
Abiotic factors controlling the establishment and abundance of the invasive golden mussel Limnoperna...
13
ORIGINAL PAPER Abiotic factors controlling the establishment and abundance of the invasive golden mussel Limnoperna fortunei Ma ´rcia D. Oliveira • De ´bora F. Calheiros • Claudia M. Jacobi • Stephen K. Hamilton Received: 9 November 2008 / Accepted: 24 August 2010 / Published online: 29 September 2010 Ó Springer Science+Business Media B.V. 2010 Abstract Limnoperna fortunei (Bivalvia, Mytilidae), a freshwater bivalve native to Southern Asia, has been an invasive species in South America since 1991. It spread upstream in the La Plata basin reaching the Paraguay River in the vicinity of the Pantanal wetland, Brazil, around 1998. The role of abiotic factors in controlling establishment and abundance of this spe- cies is not well known, making projections of its risk of further spread difficult. This study evaluates the importance of abiotic factors to L. fortunei populations established in rivers of the Pantanal, focusing on larval and juvenile densities and taking advantage of a wide range of seasonal variability in water temperature, flow, dissolved oxygen, and suspended material. Temperature, river stage (influencing several water characteristics) and water velocity are the main variables related to the larval and juvenile densities. In the Pantanal, environmental variables vary over a broader range compared with other South American locations, subjecting L. fortunei to oxygen depletion, low calcium, low pH, and high water velocity and suspended solids, associated with low chlorophyll a concentrations. The combined effect of several of these conditions may explain the relatively low densities in some Pantanal sites. However, they probably will not prevent the persistence of popula- tions in the Pantanal and the eventual establishment of viable populations in upriver systems connected to the Pantanal. These results are pertinent not only to this species but also to other aquatic invasive invertebrates whose expansion may be limited by thermal extremes, episodic oxygen depletion, and waters that are too dilute or acidic for optimal biocalcification. Keywords Aquatic invasive species Freshwater mollusks Wetlands South America Oxygen Calcium Introduction The freshwater mytilid bivalve Limnoperna fortunei (Dunker, 1857), the golden mussel, is an invasive species in South America that in many respects is comparable to the invasive dreissenid zebra mussel, Dreissena polymorpha (Pallas), in North America (Ricciardi 1998). L. fortunei was first detected in South America in 1991 in brackish waters of the La M. D. Oliveira (&) D. F. Calheiros Embrapa Pantanal, Rua 21 de Setembro, 1880, CP 109, CEP 79320 900 Corumba ´, MS, Brazil e-mail: [email protected] C. M. Jacobi Federal University of Minas Gerais, Av. Antonio Carlos, 6627, CEP 31270-901 Belo Horizonte, MG, Brazil S. K. Hamilton W.K. Kellogg Biological Station and Department of Zoology, Michigan State University, 3700 E. Gull Lake Drive, Hickory Corners, MI 49060-9516, USA e-mail: [email protected] 123 Biol Invasions (2011) 13:717–729 DOI 10.1007/s10530-010-9862-0
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Abiotic factors controlling the establishment and abundance of the invasive golden mussel Limnoperna...

ORIGINAL PAPER
Abiotic factors controlling the establishment and abundanceof the invasive golden mussel Limnoperna fortunei
Marcia D. Oliveira • Debora F. Calheiros •
Claudia M. Jacobi • Stephen K. Hamilton
Received: 9 November 2008 / Accepted: 24 August 2010 / Published online: 29 September 2010
� Springer Science+Business Media B.V. 2010
Abstract Limnoperna fortunei (Bivalvia, Mytilidae),
a freshwater bivalve native to Southern Asia, has been
an invasive species in South America since 1991. It
spread upstream in the La Plata basin reaching the
Paraguay River in the vicinity of the Pantanal wetland,
Brazil, around 1998. The role of abiotic factors in
controlling establishment and abundance of this spe-
cies is not well known, making projections of its risk of
further spread difficult. This study evaluates the
importance of abiotic factors to L. fortunei populations
established in rivers of the Pantanal, focusing on larval
and juvenile densities and taking advantage of a wide
range of seasonal variability in water temperature,
flow, dissolved oxygen, and suspended material.
Temperature, river stage (influencing several water
characteristics) and water velocity are the main
variables related to the larval and juvenile densities.
In the Pantanal, environmental variables vary over a
broader range compared with other South American
locations, subjecting L. fortunei to oxygen depletion,
low calcium, low pH, and high water velocity and
suspended solids, associated with low chlorophyll
a concentrations. The combined effect of several of
these conditions may explain the relatively low
densities in some Pantanal sites. However, they
probably will not prevent the persistence of popula-
tions in the Pantanal and the eventual establishment of
viable populations in upriver systems connected to the
Pantanal. These results are pertinent not only to this
species but also to other aquatic invasive invertebrates
whose expansion may be limited by thermal extremes,
episodic oxygen depletion, and waters that are too
dilute or acidic for optimal biocalcification.
Keywords Aquatic invasive species � Freshwater
mollusks � Wetlands � South America � Oxygen �Calcium
Introduction
The freshwater mytilid bivalve Limnoperna fortunei
(Dunker, 1857), the golden mussel, is an invasive
species in South America that in many respects is
comparable to the invasive dreissenid zebra mussel,
Dreissena polymorpha (Pallas), in North America
(Ricciardi 1998). L. fortunei was first detected in
South America in 1991 in brackish waters of the La
M. D. Oliveira (&) � D. F. Calheiros
Embrapa Pantanal, Rua 21 de Setembro, 1880, CP 109,
CEP 79320 900 Corumba, MS, Brazil
e-mail: [email protected]
C. M. Jacobi
Federal University of Minas Gerais, Av. Antonio Carlos,
6627, CEP 31270-901 Belo Horizonte, MG, Brazil
S. K. Hamilton
W.K. Kellogg Biological Station and Department
of Zoology, Michigan State University, 3700 E. Gull Lake
Drive, Hickory Corners, MI 49060-9516, USA
e-mail: [email protected]
123
Biol Invasions (2011) 13:717–729
DOI 10.1007/s10530-010-9862-0

Plata River, Argentina (Pastorino et al. 1993). It was
carried upstream in subsequent years, presumably
aided by ship and barge traffic in the Paraguay-Parana
waterway, reaching the Pantanal wetland in Brazil by
1998 (Oliveira et al. 2006). Several authors have
noted the potential for L. fortunei to invade North
American waters, where it could occupy regions that
are too warm for the dreissenid mussels (Ricciardi
1998; Oliveira et al. 2010a).
In spite of some controversy about the taxonomy
of genus Limnoperna, and consequently about its
distribution (Lee and Morton 1983; Morton 1996),
L. fortunei is thought to be native to two major river
basins in China, the Pearl (Zhujiang) and Yangtze
(Changjiang) rivers (Morton 1996). In the Yangtze
River basin, where it has a broad distribution
(Lee and Morton 1983), L. fortunei has been referred
to as Limnoperna lacustris (Xie et al. 1999; Wang
et al. 1999, 2007). Introductions into Hong Kong,
Japan, and South America were mentioned by
Morton (1975, 1977), Magara et al. (2001), and
Darrigran et al. (1999), respectively. Morton (1987)
defined L. fortunei as an r-selected species, adapted to
the invasive colonization of a wide range of aquatic
environments, with important characteristics of a
successful invader such as being short-lived, capable
of rapid growth, and having a high fecundity, aside
from broad physiological tolerance to several abiotic
factors that often limit aquatic invertebrates.
