
A novel method for preparation of HAMLET-like protein complexes
7
Research paper A novel method for preparation of HAMLET-like protein complexes Sergei E. Permyakov a, b, * , Ekaterina L. Knyazeva a, * , Marina V. Leonteva a, b , Roman S. Fadeev c , Aleksei V. Chekanov c , Andrei P. Zhadan a , Anders P. Håkansson d , Vladimir S. Akatov c , Eugene A. Permyakov a, b a Institute for Biological Instrumentation of the Russian Academy of Sciences, Pushchino, Moscow Region 142290, Russia b Department of Biomedical Engineering, Pushchino State University, Pushchino, Moscow Region 142290, Russia c Institute of Theoretical and Experimental Biophysics of the Russian Academy of Sciences, Pushchino, Moscow Region 142290, Russia d Department of Microbiology and Immunology, University at Buffalo, Buffalo, NY 14214, USA article info Article history: Received 3 March 2011 Accepted 4 May 2011 Available online xxx Keywords: a-lactalbumin Oleic acid HAMLET Cytotoxicity Bactericidal activity abstract Some natural proteins induce tumor-selective apoptosis. a-Lactalbumin (a-LA), a milk calcium-binding protein, is converted into an antitumor form, called HAMLET/BAMLET, via partial unfolding and asso- ciation with oleic acid (OA). Besides triggering multiple cell death mechanisms in tumor cells, HAMLET exhibits bactericidal activity against Streptococcus pneumoniae. The existing methods for preparation of active complexes of a-LA with OA employ neutral pH solutions, which greatly limit water solubility of OA. Therefore these methods suffer from low scalability and/or heterogeneity of the resulting a-LA e OA samples. In this study we present a novel method for preparation of a-LA e OA complexes using alkaline conditions that favor aqueous solubility of OA. The unbound OA is removed by precipitation under acidic conditions. The resulting sample, bLAeOA-45, bears 11 OA molecules and exhibits physico-chemical properties similar to those of BAMLET. Cytotoxic activities of bLAeOA-45 against human epidermoid larynx carcinoma and S. pneumoniae D39 cells are close to those of HAMLET. Treatment of S. pneumoniae with bLAeOA-45 or HAMLET induces depolarization and rupture of the membrane. The cells are markedly rescued from death upon pretreatment with an inhibitor of Ca 2þ transport. Hence, the acti- vation mechanisms of S. pneumoniae death are analogous for these two complexes. The developed express method for preparation of active a-LA e OA complex is high-throughput and suited for devel- opment of other protein complexes with low-molecular-weight amphiphilic substances possessing valuable cytotoxic properties. Ó 2011 Published by Elsevier Masson SAS. 1. Introduction Despite promin Q1 ent achievements in development of novel anticancer therapies, the need for new anticancer drug candidates is still urgent. One of the most intriguing classes of substances with antitumor therapeutic potential is a set of natural proteins, selec- tively inducing apoptosis in tumor cells: apoptin, E4orf4, NS1, mda- 7, TRAIL, HAMLET and ELOA (for reviews, see [3,4]). Some of them have shown positive results in pre-clinical and clinical studies (mda-7, TRAIL and HAMLET). Aside from direct use of these proteins for therapeutic purposes, they provide valuable informa- tion on molecular mechanisms used to defeat the biochemical machinery of malignant cells. In this respect, HAMLET is of special interest due to its ability to attack several critical organelles (mitochondrion, proteasome and nucleosome), activating different death pathways in tumor cells (for reviews, see [4,5]). HAMLET represents a complex between a small calcium-binding protein from human milk, a-lactalbumin (a-LA; for review, see [6]), and oleic acid (OA) [1]. In the lactating mammary gland, a-LA serves as a regulatory subunit of the lactose synthase enzyme complex [7]. The binding of OA to a-LA following a specific procedure converts it into a form (HAMLET, Human Alpha-lactalbumin Made LEthal to Abbreviations: OA, oleic acid (C 18:1:9cis ); CMC, critical micelle concentration; a-LA, a-lactalbumin; intact a-LA, a-lactalbumin isolated from milk; hLA, human a-lactalbumin; bLA, bovine a-lactalbumin; HAMLET(BAMLET), complex of human(bovine) a-lactalbumin with oleic acid, prepared as described in [1,2]; bLAeOA-45, complex of bovine a-lactalbumin with oleic acid, prepared as described in Materials and methods; reference bLA, sample of bovine a-lactalbumin subjected to procedures described for preparation of bLAeOA-45, skipping addition of oleic acid; MS, mass spectrometry; CD, circular dichroism; l max , maximum position of fluorescence emission spectrum; T 1/2 , mid-transition temperature; HEp- 2, human epidermoid larynx carcinoma cells; CFUs, colony-forming units. * Corresponding authors. Institute for Biological Instrumentation of the Russian Academy of Sciences, Institutskaya 7, Pushchino, Moscow Region 142290, Russia. Tel.: þ7 4967 73 41 35; fax: þ7 4967 33 05 22. E-mail addresses: [email protected] (S.E. Permyakov), elknyazeva@rambler. ru (E.L. Knyazeva). Contents lists available at ScienceDirect Biochimie journal homepage: www.elsevier.com/locate/biochi 0300-9084/$ e see front matter Ó 2011 Published by Elsevier Masson SAS. doi:10.1016/j.biochi.2011.05.002 Biochimie xxx (2011) 1e7 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108 109 110 BIOCHI3570_proof ■ 16 May 2011 ■ 1/7 Please cite this article in press as: S.E. Permyakov, et al., A novel method for preparation of HAMLET-like protein complexes, Biochimie (2011), doi:10.1016/j.biochi.2011.05.002
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of A novel method for preparation of HAMLET-like protein complexes

Q1
lable at ScienceDirect
Biochimie xxx (2011) 1e7
12345678910111213141516171819202122232425262728293031323334353637383940414243444546474849505152535455
BIOCHI3570_proof ■ 16 May 2011 ■ 1/7
Contents lists avai
Biochimie
journal homepage: www.elsevier .com/locate/biochi
56575859606162636465666768
Research paper
A novel method for preparation of HAMLET-like protein complexes
Sergei E. Permyakov a,b,*, Ekaterina L. Knyazeva a,*, Marina V. Leonteva a,b, Roman S. Fadeev c,Aleksei V. Chekanov c, Andrei P. Zhadan a, Anders P. Håkansson d, Vladimir S. Akatov c,Eugene A. Permyakov a,b
a Institute for Biological Instrumentation of the Russian Academy of Sciences, Pushchino, Moscow Region 142290, RussiabDepartment of Biomedical Engineering, Pushchino State University, Pushchino, Moscow Region 142290, Russiac Institute of Theoretical and Experimental Biophysics of the Russian Academy of Sciences, Pushchino, Moscow Region 142290, RussiadDepartment of Microbiology and Immunology, University at Buffalo, Buffalo, NY 14214, USA
6970717273747576777879
a r t i c l e i n f o
Article history:Received 3 March 2011Accepted 4 May 2011Available online xxx
Keywords:a-lactalbuminOleic acidHAMLETCytotoxicityBactericidal activity
Abbreviations: OA, oleic acid (C18:1:9cis); CMC, cra-LA, a-lactalbumin; intact a-LA, a-lactalbumin isolaa-lactalbumin; bLA, bovine a-lactalbumin; HAMhuman(bovine) a-lactalbumin with oleic acid, prepbLAeOA-45, complex of bovine a-lactalbumin widescribed in Materials and methods; reference bLA, sasubjected to procedures described for preparation of bof oleic acid; MS, mass spectrometry; CD, circularposition of fluorescence emission spectrum; T1/2, mid-2, human epidermoid larynx carcinoma cells; CFUs, c* Corresponding authors. Institute for Biological In
Academy of Sciences, Institutskaya 7, Pushchino, MoTel.: þ7 4967 73 41 35; fax: þ7 4967 33 05 22.
