
Miocene mollusks from Bowden, Jamaica. pt. 2. Gastropods ...
Upload
univ-poitiersCategory
view
1download
0
Zoological Journal of the Linnean Society
, 2006,
146
, 227–237. With 10 figures
© 2006 The Linnean Society of London,
Zoological Journal of the Linnean Society,
2006,
146
, 227–237
227
Blackwell Science, Ltd
Oxford, UK
ZOJZoological Journal of the Linnean Society
0024-4082The Lin-
nean Society of London, 2006? 2006
146
?
227237Original Article
A NEW POLYPTERID FROM THE LATE MIOCENE OF CHADO. OTERO ET AL.
*Corresponding author. E-mail: [email protected]
A new polypterid fish:
Polypterus faraou
sp. nov. (Cladistia, Polypteridae) from the Late Miocene, Toros-Menalla, Chad
OLGA OTERO
1
*, ANDOSSA LIKIUS
2
, PATRICK VIGNAUD
1
and MICHEL BRUNET
1
1
Laboratoire de Géobiologie, Biochronologie et Paléontologie humaine, CNRS UMR 6046, Faculté des Sciences fondamentales et appliquées, Université Poitiers, 40 av. du Recteur Pineau, F-86 022 Poitiers cedex, France
2
Département de Paléontologie, Université de N’Djaména, N’Djaména, Tchad
Received June 2004; accepted for publication July 2005
Polypterus faraou
sp. nov.
(Cladistia, Polypteridae) from the Late Miocene of Toros-Menalla (western Djourab,Chad) is described on the basis of a subcomplete articulated skeleton preserved in three dimensions. This is the firsttime such a complete fossil polypterid skeleton has been described. It is the only verifiable fossil record for the genus
Polypterus
.
P. faraou
closely resembles
P. bichir
and
P. endlicheri
, extant fish found in the Chad–Chari system.Intrarelationships among the polypterids are not yet resolved: however,
P. faraou
shows a primitive shape of thebody and a primitive shape of the opening of the lateral line on the scales, similar to that of three living species(
P. bichir
,
P. endlicheri
and
P. ansorgii
). © 2006 The Linnean Society of London,
Zoological Journal of the LinneanSociety
, 2006,
146
, 227–237.
ADDITIONAL KEYWORDS: anatomy – Anthracotheriid Unit – Polypteriformes – western Djourab.
INTRODUCTION
Palaeontological expeditions have been conducted inChad since the early 1930s. In 1994, the MPFT(Mission Paléoanthropologique Franco-Tchadienne)initiated a series of field missions to the Mio-Pliocenesites of the eastern Djurab Desert in northern Chad(Brunet
et al
., 1995, 1998; Brunet & MPFT, 2000) and,since 1997, to the Miocene sites of Toros-Menalla,western Djurab. The Anthracoteriid Unit in Toros-Menalla has yielded a rich vertebrate fauna (Vignaud
et al
., 2002), the oldest known species of hominid,
Sahelanthropus tchadensis
(Brunet
et al
., 2002), aswell as the first nearly complete and articulated skel-eton of a fossil polypterid. This polypterid belongs to anew species that is described here. The estimated ageof the Anthracotheriid Unit is
c
. 7 Myr (Vignaud
et al
.,2002).
Polypterids (Cladistia) are basal actinopterygianfishes (Rosen
et al
., 1981). They are represented by
two extant genera, i.e.
Polypterus
, with ten species and15 subspecies (Poll, 1941a, b, 1942; Hanssens, Teugels& Thys van der Audenaerde, 1995; Britz, 2004; thereare 11 species if
P. polli
is considered as a valid spe-cies), and the monotypic genus
Erpetoichthys
. Ninefossil genera have been described. Two have beendescribed on the basis of isolated skull bones andscales (Meunier & Gayet, 1996; Gayet & Meunier,1991), and six others have been described on the basisof the articular head of pinnules (Gayet & Meunier,1996; Werner & Gayet, 1997). Finally,
Serenoichthys
has been described on the basis of scale and finletcounts and distribution (Dutheil, 1999).
Serenoichthys
is the only other fossil articulated skeleton known sofar, but the head is missing. These fossil genera areCretaceous in age and African, except for
Dagetella
and
Latinopollia
, which are of Late Cretaceous andEarly Palaeocene age, respectively, and South Ameri-can. Post-Palaeocene polypterid history is exclusivelyAfrican. Isolated scales and skull bones attributed to
Polypterus
sp. testify to the presence of the genus inAfrican freshwaters since the Eocene. Moreover, two

228
O. OTERO
ET AL.
© 2006 The Linnean Society of London,
Zoological Journal of the Linnean Society,
2006,
146
, 227–237
types of African Cretaceous pinnules were tentativelyattributed to two fossil
Polypterus
species,
P. dageti
(Gayet & Meunier, 1996) and
P. sudanensis
(Werner &Gayet, 1997). The intrarelationships among poly-pterids have yet to be clarified; indeed, the monophylyand the intrarelationships among extant species havenot been established.
