
A comparative study of predator-induced phenotype in tadpoles across a pond permanency gradient
14
PRIMARY RESEARCH PAPER A comparative study of predator-induced phenotype in tadpoles across a pond permanency gradient Alex Richter-Boix Gustavo A. Llorente Albert Montori Received: 20 April 2006 / Revised: 15 September 2006 / Accepted: 14 October 2006 / Published online: 26 February 2007 Ó Springer Science+Business Media B.V. 2007 Abstract In a field survey the distribution of pond-breeding anuran species and their potential large predators was investigated along a freshwa- ter habitat gradient, ranging from ephemeral pools to permanent ponds. In a laboratory experiment predator-induced plasticity was examined for all tadpole species to test whether the plastic response of ephemeral and temporary pond spe- cies differs from that of permanent pond species. Desiccation and predation pose conflicting de- mands; reduced activity lowers the risk of death by predation but increases the risk of death by desiccation. It was expected that species from time-constrained habitats would display a mor- photype that would reduce vulnerability to inver- tebrate predators, thus allowing these species to maintain a high level of activity, whereas species from permanent ponds would avoid predation both morphologically and behaviourally. Species distribution and predator composition along the hydroperiod gradient differed. Variations be- tween ephemeral and temporary ponds can be attributed to hydroperiod differences and the presence of large invertebrate predators in tem- porary ponds, whereas the contrasts between temporary and permanent ponds can only be attributed to the hydroperiod, since the presence and abundance of top predators are similar in both habitat types. With the exception of bufonids, all species showed predator-induced plasticity in agreement with previous studies. Tadpole species differed in the integration of the phenotypic traits measured, but differences observed between species could not be attributed only to habitat. Species from temporary habitats showed an expected response, with a low reduction of activity in comparison with the rest of the species. The lack of general patterns in the morphological changes suggests that species within the same habitat type did not converge on similar phenotypes, perhaps due to functional constraints on differences in microhabitat use in the water column. Keywords Behavioural plasticity Á Mediterranean area Á Morphological plasticity Á Phenotypic integration Á Predation risk gradient Á Spatial distribution Á Temporary pond Á Trade-offs Introduction Models of community structure in lentic systems suggest that species composition is determined by Handling editor: K. Martens A. Richter-Boix (&) Á G. A. Llorente Á A. Montori Departament Biologia Animal, Facultat de Biologia, Universitat de Barcelona, Av. Diagonal 645, 08028 Barcelona, Spain e-mail: [email protected] 123 Hydrobiologia (2007) 583:43–56 DOI 10.1007/s10750-006-0475-7
Transcript of A comparative study of predator-induced phenotype in tadpoles across a pond permanency gradient

PRIMARY RESEARCH PAPER
A comparative study of predator-induced phenotypein tadpoles across a pond permanency gradient
Alex Richter-Boix Æ Gustavo A. Llorente ÆAlbert Montori
Received: 20 April 2006 / Revised: 15 September 2006 / Accepted: 14 October 2006 / Published online: 26 February 2007� Springer Science+Business Media B.V. 2007
Abstract In a field survey the distribution of
pond-breeding anuran species and their potential
large predators was investigated along a freshwa-
ter habitat gradient, ranging from ephemeral pools
to permanent ponds. In a laboratory experiment
predator-induced plasticity was examined for all
tadpole species to test whether the plastic
response of ephemeral and temporary pond spe-
cies differs from that of permanent pond species.
Desiccation and predation pose conflicting de-
mands; reduced activity lowers the risk of death by
predation but increases the risk of death by
desiccation. It was expected that species from
time-constrained habitats would display a mor-
photype that would reduce vulnerability to inver-
tebrate predators, thus allowing these species to
maintain a high level of activity, whereas species
from permanent ponds would avoid predation
both morphologically and behaviourally. Species
distribution and predator composition along the
hydroperiod gradient differed. Variations be-
tween ephemeral and temporary ponds can be
attributed to hydroperiod differences and the
presence of large invertebrate predators in tem-
porary ponds, whereas the contrasts between
temporary and permanent ponds can only be
attributed to the hydroperiod, since the presence
and abundance of top predators are similar in both
habitat types. With the exception of bufonids, all
species showed predator-induced plasticity in
agreement with previous studies. Tadpole species
differed in the integration of the phenotypic traits
measured, but differences observed between
species could not be attributed only to habitat.
Species from temporary habitats showed an
expected response, with a low reduction of activity
in comparison with the rest of the species. The lack
of general patterns in the morphological changes
suggests that species within the same habitat type
did not converge on similar phenotypes, perhaps
due to functional constraints on differences in
microhabitat use in the water column.
Keywords Behavioural plasticity �Mediterranean area � Morphological
plasticity � Phenotypic integration � Predation
risk gradient � Spatial distribution � Temporary
pond � Trade-offs
Introduction
Models of community structure in lentic systems
suggest that species composition is determined by
Handling editor: K. Martens
A. Richter-Boix (&) � G. A. Llorente �A. MontoriDepartament Biologia Animal, Facultat de Biologia,Universitat de Barcelona, Av. Diagonal 645, 08028Barcelona, Spaine-mail: [email protected]
123
Hydrobiologia (2007) 583:43–56
DOI 10.1007/s10750-006-0475-7

a trade-off between pond permanency and pre-
dation risk (Schneider & Frost, 1996; Wellborn
et al., 1996). Changes in predators are related to
pond permanency (Woodward, 1983; Werner &
McPeek, 1994; Gunzburger & Travis, 2004). This
model is particularly applicable to amphibian
assemblages. These two antagonistic gradients
create different selective forces for tadpole spe-
cies in ponds from different positions along the
gradient (Wellborn et al., 1996; Richardson,
2001a, b). Resource acquisition in tadpoles may
be affected by both factors: the need for timely
development in temporary ponds and the need to
avoid predators along the hydroperiod gradient.
Activity levels and foraging rates are typically
positively correlated, thus increased activity leads
to an increased predation risk (Skelly, 1994;
Anholt & Werner, 1995; Eklov & Halvarsson,
2000). Activity level is therefore expected to
reflect the differences in habitat type along the
pond permanency gradient (Woodward, 1983;
Skelly, 1996; Wellborn et al., 1996; Richardson,
2001b).
Several mesocosm and laboratory studies have
also demonstrated that tadpole species have
evolved the ability to asses predation risk and
to react in a way that reduces the likelihood of
being preyed upon (Skelly & Werner, 1990;
Chovanec, 1992; Anholt et al., 1996; McCollum
& Leimberger, 1997; Lardner, 2000; Van Buskirk,
2002; Relyea, 2004). Typical phenotypic plasticity
in the presence of predators includes a reduction
in time spent foraging and changes in morphol-
ogy which make individuals less vulnerable to
predators. All of these changes affect growth and
development, causing individuals to grow and
develop more slowly when exposed to predators
than in the absence of predators (Lardner, 2000;
Relyea, 2002; Van Buskirk, 2002). However,
species that persist in temporary ponds com-
monly face time constraints that limit their ability
to delay growth and development (Altwegg,
2002).