E-mail addresses:[email protected] (S.E. Permru (E.L. Knyazeva).
0300-9084/$ e see front matter � 2011 Published bydoi:10.1016/j.biochi.2011.05.002
8081828384858687
Please cite this article in press as: S.E. Permydoi:10.1016/j.biochi.2011.05.002
a b s t r a c t
Some natural proteins induce tumor-selective apoptosis. a-Lactalbumin (a-LA), a milk calcium-bindingprotein, is converted into an antitumor form, called HAMLET/BAMLET, via partial unfolding and asso-ciation with oleic acid (OA). Besides triggering multiple cell death mechanisms in tumor cells, HAMLETexhibits bactericidal activity against Streptococcus pneumoniae. The existing methods for preparation ofactive complexes of a-LAwith OA employ neutral pH solutions, which greatly limit water solubility of OA.Therefore these methods suffer from low scalability and/or heterogeneity of the resulting a-LA e OAsamples. In this study we present a novel method for preparation of a-LA e OA complexes using alkalineconditions that favor aqueous solubility of OA. The unbound OA is removed by precipitation under acidicconditions. The resulting sample, bLAeOA-45, bears 11 OA molecules and exhibits physico-chemicalproperties similar to those of BAMLET. Cytotoxic activities of bLAeOA-45 against human epidermoidlarynx carcinoma and S. pneumoniae D39 cells are close to those of HAMLET. Treatment of S. pneumoniaewith bLAeOA-45 or HAMLET induces depolarization and rupture of the membrane. The cells aremarkedly rescued from death upon pretreatment with an inhibitor of Ca2þ transport. Hence, the acti-vation mechanisms of S. pneumoniae death are analogous for these two complexes. The developedexpress method for preparation of active a-LA e OA complex is high-throughput and suited for devel-opment of other protein complexes with low-molecular-weight amphiphilic substances possessingvaluable cytotoxic properties.
� 2011 Published by Elsevier Masson SAS.
88
89 90919293941. Introduction
Despite prominent achievements in development of novelanticancer therapies, the need for new anticancer drug candidates
itical micelle concentration;ted from milk; hLA, humanLET(BAMLET), complex ofared as described in [1,2];th oleic acid, prepared asmple of bovine a-lactalbuminLAeOA-45, skipping additiondichroism; lmax, maximumtransition temperature; HEp-olony-forming units.strumentation of the Russianscow Region 142290, Russia.
yakov), elknyazeva@rambler.
Elsevier Masson SAS.
9596979899
100101102103104105106107108
akov, et al., A novel method
is still urgent. One of the most intriguing classes of substances withantitumor therapeutic potential is a set of natural proteins, selec-tively inducing apoptosis in tumor cells: apoptin, E4orf4, NS1, mda-7, TRAIL, HAMLET and ELOA (for reviews, see [3,4]). Some of themhave shown positive results in pre-clinical and clinical studies(mda-7, TRAIL and HAMLET). Aside from direct use of theseproteins for therapeutic purposes, they provide valuable informa-tion on molecular mechanisms used to defeat the biochemicalmachinery of malignant cells. In this respect, HAMLET is of specialinterest due to its ability to attack several critical organelles(mitochondrion, proteasome and nucleosome), activating differentdeath pathways in tumor cells (for reviews, see [4,5]).
HAMLET represents a complex between a small calcium-bindingprotein from human milk, a-lactalbumin (a-LA; for review, see [6]),and oleic acid (OA) [1]. In the lactating mammary gland, a-LA servesas a regulatory subunit of the lactose synthase enzyme complex [7].The binding of OA to a-LA following a specific procedure converts itinto a form (HAMLET, Human Alpha-lactalbumin Made LEthal to
109110
for preparation of HAMLET-like protein complexes, Biochimie (2011),

S.E. Permyakov et al. / Biochimie xxx (2011) 1e72
111112113114115116117118119120121122123124125126127128129130131132133134135136137138139140141142143144145146147148149150151152153154155156157158159160161162163164165166167168169170171172173174175
176177178179180181182183184185186187188189190191192193194195196197198199200201202203204205206207208209210211212213214215216217218219220221222223224225226227228229230231
BIOCHI3570_proof ■ 16 May 2011 ■ 2/7
Tumor cells), which induces death of tumor cells and undifferenti-ated cells, but spares healthy and mature cells in cell culture andtissue [1,8e12]. Besides its tumoricidal activity, this complex alsoexhibits bactericidal activityagainst antibiotic-sensitive and resistantstrains of Streptococcus pneumoniae [13,14]. Although a-LA is able tobindother fatty acids [15], C18:1:9cis (OA) or C18:1:11cis (vaccenic acid) isrequired to form an a-LA complex with pronounced antitumoractivity [16]. Meanwhile, human a-LA can be replaced by bovine,equine, porcine or caprine a-LA without loss of the complex activity[2]. Moreover, binding of OA to Ca2þ-binding equine lysozyme alsogives rise to tumoricidal activity, analogous to HAMLET [17]. This factand similar evidences discussed by Spolaore et al. [18] indicate thatthe cytotoxic effect ofHAMLET ismostly due toOA,which is cytotoxicby itself [19e21],while the proteinaceous component of the complexmainly serves as a carrier of the poorly water soluble OA [22e24].Nevertheless, there are evidences that a-LA within HAMLET directlyaffects some of key cellular processes [25], implying that bothcomponents of the complex are important for its cytotoxicity.
The bindingofOA to a-LA requires partial unfoldingof the protein[1], which can be achieved via Ca2þ depletion [26] or thermaldenaturation. The existing procedures for preparing tumoricidal a-LAeOAcomplexes differ in theprocedure of application ofOA, in themethod of a-LA unfolding, and in the OA concentration used. Theoriginal method employs chromatography of Ca2þ-free a-LA ona DEAE-Trisacryl� M column preconditioned with OA followed byelution of the complex by high salt concentration [1]. Othermethodsare based upon direct binding of dissolved OA by Ca2þ-depleted[21,27] or thermally denatured [28] a-LA. Only the method byKnyazeva et al. takes into account low water solubility of OA atneutral pH via use of OA concentrations below its effective criticalmicelle concentration (CMC). The absence of such a control of CMCvalues in other methods is fraught with formation of extendedphases of OA [22e24], making the a-LA e OA system highlyheterogeneous. The heterogeneity of this system is documented inthe work by Spolaore et al. [18]. Possibly this circumstance partlyrationalizes the fact that different oligomeric states of a-LA andvarious OA to a-LA molar ratios were reported for active complexesprepared using different approaches (for review, see [18]).