This paper provides the first description of a nearcomplete fossil polypterid skeleton including cranium,and it is the only clearly recognizable fossil record forthe genus
Polypterus
. The new species from the LateMiocene of Toros-Menalla (western Djurab, Chad) isassigned to the genus
Polypterus
based on its morpho-logical characteristics, which fall within the anatomi-cal range of Recent species of the genus, from which itdiffers by a unique combination of characters. We alsodiscuss the primitive or derived state of certain osteo-logical features observed in this species.
SYSTEMATIC PALAEONTOLOGY
A
CTINOPTERYGII
C
OPE
, 1887(
SENSU
R
OSEN
ET
AL
., 1981)
CLADISTIA
COPE
, 1871P
OLYPTERIDAE
G
ÜNTHER
, 1870
P
OLYPTERUS
S
T
H
ILAIRE
, 1802
Type species. Polypterus bichir
St Hilaire, 1802.
P
OLYPTERUS
FARAOU
SP
.
NOV
.
Derivation of name
. In Chadian Arabian,
faraou
means flattened. The name is given in reference to thedorso-ventrally depressed head of the fish.
Type specimen
. TM090-001-039, housed in the CentreNational d’Appui à la Recherche (CNAR, N’Djamena,Chad).
Diagnosis
. A
Polypterus
species characterized by:depressed head (as in
P. weeksii
and
P. endlicheri
only), head length approximately 20% of body length(less than 20% in other
Polypterus
species except
P. ansorgii
,
P. bichir
and
P. endlicheri
) and head widthat the level of the preoperculum reaches 70% of itslength (more than in any other
Polypterus
species);opening of the lateral line on the nasal 3 close to itsposterior border (as in
P. bichir
and
P. endlicheri
,whereas it is central in other species); large androunded snout, shorter than the interorbital distance(as in most
Polypterus
species except
P. bichir
and
P. palmas
the snout of which is longer than theinterorbital distance); superolateral position of theorbit (as in
P. ansorgii
,
P. weeksii
,
P. endlicheri
and
P. bichir
, whereas the orbit is lateral in
P. delhezi
,
P. ornatipinnis
,
P. senegalus
,
P. retropinnis
and
P. palmas
); orbit length around the half the interor-
bital space (as in
P. bichir katangae
and possibly in
P. teugelsi
, whereas it is more than 60% in
P. ansorgii
,and less than 45% in the other species) and orbitsmaller than suboperculum width (as in
P. bichir
and
P. endlicheri
only); prognathus lower jaw (as in
P. bichir
and
P. endlicheri
only); at least 5–7 prespi-racular bones (the number of prespiracular bones ispossibly equal to or above seven only in
P. weeksii
, in
P. bichir bichir
and in
P. endlicheri congicus
); deepbody with approximately 23 transversal scales, 58–60scale rows, and 14 predorsal scales (such body shapeexists elsewhere only in
P. ansorgii
,
P. bichir
and
P. endlicheri
); 15 finlets with successive pinnules ofthe finlets overlapping the base of the followingone (such dorsal fin mophology elsewhere only in
P. ansorgii
,
P. bichir
and
P. endlicheri congicus
);grooved lateral line scales with notched posterior mar-gins (as in
P. ansorgii
,
P. bichir
and
P. endlicheri
only).The holotype is 590 mm in standard length. Such
dimensions are also reached in
P. bichir
and
P. endlicheri
only.
Occurrence. Anthracotheriid Unit of Toros-Menalla,Late Miocene of Chad.
MATERIAL AND METHODS
Specimen TM090-001-039 was found lying on its back.It is preserved in three dimensions. The dorsal andlateral body surfaces have globally kept their livingshape, whereas the ventral surface has collapsed(Fig. 1A–C). Most of the scales of the body are presentand articulated with each other. All the bones of theright half of the skull are in natural position, whereassome bones of the left side are slightly displaced. Thisthree-dimension preservation allows us to observe thenatural position of the bones and the shape of the cra-nium; it also allows us to measure the dimensions ofthe specimen (Fig. 2) and to compare some dimensionsand meristic data between the specimen and Recentspecies. Jollie’s (1984) nomenclature of the bones isfollowed.