Due to the behavioural trade-offs that exist
along the pond permanence gradient, it can be
expected that levels of phenotypic plasticity in
response to predation pressure––like activity
levels––will differ among species evolving under
different selective regimes, with varying integra-
tion of the different traits (behavioural and
morphological) to fine-tune the plastic phenotype
response depending on species ecology and his-
tory (Relyea, 2004). Species from ephemeral and
temporary ponds can be expected to exhibit a
predominantly morphological, predator-induced
plasticity in order to allow the species to maintain
high levels of activity and to develop more rapidly
whilst also reducing predation risk (Relyea &
Werner, 1999; Anholt et al., 2000). In most
permanent ponds, morphological changes in spe-
cies can be accompanied by behavioural changes
in order to reduce encounters with and detection
by predators (Chovanec, 1992; Anholt et al.,
2000).
To date, only three studies have tested the
distribution and phenotypic plasticity of more
than two closely related species (Relyea &
Werner, 2000; Richardson, 2001b; Van Buskirk,
2002). The study made by Relyea & Werner
(2000) focuses only on the morphological changes
of four species along a predation risk gradient and
does not incorporate measures of behavioural
activity. In contrast, Richardson (2001b) studies
only activity levels and their plasticity without
considering morphological plasticity, whereas
Van Buskirk (2002) studies both plastic traits
but from an adaptive point of view of phenotypic
plasticity that gives no consideration to the trade-
offs that species face along the pond permanency
gradient. None of these studies address how
behavioural and morphological traits are inte-
grated along the hydroperiod gradient. The pres-
ent study evaluates the links between activity
behaviour, predator phenotypic plasticity and
habitat use of species along the pond permanency
gradient and, ultimately, explains distribution
patterns. Consequently, the distributions of six
tadpole species are compared in a field survey
with regard to what are commonly considered the
two principle sources of mortality: pond perma-
nency and predation risk. This is followed by a
laboratory experimental procedure to assess
predator-induced plasticity (morphology and
activity level) and its effects on development.
44 Hydrobiologia (2007) 583:43–56
123

Materials and methods
Site
The field study was carried out in protected
natural areas near Barcelona (NE Iberian Penin-
sula), including the Natural Park of Garraf and
the Metropolitan Park of Collserola. The climate
in this zone is Mediterranean, with hot, dry
summers, mild winters and two rainy periods,
one in spring and the other in autumn. The
amount of precipitation varies considerably from
year to year in this region (see Richter-Boix et al.,
2006b for a well description and localization of
the areas of study). The amphibian community in
this zone comprises seven anuran species (Alytes
obstetricans (Laurenti 1768), Pelobates cultripes
(Cuvier 1829), Pelodytes punctatus (Daudin
1802), Bufo calamita (Laurenti 1768), Bufo bufo
(Linne 1758), Hyla meridionalis (Boettger 1874)
and Rana perezi (Seoane 1885)) and Salamandra
salamandra (Linne 1758). We focused our study
on the anuran community but excluded Pelobates
because it is a rare species in the area and few
data are available.
Field sampling methods
We evaluated amphibian larvae and their poten-
tial invertebrate predators in four sampling peri-
ods during the spring and summer, from March to
August of 2002 in a total of 193 isolate ponds.
These localities span the range of aquatic habitats
in which the species studied breed in the area of
study, including ephemeral pools, temporary and
permanent ponds. Sampling time periods were
dictated by preliminary sampling and accounted
for temporal differences in breeding activity
among species (Richter-Boix et al., 2006b) and
ensured that all species breeding were captured.
Due to variation in hydroperiod not all sites could
be surveyed in all sampling periods, thus sample
sizes of ponds were not uniform. Both, amphibian
larvae and predaceous invertebrates were sam-
pled with dip-net sweeps (30 cm · 40 cm) to
obtain relative species densities. This is a stan-
dardized technique used to sample both groups
(e.g. Schaffer et al., 1994; Babbitt et al., 2003). A
minimum of 5–10 dip-net sweeps were taken in
each possible tadpole microhabitat following
standard techniques determined by pond size
(Schaffer et al., 1994). All tadpoles were identi-
fied in the field and photographed with a grid
background; the number of individuals of each
species were counted and finally returned to
water. Predaceous invertebrates shown in previ-
ous studies (e.g. Woodward, 1983; Travis et al.,
1985; Cronin & Travis, 1986) to prey on tadpoles
were identified, counted and photographed with a
grid background. Three types of insects were
considered as potential predators: Odonata larvae
(considering as predators aeshnid and libellulid
naiads), Heteroptera (notonectids and Nepa spp.)
and Coleoptera (larvae and adults diving beetles).
Fish presence was determined through visual
surveys in addition to dip-net captures. Indepen-
dently, for tadpole surveys, we visited ponds
every month throughout the year to establish the
data of drying and determine the position of the
pond across the hydroperiod gradient. In this
manner, we were able to evaluate the number of
days (in 30 · 30 days) that ponds retain water.
We defined hydroperiods in terms of three
categories: (1) ephemeral pools, (2) temporary
ponds and (3) permanent ponds. Ephemeral pools
(ponds containing surface water for a maximum
of two months) refilled after each rainfall period.
Temporary ponds were flooded by spring and
autumn rainfall. The shallowest temporary ponds
often dried out in winter, whereas the deepest
ponds held water until the end of spring or early
summer. It is important to consider that a pond’s
position on the gradient of the hydroperiod could
be dynamic, and especially that some temporary
ponds change position along this gradient annu-
ally as a result of climatic conditions. We did not
include a category for permanent ponds with or
without fish as in previous studies (Babbitt et al.,
2003; Stocks & McPeek, 2003; Van Buskirk,
2003), because in the study area only 6 of the
193 ponds contained fish, and these were not
included in the study, leaving 187 localities for
analysis.
Total counts for each amphibian species and
predaceous invertebrate captured in each pond
were divided by the number of dip-net sweeps
taken in each pond (Babbitt et al., 2003). This
yielded an abundance based on catch per unit
Hydrobiologia (2007) 583:43–56 45
123

effort which could be compared across the three
different habitats considered. Body lengths of
predaceous invertebrates were measured from
pictures on a computer and divided into two
groups: (1) small invertebrates from 5 to 15 mm
long, and (2) large invertebrate predators with a
body length of over 15 mm. We conducted
analyses of variance ANOVAs to determine if
amphibian species and predator group abun-
dances and relative densities varied among hy-
droperiod categories. Ponds were considered as
the units in all analyses and variables log-trans-
formed before analyses. Predation risk at which
each amphibian species was exposed was com-
pared with analysis of variance. Also, larval
amphibian species abundance was correlated with
invertebrate predator abundance, and a partial
set correlation analysis was performed to remove
the effect of hydroperiod.