The CMC values of OA do not exceed 70 mM under optimizedsolution conditions used in the method by Knyazeva et al. [21]. Thelow water solubility of OA at neutral pH limits scalability of thismethod. In the present work we present a novel method forpreparation of biologically active a-LA e OA complexes based onthe use of alkaline solvent conditions, greatly favoring aqueoussolubility of OA, thereby allowing us to overcome the scalabilitylimitation of the previous method. The developed low-cost expressmethod enables preparation under laboratory conditions of quan-tities of active sample suitable for animal testing and pre-clinicalstudies. Furthermore, the use of extreme alkaline conditionsenables the extension of this method to other proteins, thusopening up wide opportunities for preparation of proteincomplexes with fatty acids or other low-molecular-weight amphi-philic substances with valuable cytotoxic properties. The proposedmethod of dissolving of otherwise hardly water soluble cytotoxicsubstances into the hydrophobic interior of natural proteins can beexploited for cell delivery of these substances.
232233234235236237238239240
2. Materials and methods
2.1. Materials
Human epidermoid larynx carcinoma (HEp-2) cells were fromthe Russian Cell Culture Collection (Institute of Cytology, RussianAcademy of Sciences, St. Petersburg).
Please cite this article in press as: S.E. Permyakov, et al., A novel methoddoi:10.1016/j.biochi.2011.05.002
Human a-lactalbumin (hLA) was isolated frommilk as described[29,30]. Ca2þ-depleted bovine a-lactalbumin (bLA) and oleic acid(C18:1:9cis) were from SigmaeAldrich Co. Chemistry grade 96% (v/v)ethanol was treated with K2MnO4 and activated coal, followed bydouble distillation; the absence of organic impurities wasconfirmed spectrophotometrically. Stock solution of OA wasprepared by sonication of an ethanol suspension of OA for 3 min,and stored at room temperature. HAMLET and BAMLET wereprepared using the original procedure [1,2]. Concentrations of a-LAwere evaluated using extinction coefficients A1%
1 cm;280 of 20.1 and18.2 for bLA/BAMLET and hLA/HAMLET, respectively [31,32].
Ultra-grade H3BO3 was bought from Merck. Ultra-gradeammonium bicarbonate, CaCl2 standard solution, DEAE-Trisacryl�
M, DMEM culture medium, crystal violet, Ruthenium Red andpropidium iodide were from SigmaeAldrich Co. Fetal bovine serumwas from HyClone. Todd-Hewitt medium and Bacto-Yeast extractwere from BD Biosciences. DiBAC4(3) was from Invitrogen. EDTAstandard solution and dialysis tubing were from Thermo FisherScientific Inc. Water with conductivity of 18.2 MU cm was used.HPLC grade acetonitrile was from Panreac.
2.2. Estimation of the effective critical micelle concentration of OA
Estimation of the effective critical micelle concentration ofOA was performed as previously described [21]. The appearance ofOA micelles was monitored photometrically at 335 nm. The lowestestimate of OA binding capacity of a-LA, nmax, was found from thedependence of effective CMC of OA upon protein concentration, [P](CMC0 is CMC value of free OA):
CMC ¼ CMC0 þ nmax$½P� (1)
2.3. Preparation of bLAeOA-45 state
6 ml of 0.6 mM Ca2þ-free bLA at pH 12.0 (5 mM KOH, 1 mMEDTA) was titrated at 45 �C by 12 ml aliquots of 1 M OA solution in96% ethanol up to the effective CMC of OA, 22mM. The unbound OAwas separated via acidification of the solution down to pH 2.0 andcentrifugation at 12,000g for 30 min at 40 �C. The free fromunbound OAwater phase was collected, and its pH was adjusted byKOH to w5.5. The solution was triply dialyzed against 100-foldvolume of distilled water, freeze-dried and stored at�18 �C. Bovinea-lactalbumin, subjected to the described procedure, but skippingaddition of OA, will be referred to as “reference bLA”. The integrityof bLAeOA-45 and reference bLA was confirmed by SDS-PAGE.
2.4. Quantitation of the content of OA in protein samples
Quantitation of the content of OA in protein samples was per-formed using LC/MS system LCMS-2010EV (Shimadzu Co.), ESI probecoupled to a single quadrupole detector. Sample of 0.1e1 mM a-LA in50:50 (v/v)mixture of 20mMammoniumbicarbonate pH8.5 (bufferA) and acetonitrile was applied to Shimadzu Silica 100 2.5 mm RP4,4 � 50 mm column at 40 �C. OA was eluted with buffer A:aceto-nitrile 20:80 (v/v) at 50 ml/minflow rate and infused intoMSdetector(interface 4 kV, negative mode, SIM mode 281 � 0.5 m). The chro-matogramwas integrated and quantitated using a calibration curve.The calibrationwas carriedout using0.01e100mMOA in96%ethanol.
2.5. Secondary structure content in a-LA
Secondary structure content in a-LA was determined from far-UV circular dichroism (CD) spectra using the CDPro softwarepackage [33] as previously described [21].
for preparation of HAMLET-like protein complexes, Biochimie (2011),
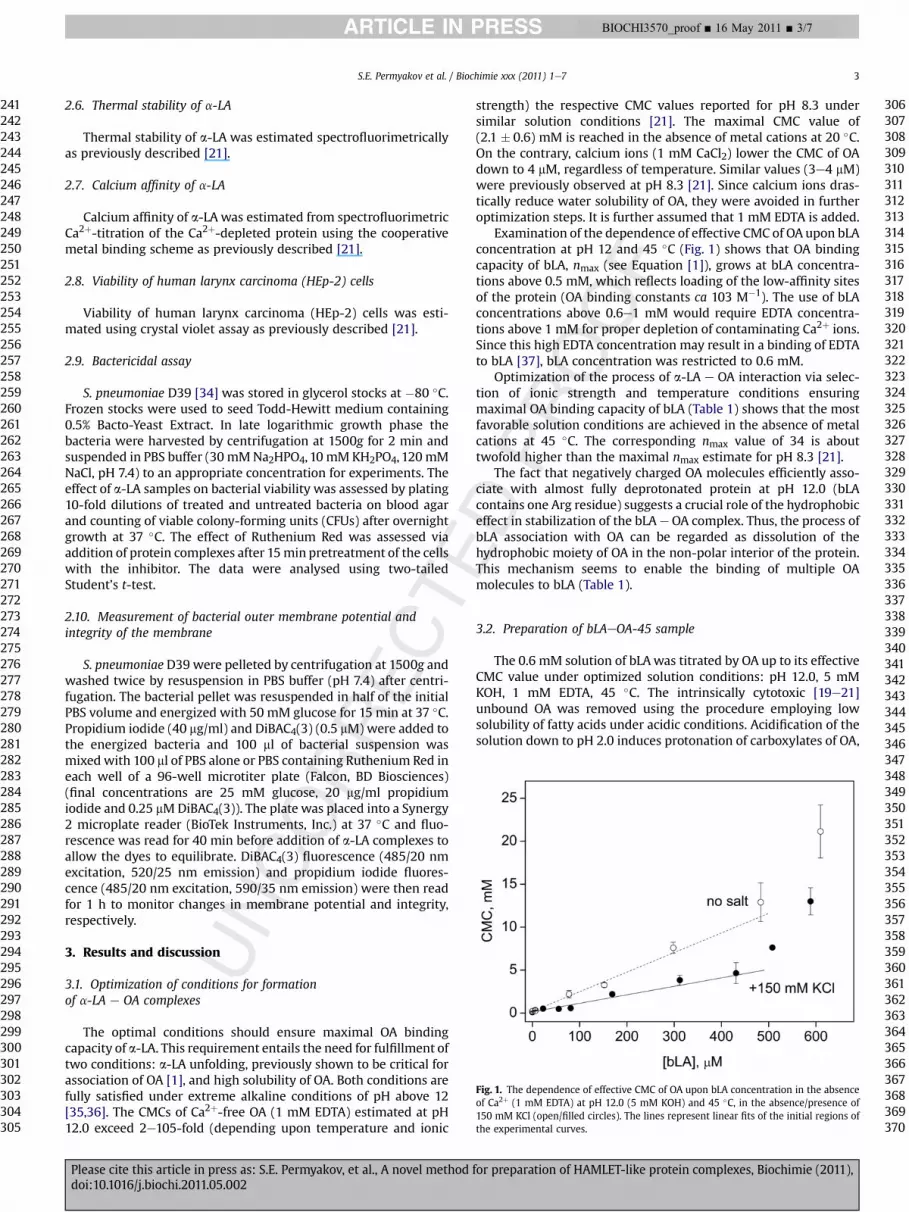
Fig. 1. The dependence of effective CMC of OA upon bLA concentration in the absenceof Ca2þ (1 mM EDTA) at pH 12.0 (5 mM KOH) and 45 �C, in the absence/presence of150 mM KCl (open/filled circles). The lines represent linear fits of the initial regions ofthe experimental curves.