ANATOMICAL DESCRIPTION
HEAD AND BODY SHAPE (FIGS 1–3)
The standard length of the specimen is 59 cm (Fig. 2,a). We observe a natural dorso-ventral depression ofthe head. The head length (from the front of the mouthto the posterior level of the opercular bone) is equiva-lent to about 20% of the standard length (Fig. 2, m).The width of the head, measured at the level of thepreoperculum, is around 70% of its length (Fig. 2, n).The distance between the lateral border of the spira-cular series at the level of the preoperculum is 45% ofthe total length of the head (Fig. 2, o). The lower jaw is
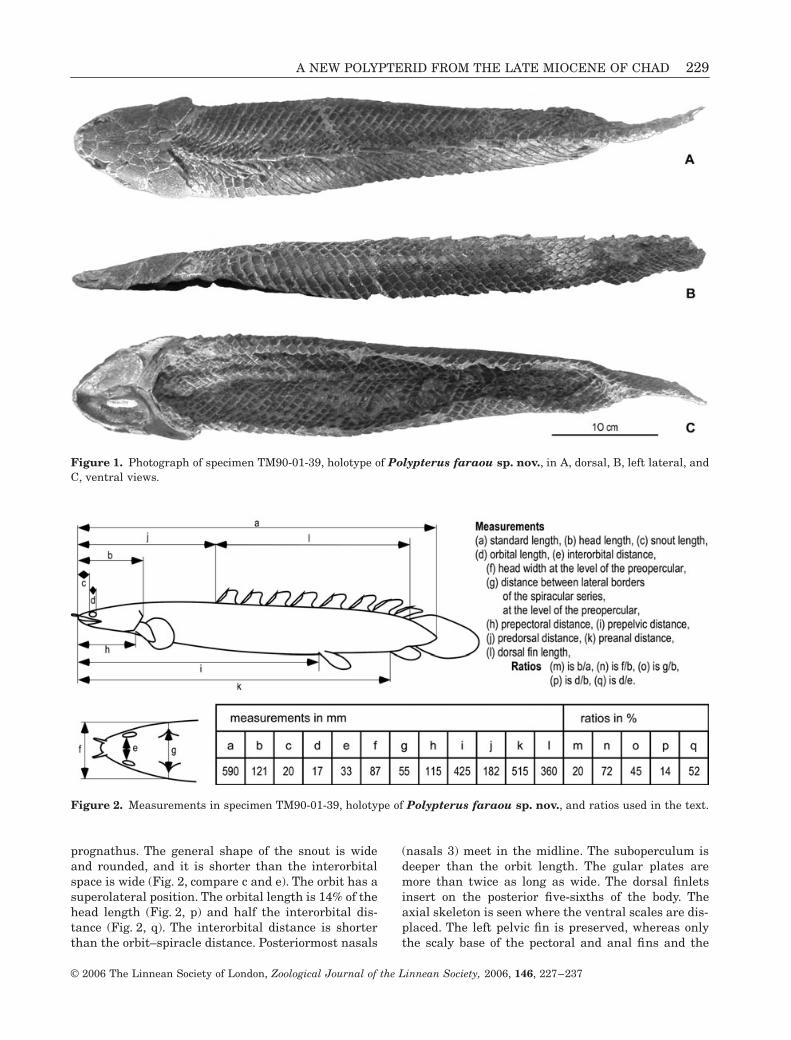
A NEW POLYPTERID FROM THE LATE MIOCENE OF CHAD 229
© 2006 The Linnean Society of London, Zoological Journal of the Linnean Society, 2006, 146, 227–237
prognathus. The general shape of the snout is wideand rounded, and it is shorter than the interorbitalspace is wide (Fig. 2, compare c and e). The orbit has asuperolateral position. The orbital length is 14% of thehead length (Fig. 2, p) and half the interorbital dis-tance (Fig. 2, q). The interorbital distance is shorterthan the orbit–spiracle distance. Posteriormost nasals
(nasals 3) meet in the midline. The suboperculum isdeeper than the orbit length. The gular plates aremore than twice as long as wide. The dorsal finletsinsert on the posterior five-sixths of the body. Theaxial skeleton is seen where the ventral scales are dis-placed. The left pelvic fin is preserved, whereas onlythe scaly base of the pectoral and anal fins and the
Figure 1. Photograph of specimen TM90-01-39, holotype of Polypterus faraou sp. nov., in A, dorsal, B, left lateral, andC, ventral views.
Figure 2. Measurements in specimen TM90-01-39, holotype of Polypterus faraou sp. nov., and ratios used in the text.

230 O. OTERO ET AL.
© 2006 The Linnean Society of London, Zoological Journal of the Linnean Society, 2006, 146, 227–237
proximal part of some caudal rays are preserved. Theposition of the fins is given (Fig. 2, h–l).
SNOUT, ORBIT AND UPPER JAW (FIG. 4)
The INTERNASAL (median rostral) is the unpaired dia-mond-shaped bone at the front of the head. It carriesthe ethmoid commissure. It lies dorsal to the rostro-premaxillae, and inserts laterally between the nasals2, and posteriorly between the nasals 3.
The paired nasal bones and the nostrils are in placeon the left side but slightly displaced on the right.NASAL 1 is a curved tubular bone, where the infraor-bital canal runs from the rostro-premaxilla to a lateralline pore posterior to the nostril. On the right side, asmall displaced piece of bone lying in the nostril open-ing could be a fragment of the missing right nasal 1.The rounded NASAL 2 lies lateral to the internasal andcarries the canal from the rostro-premaxillae to thenasals 3. NASAL 3 is roughly square and joins its anti-
Figure 3. Photograph and drawing of the skull of TM90-01-39, holotype of Polypterus faraou sp. nov., in dorsal view.
