
xדקדנס כמאפיין סטירי בעיצוב ירידת הדורות על פי "עשו" שלא מאיר שלוOfra Matzov-Cohen
Upload
khangminh22Category
view
5download
0
השפעת אינהיביציה פרסינפטית על מאפייני התגובה CRAYFISH -שריר ב-לגירוי בתדר בסינפסת עצב
מחקר לשם מילוי חלקי של הדרישות
"דוקטור לפילוסופיה"לקבלת תואר מאת
בן אלכהא
הוגש לסינאט אוניברסיטת בן גוריון בנגב
2004 אפריל ד"תשס ניסן
באר שבע

השפעת אינהיביציה פרסינפטית על מאפייני התגובה
CRAYFISH -שריר ב-לגירוי בתדר בסינפסת עצב
מחקר לשם מילוי חלקי של הדרישות
"דוקטור לפילוסופיה"לקבלת תואר
מאת
בן אלכהא
הוגש לסינאט אוניברסיטת בן גוריון בנגב
____________________ אישור המנחה
ללימודי אישור דיקן בית הספר ____________________ מןש קרייט"מחקר מתקדמים ע
2004יולי ד"אב תשס
באר שבע

יורם גרוסמן' פרופ :העבודה נעשתה בהדרכת
פזיולוגיה :במחלקה
מדעי הבריאות :בפקולטה

תוכן עיניינים
תקציר
3.......................................................................................מבוא .1 3.........................................................................................................רקע כללי .1.1 Crayfish..................................................4 - של הopener - בשריר הNMJסינפסת .1.2
4................................................................לית של הסינפסהמורפולוגיה ופיזיולוגיה כל .1.2.1 5................................................................התמיינות שונה של טרמינלים מאקסון יחיד .1.2.2 6..........................................................................הבדלים מורפולוגיים בין הטרמינלים .1.2.3 7............................................................................הבדלים פיזיולוגיים בין הטרמינלים .1.2.4 11......................................................................התגובה הסינפטית לגירוי בתדר גבוה .1.2.5
15..........................................................תפקידה של הדפולריזציה בשחרור הסינפטי .1.3 16..............................................................................מטרות המחקר והחידוש בהן .1.4
19...................................................................................שיטות .2 19..........................................................................................................פרפרט .2.1 19....................................................................................................פרמקולוגיה .2.2 20.............................................................................................אלקטרופיזיולוגיה .2.3
20..........................................................................................................גירוי אקסון .2.3.1 22...........................................................................................................גירוי ישיר .2.3.2 23................................................................................................................רישום .2.3.3 24...........................................................בעיות רישום הנובעות מגירוי בתדרים גבוהים .2.3.4
27...........................................................................................................מדידה .2.4 27....................................................................................חישוב התכולה הקואנטלית .2.4.1
29......................................................................... של התוצאותהערכה סטטיסטית .2.5
31.........................................התגובה לגירוי בתדר בתנאי ביקורת .3 31......................................................................נוהל הגירוי וניתוח התגובה: שיטות .3.1ל גבי השרירהתלות בתדר הגירוי ובמיקום הטרמינלים ע: השחרור הסינפטי במופע הרוויה .3.2
.............................................................................................................33 36.....................התלות בתדר ובמיקום הטרמינלים על גבי השריר: פוטנציאציה מקסימלית .3.2.1 37.................תלות התגובה לגירוי בתדר של טרמינלים מרכזיים במרחק מהאקסון המרכזי .3.2.2 39.................................................................................................................סיכום .3.2.3
40........................................................................השחרור הסינפטי במופע העלייה .3.3 40..................................................................כימות קינטיקת התפתחות הפוטנציאציה .3.3.1 41.........................................................סדרת גירויים ארוכה: קצב התפתחות השחרור .3.3.2 43....מאפייני השחרור הסינפטי מאוכלוסיות טרמינלים שונות בתגובה לסדרת גירויים קצרה .3.3.3 44..............................................קצב התפתחות השחרור בתגובה לסדרת גירויים קצרה .3.3.4

44..................................................עצמת הפוטנציאציה באוכלוסיות שונות של טרמינלים 45.................................................................................................................סיכום .3.3.5
46השפעות שינויים בכניסת הסידן לטרמינל על התגובה לגירוי בתדר .4 46............................................................................................................. מבוא .4.1 46..........י ישיר וההשפעה על התגובה לתדרהגברת כניסת הסידן לטרמינל באמצעות גירו .4.2
46..............................השפעת עוצמת הגירוי הישיר על שיפוע עקום התגובה לגירוי בתדר .4.2.1 49.....................................................................ההשפעה על קצב התפתחות השחרור .4.2.2
49......................ן חוץ תאי על התגובה לתדר של הטרמינליםהשפעת הפחתת ריכוז סיד .4.3 51............................................................................................................סיכום .4.4 .............................................................................................................51 דיון .4.5
51....................................................................................................יציבות הרישום .4.5.1 52.................................................סיות השונותהתמיינות הטרמינלים והומוגניות האוכלו .4.5.2 53...........................................................................שיפוע עקום התגובה לגירוי בתדר .4.5.3 54............................גורמים אפשריים להבדלים בין השחרור מאוכלוסיות טרמינלים שונות .4.5.4 57..........................................................ולת היחס הפוך בין כניסת סידן בעקבות גירוי ליכ 57.......................................................................................פוטנציאציה של לטרמינל .4.5.5 58..........................................................בעקום התגובה לגירוי בתדר" מדרגות"נוכחות .4.5.6 60.................................................אספקטים פיזיולוגיים של עקום התגובה לגירוי בתדר .4.5.7
61............השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר .5 Crayfish..........................................................61 -אינהיביציה פרסינפטית ב: מבוא .5.1
61...........................................................................................פיזיולוגיה ומורפולוגיה .5.1.1 62.................................................................................................ייםמודלים חשמל .5.1.2 62.................................................................................................אנליזה קואנטלית .5.1.3 62.....................................................................עדות למנגנונים אינהיביטוריים נוספים .5.1.4 63.................................................................................................אינהיביציה טונית .5.1.5 64.................................................................................................................סיכום .5.1.6
64................................................. וקינטיקת ההשפעהGABA -רגישות הטרמינלים ל .5.2 66............................................... על מופע הרוויה של התגובה לתדרGABAהשפעת .5.3
67..........................הפעלת אינהיביציה לא משפיעה על שיפוע עקום התגובה לגירוי בתדר .5.3.1 67........הפעלת אינהיביציה לא מקטינה את ההבדלים האיכותיים בין אוכלוסיות הטרמינלים .5.3.2 69.................................................. על ערכי הפוטנציאציה המקסימליתGABAהשפעת .5.3.3 69.................................................................................................................סיכום .5.3.4
70.......................................... על מופע העלייה של השחרור הסינפטיGABAהשפעת .5.4 70................. על קצב התפתחות השחרור בתגובה לסדרת גירויים ארוכהGABAהשפעת .5.4.1 72......... על התפתחות עצמת הפוטנציאציה בתגובה לסדרת גירויים קצרהGABAהשפעת .5.4.2 73.................................................................................................................סיכום .5.4.3
74......................י גירוי האינהיביטור על התגובה לתדר"השפעת אינהיביציה המושרית ע .5.5 74..............................................................................................................הקדמה .5.5.1 74.............ל עקום התגובה לתדר של טרמינלים פרוקסימלייםהשפעת גירוי האינהיביטור ע .5.5.2

76.............................................השפעת גירוי האינהיביטור על קצב התפתחות השחרור .5.5.3 77.................................................................................................................סיכום .5.5.4
77............................................................................................................. דיון .5.6 GABA.......................................77 -הבדל ברגישות טרמינלים מרכזיים ופרוקסימליים ל .5.6.1 78........................?מה עומד בבסיס האינהיביציה אם לא הפחתה בכניסת הסידן לטרמינל .5.6.2 81............?מלייםממה נובעים ההבדלים בין התגובה לתדר של טרמינלים מרכזיים ופרוקסי .5.6.3 81............................................הקשר בין הפעלת אינהיביציה לקצב התפתחות השחרור .5.6.4 84......העמידות היחסית של הפוטנציאציה לאינהיביציה בהשוואה לעמידות השחרור הבודד .5.6.5
GABA.............85י " של האינהיביציה המתווכת עהיבטים מנגנוניים .6 85............................................................................................................. מבוא .6.1 86....................................... על התגובות לגירוי ישיר מול גירוי האקסוןGABAהשפעת .6.2
86..............מרכזיים הסמוכים לאקסוןאינהיביציה בגירוי ישיר מול גירוי האקסון בטרמינלים .6.2.1 89...........אינהיביציה בגירוי ישיר מול גירוי אקסון בטרמינלים מרכזיים המרוחקים מהאקסון .6.2.2
90................................................ על מאפייני התגובה לתדר בגירוי ישירGABAהשפעת 91........................................................יון פרמקולוגי של המוליכות האינהיביטוריתאפ .6.3
6.3.1. GABAA.............................................................................................................91 6.3.2. GABAC.............................................................................................................93 6.3.3. GABAB.............................................................................................................94
95............................................................................................................. דיון .6.4 96....................................................האינהיביציה בטרמינלים מרכזיים הסמוכים לאקסון .6.4.1 97...............................................האינהיביציה בטרמינלים מרכזיים המרוחקים מהאקסון .6.4.2 98.................................................................אפיון הרצפטור המתווך את האינהיביציה .6.4.3
101....................................................הממצאים וחשיבותםעיקרי .7
References..............................................................................103

:רשימת איורים וטבלאות
Figure 1 : 5..............................................................................תרשים הפרפרט
Figure 2 : 24............................................................מערך הניסוי ושיטת הרישום
Figure 3 : 29...........................................................מדידת גודל הקואנטום הבודד
Figure 4 : 32....................ינלים פרוקסימליים ומרכזייםהתגובה לגירוי בתדר של טרמ
Figure 5 : 35...................................מאפייני התגובה לתדר ופוטנציאציה מקסימלית
Figure 6 : 38...................תגובות שונות לתדר בתוך אוכלוסיית הטרמינלים המרכזיים
Figure 7 : 42....התפתחות השחרור בתוך סדרת הגירויים באוכלוסיות טרמינלים שונות
Figure 8 : 48............................................התגובה לתדר בגירוי ישיר מול גירוי עצב
Figure 9 : 50..................השפעת שינויים בכניסת סידן לטרמינל על השחרור הסינפטי
Figure 10 : השפעתGABA65.. על התגובה לתדר של טרמינלים פרוקסימליים ומרכזיים
Figure 11 : השפעתGABA68............... על הפוטנציאציה המקסימלית של הטרמינלים
Figure 12 : השפעתGABA71..........................קצב התפתחות השחרור על משרעת ו
Figure 13 : 74.השפעת גירוי האינהיביטור על התגובה לתדר בטרמינלים הפרוקסימליים
Figure 14 : השפעתGABA87............ על התגובה לגירוי עצב מול התגובה לגירוי ישיר
Figure 15 : השפעותpicrotoxin ,TPMPA ,baclofen92........... על השחרור הסינפטי
Table 1 : מתן , גירוי ישיר, השפעות הפחתת ריכוז סידן חוץ תאיGABA
וגירוי האינהיביטור על מאפייני התגובה לתדר באוכלוסיות
100...........................................................................טרמינלים שונות

תקציר___________________________________________________________________
התגברות השחרור הסינפטי בתגובה למתן גירויים סמוכים הוא אחד המאפיינים הקלאסיים של
, של הפסיליטציהבמשך השנים פורסמו עבודות רבות המתארות צדדים שונים . סינפסה כימית
ותופעות הגברה נוספות , )התגברות השחרור בתגובה לשלושה גירויים סמוכים ויותר(הפוטנציאציה
למרות הידיעות הרבות שנאספו בנושא עדיין חסר . הנמשכות זמן ממושך יותר לאחר הפסקת הגירוי
בה לגירוי של השחרור בתגוקצרת הטווחתיאור מקיף המתאר את ההיבטים השונים של ההגברה
והאופן בו , טרם אופיינה התלות בין תדר ומשך הגירוי לרמות השחרור שהוא מעורר. בתדר גבוה
ל "טרם ברורים ההבדלים בהיבטים הנ. שחרורהמשפיע תדר הגירוי על קינטיקת וקצב התפתחות
נפטיות שונות כמו ולא האופן בו השפעות פרסי, )פאזיות וטוניות(בין אוכלוסיות סינפסות שונות
שינויים בריכוז הסידן החוץ תאי או הפעלת אינהיביציה פרסינפטית עשויות להשפיע על מאפייני
.עקום התגובה לגירוי בתדר של סינפסה נתונה
בשריר הפותח את (Neuromuscular Junction - NMJ)שריר -שאלות אלה נחקרו בסינפסת עצב
: הניסויים בוצעו בשתי קבוצות טרמינלים שונות. Crayfish -ברגל ההליכה הראשונה ב, הצבת
אקסוןתוך שימוש בגירוי , )בעלי מאפיינים טוניים(ומרכזיים , )בעלי מאפיינים פאזיים(פרוקסימליים
.ובשיטת הגירוי הישיר
בעבודה הנוכחית אופיין העקום המתאר את התלות בין תדר הגירוי לרמות השחרור המקסימליות
עם , (power)עקום זה נמצא מתאים לפונקצית חזקה . ) שחרור-עקום תדרלהלן (רשהוא מעור
קיים . עבור הטרמינלים המרכזיים1.68 - עבור הטרמינלים הפרוקסימליים ו1.05 -מעריך השווה ל
תאי באותם תוךל לעקום המתאר את השחרור כפונקציה של ריכוז הסידן ה"דמיון רב בין העקום הנ
מחזק זה דמיון. )1982a( .Parnas et alוהחוץ תאי , )Wright et al., 1996a, b( יםטרמינל
. אלה בטרמינליםסידן התוך תאילריכוז התדר הגירוי בין עבודות קודמות המצביעות על יחס ישר
ולהיפך , ריכוז הסידן החוץ תאי שחרור הפך תלול יותר בעקבות הפחתה ב-שיפוע העקום תדר גירוי
בעוצמה הגוררת שחרור גבוה יותר משמעותית מזה (טרמינלהשל עם מתן גירוי ישיר מתון יותר -
עשוי לשמש מדד עקיף למידת תלותה שיפועקורלציה זו מראה ש. )האקסוןהנגרם הודות לגירוי
ו של ריכוז הסידן התוך תאי בהשוואה לז, התרומה לשחרור של כניסת הסידן בתגובה לגירוי
.(Ravin et al., 1999) הממוצע

ין המלמדים ב, של אוכלוסיות טרמינלים שונות נמצאו הבדלים מהותייםשחרור -תדרבין עקומי
נקבע על ידי המשקל היחסי של תרומה זו. לשחרורהיחסית תרומתם המידה השונה של על השאר
.מעצבב את השרירתדר הירי של המוטורנוירון האקסיטטורי היחיד ה
תוך , בתגובה לסדרת גירויים קצרה בתדר גבוה השחרור התפתחותבעבודה זו ינה אופיכמו כן
שחרורהלקצב התפתחות ) מידת הכפלת התגובה בתוך סדרת הגירויים(פוטנציאציה ההבדלה בין
, הבדלה זו נעשתה בשתי אוכלוסיות הטרמינלים). גובה להגיע לרוויהמספר הגירויים הדרוש לת(
לרוב נמצא מתאם הפוך בין שני פרמטרים . ואפשרה לקבל תמונה מלאה יותר של התהליך בכללותו
מפתחים פוטנציאציה חזקה ) טוניים(טרמינלים המתאפיינים בשחרור התחלתי נמוך מאוד : אלה
שינויים . ורשים מספר גירויים גבוה יותר לצורך הגעה לרוויהאולם ד, )הכפלה מהירה של השחרור(
. שחרורהבכניסת הסידן לטרמינל השפיעו ביחס ישר על קצב התפתחות
השינויים בדפוסי השחרור בעקבות הפעלת אינהיביציה פרסינפטית על הטרמינלים האקסיטטוריים
ביטורי הפועל על השחרור שלא דרך הפחתה קיום מנגנון אינהיבממצאים קודמים בדבר תומכים
אולם , GABAA -י רצפטור בעל דמיון פרמקולוגי ל"אינהיביציה זו מתווכת ע. בכניסת סידן לטרמינלים
.ובמיוחד בטרמינלים המרכזיים, מנגנון הפעולה שלה ברמה הפיזיולוגית טרם ברור

3
מבוא .1___________________________________________________________________
רקע כללי .1.1ועד אפיון , החל ברמה של רשתות תאים, פעילות מערכת עצבית ניתן לנתח ברמות שונות
עליהם מושתת , עבודה זו עוסקת בניתוח תהליכים סינפטיים. תהליכים בודדים ברמה מולקולרית
בשריר פותח הצבת (NMJ)שריר -לסינפסת עצב. עיקר נפח התקשורת הבין תאית במערכות אלה
(opener) ברגל ההליכה ב- Crayfishמספר יתרונות ההופכים אותה לאטרקטיבית בהקשר זה :
פלסטיות ניכרת , מערכת אקסיטטורית בצד אינהיביטורית, אפיון מורפולוגי מפורט, גישה נוחה
פוץ לחקר פיזיולוגיה יתרונות אלו הביאו לכך שסינפסה זו הפכה למודל נ. ואפשרויות מודולציה רבות
. סינפטית
מתפקד באופן שונה לחלוטין מתא שריר משורטט באדם וביונקים רבים Crayfish -תא השריר ב
אלא מערכת הפועלת , הפועלת באופן של הכל או לא כלום, זו איננה מערכת בעלת סף. אחרים
עצמת . מוטורנוירון יחיד מקבלים עצבוב באמצעותopener -כל סיבי שריר ה. רציף, באופן מונוטוני
בכל , ותלויה באופן ישיר במידת הדפולריזציה על גבי הממברנה, הכיווץ של כל סיב היא הדרגתית
,Bittner, 1968; Atwood, 1976; Atwood & Wojtowicz, 1986; Araque & Buno(רגע נתון
1995; Araque et al., 1998( ,כווץ של השריר מחייבת שליטה רציפה מכאן שהבקרה על עצמת ה
שליטה זו מושגת הודות לשני מנגנונים יעילים הפועלים . על המתח הפוסט סינפטי בתאי השריר
. באופן מנוגד זה לזה
הראשון הוא הטרמינל האקסיטטורי המהווה מנגנון דפולריזטורי יעיל הודות ליכולתו להגביר
טווח השחרור . ודל בהתאם לתדר ההפעלה שלוהדרגתית את השחרור הסינפטי בשני סדרי ג
מכווץ טוני ממושך ועד להפעלת , הסינפטי הרחב מאפשר לסרטן להפעיל טווח כוח רחב בהתאמה
. כוח מהיר וחזק הדרוש לביצוע תנועות מהירות
הדומה בתכונותיו המורפולוגיות והפיזיולוגיות , המנגנון המנוגד לו הוא הטרמינל האינהיביטורי
ישירות על תא : ומספק דיכוי יעיל של כווץ השריר הודות לפעולתו הכפולה, ל האקסיטטורילטרמינ
עצמת . ועל האקסון והטרמינלים האקסיטטוריים הפרסינפטיים מאידך, השריר הפוסטסינפטי מחד
ירי בתדרים נמוכים מאפשר . הדיכוי היא פונקציה של תדר הירי של המוטורנוירון האינהיביטורי
, ירי בתדרים גבוהים מנטרל את מתח השריר וגורם להרפיה. טה טובה יותר בכוח השרירלסרטן שלי