S.E. Permyakov et al. / Biochimie xxx (2011) 1e7 3
241242243244245246247248249250251252253254255256257258259260261262263264265266267268269270271272273274275276277278279280281282283284285286287288289290291292293294295296297298299300301302303304305
306307308309310311312313314315316317318319320321322323324325326327328329330331332333334335336337338339340341342343344345346347348349350351352353354355356357358359360361362363364365366367368369370
BIOCHI3570_proof ■ 16 May 2011 ■ 3/7
2.6. Thermal stability of a-LA
Thermal stability of a-LA was estimated spectrofluorimetricallyas previously described [21].
2.7. Calcium affinity of a-LA
Calcium affinity of a-LA was estimated from spectrofluorimetricCa2þ-titration of the Ca2þ-depleted protein using the cooperativemetal binding scheme as previously described [21].
2.8. Viability of human larynx carcinoma (HEp-2) cells
Viability of human larynx carcinoma (HEp-2) cells was esti-mated using crystal violet assay as previously described [21].
2.9. Bactericidal assay
S. pneumoniae D39 [34] was stored in glycerol stocks at �80 �C.Frozen stocks were used to seed Todd-Hewitt medium containing0.5% Bacto-Yeast Extract. In late logarithmic growth phase thebacteria were harvested by centrifugation at 1500g for 2 min andsuspended in PBS buffer (30 mMNa2HPO4, 10 mMKH2PO4, 120mMNaCl, pH 7.4) to an appropriate concentration for experiments. Theeffect of a-LA samples on bacterial viability was assessed by plating10-fold dilutions of treated and untreated bacteria on blood agarand counting of viable colony-forming units (CFUs) after overnightgrowth at 37 �C. The effect of Ruthenium Red was assessed viaaddition of protein complexes after 15min pretreatment of the cellswith the inhibitor. The data were analysed using two-tailedStudent’s t-test.
2.10. Measurement of bacterial outer membrane potential andintegrity of the membrane
S. pneumoniae D39 were pelleted by centrifugation at 1500g andwashed twice by resuspension in PBS buffer (pH 7.4) after centri-fugation. The bacterial pellet was resuspended in half of the initialPBS volume and energized with 50 mM glucose for 15 min at 37 �C.Propidium iodide (40 mg/ml) and DiBAC4(3) (0.5 mM) were added tothe energized bacteria and 100 ml of bacterial suspension wasmixed with 100 ml of PBS alone or PBS containing Ruthenium Red ineach well of a 96-well microtiter plate (Falcon, BD Biosciences)(final concentrations are 25 mM glucose, 20 mg/ml propidiumiodide and 0.25 mMDiBAC4(3)). The plate was placed into a Synergy2 microplate reader (BioTek Instruments, Inc.) at 37 �C and fluo-rescence was read for 40 min before addition of a-LA complexes toallow the dyes to equilibrate. DiBAC4(3) fluorescence (485/20 nmexcitation, 520/25 nm emission) and propidium iodide fluores-cence (485/20 nm excitation, 590/35 nm emission) were then readfor 1 h to monitor changes in membrane potential and integrity,respectively.
3. Results and discussion
3.1. Optimization of conditions for formationof a-LA e OA complexes
The optimal conditions should ensure maximal OA bindingcapacity of a-LA. This requirement entails the need for fulfillment oftwo conditions: a-LA unfolding, previously shown to be critical forassociation of OA [1], and high solubility of OA. Both conditions arefully satisfied under extreme alkaline conditions of pH above 12[35,36]. The CMCs of Ca2þ-free OA (1 mM EDTA) estimated at pH12.0 exceed 2e105-fold (depending upon temperature and ionic
Please cite this article in press as: S.E. Permyakov, et al., A novel methoddoi:10.1016/j.biochi.2011.05.002
strength) the respective CMC values reported for pH 8.3 undersimilar solution conditions [21]. The maximal CMC value of(2.1 � 0.6) mM is reached in the absence of metal cations at 20 �C.On the contrary, calcium ions (1 mM CaCl2) lower the CMC of OAdown to 4 mM, regardless of temperature. Similar values (3e4 mM)were previously observed at pH 8.3 [21]. Since calcium ions dras-tically reduce water solubility of OA, they were avoided in furtheroptimization steps. It is further assumed that 1 mM EDTA is added.
Examination of the dependence of effective CMC of OA upon bLAconcentration at pH 12 and 45 �C (Fig. 1) shows that OA bindingcapacity of bLA, nmax (see Equation [1]), grows at bLA concentra-tions above 0.5 mM, which reflects loading of the low-affinity sitesof the protein (OA binding constants ca 103 M�1). The use of bLAconcentrations above 0.6e1 mM would require EDTA concentra-tions above 1 mM for proper depletion of contaminating Ca2þ ions.Since this high EDTA concentration may result in a binding of EDTAto bLA [37], bLA concentration was restricted to 0.6 mM.
Optimization of the process of a-LA e OA interaction via selec-tion of ionic strength and temperature conditions ensuringmaximal OA binding capacity of bLA (Table 1) shows that the mostfavorable solution conditions are achieved in the absence of metalcations at 45 �C. The corresponding nmax value of 34 is abouttwofold higher than the maximal nmax estimate for pH 8.3 [21].
The fact that negatively charged OA molecules efficiently asso-ciate with almost fully deprotonated protein at pH 12.0 (bLAcontains one Arg residue) suggests a crucial role of the hydrophobiceffect in stabilization of the bLA e OA complex. Thus, the process ofbLA association with OA can be regarded as dissolution of thehydrophobic moiety of OA in the non-polar interior of the protein.This mechanism seems to enable the binding of multiple OAmolecules to bLA (Table 1).
3.2. Preparation of bLAeOA-45 sample
The 0.6 mM solution of bLAwas titrated by OA up to its effectiveCMC value under optimized solution conditions: pH 12.0, 5 mMKOH, 1 mM EDTA, 45 �C. The intrinsically cytotoxic [19e21]unbound OA was removed using the procedure employing lowsolubility of fatty acids under acidic conditions. Acidification of thesolution down to pH 2.0 induces protonation of carboxylates of OA,
for preparation of HAMLET-like protein complexes, Biochimie (2011),

Table 1The dependence of OA binding capacity of bLA (nmax) at pH 12.0 (5 mM KOH) uponcalcium concentration, ionic strength and temperature. Protein concentration was0.6 mM.