Figure 4. Photograph and drawing of the snout of TM90-01-39, holotype of Polypterus faraou sp. nov., in anterolateraldorsal view.

A NEW POLYPTERID FROM THE LATE MIOCENE OF CHAD 231
© 2006 The Linnean Society of London, Zoological Journal of the Linnean Society, 2006, 146, 227–237
mere at the midline. There are no internasal super-numerary plates (they develop frequently in someliving species, e.g. P. weeksii; Poll, 1942). The supraor-bital sensory canal of either side passes posteriorlythrough nasals 1, 2 and 3 to the frontals. The openingfrom the canal within nasal 3 lies within the posteriortwo-thirds of the bone.
On each side of the skull, the ROSTRO-PREMAXILLA
carries the infraorbital canal from the midline (inter-nasal) to the maxilla, via the lacrymal. It extendsbetween the nostril and the orbit, along the ventralmargin of nasal 3, and sutures posteriorly with thefrontal. The ascending process of the rostro-premax-illa is postero-ventrally covered by the LACRYMAL. Thelacrymal is a tubular bone that carries the infraorbitalcanal from the rostro-premaxilla to the maxilla. Thelacrymal shows a preorbital process dorsally. The MAX-
ILLA reaches from the level of the nostril to the pres-piracular series. The POSTORBITAL borders the back ofthe orbit from the maxilla to the frontal, along theleading edge of the prespiracular series. It is slightlycurved backwards in its caudal part, delimiting a freenarrow space with the lateral border of the frontal atthe postero-dorsal edge of the orbit. The infraorbitalcanal runs from the lacrymal to the postorbitalthrough the maxilla and to the frontal through thepostorbital. On this latter bone, there is a tiny dorsalopening whereas the ventral one, just above the max-illa, is wide.
Nasal 3 is the only bone of the snout with a dermalornamentation of distinctly separated tubercles cov-ered by ganoine. The other snout bones are smooth,ganoine-free and with numerous small pores particu-larly in the rostral part. Caniniform teeth are devel-oped on the ventral face of the rostro-premaxilla infront of the maxilla, and on the maxilla from the ante-rior tip to the midlevel of the orbits.
DORSAL SIDE OF THE HEAD (FIGS 3 AND 4)
The FRONTALS are the largest bones of the head. Thefrontal branch of the lateral canal has two openingsabove the orbit (only one is seen on the right side of thefossil). It connects with the infra-orbital branch infront of the spiracular bones, at the level of the der-mosphenotic. According to Jollie (1984), the canal isshared by both the frontal and the dermosphenotic atthat point, and there is an opening on the dermosphe-notic. Only the opening of the dermosphenotic can beseen, but not the limits of the bone itself. It seems to beabsent or totally incorporated in the frontal. At thepostero-lateral angle of each frontal, the canal passesto the INTERTEMPORO-SUPRATEMPORAL, where it opensin a pore midway along the length of the bone. Theintertemporo-supratemporal is roughly rectangularin shape. There are six EXTRASCAPULAR PLATES. Four
are aligned along the posterior border of the inter-temporo-supratemporals, and two are lateral. Thelast-mentionned are diamond-shaped, and they liebetween the post-spiracular bones, the more medialextrascapular plates and the posttemporal. There areopenings of the lateral line between the extrascapularplates and between the lateral extrascapular plateand the posttemporal. The POSTTEMPORAL is roughlyrectangular in shape and shows a posterior openingfor the lateral-line canal. The posterior process is notseen. It is probably broken on the left side where thescales are slightly displaced (it is present in livingPolypterus but hidden by the scales). The posteriormidline scale is missing.
The PRESPIRACULAR, SPIRACULAR and POSTSPIRACU-
LAR bones border the lateral edge of the former dorsalbones of the skull, from the postorbital to the posttem-poral, above the opercular series. The spiracularbones lie along the postero-lateral border of thefrontal and the lateral border of the intertemporo-supratemporal. The spiracular formulae are, respec-tively, 5–2−3 on the left side, and 7–2−4 on the right.The SPIRACLE opens along the second spiracular bone,above the DERMOHYAL, the dorsal face of which can beseen.
All of the skull roofing bones have a dermal orna-mentation of distinctly separated tubercles covered byganoine on their dorsal face.
ORBITOSPHENOID, ECTOPTERYGOID AND LATERAL ETHMOID (FIG. 4)
Parts of these three bones and their spatial relation-ships are seen in the left orbit of the specimen.