Experimental procedure: effect of predation
risk on larval morphology and activity
Morphological and behavioural anti-predator
responses of the six species to a common predator
were tested in a short-term laboratory experi-
ment. Clutches of the six species were collected
from natural ponds. In all cases egg masses were
taken during species amplexus period to reduce
the time of exposition of embryos to predators to
avoid phenotypic plasticity induction on hatch-
ings. For Pelodytes egg samples were taken from 5
egg masses and hatched separately. For Hyla we
recollected 15 egg masses from three different
ponds. In the case of bufonids, fragments of three
different egg strings were taken for each species
(B. bufo and B. calamita). In the case of Rana,
tadpoles were obtained from 5 clutches from two
different ponds. The Alytes larvae had recently
hatched and just been deposited in the pond by
males when we recollected it. As males of
Alytes have parental care of eggs, negligible
predation on hatchings should take place during
the egg period and no defences should have been
developed.
The experiment was conducted in laboratory
tanks (30 l) filled with dechlorinated water under
approximately the same conditions as the natural
photoperiod and at a temperature of around
22�C, during the spring of 2001. Thirty tadpoles
were randomly drawn from a mixture of all
clutches and transferred from hatching aquariums
to experimental tanks when they reached Gosner
25 life stage. During the entire experiment tad-
poles were fed rabbit pellets ad libitum. Two
transparent cylindrical predator cages were
placed on both sides of each tank to prevent
predators capturing tadpoles but permitting the
flow of chemical signals. Both treatments (pred-
ator presence and absence of predator) were
replicated several times (6 for Alytes, 10 for
Pelodytes, 10 for Hyla, 7 for B. calamita, 6 for
B. bufo and 6 for Rana). In the predator presence
treatment aeshnid odonate naiads larvae were
used as predators. Aeshnid larvae are known to
feed on tadpoles and trigger defences. Each
predator was fed every day with tadpoles reared
in separate containers.
After 2 weeks from the start of the experiment
tadpole activity was measured. We sampled
activity behaviour by counting the number of
tadpoles moving in each tub the instant the tub
was first viewed (Skelly, 1995). Each tub was
observed every half hour for a total of 30
replicates during 2 weeks. This protocol was
repeated for all six species.
In order to obtain data on tadpole morphology,
4 weeks after the start of the experiment all the
tadpoles were individually photographed with a
grid background. We noted Gosner developmen-
tal stage and measured six traits which had been
taken as indications of plasticity in previous
studies (Van Buskirk & Relyea, 1998; Van
Buskirk, 2002): body length, body depth, tail fin
length, tail fin depth, tail muscle length and tail
muscle depth.
Experiment statistical analyses
To measure the behavioural response of tadpoles
in the presence of a predator the mean proportion
of active tadpoles per tub was calculated. These
proportional data were arc-sine square root
transformed previously to test the hypothesis that
the mean number of active tadpoles would differ
between treatments (presence or absence of
predator) using ANOVA analyses. Developmen-
tal stage data were log transformed, and differ-
46 Hydrobiologia (2007) 583:43–56
123

ences between treatments within species were
analysed with ANOVA analyses.
Before morphological plasticity response anal-
yses, tadpole measurements were corrected for
variation in body size. To generate size-corrected
measures, we used the residuals of the morpho-
logical measurements of log-transformed traits
after regressions against body size. We used
centroid size, obtained from landmarks, as a
measure of body size (Loy et al., 1993). Coordi-
nates of these landmarks were collected using the
TPSDIG computer program, version 1.30 (Rohlf,
2001). The centroid size, the square root of the
sum of squared distances of a set of landmarks
from their centroid (Bookstein, 1991), was calcu-
lated for each specimen and used to represent
size. After performing this correction, tadpole
morphology for each species in the two treat-
ments was tested, first with multivariate analysis
for all traits together and second with a univariate
analysis for each variable.
We measured differences in plasticity among
species (both activity and morphological traits)
and their effect on development by examining
changes in traits that occurred between treat-
ments divided by the mean value of the trait in
the absence of predator treatment ([presence of
predator – absence of predator]/absence of pred-
ator) (Van Buskirk, 2002). In the presence of
predator, positive values of plasticity reflect an
increase in the value of the trait, whereas negative
values show a decrease in this trait. The magni-
tude of plasticity in traits can be compared among
species because they are all represented in unit-
less measures of proportional change. We calcu-
lated a overall proportional change in
morphology as the mean of the absolute values
of proportional changes in the six size-corrected
traits considered (Van Buskirk, 2002). If the traits
measured affect species performance within a
habitat type (temporality and predation risk),
then species from distinct habitats should differ in
trait values. Therefore, we first tested for differ-
ences among species regardless of the kind of
habitat their occupied. We used univariate and
multivariate analyses on activity, developmental
stage and on morphological traits to determine
whether phenotypes of the species were affected
by the presence of predator. To consider the
effects of habitat we ran analyses of variance
using habitat as a fixed factor and species nested
within habitat as a random factor. Species were
nested within habitats following field data. As we
expected that the plasticity response would be in
function of habitat temporality, we considered
two groups of species: temporary ponds breeders
(species which preferably occupied ephemeral
and temporary ponds) and permanent pond
breeders (species from more predictable season-
ally and permanent water).
As species cannot be considered independent
data points because trait values could be influ-
enced by shared common ancestry (Felsenstein,
1985; Richardson, 2001a), we tested whether the
distribution of a particular species in terms of
phenotypic plasticity is correlated with its phy-
logeny. We compare proportional change in
activity for each species and the overall propor-
tional change in morphology depending on the
phylogenetic topology of the six species studied.
The phylogenetic relationships between the six
species were reconstructed using the combined
data set of three genes: 12S, 16S and cyt b.
Sequences were obtained from specimens in a
personal collection (collected and sequenced by
S. Carranza) and from the GenBank database. All
sequences were compiled, aligned and refined
manually using Sequence Navigator. Observed
distances in pairwise comparison were obtained
using PAUP software. We tested phylogenetic
independence of larval traits with the computer
program ‘‘Phylogenetic Independence 2.0’’ (Re-
eve and Abouheif, 2003). Tests For Serial Inde-
pendence (TFSI) on continuous data were
performed using the phylogenetic topology and
node distances obtained from molecular recon-
struction. Topology was randomly rotated
10,000 times to build the null hypothesis.
Results
Quantification of species abundances and
predation risk along the gradient
We surveyed 49 ephemeral pools, 85 temporary
ponds and 53 permanent ponds, with a total of
24,938 tadpoles counted during field sampling.