4 מבוא_________________________________________________________________________________
,Atwood & Kwan(בזמן סגירה מהירה של הצבת , לדוגמא לצורך הפעלת השריר האנטגוניסטי
1976( .
בעבודה זו , למרות שתדר הגירוי של הטרמינל אינו הגורם היחידי בקביעת עוצמת השחרור הסינפטי
בהתאם לבחירה זו אופיינה במסגרת העבודה התלות הרציפה בין תדר . הוא נבחר כגורם העיקרי
תוך ציון הסייגים , הגירוי לעצמת שחרור הנוירוטרנסמיטור האקסיטטורי בטרמינלים שונים
יני המשפיעים על מאפי) פיזיולוגיים ואחרים(בהמשך הופעלו גורמים שונים . הרלוונטיים לתלות זו
ל היא האינהיביציה "אחד מהגורמים הנ. תוך תאור האופן בו באות השפעות אלה לידי ביטוי, התלות
הפרסינפטית וזאת מתוך מטרה ללמוד יותר על הדינמיקה בין שחרור הנוירוטרנסמיטור
. האקסיטטורי לפעולת האינהיביציה בתדרי הגירוי השונים
Crayfish - של הopener - בשריר הNMJסינפסת .1.2, כמודל לחקר תהליכים סינפטייםCrayfish - של הopener - בשריר הNMJיתרונות סינפסת
חוקרים שנתקלו בשאלה . הביאו להצטברות נתונים רבים בנוגע לקשת רחבה של תהליכים אלה
פיזיולוגית לא פתורה פנו לעיתים קרובות למצוא רמזים לתשובה מתוך מחקר מורפולוגי של
ממצאים מורפולוגיים הולידו שאלות פיזיולוגיות רבות שנחקרו בכלים , להפךואף , הפרפרט
.לפיכך הקשר בין המבנה לתפקוד נמשך כחוט השני לאורך מרבית הפרקים בעבודה זו. המתאימים
מורפולוגיה ופיזיולוגיה כללית של הסינפסה .1.2.1
הציפורן הנעה ממוקם במקטע הדיסטלי של רגל ההליכה ואחראי על פתיחתopener -שריר ה
)Atwood, 1982; Govind, 1982( (fig.1A,B)שבקצה הרגל , בצבת
אקסיטטורי : י שני אקסונים הצמודים זה לזה לכל אורך מהלכם על פני השריר"השריר מעוצבב ע
)Common excitor ( המשחררGlutamate , ואינהיביטורי)Specific inhibitor( , המשחררGABA
(Gamma-aminobutyric acid) . קיים אקסון אינהיביטורי נוסף)Common inhibitor( , המעצבב
-אולם הוא מעצבב רק חלק בלבד מהתאים הפרוקסימליים ביותר בשריר ה, את כל שרירי הגפה
opener . סידור מבני זה מאפשר לסרטן לדכא ספציפית את שריר הopener תוך הפעלת האקסון
לדוגמא בזמן אכילה (ניע את שרירי הגפה בחופשיות גם כאשר הצבת סגורה בכוח וכך לה, הספציפי
ל מונחים על פני השריר ומתפצלים לראשונה יחדיו "שני האקסונים הנ, כאמור לעיל). או לחימה

5 מבוא_________________________________________________________________________________
Figure 1 : תרשים הפרפרט
A :שריר ה. תרשים רגל ההליכה הראשונה של הסרטן- opener בצד של הצבת , הרגל נמצא במקטע הדיסטלי של
הקו . כך שכווץ השריר מביא לפתיחתה, הגיד המרכזי של השריר מחובר לצד החיצוני של הצבת. (dactylopodite)הנעה
כפי שנחשף propodite -מקטע ה: B. המקווקו מציין את מישור החתך בו מבוצעת הדיסקציה לצורך חשיפת השריר
האקסיטטורי : שני האקסונים המעצבבים אותו ואת , האזורים השונים בוניתן לראות את השריר על. לאחר הדיסקציה
ולא ניתן להפריד ביניהם בעין , שני האקסונים צמודים זה לזה לאורך כל מהלכם. (specific inhibitor)והאינהיביטורי
.בלתי מזוינת
, השרירלאחר מספר פיצולים האקסון נצמד לתאי. ובהמשך לפיצולי משנה נוספים, Yבצורת
;varicosities ()Florey & Cahill, 1982(והטרמינלים ממוקמים לאורכו בצורת נקודות התרחבות
Atwood et al., 1984( . בכל טרמינל נמצאו מספר סינפסות)הממוקמות ) עשרות בודדות בממוצע
כ אתרי שחרור סינפטי "דובכל סינפסה ניתן למצוא ב, בנקודות המגע בין הטרמינל לסיב השריר
השוואה מורפולוגית בין הטרמינלים ).Dense bodies - כEM -מזוהים מורפולוגית ב(בודדים
אולם מספר , בכל אזור בשריר, האקסיטטוריים לאינהיביטוריים מראה שקיים איזון במספרם בכל תא
.נל אינהיביטוריהסינפסות בטרמינל אקסיטטורי גדול יותר משמעותית ממספר הסינפסות בטרמי
התמיינות שונה של טרמינלים מאקסון יחיד .1.2.2
;Atwood, 1967; Bittner, 1968(על בסיס עדויות פיזיולוגיות שהצטברו במספר עבודות
Atwood & Bittner, 1971; Parnas et al., 1982c; Thompson & Atwood, 1984; Cooper et

6 מבוא_________________________________________________________________________________
al., 1995a; Cooper et al., 1996a; Crider & Cooper, 1999, 2000( , חולקו הטרמינלים לשתי
, הקבוצה הראשונה כוללת את הטרמינלים הפרוקסימליים. קבוצות עיקריות בעלות מאפיינים נבדלים
הממוקמים במרכז , )central(והשניה את המרכזיים , הממוקמים בחלק הקרוב יותר של השריר
ובעלות תכונות האופייניות ) High output" (חזקות"קסימליות הן הסינפסות הפרו. (fig.1B)השריר
תא עצב היורה צרורות קצרים בתדר גבוה לצורך יצירת (, Phasic motorneuron -לסינפסות של
בעלות תכונות האופייניות ) Low-output" (חלשות"לעומתן הסינפסות המרכזיות הן ). תנועה חדה
התמיינות שונה ). א היורה בתדר נמוך למשך כל תנועת הרגלת (Tonic motor neuron -לאלו של
, )Atwood, 1967(של טרמינלים שמקורם מאקסון יחיד היא תופעה מוכרת מזמן בחסרי חוליות
Thomson. ותוארה גם במערכת עצבים מרכזית של יונקים, )Davis & Murphey, 1994(צרצר
)Thomson et al., 1993; Thomson, 1997( תארה פסיליטציה בקשרים בין תאים פירמידליים
בקשרים בינם לבין תאים (depression)בד בבד עם הנחתה , לאינטרנוירונים בקורטקס של חולדה
תוך רשום בו , ומה צפו תופעה ד)Markram et al. )Markram et al., 1998. פירמידליים אחרים
לדעת , תופעה זו תורמת. ושני התאים הפוסטסינפטיים, זמני מהתא הפירמידלי הפרסינפטי
ליצירת מגוון רחב של אפשרויות להעברה סינפטית של סדרת פוטנציאלי פעולה באקסון , החוקרים
ות של התמיינות שונה של טרמינלים מאקסון יחיד תוארה גם במערכות עצבים מרכזי. הפרסינפטי
.)Korn et al., 1986( ונוירונים בדג זהב )Pierce & Mendell, 1993(חתול
הניסויים שנערכו בעבודה זו , opener -מאחר וקיימת התמיינות ברורה בין הטרמינלים בשריר ה
הבנה מלאה של תוצאות . והתוצאות הושוו ביניהן, נערכו בנפרד על שתי אוכלוסיות הטרמינלים
. המובא להלן, השוואה זו דורשת רקע מפורט יותר בנוגע להבדלים בין הטרמינלים
הבדלים מורפולוגיים בין הטרמינלים .1.2.3
הממצא הבולט ביותר שהסב את תשומת לב החוקרים להבדל בין טרמינלים משני אזורי השריר
דולה בסדר גודל בקירוב באזור הג, הוא ההבדל הברור במשרעת התגובות הפוסטסינפטיות
,.Atwood, 1967; Bittner, 1968; Atwood & Bittner, 1971; Parnas et al(הפרוקסימלי
1982c; Thompson & Atwood, 1984( תופעה נוספת שתוארה באזור המרכזי היא החלשות
הסעיף העיקרי של האקסון ככל שמיקום הטרמינל הולך ומתרחק לטרלית מ, התגובה הסינפטית
הבדלים פיזיולוגיים ברורים אלה הפנו חוקרים . )Cooper et al., 1996a(המגיע לתא השריר
בעבודות אלה נערכה השוואה מורפולוגית . לחפש אחר סיבות מורפולוגיות העשויות לעמוד בבסיסם

7 מבוא_________________________________________________________________________________
,.Wojtowicz et al., 1994; Cooper et al(שיטתית בין טרמינלים מהאזורים השונים של השריר
1995a; Cooper et al., 1996a( . תוצאות המחקרים מראות שבסיבים המרכזיים יש בממוצע יותר
כמו כן , כשהגדולים מביניהם ממוקמים בקרבת סעיף האקסון העיקרי, )50% -ב(טרמינלים לסיב
נתונים אלה אינם מתיישבים עם . ט גדול יותרמספר הסינפסות הממוצע לטרמינל מרכזי מע
מאידך נמצא שבטרמינלים הפרוקסימליים יש . התגובות הסינפטיות החלשות יותר בסיבים אלה
ממוצע גבוה זה נובע מהמצאות יותר סינפסות בעלות שני . בממוצע כפליים אתרי שחרור לסינפסה
. אתרי שחרור ויותר באזור זה
מורפולוגית להפרדה הפיזיולוגית שנעשתה בין הטרמינלים בשני ממצאים אלה מספקים תמיכה
אולם עשויים להסביר באופן חלקי בלבד את השחרור הסינפטי המוגבר יחסית , אזורי השריר
מסיבה זו הופנה המחקר לחיפוש אחר הבדלים פיזיולוגיים העשויים . מהטרמינלים הפרוקסימליים
.לעמוד בבסיס התופעה
ים בין הטרמינלים הבדלים פיזיולוגי .1.2.4
פיזיולוגית ניתן להסביר את השחרור הסינפטי המוגבר בטרמינלים הפרוקסימליים על ידי אחת
, הראשונה היא פלישה יעילה יותר של פוטנציאל הפעולה לטרמינלים. או שתיים מאפשרויות הבאות
בדל בתכונות אפשרות שניה היא ה. ושחרור סינפטי מוגבר בהתאם, הגוררת כניסת סידן מוגברת
כך שפרופיל דפולריזציה זהה בשני סוגי הטרמינלים יגרור שחרור , הפרסינפטיות של הטרמינלים
.סינפטי מוגבר בטרמינלים הפרוקסימליים
המלווה , פלישה יעילה יותר לטרמינלים הפרוקסימליים: פלישת פוטנציאל פעולה לטרמינל
;Atwood, 1967(ית את ההבדל בין הטרמינלים עשויה להסביר פיזיולוג, בדפולריזציה חזקה יותר
Bittner, 1968; Atwood et al., 1969; Atwood & Bittner, 1971; Sherman & Atwood,
1972; Dudel, 1983 ,אך ראה :Zucker, 1974 #36( . למרות שלפלישה כזו קיימות עדויות
מאידך . ורמת להבדל בין תגובות הטרמינליםרוב החוקרים סבורים שהיא אכן ת, עקיפות בלבד
Parnas et(ולכן יש מקום לחפש גורמים נוספים , משקל תרומה זו לא ניתן לכימות מסיבות טכניות
al., 1982c(בין השאר הבדלים בתכונות הפרסינפטיות של שני סוגי הטרמינלים .
ה בין תכונות פרסינפטיות בטרמינלים שונים היא אחת הדרכים לערוך השווא: אנליזה קואנטלית
להתאים את מאפייני השחרור הסינפטי מהטרמינל למודל כמותי ברור ואחיד המאפשר השוואה בין
המודל המקובל ביותר בחקר תהליכים סינפטיים הוא המודל . ערכי הפרמטרים עליהם נשען המודל

8 מבוא_________________________________________________________________________________
. זה ומספר דוגמאות לשימוש שנעשה בוועל כן מובאת להלן סקירה קצרה של מודל, הקואנטלי
גורסת שהתגובה )Fatt & Katz, 1952; Del Castilo & Katz, 1954(ההיפותזה הקואנטלית
הזהה , )קואנטום(הסינפטית המעוררת מורכבת מיחידות להן גודל מינימלי הנע סביב ערך קבוע
היפותזה זו הניחה . ררות בצורה של הכל או כלוםיחידות אלה משוח. לגודל האירועים הספונטניים
על בסיס מודל סטטיסטי שנבחר כמיצג באופן הנאמן , בסיס לניתוח מתמטי של התגובה הסינפטית
מתוך המודל הסטטיסטי המתאים ביותר חולצו ערכם המספרי של . ביותר את תוצאות הניסוי
כוחה של אנליזה זו נובע . אנטליתניתוח זה נקרא אנליזה קו. הפרמטרים עליהם מושתת המודל
מיצג את ההסתברות לשחרור pלדוגמא , מהאפשרות לכמת פרמטרים בעלי מקבילה פיזיולוגית
במודל ( מייצג את מספר אתרי השחרור או הסינפסות הפעילות בטרמינל n -ו , קואנטום בכל אתר
, כלל הסינפסות בטרמינל היא תת קבוצה של(n)אוכלוסיית סינפסות פעילות זו ). הבינומי להלן
, אחד המודלים המתמטיים המוכרים יותר. הניתנת להערכה מורפולוגית כפי שצוין קודם לכן
מודל זה נשען על הנחת יסוד . המשמשים לביצוע אנליזה קואנטלית הוא המודל הבינומי הפשוט
ם מטילה הנחה זו מפשטת את המודל אול. זהה בין כל אתרי השחרור(p)שההסתברות לשחרור
. מאחר ואינה מוצדקת בהכרח פיזיולוגית, מגבלה חמורה על התאמתו לתוצאות ניסיוניות
. Pi - קואנטה iבהתאם למודל הבינומי ניתן לחשב בנפרד את ההסתברויות לקבל תגובה המכילה
כלומר תגובה מקסימלית הנובעת , n -ל, כלומר כשלון בשחרור, 0נע בין iמספר הקואנטה
. אתרי השחרורnזמני מכלל -רור בומשח
מחושבתm -התכולה הקואנטלית הממוצעת
, תוך סיכום מכפלות כלל ההסתברויות השונות
. בתכולה הקואנטלית המתאימה לכל הסתברות
המתרחש עקב ביצוע , האנליזה הקואנטלית היא כלי מקובל לכימות שינוי בתכונות הפרסינפטיות
בסרטנים נעשה בו שימוש נרחב לצורך אפיון שינויים החלים . בטרמינלמניפולציה זו או אחרת
אינהיביציה , בשחרור סינפטי מהטרמינל בעקבות הפעלת תהליכים שונים כמו פסיליטציה
,Dudel & Kuffler(ועוד , שינויי טמפרטורה, עליה בלחץ היפרברי, שינוי ריכוזי סידן, פרסינפטית
1961; Atwood et al., 1969; Bittner & Harrison, 1970; Johnson & Wernig, 1971;
( )( )ini ppini
niP −
= −−
1!
!
∑=
⋅=n
ii nPm
0

9 מבוא_________________________________________________________________________________
Wernig, 1972; Zucker, 1973; Wojtowicz & Atwood, 1986; Wojtowicz et al., 1988;
Golan et al., 1994; Cooper et al., 1995b; Strawn & Cooper, 2002; Viele et al., 2003(.
הטרמינלים הפרוקסימליים מול המרכזיים בוצעה אנליזה במטרה לכמת את ההבדל בתכונות
יחד עם ניתוח מורפולוגי של אותם אתרים , קואנטלית השוואתית בין שני סוגי טרמינלים אלה
)Wojtowicz et al., 1994; Cooper et al., 1995b; Cooper et al., 1996a( . ממצא חשוב של
אך על ההסתברות לשחרור , דומה בשניהםn)( סינפסות לטרמינל מחקרים אלה מעיד על ממוצע
(p)פירוט הבדלים אלה מופיע להלן. גבוהה יותר בסינפסות הפרוקסימליות.
כאמור לעיל המספר המחושב של הסינפסות הפעילות בטרמינל : (n)ממוצע הסינפסות לטרמינל
הוא חוסר התאמה בולט בין מספר אולם ממצא עקבי נוסף שעלה , היה דומה בין שני הטרמינלים
, המספר המורפולוגי. לבין מספר הסינפסות שנספרו מורפולוגית בטרמינל, הסינפסות הפעילות
). כ" בדn < 5(היה גבוה יותר בסדר גודל ממספר הסינפסות הפעילות , כ עשרות בודדות"בד
בית את הסינפסות מפעיל סלקטי, בהם נערך הניסוי, החוקרים משערים שגירוי בתדרים נמוכים
,.Cooper et al(העשירות יחסית באתרי שחרור ובהתאם בעלות הסתברות גבוהה לשחרור
1996b( סינפסות אלה מהוות את nאולם מהוות רק , הסינפסות הנספרות באנליזה הקואנטלית
יתכן , והים יותרבתדרי גירוי גב. ומכאן חוסר ההתאמה המספרי, חלק מכלל הסינפסות בטרמינל
. ותורמות להגברת השחרור הסינפטי, "מגויסות"והסינפסות הדלות באתרי שחרור
הסיבה להסתברות לשחרור הגבוהה יחסית בטרמינלים הפרוקסימליים : (p)הסתברות לשחרור
יתכן והיא נובעת מחפיפה מרחבית בכניסת הסידן לאתרים פרסינפטיים סמוכים . טרם ברורה
דבר המגביר את ההסתברות לשחרור , שריכוז הסידן מגיע לערכים גבוהים יותרכך, בטרמינל
)Cooper et al., 1996b( .סיבות אפשריות נוספות לכך מובאות בהמשך המבוא.
ממצאי האנליזה הקואנטלית מסבירים באופן חלקי בלבד את השחרור הסינפטי המוגבר מהסינפסות
הניסיון לאתר מקור מורפולוגי לעלייה הניכרת . רות יותר באתרי שחרורהעשי, הפרוקסימליות
והפנה חוקרים לחפש אחר סיבות פיזיולוגיות , בהסתברות לשחרור לא הניב תוצאות מספקות
. אחרות לתופעה
ההבדלים בהסתברות לשחרור : הבדלים פיזיולוגיים נוספים הקשורים בריכוז הסידן בטרמינלים
. השונים הפנתה את החוקרים לחיפוש הבדלים ברמות הסידן התוך תאי ביניהםבין הטרמינלים
העשוי להיות גבוה יותר בטרמינלים , אפשרות אחת נוגעת לרמות הסידן התוך תאי במנוחה

