
The complex policy issue of elephant ivory stockpile management
Upload
independentCategory
view
2download
0
535
Journal of Vertebrate Paleontology 21(3):535–546, September 2001q 2001 by the Society of Vertebrate Paleontology
A NEW GENUS OF MACROSCELIDEA (MAMMALIA) FROM THE EOCENE OF ALGERIA:A POSSIBLE ORIGIN FOR ELEPHANT-SHREWS
RODOLPHE TABUCE1, BRIGITTE COIFFAIT2, PHILIPPE-EMMANUEL COIFFAIT2, MOHAMED MAHBOUBI3, andJEAN-JACQUES JAEGER1
1Institut des Sciences de l’Evolution de Montpellier, UMR 5554 du CNRS, Universite Montpellier II,34095 Montpellier cedex 5, France;
2Laboratoire de Geologie des Ensembles Sedimentaires, Universite Nancy I, 54506 Vandoeuvre-les-Nancy, France;3Institut des Sciences de la Terre, Universite d’Oran, B.P. El M’naouer, Oran, Algerie
ABSTRACT—Nementchatherium senarhense, gen. et sp. nov. from the Middle–Late Eocene of Bir El Ater (Algeria)is described and assigned to the subfamily Herodotiinae (Macroscelidea). This genus and the other primitive elephant-shrews are compared with the Louisininae (Hyopsodontidae, Condylarthra) from the Early Paleogene of Europe. Thesegroups have been included in a phylogenetic analysis based on dental characters, in order to clarify the origin ofMacroscelidea. Phylogenetic reconstruction suggests that Louisininae are belonging in the polyphyletic Hyopsodontidaeexcept for Microhyus which is considered here as the sister-group the Macroscelidea. These results suggest a terrestrialinterchange between Africa and Eurasia during the Early Eocene. The phylogenetic analysis suggests also that theMacroscelidea-Microhyus clade is closely related to the Proboscidea. Like molecular phylogenies, especially thoseconcerning the African molecular clade (5Afrotheria), our results, provide evidence for a macroscelid-tethytherianrelationship. However, if the Macroscelidea emerged from European ‘‘condylarth’’ at the Early Eocene as our datasuggest, the Proboscidea are already differentiated in Africa during this period. Then, it seems that Macroscelidea andProboscidea are paraphyletic. The assumption of a unique group of condylarthran type at the origin of Afrotheria(macroscelids, tethytherians, tubulidentates, tenrecid and chrysochlorid insectivores) cannot be excluded, but the currentpaleontological data do not fit with that hypothesis.
INTRODUCTION
Living Macroscelidea or elephant-shrews are endemic to Af-rica, and fossil representatives of the order are also known onlyfrom this continent. The fossil record of macroscelids begins inthe Paleogene of North Africa, but only three species are knownbefore the Miocene. The phylogenetic status of this Africanendemic order has long been debated. Early authors placed ma-croscelids among insectivoran Menotyphla (5tupaiids and ma-croscelids) (Haeckel, 1866). Later, Butler (1956) and Patterson(1965) created a new order for elephant-shrews based on theiruniqueness among eutherians. Currently, two hypotheses aredebated for their evolutionary relationship with other mammals.
The first one, based on cladistic analysis of morphologicalcharacters, interprets macroscelids as sister group of the Glires(rodents and lagomorphs) (Novacek et al., 1988). Initially, theMacroscelidea-Lagomorpha clade (McKenna, 1975) was estab-lished on the prismatic aspect of jugal teeth; rodents were sep-arated from the rest of the eutherians and placed in incertaesedis. When the Glires concept was revived (Luckett and Har-tenberger, 1985), macroscelidids were grouped with rodents andlagomorphs on the basis of embryological and cranial charac-ters (Novacek et al., 1988). This opinion was recently assumedby Rose (1999) on postcranial characters, by McKenna and Bell(1997), and by Shoshani and McKenna (1998) in the definitionof the Grandorder Anagalida. This hypothesis implies an Asianorigin for the Macroscelidea. However, the fossil record of ma-croscelidids begins in the Eocene of Africa, and no Asian fossilseems to support such an hypothesis so far. This suggests thatcharacters shared by Glires and some macroscelidids could bethe result of convergences. Indeed the character of the prismaticjugal teeth seems to have been acquired very late (during theMiocene?) in elephant-shrews: the Miocene primitive Rhyncho-cyoninae (Miorhynchocyon) and Macroscelidinae (Pronasilio)have no prismatic jugal teeth (Butler, 1984).
The second hypothesis suggests a condylarthran origin for
the Macroscelidea (Hartenberger, 1986; Simons et al., 1991),an interpretation that followed the discovery of two supposedearly Macroscelidea, Chambius kasserinensis Hartenberger,1986 from the Early–Middle Eocene of Tunisia, and of Hero-dotius pattersoni Simons et al., 1991 from the Late Eocene ofthe Jebel El Qatrani Formation (Fayum, Egypt). As Chambiusand Herodotius are quite evolved by comparison to condylar-thrans, and as they might share dental characters with modernmacroscelidids (Butler, 1995), it seems that the condylarthranhypothesis for the origin of elephant-shrews needs further in-vestigation. Moreover, there is a strong consensus in molecularstudies to related the macroscelids to several ungulates as Tub-ulidentata and Paenungulata. This result is supported by proteinanalysis (de Jong et al., 1993), mitochondrial and nuclear DNA(Springer et al., 1997). Such a result gives some support to thepaleontological hypothesis concerning the condylarthran-ma-croscelid relationships.
SYSTEMATIC PALEONTOLOGY
Order MACROSCELIDEA Butler, 1956Family MACROSCELIDIDAE Bonaparte, 1838
Subfamily HERODOTIINAE Simons et al., 1991NEMENTCHATHERIUM gen. nov.
Type and Only Known Species N. senarhense, sp. nov.Distribution late Middle to Late Eocene, Bir El Ater (Al-
geria) (Fig. 1).Diagnosis Nementchatherium differs from Chambius in
having p4 more molarized with a more developed metaconid,P4 with a postprotocrista linked to the distal cingulum, and inhaving neither conules nor postprotocrista on M1 or 2. Ne-mentchatherium is smaller than Herodotius (holotype P4: L 52.2; W 5 2.4, P4 of Herodotius: L 5 3.1; W 5 3.3), and itdiffers from it by its less molariform p4 and its P4 having acrestiform metacone, and no hypocone. On upper molars, Her-

536 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 21, NO. 3, 2001
FIGURE 1. A, Map showing geographic location of Bir El Ater andJebel Chambi, the two known Maghrebi sites which are yielded Eoceneelephant-shrews; B, simplified geological map of the NementchasMountains in the East end of the Algerian Saharan Atlas Mountains.The marine Cretaceous limestone is overlain in concordance by theYpresian (limestone with marl intercalations) and by an other marinesuccession composed of marls and gypsum with mollusks and Lutetianostracods (Faid, 1999). The vertebrate fauna was found in a basal de-trital layer of a sandy fluvio-deltaic member discordantly overlayingthese marine deposits (Coiffait et al., 1984). The discordance seems torepresent the Lutetian Atlasic tectonic event. At last, the Middle to LateEocene deposits are overlain by the continental Miocene. These strati-graphic data indicate the Bir El Ater locality to be post-Lutetian andante-Neogene in age. Moreover, the phiomyid rodents (Jaeger et al.,1985; Holroyd, 1994) and the hyracoid Bunohyrax (Tabuce et al., 2000)from Bir El Ater, compared to those of the Early Oligocene from theJebel El Qatrani (Fayum, Egypt), also indicate a Middle–Late Eoceneage for the fossiliferous locality.
odotius shows a smaller hypocone and a lower centrocrista be-tween paracone and metacone.
Etymology From Nementcha, East-Algerian mountains,and Greek therium, beast.
NEMENTCHATHERIUM SENARHENSE, sp. nov.(Fig. 2)
Holotype Universite d’Oran (Algeria) specimen numberUON 357, right P4.
Referred Material UON 350, left p4; UON 351, left p4;UON 352, right p4; UON 353, left m1 or 2; UON 354, rightm1 or 2; UON 355, right m3; UON 356, left P4; UON 358,left M1 or 2.
Measurements (in mm) UON 350: L 5 2.4, W 5 1.5;UON 351: L 5 2.0, W 5 1.3; UON 352: L 5 2.5, W 5 1.6;UON 353: L 5 2.0, W 5 1.8; UON 354: L 5 2.2, W 5 1.7;UON 355: L 5 1.4, W 5 1.3; UON 356: L 5 2.0, W 5 2.3;UON 358: L 5 2.2, W 5 2.6; UON 357: L 5 2.2, W 5 2.4.
Diagnosis as for genus.Etymology From the Oued Senarh which is next to the
fossiliferous locality.