OPERCULAR SERIES AND CHEEK (FIG. 5)
The opercular series is well developed, with a largeSUBOPERCULUM the dorsal extremity of which reachesthe preorbital canal median opening, and an OPERCU-
LUM as large as the PREOPERCULUM. The posteroven-tral process of the preoperculum is present. TwoSUPERNUMERARY CHEEK PLATES lie anterior to theposteroventral process, along the preoperculum ven-tral border. Together with the process, they totallycover the cheek. Anteriorly, the preoperculum issutured with the maxilla. The preopercular sensorycanal has four openings. The first is dorsal and islocated between the spiracular and postspiracularseries. The second opening lies close to the posteriorborder of the bone at one-quarter of its height. Thethird is in a more ventral position, just above the ven-tral process of the preoperculum. The fourth openinglies at the lower extremity of the ventral process.
All of the cheek bones have their surface covered bythe ganoine tubercules.

232 O. OTERO ET AL.
© 2006 The Linnean Society of London, Zoological Journal of the Linnean Society, 2006, 146, 227–237
LOWER JAW AND GULAR AREA (FIG. 6)
The lower jaw is prognathus. The posterior level of thesuture between the DENTOSPLENIAL and the ANGULAR
reaches the anterior level of the supernumerary cheekplates. The angular–prearticular–articular sutures
are not seen, and nor is the quadrate–articular arti-culation; they are hidden by the gular and the cheeksupernumerary plates. The coronoids and mento-meckelian bones are not observable, owing to theirinternal position and small dimension. Caniniformteeth are developed on the edge of the dentosplenial.
Figure 5. Photograph and drawing of the opercular series and cheek of TM90-01-39, holotype of Polypterus faraou sp.nov., in laterodorsal view.
Figure 6. Photograph and drawing of the skull and pectoral girdle of TM90-01-39, holotype of Polypterus faraou sp.nov., in ventral view.

A NEW POLYPTERID FROM THE LATE MIOCENE OF CHAD 233
© 2006 The Linnean Society of London, Zoological Journal of the Linnean Society, 2006, 146, 227–237
Ventrally, the wide and long GULAR PLATES aremostly covered by a thin ganoin ornamentation exceptin their anteriormost parts. They cover nearly thewhole ventral surface between the lower jaws.Because they are slightly displaced, a medial bone isobservable which could be the urohyal.
PECTORAL GIRDLE AND BASE OF THE FINS (FIGS 6, 7)
The pectoral girdle is composed of stout bones. Dor-sally and posteriorly, the SUPRACLEITHRUM bordersthe operculum and it covers the first scale row. Later-ally, the POSTCLEITHRUM is probably represented by abony plate broken in three fragments. It is delicatelyornamented with smooth ganoine tubercles, and it islying above an aggregation of small scales which is thelateral edge of the base of the pectoral fin. Ventrally,the CLEITHRA join in the midline. They are coveredanteriorly by large clavicles that articulate with oneanother by two small processes and a notch.
SCALES AND FINLETS (FIG. 8), AXIAL SKELETON
There are 15 pinnules bearing the finlets. The firstinserts behind the 14th row of scales. Each pinnule isseparated from the following one by three scale rows.They are long enough to cover the base of the followingone when folded in life. The pair of scales with whicheach pinnule articulates are L-shaped. The distalextremity of each pinnule is forked and the articularbase is symmetrical. On one pinnule, at least two fin
rays can be seen. The caudal fin starts dorsally behindthe last finlet at the 48th scale row, and ventrally atthe 53rd scale row. The first six dorsal and first sevenventral caudal rays are preserved in their proximalpart. All scales have exposed surface covered byganoin. There are 23 scales on each transversal rowacross the body on either side: most of the scales of thelateral line bear a groove that opens posteriorly in anotch, and some present a simple pore. There are eightscales above and 14 below the lateral line in a trans-versal row. There are 58–60 longitudinal scales, andthus probably about 60 vertebrae (Daget & Desoutter,1983). This agrees with the count of centra madewhere the covering scales are displaced. The centraare typical, rounded with one median and two lateraldeep gutters ventral to the lateral apophysis.
PELVIC FIN (FIG. 9) AND ANAL FIN
The scaly base of the left pelvic fin and the proximalpart of the rays are preserved. The fin inserts behindthe 31st transversal row of scales. There are at least11 pelvic rays. The position of the anal fin is assumedbased on the preceding small square scales known inliving polypterids.
SYSTEMATIC AFFINITIES AND DISCUSSION
Polypterus faraou has a character unique to the cla-distians: the dorsal fin is composed of several finlets,each consisting of a pinnule articulated with one or
Figure 7. Photograph and drawing of the operculum, cleithrum and postcleithrum of TM90-01-39, holotype of Polypterusfaraou sp. nov., in lateral view.
Figure 8. Photograph and drawing of pinnules 1–4 in place with the surrounding scales of TM90-01-39, holotype ofPolypterus faraou sp. nov., in dorsal view.