L. fortunei is recognized as a problem since it
promotes large-scale fouling in water supply systems
and power plants, attaining densities greater than
50,000 ind m-2 (Darrigran et al. 2003). Morton
(1975) suggested that L. fortunei has a potential for
causing problems even in its native environments,
being considered a pest that fouls boats and jetties in
the Pearl River. There are no reports of fouling by
Limnoperna in the Yangtze River, where only
1.0–40.0 ind m-2 of L. lacustris (L. fortunei accord-
ing to Morton 1996) were recorded by Wang et al.
(2007).
In general, exotic species with broad distribution
in their native range should also achieve broad
distributions in comparable introduced habitats if
given enough time (Guo 2006). Modeling the distri-
bution potential based on their ecological niche has
been widely used to predict the spread of invasive
species (Drake and Bossenbroek 2004; Bossenbroek
et al. 2007; Kluza and McNyset 2005). Therefore, to
build realistic models and validate them it is funda-
mental to have reliable environmental data and
scientific understanding of the species biology
(Peterson and Vieglais 2001). Information on abiotic
limiting factors is essential to predict where an
invasive species is likely to flourish. In the case of
L. fortunei its broad tolerance to chemical and
physical variables that usually inhibit other aquatic
invertebrates makes it difficult to identify limiting
abiotic factors that would control or prevent its spread.
Ricciardi (1998) suggested some limits of L. fortunei
tolerance to chemical and physical aquatic variables,
based on characteristics of habitats invaded by this
species in eastern Asia and southern South America.
Inferences about these abiotic limits are mostly based
on observations of adult mussels. However, larvae and
juveniles may be more sensitive than adults to some of
these variables, and there is very little information on
how abiotic factors affect immature stages in natural
environments, with direct implications for adult
mussel density and invasive potential.
The objective of this study is to test our current
understanding of the environmental tolerance of
L. fortunei, with an emphasis on initial colonization
and immature stages. We examine the population
dynamics of L. fortunei in rivers of the Pantanal
wetland, a tropical floodplain-river complex where
abiotic factors such as temperature, oxygen and
calcium concentrations vary over a greater range than
has been reported for any other South American
location containing this mussel. We describe the
environment where L. fortunei is established in the
Pantanal, compare water conditions with the tolerance
limits presented in the literature, and examine the
influence of abiotic factors on L. fortunei colonization
and density. These results are pertinent not only to this
species but also to other aquatic invasive invertebrates
whose expansion may be limited by thermal extremes,
episodic oxygen depletion, and waters that are too
dilute or acidic for optimal biocalcification.
Methods
Study area
The Brazilian portion of the upper Paraguay River basin
covers about 362,000 km2, within which the Pantanal
wetland, the largest contiguous floodplain area in the
718 M. D. Oliveira et al.
123

world, extends over about 140,000 km2 in Brazil
(PCBAP (Plano de Conservacao da Bacia do Alto
Paraguai) 1997) (Fig. 1). The climate in the region is
tropical with marked wet and dry seasons. The maxi-
mum daily temperature averages 32�C in the Pantanal,
ranging from 28 to 34�C. The minimum monthly
temperature averages about 20�C (PCBAP 1997).
The Paraguay River (mean depth 8.6 m and width
varying from 100 to 500 m) flows over 3,800 km
from north to south collecting the waters of large
tributaries on its left margin, including the Cuiaba,
Taquari and Miranda rivers (Fig. 1). Its mean annual
discharge is about 300 m3 s-1 in the northern Pant-
anal and 1,555 m3 s-1 after it receives water from its
major tributaries in the Pantanal.
The rivers and floodplains of the region exhibit a
marked and protracted seasonal flood pulse, with
water levels varying seasonally over a range of 3–5 m
in the southern Pantanal. Flooding occurs as a result
of river overflow, local precipitation or a combination
of both. The contact between the Paraguay River and
its tributaries with the extensive floodplains at the
beginning of the rising water phase often causes
dramatic changes in water chemistry, a natural
phenomenon labeled as ‘oxygen depletion events’
by Hamilton et al. (1997) and Calheiros and Hamilton
(1998) in reference to one of its main consequences.
Oxygen depletion to hypoxic or anoxic conditions,
accompanied by a decrease in pH and other chemical
changes, extends to the Paraguay River and some-
times results in massive fish kills in the rivers and
associated floodplain waters.
The Miranda River is one of the main tributaries of
the Paraguay River within the Pantanal and drains
about 47,000 km2 (PCBAP 1997), including upland
tributaries with clear waters that originate in lime-
stone formations of the Bodoquena hills. The lower
Miranda river is about 150 m wide and its discharge
is around 86 m3 s-1 (PCBAP 1997). Its flow regime
responds more directly to rain events than the
Paraguay River, so the flooding phase takes place
earlier, from November or December to April, and is
influenced by backwater effects of the Paraguay
River as well as rainfall in its drainage basin.
Data collection and analysis
The study area included two reaches of the Paraguay
River (Porto Esperanca site/PR-PE, 19�360S and
57�260W, and Corumba site/PR-CR, 18�590S and
57�420W), and one section in the lower Miranda
River (Passo do Lontra, MR, 19�340S and 57�14.70W)
close to the confluence with the Paraguay River.
These sites represent a gradient of L. fortunei density
and calcium concentration, with the highest densities
of juveniles and adults at Porto Esperanca, and the
lowest in the Miranda river, which is rich in calcium
compared to the Paraguay River (Oliveira 2009). The
establishment of L. fortunei was confirmed in 1998 in
the Paraguay River and 2002 in the Miranda River.
River stage (water level) data for the Paraguay River
(19�020S, 57�330W) were obtained from daily readings
carried out by the Brazilian Navy and by the National
Water Agency (ANA), and stage data for the Miranda
River were obtained from the Federal University of
Mato Grosso do Sul. Substrata for larval settlement
were available at all sites, in the form of hard natural
rock surfaces or artificial structures, and thus substrata
were not considered a limiting factor.
Samples for analysis of limnological variables and
larval and juvenile density were taken at each site
monthly from January 2004 to November 2007. For
water analysis, subsurface water samples were taken
with a Van Dorn sampler. Water temperature,
dissolved oxygen concentration (DO), electrical con-
ductivity and pH were measured in situ with a water
quality sonde (YSI). Water transparency was esti-
mated using a Secchi disc. Total alkalinity was
analyzed by the Gran titration method (Gran 1952)
and free CO2 was calculated following Kempe
(1982), with modifications by Hamilton et al.
(1995). Chlorophyll a was analyzed following
Marker et al. (1981). Total suspended solids (TSS),
inorganic (IM) and organic matter (OM), and organic
matter percentage were determined by gravimetric
and loss-on-ignition methods, and calcium concen-
tration was analyzed by atomic absorption, all based
on APHA (1998). Total organic carbon (TOC) was
analyzed using a Shimadzu Carbon Analyzer. Water
velocity was measured with a portable flowmeter
(Marsh-McBirney, Inc, model 2000).