Hydrobiologia (2007) 583:43–56 47
123

Species richness ranged from 0 to 6 species per
locality, without significant statistical differences
in the three habitats considered (F2, 184 = 2.537;
P = 0.0818; Fig. 1a). Total tadpole abundance did
not vary significantly among hydroperiod catego-
ries (F2, 184 = 1.227; P = 0.2954; Fig. 1a). Relative
abundances of larval amphibians, predators taxo-
nomically groups and the predator size categories
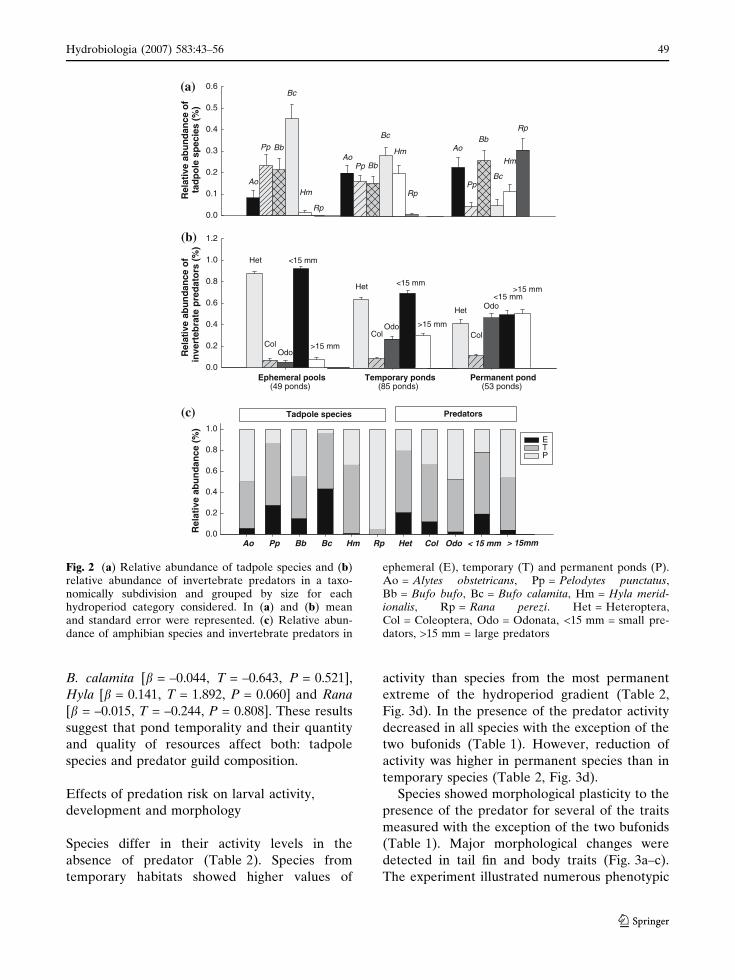
varied among hydroperiod categories (Figs. 2a, b,
c). Abundances of most amphibian species were
determined by the hydroperiod category, showing
a segregation of species along the hydroperiod
gradient. Pelodytes and B. calamita were signifi-
cantly related to ephemeral and temporary ponds
(F2, 184 = 4.69; P = 0.0102 and F2, 184 = 12.42;
P < 0.001 respectively); Alytes, Hyla and B. bufo
were more abundant in temporary and permanent
ponds (F2, 184 = 12.44; P < 0.001; F2, 184 = 9.81;
P < 0.001 and F2, 184 = 4.35; P = 0.014 respec-
tively); and Rana mainly occupied permanent
ponds (F2, 184 = 34.10; P < 0.001). Based on these
preferences we classified B. calamita and Pelo-
dytes as typical temporary species (Habi-
tat = Temporary), and Alytes, B. bufo, Hyla and
Rana as permanent species (Habitat = Perma-
nent) to conduct nested analyses.
The groups of invertebrate predators also
differed along the gradient. Only 8 of the 49
ephemeral ponds (16.3%) were predator-free,
whereas none of the temporary and permanent
ponds were free of predators. Heteroptera and
small predators were more abundant in tempo-
rary ponds (F2, 184 = 56.48; P < 0.001 and
F2, 184 = 13.02; P < 0.001 respectively), whereas
the abundance of Odonata and large predators
increased along the hydroperiod gradient, with
higher values in permanent ponds (F2, 184 = 53.19;
P < 0.001 and F2, 184 = 34.10; P < 0.001 respec-
tively) (Fig. 2b). No differences between habitats
was found for Coleoptera (F2, 184 = 2.44;
P = 0.089). Abundance of predators at which
each amphibian species or species grouped by
habitat (temporary vs. permanent) was exposed
in field varies (F5, 369 = 4.14; P = 0.001 and
F1, 369 = 18.11; P < 0.001 respectively), but not
for species nested within habitat (F4, 369 = 0.06;
P = 0.505). Species from permanent habitats were
exposed to more predators than species from
temporary habitats (Fig. 1b).
Larval amphibian species abundance was cor-
related with invertebrate predator abundance.
Pelodytes and B. calamita were negatively corre-
lated with invertebrate predators (r = –0.168;
P = 0.021; N = 187 and r = –0.271; P < 0.001;
N = 187 respectively), whereas Hyla and Rana
were positively correlated with large predators
(r = 0.188; P = 0.01; N = 187 and r = 0.229;
P = 0.002; N = 187 respectively). Partial set cor-
relation analysis don’t showed any correlation
between tadpole abundance and predator abun-
dance after removing the effect of hydroperiod
(Pelodytes [b = –0.019, T = –0.275, P = 0.783],
Fig. 1 (a) Larval amphibian abundances using catch perunit effort (black boxes) and larval amphibian speciesrichness (white boxes) in the three hydroperiods consid-ered. (b) Abundance of potentially predators at ponds inwhich tadpoles of the different amphibian species were
found. Mean and standard error were represented.Ao = Alytes obstetricans, Pp = Pelodytes punctatus,Bb = Bufo bufo, Bc = Bufo calamita, Hm = Hyla merid-ionalis, Rp = Rana perezi
48 Hydrobiologia (2007) 583:43–56
123
B. calamita [b = –0.044, T = –0.643, P = 0.521],
Hyla [b = 0.141, T = 1.892, P = 0.060] and Rana
[b = –0.015, T = –0.244, P = 0.808]. These results
suggest that pond temporality and their quantity
and quality of resources affect both: tadpole
species and predator guild composition.
Effects of predation risk on larval activity,
development and morphology
Species differ in their activity levels in the
absence of predator (Table 2). Species from
temporary habitats showed higher values of
activity than species from the most permanent
extreme of the hydroperiod gradient (Table 2,
Fig. 3d). In the presence of the predator activity
decreased in all species with the exception of the
two bufonids (Table 1). However, reduction of
activity was higher in permanent species than in
temporary species (Table 2, Fig. 3d).
Species showed morphological plasticity to the
presence of the predator for several of the traits
measured with the exception of the two bufonids
(Table 1). Major morphological changes were
detected in tail fin and body traits (Fig. 3a–c).