DESCRIPTION
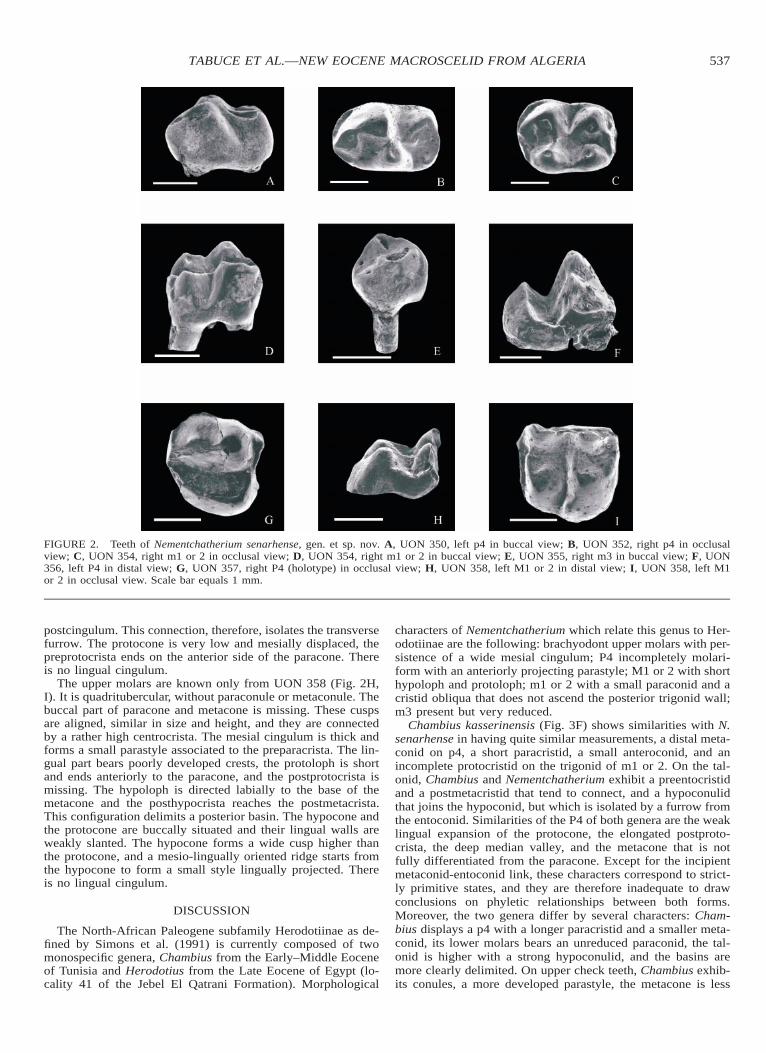
The three p4 (Fig. 2A, B) are clearly molariform, they dis-play an elongated occlusal outline and they are slightly narrow-er and longer than lower molars. The paraconid is as large asthe metaconid, anteriorly projected and centrally placed, low onthe crown, and it is connected to a large protoconid by a shortparacristid. On UON 351 and UON 352, the metaconid is lowerand narrower and more distal than the protoconid. These twonearly twinned cusps are linked by a weak protocristid. Thedistal side of the protoconid and metaconid constitute a verticalwall, this character being well observed in buccal view on theless worn lower premolar UON 352. The trigonid basin issmall, very lingually displaced, and only a slight mesial furrowcan be seen at the base of the protoconid. On UON 352, thisbasin is hardly delineated and widely open on the lingual side;the paraconid is more developed and its most mesial part isconnected to the anterior cingulid. The hypoflexid forms a deepsinus that ends on the buccal side of the metaconid. The talonidof p4 is lower and at least twice shorter than the trigonid. Thetalonid is also characterized by a robust cristid obliqua that runsanterolingually from the hypoconid to the posterior wall of themetaconid. The length of the cristid obliqua is variable: onUON 352 it is short, but on UON 350 it ends very high on themetaconid. The talonid basin is reduced by the proximity of thelarge hypoconid to the small entoconid; both cusps are trans-versely in line and joined by a weak postcristid; there is nothypoconulid.
Both m1 or 2 (Fig. 2C, D) have the same morphology andthey are characterized by their rectangular outline, a high buccalcoronal elevation, and a slight exodaenodonty (the labial lobesof enamel extend ventrally over the roots). The mesial cingulidbetween metaconid and paraconid forms a small anteroconid.The paraconid is present but crestiform, it is buccally displaced,and very close to the protoconid. The protoconid and the meta-conid are subequal in size and transversely aligned; these cuspsare connected by an incipient protocristid. The basin of thetrigonid is short and open on the mesial cingulid. The talonidis narrower and slightly lower than the somewhat procumbenttrigonid. On the talonid, the hypoflexid is deep, the cristid ob-liqua is short, and it ends at the base of the posterior trigonidwall. The entoconid and hypoconid are transversely aligned andsubequal in size. The entoconid end metaconid are similar inmorphology and they are vertical whereas the buccal cusps havetheir distal side anteriorly inclined. The hypoconulid is reduced,it is situated close to the longitudinal axis, and it bears a weakpostcristid that does not reach the entoconid. There is a notchbetween the metaconid and the entoconid, thus the postmeta-cristid is not entirely connected to the entocristid. The distalcingulid is absent, and only a small style occurs on the buccalside of the hypoconulid.
The m3 (Fig. 2E) is two-rooted, circular in shape, consider-ably reduced and has a very simplified morphology. On thetrigonid, the protoconid and metaconid are mesiodistally elon-gated and subequal in size; the paraconid is linked to the pro-toconid. The talonid is very reduced, with neither hypoconulidnor posterior lobe; the cristid obliqua is absent, and the ento-conid and hypoconid are tiny and worn.
Both P4 (Fig. 2F, G) are slightly molariform, and they lackconules. They exhibit a moderate parastyle, anteriorly projectedand connected to a short mesial cingulum. The paracone is tallerthan the metacone. On the holotype, the metacone is more in-dividualized than in UON 356. The paracone, the metacone anda small metastyle are in line, and they are joined by the cen-trocrista and the postmetacrista. This strong buccal structuredominates a sharp mesiodistally oriented postprotocrista. Thiscrest forms a postero-internal swelling (a pseudo hypocone),and it curves in its distal part to reach the metastyle and the
537TABUCE ET AL.—NEW EOCENE MACROSCELID FROM ALGERIA
FIGURE 2. Teeth of Nementchatherium senarhense, gen. et sp. nov. A, UON 350, left p4 in buccal view; B, UON 352, right p4 in occlusalview; C, UON 354, right m1 or 2 in occlusal view; D, UON 354, right m1 or 2 in buccal view; E, UON 355, right m3 in buccal view; F, UON356, left P4 in distal view; G, UON 357, right P4 (holotype) in occlusal view; H, UON 358, left M1 or 2 in distal view; I, UON 358, left M1or 2 in occlusal view. Scale bar equals 1 mm.
postcingulum. This connection, therefore, isolates the transversefurrow. The protocone is very low and mesially displaced, thepreprotocrista ends on the anterior side of the paracone. Thereis no lingual cingulum.
The upper molars are known only from UON 358 (Fig. 2H,I). It is quadritubercular, without paraconule or metaconule. Thebuccal part of paracone and metacone is missing. These cuspsare aligned, similar in size and height, and they are connectedby a rather high centrocrista. The mesial cingulum is thick andforms a small parastyle associated to the preparacrista. The lin-gual part bears poorly developed crests, the protoloph is shortand ends anteriorly to the paracone, and the postprotocrista ismissing. The hypoloph is directed labially to the base of themetacone and the posthypocrista reaches the postmetacrista.This configuration delimits a posterior basin. The hypocone andthe protocone are buccally situated and their lingual walls areweakly slanted. The hypocone forms a wide cusp higher thanthe protocone, and a mesio-lingually oriented ridge starts fromthe hypocone to form a small style lingually projected. Thereis no lingual cingulum.
DISCUSSION
The North-African Paleogene subfamily Herodotiinae as de-fined by Simons et al. (1991) is currently composed of twomonospecific genera, Chambius from the Early–Middle Eoceneof Tunisia and Herodotius from the Late Eocene of Egypt (lo-cality 41 of the Jebel El Qatrani Formation). Morphological
characters of Nementchatherium which relate this genus to Her-odotiinae are the following: brachyodont upper molars with per-sistence of a wide mesial cingulum; P4 incompletely molari-form with an anteriorly projecting parastyle; M1 or 2 with shorthypoloph and protoloph; m1 or 2 with a small paraconid and acristid obliqua that does not ascend the posterior trigonid wall;m3 present but very reduced.
Chambius kasserinensis (Fig. 3F) shows similarities with N.senarhense in having quite similar measurements, a distal meta-conid on p4, a short paracristid, a small anteroconid, and anincomplete protocristid on the trigonid of m1 or 2. On the tal-onid, Chambius and Nementchatherium exhibit a preentocristidand a postmetacristid that tend to connect, and a hypoconulidthat joins the hypoconid, but which is isolated by a furrow fromthe entoconid. Similarities of the P4 of both genera are the weaklingual expansion of the protocone, the elongated postproto-crista, the deep median valley, and the metacone that is notfully differentiated from the paracone. Except for the incipientmetaconid-entoconid link, these characters correspond to strict-ly primitive states, and they are therefore inadequate to drawconclusions on phyletic relationships between both forms.Moreover, the two genera differ by several characters: Cham-bius displays a p4 with a longer paracristid and a smaller meta-conid, its lower molars bears an unreduced paraconid, the tal-onid is higher with a strong hypoconulid, and the basins aremore clearly delimited. On upper check teeth, Chambius exhib-its conules, a more developed parastyle, the metacone is less

538 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 21, NO. 3, 2001
FIGURE 3. Teeth of Louisininae and Macroscelidea. A, Dipavali pe-tri, M2 and m2–m3; B, Louisina mirabilis, P4–M3 and p4–m3; C,Monshyus praevius, M1 or 2; D, Microhyus musculus, M1 or 2; E,Microhyus reisi, composite M1–M3 and p4–m3; F, Chambius kasser-inensis, P4–M3 and p4–m3; G, Nementchatherium senarhense, com-posite P4–M1 or 2 and p4–m3; H, Herodotius pattersoni, P4–M3 andp4–m3; I, Miorhynchocyon, composite P4–M1 and p4–m1; J, Metol-dobotes, M2 and p4–m2. (F, H, I modified from Butler, 1984 and 1995).Scale bar equals 1 mm.
individualized from the paracone; on P4 the postprotocrista isnot connected to the distal cingulum, and on molars the centro-crista is lower, and the hypocone is more reduced.