To determine juvenile density, we counted indi-
viduals of 0.3–3.0 mm shell length settled after about
30 days onto artificial substrata. Each substratum was
a 100 cm2 Nylon net (1.0 mm mesh) installed inside
a PVC tube with a diameter of 12 cm (or a PET
bottle), closed with a 5.0-mm mesh net to prevent fish
predation (predation represents up to 35% of total
Abiotic factors controlling the establishment 719
123
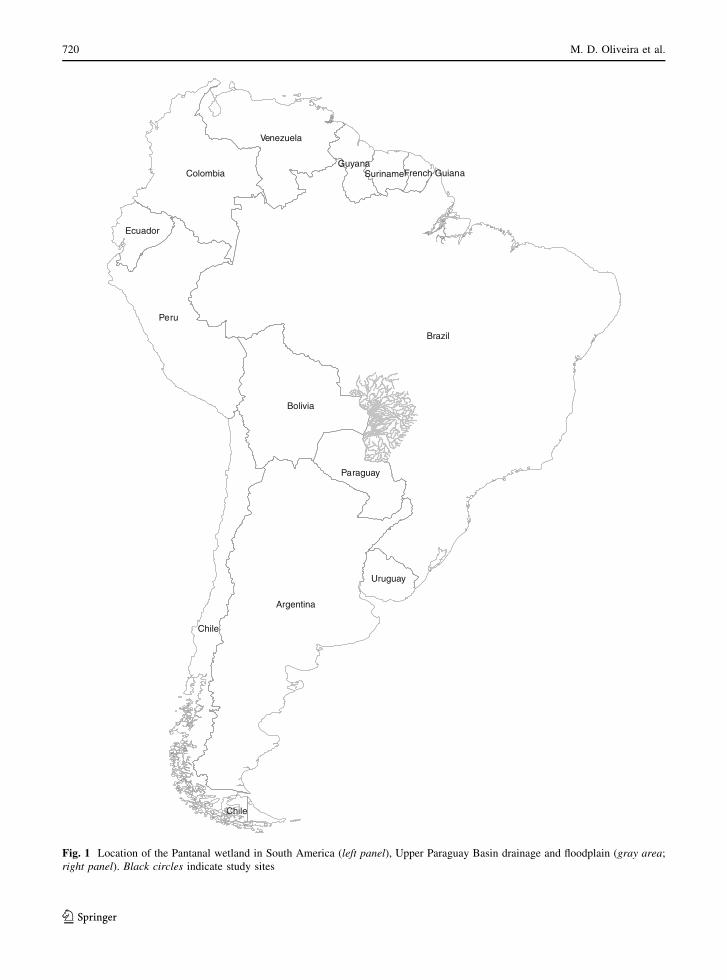
Brazil
Argentina
Peru
Bolivia
Chile
Colombia
Venezuela
Paraguay
Ecuador
Uruguay
GuyanaSuriname
Chile
French Guiana
Fig. 1 Location of the Pantanal wetland in South America (left panel), Upper Paraguay Basin drainage and floodplain (gray area;
right panel). Black circles indicate study sites
720 M. D. Oliveira et al.
123
losses after settlement; Oliveira, unpublished data).
Six of these units were attached to supports of
existing bridges or docks in each site. The Nylon net
was replaced each month and settled organisms were
preserved in 70% ethanol for counting.
For larval counts, three samples of 300 l were
concentrated in a 36-lm mesh plankton net, and
preserved in 70% ethanol. Five aliquots of 2-ml
subsamples were taken using a Hensen-Stempel pipette
and all larvae in each subsample were counted in a
Fig. 1 continued
Abiotic factors controlling the establishment 721
123
reticulated camera, with the aid of an 809 stereomicro-
scope (Wetzel and Likens 2001).
The adult condition index, shell length:weight
ratio, and shell growth rate were measured to
compare the condition of adult mussels in the
Miranda river with those in the Paraguay River-PE,
where adult density is much higher. To calculate the
Condition Index (CI = tissue weight 9 100/shell
length) we measured shell length of mussels larger
than 13.0 mm (average length of 14.2 mm in the
Paraguay River-PE and 17.2 mm in the Miranda
River). Mussel tissue was separated from the shell
and both were dried to constant weight at 80�C. We
also conducted in situ assays of shell growth rate,
placing 30 mussels ranging between 6.0 and 10.0 mm
in shell length in a cage protected with Nylon mesh in
the Paraguay River-PE and Miranda River. Mussel
shell growth was were measured every 30 days over
210 days in the Paraguay River and 320 days in the
Miranda River, from June 2006 to July 2007.
To examine how the flood pulse affected mussel
densities we estimated the Spearman correlation
between juvenile density and limnological variables,
combining data from all sites. We also examined the
relationship between river stage and other variables.
We were particularly interested in how water temper-
ature and oxygen depletion events affect L. fortunei
recruitment (measured as juvenile density). These two
variables were combined in a two-factor logistic
regression to evaluate the probability of L. fortunei
recruitment. Analyses and graphs were performed
using SYSTAT 11 software (Wilkinson 2004).
Results
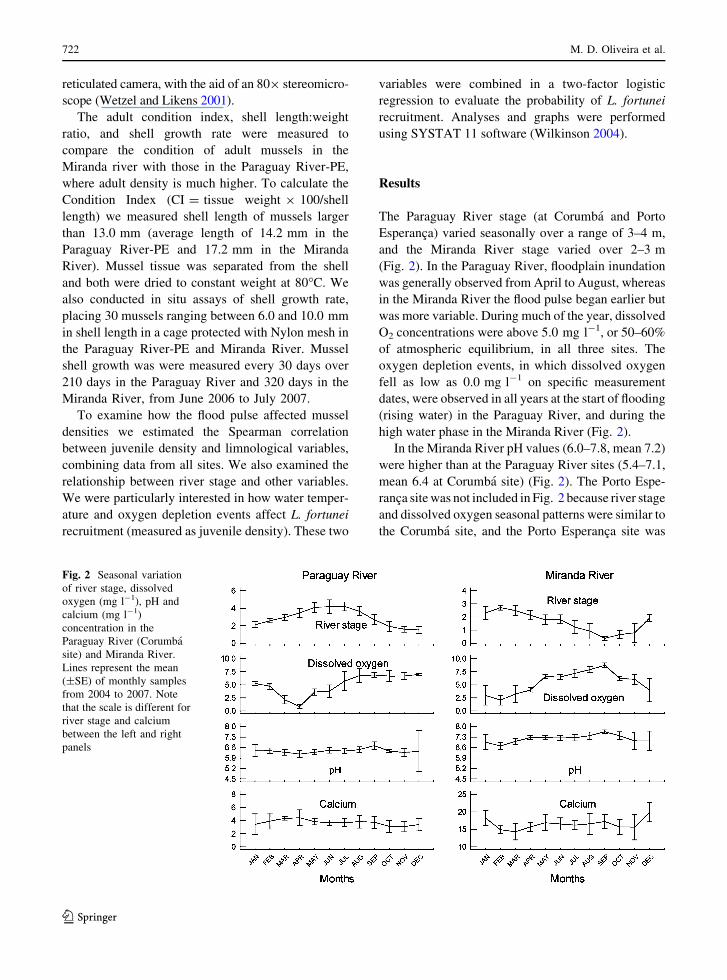
The Paraguay River stage (at Corumba and Porto
Esperanca) varied seasonally over a range of 3–4 m,
and the Miranda River stage varied over 2–3 m
(Fig. 2). In the Paraguay River, floodplain inundation
was generally observed from April to August, whereas
in the Miranda River the flood pulse began earlier but
was more variable. During much of the year, dissolved
O2 concentrations were above 5.0 mg l-1, or 50–60%
of atmospheric equilibrium, in all three sites. The
oxygen depletion events, in which dissolved oxygen
fell as low as 0.0 mg l-1 on specific measurement
dates, were observed in all years at the start of flooding
(rising water) in the Paraguay River, and during the
high water phase in the Miranda River (Fig. 2).