Temperature 1 mM CaCl2 1 mM EDTA
nmax, 150 mM KCl nmax, no salt nmax, 150 mM KCl
20 �C 0 18 � 1 26 � 245 �C 0 34 � 5 22 � 3
S.E. Permyakov et al. / Biochimie xxx (2011) 1e74
371372373374375376377378379380381382383384385386387388389390391392393394395396397398399400401402403404405406407408409410411412413414415416417418419420421422423424425426427428429430431432433434435
436437438439440441442443444445446447448449450451452453454455456457458459460461462463464465466467468469470471472473474475476477478479480481482483484485486487488489490
BIOCHI3570_proof ■ 16 May 2011 ■ 4/7
resulting in its precipitation. The precipitates were removed viacentrifugation. The resulting sample was adjusted to pH w5.5,dialyzed against water and freeze-dried. The bLA sample subjectedto the same procedures, but without addition of OA, will be referredto as “reference bLA”.
3.3. Physico-chemical characterization of bLAeOA-45 complex
The number of OA molecules per a-LA molecule in bLAeOA-45sample estimated by LC/MS is 11, which is comparable to thecontent of OA in an LAeOA-45 sample of human a-LA [21]. Thus,about one third of the initially bound OA molecules are retained inbLAeOA-45 sample. Notably, examination of the samples of otherproteins binding OA only non-specifically, which were subjected tothe same procedures used for preparation of bLAeOA-45 sample,have shown for some of them at least an order of magnitude lowerOA content. This fact confirms efficacy of the acidic method of OAremoval and indicates that bLAeOA-45 sample retains OA mole-cules due to their specific binding.
The CD estimates of a-helical content for Ca2þ-depleted forms ofdifferent bLA states at 5 �C (pH 8.3) are shown in Table 2. ReferencebLA exhibits a-helical content lowered by (4.6 � 0.7)%, evidencingsome structural perturbations caused by the procedures used forpreparation of bLAeOA-45 sample. At the same time, the bLAeOA-45 state has the same a-helical content as intact protein. Hence,modification of bLA with OA improves its a-helicity. An analogouseffect is observed for BAMLET (Table 2) and hLA e OA complexes[21]. This effect is also observed at 45 �C.
Spectrofluorimetric measurement of thermal unfolding of theCa2þ-free bLAeOA-45 shows that its thermal stability is equivalentto that of the BAMLET state, being lowered by about 8 �C withrespect to both intact and reference bLA samples (Table 2). Thiseffect is in agreement with the previous data, evidencing a decreasein stability of apo-a-LA upon association with OA [21,38]. Thelowered stabilities of the Ca2þ-free bLA complexes with OA are inline with red shifts (9e12 nm) of their fluorescence emissionspectra (Table 2), which indicate higher accessibility of emittingtryptophans to the solvent. A similar, but less pronounced, effect(7 nm red shift) is observed for reference bLA. Calcium-binding(1 mM CaCl2) lowers this effect to 5 nm. The structural perturba-tions in reference bLA affect protein’s secondary structure and theenvironment of emitting tryptophans, but do not change thermalstability of bLA.
Calcium binding (1 mM CaCl2) renders all bLA states studiedequally stable (mid-transition temperatures of 61e62 �C). Despite
Table 2Physico-chemical properties of various states of bLA: a-helical content, maximum positioCa2þ-depleted protein (1 mM EDTA), equilibrium Ca2þ association constant (K) and Hill c
Protein state a (5 �C), % lmax (10 �C), nm
Intact bLA 31.0 � 0.5 324.6 � 0.1Reference bLA 26.4 � 0.5 331.4 � 0.1bLAeOA-45 31.0 � 0.5 336.2 � 0.1BAMLET 38.4 � 0.5 333.6 � 0.1
Please cite this article in press as: S.E. Permyakov, et al., A novel methoddoi:10.1016/j.biochi.2011.05.002
that, fluorescence spectrum of bLAeOA-45 is red-shifted by 11 nmin comparison with intact bLA and BAMLET. Hence, in the case ofbLAeOA-45, the association with OA increases solvent accessibilityof emitting tryptophans of the Ca2þ-bound protein, withoutaffecting its thermal stability.
Spectrofluorimetric measurements of calcium affinity of variousbLA states at 45 �C show that association of OA does not affectprotein calcium affinity, but decreases cooperativity of Ca2þ-binding by bLAeOA-45 (Table 2). Since calcium affinity data forbLAeOA-45 and reference bLA are indistinguishable, the modifi-cation of the protein by OA does not affect calcium affinity of bLA.
Overall, our data show that conformational and physico-chemical properties of bLAeOA-45 state are similar to those ofBAMLET.
3.4. Cytotoxic effects of bLAeOA-45 upon HEp-2and S. pneumoniae D39 cells
The concentration dependencies of the cytotoxicites of bLAeOA-45, HAMLET and respective control samples on human larynxcarcinoma (HEp-2) cells in vitro are shown in Fig. 2. Intact andreference bLA at concentrations up to 1 mMwere shown to be non-toxic. Oleic acid exerts toxic effect on HEp-2 cells only at concen-trations above 300 mM. The IC50 values for bLAeOA-45 (about20 mM) and HAMLET (40 mM) differ insignificantly.
The comparison of bactericidal activities of bLAeOA-45 andHAMLET against S. pneumoniae D39 is shown in Table 3. In therange of protein concentrations of 50e150 mg/ml (3.5e10.6 mM)bLAeOA-45 reduces viability of the bacteria by 2.4e6.2 orders ofmagnitude, which is close to the bactericidal activity of HAMLET.Intact a-LA at a concentration of 150 mg/ml does not exert anybactericidal activity. To evaluate the specific contribution of OA tothe bactericidal activity, we exposed bacteria to a concentration ofOA, equivalent to the content of OA in 10.6 mM of bLAeOA-45sample. An OA concentration of 106 mM exhibits a bactericidalactivity of 6.3� 0.2 orders of magnitude, which is similar to activityfor 10.6 mM of bLAeOA-45. Hence, the cytotoxic effect of bLAeOA-45 sample could be ascribed primarily to the toxic action of OA. Thesame conclusion follows from the cytotoxicity data presented inFig. 2 and the previous data on the hLA e OA complexes [21]. Thesefacts favor the view that the proteinaceous component of the LA e
OA complexes mainly serves as a carrier of the otherwise poorlysoluble cytotoxic component, oleic acid.
Summarizing, bLAeOA-45 and HAMLET samples are equallyeffective with respect to both HEp-2 and S. pneumoniae D39 cells.
3.5. The initial stages of bLAeOA-45 actionupon S. pneumoniae D39 cells
Pretreatment of S. pneumoniae D39 cells with 30 mM RutheniumRed, an inhibitor of Ca2þ transport, significantly prevents the bLAeOA-45einduced death of the cells, indicated by a reduction of log-death from 3.1 � 0.5 to 1.48 � 0.17 (protein concentration is 75 mg/ml), which suggests that Ca2þ transport is directly involved in thecytotoxic action of this complex. This is analogous to the 51% log-
n of fluorescence emission spectrum (lmax) and mid-transition temperature (T1/2) foroefficient (n) at 45 �C. Buffer conditions: pH 8.3, 20 mM H3BO3eKOH, 150 mM NaCl.