10 מבוא_________________________________________________________________________________
מתוך בנייה )Parnas et al. )Parnas et al., 1982cאפשרות זו הועלתה על ידי . הפרוקסימליים
ל טרם נמצא ביסוס בעבודות שונות שבדקו בין השאר השערה זו "לאפשרות הנ. תיאורטישל מודל
(Cooper et al., 1995a) .
בין אם הודות לפלישה סמוכה , אפשרות שניה היא כניסת סידן חזקה יותר לטרמינל בתגובה לגירוי
יית סידן בשני סוגי הטרמינלים הדמ. או לריבוי של תעלות סידן, יותר של פוטנציאל הפעולה
הדגימה עליה חדה יותר בריכוז , )Calcium green 2, 5N(באמצעות אינדיקטורים פלואורצנטיים
אין בכוחו של .)Cooper et al., 1995a(הסידן בטרמינלים הפרוקסימליים עם הגברת תדר הגירוי
או , וז גבוה יותר של תעלות סידן בטרמינלים אלהלמשל ריכ, ממצא זה לבודד גורם יחיד לתופעה
ממצא דומה . דפולריזציה ניכרת יותר הודות לפלישה יעילה יותר של פוטנציאל הפעולה לטרמינל
). Main leg extensor muscle(התקבל מניסוי מעט יסודי יותר שנערך באותו פרפרט בשריר אחר
האחד פאזי והשני , ורי משני מוטורנוירוניםשריר זה שונה מהותית בכך שמקבל עיצבוב אקסיטט
Msghina et(גירוי בתדר גבוה גרם לעליה חדה פי חמש בריכוז הסידן בטרמינלים הפאזיים . טוני
al., 1999( . השוואת השחרור הסינפטי בין הטרמינלים השונים בריכוזי סידן תוך תאיים זהים
עובדה , לים הפאזיים עדיין הייתה גבוהה יותר משמעותיתהראתה שהתכולה הקואנטלית בטרמינ
. המצביעה לדעת החוקרים על רגישות מוגברת של מנגנון השחרור לסידן בטרמינלים הפאזיים
עבודות אלה מצביעות בבירור על הצטברות מהירה יותר של סידן בטרמינלים הפאזיים בתגובה
. התופעהאולם לא עומדות על הגורם שבבסיס, לגירוי ממושך
אפשרות נוספת היא שהצטברות מואצת זו נובעת מהבדל ביעילות המנגנונים האחראים על פינוי
הבדלים כאלה נרשמו בין טרמינלים . הסידן מציטופלסמת הטרמינל במהלך גירויים מתמשכים
& Crayfish (Fengler -פאזיים וטוניים מתרביות שנלקחו מהשריר הסוגר את הצבת הגדולה ב
Lnenicka, 2002) .6 -בעבודה זו הסידן הצטבר בטרמינלים הפאזיים במהירות הגבוהה פי
ממצאי העבודה גם הצביעו . למרות כניסת סידן דומה מאוד בשניהם, בהשוואה לטרמינלים הטוניים
יים גירוי טרמינלים פאז. קצב פינוי הסידןקביעת על תדר הירי כעל גורם בעל השפעה מכרעת על
הביא להאצה משמעותית ) פיזיולוגית מתאפיינים בתדר ירי נמוך מאוד בפרפרט זה(בתדר גבוה
עדות מורפולוגית התומכת בקיום הבדלים . בדומה לזה של הטרמינלים הטוניים, בקצב פינוי הסידן
ראי איבר תוך תאי האח, בקצב פינוי הסידן נרשמה בדמות ריבוי המיטוכונדריות בטרמינלים הטוניים
. בין השאר על קליטת הסידן מציטופלסמת הטרמינל במהלך סדרות גירויים ארוכות

11 מבוא_________________________________________________________________________________
בעיקר כיוון , היא בעייתית) opener -שריר ה(השלכת ממצאים אלה על מערכת הנוכחית
ולפיכך תדר הגירוי של כלל , נובעים מאקסון יחידopener -שהטרמינלים השונים בשריר ה
בנוסף לכך אין עדויות להבדלים משמעותיים בצפיפות המיטוכונדריות . הטרמינלים בשריר הוא זהה
. opener -בין טרמינלים מאזורים שונים של שריר ה
התגובות הסינפטיות הנמדדות בתגובה לירי המוטורנוירון היחיד המעצבב את השריר : סיכום
יים קרובים יותר הטרמינלים הפרוקסימל. תלויות באופן ברור במיקום הסינפסה על גבי השריר
ומאופיינים בסינפסות עשירות יחסית באתרי שחרור , שמתפצל פחות באזור זה, לאקסון המרכזי
צירוף אלה מעניק לטרמינלים הפרוקסימליים תכולה קואנטלית . ובעלות הסתברות גבוהה לשחרור
ורמים לו ות, הגורמים להבדל בהסתברות לשחרור לא ברורים במלואם. גבוהה כבר בתדרים נמוכים
עליה חדה יותר בריכוז הסידן בטרמינל , כנראה פלישה יעילה יותר של פוטנציאל הפעולה לטרמינל
ויתכן שאף רגישות גבוהה יותר של מנגנון , )אולי גם הודות לפינוי סידן איטי יותר(בתגובה לגירוי
.השחרור לסידן
התגובה הסינפטית לגירוי בתדר גבוה .1.2.5
, opener -פולוגי מפורט של ארגון הטרמינלים השונים בשריר העד עתה ניתן תיאור מור
מאחר ויכולת . והסיבות האפשריות להבדלים אלה, ההבדלים הפיזיולוגיים בין תגובות הטרמינלים
ועבודה זו עוסקת , ל"הפוטנציאציה המרשימה היא אחת התכונות הבולטות של כל הטרמינלים הנ
.א להלן הרקע המדעי עליו מסתמכת העבודה בנושא זהמוב, בתגובת הטרמינלים לגירוי בתדר
לצורך הפעלת , בפרטcrayfish -טווח תדרי הירי של מוטורנוירונים אקסיטטוריים בחסרי חוליות וב
& Grossman et al., 1979; Crider(נע בין פעמים ספורות עד ליותר ממאה לשניה , השריר
Cooper, 1999, 2000( .ן שלצורך הבנת הפיזיולוגיה הסינפטית הקשורה בתגובה לגירוי מכא
אופי התגובה הסינפטית לגירוי בתדר שונה כמובן בין . בתדר יש להפעיל גירוי בטווח תדרים דומה
ובנוכחות מולקולות בעלות , במספר הגירויים, ותלוי בתדר הגירוי עצמו, פרפרטים ומערכות שונות
ניתן לחלק את השפעות הגירוי בתדר לשתי קבוצות באופן . השחרורהשפעה מדולטורית על מנגנון
עד שעות וימים לאחר הפסקת , השפעה הגורמת לשינוי תכונות פרסינפטיות לאורך דקות רבות: גס
והשפעות בהן שינוי התכונות הסינפטיות מוגבל , LTP (Long Term Potentiation)כדוגמת , הגירוי
Fcilitation, Potentiation, Augmentation, Postכמו , ל היותרעד דקות בודדות לכ, לזמן קצר
tetanic potentiation . הקבוצה האחרונה מכונה לרובShort term synaptic plasticity , וקיימת

12 מבוא_________________________________________________________________________________
Fisher et(באופן שמור יחסית במערכות עצבים פריפריות ומרכזיות במינים שונים של בעלי חיים
al., 1997; Zucker & Regehr, 2002( . המאפיין קבוצה זו הם שינויים פרסינפטיים הקשורים
ואינם תלויים בפעולת נוירומודולטורים או בשינוי תכונות , בעליה בריכוז וקישור סידן בטרמינל
המאופיינת בשינויים פרסינפטיים בעלי טווח זמן קצר , עבודה זו עוסקת בעיקר בפוטנציאציה. תעלות
. זוNMJהאופיינית למערכת ,בתגובה לסדרת גירויים קצרה יחסית , ) בודדות msאות עד מ(
הגברת : התופעה הפרסינפטית הראשונה שנחקרה בהקשר זה היא הפסיליטציה: פסיליטציה
,Del Castilo & Katz(השחרור הסינפטי בתגובה לגירוי השני מתוך צמד גירויים סמוכים בזמן
כלומר הגברת השחרור הסינפטי , פוטנציאציה היא הרחבה של התופעה על ציר הגירויים. )1954
. בתגובה לשלושה גירויים סמוכים בזמן ויותר
בהצטברות סידן בתוך האתר בכל הסינפסות שנחקרו בפרפרטים השונים הפסיליטציה קשורה
,Katz & Miledi(" דן השאריתיתיאוריית הסי"ומכאן השם , הפרסינפטי הודות לסמיכות הגירויים
בנוסף לאמור לעיל .ל להגברת השחרור שנוי במחלוקת"האופן המדויק בו פועל הסידן הנ. )1968
אוגמנטציה ועליה בתדר , הוכחה מעורבות ישירה של הסידן גם בתהליכים נוספים כמו פוטנציאציה
,Rosenthal, 1969; Miledi & Thies, 1971; Weinreich(אירועים ספונטניים לאחר הגירוי
1971; Kamiya & Zucker, 1994; Felmy et al., 2003(.
יש לציין שקיימים הבדלים רבים במאפייני הקשר בין עליית ריכוז הסידן והתפתחות , עם זאת
י המורפולוגי והפיזיולוגי הבדלים אלה נגזרים בין השאר מהשונ. הפסיליטציה בין הסינפסות השונות
הסקירה מתמקדת , אור אחיד של העומד בבסיס התופעהימאחר ולא ניתן לספק ת. הניכר ביניהן
בתיאור הפסיליטציה על סמך עבודות שנעשו בטרמינלים זהים לאלה שנחקרו בעבודה בעיקר
.הנוכחית
ריכוז לאסי הקושר בין התיאוריה הקלאסית בנוגע למעורבות סידן בפסיליטציה משולבת במודל הק
ל "הקשר הנלפי מודל זה . )Dodge & Rahamimoff, 1967( לשחרור הסינפטי הסידן החוץ תאי
יש לציין כי אחד או יותר מהתהליכים החל מכניסת הסידן ועד (בעל ערך של ארבע, קואופרטיביהוא
ואופרטיביות ייחסו את הקהכותבים ). זהשחרור עצמו עשויים לתרום לערךקשירתו למנגנון הל
מתרחשת הפסיליטציה לפי המודל הקלאסי. הסינפטי השחרורמנגנוןל ארבעה יוני סידן ה שלקשירל
. עם הגעת הגירוישחרורהארבעה אתרי מנותר קשור לאחד או יותרעדיין כאשר הסידן השאריתי
בהמשך הצטברו . תתרחש פסיליטציה, מעל לריכוז מסוים, מכאן שכל עוד קיים סידן חופשי בטרמינל

13 מבוא_________________________________________________________________________________
חלק מעדויות אלה מבוססות על הדמיית ריכוז סידן . עדויות רבות המפריכות את המודל הקלאסי
;Delaney et al., 1989; Delaney & Tank, 1994(, ומודלים מתמטיים להתפשטותו בטרמינל
Winslow et al., 1994( וחלקן מתוך תוצאות ניסויים המדגימים עמידות מרשימה של פסיליטציה
ביחס לרגישותו הרבה של מנגנון השחרור הסינפטי למניפולציות דומות , לירידה בריכוז סידן
)Balnave & Gage, 1974; Younkin, 1974; Bittner & Schatz, 1981; Dudel et al., 1982;
Stanley, 1986; Blundon et al., 1993; Winslow et al., 1994; Liu & Stanley, 1995;
Wright et al., 1996a; Vyshedskiy & Lin, 1997; Vyshedskiy et al., 2000( תוצאות אלה
ה להתקיים גם בשעה שהסידן השאריתי חוזר לרמות הצביעו על כך שהפסיליטציה ממשיכ
עדות זאת הובילה להנחה שבבסיס הפסיליטציה עומד סידן שנותר קשור . ואף למטה מהן, הביקורת
& Delaney(לאתר או אתרים בעלי זיקה גבוהה יותר מאלו המביאים לשחרור הסינפטי עצמו
Tank, 1991; Hochner et al., 1991; Van der Kloot & Molgo, 1993; Kamiya & Zucker,
1994; Ravin et al., 1999; Tang et al., 2000( . עדויות אלה הביאו להחלפת המודל הקלאסי
אולם הדעה חלוקה בנוגע , תוצאות הניסויים וההדמיותמ התואמים חלק ניכר יותרבמודלים חדשים
באופן כללי קיימות שתי אפשרויות . האחראים לפסיליטציה, הקישור אלהלטיבם ומיקומם של אתרי
: לשלבם במודל חדש
השחרור הסינפטי : אפשרות ראשונה מניחה קיום של אתר בודד האחראי לשני התהליכים
עם צירוף הנחות יסוד נוספות בדבר זיקה , אפשרות זו דומה יותר למודל הקלאסי. והפסיליטציה
האתרים . וקבועי קשירה והתנתקות שונים, אחד מארבעת אתרי הקישורשונה לסידן של כל
עם קבועי , האחראים לפסיליטציה יהיו אלה בעלי הזיקה הגבוהה יותר לסידן מתוך הארבעה
;Stanley, 1986; Segev, 1990(התואמים את קבועי דעיכת הפסיליטציה , התנתקות ממושכים
Liu & Stanley, 1995; Bertram et al., 1996( . עשויה מקסימלית סיליטציהפ לפי מודל זה
,Stanley(כן קיימות עדויות לכך וא, היא מינימליתהשחרור הסידן לאתר להתרחש כאשר קשירת
1986; Wright et al., 1996a; Vyshedskiy & Lin, 1997( . מודל דומה מציע אפשרות בה
באופן אנלוגי לקשירת , קשירת סידן לאתר בעל זיקה גבוהה מגביר את הזיקה של האתרים הנותרים
עבודה שנעשתהלציין שיש .)Vyshedskiy et al., 2000(י מולקולת ההמוגלובין "חמצן ע
Felmy et( תומכת באפשרות זולא בחולדה(Calyx of Held)מערכת עצבים מרכזית בטרמינלים ב
al., 2003( . ולכן מנגנון השחרור לסידן בזמן פסיליטציה כל שינוי ברגישות הודגםלא ל"הנבעבודה

14 מבוא_________________________________________________________________________________
םלטענת. גנון פסיליטטורי המבוסס על סידן שאריתי הקשור למנגנון השחרורהכותבים שוללים מנ
. דן בסביבה הסמוכה לאתרי השחרורליניארי של הסי-הפסיליטציה נגרמת מסיכום סופרהעיקר
אולם בהתחשב בכך , מהתוצאות שהוצגו קודם לכןרבותלכאורה תוצאות עבודה זו סותרות
שחרור סידן תוך תאי :תוך שימוש בשיטות גירוי שונות( מפרפרט שונהשתוצאות אלה הופקו
(caged)תר לראות הבדלים אלה אפשרי ונכון יו, ) גירוי דפולריזטורימול אור בהבזקי גירוי" ע
.ל לזו של חסרי חוליות הנחקרת כאן"כמייצגים את השוני המהותי בין הסינפסה המרכזית הנ
פסיליטטורי אתרומניחה קיום, יה מפרידה בין אתר השחרור לאתר הפסיליטציהיהאפשרות השנ
קיים לפחות לפי מודל זה. נפרד מרחבית ובעל רגישות גבוהה יותר לסידן בהשוואה לאתר השחרור
משמעותיתנמוכים סידן נחשף לריכוזי ה, המרוחק מאתר השחרור ומתעלות הסידן, אתר אחד נוסף
;Van der Kloot & Molgo, 1993; Tang et al., 2000( מאלה להם נחשף אתר השחרור
Matveev et al., 2002(. אתר השחרור נחשף לריכוזי שהגורסת עומדת הנחהאפשרות זו בבסיס
נעשו מספר עבודות בהן נמדד השנים האחרונותבמהלך .µMמאות בודדות של עד סידן של
.crayfish (Ravin et al - חלקן ב, בו זמנית עם מדידת ריכוז הסידן בטרמינל הסינפטירורהשח
,.Bollmann et al) ,(calyx of Held) תמרכזיממערכת עצבים טרמינל רובן בו (1997,1999,
2000; Schneggenburger et al., 2000; Felmy et al., 2003 Meinrenken et al.,
של מנגנון השחרור לריכוז (10µM~)עבודות אלה ככלל הדגימו רגישות גבוהה .(2002,2003
, ובניגוד להערכות קודמות ,Aharon et al. ,(1994) בהסכמה עם המודל של ,הסידן התוך תאי
Simon and Llina´s,1985;Yamada and)בכסדר גודלרגישות נמוכה יותר שגרסו
Zucker,1992;Roberts,1994). טרם ברור כיצד ליישב את הסתירה בין עבודות אלה לקודמות ,
ריכוז הסידן המתפתח בסביבת אתר הקשירה של מנגנון השחרור תלוי במידה רבה ומאחר ו
יש לשער שהתוצאות השונות , (Meinrenken et al., 2002) במרחקו מתעלות הסידן הממברנליות
ובהתאם , שוניםערכים התפתח לריכוז הסידן לעשוי מצביעות על ספקטרום של סינפסות בהןלהלן
).Augustin GJ) 2001לצורך עיון ראה . רגישות מנגנון השחרור לסידן עשוייה להיות שונה, לכך
אנליזה קואנטלית של השחרור הסינפטי שנעשתה בביקורת ובמהלך פסיליטציה הדגימה עליה
,Dudel & Kuffler, 1961; Wernig, 1972; Zucker, 1973; Smith(או שניהם , n ,p -בפרמטרים
1983b; Viele et al., 2003(

15 מבוא_________________________________________________________________________________
, מדידת הפסיליטציה כפי שנעשתה באופן מסורתי :סיליטציה באוכלוסיות טרמינלים שונותפ
, סיפקה עדות חשובה בנוגע ליכולת של הטרמינל להגביר את השחרור הסינפטי בתגובה לגירוי נוסף
כחלק מהגדרת ההבדלים בין אוכלוסיות . מדד שלא ניתן היה להפיק ממאפייני התגובה הבודדת
שהגדלת המרווח הזמן בין זוג הגירויים )Linder )Linder TM, 1974טרמינלים מצא שונות של
Parnas etבהמשך מצאו . בהשוואה למרכזיים, מביא להגברה בפסיליטציה בטרמינלים הדיסטליים
al. )Parnas et al., 1982c(בטרמינלים הדיסטלייםבדמות פסיליטציה גבוהה יותר , תמיכה לכך ,
עבודות אלה הביאו להנחה שהטרמינלים . בשריר הפותח את הצבת, בהשוואה לפרוקסימליים
,Crider & Cooper )Crider & Cooper, 1999. המשחררים פחות נוטים לפסיליטציה מוגברת
מוש בשלושה נהלי גירוי תוך שי, בדקו סברה זו על הטרמינלים הפרוקסימליים והמרכזיים)2000
בעבודתם הראו שבכל נהלי הגירוי הפסיליטציה הייתה גבוהה יותר דווקא בטרמינלים . שונים
תוצאות אלה משאירות את סוגיית . בעלי התכולה הקואנטלית הגבוהה יותר, הפרוקסימליים
עקב ההשוואה בין תוצאות העבודות השונות מסובכת. הפסיליטציה בטרמינלים השונים פתוחה
.ושימוש בנהלי גירוי שונים , בחירה סובייקטיבית של טרמינלים
היא תופעה הקשורה באופן ברור לעליה בסידן Crayfish -פסיליטציה או פוטנציאציה ב: סיכום
הסידן נקשר לאתר או אתרים בעלי זיקה גבוהה וקינטיקת שחרור . תוך תאי בעקבות גירויים קודמים
אין עדויות ברורות בנוגע למיקומם ומספרם של אתרים . את השחרור הסינפטיומגביר , איטית יחסית
קיים עניין כללי רב בחקר . והאופן בו קשירת סידן לאתרים אלה מגבירה את השחרור הסינפטי, אלה
קיימות עדויות . ובעיקר בפרפרט זה, הפוטנציאציה הודות לחשיבות הפיזיולוגית המכרעת שלה
, פוטנציאציית התגובה הסינפטית בין טרמינלים באזורים שונים בשרירהמצביעות על הבדלים ב
.אולם אין תיאור כמותי ברור של הבדלים אלה ומשמעותם הפיזיולוגית
תפקידה של הדפולריזציה בשחרור הסינפטי .1.3הוא , הגורסת שגורם בודד כלומר העלאת ריכוז סידן תוך תאי בטרמינל, התיאוריה המקובלת
מתקשה להסביר חלק מהתופעות האופייניות , על מנת להביא לשחרור סינפטימספיק והכרחי
שריר בעכבר - הראו בסינפסת עצב)Datyner et al. )Datyner & Gage, 1980. לשחרור זה
שהפחתת ריכוז הסידן החוץ תאי למחצית לא השפיעה כלל על חלון הזמן בו מתרחש השחרור
ממצאים אלה עודדו חוקרים שונים לחפש אחר גורם נוסף מלבד . (release timecourse)הסינפטי