In the Miranda River pH values (6.0–7.8, mean 7.2)
were higher than at the Paraguay River sites (5.4–7.1,
mean 6.4 at Corumba site) (Fig. 2). The Porto Espe-
ranca site was not included in Fig. 2 because river stage
and dissolved oxygen seasonal patterns were similar to
the Corumba site, and the Porto Esperanca site was
Fig. 2 Seasonal variation
of river stage, dissolved
oxygen (mg l-1), pH and
calcium (mg l-1)
concentration in the
Paraguay River (Corumba
site) and Miranda River.
Lines represent the mean
(±SE) of monthly samples
from 2004 to 2007. Note
that the scale is different for
river stage and calcium
between the left and right
panels
722 M. D. Oliveira et al.
123

intermediate in pH and calcium values (5.9–7.0 and
1.8–8.9 mg l-1, respectively) because it is below the
confluence of the Miranda River. A clear decrease of
pH to 6.0 was observed in the Miranda River at the time
of its lowest oxygen concentration. In the Paraguay
River the minimum pH was 5.4 at Corumba and 5.9 at
Porto Esperanca. Some acidification was also observed
during falling water (November to December) in the
Paraguay River.
Free CO2 was highest during oxygen depletion
events and is the predominant factor controlling pH
changes. The maximum free CO2 concentrations
were 68 mg l-1 (Porto Esperanca site) and 87 mg l-1
(Corumba site) in the Paraguay River, and
153 mg l-1 in the Miranda River. The normal CO2
concentration throughout the year, i.e. excluding
oxygen depletion events, was below 20 mg l-1.
Calcium concentrations were relatively high in the
Miranda River, varying between 10 and 23 mg l-1.
Calcium concentrations were much lower over the
course of the year in the Paraguay River (between 1.0
and 6.0 mg l-1 at the Corumba site, Fig. 2), and were
lowest from October to February.
Water transparency in the Miranda River
(10–90 cm Secchi depths) was lower than in the
Paraguay River (20–190 cm). Concentrations of TSS
up to 120 mg l-1 occurred from October to December
in the Miranda River, although the mean values were
between 10 and 50 mg l-1. Both Paraguay River sites
showed TSS concentrations of less than 20 mg l-1
along the year, and reached around 50 mg l-1 from
October through December. Ratios of organic matter
(OM) to inorganic matter (IM) indicate predominance
of particulate inorganic material throughout the year
(Paraguay River = 4.5 ± 2.4 mg l-1 at Corumba
and 4.3 ± 2.1 mg l-1 at Porto Esperanca; Miranda
River = 5.2 ± 1.9 mg l-1).
The concentration of chlorophyll a was low at all
sites, averaging less than 1.0 lg l-1 (Paraguay River-
PE: mean 0.79 ± 0.76 lg l-1; CR: 0.78 ± 0.98 lg l-1
and Miranda River: 0.98 ± 0.98 lg l-1). TOC was
slightly higher in the Miranda River (mean
1.7 ± 0.9 mg l-1) compared to the Paraguay River at
Porto Esperanca (1.3 ± 1.0 mg l-1) and Corumba
(1.2 ± 0.8 mg l-1). Ratios of TOC to chlorophyll
a indicate predominance of detritus in both Miranda
(82,204 ± 21,084) and Paraguay rivers (115,033 ±
36,128 at Corumba and 17,715 ± 75,565 at Porto
Esperanca).
Water velocity, usually between 0.6 and
0.8 m s-1, was relatively high in the Miranda River.
At Porto Esperanca, on the other hand, mean monthly
values were low (\0.2 m s-1) and fairly uniform.
Comparatively, water velocity at Corumba fluctuated
more, ranging between 0.1 and 0.6 m s-1 (Fig. 3).
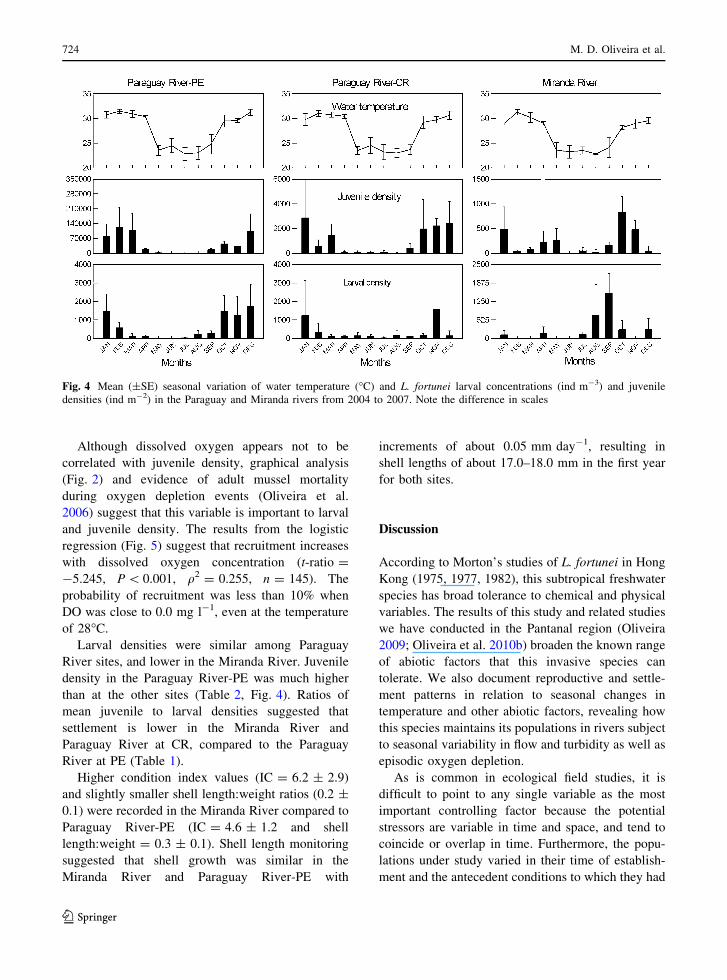
Both planktonic larval density and juvenile settle-
ment lasted around seven to 8 months (Fig. 4) in both
Paraguay River sites, associated with seasonally high
water temperature, but showed a poorly defined
pattern in the Miranda River. From February to
March no larvae or juveniles were recorded in the
Miranda River. In general, larval settlement was
observed from September to April with peaks from
October to February. Between May and August
settlement usually was not observed. After Septem-
ber, when water temperature rose above 8�C, larvae
and juveniles were again observed, except in the
Miranda River, where larvae occurred in August and
September. In general, the reproductive period was
coincident with temperatures between 28.0 and
32.5�C (maximum water temperatures of 35�C were
recorded in the Paraguay River in October and
November).
The relationship between juvenile density and
most limnological variables was weak, as shown by
Spearman’s correlation (Table 1). The best correla-
tions with juvenile density were observed for water
velocity, water temperature and river stage. River
stage covaries with variables such as dissolved
oxygen, pH, free CO2, total suspended solids and
organic matter percentage, as shown by the correla-
tions in Table 1.
Fig. 3 Water velocity in the Paraguay and Miranda rivers.