Herodotius pattersoni (Fig. 3H) and N. senarhense share re-duced paraconid (1) and hypoconulid (2) on m1 or 2; a short-ened paracristid (3), a well-individualized metaconid (4), abroad cristid obliqua (5), and the absence of hypoconulid (6)on the molariform p4. On P4–M2 conules are lacking (7), onM1–2 a short hypoloph and protoloph are developed (8), thehypocone is enlarged (9), and the postprotocrista is lacking(10). According to Butler (1995), most of these characters (1,2, 4, 7, 8, 9, 10) can be considered as synapomorphies. How-ever, Nementchatherium differs from Herodotius by a sharppostmetacristid on m1 or 2, a strong connection between theparaconid and protoconid, a distal metaconid, and a deeper hy-poflexid on p4. The P4 of Herodotius is more molariform witha distinct hypocone, a more distinct metacone as high as the
paracone. The upper molars of the Fayum genus exhibit asmaller hypocone and a lower centrocrista between the buccalcusps.
Following these comparisons, it appears that Nementchath-erium constitutes a morphological intermediate between Cham-bius and Herodotius, thus filling the gap between these twoprimitive elephant-shrews. The set of characters observed inNementchatherium suggests affinities with Herodotius, but a di-rect filiation cannot yet be supported. The reason is that theAlgerian species is too peculiar, by its mosaic of primitive(small size and weak molarization of P4) and derived characters(M1 or 2 with hypoloph and protoloph, and hypocone higherthan protocone) to be referred to Herodotius.
The third known Paleogene Macroscelidea is Metoldobotesstromeri Schlosser, 1910 from the Early Oligocene of the Fa-yum, which constitutes the unique member of the subfamilyMetoldobotinae. This taxon was initially described from a man-dible and, more recently, Simons et al. (1991) have referred anisolated M2 to the genus (Fig. 3J). Its m1, as in Nementchath-erium, exhibits a buccal coronal elevation, the lack of distalcingulid and similar metaconid and entoconid. However, thelower teeth of Metoldobotes are distinctively different fromthose of Herodotiinae by the lost of m3 as in some living forms,the great reduction of m2, and the cristid obliqua that runs tothe center of the posterior face of the trigonid on the m1. TheM2 of Metoldobotes differs from that of Herodotius and Ne-mentchatherium in having retained a metaconule, by its narrow-er posterior half, a long protoloph that reaches the parastyle,and by the absence of cingulum. Thus the position of Metol-dobotes within the primitive macroscelidids remains problem-atic, and only the examination of recently discovered skulls inEgypt (Butler, 1995) might help to shed light on the problemof the status of this very specialized genus.
According to Butler (1995), the teeth of the Herodotiinaereproduce in a more primitive way some characters of Neogenemacroscelids. The m3/M3 are very reduced in size as in theRhynchocyoninae, Myohyracinae, and some Macroscelidinae.The long and fully molariform p4, the occurrence of protolophand hypoloph on upper molars are common characters betweenNementchatherium, Herodotius and several latter macroscelids.Moreover, the Rhynchocyoninae, especially Miorhynchocyonfrom the Miocene of Kenya (Butler, 1984) (Fig. 3I) shares sev-eral characters with Nementchatherium and Herodotius: on theirp4, the trigonid is widened, the metaconid is strong, the cristidobliqua ends high on the posterior wall of the metaconid, thehypoconulid is absent, and the entoconid is connected to thehypoconid. This morphology might indicate phylogenetic linksbetween Herodotiinae and Rhynchocyoninae. This Neogenesubfamily is regarded as having an early origin in the Paleogene(Novacek, 1984), but the characters of p4 are often convergentin eutherian mammals, and according to Butler’s statement(1995), the protocone-metaconule link (postprotoloph) on uppermolars of Miorhynchocyon seems to exclude a close relation-ship between herodotiines and rhynchocyonines.
Early Paleogene African faunas that have yielded taxa withbunodont quadritubercular teeth are scarce, and comparisonswith the Herodotiinae are thus limited. An indeterminate ‘‘con-dylarth’’ from the Thanetian of Adrar Mgorn (Ouarzazate Ba-sin, Morocco) suggests affinities with some macroscelidids(Gheerbrant, 1995) on the basis of its size, its robustness, itsprocumbent trigonid, and comparable proportions of trigonidand talonid. However, this lower molar is too fragmentary tobe referred to Herodotiinae. The Early Eocene controversial pri-mate Azibius trerki Sudre, 1975 of Gour Lazib (Algeria) mightbe compared to N. senarhense. Indeed, Szalay (1975) suggeststhat this taxon could be a hyopsodontid-like ‘‘condylarth,’’whereas Patterson (in Sudre, 1979) noted similarities with Me-toldobotes. Finally, Hartenberger et al. (1997) compared it to
539TABUCE ET AL.—NEW EOCENE MACROSCELID FROM ALGERIA
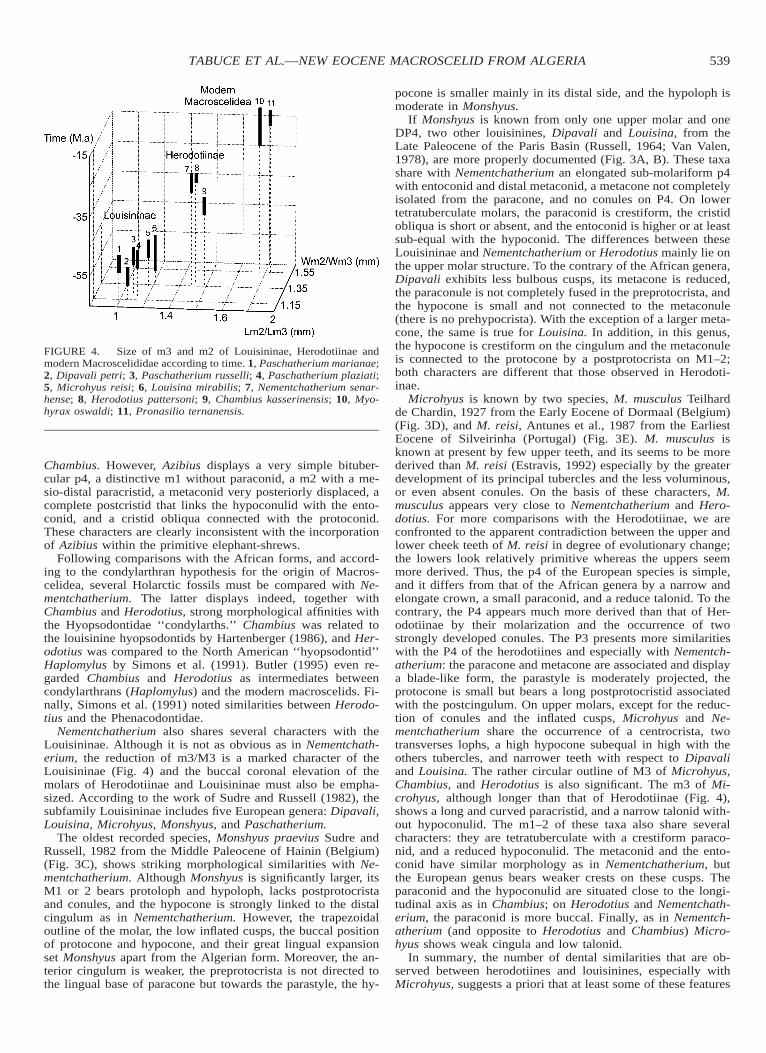
FIGURE 4. Size of m3 and m2 of Louisininae, Herodotiinae andmodern Macroscelididae according to time. 1, Paschatherium marianae;2, Dipavali petri; 3, Paschatherium russelli; 4, Paschatherium plaziati;5, Microhyus reisi; 6, Louisina mirabilis; 7, Nementchatherium senar-hense; 8, Herodotius pattersoni; 9, Chambius kasserinensis; 10, Myo-hyrax oswaldi; 11, Pronasilio ternanensis.
Chambius. However, Azibius displays a very simple bituber-cular p4, a distinctive m1 without paraconid, a m2 with a me-sio-distal paracristid, a metaconid very posteriorly displaced, acomplete postcristid that links the hypoconulid with the ento-conid, and a cristid obliqua connected with the protoconid.These characters are clearly inconsistent with the incorporationof Azibius within the primitive elephant-shrews.
Following comparisons with the African forms, and accord-ing to the condylarthran hypothesis for the origin of Macros-celidea, several Holarctic fossils must be compared with Ne-mentchatherium. The latter displays indeed, together withChambius and Herodotius, strong morphological affinities withthe Hyopsodontidae ‘‘condylarths.’’ Chambius was related tothe louisinine hyopsodontids by Hartenberger (1986), and Her-odotius was compared to the North American ‘‘hyopsodontid’’Haplomylus by Simons et al. (1991). Butler (1995) even re-garded Chambius and Herodotius as intermediates betweencondylarthrans (Haplomylus) and the modern macroscelids. Fi-nally, Simons et al. (1991) noted similarities between Herodo-tius and the Phenacodontidae.