234 O. OTERO ET AL.
© 2006 The Linnean Society of London, Zoological Journal of the Linnean Society, 2006, 146, 227–237
more soft rays. It also shows polypteriform character-istics such as absence of branchiostegals and a maxillathat carries the infraorbital sensory canal. It haspolypterid features: infraorbital canals fused to themaxilla and fusion of lateral rostral bones to the pre-maxilla (Pehrson, 1947; Jollie, 1984), the presence ofspiracular bones, the preoperculum covering the cheekand three separate nasal bones. The lateral-line sys-tem on the head of Polypterus faraou conforms to thecommon pattern described by Jollie (1984) for livingpolypterids.
Polypterus faraou has symmetrical pinnules as inRecent Polypterus species and in Erpetoichthys,whereas a different structure of the articular head ofthe pinnules characterized the fossil genera describedby Gayet & Meunier (1996) and Werner & Gayet(1997) (see Introduction). It also differs from Ser-enoichthys, in which the body is much shorter thanin Recent genera (Dutheil, 1999). However, it doesnot show the elongation of the body as observed inErpetoichthys, nor the reductive characters of thatgenus, e.g. no suboperculars and no supernumerarycheek plates (Smith, 1865; Poll, 1941b; O. Otero, pers.observ.). Moreover, at least one of the pinnule bears atleast two rays as in Recent Polypterus species, by con-trast with Erpetoichthys, the pinnules of which bearone single fin ray (Daget, 1950; O. Otero, pers. observ.).From this evidence, we assign the new species to thegenus Polypterus.
Poll (1941a, b, 1942, 1965) described the Recentpolypterid species and defined six anatomically basedclusters of species and subspecies organized on ana-tomical trends deriving from a central type (Fig. 10A).The possible phylogenetic significance of these clustershas not been established; they only have a descriptivevalue. Polypterus faraou belongs to the P. bichir –P. endlicheri cluster with: (1) head length about 20% ofbody length; (2) head depressed; (3) prognathus jaw; (4)opening of the lateral line on the nasal 3 close to its pos-terior border; (5) orbit length smaller than the subo-
perculum width; (6) successive pinnules overlappingthe base of the following ones when folded in life; (7)grooved lateral line scales with notched posterior mar-gin. It also has other characteristics observed inP. bichir, P. endlicheri and also in some other species:(1) superolateral position of the orbit (P. ansorgii,P. weeksii); (2) 5–7 prespiracular bones (P. ornatipin-nis, P. weeksii); (3) 15 finlets (P. ansorgii); (4) deep body,with 46 transversal scales and 58 scale rows(P. ornatipinnis, P. ansorgii, P. weeksii); (5) only 14 pre-dorsal scales (P. delhezi); (6) known maximum stan-dard length of 590 mm (P. ornatipinnis). Moreover,Polypterus faraou differs from P. bichir and resemblesP. endlicheri based on its depressed head, and largeand rounded snout which is shorter than the interor-bital distance; it differs from P. endlicheri and resem-bles P. bichir katangae based on its orbital length,which is 50% of the interorbital space in adults (lessthan 40% in adults of P. endlicheri subspecies andP. bichir subspecies bichir and lapradei). Finally,Polypterus faraou is also the only Polypterus specieswith such a wide head: the head width at the level ofthe preoperculum is about 70% of the length; this canalso be expressed by the distance between the lateralborders of the spiracular series (at the level of the pre-operculum), which is 45% of the head length whereasit is around one-third in large specimens of P. bichir.
In conclusion, the new taxon is assigned to thegenus Polypterus based on its morphological charac-teristics, which fall into the range observed in the spe-cies of the genus, particularly the cluster P. endlicheri– P. bichir, from which it differs by a unique combina-tion of characters. The suggestion that the clusterformed by P. faraou and the Recent P. endlicheri andP. bichir (Fig. 10A) is a natural group will need to betested as intrarelationships among Polypterus speciesare still unknown. Nevertheless, this hypothesis is inagreement with the distribution of extant species:among the three polypterid fishes living in the Chad–Chari system, P. endlicheri and P. bichir are present
Figure 9. Photograph and drawing of the left pelvic fin of TM90-01-39, holotype of Polypterus faraou sp. nov., in lateralview.

A NEW POLYPTERID FROM THE LATE MIOCENE OF CHAD 235
© 2006 The Linnean Society of London, Zoological Journal of the Linnean Society, 2006, 146, 227–237
(Fig. 10B), as represented by P. endlicheri endlicheriand P. bichir bichir; the third one is P. senegalus(Gosse, 1990). However, the phylogenetic relation-ships of Recent polypterids must be established beforewe can draw any biogeographical inferences. A cladis-tic analysis on the basis of anatomical characters willrequire further anatomical review of the species,which seem morphologically so close to each other anddiffer mainly in their combination of characteristics.Moreover, difficulty exists in choosing an outgroupbecause (1) the closely related Erpetoichthys must beincluded as an ingroup in such an analysis (this mono-typic genus is characterized by reductive charactersand the elongation of the body; it thus could be moreclosely related to one of the species of Polypterus), and(2) the related fossils are only known by their pinnulesand/or scales, except Serenoichthys (see Introduction).However, two anatomical characters have been dis-cussed previously in terms of primitive/derived states,
and for the first time they are both observed in a fossilattributed to the genus Polypterus:
1. The scales of the lateral line bear a groove thatopens posteriorly in a notch and this is regarded asa primitive state for polypteriforms. The derivedstate is thought to be lateral line pores piercing thebody of the scale. This assumption was made byPoll on the basis of ontogenetic arguments.P. faraou shows this character state, as do P. bichir,P. endlicheri and P. ansorgii.