Lines are the means of monthly samples from 2004 to 2007,
indicated by symbols: PR-PE (open square), PR-CR (fulltriangle) and MR (cross)
Abiotic factors controlling the establishment 723
123
Although dissolved oxygen appears not to be
correlated with juvenile density, graphical analysis
(Fig. 2) and evidence of adult mussel mortality
during oxygen depletion events (Oliveira et al.
2006) suggest that this variable is important to larval
and juvenile density. The results from the logistic
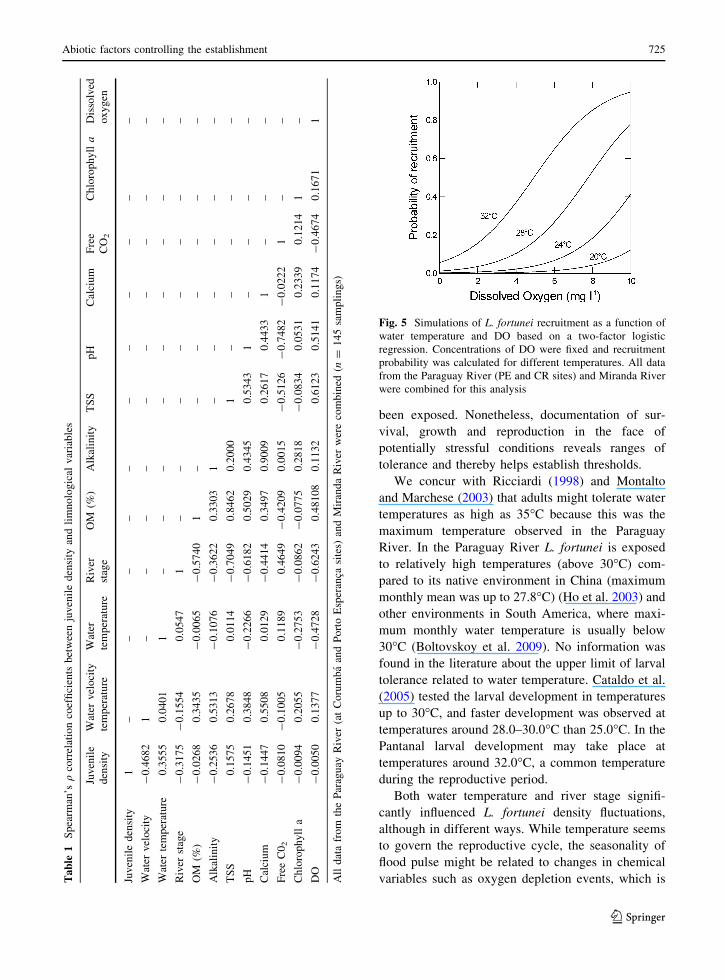
regression (Fig. 5) suggest that recruitment increases
with dissolved oxygen concentration (t-ratio =
-5.245, P \ 0.001, q2 = 0.255, n = 145). The
probability of recruitment was less than 10% when
DO was close to 0.0 mg l-1, even at the temperature
of 28�C.
Larval densities were similar among Paraguay
River sites, and lower in the Miranda River. Juvenile
density in the Paraguay River-PE was much higher
than at the other sites (Table 2, Fig. 4). Ratios of
mean juvenile to larval densities suggested that
settlement is lower in the Miranda River and
Paraguay River at CR, compared to the Paraguay
River at PE (Table 1).
Higher condition index values (IC = 6.2 ± 2.9)
and slightly smaller shell length:weight ratios (0.2 ±
0.1) were recorded in the Miranda River compared to
Paraguay River-PE (IC = 4.6 ± 1.2 and shell
length:weight = 0.3 ± 0.1). Shell length monitoring
suggested that shell growth was similar in the
Miranda River and Paraguay River-PE with
increments of about 0.05 mm day-1, resulting in
shell lengths of about 17.0–18.0 mm in the first year
for both sites.
Discussion
According to Morton’s studies of L. fortunei in Hong
Kong (1975, 1977, 1982), this subtropical freshwater
species has broad tolerance to chemical and physical
variables. The results of this study and related studies
we have conducted in the Pantanal region (Oliveira
2009; Oliveira et al. 2010b) broaden the known range
of abiotic factors that this invasive species can
tolerate. We also document reproductive and settle-
ment patterns in relation to seasonal changes in
temperature and other abiotic factors, revealing how
this species maintains its populations in rivers subject
to seasonal variability in flow and turbidity as well as
episodic oxygen depletion.
As is common in ecological field studies, it is
difficult to point to any single variable as the most
important controlling factor because the potential
stressors are variable in time and space, and tend to
coincide or overlap in time. Furthermore, the popu-
lations under study varied in their time of establish-
ment and the antecedent conditions to which they had
Fig. 4 Mean (±SE) seasonal variation of water temperature (�C) and L. fortunei larval concentrations (ind m-3) and juvenile
densities (ind m-2) in the Paraguay and Miranda rivers from 2004 to 2007. Note the difference in scales
724 M. D. Oliveira et al.
123
been exposed. Nonetheless, documentation of sur-
vival, growth and reproduction in the face of
potentially stressful conditions reveals ranges of
tolerance and thereby helps establish thresholds.
We concur with Ricciardi (1998) and Montalto
and Marchese (2003) that adults might tolerate water
temperatures as high as 35�C because this was the
maximum temperature observed in the Paraguay
River. In the Paraguay River L. fortunei is exposed
to relatively high temperatures (above 30�C) com-
pared to its native environment in China (maximum
monthly mean was up to 27.8�C) (Ho et al. 2003) and
other environments in South America, where maxi-
mum monthly water temperature is usually below
30�C (Boltovskoy et al. 2009). No information was
found in the literature about the upper limit of larval
tolerance related to water temperature. Cataldo et al.
(2005) tested the larval development in temperatures
up to 30�C, and faster development was observed at
temperatures around 28.0–30.0�C than 25.0�C. In the
Pantanal larval development may take place at
temperatures around 32.0�C, a common temperature
during the reproductive period.