T1/2, �C K, M�1 n
37.4 � 0.8 (1.1 � 0.2) � 106 0.95 � 0.0238.1 � 1.0 (2.0 � 0.3) � 106 0.65 � 0.0830.0 � 0.7 (2.0 � 0.3) � 106 0.68 � 0.0429.5 � 1.4 (1.2 � 0.2) � 106 0.98 � 0.01
491492493494495496497498499500
for preparation of HAMLET-like protein complexes, Biochimie (2011),
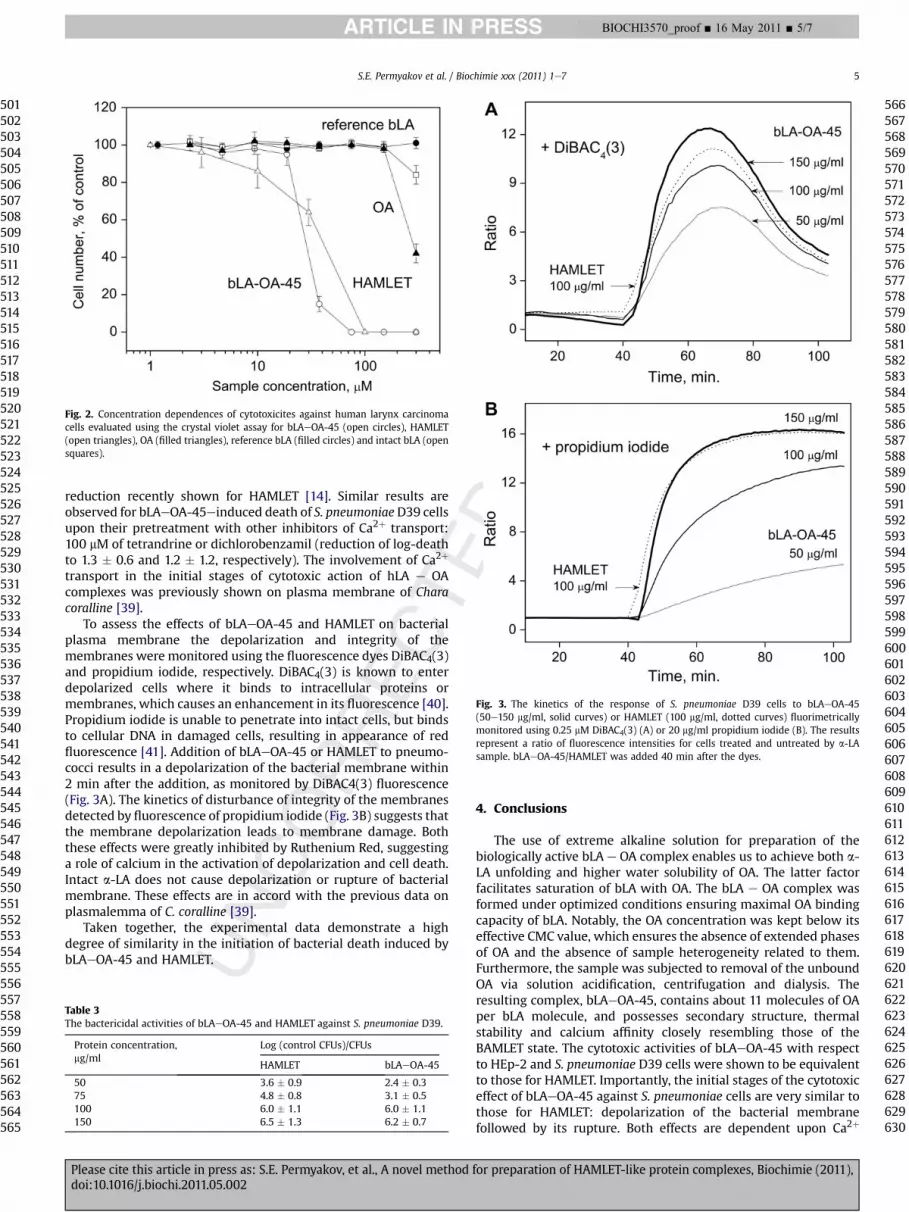
Fig. 3. The kinetics of the response of S. pneumoniae D39 cells to bLAeOA-45(50e150 mg/ml, solid curves) or HAMLET (100 mg/ml, dotted curves) fluorimetricallymonitored using 0.25 mM DiBAC4(3) (A) or 20 mg/ml propidium iodide (B). The resultsrepresent a ratio of fluorescence intensities for cells treated and untreated by a-LAsample. bLAeOA-45/HAMLET was added 40 min after the dyes.
Fig. 2. Concentration dependences of cytotoxicites against human larynx carcinomacells evaluated using the crystal violet assay for bLAeOA-45 (open circles), HAMLET(open triangles), OA (filled triangles), reference bLA (filled circles) and intact bLA (opensquares).
S.E. Permyakov et al. / Biochimie xxx (2011) 1e7 5
501502503504505506507508509510511512513514515516517518519520521522523524525526527528529530531532533534535536537538539540541542543544545546547548549550551552553554555556557558559560561562563564565
566567568569570571572573574575576577578579580581582583584585586587588589590591592593594595596597598599600601602603604605606607608609610611612613614615616617618619620
BIOCHI3570_proof ■ 16 May 2011 ■ 5/7
reduction recently shown for HAMLET [14]. Similar results areobserved for bLAeOA-45einduced death of S. pneumoniaeD39 cellsupon their pretreatment with other inhibitors of Ca2þ transport:100 mM of tetrandrine or dichlorobenzamil (reduction of log-deathto 1.3 � 0.6 and 1.2 � 1.2, respectively). The involvement of Ca2þ
transport in the initial stages of cytotoxic action of hLA e OAcomplexes was previously shown on plasma membrane of Characoralline [39].
To assess the effects of bLAeOA-45 and HAMLET on bacterialplasma membrane the depolarization and integrity of themembranes were monitored using the fluorescence dyes DiBAC4(3)and propidium iodide, respectively. DiBAC4(3) is known to enterdepolarized cells where it binds to intracellular proteins ormembranes, which causes an enhancement in its fluorescence [40].Propidium iodide is unable to penetrate into intact cells, but bindsto cellular DNA in damaged cells, resulting in appearance of redfluorescence [41]. Addition of bLAeOA-45 or HAMLET to pneumo-cocci results in a depolarization of the bacterial membrane within2 min after the addition, as monitored by DiBAC4(3) fluorescence(Fig. 3A). The kinetics of disturbance of integrity of the membranesdetected by fluorescence of propidium iodide (Fig. 3B) suggests thatthe membrane depolarization leads to membrane damage. Boththese effects were greatly inhibited by Ruthenium Red, suggestinga role of calcium in the activation of depolarization and cell death.Intact a-LA does not cause depolarization or rupture of bacterialmembrane. These effects are in accord with the previous data onplasmalemma of C. coralline [39].
Taken together, the experimental data demonstrate a highdegree of similarity in the initiation of bacterial death induced bybLAeOA-45 and HAMLET.
Table 3The bactericidal activities of bLAeOA-45 and HAMLET against S. pneumoniae D39.