The experiment illustrated numerous phenotypic
(a)
(b)
(c)
Rel
ativ
e ab
un
dan
ce o
f t
adp
ole
sp
ecie
s (%
)R
elat
ive
abu
nd
ance
of
inve
rteb
rate
pre
dat
ors
(%
)
Ephemeral pools(49 ponds)
Temporary ponds(85 ponds)
Permanent pond(53 ponds)
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Ao
Pp Bb
Bc
Hm
Rp
PpAo
Bb
Hm
Rp
PpBc
Hm Rp
AoBb
Bc
Het
Het
Het
Col >15 mm
Col Col
Odo
Odo
Odo
<15 mm
<15 mm
<15 mm
>15 mm
>15 mm
Ao Pp Bb Bc Hm Rp Het Col Odo0.0
0.2
0.4
0.6
0.8
1.0E T P
Rel
ativ
e ab
un
dan
ce (
%)
Tadpole species Predators
< 15 mm > 15mm
Fig. 2 (a) Relative abundance of tadpole species and (b)relative abundance of invertebrate predators in a taxo-nomically subdivision and grouped by size for eachhydroperiod category considered. In (a) and (b) meanand standard error were represented. (c) Relative abun-dance of amphibian species and invertebrate predators in
ephemeral (E), temporary (T) and permanent ponds (P).Ao = Alytes obstetricans, Pp = Pelodytes punctatus,Bb = Bufo bufo, Bc = Bufo calamita, Hm = Hyla merid-ionalis, Rp = Rana perezi. Het = Heteroptera,Col = Coleoptera, Odo = Odonata, <15 mm = small pre-dators, >15 mm = large predators
Hydrobiologia (2007) 583:43–56 49
123

differences among species and for species nested
within the appropriate habitat affiliation (Ta-
ble 2), with higher values of morphological
changes for temporary habitat species. Species
were highly variable in predator-induced mor-
phological phenotypic plasticity but with a com-
mon response in activity, which varied in
magnitude depending on the species and habitat
occupied by species.
Also, developmental rate differed between
treatments within species with the exception of
Pelodytes (Table 1). Species and the two groups
considered in function of habitat differed in
proportional developmental stage (Table 2). Spe-
cies from permanent habitats showed a great
reduction of development whereas development
of species from temporary ponds remains unal-
tered. Proportional developmental stage change
was correlated with proportional change in activ-
ity (r = 0.835; P = 0.039; N = 6; Fig. 4b), but not
with overall proportional morphological change
(r = –0.327; P = 0.526; N = 6; Fig. 4a).
The TFSI showed no significant phylogenetic
autocorrelation among species for the morphol-
ogy data set (C-statistics = 0.0930; P = 0.2820)
(Fig. 4c), or for proportional change in activity
(a) (b)
(d)(c)
Bc
PpBb
HmAo
Rp
Activity in absence of predator (%)
Act
ivity
in p
rese
nce
of p
reda
tor
(%)
Pp
Bb
Rp
Hm
Ao
Bc
0.1
0.08
0.06
0.04
0.02
0
-0.02-0.01 0 0.01 0.02 0.03 0.04
Proportional change in body length
Pro
port
iona
l cha
nge
in b
ody
dept
hPp
Rp
Bb
Bc
Ao
Hm
-0.06 -0.04 -0.02 0 0.02 0.04
0.04
0.03
0.02
0.01
0
-0.01
-0.02
0.05
Pp
RpAoHm
Bc
Bb
-0.01 0 0.01 0.02 0.03 0.04
0.1
0.08
0.06
0.04
0.02
0
-0.02
Proportional change in tail fin length
Pro
port
iona
l cha
nge
in ta
il fin
dep
th
Proportional change in tail muscle length
Pro
port
iona
l cha
nge
in ta
il m
uscl
e de
pth
0.0 0.2 0.4 0.6 0.8 1.00.0
0.2
0.4
0.6
0.8
1.0
Fig. 3 Predator-induced plasticity in the body (a), tail fin(b) and tail muscle (c) shape of amphibian species. Figureshows changes in shape due to the presence of predator(mean and standard error). Dashed lines indicate the casein which there was no plasticity. Fig. (d) shows proportionof time active in the presence and absence of predator.
Species near the dashed line showed little behaviouralresponse to predator. Ao = Alytes obstetricans, Pp = Pe-lodytes punctatus, Bb = Bufo bufo, Bc = Bufo calamita,Hm = Hyla meridionalis, Rp = Rana perezi. Open circlesrepresent Temporary habitat species, full circles Perma-nent habitat species
50 Hydrobiologia (2007) 583:43–56
123

(C-statistics = 0.1451; P = 0.2882) (Fig. 4d).
These results indicated little correlation between
these traits and their phylogenetic story for the six
species studied.
Discussion
Quantitative field data showed a turnover in
anuran larvae species along the hydroperiod
Table 1 Summary of univariate and multivariate ANOVA on plasticity in activity, developmental stage and morphology intadpoles of the six species
Source ofvariation
Univariate ANOVA on
Activity level Developmental stage
d.f. F P d.f. F P
Alytes 1, 9 27.69 0.0005 1, 9 7.2 0.0251Pelodytes 1, 17 17.79 0.0005 1, 17 0.1 0.7454B. bufo 1, 10 0.72 0.4136 1, 10 6.2 0.0318B. calamita 1, 12 2.18 0.1655 1, 12 15.2 0.0021Hyla 1, 17 14.31 0.0014 1, 17 9.8 0.0061Rana 1, 9 29.37 0.0004 1, 9 14.41 0.0042
Source ofvariation
d.f. MANOVA onmorphology
Univariate ANOVA on
F P Bodylength
Bodydepth
Tail finlength
Tail findepth
Tail musclelength
Tail muscledepth
Alytes 6, 4 2.08 0.2485 8.15* 2.14 1.19 0.49 1.61 12.97*Pelodytes 6, 12 39.46 <0.0001 19.13* 23.99* 38.61* 76.58* 14.99* 48.97*B. bufo 6, 5 3.27 0.1068 0.86 10.41* 0.88 1.39 0.87 0.81B. calamita 6, 7 1.63 0.2663 4.32 0.41 0.24 0.59 2.93 0.02Hyla 6, 12 20.48 <0.0001 2.27 0.01 1.47 97.87* 0.03 0.45Rana 6, 4 3.83 0.1069 0.43 0.78 14.77* 33.88* 0.30 0.91
Entries for individual morphological traits are F-values in univariate ANOVA, asterisk indicates statistical significance atalpha < 0.05
Table 2 ANOVA of the eight standardized variables (unitless magnitude of the phenotypic plasticity) and overall pro-portional change in morphology between species, habitats and with species nested within habitat
Variable Source of variation
Species (SP) (df = 5)(error = 36)
Habitat (HAB) (df = 1)(error = 40)
Species (SP[HAB]) (df = 4)(error = 36)
Activity level (absence ofpredator)
13.788** 42.097** 4.463*
Proportional change in activitylevel
9.985** 9.439* 8.380**
Developmental stage 12.938** 11.780* 10.445**Body length 17.076** 14.824** 13.139**Body depth 23.274** 13.346** 19.562**Tail fin length 46.138** 13.491** 40.857**Tail fin depth 61.708** 0.575 75.913**Tail muscle length 16.418** 8.183* 15.508**Tail muscle depth 73.985** 21.765** 56.720**Overall proportional change in
morphology73.879** 51.557** 73.420**
Notes: Six species, and Ephemeral-Temporary (Pelodytes and B. calamita) versus Temporary-Permanent habitat species(Alytes, B. bufo, Hyla and Rana) are compared (Habitat). Table entries are F ratios; * P < 0.05; ** P < 0.001
Hydrobiologia (2007) 583:43–56 51
123

habitat gradient as documented in previous stud-
ies (Snodgrass et al., 2000; Babbitt et al., 2003).