Both water temperature and river stage signifi-
cantly influenced L. fortunei density fluctuations,
although in different ways. While temperature seems
to govern the reproductive cycle, the seasonality of
flood pulse might be related to changes in chemical
variables such as oxygen depletion events, which isTa
ble
1S
pea
rman
’sq
corr
elat
ion
coef
fici
ents
bet
wee
nju
ven
ile
den
sity
and
lim
no
log
ical
var
iab
les
Juv
enil
e
den
sity
Wat
erv
elo
city
tem
per
atu
re
Wat
er
tem
per
atu
re
Riv
er
stag
e
OM
(%)
Alk
alin
ity
TS
Sp
HC
alci
um
Fre
e
CO
2
Ch
loro
ph
yll
aD
isso
lved
ox
yg
en
Juv
enil
ed
ensi
ty1
––
––
––
––
––
–
Wat
erv
elo
city
-0
.46
82
1–
––
––
––
––
–
Wat
erte
mp
erat
ure
0.3
55
50
.04
01
1–
––
––
––
––
Riv
erst
age
-0
.31
75
-0
.15
54
0.0
54
71
––
––
––
––
OM
(%)
-0
.02
68
0.3
43
5-
0.0
06
5-
0.5
74
01
––
––
––
–
Alk
alin
ity
-0
.25
36
0.5
31
3-
0.1
07
6-
0.3
62
20
.33
03
1–
––
––
–
TS
S0
.15
75
0.2
67
80
.01
14
-0
.70
49
0.8
46
20
.20
00
1–
––
––
pH
-0
.14
51
0.3
84
8-
0.2
26
6-
0.6
18
20
.50
29
0.4
34
50
.53
43
1–
––
–
Cal
ciu
m-
0.1
44
70
.55
08
0.0
12
9-
0.4
41
40
.34
97
0.9
00
90
.26
17
0.4
43
31
––
–
Fre
eC
02
-0
.08
10
-0
.10
05
0.1
18
90
.46
49
-0
.42
09
0.0
01
5-
0.5
12
6-
0.7
48
2-
0.0
22
21
––
Ch
loro
ph
yll
a-
0.0
09
40
.20
55
-0
.27
53
-0
.08
62
-0
.07
75
0.2
81
8-
0.0
83
40
.05
31
0.2
33
90
.12
14
1–
DO
-0
.00
50
0.1
37
7-
0.4
72
8-
0.6
24
30
.48
10
80
.11
32
0.6
12
30
.51
41
0.1
17
4-
0.4
67
40
.16
71
1
All
dat
afr
om
the
Par
agu
ayR
iver
(at
Co
rum
ba
and
Po
rto
Esp
eran
casi
tes)
and
Mir
and
aR
iver
wer
eco
mb
ined
(n=
14
5sa
mp
lin
gs)
Fig. 5 Simulations of L. fortunei recruitment as a function of
water temperature and DO based on a two-factor logistic
regression. Concentrations of DO were fixed and recruitment
probability was calculated for different temperatures. All data
from the Paraguay River (PE and CR sites) and Miranda River
were combined for this analysis
Abiotic factors controlling the establishment 725
123

known to cause adult mussel die-offs (Oliveira et al.
2006), directly affecting density.
Both larval and juvenile densities indicated almost
continuous reproduction for about 7–8 months in the
Pantanal, a span that resembles that of other popu-
lations established in South America (Boltovskoy and
Cataldo 1999, Darrigran et al. 2003; Santos et al.
2005). Continuous occurrence of planktonic larvae
6–10 months of the year was observed by Boltovskoy
et al. (2009) in most lentic water bodies. These
authors suggested that larval density might be
regulated by the time of reproduction, which in turn
is influenced by water temperature and food supply.
The reproductive pattern of L. fortunei in South
America is quite different from results described by
Iwasaki and Uryu (1998), who compared data from
central China, Hong Kong and Japan. The processes
of gametogenesis and spawning of L. fortunei have
been related to maximum and minimum temperature
or to abrupt changes in water temperature (Morton
1982; Darrigran et al. 2003; Santos et al. 2005). We
suggest that abrupt changes, rather than absolute
values, may elicit the spawning of L. fortunei in the
Pantanal, since mean winter temperatures (which
remained above 20�C) were not limiting.
Although river stage has no direct effect on
density, unless the mussel habitat is exposed to air
during low waters, the seasonal flood pulse causes
important changes in the chemical variables. Oxygen
depletion events usually occur at the end of the
reproductive period in the Paraguay River and in the
middle of the reproductive period (February to
March) in the Miranda River, when low densities or
absence of larvae and juveniles were recorded. In the
latter case, the effect might be more severe -stopping
reproduction- and could explain the low densities in
the Miranda River; we observed that the probability
of reproduction was less than 10% when DO was
close to 0.0 mg l-1. In the Pantanal low DO, possibly
exacerbated by high free CO2, was considered the
main cause of fish mortality during oxygen depletion
events (Hamilton et al. 1995; Calheiros and Hamilton
1998). Information about free CO2 concentration is
not provided for any place where L. fortunei is
present. Oxygen depletion events have been shown to
affect L. fortunei density via direct mortality of adults
(Oliveira et al. 2006). Effects of anoxic conditions on
larval development have not been documented in the
literature, but low dissolved oxygen levels affecting
L. fortunei gametogenesis and spawning were cited
by Morton (1982). The tolerance limits of each
variable were difficult to estimate because of the
interaction among factors, so the minimum dissolved
oxygen requirements of young and adults of
L. fortunei in the Pantanal wetland remain unclear.
L. fortunei is well established in the Paraguay
River (Corumba site) where mean pH is around 6.4,
the lower limit suggested by Ricciardi (1998),
although it may drop to 5.5 for periods of days to a
few weeks during oxygen depletion events. Kara-
tayev et al. (2007) suggested 5.5 as the lower pH limit
to L. fortunei but provided no example of established
populations in habitats with similar characteristics,
and Montalto and Marchese (2003) recorded 80% of
adult survival in pH 5.0 after 96 h of laboratory tests,
but this period is too short to extrapolate to a natural
environment.
The Pearl River in China, which is thought to be a
native environment of L. fortunei, carries about
33 mg l-1 of calcium (Zhang et al. 2007). The lower
limit of 3.0 mg l-1 proposed by Ricciardi (1998) was
likely based on data from Sum Chum Reservoir (China),
the site where L. fortunei has been reported at the lowest
calcium concentrations: 2.4 and 4.8 mg l-1 with a mean
Table 2 Larval, juvenile, and adult density, and juvenile to larva ratio
Larval density Juvenile density (30-day recruitment) Adult density* Juvenile:
larval ratioN ind m-3 N ind m-2 N ind m-2
Paraguay, Corumba 46 1.629 ± 4.571 46 3.149 ± 6.928 12 6.539 ± 5.904 1.93
Paraguay, Porto
Esperanca
41 1.772 ± 4.033 46 88.523 ± 153.349 12 143.134 ± 62.692 49.96
Miranda 42 990 ± 2.292 46 377 ± 682 12 585 ± 358 0.38
Data are mean ± SD of monthly samples from Paraguay River, at Corumba and Porto Esperanca sites, and Miranda River, from 2004
to 2007
*Source: Oliveira (2009)
726 M. D. Oliveira et al.
123

pH of 6.4 (Morton 1975; Ricciardi 1998). In the
Paraguay River at Corumba calcium concentrations
were quite low, varying between 1.0 and 6.0 mg l-1,
and pH can occasionally be more acidic than at Sum
Chum. Indications that mussels are tolerant to ionically
dilute water over short time intervals come from lab
experiments by Deaton et al. (1989) who verified that
mussels retained byssal attachments and filtering activ-
ities by the end of 1 week in deionized water.
In the relatively calcium-rich water of the Miranda
River, other negative factors such as high water
velocity, low availability of food, and high concen-
tration of suspended sediments might explain the low
larval settlement. The recent colonization in the
Miranda River (around 5 years) might contribute to
low juvenile density, but does not explain low density
in the Corumba site (colonization time about
7–10 years). Besides that, no clear increases in larval
or juvenile densities observed from 2004 to 2007
(Oliveira 2009).
The effects of turbidity and low food quality and
quantity on the establishment of Dreissena polymor-
pha are well known (Madon et al. 1998; Schneider
et al. 1998; Allen et al. 1999; Baines et al. 2005,
2007). Schneider et al. (1998) suggested that an
organic:inorganic matter ratio lower than 0.5 can be
considered low food quality. This threshold is related
to the inability of D. polymorpha to selectively ingest
organic material as food quality declines. Availability
of food was proposed by Boltovskoy et al. (2009) to
be a major factor controlling the reproductive activity
of L. fortunei, although other factors such as pH and
calcium were not explored. These authors also
reported low larval density in turbid water (transpar-
ency around 0.1–0.4 m) with scarce phytoplankton.