Nementchatherium also shares several characters with theLouisininae. Although it is not as obvious as in Nementchath-erium, the reduction of m3/M3 is a marked character of theLouisininae (Fig. 4) and the buccal coronal elevation of themolars of Herodotiinae and Louisininae must also be empha-sized. According to the work of Sudre and Russell (1982), thesubfamily Louisininae includes five European genera: Dipavali,Louisina, Microhyus, Monshyus, and Paschatherium.
The oldest recorded species, Monshyus praevius Sudre andRussell, 1982 from the Middle Paleocene of Hainin (Belgium)(Fig. 3C), shows striking morphological similarities with Ne-mentchatherium. Although Monshyus is significantly larger, itsM1 or 2 bears protoloph and hypoloph, lacks postprotocristaand conules, and the hypocone is strongly linked to the distalcingulum as in Nementchatherium. However, the trapezoidaloutline of the molar, the low inflated cusps, the buccal positionof protocone and hypocone, and their great lingual expansionset Monshyus apart from the Algerian form. Moreover, the an-terior cingulum is weaker, the preprotocrista is not directed tothe lingual base of paracone but towards the parastyle, the hy-
pocone is smaller mainly in its distal side, and the hypoloph ismoderate in Monshyus.
If Monshyus is known from only one upper molar and oneDP4, two other louisinines, Dipavali and Louisina, from theLate Paleocene of the Paris Basin (Russell, 1964; Van Valen,1978), are more properly documented (Fig. 3A, B). These taxashare with Nementchatherium an elongated sub-molariform p4with entoconid and distal metaconid, a metacone not completelyisolated from the paracone, and no conules on P4. On lowertetratuberculate molars, the paraconid is crestiform, the cristidobliqua is short or absent, and the entoconid is higher or at leastsub-equal with the hypoconid. The differences between theseLouisininae and Nementchatherium or Herodotius mainly lie onthe upper molar structure. To the contrary of the African genera,Dipavali exhibits less bulbous cusps, its metacone is reduced,the paraconule is not completely fused in the preprotocrista, andthe hypocone is small and not connected to the metaconule(there is no prehypocrista). With the exception of a larger meta-cone, the same is true for Louisina. In addition, in this genus,the hypocone is crestiform on the cingulum and the metaconuleis connected to the protocone by a postprotocrista on M1–2;both characters are different that those observed in Herodoti-inae.
Microhyus is known by two species, M. musculus Teilhardde Chardin, 1927 from the Early Eocene of Dormaal (Belgium)(Fig. 3D), and M. reisi, Antunes et al., 1987 from the EarliestEocene of Silveirinha (Portugal) (Fig. 3E). M. musculus isknown at present by few upper teeth, and its seems to be morederived than M. reisi (Estravis, 1992) especially by the greaterdevelopment of its principal tubercles and the less voluminous,or even absent conules. On the basis of these characters, M.musculus appears very close to Nementchatherium and Hero-dotius. For more comparisons with the Herodotiinae, we areconfronted to the apparent contradiction between the upper andlower cheek teeth of M. reisi in degree of evolutionary change;the lowers look relatively primitive whereas the uppers seemmore derived. Thus, the p4 of the European species is simple,and it differs from that of the African genera by a narrow andelongate crown, a small paraconid, and a reduce talonid. To thecontrary, the P4 appears much more derived than that of Her-odotiinae by their molarization and the occurrence of twostrongly developed conules. The P3 presents more similaritieswith the P4 of the herodotiines and especially with Nementch-atherium: the paracone and metacone are associated and displaya blade-like form, the parastyle is moderately projected, theprotocone is small but bears a long postprotocristid associatedwith the postcingulum. On upper molars, except for the reduc-tion of conules and the inflated cusps, Microhyus and Ne-mentchatherium share the occurrence of a centrocrista, twotransverses lophs, a high hypocone subequal in high with theothers tubercles, and narrower teeth with respect to Dipavaliand Louisina. The rather circular outline of M3 of Microhyus,Chambius, and Herodotius is also significant. The m3 of Mi-crohyus, although longer than that of Herodotiinae (Fig. 4),shows a long and curved paracristid, and a narrow talonid with-out hypoconulid. The m1–2 of these taxa also share severalcharacters: they are tetratuberculate with a crestiform paraco-nid, and a reduced hypoconulid. The metaconid and the ento-conid have similar morphology as in Nementchatherium, butthe European genus bears weaker crests on these cusps. Theparaconid and the hypoconulid are situated close to the longi-tudinal axis as in Chambius; on Herodotius and Nementchath-erium, the paraconid is more buccal. Finally, as in Nementch-atherium (and opposite to Herodotius and Chambius) Micro-hyus shows weak cingula and low talonid.
In summary, the number of dental similarities that are ob-served between herodotiines and louisinines, especially withMicrohyus, suggests a priori that at least some of these features

540 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 21, NO. 3, 2001
are synapomorphies. Therefore, McKenna and Bell (1997) haveincluded Chambius into the condylarthran louisinines. However,Herodotius is still referred to true macroscelidids which arerelated to the Glires. Hartenberger (1986) considered the incip-ient lophodonty and the nearly identical height of both talonidand trigonid of Chambius as derived characters, heralding mod-ern macroscelidids. However, lophodonty appears to be a dis-tinct character in some louisinines (Microhyus and Monshyus),and Nementchatherium displays a talonid that is lower than thetrigonid. Then, the assumption of McKenna and Bell (1997)which attributes Chambius, and thus Nementchatherium, to thecondylarthrans could be defended. Nevertheless, we are in dis-agreement with that opinion because it discounts the possibilityof a convergent evolution between the Herodotiinae and theLouisininae. We therefore propose to test if Herodotiinae–Louisininae and Herodotiinae–Macroscelidea similarities arestrictly homoplasic or if they reveal phylogenetic relationships.Furthermore, the following analysis could also clarify phylo-genetic relationships between Ungulata and Macroscelidea.
PHYLOGENETIC ANALYSIS
Choice of Characters and Taxa (see also Appendix 1)
Phylogenetic relationships between archaic and modern un-gulates are the subject of numerous discussions (see Janis etal., 1998, for an overview). Our aim is not to propose a newphylogeny of condylarthrans, such a study would require amore detailed work. Nevertheless it was necessary to test thecondylarthran hypothesis for the origin of elephant-shrews, in-cluding at least one representative of each ‘‘condylarth’’ familyin the analysis.
Taxa included in the analysis correspond to archaic ungulatesfamilies: Protungulatum (Ungulata incertae sedis), Loxolophus(Arctocyonidae), Hyopsodus, Litomylus, Haplaletes and Hap-lomylus (Hyopsodontidae), Mioclaenus (Mioclaenidae), Perip-tychus (Periptychidae), Eoconodon (Hapalodectidae), and final-ly Ectocion and Phenacodus (Phenacodontidae). Diacodexis(Artiodactyla), Hyracotherium (Perissodactyla), Numidotherium(Proboscidea), and Bunohyrax (Hyracoidea) characterize mod-ern ungulates.
We also included the Louisininae: Dipavali, Louisina, Mi-crohyus and Paschatherium. McKenna and Bell (1997) addedKhamsaconus in the subfamily. This genus known from theYpresian of Morocco, described as a possible Louisininae bySudre et al. (1993), is now related to tethytherians by Gheer-brant et al. (1998). In any case, Monshyus and Khamsaconus,described from only one tooth, were not included in the phy-logenetic analysis.
Today, the Herodotiinae include three genera previously dis-cussed: Chambius, Herodotius and Nementchatherium. Metol-dobotes (Metoldobotinae) from the Oligocene of Fayum, Myo-hyrax (Myohyracinae), Miorhynchocyon (Rhynchocyoninae),and Pronasilio (Macroscelidinae) from the Miocene of East Af-rica represent the modern macroscelidids. As Pronasilio isknown only by its lower dentition, the characters of the upperteeth of the macroscelidines used here are those of Elephantulusfrom the Early Pleistocene of Olduvai Bed I.
Rooting and outgroup comparisons are ensured by Palae-chthon (Plesiadapiform) from the Middle Paleocene of NorthAmerica and by two Cretaceous genera stemming in Zhelesti-dae (Aspanlestes and Eoungulatum), supposed to represent thesister-group of ungulates (Nessov et al., 1998).
Herodotiinae and the Miocene macroscelidids are describedonly by dental characters. Therefore, only these characters (Ap-pendix 1) are taken into account in the following analysis. Insuch an investigation, dental and postcranial characters are of-ten judged to be less informative than cranial data because theycould be more homoplasic in reference to strong selective pres-
sures on feeding and locomotion. Cranial characters are thusconsidered more valid to solve phylogenetic problems. How-ever, Sanchez-Villagra and Williams (1998) have statisticallydemonstrated that the rates of homoplasy in dental, postcranialand cranial characters were not significantly different. Further-more, as in all phylogenetic analysis, the choice of charactersis a major difficulty, and their homology constitutes a startingassumption.
Analysis and Results
The data matrix of 44 characters for 29 taxa (Appendix 2)has been processed with MacClade 3.01 and analyzed with theheuristic option of the PAUP 3.1.1. algorithm by stepwise ad-dition (random addition sequences, 100 replications). All char-acters are coded as reversible and multistate characters as un-ordered. The cladistic analysis generated 6 equally parsimoni-ous trees of 285 steps (consistency index, CI 5 0.530 and re-tention index RI 5 0.601). The consensus tree built from thesesix topologies is represented in Figure 5. As our objective isnot to discuss the phylogeny of condylarthrans, we will consid-er only the intra- and interordinal relationships of Macrosceli-dea in our work, especially with living ungulates. The simpli-fied cladogram is provided in Figure 6.