16 מבוא_________________________________________________________________________________
Parnas et al. )Parnas et. המעורב בקביעת חלון הזמן המאפיין את השחרור הסינפטי, הסידן
al., 1986; Parnas & Parnas, 1986( הציעו תיאוריה חדשה לפיה לדפולריזציה של ממברנת
והפעלת מולקולה ממברנלית , פתיחת מוליכות הסידן בטרמינל: תפקידים בלתי תלוייםהטרמינל שני
. (calcium-voltage hypothesis)רגישה למתח ההופכת את מנגנון השחרור הסינפטי לאקטיבי
וטרם הושג , ל ואחרות לא"חלקן תומכות בתיאוריה הנ, במשך השנים פורסמו עבודות שונות בנושא
בתנאי crayfish-בעבודות שנעשו בטרמינלים של ה. )Parnas et al., 2000a(ין זה קונצנזוס בעני
voltage-clamp ,ל"תנאים המאפשרים לבדוק את תקפות התיאוריה הנ ,)Wright et al., 1996a,
b(עבודות שנעשו בשנים האחרונות פורסמו מספר. לא נמצאו עדויות ברורות התומכות בה
בהן עוררו התגובות באמצעות שחרור סידן (Calyx of Held)בסינפסות במערכת עצבים מרכזית
& Bollmann et al., 2000; Schneggenburger(ללא דפולריזציה , ממולקולות תוך תאיות
Neher, 2000( .ה הציעו אולם לאחרונ, עבודות אלה הטילו ספק בתקפות תיאוריה זוParnas et
al. המופיע באופן , להתייחס לשחרור הסינפטי המעורר ללא דפולריזציה כאל שחרור ספונטני
י דפולריזציה "בניגוד לשחרור סינפטי המעורר ע, מסונכרן הודות לריכוז הסידן הגבוה בטרמינל
)Parnas et al., 2002( . הנגרמות הודות , צורות שונותהפרדה זו של השחרור הסינפטי לשתי
. ומותירה מקום למחקרים נוספים בנושא, מפחיתה מעט מחילוקי הדעות בעניין, לגירויים שונים
בעבודה הנוכחית לא נעשה שימוש בטכניקות גירוי המפרידות בין דפולריזציה לכניסת הסידן
.לדחות אותהל או "ולפיכך תוצאות העבודה לא מאפשרות לצדד בתיאוריה הנ, לטרמינל
מטרות המחקר והחידוש בהן .1.4
התגובה לגירוי בתדר בתנאי ביקורת
בחלק זה מתוארים מאפייני העקום המתאר את התלות בין השחרור הסינפטי האקסיטטורי
עד כה הודגמו שינויים ברורים בשחרור הסינפטי בין אזורים . באזורי השריר השונים, לתדר הגירוי
וב העבודות שנעשו עד כה הושווה השחרור הסינפטי תוך שימוש בנוהל אולם בר, שונים של השריר
שהופעל על טרמינלים שנבחרו , )Crider & Cooper, 1999, 2000(גירויים מצומצם באופן יחסי
החסרון בעבודות אלה הוא שאינן מספקות מידע בנוגע לאופן הרציף בו מתפתח . באופן סובייקטיבי
תלות רציפה זו . כתלות בספקטרום הפיזיולוגי הרחב של תדרי הגירוי, ינפטי מהטרמינלהשחרור הס
וההשוואה ביניהם מאפשרת להטיל אור על מנגנוני השחרור , שונה בין אזורים שונים בשריר

17 מבוא_________________________________________________________________________________
והאופן בו מתורגם תדר הירי של מוטורנוירון בודד להפעלה הדרגתית של השריר , הסינפטי בכלל
.כולו בפרט
ות שינויים בכניסת הסידן לטרמינל על התגובה לגירוי בתדרהשפע
בחלק זה נבחנת השפעת שינויים בכניסת הסידן לטרמינל על מאפייני התגובה לתדר של
וניתן , כניסת הסידן לתוך הטרמינל תופסת מקום מרכזי בתהליך השחרור הסינפטי. הטרמינל
או הפחתת ריכוז סידן חוץ , יר של הטרמינלכמו גירוי יש, להשפיע עליה באמצעים פשוטים יחסית
מטרת פרק זה היא לבחון אילו שינויים במאפייני התגובה לתדר של טרמינל נתון ניתן לייחס . תאי
ובאיזו מידה שינויים אלה עשויים להסביר את ההבדלים בין עקומי התגובה , לשינויים בכניסת הסידן
.לתדר של טרמינלים מאזורים שונים של השריר
השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר
המושרית באמצעות מתן , בחלק הראשון של פרק זה מתוארת השפעת אינהיביציה פרסינפטית .1
GABA ,ההשפעה . על המאפיינים השונים של התגובה לתדר כפי שתוארו בפרק הראשון
בהמשך הפרק נעשית . אקסוןתוך גירוי ה, נבחנת בנפרד על טרמינלים פרוקסימליים ומרכזיים
, על התגובה לתדר להשפעה של הפחתה בכניסת סידן לטרמינלGABAהשוואה בין השפעת
באופן נפרד GABAכמו כן נבחנת השפעת . במטרה להבין טוב יותר את פעולת האינהיביציה
מתוך מטרה ללמוד על של התגובה קצב התפתחות השחרורעל השחרור הסינפטי ועל עצמת ו
. התלות ביניהם
י גירוי האקסון האינהיביטורי על "בחלק זה מתוארת השפעת אינהיביציה פרסינפטית המעוררת ע .2
הפעלה מוקדמת של האקסון האינהיביטורי מנחיתה את השחרור . מאפייני התגובה לתדר
כלור בטרמינל והאקסון הודות להגדלת מוליכות לר יתהבין , הסינפטי מהטרמינל האקסיטטורי
בחלק זה של הפרק האינהיביציה הפרסינפטית מעוררת באמצעות גירוי בו זמני . הפרטרמינלי
באופן זה הגדלת המוליכות של הטרמינל . של הטרמינל האינהיביטורי עם האקסיטטורי
. ולכן יעילות האינהיביציה פוחתת, האקסיטטורי לא מקדימה את הדפולריזציה הפולשת אליו
רקאידך הפעלה זו מאפשרת לבחון את השפעתה של אינהיביציה פיזיולוגית שאיננה מבוססת מ
השפעת אינהיביציה זו . על התגובה לתדר של הטרמינלים , הקלאסיshunt -על מנגנון ה
. לעילGABAי "מושווית לזו המושרית ע

18 מבוא_________________________________________________________________________________
GABAי "היבטים מנגנוניים של האינהיביציה המתווכת ע
תוך שימוש בשתי שיטות גירוי , על השחרור הסינפטיGABAן נבחנת השפעת בחלק הראשו .1
שימוש בגירוי ישיר מאפשר שליטה טובה יותר במתח . גירוי אקסון וגירוי ישיר: שונות יחדיו
כך שהשוואת יעילות האינהיביציה תוך הפעלת שתי שיטות הגירוי , הממברנה של הטרמינל
יעיל להבנה טובה יותר של המנגנונים העומדים השונות על אותו טרמינל משמשת כלי
.בבסיסה
בחלק השני מאופיינת פרמקולוגית המוליכות המתווכת את האינהיביציה הפרסינפטית תוך .2
חלק זה כולל . GABAC, GABAA GABAB -שימוש בפרמקולוגיה ספציפית לרצפטורים מסוג
תוצאות אלה . הוות חידושואינן מ, גם חזרה על ניסויים שתוצאותיהם פורסמו זה מכבר
.מובאות בקצרה

19
שיטות .2___________________________________________________________________
פרפרט .2.1 במקטע הדיסטלי של רגל opener -שריר בשריר ה-כל הניסויים בעבודה זו נערכו בסינפסת עצב
נרכשו ) אורך בטן(מ " ס4-6חיות בגודל ). Crayfish) Procambarus clarkiiההליכה הראשונה של
הסרטנים הוחזקו באמבט מים . ב"ארה, לואיזיאנה.Atchafalaya biological supply Co -מ
והואכלו בפילה דג אחת , שעות12חושך של -תוך שמירה על מחזורי אור, מתוקים עם בעבוע אויר
. יםמי האמבט נוקו באמצעות הזרמה דרך מערכת מסננ. לשבוע
. )Atwood, 1982( על העיצבוב המוטורי שלו נחשף תוך ביצוע ניתוח מתאים opener -שריר ה
, אקסון אקסיטטורי: באופן זה נפרש השריר כשעליו מונחים האקסונים המוטוריים המעצבבים אותו
הועבר לאמבט הניסוי הפרפרט . specific inhibitor :fig.1B)(ובצמוד אליו אקסון אינהיביטורי
: mM -הרכב התמיסה ב ). Van Harreveld(בו הוזרמה תמיסה פיזיולוגית , ק" סמ1בנפח של
NaCl 200 ,KCl 5.4 ,CaCl2 13.5 ,MgCl2 2.6 ,Tris. HCl 10 , טמפרטורת חדר(22-24°C) ,
pH=7.4 .לרי של בוצע פיצוי מו, בניסויים בהם נעשה שימוש בתמיסות בעלות ריכוז סידן מופחת
חלק מסדרות הניסויים נערכו . בתמיסת הניסוי הוזרם אויר בבעבוע. NaCl -הריכוז תוך שימוש ב
.כפי שמצוין בהמשך, )17°C (בטמפרטורות נמוכות יותר
פרמקולוגיה .2.2כל החומרים הומסו במים לתוך התמיסה . Sigmaרוב החומרים הפרמקולוגיים נרכשו מחברת
זמן החלפת הנוזל באמבט . ק לשעה" סמ200 -בט במהירות זרימה של כוהוזרמו לאמ, הפיזיולוגית
דקות מתחילת ההזרמה הונח מצב של שווי משקל בין 10כעבור . שניות18אינו עולה לפיכך על
.שטיפת חומרים נעשתה במהירויות זרימה זהות. התמיסה והפרפרט

20 שיטות_________________________________________________________________________________
אלקטרופיזיולוגיה .2.3
אקסוןגירוי .2.3.1
קסון האקסיטטורי באלקטרודה ביפולארית העשויה מפיפטת זכוכית עם גורה הא אלהניסוייםב
. (fig.2A)שהונחה על גבי האקסון באזור הכניסה למקטע הדיסטלי , 20-30µmמפתח בקוטר של
האלקטרודה מפתח זהיר של מיקוםתוך האקסונים סלקטיבית משני באופן זה ניתן לגרות את אחד
שיטה זו מנצלת את ההבדלים הקיימים . של עצמת הגירויהגברה הדרגתית ו, בסמוך יותר אליו
באופן זה ניתן לגרות .ולפיכך בערך הסף שלהם לעירור, בהתנגדויות הכניסה בין שני האקסונים
הלאחר הגברוזאת , יחדיואו את שניהם, או האינהיביטוריאת האקסון האקסיטטורי סלקטיבית
של חמישה רצף. סף העירור של שני האקסוניםאתעבור ת כך ש, עצמת הגירוי נוספת שלהדרגתית
ראה לשם דוגמא (עורר תמיד תגובה פסיליטטורית אופיינית 100Hzבתר של םיי אקסיטטורםייגירו
fig.15Aם פרסינפטי בלבדהניב רישום של זר ו האינהיביטור לבד באופן דומה שלגירוי. ) משמאל-
זרם נוכחות ללא ,ENTC (End Nerve Terminal Current) או זרמי (AP)פוטנציאל הפעולה
קרבת פוטנציאל הממברנה לפוטנציאל ההיפוך של היעדר זרם פוסטסינפטי נובע מ .יפוסטסינפט
הגברת עצמת הגירוי .).fig -לא מוצג ב (עיקריתההאינהיביטורית פוסטסינפטית המוליכות ה, הכלור
ולהנחתה ברורה של הזרמים , נפטייםפרסייחדיו הביא לסיכום הזרמים הגירוי שני האקסונים ו
הונחה בהם ו, אלה נערכו בטרמינלים פרוקסימלייםניסויים .) מימיןfig.15Aראה (הפוסטסינפטיים
בו זמני מדויק של הולכה על מנת לאפשר תזמון ,פיפטת הגירוי בסמוך לאתר הרישום ככל הניתן
. םשני האקסוני

21 שיטות_________________________________________________________________________________
Figure 2 : מערך הניסוי ושיטת הרישום
A: כולל מיקום פיפטות הגירוי והרישום, מערך הניסוי ,A- Amplifier . בהגדלה תרשים פיפטת הרישום המונחת על גבי
דיאגרמת המעגל : B. פיפטהי ה"הנקלטים ע, (Ipre, Ipost)ים עם איור הזרמים הפרסינפטיים והפוסטסינפטי, הטרמינל
התנגדות האטימה -RS, התנגדות האלקטרודה-macropatch-clamp .RE -החשמלי הנוצר בזמן הרישום בשיטת ה
התנגדות פיסת -Rpatch, התנגדות וקיבול ממברנת תא השריר-Rm, Cm, (seal)הנוצרת בין האלקטרודה לממברנה
אחת , במהלך ניסויRSבחלק העליון דוגמא למדידת ערך : C. קוטר האלקטרודה-Ø . צמודה לאלקטרודההממברנה ה
ההסטוגרמה הסימטרית והצרה מעידה על . בחלק התחתון הסטוגרמת הערכים שנמדדו בחלק העליון. לשתי שניות
.רישום יציב

22 שיטות_________________________________________________________________________________
גירוי ישיר .2.3.2
מאפשרת גירוי ורישום בו )Dudel )Dudel, 1981י "מערכת הגירוי הישיר של הטרמינל שהוצגה ע
גירוי בזרם שלילי יוצר היפרפולריזציה בתוך האלקטרודה ביחס לנוזל החוץ . זמני מאלקטרודה יחידה
וכך מקרב את הפוטנציאל שמחוץ לפיסת ממברנת התא הצמודה לאלקטרודה לפוטנציאל , תאי
, גירוי זה. את הפרש הפוטנציאלים על פני הממברנהבאפן זה מצטמצם באופן מקומי. בתוך התא
, משפעל מרכיבי ממברנה התלויים במפל המתח על פניהם כמו תעלות סידן, בתלות בעצמתו למשכו
הגירוי הישיר מטבע הדברים אינו מושפע באופן מהותי מתכונות הממברנה אותה הוא . נתרן וכדומה
. י כמעט ללא הזרקת זרם דרך הממברנהמגרה מאחר והוא מסיט את הפוטנציאל החוץ תא
עות ובקבועי שרהארטיפקט הנוצר עקב הגירוי מאוזן באמצעות שני מעגלים המזרימים זרם במ
ל מבטל את חלק "הכוונון נעשה כך שהזרם הנ. דעיכה אקספוננציאליים הניתנים לכוונון באופן ידני
המדדים . אות מייד לאחר הגירויובכך מאפשר זיהוי ומדידה של תגובות הב, הארי של הארטיפקט
ניסויי הגירוי . fig.10A -דוגמאות לרישומים ניתן לראות ב. הנשלטים הם משך הגירוי והמשרעת שלו
. 0.5-1ms למשך זמן של 0.5-2µAעצמות הגירוי נעו סביב . 170Cהישיר נערכו בטמפרטורה של
.)Dudel - )Dudel, 1981, 1983 לצורך פירוט נוסף של השיטה ראה
כפי שמצוין , בחלק מניסויי הגירוי הישיר : וסיווג טרמינלים לפסיבי ואקטיביTTX -שימוש ב
. את ממברנת הטרמינל לפסיבית ההופך, 0.5µM בריכוז של TTX-נעשה שימוש ב, המשךב
נערכה , סירוגין עקב הצורך בגירוי ישיר וגירוי אקסון לTTX -בנעשה שימוש לא בניסויים בהם
גירוי צמות ועמספר ב נרשמה התגובה לגירוי ישיר במהלכה , טרם הניסוי עצמובדיקה מקדימה
טרמינל ממברנת . או פסיבי בדיקה זו אפשרה לסווג את הטרמינל כאקטיבי.מתגברותהולכות ו
כך שגירוי ישיר בעצמה חזקה מסף זה מעוררת אלמנטים ממברנליים , גירויסף בנתאקטיבי מתאפיי
fig.3ראה , "הכל או כלום"הדומה ל שחרור בעל אופי יחס עצמת גירוי אריים ומתקבליי לינבלת
מתאפיין ו,שחרור הופך רציף- העקום עוצמת גירויTTX -לאחר טיפול ב. עיגולים מלאים, מימין
לאמבט לא משפיעה על TTXבטרמינל פסיבי הוספת . )עיגולים פתוחים (שיפוע אחיד בקירובב
רובם . משמאלfig.3ראה , ללא תלות בעצמת הגירוי, קואנטלית בתגובה לגירוי ישירהתכולה ה
בניגוד לכלל , ם הטרמינלים המרכזיים המרוחקים מהאקסון המרכזי נמצאו פסיבייהמוחלט של
.ו התנהגות חשמלית אקטיביתציגוהטרמינלים הפרוקסימליים שה

23 שיטות_________________________________________________________________________________
Figure 3 : תוך גירוי ישיר בעוצמה מתגברת הפרדה בין טרמינל אקטיבי לפסיבי
פולס תכולה קואנטלית של שני טרמינלים בתגובה ל
. 0.3-1.8µAבעוצמות בין , 0.7msבמשך גירוי ישיר
גירויים 200 -מוצעה התגובה לעבור כל נקודה
לאחר הביקורת הוסף . 0.5Hzבתדר עוקבים
והניסוי נערך בשנית, TTX (0.5µM)לאמבט
נותר קבוע במהלך RS. )וחים פתעיגולים/ריבועים(
.הניסוי כולו
בעל התנהגות , מימין טרמינל מרכזי אופייני
משמאל טרמינל פרוקסימלי . חשמלית פסיבית
שגירוי 0.4µAסביב ניתן לראות סף גירוי .אופייני
ממברנת התנהגות אקטיבית של חזק ממנו מעורר
- לאחר טיפול ב.רמינל או האקסון הפרטרמינליטה
TTXבדומה לטרמינל , ינל הופך פסיבי הטרמ
.המרכזי משמאל
רישום .2.3.3
Dudelי " שתוארה עMacropatch-clamp -הזרמים הסינפטיים נרשמו חוץ תאית בשיטת ה
)Dudel, 1981( ,תוך שימוש במגבר- Zeitz-instruments .מערך הניסוי מופיע ב- fig.2A,B .
עבר , ולאחר מכן הקצה הוחלק תוך שיוף ,25µm -מי של כפיפטות זכוכית נמתחו לקוטר פני
נמדדה בתחילת כל RE - התנגדות הפיפטה.15µm -והותאם לקוטר פנימי של כ, התעגלות בחום
לרוב שימשה פיפטה בודדת לרישום במספר רב של . 100-200kΩ (fig.2B) ונעה בין , ניסוי ובסופו
רמינל וממברנת השריר לצורך רישום הזרמים הביאה ליצירת הנחת הפיפטה על גבי הט. ניסויים
התנגדות זו נמדדה באופן . (Rs: Seal resistance)התנגדות בין תוך הפיפטה לתווך החוץ תאי
, של הפרש המתחים על פני הפיפטה לערך פנימי חיובי(10ms)י קיבוע קצר "רציף במהלך הניסוי ע
.50-200kΩנעו בין RSי ערכ. י המגבר"ומדידת הזרם המוזרם ע
אולם במידה וחל שינוי עקבי , ברוב המכריע של הניסויים(fig.2C) היו יציבים על ציר הזמן RSערכי
. הרישום הופסק והתוצאות נזנחו, מערך ההתנגדות10%של למעלה מ
תוך , ום הרצוי בעזרת הפיפטהחיפוש הטרמינל התבצע על ידי סריקת תאי השריר באזור הריש
במיקום בו נרשמו . אחת לשתי שניות100Hzהפעלת פרוטוקול גירוי שכלל חמישה גירויים בתדר
תוך תיקון המיקום שלה במישור המקביל לממברנת , זרמים שליליים הוצמדה מעט הפיפטה לשריר
0.2 0.5 10.01
0.1
1
10
proximalcentral
m
direct stim. amplitude (-µA)
control+TTX
0.2 0.5 10.01
0.1
1
10
control+TTX