In the Miranda River mussel reproduction (October to
February) might have been negatively affected by
high concentration of suspended solids, low
organic:inorganic matter ratio (0.2), and low chloro-
phyll a occurring at the same time. However,
L. fortunei attains high densities in the Guaiba Lake
(Southern Brazil) where suspended solids concentra-
tions range up to 214 mg l-1 and water transparency
is around 0.4 m. The difference in settlement could
be because chlorophyll a is higher in Guaiba Lake
(0–23.9, mean 3.5 lg l-1; Santos et al. 2005) than in
Pantanal waters.
Shell growth at rates similar to those reported for
Argentine populations (Boltovskoy and Cataldo 1999)
and similar condition indices in the Miranda and
Paraguay River at PE suggest that the environmental
conditions are suitable for adults in both sites. In the
Miranda River the condition index is higher than those
of mussels from the Itaipu reservoir in Brazil, where
chlorophyll a ranges between 4 and 22 lg l-1 (Silva
2006), suggesting that L. fortunei uses other sources of
energy besides phytoplankton in the Pantanal rivers,
where this resource is scarce (Oliveira and Calheiros
2000). Detrital organic carbon can provide another
source of energy to L. fortunei in the Miranda River,
considering that concentrations of TOC in the Pantanal
waters are around 1–2 mg l-1, analogous to the Parana
River (Depretris and Kempe 1993) where L. fortunei is
successfully established.
In the Miranda River, the good mussel condition
index and greater shell length support the idea that
low food combined with high concentration of
inorganic sediments are probably not a limiting
factor for growth of adults. However, they might be
detrimental to larval stages, where survival seems to
be low in the Miranda River compared to the
Paraguay River. Another negative factor in the
Miranda River might be the higher water velocity,
which ranged from 0.4 to 1.1 m s-1. Although not
harmful to adults it seems to have hindered settlement
onto artificial substrata. Nagaya et al. (2001) found
that a fluid velocity around 1.0 m s-1 affected the
attachment of L. fortunei juveniles.
We confirmed that the environments where
L. fortunei lives in the Paraguay and Miranda rivers
are quite extreme compared to other places where
L. fortunei occurs. Larval density was relatively
lower in the Pantanal compared to data summarized
in Boltovskoy et al. (2009), which likely contribute to
the lower juvenile and adult densities. The effects of
multiple abiotic stressors on larval development in
natural environments are poorly known, and further
research is necessary. Most studies are more con-
cerned with adults than larval stages. Additional
information about larval tolerance or the effects of
environmental variables on reproductive phases such
as gametogenesis and spawning is critical in order to
formulate effective measures to control L. fortunei.
We suggest that L. fortunei can withstand lower
limits of pH and calcium than those proposed by
Ricciardi (1998). The upper water temperature limits
were in the range suggested by Ricciardi (1998), but
tolerance to low dissolved oxygen was not possible to
Abiotic factors controlling the establishment 727
123

determine because oxygen depletion events coincided
with changes in other variables. Extreme conditions
in the Pantanal are likely responsible for maintaining
relatively low densities of L. fortunei, but the
populations persist and can lead to further expansion
into the more hospitable waters in upper Paraguay
River basin locations where constant vessel traffic
exerts a considerable propagule pressure and seasonal
extremes may be less marked.
Acknowledgments This work was supported by The National
Council for Scientific and Technological Development (PELD
and CTHIDRO programs), and Embrapa Pantanal. We are
grateful to the Pantanal Matogrossense National Park for the field
assistance, and the Federal University of Minas Gerais/ECMVS
and US Fish and Wildlife Service for support. We are also
grateful to Maria D. Oliveira, Egidia do Amaral, Waldomiro L.
Silva, and Isac Teixeira for technical assistance, and to the
students Viviane Eilers, Izabella Xavier, Claudiane Santos, and
Tatiane Cielo for the great collaboration in field work and
laboratory analyses. The valuable suggestions of two anonymous
referees are gratefully acknowledged.
References
Allen YC, Thompson BA, Ramcharan CW (1999) Growth and
mortality rates of the zebra mussel, Dreissena polymor-pha, in the lower Mississippi River. Can J Fish Aquat Sci
56:748–759
APHA (1998) Standard methods for the examination of water
and wastewater, 20th edn. American Public Health
Association, American Water Works Association, and
Water Environment Federation, Washington, DC, 1268 p
Baines SB, Fisher NS, Cole JJ (2005) Uptake of dissolved
organic matter (DOM) and its importance to metabolic
requirements of the zebra mussel, Dreissena polymorpha.
Limmnol Oceanogr 50:36–47
Baines SB, Fisher NS, Cole JJ (2007) Dissolved organic matter
and persistence of the invasive zebra mussel (Dreissenapolymorpha) under low food conditions. Limnol Ocea-
nogr 52:70–78
Boltovskoy D, Cataldo D (1999) Population dynamics of
Limnoperna fortunei, an invasive fouling mollusc, in
the Lower Parana River (Argentina). Biofouling 14:
255–263
Boltovskoy D, Sylvester F, Otaegui A et al (2009). Environ-
mental modulation of reproductive activity of the invasive
mussel Limnoperna fortunei: implications for antifouling
strategies. Austral Ecol. doi:10.1111/j.1442-9993.2009.
01974.x
Bossenbroek JM, Johnson LE, Peters B et al (2007) Forecast-
ing the expansion of Zebra Mussels in the United States.
Conserv Biol 21(3):800–810
Calheiros DF, Hamilton SK (1998) Limnological conditions
associated with natural fish kills in the Pantanal Wetland
of Brazil. Verh Internat Verein Limnol 26:2189–2193
Cataldo D, Boltovskoy D, Hermosa JL et al (2005) Tempera-
ture-dependent rates of larval development in Limnopernafortunei (Bivalvia:Mytilidae). J Moll Stud 71:41–46
Darrigran G, Penchaszadeh P, Damborenea MC (1999) The
reproductive cycle of Limnoperna fortunei (Dunker, 1857)
(Mytilidae) from a neotropical temperate locality.