Protein concentration,mg/ml
Log (control CFUs)/CFUs
HAMLET bLAeOA-45
50 3.6 � 0.9 2.4 � 0.375 4.8 � 0.8 3.1 � 0.5100 6.0 � 1.1 6.0 � 1.1150 6.5 � 1.3 6.2 � 0.7
621622623624625626627628629630
Please cite this article in press as: S.E. Permyakov, et al., A novel methoddoi:10.1016/j.biochi.2011.05.002
4. Conclusions
The use of extreme alkaline solution for preparation of thebiologically active bLA e OA complex enables us to achieve both a-LA unfolding and higher water solubility of OA. The latter factorfacilitates saturation of bLA with OA. The bLA e OA complex wasformed under optimized conditions ensuring maximal OA bindingcapacity of bLA. Notably, the OA concentration was kept below itseffective CMC value, which ensures the absence of extended phasesof OA and the absence of sample heterogeneity related to them.Furthermore, the sample was subjected to removal of the unboundOA via solution acidification, centrifugation and dialysis. Theresulting complex, bLAeOA-45, contains about 11 molecules of OAper bLA molecule, and possesses secondary structure, thermalstability and calcium affinity closely resembling those of theBAMLET state. The cytotoxic activities of bLAeOA-45 with respectto HEp-2 and S. pneumoniae D39 cells were shown to be equivalentto those for HAMLET. Importantly, the initial stages of the cytotoxiceffect of bLAeOA-45 against S. pneumoniae cells are very similar tothose for HAMLET: depolarization of the bacterial membranefollowed by its rupture. Both effects are dependent upon Ca2þ
for preparation of HAMLET-like protein complexes, Biochimie (2011),

S.E. Permyakov et al. / Biochimie xxx (2011) 1e76
631632633634635636637638639640641642643644645646647648649650651652653654655656657658659660661662663664665666667668669670671672673674675676677678679680681682683684685686687688689690691692693694695
696697698699700701702703704705706707708709710711712713714715716717718719720721722723724725726727728729730731732733734735736737738739740741742743744745746747748749750751752753754755756757758759760
BIOCHI3570_proof ■ 16 May 2011 ■ 6/7
transport. Overall, the bLAeOA-45 sample is in many ways equiv-alent to HAMLET/BAMLET. The developed low-cost express methodfor preparation of the HAMLET-like complex lacks the scalabilitylimitation of the previous method and is well suited for preparationunder regular laboratory conditions of quantities of the activecomplex necessary to conduct animal testing and pre-clinicalstudies.
The shown crucial role of hydrophobic effect in stabilization ofthe a-LA e OA complex opens up the possibility to prepare similarcomplexes of alkaline-denatured proteins with fatty acids andother low-molecular-weight amphiphilic substances, possessingcytotoxic properties with respect to malignant cells. The possibilityof dissolving cytotoxic hydrophobic substances in the non-polarinterior of natural proteins opens up a new avenue for celldelivery of these substances. Further efforts aimed at developmentof the complexes of this kind are required.
Conflict of interestsWe confirm that there is no conflict of interest in this
manuscript.
Author contributions
Sergei E. Permyakov was involved in experiment design, datatreatment and analysis, and manuscript preparation. Ekaterina L.Knyazeva was involved in data collection, data treatment andanalysis, manuscript preparation. Marina V. Leonteva, Roman S.Fadeev, Aleksei V. Chekanov and Andrei P. Zhadanwere involved indata collection, data treatment and analysis. Anders P. Håkanssonwas involved in experiment design, data collection, data treatmentand analysis, and manuscript preparation. Vladimir S. Akatov wasinvolved in experiment design. Eugene A. Permyakov was involvedin experiment design, and manuscript preparation.
Acknowledgements
This work was supported by grant to P.E.A. from the Program ofthe Russian Academy of Sciences «Molecular and Cellular Biology»,grant to K.E.L. from the Dynasty Foundation, and grants to A.P.H.from the Bill and Melinda Gates Foundation (Grant 53085), the JROishei Foundation, and The American Lung Association (Grant RG-123721-N).
References
[1] M. Svensson, A. Hakansson, A.K. Mossberg, S. Linse, C. Svanborg, Conversion ofalpha-lactalbumin to a protein inducing apoptosis, Proc. Natl. Acad. Sci. U S A97 (2000) 4221e4226.
[2] J. Pettersson, A.K. Mossberg, C. Svanborg, alpha-Lactalbumin species variation,HAMLET formation, and tumor cell death, Biochem. Biophys. Res. Commun.345 (2006) 260e270.
[3] M.H. Noteborn, Proteins selectively killing tumor cells, Eur. J. Pharmacol. 625(2009) 165e173.
[4] A.K. Mossberg, K. Hun Mok, L.A. Morozova-Roche, C. Svanborg, Structure andfunction of human alpha-lactalbumin made lethal to tumor cells (HAMLET)-type complexes, FEBS J. 277 (2010) 4614e4625.
[5] K.H. Mok, J. Pettersson, S. Orrenius, C. Svanborg, HAMLET, protein folding, andtumor cell death, Biochem. Biophys. Res. Commun. 354 (2007) 1e7.
[6] E.A. Permyakov, a-Lactalbumin. Nova Science Publishers, New York, 2005.[7] R.L. Hill, K. Brew, Lactose synthetase, Adv. Enzymol. Relat. Areas Mol. Biol. 43
(1975) 411e490.[8] C. Svanborg, H. Agerstam, A. Aronson, R. Bjerkvig, C. Duringer, W. Fischer,
L. Gustafsson, O. Hallgren, I. Leijonhuvud, S. Linse, A.K. Mossberg, H. Nilsson,J. Pettersson, M. Svensson, HAMLET kills tumor cells by an apoptosis-likemechanismecellular, molecular, and therapeutic aspects, Adv. Cancer Res.88 (2003) 1e29.
[9] L. Gustafsson, I. Leijonhufvud, A. Aronsson, A.K. Mossberg, C. Svanborg,Treatment of skin papillomas with topical alpha-lactalbumin-oleic acid,N. Engl. J. Med. 350 (2004) 2663e2672.
[10] W. Fischer, L. Gustafsson, A.K. Mossberg, J. Gronli, S. Mork, R. Bjerkvig,C. Svanborg, Human alpha-lactalbumin made lethal to tumor cells (HAMLET)
Please cite this article in press as: S.E. Permyakov, et al., A novel methoddoi:10.1016/j.biochi.2011.05.002
kills human glioblastoma cells in brain xenografts by an apoptosis-likemechanism and prolongs survival, Cancer Res. 64 (2004) 2105e2112.
[11] A.K. Mossberg, B. Wullt, L. Gustafsson, W. Mansson, E. Ljunggren,C. Svanborg, Bladder cancers respond to intravesical instillation of HAMLET(human alpha-lactalbumin made lethal to tumor cells), Int. J. Cancer 121(2007) 1352e1359.
[12] A.K. Mossberg, Y. Hou, M. Svensson, B. Holmqvist, C. Svanborg, HAMLETtreatment delays bladder cancer development, J. Urol. 183 (2010) 1590e1597.
[13] A. Hakansson, M. Svensson, A.K. Mossberg, H. Sabharwal, S. Linse, I. Lazou,B. Lonnerdal, C. Svanborg, A folding variant of alpha-lactalbumin withbactericidal activity against Streptococcus pneumoniae, Mol. Microbiol. 35(2000) 589e600.
[14] A. Hakansson, H. Roche-Hakansson, A.K. Mossberg, C. Svanborg, Apoptosis-like death in bacteria induced by HAMLET, a human milk lipid-proteincomplex, PLoS One (2011).
[15] K.M. Cawthern, M. Narayan, D. Chaudhuri, E.A. Permyakov, L.J. Berliner,Interactions of alpha-lactalbumin with fatty acids and spin label analogs,J. Biol. Chem. 272 (1997) 30812e30816.