2. A deep and short body (low vertebrae count) is aprimitive state for polypteriforms. This assumption(Daget & Desoutter, 1983) is congruent with thebody shape of the Cretaceous Serenoichthys (asnoted by Dutheil, 1999), and also with the generalobservations made on the elongation of the bodyamong actinopterygians. P. faraou has a short anddeep body with 46 scales in a transversal row, as
Figure 10. The Recent polypterid species and Polypterus faraou sp. nov. (A) Position of the species in the anatomicallybased clusters of species, as defined by Poll, P. teugelsi not included (see Britz, 2004, for discussion), and (B) distribution inthe African hydrographical basins; data from FishBase (Froese & Pauly, 2005) and map of the ichthyofaunic regions ofAfrica from Roberts (1975).

236 O. OTERO ET AL.
© 2006 The Linnean Society of London, Zoological Journal of the Linnean Society, 2006, 146, 227–237
in P. bichir, P. endlicheri, P. weeksii and P. ansorgii,and with fewer than 60 scales in a longitudinal row,as in all Recent species except P. ornatipinnis,P. bichir and Erpetoichthys. We can also considerthe trend to anguilliform shape through the posi-tion and the count of the dorsal finlets. On thisbasis, P. faraou has no anguilliform character, thenumber of pinnules being greater than 12, and thecount of predorsal scales being lower than 15 (as inP. bichir, P. endlicheri and P. ansorgii).
The polypterid from Toros-Menalla (westernDjurab, Chad) described here is the first fossil speci-men known by a near complete articulated skeletonincluding a cranium. It is assigned to the genusPolypterus and recognized as a new species, Polypterusfaraou. It is c. 7 Myr in age (Vignaud et al., 2002).
ACKNOWLEDGEMENTS
We thank the Chadian authorities (Ministère del’Education Nationale et de la Recherche, Universitéde N’Djaména, CNAR). For their support, we extendour gratitude to the French Ministère de l’Enseigne-ment supérieur, de la Recherche et de la Technologie(CNRS; Université de Poitiers) and the Ministèredes Affaires Etrangères (DCSUR, Paris; CoopérationSCAC, N’Djaména), to the Région Poitou-Charentesand the Département de la Vienne. The MPFT field-work is also supported by an NSF/RHOI Grant. Wethank Emmanuel Fara (CNRS, Poitiers) for discus-sion, the referees for their reviews and comments, andalso all members of the MPFT. Drawings and photo-graphs by Olga Otero.
REFERENCES
Britz R. 2004. Polypterus teugelsi, a new species of bichir fromthe Upper Cross River system in Cameroon (Actinopterygii:Cladistia: Polypteridae). Ichthyological Explorations ofFreshwaters 15: 179–186.
Brunet M, MPFT. 2000. Chad: discovery of a vertebratefauna close to the Mio-Pliocene boundary. Journal of Verte-brate Paleontology 20: 205–209.
Brunet M, Beauvilain A, Coppens Y, Heintz E, MoutayeEHA, Pilbeam D. 1995. The first australopithecine 2500kilometers west of the Rift Valley (Chad). Nature 378: 273–275.
Brunet M, Beauvilain A, Geraards D, Guy F, Kasser M,Mackaye HT, MacLatchy L, Mouchelin G, Sudre J,Vignaud P. 1998. Tchad: découverte d’une faune demammifères du Pliocène inférieur. Comptes Rendus del’Académie des Sciences, Paris, Série IIa 326: 153–158.
Brunet M, Guy F, Pilbeam D, Mackaye HT, Likius A,Djimdoumalbaye A, Beauvilain A, Blondel C, Bocher-ens H, Boisserie JR, de Bonis L, CoppensY, Dejax J,Denys C, Duringer P, EisenmannV, Gongdibé F,
Fronty P, Geraads D, LehmannT, Lihoreau F,Louchart A, Adoum M, Merceron G, Mouchelin G,Otero O, Pelaez Campomanes P, Ponce de Leon M,Rage JC, Sapanet M, Schuster M, Sudre J, Tassy P,Valentin X, Vignaud P, Viriot L, Zazzo A. 2002. A newhominid from the Upper Miocene of Chad, Central Africa.Nature 418: 145–151.