J Shellfish Res 18(2):361–365
Darrigran G, Damborenea MC, Penchaszadeh P et al (2003)
Reproductive stabilization of Limnoperna fortunei (Biv-
alvia Mytilidae) after ten years of invasion in the Amer-
icas. J Shellfish Res 22:141–146
Deaton LE, Derby JGS, Subhedar N et al (1989) Osmorregu-
lation and salinity tolerance in two species of bivalve
mollusk: Limnoperna fortunei and Mytilopsis leucophae-ta. J Exp Mar Biol Ecol 133:67–79
Depretris PJ, Kempe S (1993) Carbon dynamics and sources in
the Parana River. Limnol Oceanogr 38:382–395
dos Santos CP, Wurdig NL, Mansur MCD (2005) Fases larvais
do mexilhao dourado Limnoperna fortunei (Dunker)
(Mollusca, Bivalvia, Mytilidae) na Bacia do Guaıba, Rio
Grande do Sul, Brasil. Rev Bras Zool 22:702–708
Drake JM, Bossenbroek JM (2004) The potential distribution
of Zebra Mussels in the United States. Bioscience
54(10):931–941
Gran G (1952) Determination of the equivalence point in
potentiometer titrations, Part II. Analyst 77:661–671
Guo Q (2006) Intercontinental biotic invasions: what can we
learn from native populations and habitats? Biol Invasions
8:1451–1459
Hamilton SK, Sippel SJ, Melack JM (1995) Oxygen depletion
and carbon dioxide and methane production in waters of
the Pantanal wetland of Brazil. Biogeochemistry
30:115–141
Hamilton SK, Sippel SJ, Calheiros DF et al (1997) An oxygen
depletion event and other biogeochemical effects of the
Pantanal wetland on the Paraguay River. Limnol Ocea-
nogr 42:257–272
Ho KC, Chow YL, Yau JTS (2003) Chemical and microbio-
logical qualities of the East River (Dongjiang) water, with
particular reference to drinking water supply in Hong
Kong. Chemosphere 52:1441–1450
Iwasaki K, Uryu Y (1998) Life cycle of a freshwater mytilid
mussel, Limnoperna fortunei, in Uji River, Kyoto. Jpn J
Malacol 57(2):105–113
Karatayev AY, Boltovskoy D, Padilla D et al (2007) Theinvasive bivalves Dreissena polimorpha and Limnopernafortunei: parallels, contrasts, potential spread and invasion
impacts. J Shellfish Res 26:205–213
Kempe S (1982) Long-term records of CO2 pressure fluctua-
tions in fresh waters. SCOPE/UNEP Sonderband
52:91–332
Kluza DA, McNyset M (2005) Ecological niche modeling of
aquatic invasive species. Aquat Invaders 16(1):1–7
Lee SY, Morton B (1983) The Hong Kong Mytilidae. In:
Morton B, Dudgeon D (eds) The malacofauna of Hong
Kong and Southern China II. Proceedings of the second
international workshop on the malacofauna of Hong Kong
and Southern China. Hong Kong University Press. Hong
Kong, pp 49–76
Madon SP, Schneider DW, Stoeckel JA et al (1998) Effects of
inorganic sediment and food concentrations on energetic
728 M. D. Oliveira et al.
123

processes of the zebra mussel, Dreissena polymorpha:
implications of growth in turbid rivers. Can J Fish Aquat
Sci 55:401–413
Magara Y, Matsui Y, Goto Y et al (2001) Invasion of the non
indigenous nuisance mussel, Limnoperna fortunei, into
water supply facilities in Japan. J Water Supply Res
Technol - AQUA 50:113–124
Marker AFH, Nush EA, Rai H et al (1981) The measurement of
photosynthetic pigments in freshwaters and standardiza-
tion of methods: Conclusions and recommendations. Arch
Hydrobiol Beih Ergebn Limnol 14:91–106
Montalto L, Marchese M (2003) Limnoperna fortunei (Dunker,
1857) (Bivalvia:Mytilidae) tolerance to temperature and
pH in experimental conditions. Neotropica 49:26–34
Morton B (1975) The colonization of Hong Kong’s water
supply system by Limnoperna fortunei (Dunker 1857)
(Bivalvia: Mytilacea) from China. Mal Rev 8:91–105
Morton B (1977) The population dynamics of Limnopernafortunei (Dunker 1857) (Bivalvia: Mytilacea) in Plover
Cove reservoir, Hong Kong. Malacologia 16:165–182
Morton B (1982) The reproductive cycle in Limnoperna for-tunei (Dunker 1857) (Bivalvia: Mytilidae) fouling Hong
Kong’s raw water supply system. Oceanol Limnol Sinica
13:312–325
Morton B (1987) Comparative life history tactics and sexual
strategies of the fresh and brackish water bivalve fauna of
Hong Kong and Southern China. Am Malacol Bull
5:91–99
Morton B (1996) The aquatic nuisance species problem: a
global perspective and review. In: D’Itri FM (ed) Zebra
mussels and other aquatic nuisance species. Ann Arbor
Press, Chelsea, pp 1–54
Nagaya K, Matsui Y, Ohira H et al (2001) Attachment strength
of an adhesive nuisance mussel, Limnoperna fortunei,against water flow. Biofouling 17:263–274
Oliveira MD (2009) Factors controlling the density and
potential distribution of golden mussel (Limnoperna for-tunei Dunker 1857) in the Upper Paraguay Basin. Ph.D.
thesis, Universidade Federal de Minas Gerais, Belo Hor-
izonte, Brazil
Oliveira MD, Calheiros DF (2000) Flood pulse influence on a
phytoplankton community in south Pantanal floodplain,
Brazil. Hydrobiologia 427:101–112
Oliveira MD, Hamilton SK, Jacobi CM (2010a) Forecasting
the expansion of the invasive golden mussel Limnopernafortunei in Brazilian and North American rivers based on
its occurrence in the Paraguay River and Pantanal wetland
of Brazil. Aquat Invasions 5(1):59–73
Oliveira MD, Hamilton SK, Calheiros DF, Jacobi CM (2010b)
Oxygen depletion events control the invasive golden mus-
sel (Limnoperna fortunei) in a tropical floodplain. Wetlands
30(4):705–716. doi:10.1007/s13157-010-0081-3
Oliveira MD, Takeda AM, Barros LF et al (2006) Invasion by
Limnoperna fortunei (Dunker, 1857) (Bivalvia Mytilidae)
of the Pantanal wetland, Brazil. Biol Invasions 8:97–104
Pastorino G, Darrigran G, Martin SM et al (1993) Limnopernafortunei (Dunker, 1857) (Mytilidae), nuevo bivalvo in-
vasor en aguas del Rio de La Plata. Neotropica 39:34
PCBAP (Plano de Conservacao da Bacia do Alto Paraguai)
(1997) Analise Integrada e prognostico da Bacia do alto
Paraguai.Programa Nacional do Meio Ambiente. Brasılia:
PNMA. V.3. 367 p
Peterson AT, Vieglais DA (2001) Predicting species invasions
using ecological niche modeling: new approaches from
bioinformatics attack a pressing problem. Bioscience
51:363–371
Ricciardi A (1998) Global range expansion of the Asian mussel
Limnoperna fortunei (Mytilidae): another fouling threat to
freshwater systems. Biofouling 13:97–106
Schneider DW, Madon SP, Stoeckel JA et al (1998) Seston
quality controls zebra mussel (Dreissena polymorpha)
energetics in turbid rivers. Oecologia 117:331–341
Silva DP (2006) Aspectos bioecologicos do mexilhao dourado
Limnoperna fortunei (Bivalvia, Mytilidae) (Dunker,
1857). Ph.D. thesis. Universidade Federal do Parana.
Curitiba, 138 p
Wang HZ, Xie Z, Wu XP et al (1999) A preliminary study of
zoobenthos in the Poyang Lake, the largest freshwater
lake of China, and its adjoining reaches of Changjiang
River. Acta Hydrobiol Sinica 23:132–138
Wang HZ, Xu QQ, Cui YD et al (2007) Macrozoobenthic
community of Poyang Lake, the largest freshwater lake of
China, in the Yangtze floodplain. Limnology 8:65–71
Wetzel RA, Likens GE (2001) Limnological analyses.
Springer, New York 391 p
Wilkinson L (2004) Systat 11. SYSTAT Software Inc, San Jose
Xie Z, Liang YL, Wang J et al (1999) Preliminary studies of
macroinvertebrates of the Mainstream of the Changjiang
(Yangtze) River. Acta Hydrobiol Sinica 23:148–157
Zhang SR, Lu XX, Higgitt DL et al (2007) Water chemistry of
the Zhujiang (Pearl River): natural processes and anthro-
pogenic influences. J Geophys Res Earth Surf 112:1–17
Abiotic factors controlling the establishment 729
123




