The monophyly of the order Macroscelidea is observed in alltopology. In four trees, the subfamily Herodotiinae is mono-phyletic, and Nementchatherium has a basal position. In theother topologies, the Herodotiinae are paraphyletic (successivedichotomies) or diphyletic (Chambius roots the Miocene ele-phant-shrews and Nementchatherium-Herodotius root Metol-dobotes). Among the Louisininae, Microhyus is the only onegenus related to the Macroscelidea. The other louisinines, Pas-chatherium, Dipavali and Louisina, are monophyletic (Fig. 6,node 1) and included in the polyphyletic Hyopsodontidae.
The Macroscelidea-Microhyus cluster (Fig. 6, node 2) is root-ed by Numidotherium. This genus, representing the Probosci-dea, is closely related to Hyracotherium (Perissodactyla), Bu-nohyrax (Hyracoidea) and Phenacodontidae (Phenacodus andEctocion) (Fig. 6, node 3). This relationship between the Phen-acodontidae and the Altungulata (Perissodactyla–Hyracoidea–Proboscidea–Sirenia–Embrithopoda–Desmostylia) is obviouslycongruent with all data. Current results, well established byother dental, postcranial, and cranial characters (Prothero et al.,1988; Thewissen and Domning, 1992), call for the union ofPhenacodontidae and Altungulata in the Taxeopoda (Archibald,1998). In our analysis Perissodactyla and Hyracoidea display asister-group relationship in 67% of the cases, but successivedichotomies between Perissodactyla–Hyracoidea–Proboscideacan also be observed.
Discussion
Novacek (1984) presented a phylogenetic tree for the Ma-croscelidea based on dental and cranial characters. For this au-thor, a multifurcation including Metoldobotes, the Rhyncho-cyoninae, the Myohyracinae, and the Macroscelidinae appearsduring the Paleocene–Eocene. Our analysis and the sister grouprelationships between Herodotiinae and the modern elephant-shrews confirm this age of differentiation. The dental charactersof Herodotiinae (derived with respect to those of ‘‘condy-larths,’’ and primitive compared to those of Miocene elephant-shrews) allow to shed light on the phylogeny of modern ele-phant-shrews at the subfamily level: the Rhynchocyoninae rootsthe Myohyracinae-Macroscelidinae clade. The Herodotiinae aremonophyletic and Nementchatherium has a basal position bythe trigonid higher than the talonid on m1–2 (Character 10-state1: Ch10-1) and the small parastyle on P4 (Ch26-2). Chambiusacquired a primitive morphology by reversion of three charac-ters (return to primitive states): the postprotocrista is present on

541TABUCE ET AL.—NEW EOCENE MACROSCELID FROM ALGERIA
FIGURE 5. Cladogram showing the phylogenetic relationships of Macroscelidea, archaic and modern ungulates. This topology is the majority-rule consensus of the six most parsimonious trees based on 44 unweighted characters (see Appendices 1 and 2). Length, 285 steps; consistencyindex, 0.530; retention index, 0.601. The values indicate, when they are not 100%, the frequency of occurrence of groupings.
M1–2 (Ch41-1), the paraconid is well developed (Ch11-0), andthe protoconid is anterior on m1–2 (Ch13-1). Similar changesof character states are also observed, probably because of thecolonization of the insectivorous niche, among Miorhyncho-cyon, Pronasilio, and Myohyrax. Therefore, only one treeshows that the Herodotiinae are diphyletic and Chambius rootsthese modern elephant-shrews. These uncertain relationships forthe herodotiines can be explained by the lack of data concerningthe oldest macroscelids. They were certainly more diversifiedthan actually illustrated by the known fossil record (Fig. 6).Indeed, Nementchatherium seems to illustrate, by its peculiarmosaic of characters (less molariform P4 and evolved M1–2),the diversity of Macroscelidea during the Paleogene in Africa.Morphological comparisons of these primitive elephant-shrewsrelate the Herodotiinae to the louisinine condylarthrans (molar-ization of p4, morphology of M1–2, and strong reduction ofm3/M3). But the phylogenetic analysis demonstrates a conver-gent evolution with the exception of Microhyus. Therefore, ouranalysis supports Sudre and Russell (1982) who consider theLouisininae as Hyopsodontidae having achieved a primitivestage corresponding to the morphology seen in Haplaletes.However, we consider that the louisinine clade (Node 2) in-cludes only Paschatherium, Dipavali, and Louisina. Althoughsome authors consider that Paschatherium is an erinaceomorph-like insectivore (Van Valen, 1967; Collinson and Hooker, 1987),our cladogram argues for a condylarthran status for this genus.This interpretation is also supported by the functional analysisof its talus (Godinot et al., 1996), by other dental data (Denysand Russell, 1981) and by its strongly posteriorly displacedmental foramen (Estravis and Russell, 1992). Another genus,Microhyus, seems to be excluded from the louisinine subfamilyand is considered here to be the sister group of the Macroscel-idea. This clade is supported by the weak lingual slope of theprotocone on M1–2 (Ch39-1), the trigonid larger than the tal-onid on m3 (Ch21-2) (two convergent characters with Louisi-na), and the lack of the hypoconulid on m3 (Ch19-2).
The relationships between the Macroscelidea-Microhyusclade and the Taxeopoda (Phenacodontidae–Altungulata) is wellsupported by the decay index because 11 steps are necessary
to exclude the modern elephant-shrews from the ‘‘ungulateclade.’’ Moreover, several dental characters support this rela-tionship between Macroscelidea and Taxeopoda. These groupsshare a developed metacone on P4 (Ch23-2) (reversion forNumidotherium) and an enlarged metaconid on p4 (Ch7-2)(convergent character with Paschatherium, reversion for Cham-bius and Microhyus). Also, Macroscelidea and Altungulatashare on M1–2 aligned protocone and paracone (Ch38-1) (con-vergent with Dipavali and Louisina), a hypocone sub-equal orhigher than the protocone (Ch42-2), a hypoloph (Ch43-1) (re-version for Metoldobotes and Miorhynchocyon), and a cresti-form hypoconulid on m1–2 (Ch16-1) (reversion for Microhyusand strong hypocristid for the Miocene elephant-shrews). Ac-cording to other morphological data, these altungulates, repre-sented by the Perissodactyla, Hyracoidea, and Proboscidea inour analysis, form a quite homogeneous unit (Shoshani and Mc-Kenna, 1998). But the position of the hyraxes is still uncertain.Two alternative hypotheses have been tested and there is evi-dence in favor of both (Fischer, 1996). The first hypothesis sup-ports a close relationship between hyraxes and perissodactyls(Fischer and Tassy, 1993) and the second hypothesis supportsa hyrax-tethytherian relationship (Novacek et al., 1988). In ouranalysis, four trees support the first assumption, but the twoother most parsimonious trees support the Paenungulate conceptwhich includes the tethytherians (Proboscidea and Sirenia) andthe hyraxes.
In all retained topologies, the Macroscelidea and Microhyusappear as the sister-group of the proboscidean Numidotherium.Then, when the Paenungulate clade is observed, the elephant-shrews are included in it. Such a result would corroborate themolecular African clade (5Afrotheria) which assigned macros-celids to the paenungulates (de Jong et al., 1993). However, thisconsensus on molecular and morphological data is not validwhen molecular phylogenies also include the chrysochlorids(Springer et al., 1997) and the tenrecids (Stanhope et al., 1998a)in the African clade. The lack of homologous dental charactersbetween ungulates and tenrecids-chrysochlorids does not allowus to include these insectivores in our analysis. Moreover, earlytenrecids and chrysochlorids are unknown before the Miocene.

542 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 21, NO. 3, 2001
FIGURE 6. Simplified phylogenetic relationships and fossil record of primitive Macroscelidea in the ‘‘ungulate complex.’’ The node 1 representsthe subfamily Louisininae, the node 2 symbolizes the Macroscelidea–Microhyus clade, and the node 3 corresponds to the Macroscelidea–Taxeopodaclade.
Currently, there is no morphological or paleontological evi-dence to link these lipotyphlans with African ungulates.
The difference of opinion between molecularists and pale-ontologists also concerns the monophyly of ungulates. Paleon-tological results indicate that modern ungulates and ‘‘condy-larths’’ form a monophyletic group. In contrast, the moleculardata suggest a diphyly for the living ungulates. The phylogeniespublished so far associate perissodactyls and cetartiodactylswith carnivores (Graur et al., 1997), the Paenungulates beingincluded in the Afrotheria clade. The molecular consensus clad-ogram (Waddell et al., 1999) considers that both groups arevery different in terms of phylogeny. These results either implythe convergent evolution of characters of ungulates in two lin-eages (perissodactyls–cetartiodactyls and tethytherians–hyrax-es), or else they infer a secondary loss of these characters inall other mammals. The latter point of view requires the ‘‘con-dylarths’’ to be considered as the stem group of many placen-
tals, as already thought at the end of the 19th century. Then,the first hypothesis appears more parsimonious: the ‘‘condy-larths’’ would form two very distinct groups, one of them beingat the origin of the perissodactyls–cetartiodactyls clade, the oth-er group rooting the tethytherians–hyraxes–macroscelids clade.This molecular theory invalidates our results, as well as allthose based on morphological and paleontological data (Shos-hani and McKenna, 1998).