Daget J. 1950. Révision des affinités phylogénétiques desPolyptéridés. Mémoires de l’Institut Français d’Afrique Noire11: 1–178.
Daget J, Desoutter M. 1983. Essais de classification cladis-tique des polyptéridés (Pisces; Brachiopterygii). Bulletin duMuséum National d’Histoire Naturelle, Paris, Série A 4: 661–674.
Dutheil D. 1999. The first articulated fossil cladistian: Ser-enoichthys kemkemensis gen. et sp. nov., from the Creta-ceous of Morocco. Journal of Vertebrate Paleontology 19:243–246.
Froese R, Pauly D (eds). 2005. FishBase. World Wide Webelectronic publication. www.fishbase.org.
Gayet M, Meunier F. 1991. First discovery of Polypteridae(Pisces, Cladistia, Polypteriformes) outside of Africa. Géobios24: 463–467.
Gayet M, Meunier F. 1996. Nouveaux Polypteriformes duConiacien-Sénonien d’In Becetem (Niger). Comptes Ren-dus de l’Académie des Sciences, Paris, Série IIa 322: 701–707.
Gosse J-P. 1990. Polypteridae. In: Lévêque C, Paugy D, Teu-gels GG, eds. Faune des poissons d’eaux douces et saumâtresd’Afrique de l’Ouest I. MRAC Orstom, collection faune trop-icale, 79–87.
Hanssens MM, Teugels GG, Thys van den AudenaerdeDFE. 1995. Suspecies in the Polypterus palmas complex(Brachiopterygii; Polypteridae) from West and CentralAfrica. Copeia 1995: 694–705.
Jollie M. 1984. Development of the head and pectoral skeletonof Polypterus with a note on scales (Pisces: Actinopterygii).Journal of Zoology, London 204: 469–507.
Meunier F, Gayet M. 1996. A new Polypteriform from theLate Cretaceous and the middle Paleocene of South America.In: Arratia G, Viohl G, eds. Mesozoic Fishes, 1. Systematicsand paleoecology. Proceedings of the 1st International Meet-ing on Mesozoic Fishes, Eichstätt, 1993. München, Germany:Verlag Dr. F. Pfeil, 93–103.
Pehrson T. 1947. Some new interpretations of the skull inPolypterus. Acta Zoologica, Stockholm 28: 241–258.
Poll M. 1941a. Contribution à l’étude systématique des Poly-pteridae (première partie). Revue de Zoologie et de BotaniqueAfricaines 35: 141–179.
Poll M. 1941b. Les tendances évolutives des Polyptèresd’après l’étude systématique des espèces. Annales de laSociété Royale Zoologique de Belgique 72: 157–173.
Poll M. 1942. Contribution à l’étude systématique des Poly-pteridae (suite et fin). Revue de Zoologie et de BotaniqueAfricaines 35: 269–308.
Poll M. 1965. Anatomie et systématique des Polyptères. Bul-letin de l’Académie Royale de Belgique, Sciences 1965: 553–569.

A NEW POLYPTERID FROM THE LATE MIOCENE OF CHAD 237
© 2006 The Linnean Society of London, Zoological Journal of the Linnean Society, 2006, 146, 227–237
Roberts TR. 1975. Geographical distribution of African fresh-water fishes. Zoological Journal of the Linnean Society 57:249–319.
Rosen DE, Forey PL, Gardiner B, Patterson C. 1981.Lungfishes, tetrapods, paleontology and plesiomorphy. Bul-letin of the American Museum of Natural History 167: 159–276.
Smith JA. 1865. Description of Erpetoichthys, a new genus ofganoid fish, from Old Calabar, Western Africa; forming anaddition to the Family Polypterini. Proceedings of the RoyalSociety of Edinburgh 1865–1866: 654–659.
Vignaud P, Duringer P, Mackaye HT, Likius A, BlondelC, Boisserie JR, de Bonis L, Eisenmann V, GeraadsD, Guy F, Lehmann T, Lihoreau F, Lopez-MartinezN, Mourer-Chauviré C, Otero O, Rage J-C, SchusterM, Viriot L, Zazzo A, Brunet M. 2002. Geology andpalaeontology of the Upper Miocene Toros-Menalla fossilif-erous area, Djurab Desert, Northern Chad. Nature 418:152–155.
Werner C, Gayet M. 1997. New fossil Polypteridae from theCenomanian of Sudan. An evidence of their high diversity inthe Early Late Cretaceous. Cybium 21: 67–81.
Copyright © 2022 FDOKUMEN


![GÉOPOLITIQUE ET POPULATIONS AU TCHAD [Geopolitics and populations in Chad]](https://static.fdokumen.com/doc/165x107/631378e5fc260b71020f1c3f/geopolitique-et-populations-au-tchad-geopolitics-and-populations-in-chad.jpg)

