Our results differ significantly with respect to the
mechanisms that shape community structure
along the freshwater gradient proposed by Well-
born et al. (1996), where it was assumed that
permanence transition (between temporary hab-
itats and permanent habitats) also coincides with
the shift from communities without predators to
communities with large invertebrate top preda-
tors. The present study showed that the relative
abundance of large invertebrate predators is
greater in permanent ponds, but temporary ponds
could not be considered predator-free habitats.
The transition towards communities dominated
by large invertebrate predators occurs between
ephemeral pools and temporary ponds (Skelly,
1996; Wilbur 1997; Stoks & McPeek, 2003).
Tadpole assemblage may differ in the first tran-
sition due to the high risk of ephemeral pools
drying up and the different predator assemblages
in the two habitats: ephemeral and temporary
ponds. In the second transition detected in the
system studied (from temporary to permanent
ponds) the hydroperiod also seems to be the most
important factor. In the Mediterranean system of
isolated ponds studied, differences between tad-
pole assemblages could be attributed to factors
other than an increase in the abundance of
predators. Tadpole species assemblages can be
explained by the interplay of species differences
in developmental plasticity in response to pond
drying (Richter-Boix et al., 2006a), differences in
phenological breeding between species (Richter-
Boix et al., 2006b), strong interspecific interac-
tions (Richter-Boix et al., 2007) and vulnerability
Proportional change in morphology0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7
Pro
port
iona
l cha
nge
inde
velo
pmen
tals
tage
-0.20
-0.15
-0.10
-0.05
0.00
0.05
0.10
Bc
Bb
PpHm
Ao
Rp
Pp
Bc
Bb
AoHm
Rp
(b)
(c) (d)
Ao
Pp
Hm Bc
Bb
Rp Ao
Pp
Hm Bc
Bb
Rp
(a)
Proportional change in activity-0.7 -0.6 -0.5 -0.4 -0.3 -0.2 -0.1 0.0 0.1
Fig. 4 (a) Relationship between morphological plasticityand change in developmental stage, and (b) betweenbehavioural plasticity and change in developmental stage.All proportions are calculated in comparison to thepredator-free treatment, bars indicate standard error.Hypothesized phylogenetic relationships among the 6anuran species, where branch lengths did not correspond
to the real molecular data estimated. Sizes of full circlesrepresent magnitude of plasticity for morphology (c) andfor behaviour (d). Ao = Alytes obstetricans, Pp = Pelo-dytes punctatus, Bb = Bufo bufo, Bc = Bufo calamita,Hm = Hyla meridionalis, Rp = Rana perezi. Open circlesrepresent Temporary habitat species, full circles Perma-nent habitat species
52 Hydrobiologia (2007) 583:43–56
123

to predation. This system contains a low number
of predator-free ponds and, with the exception of
B. calamita and Pelodytes, the other species never
occur in predator-free ponds, although high vari-
ations were observed in predator composition and
concentration between ponds. The high spatial
and temporal variability in predator communities
to which tadpole species were subjected (Van
Buskirk & Arioli, 2005) supports the theory that
phenotypic plasticity is developed and maintained
in species or populations exposed to heteroge-
neous and unpredictable environments (Via et al.,
1995; Schlichting & Pigliucci, 1998; DeWitt &
Scheiner, 2004).
The laboratory experiments demonstrated that
four of the six anuran species exhibited predator-
induced plasticity in behavioural and morpholog-
ical traits, results that are consistent with previous
studies (e.g. Lardner, 2000; Van Buskirk, 2002).
The results of the experiments, in conjunction
with field observations of species habitat prefer-
ences and predation risk, confirm in part the
expected link between phenotypic plasticity and
habitat use according to temporality and preda-
tion risk. Species from the most temporary part of
the hydroperiod range, commonly exposed to the
development/predation trade-off, exhibited dif-
ferent integration of the plastic traits evaluated to
species from the most permanent ponds. Tempo-
rary species showed higher values of morpholog-
ical changes (Relyea & Werner, 2000) and low
activity reduction. Morphological changes (dee-
per and larger tails) were consistent with the
adaptive changes observed in several larval
amphibian species (Van Buskirk et al., 1997;
Van Buskirk & Relyea, 1998). Species could
therefore maintain a high level of activity while
responding morphologically rather than behavio-
urally to the presence of predators. Presumably,
these species must maintain activity levels in
order to achieve sufficient growth and develop-
mental rates to allow them to metamorphose
before ponds dry up (Laurila & Kujasalo, 1999;
Anholt et al., 2000). The positive correlation
between developmental change and activity
response supports this idea. In ephemeral ponds
the cost of greater activity (more rapid growth
and development) is a smaller increase in preda-
tion risk compared to the situation in more
permanent ponds, in which predator densities
are higher (Skelly, 1995). Unlike species from
temporary habitats, those from the most perma-
nent part of the range may tip the balance in
favour of predation risk as they are able to remain
in the aquatic habitat as larvae for a longer period
before metamorphosis (Anholt et al., 2000). As a
result, they showed a greater plasticity in activity
than typical temporary pond species, which in
turn leads to a greater reduction in developmen-
tal rates. Development stage change was strongly
correlated with activity levels but not with mor-
phological changes, which suggests that morpho-
logical changes have little impact on
developmental rates. Contrary to the results of
the present study, previous works have suggested
that morphological defences in response to pre-
dators have come at the cost of decreased
developmental rates (Relyea, 2002; Teplitsky
et al., 2005) and growth rates (Van Buskirk,
2000; Relyea, 2002; Teplitsky et al., 2005). Unfor-
tunately no data about growth rates was available
to estimate the effect of plasticity on growth
among species and to make comparisons with
other studies. However, the detected impact of
morphological phenotypic plasticity on develop-
ment is weak with respect to activity in the work
of Relyea (2002) after Bonferroni corrections,
thus supporting our results. Teplitsky et al. (2005)
did not measure activity and attributed all devel-
opmental cost to morphological changes without
including behavioural information, when is well
known that tadpoles exhibit both morphological
and behavioural inducible defences in response to
predators (e.g. Lardner, 2000; Van Buskirk, 2002)
and it is possible that part of the developmental
cost observed was related to changes in activity.