Stanhope et al. (1998b) suggested that the macroscelids werethe first to differentiate within the Afrotheria clade. If ‘‘Afro-therians’’ constitute a natural group, our data suggest a ‘‘con-dylarth’’ origin for this super-ordinal clade. However, the oc-currence of an African group with diversified dental morphol-ogies, at the origin the Afrotheria clade is now mere conjecture:if the macroscelids seem to have differentiated in the earliestEocene, the fossil record of hyraxes and proboscideans beginsin the Earliest Eocene, and that of tenrecids and chrysochlorids

543TABUCE ET AL.—NEW EOCENE MACROSCELID FROM ALGERIA
in the Miocene. Thus, the discovery of further new key-taxa inPaleocene and Early Eocene sediments of Africa becomes nec-essary for an efficient comparison of the molecular and mor-phological phylogenies.
PALEOBIOGEOGRAPHIC CONCLUSIONS
The African mammalian fauna presents a great degree ofendemism. This endemism culminates during the Paleogenewith the appearance of endemic orders such as Proboscidea(Gheerbrant et al., 1996), Hyracoidea (Sudre, 1979) and Ma-croscelidea (Hartenberger, 1986; this paper). In addition, mod-ern chiropterans (Sige, 1991), endemic rodents (Jaeger et al.,1985; Vianey-Liaud et al., 1994) and extinct endemic taxa likethe ptolemaiids (Bown and Simons, 1987) and Helioseus (Sud-re, 1979) are also recorded during the Paleogene. However, theArabo-African continent was not a strictly isolated land massduring these times and several Paleogene terrestrial interchang-es with the North Tethyan regions are documented (see Mah-boubi et al., 1997, for an overview). The paleoryctid and adap-isoriculid ‘‘insectivores’’ from the Thanetian of the AdrarMgorn locality in Morocco indicate early faunal exchanges withNorth America and Europe around the K/T boundary (Gheer-brant, 1987). Adrar Mgorn has also yielded possible hyaeno-dontid creodonts (Gheerbrant, 1995) and the oldest primateknown (Sige et al., 1990), which testify Paleocene/Eocene in-terchanges with Europe (and perhaps Asia) at the Paleocene/Eocene boundary (Gheerbrant, 1990). Gheerbrant (1990) con-sidered that these late trans-Tethyan dispersal events are a mon-odirectional northward trend, but Vianey-Liaud et al. (1994)also suggest a North-South communication to explain the oc-currence of the zegdoumyid rodents in Maghreb.
The phylogenetic relationships between Proboscidea, Mi-crohyus and Macroscelidea represent another indication in sup-port of Africa–Eurasia faunal exchange around the Paleocene-Eocene limit. The topology of the cladogram suggests two pos-sible paleobiogeographic scenarios. The first hypothesis impliesa South to North communication if the ancestor of Microhyusappears to be an African ‘‘condylarth.’’ The discovery of prim-itive ungulates in the Thanetian–Ypresian of Morocco and Sen-egal (Sudre et al., 1993) supports this hypothesis. One of them,Khamsaconus bulbosus from N’Tagourt 2 (Ypresian, Morocco),now clearly attributed to the tethytherians, was initially attri-buted by morphological convergences to the ‘‘louisinines’’ andwas compared with Microhyus and Monshyus.
However, the absence of macroscelids in the Late Thanetian–Ypresian African fossil record limits these conclusions. TheMiddle Paleocene age of Monshyus does not exclude anotherpaleobiogeographic scenario and the existence of a European‘‘condylarth’’ lineage from which elephant-shrews originated.In the same way, most of the analyzed characters of Microhyusare those of M. reisi from the Earliest Eocene of Silveirinha(Portugal). Thus, this Portuguese species would be morpholog-ically close to the European ‘‘condylarth’’ lineage that migratedto Africa. Such a wave of migration during the Early Eocene,via the Iberian plate (?), has already been suggested for theTunisian fauna from Chambi (Hartenberger et al., 1985).
During this period, the Proboscidea are already recorded inAfrica and they are well-individualized (Gheerbrant et al.,1998). Then if Macroscelidea originated from a European Eo-cene ‘‘condylarth,’’ it seems that tethytherians and elephant-shrews are paraphyletic. Therefore, we have to admit severalwaves of migration, and it is likely that the migration of formsat the origin of the Proboscidea is earlier than the migration offorms from which Macroscelidea originated.
Thus, the polarity of these migrations are not well-knownyet. They must have concerned the whole circumference of theTethys which, during the early Tertiary, extended from the In-
dian sub-continent in the East and to the Iberian plate in theWest. Nevertheless, the role of Africa must not be neglectedwith regard to the origin of modern orders of mammals. If en-demism is well-established during the Oligocene, the knowl-edge of the African fossil mammals from the Paleocene–Eocenetimes is now capital for both paleobiogeographic and phylo-genetic reconstructions.
ACKNOWLEDGMENTS
We thank especially J. Sudre for general discussion aboutearly ungulates and the systematic part of this paper; P. M.Butler, P. A. Holroyd, J. J. Hooker, and F. Senegas for providingus with casts of Macroscelididae, and E. Gheerbrant gave ac-cess to comparative material of Louisininae. We are grateful toS. Ducrocq and B. Marandat for reading a first version of thepaper and for their useful comments. This is ISEM contributionnumber 2001-018.
LITERATURE CITED
Antunes, M. T., C. Estravis, and D. E. Russell. 1987. A new condylarth(Mammalia) from the Early Eocene of Silveirinha, Portugal.Munchner Geowissenschaftliche Abhandlungen A(10):219–224.
Archibald, J. D. 1998. Archaic ungulates (‘‘Condylarthra’’); pp. 292–329 in C. M. Janis, K. M. Scott, and L. L. Jacobs (eds.), Evolutionof Tertiary Mammals of North America. Cambridge UniversityPress, Cambridge.
Bonaparte, C. L. 1838. Synopsis vertebratorum systematis. Nuovi An-nali delle Scienze Naturali 1:105–133.
Bown, T. M., and E. L. Simons. 1987. New Oligocene Ptolemaiidae(Mammalia:? Pantolesta) from the Jebel Qatrani Formation, FayumDepression, Egypt. Journal of Vertebrate Paleontology 7:311–324.
Butler, P. M. 1956. The skull of Ictops and the classification of theInsectivora. Proceedings of the Zoological Society of London 126:453–481.
——— 1984. Macroscelidea, Insectivora and Chiroptera from the Mio-cene of East Africa. Palaeovertebrata 14:117–198.
——— 1995. Fossil Macroscelidea. Mammal Review 25:3–14.Coiffait, P.-E., B. Coiffait, J.-J. Jaeger, and M. Mahboubi. 1984. Un
nouveau gisement a mammiferes fossiles d’age Eocene superieursur le versant sud des Nementchas (Algerie orientale): decouvertedes plus anciens rongeurs d’Afrique. Comptes Rendus del’Academie des Sciences 299:893–898.
Collinson, M. E., and J. J. Hooker. 1987. Vegetational and mammalianfaunal changes in the early Tertiary of southern England; pp. 259–303 in M. E. Collinson and J. J. Hooker (eds.), Origins of Angio-sperms and Their Biological Consequences. Cambridge UniversityPress, Cambridge.
de Jong, W. W., J. A. M. Leunissen, and G. J. Wistow. 1993. Eye lenscrystallins and the phylogeny of placental orders: evidence for aMacroscelid–Paenungulate clade?; pp. 5–12 in F. S. Szalay, M. J.Novacek, and M. C. McKenna (eds.), Mammal Phylogeny. Spring-er Verlag, New York.
Denys, C., and D. E. Russell. 1981. Etude de la variabilite dentaired’une population de Paschatherium (condylarthre hyopsodontide),provenant de la localite sparnacienne de Dormaal (Belgique). Bul-letin d’information sur la Geologie du Bassin de Paris 18:37–45.
Estravis, C. 1992. Estudo dos mameferos do Eocenico inferior de Sil-veirinha (Baixo Mondego), Ph.D. dissertation, Universidade Novade Lisboa, Lisboa, 248 pp.
———, and D. E. Russell. 1992. Paschatherium marianae, un nouveauCondylarthra de Silveirinha, Eocene inferieur du Portugal. Bulletindu Museum National d’Histoire Naturelle 14:185–203.
Faid, N. 1999. Ostracodes de l’Eocene inferieur-moyen de l’Atlas Sa-harien (Algerie): interpretation environnementale et paleogeograph-ique. Geobios 32:459–481.
Fischer, M. S. 1996. On the position of Proboscidea in the phylogeneticsystem of Eutheria: a systematic review; pp. 35–38 in J. Shoshaniand P. Tassy (eds.), The Proboscidea: Evolution, Palaeoecology ofElephants and Their Relatives. Oxford University Press, Oxford.
———, and P. Tassy. 1993. The interrelation between Proboscidea, Sir-enia, Hyracoidea, and Mesaxonia: the morphological evidence; pp.

544 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 21, NO. 3, 2001
217–234 in F. S. Szalay, N. M. J. McKenna, and M. C. McKenna(eds.), Mammal Phylogeny. Springer Verlag, New York.
Gheerbrant, E. 1987. Les vertebres continentaux de l’Adrar Mgorn(Maroc, Paleocene); une dispersion de mammiferes transtethysien-ne aux environs de la limite mesozoıque/cenozoıque? GeodinamicaActa 1:233–246.
——— 1990. On the early biogeographical history of the African pla-centals. Historical Biology 4:107–116.
——— 1995. Les mammiferes paleocenes du Bassin d’Ouarzazate(Maroc). III Adapisoriculidae et autres mammiferes (Carnivora,?Creodonta, Condylarthra,? Ungulata et incertae sedis). Palaeonto-graphica 237:39–132.