24 שיטות_________________________________________________________________________________
ENTC (Endנוכחות : הםהקריטריונים לרישום . עד להשגת רישום אופטימלי, וחוזר חלילה, השריר
Nerve Terminal Current) קלה לזיהוי (fig.5B) , מ גבוהיםשיפועי זרמים סינפטיים- nA/ms1 ,
והחשוב מכולם , שונות ברורה בין הזרמים הבאה לידי ביטוי בקפיצות בעלות אופי קונטלי במשרעות
.(fig.5B)הפרדה ברורה מאוד לעין בין כישלון לשחרור
בתדר ( תגובות באופן רציף 500נרשמו ): baseline(בהמשך נבדקה יציבות הרישום בתנאי בקורת
במידה והתקבלה ירידה . מוצעו כנקודה עם סטיות תקן תגובות 50כאשר כל , )1Hzגירוי של
פעמיים רישום אירועים ספונטניים נעשה . מונוטונית של מתחת לסטיית תקן אחת הופסק הרישום
ללא נעשה זה רישום לרוב . דקות15למשך זמן רצוף של , )בהתחלה ובסיום (בכל ניסוילפחות
תעוררות פוטנציאל פעולה ספונטני באקסון כיוון שניתן ללא כל קושי לזהות הTTX -שימוש ב
ולכן להגדיר את התגובה הבאה אחריו , ברישוםAP / ENTCהאקסיטטורי באמצעות זיהוי
הרישום עבר דיגיטציה . ספונטני היו ככלל נדירים ביותר אם בכללAPאירועים אלה של ". מעוררת"כ
כל תוכנות הדגימה נכתבו בהתאם . ל מחשבונרשם ישירות לדיסק קשיח ש, 20kHzבתדר של
.LabVIEWלדרישות הספציפיות בתוכנת
בעיות רישום הנובעות מגירוי בתדרים גבוהים .2.3.4
בתדרי גירוי גבוהים קיימת נטייה של תאי השריר להתכווץ בתלות בתדר הגירוי : כווץ השריר
כווצות השריר לא אפשרה בחלק מהפרפרטים הת. בעיקר בתאים הפרוקסימליים, ובמספר הגירויים
, ברוב המקרים למרות כווץ מינימלי של השריר בתדרים הגבוהים. רישום של תגובות בתנאים אלה
שימש כפולס AP/ENTC -במהלך גירוי בתדרים הגבוהים ה. הרישום נותר יציב לאורך שעות
ע ויציב לאורך סדרת ומדידתו הראתה שגודלו נותר קבו, )במידה והיה גדול די צורכו(קליברציה טבעי
היבטים נוספים . לא מלווה בשגיאה משמעותיEPSC -מכאן ניתן להניח שגם מדידת ה. הגירויים
בחלק מהמקרים כווץ . בנוגע ליציבות הרישום בתנאים אלה מופיעים בדיון בסוף הפרק הרביעי
של , יחסיתובעל קבוע זמן ארוך, EPSC -השריר הכניס זרם ביפאזי בעל משרעת נמוכה יחסית ל
זרם זה לא הפריע למדידת הזרמים הסינפטיים הודות לקינטיקה האיטית והמשרעת . msעשרות
.הנמוכה המאפיינים אותו
בתדרים ) עשרות(סדרת גירויים ארוכה : נוספות רישום ובעיות סיכום לא ליניארי של תגובות
העיקריים להלן פירוט.מי הזרםרישוביוותים אפשריים עלמספרעשויה להביא ) Hz50>(גבוהים
:בהם

25 שיטות_________________________________________________________________________________
הכוח המניע יקטןבמקרה זה :תא הפוסט סינפטיה ממברנת שלמצטברת פולריזציהד .1
תופעה זו רלוונטית יותר לסיבים הפרוקסימליים המתאפיינים . הנרשםסינפטיפוסטלזרם ה
במידה ומתרחשת עליה . ניסה גבוהה יותרובהתנגדות כ, בתגובות סינפטיות גדולות יחסית
הדרך . משמעותית בפוטנציאל התא עשוי להתקבל סיכום לא ליניארי של תגובות
הקיבוע מחייב החדרת זוג . האופטימלית למנוע תופעה זו היא לקבע את המתח בתא
אולם תזוזות קלות עקב התכווצות התא עשויות לפגוע בשלמות , אלקטרודות לתא
בעבודה זו לא נעשה קיבוע מתח שאמנם לא . ר בו הוחדרו האלקטרודותהממברנה באזו
מסיבות אלה בתדרי הגירוי הגבוהים יתכן והתכולה . נחוץ ברוב תדרי הגירוינמצא
נמוכה מהאמיתית עקב הסיכום הלא הקואנטלית שחושבה בסיבים הפרוקסימליים היא
לפחות באופן , נגרמת גירוי גבוהבתגובה לתדר ויה בשחרורוויתכן שהופעת הר, ליניארי
הודות לתגובות פחות משמעותית ל "הנ בסיבים המרכזיים התופעה .מגורם זה, חלקי
.והתנגדות הכניסה הנמוכה, הנמוכות האופייניות לסיבים אלה
במהלך גירוי מצב זה עשוי לעוות את הרישום בעיקר :ינל ממברנת הטרמדפולריזציה של .2
ולפיכך דפולריזציה של , גורם להיפרפולריזציה חוץ תאיתהישירגירוי ה .ישיר של הטרמינל
תופעות אלה עשויות להפחית את .הטרמינל מפחיתה למעשה את עצמת הגירוי הישיר
, קדמת של רוויה בשחרורולתרום להופעה מו, השחרור הסינפטי בתדרי הגירוי הגבוהים
בטרמינל עובדה שגירוי ישיר וגירוי האקסון ה.שאינה נובעת מתכונות מנגנון השחרור עצמו
וללא הופעת ) תוך כוון זהיר של עוצמת הגירוי הישיר( שחרור זהה-הניבו עקום תדריחיד
על שהגירוי הישיר הוסיף מרכיב דפולריזטורי מצטבר שוללת אפשרות ,(fig.10B) רוויה
לא ניתן לשלול קיום עוות זה מאידך .מעבר לזה שמוסיף גירוי האקסון, ממברנת הטרמינל
.בעוצמות גירוי חזקות יותר
מאחר וזו מצריכה שליטה , לא נעשתה הערכה מדויקת של השפעת עיוותים אפשריים אלה
קיימות עדויות לכךמאידך . במתח ממברנת הטרמינל ורישום מדויק של הזרמים דרכה
פירוט בנוגע לכך , משמעותי של התוצאותשינוי לים לא גורמיםבמידה וקיימ, אלהשעוותים
נעשה שימוש בספקטרום כמו כן לאורך כל העבודה. בסוף הפרק ברביעיראה בדיון
הם למעשה , בגירוי האקסוןבמידה וקיימים, כך שעיוותים אלה, פיזיולוגי של תדרי גירוי
וכך יש להתייחס לתוצאות המתארות , ינפטי בפרפרט זהחלק בלתי נפרד מהשחרור הס
.שחרור בתדרי הגירוי הגבוהים
26 שיטות_________________________________________________________________________________
או /תופעה זו עשויה להוביל לדפולריזציה של האקסון ו :הצטברות אשלגן בתווך החוץ תאי .3
עלולה להתפתח אינאקטיבציה של תעלות נתרן וסידן ובעקבות במקרים אלה . הטרמינל
זהכשל .כשל בהולכת האקסוןפעה זו מופיע בדמות סממן של תו. זאת ירידה בשחרור
, כ בטרמינלים שהיו מרוחקים מורפולוגית מהאקסון המרכזי"בד, אירע לעיתים נדירות בלבד
במקרים אלה במהלך סדרת הגירויים החלה האטה הדרגתית . ובתדרי גירוי גבוהים
, )ט הגירוי מארטיפקENTC -התרחקות הדרגתית של אות ה(במהירות ההולכה באקסון
בעיה זו לא . עד להעלמות הסיגנל הפרסינפטי, ENTC -בד בבד עם הקטנת משרעת ה
ואותם , תדרי הגירוי הם בטווח הפיזיולוגימאחר ויתכן , היוותה מכשול במחקר הנוכחי
. תוצאות הניסוי נזנחוו, היו קלים לזיהויהתרחשה כן מיקרים בודדים בהם
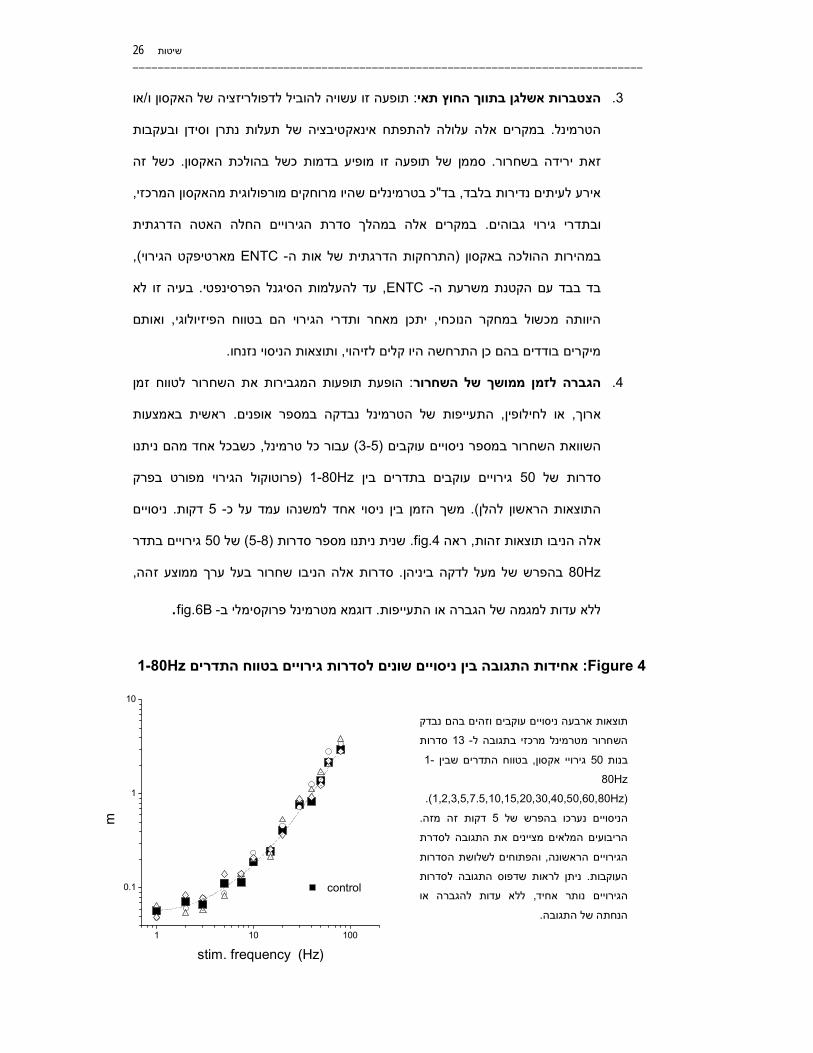
המגבירות את השחרור לטווח זמן ופעות ופעת תה: הגברה לזמן ממושך של השחרור .4
באמצעות ראשית . במספר אופניםנבדקה התעייפות של הטרמינל , או לחילופין, ארוך
ניתנו כשבכל אחד מהם, כל טרמינלעבור )3-5 (ניסויים עוקביםמספר ב ת השחרורהשווא
רט בפרק פרוטוקול הגירוי מפו (1-80Hzים בין בתדרים עוקבים גירוי 50של סדרות
ניסויים . דקות5 -משך הזמן בין ניסוי אחד למשנהו עמד על כ. )התוצאות הראשון להלן
בתדר ים גירוי50 של )5-8(ות סדרו מספרניתנשנית . fig.4ראה , אלה הניבו תוצאות זהות
80Hzזההממוצע רך ל עבעסדרות אלה הניבו שחרור . בהפרש של מעל לדקה ביניהן ,
.fig.6B -ב מטרמינל פרוקסימלי דוגמא. הגברה או התעייפותמגמה שלללא עדות ל
Figure 4 : 1-80גירויים בטווח התדרים לסדרות בין ניסויים שונים התגובה אחידותHz
בהם נבדק עוקבים וזהים תוצאות ארבעה ניסויים
סדרות 13 -שחרור מטרמינל מרכזי בתגובה לה
-1 ן בטווח התדרים שבי, אקסוןגירויי 50בנות
80Hz
(1,2,3,5,7.5,10,15,20,30,40,50,60,80Hz) .
. דקות זה מזה5הניסויים נערכו בהפרש של
הריבועים המלאים מציינים את התגובה לסדרת
לשלושת הסדרות והפתוחים, הגירויים הראשונה
ת ו ניתן לראות שדפוס התגובה לסדר.העוקבות
ללא עדות להגברה או , הגירויים נותר אחיד
.של התגובההנחתה
1 10 100
0.1
1
10
m
stim. frequency (Hz)
control

27 שיטות_________________________________________________________________________________
ככלל תדר האירועים הספונטניים בכל :תדר אירועים ספונטניים ושחרור אסינכרוני .5
80Hzלקראת סופה של סדרת גירויים בתדר . 0.01-0.1Hz: ונע בין, הטרמינלים היה נמוך
, שחרור המופיע לאחר חלון הזמן של השחרור המעורר( שחרור אסינכרוני לעיתיםהופיע
מקור . Ravin et al. 1997,1999): ראה, עבור חלון זה5ms -ווה לכאן נלקח ערך הש
-ss (steady[+Ca2]ל בעליית ריכוז הסידן התוך תאי הממוצע"השחרור האסינכרוני הנ
state average intracellular Ca2+ concentration) , בעקבות סדרת גירויים ממושכת
ברוב המקרים היה מדובר ). 40Hz) Ravin et al. 1997,1999 -בתדרים של מעל ל
כלומר מספר אירועים נמוך , במהלך סדרת הגירוייםוקלים לזיהוי באירועים בודדים
ולכן נוכחות שחרור זה לא גרמה לטעות או עוות במדידת , משמעותית בהשוואה למעורר
.רמות השחרור המעורר
מדידה .2.4נעשה תוך מדידת המדדים ) ENTC, EPSC(סינפטיים והפרסינפטיים פוסטכימות הזרמים ה
בנקודת המקסימום של נגזרת הזרם לפי (שיפוע מקסימלי , אינטגרל לפי זמן, משרעת: הבאים
נמדד baseline. כמו כן נמדד האיחור הסינפטי. זמן עליה למקסימום וזמן הדעיכה למחצית, )הזמן
כל תוכנות המדידה נכתבו . לפני ובסמוך לתגובה הסינפטית3ms -כ מתוך חלון זמן של כ"בד
. LabVIEWבהתאם לדרישות הספציפיות בתוכנת
חישוב התכולה הקואנטלית .2.4.1
הראשונה מתוך התפלגות משרעות אירועים . מדידת גודל הקואנטום נעשתה בשתי דרכים
ההתפלגות המתקבלת . 0.1Hz -ר נמוך של מתחת להמופיעים ברוב הטרמינלים בתד, ספונטניים
המתמשך לכוון המשרעות הגבוהות " זנב"עם , בטרמינלים אלה היא נורמלית סביב ערך הקואנטום
מהתפלגות זו . המורכבים מיותר מקואנטום בודד, זה מייצג אירועים ספונטניים נדירים" זנב. "יותר
ים טרמינלים בהם תדר האירועים הספונטניים גבוה יש לציין שקיימ. קל להעריך את גודל הקואנטום
טרמינלים אלה לא נכללו בניסויים המתוארים . וכך גם שכיחותם של האירועים הגדולים יותר, יותר
. בפרק זה

28 שיטות_________________________________________________________________________________
cumulative)יה היא על סמך ניתוח סטטיסטי של התפלגות מצטברת של תגובות מעוררות ידרך שנ
distribution)) ראהfig.5A .(ואת זו הבאה , מהתפלגות זו קל לאתר את פרקציית הכישלונות
כעת מורכבת הסטוגרמה המאפשרת למדוד במדויק . המייצגת את אירועי הקואנטום הבודד, אחריה
מחושבת על ידי חלוקת ממוצע המשרעות (m)התכולה הקואנטלית . (fig.5C)את גודל הקואנטום
.ל"אנטום הנשל התגובות הסינפטיות בגודל הקו
Figure 5 : מדידת גודל הקואנטום הבודד
A : 500דוגמא להסטוגרמת שכיחות מצטברת של
הקווים המקווקוים מציינים את . משרעות אירועי ביקורת
ערך המשרעות בהן מתקבל שיפוע מקסימלי על גבי
רישומי תגובות מתוך 15דוגמא של : B .העקומה
ל לזהות את הכישלונות ק. A-האירועים המופיעים ב
ואת הקפיצות בעלות האופי הקואנטלי , בשחרור
ENTC - End Nerve Terminal. במשרעות הזרמים
Current. C: היסטוגרמת עמודות של משרעות
. 40pA - שווה לbin -גודל ה. A -האירועים המופיעים ב
ניתן . A -הקווים המקווקוים ממשיכים את אלה שב
בערכים (peaks)שלושה שיאים לראות בבירור לפחות
הקו הרציף מתאר התאמה . nA :(0 ,0.45 ,0.9-ב(
לסכום שלוש עקומות נורמליות בעלות ממוצע השווה
. גודל הקואנטום הבודד-q .לערכים אלה
0.0 0.5 1.0 1.5 2.00.0
0.2
0.4
0.6
0.8
1.0
cum
ulat
ive p
roba
bilit
y
amplitude nA
!
1ms
0.25nA
EPSC
ENTC
0.00 0.45 0.90 1.35 1.800
20
120
q
C.
B.A.
amplitude nA
No.
of e
vent
s

29 שיטות_________________________________________________________________________________
הערכה סטטיסטית של התוצאות .2.5ון על על מנת להעריך האם קיימת השפעה פרמקולוגית מובהקת של חומר נת:טרמינל בודד
השחרור הסינפטי מטרמינל בודד נערכה השוואה בין ערכי השחרור הסינפטי בביקורת ובמהלך
בכל הניסויים הכיל . למדגמים בלתי מזווגים, מוטה לשני הצדדיםT-test -תוך שימוש ב, הטיפול
ניתוח . לרוב הרבה מעבר לדרוש לצורך השגת מובהקות סטטיסטית, המדגם מאות תגובות
קצב , סטטיסטי בנוגע להשפעה על מאפיינים אחרים של התגובה כמו פוטנציאציה מקסימלית
פועי עקומים לא נבדקו בניסוי בודד מאחר ומאפיינם אלה מחושבים מתוך ושיהתפתחות השחרור
.ממוצעי המדידות בכל ניסוי
ולאחר , בכל טרמינל חושבה התכולה הקואנטלית של התגובות כפי שתואר לעיל: סדרת ניסויים
בין ממוצע זה מכיל את השונות הפיזיולוגית. מכן מוצעו התוצאות מכל הטרמינלים בסדרת הניסויים
שהייתה בעלת חשיבות עקרונית מאחר ובמהלך העבודה נערכה , הטרמינלים באותה סדרת ניסויים
בחלק מניסויי הפרמקולוגיה נעשה המיצוע לאחר נרמול . השוואה בין אוכלוסיות שונות של טרמינלים
מתוך מטרה לנטרל את השונות בין , ערכי השחרור הסינפטי בכל טרמינל לערך הביקורת
. לים ולבטא את השונות של הטיפול הפרמקולוגיהטרמינ
בדיקת מובהקות ההשפעה של טיפולים פרמקולוגיים שונים על מאפיינים נבחרים של השחרור
למדגמים , מוטה לשני הצדדים T-test -הסינפטי באוכלוסיית טרמינלים נעשתה תוך שימוש ב
בין ) מופיע בהמשך, ½St.n(ה בדיקת מובהקות ההבדל בין מדדים המתארים קינטיק. מזווגים
לא נשענה ) או לחילופין בביקורת ולאחר טיפול פרמקולוגי באותה קבוצה(קבוצות טרמינלים שונות
מסיבה זו נבדקה ראשית הומוגניות השונות של . על ההנחה שמדדים אלה מתפלגים באופן נורמלי
וההבדל היה מובהק במידה . (Homogeneity of variance)המדד בשתי הקבוצות הנבדקות
(p<0.01)פרמטרי - בוצע מבחן א(Mann-Whitney) מוטה לשני הצדדים לקביעת מובהקות ההבדל
.שנרשם בערך המדד הנבדק בין שתי הקבוצות
לעקומים המתארים את השחרור הסינפטי כפונקציה של תדר הגירוי : התאמת פונקציות לתוצאות
: הותאמה פונקציה מעריכית) שחרור- תדראו עקומי, עקומי התגובה לגירוי בתדר(
m(f) : התכולה הקואנטלית כפונקציה של תדר הגירוי .
m1 : 1התכולה הקואנטלית בתדר גירוי שלHz.
K : קבוע המקשר בין יחידותHzליחידות קואנטה .
f : תדר הגירוי(Hz).
nf fKmm ⋅+= 1)(