Tadpole species differed in the integration of
the phenotypic traits measured, but differences
observed between species could not be attributed
only to habitat. It is possible that taxonomy and
habitat interact in determining anti-predator
responses (Richardson, 2001a; Van Buskirk,
2002). This may reflect the presence of alternative
strategies available for persistence in a given
habitat type (Richardson 2001b, 2002a). Our
results did not reveal any phylogenetic correla-
tion between the traits studied, although the small
number of species considered and the great
Hydrobiologia (2007) 583:43–56 53
123

phylogenetic distances between them may ob-
scure any possible relationship. In addition, the
similar response in activity and morphology
between the two bufonid species suggest some
phylogenetic constraints. Previous studies with a
greater number of species have demonstrated
potential constraint in the evolution of predator-
induced plasticity in anurans, with some phyloge-
netic inertia in behavioural and morphological
trait values (Richardson, 2001b, 2002a; Van Bus-
kirk, 2002). Species from the same habitat type do
not always converge on similar morphological or
behavioural phenotypes, which suggests that lin-
eages in this system can evolve to one of many
different evolutionary solutions (Richardson,
2001a; Van Buskirk, 2002). The absence of or
weak responses to the presence of a predator
detected in bufonids suggest that Bufo use an
anti-predator mechanism other than reduction of
activity levels or morphological changes, such as
chemical deterrents or the formation of cohesive
swarms (Watt et al., 1997). Differences in mor-
phological changes between species from a sim-
ilar habitat (Alytes, Hyla and Rana) could in part
reflect differences in microhabitat use in the
water column and biomechanical constraints
(Richardson, 2002b). Microhabitat use differs
between these species (Dıaz-Paniagua, 1987a, b)
and it is plausible that morphological changes
affect swimming speed, stability and manoeuvra-
bility. Studies of the phenotypic integration of
morphological traits in different environments
and the consequences and ecology of these traits
are needed to understand differences in morpho-
logical plasticity between species.
A potential weakness of this study in terms of
its ability to resolve possible trade-offs between
species along the pond permanency and predator
risk gradients is the lack of consideration given to
the effects of pond drying on predator-induced
phenotype. We know that species differ in their
developmental phenotypic plasticity in response
to pond drying (Richter-Boix et al., 2006a) and in
their morphological and behavioural plasticity in
response to a predation risk (present study). To
date only three studies have addressed the non-
lethal presence of predators under time con-
straints (Laurila & Kujasalo, 1999; Altwegg, 2002;
Bridges, 2002) and a discrepancy exists between
the findings of these studies. Rana temporaria, a
typical temporary pond breeder, reduced their
predation reaction when exposed to a drying
environment (Laurila & Kujasalo, 1999), whereas
the anti-predator responses of Rana lessonae and
Pseudacris triseriata do not alter under time
constraints (Altwegg, 2002; Bridges, 2002). It is
possible that species from temporary ponds with
fewer predators prioritise the desiccation risk
over the predation risk, whereas species from
permanent ponds and exposed to a greater
abundance of predators perceive predation as a
greater immediate threat than pond drying and
consequently do not take greater risks. Future
experimental research into the joint effects of
pond drying and predators on the development
and phenotypic plasticity of tadpoles is required
in order shed light on these relationships and the
distribution of species along the hydroperiod
gradient.
Acknowledgements We sincerely thank Nuria Garrigafor help during laboratory experiments and for usefulcomments. We thank Laura Perez and Santi Escartınfor assisting with field surveys, and S. Llacuna andF. Llimona for providing the corresponding permits andfieldwork facilities in the Natural Parks. We also thankS. Carranza for their assistance in the phylogenetictopology reconstruction of the species object of studyand providing personal data for this purpose. We extendour gratitude to two anonymous referees for providinguseful comments and suggestions. Permission to capturewas granted by the Departament de Medi Ambient de laGeneralitat de Catalunya.
References
Altwegg, R., 2002. Predator-induced life-history plasticityunder time constraints in pool frogs. Ecology 83:2542–2551.
Anholt, B. R. & E. E. Werner, 1995. Interaction betweenfood availability and predation mortality mediated byactivity. Ecology 76: 2235–2239.
Anholt, B. R., D. K. Skelly & E. E. Werner, 1996. Factorsmodifying antipredator behaviour in larval toads.Herpetologica 52: 301–313.
Anholt, B. R., E. E. Werner & D. K. Skelly, 2000. Effect offood and predators on the activity of four larval ranidfrogs. Ecology 81: 3509–3521.
Babbitt, K. J., J. B. Matthew & T. L. Tarr, 2003. Patternsof larval amphibian distribution along a wetlandhydroperiod gradient. Canadian Journal of Zoology81: 1539–1552.
54 Hydrobiologia (2007) 583:43–56
123

Bookstein, F. L., 1991. Morphometrics Tool for LandmarkData: Geometry and Biology. Cambridge UniversityPress, Cambridge.
Bridges, C. M., 2002. Tadpoles balance foraging andpredator avoidance: effects of predation, pond drying,and hunger. Journal of Herpetology 36: 627–634.
Chovanec, A., 1992. The influence of tadpole swimmingbehaviour on predation by dragonfly nymphs.Amphibia-Reptilia 13: 341–349.
Cronin, J. T. & J. Travis, 1986. Size-limited predation onlarval Rana aerolata (Anura: Ranidae) by two speciesof backswimmers (Heteroptera: Notonectidae). Her-petologica 42: 171–174 .
DeWitt, T. J. & S. M. Scheiner, 2004. Phenotypic Plastic-ity: Functional and Conceptual Approaches. OxfordUniversity Press, New York, USA.
Dıaz-Paniagua, C., 1987a. Estudio en cautividad de laactividad alimenticia de las larvas de siete especies deanuros. Revista Espanola de Herpetologıa 2: 189–197.
Dıaz-Paniagua, C., 1987b. Tadpole distribution in relationto vegetal heterogeneity in temporary ponds. Herpe-tological Journal 1: 167–169.
Eklov, P. & C. Halvarsson, 2000. The trade-off betweenforaging activity and predation risk for Rana tempo-raria in different food environments. Canadian Jour-nal of Zoology 78: 734–739.
Felsenstein, J., 1985. Phylogenies and the comparativemethod. American Naturalist 125: 1–13.
Gunzburger, M. S. & J. Travis, 2004. Evaluating predationpressure on green treefrog larvae across a habitatgradient. Oecologia 140: 422–429.
Laurila, A. & J. Kujasalo, 1999. Habitat duration, preda-tion risk and phenotypic plasticity in common frog(Rana temporaria) tadpoles. Journal of Animal Ecol-ogy 68: 1123–1132.
Lardner, B., 2000. Morphological and life historyresponses to predators in larvae of seven anurans.Oikos 88: 169–180.
Loy, A., M. Corti & L. F. Marcus, 1993. Landmarks data:size and shape analysis in systematics. A case studyon old world talpidae (Mammalia, Insectivora). InMarcus, L. F., E. Bello & A. Garcıa-Valdecasas (eds),Contributions to Morphometrics. Monografıas delMuseo Nacional de Ciencias Naturales, Madrid,215–239.