[16] M. Svensson, A.K. Mossberg, J. Pettersson, S. Linse, C. Svanborg, Lipids ascofactors in protein folding: stereo-specific lipid-protein interactions arerequired to form HAMLET (human alpha-lactalbumin made lethal to tumorcells), Protein Sci. 12 (2003) 2805e2814.
[17] K. Wilhelm, A. Darinskas, W. Noppe, E. Duchardt, K.H. Mok, V. Vukojevic,J. Schleucher, L.A. Morozova-Roche, Protein oligomerization induced by oleicacid at the solid-liquid interfaceeequine lysozyme cytotoxic complexes, FEBSJ. 276 (2009) 3975e3989.
[18] B. Spolaore, O. Pinato, M. Canton, M. Zambonin, P. Polverino de Laureto,A. Fontana, alpha-Lactalbumin forms with oleic acid a high molecular weightcomplex displaying cytotoxic activity, Biochemistry 49 (2010) 8658e8667.
[19] Y. Zhu, S. Schwarz, B. Ahlemeyer, S. Grzeschik, S. Klumpp, J. Krieglstein, Oleicacid causes apoptosis and dephosphorylates Bad, Neurochem. Int. 46 (2005)127e135.
[20] M.F. Cury-Boaventura, C. Pompeia, R. Curi, Comparative toxicity of oleic acidand linoleic acid on Jurkat cells, Clin. Nutr. 23 (2004) 721e732.
[21] E.L. Knyazeva, V.M. Grishchenko, R.S. Fadeev, V.S. Akatov, S.E. Permyakov,E.A. Permyakov, Who is Mr. HAMLET? Interaction of human alpha-lactalbumin with monomeric oleic acid, Biochemistry 47 (2008)13127e13137.
[22] D.P. Cistola, J.A. Hamilton, D. Jackson, D.M. Small, Ionization and phasebehavior of fatty acids in water: application of the Gibbs phase rule,Biochemistry 27 (1988) 1881e1888.
[23] H. Fukuda, A. Goto, H. Yoshioka, R. Goto, K. Morigaki, P. Walde, Electron spinresonance study of the pH-induced transformation of micelles to vesicles inan aqueous oleic acid/oleate system, Langmuir 17 (2001) 4223e4231.
[24] K. Edwards, M. Silvander, G. Karlsson, Aggregate structure in dilute aqueousdispersions of oleic acid/sodium oleate and oleic acid/sodium oleate/eggphosphatidylcholine, Langmuir 11 (1995) 2429e2434.
[25] C. Duringer, A. Hamiche, L. Gustafsson, H. Kimura, C. Svanborg, HAMLETinteracts with histones and chromatin in tumor cell nuclei, J. Biol. Chem. 278(2003) 42131e42135.
[26] D.B. Veprintsev, S.E. Permyakov, E.A. Permyakov, V.V. Rogov, K.M. Cawthern,L.J. Berliner, Cooperative thermal transitions of bovine and human apo-alpha-lactalbumins: evidence for a new intermediate state, FEBS Lett. 412 (1997)625e628.
[27] P. Polverino d.L., E. Frare, R. Gottardo, A. Fontana, Molten globule of bovinealpha-lactalbumin at neutral pH induced by heat, trifluoroethanol, and oleicacid: a comparative analysis by circular dichroism spectroscopy and limitedproteolysis, Proteins 49 (2002) 385e397.
[28] T. Kamijima, A. Ohmura, T. Sato, K. Akimoto, M. Itabashi, M. Mizuguchi,M. Kamiya, T. Kikukawa, T. Aizawa, M. Takahashi, K. Kawano, M. Demura,Heat-treatment method for producing fatty acid-bound alpha-lactalbuminthat induces tumor cell death, Biochem. Biophys. Res. Commun. 376 (2008)211e214.
[29] R.I. Kaplanas, A.I. Antanavichyus, Isolation and purification of a-lactalbumin,a component of lactose synthetase, Biochemistry (Mosc.) 40 (1975) 493e495.
[30] L. Lindahl, H.J. Vogel, Metal-ion-dependent hydrophobic-interaction chro-matography of alpha-lactalbumins, Anal. Biochem. 140 (1984) 394e402.
[31] M.J. Kronman, R.E. Andreotti, Inter and intramolecular interactions of -lact-albumin. I. The apparent heterogeneity at acid pH, Biochemistry 3 (1964)1145e1151.
[32] M. Nozaka, K. Kuwajima, K. Nitta, S. Sugai, Detection and characterization ofthe intermediate on the folding pathway of human a-lactalbumin, Biochem-istry 17 (1978) 3753e3758.
[33] N. Sreerama, S.Y. Venyaminov, R.W. Woody, Estimation of protein secondarystructure from circular dichroism spectra: inclusion of denatured proteinswith native proteins in the analysis, Anal. Biochem. 287 (2000) 243e251.
[34] O.T. Avery, C.M. Macleod, M. McCarty, Studies on the chemical nature of thesubstance inducing transformation of pneumococcal types: induction oftransformation by a desoxyribonucleic acid fraction isolated from pneumo-coccus type III, J. Exp. Med. 79 (1944) 137e158.
[35] J.R. Kanicky, D.O. Shah, Effect of degree, type, and position of unsaturation onthe pKa of long-chain, J. Colloid Interface Sci. 256 (2002) 201e207.
[36] E.A. Permyakov, L.A. Morozova, E.A. Burstein, Cation binding effects on the pH,thermal and urea denaturation transitions in alpha-lactalbumin, Biophys.Chem. 21 (1985) 21e31.
for preparation of HAMLET-like protein complexes, Biochimie (2011),

S.E. Permyakov et al. / Biochimie xxx (2011) 1e7 7
761762763764765766767
768769770771772773
BIOCHI3570_proof ■ 16 May 2011 ■ 7/7
[37] E.A. Permyakov, K. Murakami, L.J. Berliner, On experimental artifacts in theuse of metal ion chelators for the determination of the cation bindingconstants of alpha-lactalbumin. A reply, J. Biol. Chem. 262 (1987) 3196e3198.
[38] J. Fast, A.K. Mossberg, C. Svanborg, S. Linse, Stability of HAMLETeA kineticallytrapped {alpha}-lactalbumin oleic acid complex, Protein Sci. 14 (2005)329e340.
[39] O.M. Zherelova, A.A. Kataev, V.M. Grishchenko, E.L. Knyazeva, S.E. Permyakov,E.A. Permyakov, Interaction of antitumor alpha-lactalbumin-oleic acid
Please cite this article in press as: S.E. Permyakov, et al., A novel methoddoi:10.1016/j.biochi.2011.05.002
complexes with artificial and natural membranes, J. Bioenerg. Biomembr. 41(2009) 229e237.
[40] D.E. Epps, M.L. Wolfe, V. Groppi, Characterization of the steady-state anddynamic fluorescence properties of the potential-sensitive dye bis-(1,3-dibutylbarbituric acid)trimethine oxonol (Dibac4(3)) in model systems andcells, Chem. Phys. Lipids 69 (1994) 137e150.
[41] D.J. Arndt-Jovin, T.M. Jovin, Fluorescence labeling and microscopy of DNA,Methods Cell Biol. 30 (1989) 417e448.
774
for preparation of HAMLET-like protein complexes, Biochimie (2011),




