30 שיטות_________________________________________________________________________________
n : מעריך.
הרגרסיה הלא ליניארית המבוססת על האלגוריתם של ל נקבעו לפי שיטת"ערכי הפרמטרים הנ
Levenberg-Marquardt . בכל התאמה של הפונקציה לעקום התגובה לגירוי בתדר של טרמינל
. ערכי התחלה זהיםm1, K, n: נתון נקבעו לכל הפרמטרים
.r2 0.98 (coefficient of determination)בכל המקרים הושגה התאמה של

31
התגובה לגירוי בתדר בתנאי ביקורת .3___________________________________________________________________
נוהל הגירוי וניתוח התגובה: שיטות .3.1 תוך שימוש בטווח תדרים רחב, האקסון האקסיטטוריבפרק זה השחרור עורר בתגובה לגירוי
נוהל הגירוי הורכב .)Crider & Cooper, 1999, 2000(הגת מחיה מתנEMGעל מדידות המבוסס
, 80Hz - 1ונע בטווח של , כשתדר הגירוי בתוך כל סידרה היה שונה, מסדרות של חמישים גירויים
, Hz :(1 ,2 ,3 ,5 ,7.5 ,10 ,15 ,20 ,30 ,40 ,50 -ב(בסדר עולה , סדרות בתדרים שונים13כ "בסה
במטרה למנוע התפתחות , דקהוח הזמן בין סדרות הגירויים בתדר גבוה עלה על מרו. 80, 60
, שניות7 -תהליך המתאפיין בקבוע דעיכה של כ(תהליכי הגברה ארוכי טווח כדוגמת אוגמנטציה
התגובות הראשונות מסדרת 15 דוגמא לרישום fig.6A -ב). Zucker & Regehr 2002ראה
ניתן להתרשם מהיעדר כמעט . מרכזי ופרוקסימלישל טרמינל, 80Hz - ו 1Hzם הגירויים בתדרי
-ב. ומתכולה קואנטלית גבוהה יחסית, מוחלט של כישלונות בשחרור מהטרמינל הפרוקסימלי
fig.6B 80 גירויים בתדר 50 דוגמא לתגובה ממוצעת של טרמינל פרוקסימלי לסדרות בנותHz.
חולק באופן מלאכותי לשני חלקים ניתוח התגובה :גבוהיםירוי בתדרים ניתוח התגובה לגאופן
.ומפורט להלן fig.6B -ב שמצוין מעל לתגובה כפי, plateau phase 2 (rising phase )1: נפרדים
הנמדד לאחר , ערך הרוויה של השחרור הסינפטי:(plateau) או מופע תלוי תדרמופע הרוויה .1
כאשר התגובות מתייצבות סביב ערך קבוע ערך זה מציין את מקסימום השחרור , העלייהשלב
כלומר הערך אליו יגיע השחרור הסינפטי אם יינתנו , הסינפטי של הטרמינל עבור תדר גירוי נתון
. במופע הרוויה אין התייחסות למהירות בה מתפתח השחרור הסינפטי. מספיק גירויים בתדר זה
התגובות 3020כ "בד, דד על ידי מיצוע התגובות לאחר ההגעה לרוויהערך הרוויה נמ
מתוך ערכי הרוויה מחושבת הפוטנציאציה המקסימלית בכל . האחרונות בכל סדרת גירויים
.1Hzבאמצעות היחס בינם לתגובה הממוצעת בתדר גירוי של , תדר
ברה של השחרור הסינפטי מופע זה מתייחס לשלב ההג:(rise) או מופע תלוי זמןמופע העלייה .2
עצמת (מאפיינים אותו משרעת הפוטנציאציה בכל תגובה . תוך מתן סדרת גירויים בתדר גבוה
המבוטא כמותית במספר התגובה בה , קצב התפתחות השחרורו, )התפתחות הפוטנציאציה
). קו מקווקו(רוי נתון עבור תדר גי, מגיע השחרור הסינפטי למחצית מערך הרוויה בממוצע
.מפורטת בהמשך, St.n½ (Stimulus number ½)המכונה להלן , ההתייחסות למדד זה

32 התגובה לגירוי בתדר בתנאי ביקורת_________________________________________________________________________________
. הפרוקסימלי והמרכזי-נוהל גירוי זה הופעל על טרמינלים משני אזורים עיקריים בשריר
Figure 6 :לים פרוקסימליים ומרכזיים התגובה לגירוי בתדר של טרמינ
A :רישומי חמש עשרה התגובות הראשונות של הטרמינלים מ- B1 בתדרים של אקסוןלסדרות גירויי, לעילHz ,ו-
80Hz .הכוכבית מציינת את אות ה- ENTCבכל טרמינל .B :ותדוגמא לתגובה ממוצעת של טרמינל פרוקסימלי לסדר
סטיית התקן הנמוכה יחסית מעידה על יציבות התגובה לסדרות גירויים . 80Hz (mean±sd;n=8) גירויים בתדר 50של
50% -הקו המקווקו מציין ערך השווה ל. שלב הרוויה של התגובה-plateau, שלב התפתחות התגובה-rise. חוזרות
מקסימום י מנורמל לומרכז, )ריבועים(מרכזי , )עיגולים(דוגמא לתגובה לתדר של טרמינל פרוקסימלי : C. מערך הרוויה
השוואה בין העקום הפרוקסימלי והמרכזי המנורמל מדגימה את האיחור על ציר התדר ). הקו המקווקו (פרוקסימליה
יש . הראשונה בשחרור" מדרגה"החצים מסמנים את התדרים בהם חלה ה. בהתפתחות התגובה של הטרמינל המרכזי
כפי שמפורט בדיון בסוף הפרק , בשחרור הופיעו רק בחלק קטן מהטרמינלים הפרוקסימליים" מדרגות"לציין ששלושת ה
.הרביעי
0 10 20 30 40 500
1
2
3
C.B.
50%
plateaurise
ampl
itud
nA
stimulus No.1 10 100
0
2
4
6
8
10
12
""
stim. frequency Hz
quan
tal c
onte
nt proximal terminal central terminal central, normalized
A.
*
1Hz
Central:
1ms
0.3nA80Hz
*Proximal:

33 התגובה לגירוי בתדר בתנאי ביקורת_________________________________________________________________________________
התלות בתדר הגירוי ובמיקום : השחרור הסינפטי במופע הרוויה .3.2 הטרמינלים על גבי השריר
הכוונה הייתה למדוד במדויק . ים לא הייתה בחירה מקריתיהבחירה בסדרות בנות חמישים גירו
לשם כך נוסו גם סדרות בנות . בכל אחד מתדרי הגירוי, הרוויה היציב של השחרור הסינפטיאת מצב
: בסדרות אלה הופיעה לעיתים תופעה של מצב רוויה בלתי יציב. מאה ומאתיים גירויים לסדרה
לאחר כחמישים , אולם בהמשך, השחרור הסינפטי התייצב סביב ערך רוויה לאחר כעשרים גירויים
תופעה . גבוה יותר מקודמו, ובהמשך התייצב סביב ערך רוויה חדש, ספים החל לעלות שובגירויים נו
, ולכן ההתייחסות הייתה תמיד למצב הרוויה היציב הראשון, זו לא הייתה עקבית בכל הטרמינלים
בחירה זו במצב הרוויה הראשון נועדה . המושג לרוב כעבור עשרים עד שלושים הגירויים הראשונים
והיא התפתחות הגברה בעלת , עיה נוספת המתעוררת עם שימוש בסדרות גירויים ארוכותלמנוע ב
. (Augmentation, Post Tetanic Potentiation)של התגובה ) שניות עד דקות(טווח זמן בינוני
פעה של ומנע הו, שימוש בסדרות בנות חמישים גירויים אפשר מדידה אמינה של מצב הרוויה מחד
כך שתוצאות מדידות חוזרות של התגובה לתדר באותו טרמינל , תופעות הגברה של התגובה מאידך
. היו עקביות
ניתן לראות דוגמא לתגובה לתדר של שני טרמינלים מייצגים מהאזורים הפרוקסימלי fig.6C -ב
ת שהשחרור הסינפטי מתפתח ניתן לראו). fig.6A -אותם טרמינלים שתגובתם מיוצגת ב( והמרכזי
ובעיקר , מהטרמינלים50% -תופעה זו חזרה על עצמה בכ. שלוש במספר בכל טרמינל, "מדרגות"ב
.התייחסות נוספת לתופעה זו מופיעה מאוחר יותר בדיון. נראתה באזור המרכזי
:ניתן לעמוד על שלושה הבדלים עיקריים בתגובה לתדר של שני סוגי הטרמינלים
ינפטי מהטרמינל המרכזי בתדרים הנמוכים נמוך יותר בסדר גודל אחד לפחות השחרור הס .1
).fig.6A -1Hz: ראה, בטרמינל זה רוב התגובות לגירוי בתדר נמוך הן כישלונות(
התדר בו מתחילה . השחרור הסינפטי מהטרמינל המרכזי מתחיל להתפתח בתדרים גבוהים יותר .2
חצים (ה כפליים בקירוב בהשוואה לטרמינל הפרוקסימלי עליה משמעותית בשחרור הסינפטי גבו
). בהתאמה10Hz ,5Hz בתדרים fig.6C -ב
בניגוד לרוויה , השחרור הסינפטי מהטרמינל המרכזי לא מתקרב לרוויה בתדרים הגבוהים .3
. ומעלה50Hzהמושגת בטרמינל הפרוקסימלי בתדרי גירוי של

34 התגובה לגירוי בתדר בתנאי ביקורת_________________________________________________________________________________
, בתדרים גבוהים יחסית" מתעורר", ד בתדרים נמוכיםלסיכום הטרמינל המרכזי משחרר מעט מאו
.ולא מגיע לרוויה בתדרים הגבוהים
. מרכזיים16 - טרמינלים פרוקסימליים ו8עבור , ניתן לראות תגובה ממוצעת לתדרfig.7A -ב
היות ובכל טרמינל הן מתרחשות בתדרים מעט, בשחרור הסינפטי נעלמו עקב המיצוע" מדרגות"ה
, השחרור הסינפטי מהטרמינלים הפרוקסימליים גבוה יותר אבסולוטית בכל תדרי הגירוי .שונים
גבוה יותר 12פי : 10Hzהיחס המקסימלי מתקבל בגירוי בתדר . ובעיקר בתדרים הנמוכים
הטרמינלים הפרוקסימליים מגיעים למחצית מהתכולה הקואנטלית המקסימלית . בפרוקסימליים
שלא מגיעים לרוויה , במרכזיים50Hzלעומת , 25Hz בתדר גירוי של כבר) 80Hzבתדר גירוי של (
השיפוע הלוגריתמי של עקום התגובה לתדר בטרמינלים המרכזיים גבוה . גם בתדרים הגבוהים
). בהתאמה, 1.05 מול 1.68(כמעט כפליים בהשוואה לפרוקסימליים
רכזי בתיאור עקום התגובה מדד מכאן המהווה , ל"על מנת לעמוד על משמעות שיפוע העקום הנ
בשני ,(m)התכולה הקואנטלית המעוררת תלות אופי על כאן להרחיב מעט מקוםיש , לגירוי בתדר
התוך תאי הממוצע המתפתח הודות לגירוי סידןה ריכוז ,(f) גירויהתדירות : המרכבים המרכזיים
פירוט נוסף ((f » [Ca2+]ss » m)שחרור -סידן-תדר: כלומר התלות המשולשת. ss[+Ca2]מתמשך
ss » m.Dodge & Rahamimoff (1967)[+Ca2]: ראשית נתייחס לתלות. )מופיע מאוחר יותר בדיון
,לשחרור סינפטי החוץ תאי המתאר את התלות בין ריכוז סידן , שיפוע העקום הלוגריתמיייחסו את
, מאידך .(Hill Coefficient) 4 -וקבעו לו ערך השווה ל, רהשחרולתכונה קואופרטיבית של מנגנון
וריכוז הסידן התוך תאי הממוצע במהלך סדרות גירויים בתדר גבוה הניבה ערכים נמוכים mמדידת
היא פונקציה של סכום ריכוז m -הסבירו זאת בכך ש) Ravin et al.) 1999. יותר של קבוע זה
ר השחרור הודות לשטף וריכוז הסידן המושג באזור את,(ss[+Ca2])הסידן הממוצע הנמדד לעיל
ריכוז סידן זה ). לצורך הבהרה נוספתRavin et al., 1999ראה ((Y)כניסת הסידן בעקבות הגירוי
Yמסיבה זו יש להתייחס לערכים נמוכים יותר אלה . הוא בעל אופי ארעי וקשה טכנית למדידה
כניות לא מופיעה שמסיבות טYהנובעת מתרומת , בלבדHill Coefficientכאל תת הערכה של
משקף תלות חזקה של השחרור (4~)לפיכך שיפוע בעל ערך קרוב למקסימלי . I » m[*Ca2]בתלות
ואילו שיפוע נמוך משקף תלות חזקה בסידן ממקור חיצוני בעקבות , בריכוז הסידן הממוצע בטרמינל
.ss » m[+Ca2]עד כאן בנוגע לתלות . הגירוי

35 התגובה לגירוי בתדר בתנאי ביקורת_________________________________________________________________________________
Figure 7 : התגובה לתדר ופוטנציאציה מקסימלית מאפייני
A : תגובה לתדר של טרמינלים פרוקסימליים(n=8) , ומרכזיים(n=16) ,(mean±se) .הקו הרציף הוא פונקציה מעריכית ,
בתגובה , עקומי פוטנציאציה מקסימלית ממוצעת: B. שני הצירים הם לוגריתמיים. מעריך החזקה מצוין ליד כל עקומה
-בערך השווה ל, הפוטנציאציה משתווה בין הטרמינלים10Hzבתדר של . A -י קבוצות הטרמינלים מבשת, לגירוי בתדר
2.3 C : נגזרת העקומים מ(נגזרת השחרור לפי תדר של הטרמינלים הפרוקסימליים והמרכזיים- A .( ניתן לראות שקצב
. quanta/Hz 0.28 -ושווה ל, 50Hzקבוע עד לתדר של ) עיגולים(התפתחות השחרור בטרמינלים הפרוקסימליים
יכולת הגברת הפוטנציאציה בכל : D. מתגבר עם עליית התדרקצב התפתחות השחרור) ריבועים(בטרמינלים המרכזיים
תדר הגירוי מסומן בצד כל נקודה על (1-40Hz הפוטנציאציה עבור התדרים Xבציר . בתגובה להכפלת תדר הגירוי, תדר
הטרמינלים 10Hzניתן לראות שהחל מתדר של . מידת הגברת הפוטנציאציה הודות להכפלת תדר הגירויYבציר ). רףהג
הכוכבית מסמנת את התדר בו רגישות . משלשים את הפוטנציאציה בתגובה להכפלת תדר הגירוי) ריבועים(המרכזיים
.15Hz -ל שווה ל"ם התדר הנבשני סוגי הטרמינלי. הטרמינל להכפלת תדר הגירוי היא מקסימלית
כלומר, עבודות שונות בפרפרט הנוכחי במספר ת כליניאריתמתואר f » [Ca2+]ss, נה הראשוהתלות
עם הגברת תדר הגירוי תליניארי עולה ) הרוויהךער (בטרמינלהממוצע ריכוז הסידן התוך תאי
)Cooper et al., 1995a; Tank et al., 1995; Msghina et al., 1999; Ravin et al., 1999 ( .
שחרור -כלומר שיפוע התלות תדר גירוי, ss » m[+Ca2] תלות דומה איכותית לf » m תלותשהמכאן
1 10 100
0.1
1
10
20
D.C.
slope 1.68
slope 1.05
B.A.proximalcentral
quan
tal c
onte
nt
stim. frequency Hz1 10 100
0.1
1
2.3
10
100
7.5
*
*
1Hz
3 510
15 2030
5
proximal central
40Hz
2
3
7.5
1015 20
3040Hz
max
. pot
entia
tion
stim. frequency Hz
1 10 100
0.01
0.1
0.28
1
d(qu
anta
l con
tent
)/df
proximal central
stim. frequency Hz1 10 40
1.0
1.5
2.0
2.5
3.0
3.5
4.0
doub
led
stim
. fre
q. p
oten
tiatio
n
proximal central
max. potentiation

36 התגובה לגירוי בתדר בתנאי ביקורת_________________________________________________________________________________
ואכן ערכי השיפועים שנמצאו בעבודה זו זהים , ss » m[+Ca2]צפוי להיות דומה לשיפוע התלות
מכאן שמתוך שיפוע .(Ravin et al., 1999) בפרפרט זה ss» m[+Ca2] את התלותלאלה המתארים
הארעיתשל כניסת הסידןלשחרור שחרור ניתן ללמוד בפרפרט זה על התרומה -התלות תדר גירוי
העקום שיפוע ניתן לראות ש בטרמינלים המרכזיים .ביחס לזו של ריכוז הסידן התוך תאי הממוצע
ניתן לומר , לעילהנחה תקפות העל בסיס , ולכן, פרוקסימלייםשל הזה כמעט כפול ביחס ל
3~ פי(גבוהה יותר , בעקבות גירוי לשחרור כניסת הסידןה שלרומהתשבטרמינלים הפרוקסימליים
חיזוק להנחה זו מופיע בהמשך .בטרמינלים המרכזייםאשר מ, )Ravin et al., 1999 בהסתמך על
.ויים בהם הונמך ריכוז הסידן החוץ תאיובניס, בניסויי הגירוי הישיר
התלות בתדר ובמיקום הטרמינלים על גבי השריר: פוטנציאציה מקסימלית .3.2.1
הפוטנציאציה ברוויה עבור כל תדר מחושבת כיחס בין משרעת התגובה הסינפטית הממוצעת
, )1Hz בתדר גירוי של התגובה הממוצעת(למשרעת תגובה הממוצעת הראשונה , בכל תדר, ברוויה
. בהמשך הסעיף הזה המונח פוטנציאציה מתייחס לפוטנציאציה המקסימלית להלן. פחות אחד
היות ואי , ל יש צורך במדידה מדויקת ככל האפשר של התגובה הראשונה"לצורך חישוב היחס הנ
ים בחלק מהטרמינל. דיוק קל במדידה זו עשוי לגרום לשגיאה גדולה בחישוב הפוטנציאציה
כיוון שרוב או כל התגובות לגירוי בתדר נמוך הן , המרכזיים ביצוע מדידה מדויקת זו הוא עניין מורכב
ויש , עבור טרמינלים אלה לא ניתן לחשב כלל בנפרד את הפוטנציאציה באופן הקלאסי. כישלונות
מסיבה זו . צורך במדד אחר על מנת לכמת את האופן בו מתגבר השחרור הסינפטי עם מתן הגירויים
.והפוטנציאציה חושבה על פי הממוצע, מוצעו התכולות הקואנטליות של כל הטרמינלים המרכזיים
. (fig.7B)השוואת הפוטנציאציה המקסימלית בין שני סוגי הטרמינלים מראה תופעה מעניינת
בערך כפליים מאשר, בטרמינלים הפרוקסימליים הפוטנציאציה בתדרים נמוכים גבוהה יותר
ערכי 10Hzבתדר של . גבוה יותרקצב התפתחות השחרוראולם באחרונים , במרכזיים
ובתדרים גבוהים יותר , )2.3 -בערך השווה ל(הפוטנציאציה בשני סוגי הטרמינלים משתווים
בקירוב 6 -הפוטנציאציה בטרמינלים המרכזיים ממשיכה להתפתח עד שמגיעה לערך הגבוה פי
הבדל בין שיפועי היפוך זה מוסבר הודות ל). 13.4 מול 80.5(מאשר בטרמינלים הפרוקסימליים
על הבדלים בתרומת כניסת כאמור המעידה, שחרור בין שתי אוכלוסיות הטרמינלים-עקומי תדר
מראה שהעלייה fig.9C -התבוננות בנגזרת התגובה לתדר ב. קבות גירוי לשחרורעהסידן ב
בשחרור הסינפטי עם עליית תדר הגירוי בטרמינלים הפרוקסימליים היא קבועה עד לתדר של

