assessing behavioral interactions607in individual toadfish, T. Am. Fish. Soc. 137 (2008) 627–637, http://dx.doi.org/10.6081577/T04-134.1.
609
7M.C.P. Amorim et al. / Physiology & Behavior xxx (2015) xxx–xxx
Please cite this article as: M.C.P. Amorim, et al., Agonistic sounds signal male quality in the Lusitanian toadfish, Physiol Behav (2015), http://dx.doi.org/10.1016/j.physbeh.2015.06.002