McCollum, S. A. & J. D. Leimberger, 1997. Predator-induced morphological changes in an amphibian:predation by dragonflies affects tadpole shape andcolor. Oecologia 109: 612–615.
Reeve, J. & E. Abouheif, 2003. Phylogenetic Indepen-dence. Version 2.0, Department of Biology, McGillUniversity. Distributed freely by the authors onrequest.
Relyea, R.A., 2002. Cost of phenotypic plasticity. Amer-ican Naturalist 159: 272–282.
Relyea, R. A. 2004. Integrating phenotypic plasticity whendeath is on the line: insights from predator–preysystems. In Pigliucci, M. & K. Preston (eds), Pheno-typic Integration: Studying the Ecology and Evolutionof Complex Phenotypes. Oxford University Press,New York, USA.
Relyea, R. A. & E. E. Werner, 1999. Quantifying therelation between predator-induced behavior andgrowth performance in larval anurans. Ecology 80:2117–2124.
Relyea, R. A. & E. E. Werner, 2000. Morphologicalplasticity in four larval anurans distributed along anenvironmental gradient. Copeia 2000: 178–190.
Richardson, J. M. L., 2001a. The relative roles of adapta-tion and phylogeny in determination of larval traits indiversifying anuran lineages. American Naturalist 157:282–299.
Richardson, J. M. L., 2001b. A comparative study ofactivity levels in larval anurans and response to thepresence of different predators. Behavioural Ecology12: 51–58.
Richardson, J. M. L., 2002a. A comparative study of pheno-typic traits related to resource utilization in anurancommunities. Evolutionary Ecology 16: 101–122.
Richardson, J. M. L., 2002b. Burst swim speed in tadpolesinhabiting ponds with different top predators. Evolu-tionary Ecology Research 4: 627–642.
Richter-Boix, A., G. A. Llorente & A. Montori, 2006a. Acomparative analysis of the adaptive developmentalplasticity hypothesis in six Mediterranean anuranspecies along a pond permanency gradient. Evolu-tionary Ecology Research 8: 1139–1154.
Richter-Boix, A., G. A. Llorente & A. Montori, 2006b.Breeding phenology of an amphibian community in aMediterranean area. Amphibia-Reptilia 27: 544–559.
Richter-Boix, A., G. A. Llorente & A. Montori, 2007.Hierarchical competition in pond-breeding anuranlarvae in a Mediterranean area. Amphibia-Reptilia28: in press.
Rohlf, F. J., 2001. Thin-Plate Spline (TPS) Software. Freeavailable on:http://life.bio.sunysb.edu/morph/ (lastmodified October 31, 2005).
Schaffer, H. B., R. A. Alford, B. D. Woodward, S. J.Richards, R. G. Altig & C. Gascon, 1994. Quantitativesampling of amphibian larvae. In Heyer, W. R., M. A.Donnelly, R. W. McDiarmid, L.C. Hayek & M. S.Foster (eds), Measuring and Monitoring BiologicalDiversity Standard Methods for Amphibians. Smith-sonian Institution Press, Washington, 130–141.
Schlichting, C. & M. Pigliucci, 1998. Phenotypic Evolution:A Reaction Norm Perspective. Sinauer, Sunderland,400 pp.
Schneider, D. W. & T. M. Frost, 1996. Habitat duration andcommunity structure in temporary ponds. Journal ofthe North American Benthological Society 15: 64–86.
Skelly, D. K., 1994. Activity level and the susceptibility ofanuran larvae to predation. Animal Behaviour 47:465–468.
Skelly, D. K., 1995. A behavioural trade-off and itsconsequences for the distribution of Pseudacris tree-frog larvae. Ecology 76: 150–164.
Skelly, D. K., 1996. Pond drying, predators, and thedistribution of Pseudacris treefrog tadpoles. Copeia1996: 599–605.
Skelly, D. K. & E. E. Werner, 1990. Behavioral and life-historical responses of larval American toads to anodonate predator. Ecology 71: 2313–2322.
Hydrobiologia (2007) 583:43–56 55
123

Snodgrass, J. W., A. L. Bryan & J. Burger, 2000.Development of expectations of larval amphibianassemblage structure in southeastern depression wet-lands. Ecological Applications 10: 1219–1229.
Stocks, R. & M. A. McPeek, 2003. Predators and lifehistories shape Lestes damselfly assemblages along afreshwater habitat gradient. Ecology 84: 1576–1587.
Teplitsky, C., S. Plenet & P. Joly, 2005. Cost and limits ofdosage response to predation risk: to what extent cantadpoles invest in anti-predator morphology? Oeco-logia 145: 364–370.
Travis, J., W. H. Keen & J. Juilianna, 1985. The role ofrelative body size in a predator–prey relationshipbetween dragonfly naiads and larval anurans. Oikos45: 59–65.
Van Buskirk, J., 2000. The cost of an inducible defense inanuran larvae. Evolution 81: 2813–2821.
Van Buskirk, J., 2002. A comparative test of the adaptiveplasticity hypothesis: relationships between habitatand phenotype in anuran larvae. American Naturalist160: 87–102.
Van Buskirk, J., 2003. Habitat partitioning in Europeanand North American pond-breeding frogs and toads.Diversity and Distributions 9: 399–410.
Van Buskirk, J. & M. Arioli, 2005. Habitat specializationand adaptive phenotypic divergence of anuranpopulations. Journal of Evolutionary Biology 18:596–608.
Van Buskirk, J. & R. A. Relyea, 1998. Natural selectionfor phenotypic plasticity: predator-induced responsesin tadpoles. Biological Journal of the Linnean Society65: 301–328.
Van Buskirk, J., S. A. McCollum & E. E. Werner, 1997.Natural selection for environmentally-induced phe-notypes in tadpoles. Evolution 52: 1983–1992.
Via, S., R. Gomulkiewicz, G. De Jong, S. Scheiner, C. D.Schlichting & P. H. Van Tienderen, 1995. Adaptivephenotypic plasticity: consensus and controversy.Trends in Ecology and Evolution 10: 212–217.
Watt, P. J., S. F. Nottingham & S. Young, 1997. Toadtadpole aggregation behaviour: evidence for a pred-ator avoidance function. Animal Behaviour 54:865–872.
Wellborn, G. A., D. K. Skelly & E. E. Werner, 1996.Mechanism creating community structure across afreshwater habitat gradient. Annual Review in Ecol-ogy and Systematics 27: 337–363.
Werner, E. E. & M. A. McPeek, 1994. Direct and indirecteffects of predators on two anuran species along anenvironmental gradient. Ecology 75: 1368–1382.
Wilbur, H. M., 1997. Experimental ecology of food webs:complex systems in temporary ponds. Ecology 78:2279–2302.
Woodward, B. D., 1983. Predator–prey interactions andbreeding-pond use of temporary-pond species in adesert anuran community. Ecology 64: 1549–1555.
56 Hydrobiologia (2007) 583:43–56
123




