37 התגובה לגירוי בתדר בתנאי ביקורת_________________________________________________________________________________
50Hz ,קואנטה בממוצע לכל 0.28 -ושווה ל- Hz , מעבר לתדר . קואנטה0.046עם סטיית תקן של
, בטרמינלים המרכזיים השחרור הסינפטי לא מתפתח בקצב קבוע, יגוד לכךבנ. זה מתפתחת רוויה
. אלא בקצב ההולך ומתגבר עם עליית תדר הגירוי
יכולתו של הטרמינל : ניתן לאפיין את התפתחות הפוטנציאציה בציר התדר תוך שימוש במדד נוסף
ניתן לראות את fig.9D -ב. להגביר את השחרור הסינפטי בעקבות הכפלת תדר הגירוי שלו
ל באם " את הגברת הפוטנציאציה הנYובציר , Xעל גבי ציר , הפוטנציאציה של התגובה בכל תדר
2-3Hzניתן לראות שהטרמינלים הפרוקסימליים מתייצבים כבר בתדרים של . תדר הגירוי מוכפל
תדר הגירוי כלומר יכולת הגברת השחרור הסינפטי בתגובה להכפלת, 2 -סביב ערך קבוע הקרוב ל
הטרמינלים המרכזיים . אולם היא מושגת כבר בתדרים נמוכים, אינה גבוהה בטרמינלים אלה
אולם יכולת זו כפולה כמעט , 10Hz -מפתחים יכולת הגברה המקסימלית רק בתדרים של מעל ל
שני סוגי הטרמינלים מפתחים 15Hzסביב תדר של . בהשוואה לזו של הטרמינלים הפרוקסימליים
. לת הגברה מקסימלית בתגובה להכפלת תדר הגירוייכו
של הטרמינלים השונים (proximal to distal) תוצאות אלה מדגימות ציר הפעלה אורכי ברור
, בתדרי גירוי נמוכים עיקר השחרור הסינפטי נתרם על ידי הטרמינלים הפרוקסימליים. בשריר
.לים המרכזיים הולכת והופכת מהותית יותרתרומת הטרמינ) 10Hzמעל (ובתדרי גירוי גבוהים יותר
תלות התגובה לגירוי בתדר של טרמינלים מרכזיים במרחק מהאקסון המרכזי .3.2.2
בעיקר בסף התדר שמעליו , קיימים הבדלים ניכרים בתגובה לתדר של הטרמינלים המרכזיים
של שני טרמינלים לתדרהתגובה ים עקומי מתוארfig.8A -ב. השחרור הסינפטי הופך משמעותי
, 10Hzהאחד מתחיל לשחרר כבר בתדר של . )גירוי אקסון (מרכזיים שונים על גבי אותו סיב שריר
בנוסף לכך שני הטרמינלים נבדלים במיקומם על גבי . 40Hz -ואילו השני רק בתדרים הגבוהים מ
ואילו האחרון , הטרמינל הראשון ממוקם בסמוך לסעיף המרכזי של האקסון לאותו תא: תא השריר
.5, 1מספרים , fig.8B - במופיע באיור סיב השריר מיקום טרמינלים אלה. ממוקם רחוק מהאקסון
בצורה דומה נרשמה .תא שריר מרכזי יחידעל גבי שונים בניסוי זה נערך רישום משמונה טרמינלים
טרמינלים 12 - ל8בין (שריר מרכזיים שונים סיבי 6 - טרמינלים מרכזיים מ55 -התגובה לתדר מ
רישומים משני .החל מאלו הקרובים ביותר לסעיף האקסון ועד למרוחקים ביותר, )לכל סיב
. fig.8C -טרמינלים מייצגים ראה ב

38 התגובה לגירוי בתדר בתנאי ביקורת_________________________________________________________________________________
Figure 8 : תדר בתוך אוכלוסיית הטרמינלים המרכזיים בהתגובה לגירוי שונות
A :מעויינים(והשני מרוחק ממנו , )ריבועים(האחד קרוב לאקסון , דוגמא לתגובה לתדר של שני טרמינלים מרכזיים .(B :
באחד מהניסויים , ביחס לסעיף האקסון המרכזי) שמונה במספר( את מיקום הטרמינלים על גבי השריר איור המתאר
מופיעה 5 - ו1התגובה לתדר של הטרמינלים . מהאקסון המרכזי400µm מרוחק 7טרמינל מספר . שבוצעו בסיב מרכזי
טרמינל.50Hzלסדרת גירויי אקסון בתדר בתגובה מטרמינלים אופיינייםעוקבים רישומים 15 -דוגמא ל: C. לעילA -ב
של שתי אוכלוסיות (mean.±se) הממוצעתהתגובה לתדר עקומי : D. ומרוחק מימין, משמאלהמרכזי קרוב לאקסון
עקומי הפוטנציאציה :E). מעויינים(והמרוחקת ממנו ) ריבועים(הקרובה לאקסון : בגירוי אקסוןטרמינלים מרכזיים
.D -ל שתי אוכלוסיות הטרמינלים המתוארות בש הממוצעתהמקסימלית
פיזיולוגית בארגון ומבנה הטרמינלים על גבי סיבי -ממצא זה העלה שאלה בדבר קורלציה מורפולוגית
שלהם לסעיף הפיזית הטרמינלים חולקו לשלוש קבוצות בהתאם לקרבה בהמשך . השריר המרכזיים
1 10 1000
1
2
3
4
5
quan
tal c
onte
nt
stim. frequency Hz
close to axon remote from axon
1 10 100
0.1
1
10
stim. frequency Hz
quan
tal c
onte
nt
slope 1.75
slope 1.35close to axonremote from axon
1 10 100
0.1
1
10
E.D.
C.
B.A.
pote
ntia
tion
stim. frequency Hz
close to axon remote from axon
close toaxon
0.5nA
1ms
remote from axon

39 התגובה לגירוי בתדר בתנאי ביקורת_________________________________________________________________________________
הממוצעת של שתי קבוצות הטרמינלים ניתן לראות את התגובה לתדר fig.8D -ב. האקסון
קיים ההבדל . טרמינלים המרוחקים ממנו19 -ו, טרמינלים הקרובים ביותר לאקסון18: הקיצוניות
בתדרים הנמוכים ההסתברות לשחרור . מובהק בתכולה הקואנטלית בין הקבוצות בכל התדרים
חלק . נלים אלה הן כישלונותובפועל כל התגובות בטרמי, סינפטי בטרמינלים המרוחקים היא אפסית
מעל לתדרים אלה . 40Hz -מהטרמינלים המרוחקים לא משחררים כלל עד לתדרי גירוי של מעל ל
אך לא , הטרמינלים המרוחקים משחררים בהסתברות ההולכת וגוברת עם עליית תדר הגירוי
חין בהבדל כמו כן ניתן להב. מגיעים לרמות השחרור הסינפטי של הטרמינלים הקרובים לאקסון
הבדל זה בתוך קבוצת הטרמינלים . הברור בין שיפוע עקומי התגובה לתדר בין שתי הקבוצות
.המרכזיים דומה מעט להבדל הקיים בין הטרמינלים המרכזיים לפרוקסימליים
תלות הפוטנציאציה המקסימלית במרחק מהאקסון המרכזי
ים אין הבדל עקרוני בקינטיקת התפתחות רמות הרוויה של השחרור הסינפטי בין טרמינל
וקרוב לודאי ששני ממוצעים אלה מייצגים ספקטרום רציף של כל , הקרובים לאקסון למרוחקים ממנו
ערכי הפוטנציאציה הנמוכים בטרמינלים המרוחקים . (fig.8E)אוכלוסיית הטרמינלים המרכזיים
בהתפתחות התגובות בטרמינלים )על ציר התדר(בתדרים הנמוכים מבטאים את האיחור היחסי
הערכים המוחלטים של הפוטנציאציה אינם מדויקים עבור טרמינלים אלה כיוון שהתכולה . אלה
למרות זאת ניתן לקבל מעקום הפוטנציאציה מדד . הקואנטלית שלהם בתדרים נמוכים שואפת לאפס
השחרור הסינפטי בטרמינלים . לגבי אופן התפתחות השחרור הסינפטי כתלות בתדר הגירוי
אך מתפתח בקצב מעט גבוה יותר ביחס , כאמור בתדרים גבוהים יותרהמרוחקים מתחיל
ההבדל בין שתי תת אוכלוסיות אלה דומה מעט להבדל בין . לטרמינלים הקרובים לאקסון
פרט לכך שהשחרור הסינפטי מכלל הטרמינלים משתי תת , הטרמינלים הפרוקסימליים למרכזיים
ניראה שקיים יחס ישר בין מרחק הטרמינל . וי גבוהיםל לא מגיע לרוויה בתדרי גיר"האוכלוסיות הנ
או לחילופין שיפוע עקום התגובה לתדר , והתדר בו הוא מתחיל לשחרר, מסעיף האקסון המרכזי
.המאפיין אותו
_______________________
סיכום .3.2.3
תדר -קיים ציר הפעלה תלוי. התגובה לתדר של הטרמינלים השונים על גבי השריר איננה אחידה
ועם עליית , כך שבתדרים נמוכים השחרור הסינפטי הוא בעיקר בסיבים פרוקסימליים, אורכי ברור

40 התגובה לגירוי בתדר בתנאי ביקורת_________________________________________________________________________________
כשנקודת המפנה היא , תדר הגירוי השחרור הסינפטי עובר בהדרגה לטרמינלים בסיבים המרכזיים
. בכל סיב וסיב באזור המרכזי של השריר, רוחבי, קיים ציר הפעלה נוסף. 10Hzסביב תדר של
ומתחילים לשחרר , רמינלים המרכזיים הקרובים יותר לאקסון רגישים יותר לגירוי בתדרים נמוכיםהט
בתדרים גבוהים יותר מגויסים באופן הדרגתי הטרמינלים המרוחקים . מוקדם יותר על ציר התדר
טרמינלים אלה מתאפיינים בסף תדר גבוה יחסית עם תגובה מעט חדה ). לטרלית לאקסון(יותר
.הבאה לביטוי בשיפוע החד יותר של עקום התגובה לתדר, עבר לסף זהיותר מ
השחרור הסינפטי במופע העלייה .3.3, החלק הקודם של הפרק התייחס לערכי הרוויה של השחרור הסינפטי כפונקציה של תדר הגירוי
מהשריר EMGרישומי . גירויים ויותר ברציפות10 -אולם על מנת להגיע לערכים אלה נדרשים כ
המוטורנוירון יורה בתדרים גבוהים למשך פרקי זמן קצרים , מראים שבזמן פתיחה מהירה של הצבת
ירי מהיר . )Crider & Cooper, 1999(המכילים לכל היותר מספר תגובות בודדות ברציפות , בלבד
ומכאן , הקודםוקצר זה לא מספיק על מנת לאפשר לתגובה להתפתח לרמות הרוויה שתוארו בחלק
לאפיון מופע הרוויה של השחרור . החשיבות הרבה של תאור מופע העלייה של השחרור הסינפטי
חשיבות רבה כיוון שהוא מצביע בצורה ברורה ופשוטה על , כפי שנעשה בחלק הקודם, הסינפטי
של " כוח המניע"ובכך נותן מדד ל, רמת השחרור הסינפטי אליה שואף טרמינל נתון בכל תדר גירוי
על מנת להבין בתנאים פיזיולוגיים את ההבדל , למרות זאת. השחרור הסינפטי כתלות בתדר הגירוי
בתרומה של הטרמינלים השונים לשחרור הסינפטי וליצירת המתח המכאני בשריר חשוב לנתח
ניתוח זה . ולכמת את הקינטיקה בה מתפתח השחרור הסינפטי בטרם הוא מגיע לרמת הרוויה שלו
. בחלק הנוכחי של הפרק עבור טרמינלים פרוקסימליים ומרכזייםנעשה
כימות קינטיקת התפתחות הפוטנציאציה .3.3.1
. נשאלת השאלה כיצד לכמת את קינטיקת התפתחות השחרור הסינפטי עם מתן סדרת גירויים
י "השחרור הסינפטי מתפתח באופן הניתן לתיאור ע, בתדרים הגבוהים יותר, בחלק מהטרמינלים
מאידך . מפונקציה זו ניתן לחלץ את קבוע הזמן כמדד לקינטיקת התהליך. ציה אקספוננציאליתפונק
בהם התהליך מתאים יותר , בעיקר אלה בעלי הסתברות נמוכה לשחרור, קיימים טרמינלים רבים

41 התגובה לגירוי בתדר בתנאי ביקורת_________________________________________________________________________________
קיימים . המתחילה בקינטיקה איטית דרך קצב מקסימלי ומסתיימת ברוויה, Boltzmannלפונקציית
. בהם ההתאמה לפונקציה פשוטה אינה משביעת רצוןאף מקרים
מסיבות אלה נעשה שימוש במדד אמפירי אחיד ופשוט והוא מספר הגירויים המביא לתגובה השווה
ללא התייחסות ספציפית , (½Stimulus number ½ - St.n)למחצית מערך הרוויה בתדר נתון
עבור תדר גירוי "קצב התפתחות השחרור"מדד זה משמש להלן לתיאור . לפונקציה זו או אחרת
או מספר הגירויים עבורם הפוטנציאציה של השחרור , ל מתאר רק את משך הזמן"המדד הנ. נתון
מידת הכפלת השחרור הסינפטי מגירוי (ולא את העצמה בה מתפתחת , הסינפטי ממשיכה להתפתח
:המחושב לפי הנוסחה, ציאציה הקלאסיולכן אינו מחליף את עקום הפוטנ, )לגירוי
Pn היא הפוטנציאציה של התגובה ה n.
mn - תכולה קואנטלית של התגובה ה n.
m1 - תכולה קואנטלית של התגובה הראשונה.
ל "שני המדדים הנ. להלןהפוטנציאציה ל מתאר את"עקום הפוטנציאציה הנ, במטרה למנוע בלבול
ולכן דרושים שניהם לצורך תיאור מדויק של מופע העלייה של , תלויים באופן עקרוניהם בלתי
.השחרור הסינפטי
סדרת גירויים ארוכה: קצב התפתחות השחרור .3.3.2
ושני , של טרמינל פרוקסימלי80Hz ניתן לראות תגובה לסדרת גירויים בתדר fig.9A -ב
ניתן . ערכי הרוויה של השחרור הסינפטי מהטרמינלים מנורמלים לאחד. פיינייםטרמינלים מרכזיים או
הטרמינל הפרוקסימלי מגיע : לראות הבדל ברור בקצב התפתחות התגובה בין הטרמינלים השונים
בעוד שלטרמינלים המרכזיים , למחצית מהשחרור הסינפטי המקסימלי לאחר ארבע תגובות בלבד
ניתן לראות תגובה fig.9B -ב). קו מקווקו(ע למחצית ערך הרוויה תגובות להגי14 - ו8לוקח
ערכי . 80Hz מרכזיים לסדרת גירויים בתדר של 12 - טרמינלים פרוקסימליים ו8ממוצעת של
, תגובות4הטרמינלים הפרוקסימליים מגיעים לחצי מערך הרוויה לאחר . 1 -הרוויה מנורמלים ל
ההסתברות לשחרור הנמוכה מאוד בטרמינלים . גירויים9רושים בעוד שלטרמינלים המרכזיים ד
המרכזיים הביאה לכך שהשחרור הסינפטי מטרמינלים אלה התחיל להתפתח רק לאחר מספר
מסבירה את ההתפתחות , שלא נראתה בטרמינלים הפרוקסימליים, תופעה זו. כישלונות בשחרור
. האיטית של השחרור הסינפטי בטרמינלים אלה
1mm
nP n=

42 התגובה לגירוי בתדר בתנאי ביקורת_________________________________________________________________________________
Figure 9 :התפתחות השחרור בתוך סדרת הגירויים באוכלוסיות טרמינלים שונות
A : 80דוגמא לתגובה ממוצעת של טרמינל פרוקסימלי ושני טרמינלים מרכזיים לסדרת גירויים בתדרHz) כל תגובה היא
) עיגולים(ניתן לראות שהטרמינל הפרוקסימלי . 1 -ערך הרוויה של כל התגובות מנורמל ל). סדרות גירויים זהות8ממוצע של
ריבועים ( בטרמינלים המרכזיים 14 - ו8 -בהשוואה ל, גירויים בלבד4ך תו) קו מקווקו(מגיע למחצית התגובה המקסימלית
לסדרת גירויים ) n=12, ריבועים(ומרכזיים ) n=8, עיגולים( של טרמינלים פרוקסימליים 1 -תגובה מנורמלת ל: B). ומעויינים
גובה לסדרות של חמישה בת, ) לעילA -מ(דוגמא לרישומים מטרמינל פרוקסימלי ומרכזי : 80Hz (mean±sd) .Cבתדר
: D. ניתן לראות את ריבוי הכישלונות היחסי בטרמינל המרכזי. ENTC -הכוכבית מציינת את אות ה. 100Hzגירויים בתדר
נגזרת על ציר : E (mean±se)משלוש אוכלוסיות שונות של טרמינלים ) לעילC( בתגובה לסדרת גירויים קצרה (m)השחרור
ניתן לראות את ההבדל . בתגובה החמישית1 -הנגזרות מנורמלות להתחלה מאפס והגעה ל. D -הגירויים של התגובות מ
אולם , לגירויquantaהטרמינלים הפרוקסימליים צוברים יותר : הברור בקינטיקת התפתחות השחרור בין קבוצות הטרמינלים
הטרמינלים המרכזיים המרוחקים עקומת. D -עקומות הפוטנציאציה של התגובות מ: F. מתקרבים מהר יותר לרוויה
.ומגיעה לערכים גבוהים יותר, היא הקעורה ביותר מבין שלושת העקומות) מעויינים(
0 10 20 300.0
0.5
1.0
stimulus No.
norm
aliz
ed re
leas
e
proximal terminalcentral terminal 1central terminal 2
0 10 20 300.0
0.5
1.0
norm
aliz
ed re
leas
e
stimulus No.
proximal terminalscentral terminals
1 2 3 4 502468101214
stimulus No.
m
proximal "close" central "remote" central
1 2 3 4 50.00.20.40.60.81.01.2
m n
or. d
eriv
ativ
e
stimulus No.1 2 3 4 5
01
5
10
15
B.A.
*
central:
proximal:
pote
ntia
tion
stimulus No.
F.E.D.
C.
2ms
0.5nA
100Hz
*