———, J. Sudre, and H. Cappetta. 1996. A Palaeocene proboscideanfrom Morocco. Nature 383:68–70.
———, ———, ———, and G. Bignot. 1998. Phosphatherium es-cuilliei du Thanetien du Bassin des Ouled Abdoun (Maroc), plusancien proboscidien (Mammalia) d’Afrique. Geobios 30:247–269.
Godinot, M., T. Smith, and R. Smith. 1996. Mode de vie et affinites dePaschatherium (Condylarthra, Hyopsodontidae) d’apres ses os dutarse. Palaeovertebrata 25:225–242.
Graur, D., M. Gouy, and L. Duret. 1997. Evolutionary affinities of theorder Perissodactyla and the phylogenetic status of the superordinaltaxa Ungulata and Altungulata. Molecular Phylogenetics and Evo-lution 7:195–200.
Haeckel, E. 1866. Generelle Morphologie der Organismen. Georg Rei-mer, Berlin, 462 pp.
Hartenberger, J.-L. 1986. Hypothese paleontologique sur l’origine desMacroscelidea (Mammalia). Comptes Rendus de l’Academie desSciences 302:247–249.
———, J.-Y. Crochet, C. Martinez, M. Feist, M. Godinot, B. MannaiTayech, B. Marandat, and B. Sige. 1997. Le gisement de mammi-feres de Chambi (Eocene, Tunisie centrale) dans son contexe geo-logique. Apport a la connaissance de l’evolution des mammiferesen Afrique; pp. 263–274 in J.-P. Aguilar, S. Legendre, and J. Mi-chaux (eds.), Actes du congres BiochroM’97.
———, C. Martinez, and A. Ben Saıd. 1985. Decouverte de mammi-feres d’age Eocene inferieur en Tunisie centrale. Comptes Rendusde l’Academie des Sciences 301:649–652.
Holroyd, P. A. 1994. An examination of dispersal origins for Fayummammals, Ph.D. dissertation, Duke University, Durham, 328 pp.
Jaeger, J.-J., C. Denys, and B. Coiffait. 1985. New Phiomorpha andAnomaluridae from the Late Eocene of north-west Africa: phylo-genetic implications; pp. 567–588 in W. P. Luckett and J.-L. Har-tenberger (eds.), Evolutionary Relationships among Rodents, aMultidisciplinary Analysis. Plenum Press, London.
Janis, C. M., J. D. Archibald, R. L. Cifelli, S. G. Lucas, C. R. Schaff,R. M. Schoch, and T. E. Williamson. 1998. Archaic ungulates andungulatelike mammals; pp. 247–259 in C. M. Janis, K. M. Scott,and L. L. Jacobs (eds.), Evolution of Tertiary Mammals of NorthAmerica. Cambridge University Press, Cambridge.
Luckett, W. P., and J.-L. Hartenberger (eds.). 1985. Evolutionary Re-lationships among Rodents, a Multidisciplinary Analysis. PlenumPress, London, 721 pp.
Mahboubi, M., F. Mebrouk, and J.-J. Jaeger. 1997. Consequences pa-leobiogeographiques tirees a partir de l’etude de quelques gise-ments paleogenes du Maghreb (Mammiferes, Gasteropodes, Char-ophytes); pp. 275–284 in J.-P. Aguilar, S. Legendre, and J. Michaux(eds.), Actes du congres BiochroM’97.
McKenna, M. C. 1975. Toward a phylogenetic classification of theMammalia; pp. 21–46 in W. P. Luckett and F. S. Szalay (eds.), APhylogeny of the Primates. A Multidisciplinary Approach. PlenumPress, New York.
———, and S. K. Bell. 1997. Classification of Mammals above theSpecies Level. New York, 631 pp.
Nessov, L. A., J. D. Archibald, and Z. Kielan-Jaworowska. 1998. Un-gulate-like mammals from the Late Cretaceous of Uzbekistan anda phylogenetic analysis of Ungulatomorpha. Bulletin of CarnegieMuseum of Natural History 34:40–88.
Novacek, M. J. 1984. Evolutionary stasis in the elephant-shrew, Rhyn-chocyon; pp. 4–31 in N. Eldredge and S. M. Stanley (eds.), LivingFossils. Springer Verlag, New York.
———, A. R. Wyss, and M. C. McKenna. 1988. The major groups ofeutherian mammals; pp. 31–71 in M. J. Benton (ed.), The Phylog-eny and Classification of the Tetrapods. Clarendon Press, Oxford.
Patterson, B. 1965. The fossil elephant-shrews (family Macroscelidi-
dae). Bulletin of the Museum of Comparative Zoology 133:295–335.
Prothero, D. R., E. M. Manning, and M. S. Fischer. 1988. The phylog-eny of the ungulates; pp. 201–234 in M. J. Benton (ed.), The Phy-logeny and Classification of the Tetrapods. Clarendon Press, Ox-ford.
Rose, K. D. 1999. Postcranial skeleton of Eocene Leptictidae (Mam-malia), and its implications for behavior and relationships. Journalof Vertebrate Paleontology 19:355–372.
Russell, D. E. 1964. Les mammiferes paleocenes d’Europe. Memoiredu Museum National d’Histoire Naturelle C(8):1–324.
Sanchez-Villagra, M. R., and B. A. Williams. 1998. Levels of homo-plasy in the evolution of the mammalian skeleton. Journal of Mam-malian Evolution 5:113–126.
Schlosser, M. 1910. Uber einige fossile Saugetiere aus dem Oligocanvon Agypten. Zoologische Anzeige 35:500–508.
Shoshani, J., and M. C. McKenna. 1998. Higher taxonomic relation-ships among extant mammals based on morphology, with selectedcomparisons of results from molecular data. Molecular Phyloge-netics and Evolution 9:572–584.
Sige, B. 1991. Rhinolophoidea et Vespertilionoidea (Chiroptera) deChambi (Eocene inferieur de Tunisie). Aspects biostratigraphique,biogeographique et paleoecologique de l’origine des chiropteresmodernes. Neues Jahrbuch fur Geologie und Palaontologie 182:355–376.
———, J.-J. Jaeger, J. Sudre, and M. Vianey-Liaud. 1990. Altiatlasiuskoulchii n. gen. et sp., primate omomyide du Paleocene superieurdu Maroc, et les origines des euprimates. Palaeontographica 214:31–56.
Simons, E. L., P. A. Holroyd, and T. M. Bown. 1991. Early Tertiaryelephant-shrews from Egypt and the origin of the Macroscelidea.Proceedings of the National Academy of Sciences USA 88:9734–9737.
Springer, M. S., A. Burk, J. R. Kavanagh, V. G. Waddell, and M. J.Stanhope. 1997. The interphotoreceptor retinoid binding proteingene in therian mammals: implications for higher level relation-ships and evidence for loss of function in the marsupial mole. Pro-ceedings of the National Academy of Sciences USA 94:13754–13759.
Stanhope, M. J., V. G. Waddell, O. Madsen, W. W. de Jong, S. B.Hedges, G. C. Clevens, D. Kao, and M. S. Springer. 1998a. Mo-lecular evidence for multiple origins of Insectivora and for a neworder of endemic African insectivore mammals. Proceedings of theNational Academy of Sciences USA 95:9967–9972.
———, O. Madsen, V. G. Waddell, G. C. Clevens, W. W. de Jong, andM. S. Springer. 1998b. Highly congruent molecular support for adiverse superordinal clade of endemic African mammals. Molecu-lar Phylogenetics and Evolution 9:501–508.
Sudre, J. 1975. Un prosimien du Paleogene ancien du Sahara nord-occidental: Azibius trerki n. g. n. sp. Comptes Rendus del’Academie des Sciences 280:1539–1542.
——— 1979. Nouveaux mammiferes eocenes du Sahara occidental.Palaeovertebrata 9:83–115.
———, J.-J. Jaeger, B. Sige, and M. Vianey-Liaud. 1993. Nouvellesdonnees sur les condylarthres du Thanetien et de l’Ypresien duBassin d’Ouarzazate (Maroc). Geobios 26:609–615.
———, and D. E. Russell. 1982. Les mammiferes montiens de Hainin(Paleocene moyen de Belgique). Part II: les condylarthres. Palaeo-vertebrata 12:173–184.
Szalay, F. S. 1975. Phylogeny, Adaptations, and dispersal of the tarsi-iform primates; pp. 357–404 in W. P. Luckett and F. S. Szalay(eds.), A Phylogeny of the Primates. A Multidisciplinary Approach.Plenum Press, New York.
Tabuce, R., B. Coiffait, P.-E. Coiffait, M. Mahboubi, and J.-J. Jaeger.2000. A new species of Bunohyrax (Hyracoidea, Mammalia) fromthe Eocene of Bir El Ater (Algeria). Comptes Rendus del’Academie des Sciences 331:61–66.
Teilhard de Chardin, P. 1927. Les mammiferes de l’Eocene inferieur dela Belgique. Memoires du Musee Royal d’Histoire Naturelle deBelgique 36:1–26.
Thewissen, J. G. M., and D. P. Domning. 1992. The role of phenaco-dontids in the origin of the modern orders of ungulate mammals.Journal of Vertebrate Paleontology 12:494–504.
Van Valen, L. 1967. New Paleocene insectivores and insectivore clas-

545TABUCE ET AL.—NEW EOCENE MACROSCELID FROM ALGERIA
sification. Bulletin of the American Museum of Natural History135:217–284.
——— 1978. The beginning of the age of mammals. Evolutionary The-ory 4:45–80.
Vianey-Liaud, M., J.-J. Jaeger, J.-L. Hartenberger, and M. Mahboubi.1994. Les rongeurs de l’Eocene d’Afrique nord-occidentale—GlibZegdou (Algerie) et Chambi (Tunisie)—et l’origine des Anomal-uridae. Palaeovertebrata 23:93–118.
Waddell, P. J., N. Okada, and M. Hasegawa. 1999. Towards resolvingthe interordinal relationships of placental mammals. Systematic Bi-ology 48:1–5.
Received 1 November 1999; accepted 15 February 2001.
APPENDIX 1
List of characters used in the phylogenetic analysis.
1. Enamel surface: unwrinkled (0); wrinkled (1).2. p4 much less wide than m1 (0); not inflated or less inflated than
m1 (1); swollen (2).3. p4 shorter than m1 (0); sub-equal or longer (1).4. Trigonid of p4 more developed than talonid (0); talonid quite de-
veloped (1); talonid developed like that of a molar (2).5. Trigonid basin of p4 absent or reduced (0); well individualized (1).6. Paraconid on p4 absent or well reduced (0); anteriorly projected
(1); lingual (2).7. Metaconid on p4 absent (0); small (1); enlarged (2).8. Metaconid on p4 distally placed than protoconid (0); directly op-
posed (1).9. Trigonid on m1–2 shorter than talonid (0); sub-equal (1); longer
(2).10. Trigonid on m1–2 much higher than talonid (0); higher (1); sub-
equal (2).11. Paraconid on m1–2 well developed (0); crestiform or absent (1).12. Paracristid on m1–2 absent or short (0); strong (1).13. Protoconid on m1–2 directly opposed from metaconid (0); anterior
(1); strongly anterior (2).14. Metaconid on m1–2 at least as wide as the protoconid (0); less
wide than the protoconid (1).15. Cristid obliqua on m1–2 ending at the base of the posterior trigonid
wall (0); cristid obliqua ending on the trigonid wall (1); connectedto the metaconid (2).
16. Hypoconulid on m1–2 cusped (0); crestiform (1); forming a stronghypocristid (2).
17. m2 sub-equal to m1 (0); larger (1); narrower (2).18. m3 longer or equal to m2 (0); reduced (1); vestigial (2); absent (3).19. Hypoconulid on m3 cusped (0); lobed (1); absent(2).20. Talonid on m3 longer than trigonid (0); sub-equal (1); shorter (2).21. Trigonid on m3 as large as talonid (0); larger (1); more larger (2).22. P3 not premolariform (0); premolariform (1); molariform (2).23. Metacone on P4 reduced but cusped (0); absent (1); well individ-
ualized (2).24. Metaconule on P4 absent (0); reduced (1); strong (2); associated
with a loph (3).25. Paraconule on P4 strong (0); reduced (1); absent (2); associated
with a loph (3).26. Parastyle on P4 anteriorly projected (0); less projected (1); small
or not prominent (2).27. Protocone on P4 high (0); reduced (1).28. Postprotocrista on P4 long and buccal (0); short (1); distal and
connected with the cingulum (2); absent (3).29. Hypocone on P4 absent or reduced (0); small (1); strong (2).30. Ectoflexus on M2 deep (0); shallow (1); absent (2).31. Stylar shelf on M2 extended (0); reduced (1); absent (2).32. Buccal expansion under the paracone on M2 absent (0); present (1).33. Cuspules of the parastylar lobe on M1–2 present (0); absent (1).34. Mesostyle on M1–2 absent (0); connected with the centrocrista (1);
isolated (2).35. Paracone on M1 not higher than the metacone (0); higher (1).36. Premetaconule crista on M1–2 present (0); absent (1).37. Paraconule on M1–2 present (0); absent (1); included in the pro-
tocone (2).38. Protocone on M1–2 situated between the paracone and the meta-
cone (0); aligned with the paracone (1).39. Lingual side of the protocone on M1–2 greatly pronounced (0);
weak (1).40. protoloph on M1–2 absent (0); short (1); long (2); forming a strong
crest (3).41. Postprotocrista on M1–2 present (0); absent (1); short and oriented
towards the hypocone (2).42. Hypocone on M1–2 absent or incipient (0); reduced (1); sub-equal
or higher than the protocone (2).43. Hypoloph on M1–2 absent (0); present (1).44. Lingual cingulum on M1–2 absent (0); present (1).

546 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 21, NO. 3, 2001
AP
PE
ND
IX2
Dis
trib
utio
nof
44de
ntal
char
acte
rssu
bjec
ted
toPA
UP
anal
ysis
.C
hara
cter
sar
eli
sted
num
eric
ally
toco
rres
pond
toA
ppen
dix
1.‘‘
0’’
repr
esen
tsth
epr
imit
ive
oran
cest
ral
stat
e,‘‘
1’’
to‘‘
3’’
repr
esen
tde
rive
dch
arac
ters
,‘‘
?’’
indi
cate
sth
atth
ech
arac
ter
isun
obse
rved
,‘‘
-’’
indi
cate
sth
atth
ech
arac
ter
isno
tap
plic
able
.T
hepo
lym
orph
ism
isre
pres
ente
dby
A(0
/1),
B(1
/2),
C(1
/3),
D(0
/1/
2),
E(0
/2).
Tax
a
Cha
ract
er
1–10
11–20
21–30
31–40
41–44
Asp
anle
stes
00
00
00
10
00
00
01
00
01
00
0?
00
00
00
00
00
00
00
00
00
00
00
Eou
ngul
atum
00
??
??
??
00
00
00
00
?0
00
0?
0?
?1
00
02
10
00
?0
00
00
01
00
Pal
aech
thon
00
00
00
0-
10
00
00
02
00
10
00
20
01
03
00
20
10
00
00
00
00
00
Pro
tung
ulat
um0
00
00
21
0B
00
01
10
01
01
00
01
01
00
00
01
01
00
10
00
00
10
0L
oxol
ophu
s1
10
00
21
01
10
01
00
01
01
00
01
02
00
00
11
01
00
00
00
00
10
1H
yops
odus
00
00
02
10
12
10
11
B0
1A
10
0A
10
22
00
02
21
10
01
0A
02
B1
00
Hap
lale
tes
01
00
01
10
11
10
00
00
1A
AA
11
10
22
00
01
20
10
00
00
00
01
00
Lit
omyl
us0
10
00
00
-1
11
01
00
00
10
10
01
02
00
00
12
01
00
00
00
00
10
0M
iocl
aenu
s1
20
00
00
-2
11
01
00
01
1A
01
11
02
20
30
22
11
20
10
00
01
00
0P
erip
tych
us1
20
00
21
12
20
01
11
00
01
00
11
20
21
30
22
11
00
10
00
01
10
0E
ocon
odon
00
00
00
11
21
10
A0
10
10
10
0?
10
20
00
01
20
10
01
00
00
01
0A
Dia
code
xis
01
10
01
0-
A1
00
10
00
10
10
00
10
2A
03
01
20
10
00
00
00
00
00
Phe
naco
dus
01
01
AA
2A
A1
10
00
A0
A0
A0
A0
2A
B2
10
02
21
11
A1
0A
00
A1
01
Ect
ocio
n0
10
1A
A2
AA
11
0A
0A
0A
0A
0A
02
DA
21
00
22
11
1A
A0
A0
0A
10
AH
yrac
othe
rium
01
01
A0
21
01
10
A0
A1
00
10
01
20
0B
01
02
21
10
01
01
02
12
11
Bun
ohyr
ax0
10
1A
12
11
11
00
10
10
01
01
12
02
10
20
12
01
10
-1
10
12
21
1N
umid
othe
rium
01
10
00
20
11
10
00
01
10
00
01
10
20
03
01
21
10
01
11
03
12
10
Dip
aval
i0
11
10
01
0B
B1
00
10
00
10
11
??
??
??
??
22
11
01
10
10
22
10
0H
aplo
myl
us0
A1
A0
11
0A
1A
01
00
01
11
00
?0
0B
01
10
22
01
00
00
00
02
10
0L
ouis
ina
01
0A
00
10
21
10
01
00
11
0B
20
00
22
11
02
21
10
10
01
10
01
00
Mic
rohy
us0
11
00
0D
-1
11
0A
00
01
12
12
12
20
21
11
22
11
00
1E
11
A1
2A
0P
asch
athe
rium
01
A0
00
21
11
A0
0A
00
A1
01
10
01
10
10
0B
B0
10
1A
0A
0A
01
00
Cha
mbi
us0
10
00
11
01
20
01
10
1?
22
22
?2
11
11
10
22
11
00
10
11
10
21
0H
erod
otiu
s0
11
11
12
11
21
00
10
10
22
22
?2
02
11
11
22
11
0?
-2
11
11
21
0N
emen
tcha
ther
ium
01
A1
01
20
11
10
01
01
?2
22
2?
20
22
12
0?
??
?0
?-
21
11
12
10
Met
oldo
bote
s0
11
21
12
01
21
00
01
12
3-
--
??
??
??
??
22
11
0?
12
11
31
20
0M
iorh
ynch
ocyo
n0
11
21
12
01
10
12
10
22
3-
--
02
02
20
12
12
-1
00
-2
10
30
20
0M
acro
scel
idin
ae0
10
21
12
01
10
12
02
22
22
22
22
33
20
12
12
11
00
10
10
30
21
0M
yohy
rax
01
02
11
20
11
01
20
22
02
22
22
23
31
03
21
21
10
01
01
03
12
10
Copyright © 2022 FDOKUMEN




