43 התגובה לגירוי בתדר בתנאי ביקורת_________________________________________________________________________________
מתחת (30Hz -קצב התפתחות זה היה איטי יותר במידה דומה בכל טווח התדרים הגבוהים מ
מגמת לתדר זה המדידה פחות מדויקת מאחר והשונות הגבוהה בין התגובות בסדרה מטשטשת את
בתדרים אלה (½St.n)מספר הגירויים למחצית תגובת הרוויה ).העלייה הכללית בשחרור הסינפטי
והיה גבוה יותר באופן מובהק , (8.5±0.64) בטרמינלים המרכזיים 7-15נע בין
(mean±se;p<0.001;Mann-Whitney)3-7שנע בין , מזה של הטרמינלים הפרוקסימליים
בין שתי האוכלוסיות הדגימה שונות גבוהה יותר באופן משמעותי ½St.nת השוואת שונו. (3.9±0.1)
מקור שונות . (p<0.001; Homogeneity of variance test)באוכלוסיית הטרמינלים המרכזיים
מוגברת זו טמון קרוב לודאי בהבדלים הניכרים בשחרור הסינפטי בין הטרמינלים המרכזיים כתלות
יש לציין שבטרמינלים הפרוקסימליים קצב ). לעיל3.2.2(רכזי בריחוקם מסעיף האקסון המ
עם עליית תדר הגירוי ) היו דרושים פחות גירויים(התפתחות התגובות בסדרה הפך מהיר יותר
.30Hz -בטווחים של מעל ל
מאפייני השחרור הסינפטי מאוכלוסיות טרמינלים שונות בתגובה לסדרת גירויים קצרה .3.3.3
בין הטרמינלים השונים יש למצע סדרות קצב התפתחות השחרורקת של לצורך השוואה מדוי
ראשית לא ניתן לחזור . אולם גירוי אינטנסיבי בתדרים גבוהים עלול ליצור מספר בעיות, גירויים רבות
ליכי מספר פעמים רב על סדרות גירויים אלה מאחר וגירוי ממושך בתדר גבוה עשוי לגרום לתה
שנית גירוי כזה גורם לכווץ של . כפי שהוזכר מוקדם יותר, הגברת שחרור בעלי קבוע דעיכה ארוך
על מנת . כפי שהוזכר במבוא, תא השריר העשוי לפגום ביציבות הרישום או בשלמות הטרמינל
ת להימנע מתופעות בלתי רצויות אלה נעשה שימוש בנוהל גירוי נוסף המורכב מסדרות גירויים קצרו
דוגמא לתגובות טרמינל פרוקסימלי ומרכזי לסדרות גירויים . 100Hzבנות חמישה גירויים בתדר
.fig.9C -אלה ניתן לראות ב
מאחר וקצב התפתחות השחרור הסינפטי בטרמינלים המרכזיים היה איטי יותר בכל טווח התדרים
דע שניתן להכלילו על טווח מספקת מי100Hzהבחירה בסדרת גירויים בתדר של , 30-80Hzשבין
על , סדרות בכל ניסוי500-800כ בין "סה, הגירויים ניתנו אחת לשתי שניות. רחב יותר של תדרים
מרכזיים , (n=8)פרוקסימליים : שתי הקבוצות הקודמות. שלוש אוכלוסיות שונות של טרמינלים
(n=12) ,ינפטי גבוה יחסית וקבוצה נוספת של טרמינלים מרכזיים שנבחרו הודות לשחרור ס
(n=18) .ונבחרו , הקבוצה האחרונה הורכבה מטרמינלים מרכזיים שהיו ממוקמים בסמיכות לאקסון
טרמינלים אלה נקראים להלן טרמינלים . כקבוצת ביניים לצורך השוואה לטרמינלים הפרוקסימליים

44 התגובה לגירוי בתדר בתנאי ביקורת_________________________________________________________________________________
ולה הקואנטלית כלומר התכ, ן התגובה הייתה יציבה בזמןבכל הטרמינלים להל ".חזקים"מרכזיים
.שמרה על ממוצע קבוע לאורך כל סדרת הגירויים
ניתן לראות שהתכולה הקואנטלית בין הטרמינלים הפרוקסימליים למרכזיים נעה בטווח fig.9D -ב
1.16-9.1ערכי התכולה הקואנטלית של התגובה הראשונה עד לחמישית נע בין . של סדר גודל
השחרור הסינפטי . במרכזיים0.07-0.95 -בהשוואה ל, קואנטה בטרמינלים הפרוקסימליים
0.6-6.9עם ערכים בין , דומה יותר לזה של הפרוקסימליים" חזקים"מהטרמינלים המרכזיים ה
. קואנטה
בתגובה לסדרת גירויים קצרה קצב התפתחות השחרור .3.3.4
ונורמלו לערך , fig.9D -חושבו נגזרות התגובות מ שחרורההתפתחות קצבעל מנת להשוות בין
היתרון בעקום הנגזרת טמון בכך שהוא . (fig.9E) 1 - וערך סופי השווה ל0 -התחלתי השווה ל
ובנוסף את קצב , בדמות הערכים המוחלטים שלו-מכמת את העלייה בעצמת הפוטנציאציה
עקום נגזרת תגובת הטרמינלים הפרוקסימליים מגיע . העקום בדמות מיקום השיא של-ההתפתחות
. כלומר מתגובה זו מתחיל השחרור הסינפטי להתקרב יותר לערכי הרוויה, לשיא בתגובה הרביעית
כפי שצוין , ממצא זה מתיישב עם הגעה לחצי מערך הרוויה תוך ארבעה גירויים בטרמינלים אלה
אולם , מתנהגים באופן דומה" חזקים"מינלים המרכזיים ההטר. 80Hzקודם לכן עבור תדר גירוי של
התבוננות . כלומר התגובה טרם הגיעה לקצב ההתפתחות המקסימלי, הנגזרת לא מגיעה למקסימום
בנגזרת השחרור הסינפטי מהטרמינלים המרכזיים מלמדת שהתגובה בהם מתפתחת בקצב איטי
יש לציין שחלק . מצאים הקודמיםומאששת את המ, יותר בהשוואה לשתי הקבוצות הקודמות
מהטרמינלים המרכזיים לא נכללו בממוצע האחרון כיוון שרוב התגובות מתוך החמש היו כישלונות
. כלומר קצב ההתפתחות בפועל הוא אף איטי יותר מהמופיע בממוצע זה, )כרבע מהטרמינלים(
עצמת הפוטנציאציה באוכלוסיות שונות של טרמינלים
וכלוסיות שונות של טרמינלים מראה שהפוטנציאציה בטרמינלים המרכזיים השוואה בין א
קרוב לודאי שנטייה זו נובעת . (fig.9F)מתפתחת לערכים גבוהים יותר מאשר בפרוקסימליים
מכנה . לתגובה הראשונה בסדרה, מההגדרה המתמטית של פוטנציאציה כיחס בין תגובה נתונה
ועקב כך , ך מאד בהתאם לבחירה הסובייקטיבית של הטרמינלל עשוי לקבל ערך נמו"היחס הנ
התופעה הייתה . הפוטנציאציה מקבלת ערכים גבוהים למרות התגובות הנמוכות באופן מוחלט
. בולטת בעיקר בטרמינלים המרכזיים המרוחקים מהאקסון

45 התגובה לגירוי בתדר בתנאי ביקורת_________________________________________________________________________________
_______________________
סיכום .3.3.5
כלומר במהירות בה , בין הטרמינלים השוניםצב התפתחות השחרורקקיים הבדל משמעותי ב
משך הזמן הדרוש להתפתחות התגובה פרופורציוני לתדר . מגיעה התגובה לערך הרוויה שלה
מספר הגירויים הדרוש להגיע למחצית מערך הרוויה נע 40-80Hzכך שבטווח התדרים בין , הגירוי
קסימליים קיימת נטייה לירידת מספר הגירויים למחצית בטרמינלים הפרו(סביב ערך קבוע בקירוב
גירויים לעומת 3.9±0.1בטרמינלים הפרוקסימליים דרושים ). התגובה עם עליית תדר הגירוי
השוואה בין שלוש קבוצות טרמינלים מדגימה קשר ישיר בין . בטרמינלים המרכזיים8.5±0.64
התגובה , יותר" חזקים"רמינלים ה בט-התכולה הקואנטלית לקינטיקת התפתחות התגובה
עצמת הפוטנציאציה בטרמינלים המרכזיים הופכת מהתגובה השלישית . מתפתחת בקצב מהיר יותר
הודות לתגובה ראשונה בעלת ערך , ואילך לגבוהה ביותר בהשוואה לקבוצות הטרמינלים האחרות
וצות הטרמינלים השונות כאל אפשר ונכון יותר להתייחס לשוני בין עקומי הפוטנציאציה של קב. אפסי
השחרור מהטרמינלים : הסטה של נקודת ההתחלה ממנה מתחיל להתפתח השחרור הסינפטי
ומגיע מוקדם יותר , הפרוקסימליים מוסט שמאלה כלומר מתחיל להתפתח מערכים גבוהים יותר
.לרוויה
.הדיון בממצאי פרק זה מופיע במאוחד עם הדיון בממצאי הפרק הבא

46
השפעות שינויים בכניסת הסידן לטרמינל על התגובה .4 לגירוי בתדר
___________________________________________________________________
מבוא .4.1וקיימים תימוכין , כניסת הסידן לתוך הטרמינל תופסת מקום מרכזי בתהליך השחרור הסינפטי
הסידן לתוך הטרמינל בתגובה לגירוי גבוה יותר ביחס רבים לכך שבטרמינלים הפרוקסימליים שטף
נשאלת השאלה אילו ). הבדלים פיזיולוגיים נוספים 1.2.4: ראה מבוא(לזה שבטרמינלים המרכזיים
מההבדלים שתוארו בעקום התגובה לתדר בין הטרמינלים הפרוקסימליים למרכזיים ניתן לייחס
הדרך . להשפיע על עוצמת כניסת הסידן לטרמינלקיימות מספר דרכים . לשינויים בכניסת הסידן
דרך זו יעילה בעיקר לצורך הפחתה בכניסת . הקלה היא באמצעות שינוי בריכוז הסידן החוץ תאי
הגדלת הריכוז החוץ תאי של סידן אינה מגבירה באופן . ואכן מנוצלת כאן למטרה זו, הסידן לטרמינל
שחרור מתקרבת לרוויה בריכוזי סידן העולים על -ןמשמעותי את השחרור הסינפטי היות והתלות סיד
דרך שונה המאפשרת להתגבר . )Parnas et al., 1982c(הריכוז הפיזיולוגי בטרמינלים המרכזיים
הגירוי הישיר מאפשר שליטה על עצמת ומשך . ל היא גירוי ישיר של הטרמינל"על המגבלה הנ
, )Dudel, 1983(כך שליטה על שטף הסידן לתוך הטרמינל ובהתאם ל, הדפולריזציה בטרמינל
לפיכך השימוש בגירוי ישיר מנוצל כאן לצורך בדיקת השפעות ההגברה בכניסת . והשחרור ממנו
.סידן על התגובה לתדר של הטרמינלים
הגברת כניסת הסידן לטרמינל באמצעות גירוי ישיר וההשפעה על .4.2 התגובה לתדר
התגובה לגירוי בתדרעל שיפוע עקום הגירוי הישיר עת עוצמתהשפ .4.2.1
לאחר . עקום התגובה לתדר תוך שימוש בגירוי האקסוןמהטרמינל בניסויים אלה ראשית נרשם
ונרשם עקום התגובה לתדר של הטרמינל במספר , TTX (0.5µM)מכן נעשתה אפליקציה של
, 0.5msעמד על הישירמשך הגירוי . 2µA - ועד ל0.5µA בין הטווחים, ישיררויעוצמות גי
עשויה להתרחש בזמן ה ,במטרה למנוע ככל הניתן הופעה של תופעות כמו פסיליטציה מוקדמת

47 השפעות שינויים בכניסת הסידן לטרמינל על התגובה לגירוי בתדר_________________________________________________________________________________
לתגובות המעוררות לסירוגין על דוגמא . )Katz & Miledi, 1971; Dudel, 1983( הגירוי עצמו
בגירוי ביקורתרישום לאחר . fig.10A -רוי עצב וגירוי ישיר בטרמינל מרכזי ניתן לראות בידי גי
הגוררת עקום תגובה ישיר גירוי נבחרה עצמת ראשית וTTXנעשתה אפליקציה של אקסון
ביקורת זו נועדה לוודא שאכן שני סוגי הגירוי ). fig.10Bראה ( לזה של גירוי האקסון זהה לתדר
עקום זה הושג לרוב תוך שימוש . מניבים עקום תגובה לתדר זהה) ון וישיראקס(השונים
שחרור תגביר את הלאחר מכן הוגברה עצמת הגירוי הישיר כך ש .0.5-0.8µAבעוצמות גירוי בין
לשם .כלומר ביחס לגירוי האקסון, זה שעורר קודם לכןביחס ל לפחות )2-5 -פי(מהטרמינל
. fig.10C,D, למטהfig.10A -לכך ניתן לראות בדוגמא . 1.5-2 -הועלתה עוצמת הגירוי פי
את עקום התגובה לתדר " להפוך"מתוך ניסיון , ניסויים אלה נערכו על שישה טרמינלים מרכזיים
הרישום נעשה במתכוון . של טרמינלים אלה לדומה יותר לזה של טרמינלים פרוקסימליים
ההבדלים בין מאפייני עקום : סיבות עיקריותמאוכלוסיית הטרמינלים המרוחקים מהאקסון משתי
השחרור בתדר של אוכלוסיית טרמינלים זו לאלה של עקום הטרמינלים הפרוקסימליים הם
ורחוקה , והתכולה הקואנטלית של הטרמינלים המרוחקים היא הנמוכה ביותר, ביותרקיצונייםה
מראה שחיזוק עצמת (fig.10C,D)שחרור -התבוננות בשיפוע העקום תדר. מהגעה לערכי רוויה
שהייתה הפיכה עם הפחתת עוצמת , הביאה לירידה בערך השיפוע, הגירוי תוך הגברת השחרור
ומגיע לערכים סופיים , )בתדרים נמוכים יותר(השחרור הסינפטי מתפתח מוקדם יותר . הגירוי
ומצביעה , בהסתמך על מסקנות הפרק הקודם, ירידה זו בערך השיפוע היא צפויה. גבוהים יותר
ביחס , )הודות לגירוי החזק יותר(על התחזקות תרומת ריכוז הסידן שבעקבות הגירוי לשחרור
ממצא זה מחזק את מסקנות הפרק הקודם בדבר . לתרומת ריכוז הסידן הממוצע בטרמינל
אוכלוסיות שבין הבדלים שינויים אלה דומים ל.שחרור-משמעותו של שיפוע העקום תדר
עוצמת הגירוי הישיר נבחרה כך . מהותימלבד הבדל אחד , קסימליים למרכזייםהטרמינלים הפרו
בתנאים אלה . שתכפיל ואף יותר מכך את השחרור הסינפטי מהטרמינלים בתדרים הגבוהים
לזה של חס ביאולם עדיין נותר נמוך , השחרור בתדרים הנמוכים עולה בסדר גודל דומה
לשם השוואה בתדר גירוי של . (50%<)בוי כישלונות ומתאפיין ברי, הטרמינלים הפרוקסימליים
1Hz , ראהfig.10A שמאלי תחתון מול fig.6Aבכל ששת הפרפרטים בהם בוצעו . שמאלי תחתון
.) ריבועים מלאיםfig.11Aראה (הניסויים בגירוי ישיר התקבלו תוצאות דומות

48 השפעות שינויים בכניסת הסידן לטרמינל על התגובה לגירוי בתדר_________________________________________________________________________________
Figure 10 :צב התגובה לתדר בגירוי ישיר מול גירוי ע
A : אקסון וישיר עם דוגמאות לרישומי חמישה עשר התגובות הראשונות לסדרות בגירויTTX , 1בתדרים שלHz ,ו-
80Hz .הרישומים לקוחים מהטרמינל ב- B,C0.5הגירוי ועוצמת משך , לעילms ,1.2µA . הסיגנל הכוכבית מציינת את
עקום תגובה : B. קיימים כישלונות מרובים גם בגירוי ישיר1Hz שבתדר גירוי של ניתן לראות. בגירוי העצבהפרסינפטי
עוצמת הגירוי הישיר נקבעה כך שתניב תגובה זהה ככל . )0.5ms ,0.6µA (בגירוי עצב וישיר, לתדר של טרמינל מרכזי
לאחר חיזוק עוצמת הגירוי , B - בגירוי עצב וישיר של הטרמינל בעקומי תגובה לתדר: C.האפשר לזו של גירוי העצב
המצוינת , ניתן לראות את הירידה בשיפוע הלוגריתמי של עקומת הגירוי הישיר עם העלייה בשחרור. 1.2µA -להישיר
הגירוי הישיר עוצמתמשך ו. בגירוי עצב וישיר, דוגמא לתגובת טרמינל מרכזי נוסף: D. באחוזים ביחס לביקורת
(1.4µA;0.5ms) 80 - כך שהתגובה בונקבעHzהעקום המקווקו . ויותר מהתגובה לגירוי העצב3 - תהיה גבוהה יותר פי
.ממחיש את הירידה בשיפוע הלוגריתמי של עקום התגובה לתדר החל עם מתן הגירוי הישיר
מעלה את השחרור תוך שימוש בגירוי ישיר אם כן שהגברת כניסת הסידן לטרמינל סכםניתן ל
העלייה מאידך. שחרור בהתאם-ומורידה את שיפוע העקום תדר, הסינפטי בכל התדרים ביחס דומה
1 10 1000
1
2
3
93%
stim. frequency Hz
quan
tal c
onte
nt slope 1.45
Central terminal 1nerve stim."weak" direct stim.
1 10 100
0
2
4
6
8
10
12
Central terminal 1nerve stim."strong" direct stim.
stim. frequency Hz
525%
slope 1.45
slope 1.0
1 10 100
0
2
4
6
8
10
12
Central terminal 2nerve stim."strong" direct stim.normalized nerve stim.
D.C.B.
A.
stim. frequency Hz
slope 1.6
slope 1.3
400%
*
80Hz1Hz
Central terminal
Direct stim.:
Nerve stim.:
1ms
0.5nA

49 השפעות שינויים בכניסת הסידן לטרמינל על התגובה לגירוי בתדר_________________________________________________________________________________
שתי את ההבדל המשמעותי בשחרור הסינפטי בתדרים הנמוכים ביןצמצם למתקשהבכניסת סידן
.מינלים הטראוכלוסיות
ההשפעה על קצב התפתחות השחרור .4.2.2
גירוי ישיר של הטרמינלים מאפשר לבדוק את ההשפעה של כניסת סידן מוגברת על קצב
בדיקה זו מאפשרת להצביע על חלק מהגורמים . עד להגעה לשלב הרוויה, שחרורהתפתחות ה
ביחס למרכזיים שעשויים להסביר את ההתפתחות המהירה של התגובה בטרמינלים הפרוקסימליים
(fig.9A,B) .ב- fig.11B 80 מתוארת דוגמא של תגובה ממוצעת לסדרת גירויים בתדר שלHz ,
גירויים 7ונדרשו , תחת גירוי ישיר התכולה הקואנטלית הוכפלה. בגירוי אקסון וישיר באותו טרמינל
תגובת גירוי לצורך ההשוואה נורמלה ( בתגובת גירוי האקסון 12 -בהשוואה ל, להגיע למחציתה
בכל הטרמינלים שגורו ישירות בעצמה ). ומופיעה כעקומה מקווקוות, האקסון לתגובת הגירוי הישיר
(½St.n) קצב התפתחות השחרור, המעוררת שחרור סינפטי גבוה יותר מזה שעורר גירוי האקסון
גירויים 7.3±1.7 -ל, גירויים למחצית ערך הרוויה12.6±2.2 -ובממוצע מ, ירד באופן ברור ומובהק
ממצאים אלה מראים שכניסת סידן מוגברת מעלה את רמות . (mean±se;n=5;p=0.005)בלבד
כלומר במובן זה , תוך האצת מובהקת של קצב התפתחות השחרור, השחרור הסינפטי ברוויה
.דוחפת את דפוס השחרור של הטרמינלים המרכזיים לזה של הטרמינלים הפרוקסימליים
יכוז סידן חוץ תאי על התגובה לתדר של הטרמינליםהשפעת הפחתת ר .4.3בניסויים אלה הופחת ריכוז הסידן החוץ תאי מתוך כוונה לבחון האם הפחתת כניסת הסידן
ניסויים אלה נערכו בגירוי האקסון . לטרמינל תגרור אפקט סימטרי והפוך לזה שתואר לעיל
כך שהתאפשרה הפחתה , ומעלה" ממוצע"בטרמינלים מרכזיים שהתאפיינו בשחרור סינפטי
לאחר הפחתת ריכוז , התגובה לתדר נבדקה בביקורת. בשחרור מבלי להגיע לרמות תגובה אפסיות
מידה והפחתת ריכוז הסידן למחצית לא גררה שינוי מובהק מספיק ב( או רבעמחציתכדי הסידן ל
, ינל מרכזי בביקורת מתוארת תגובה לתדר אופיינית של טרמ fig.11C -ב. ולאחר שטיפה)בתגובה
37% -השחרור הסינפטי בתדרים הגבוהים ירד ל. ולאחר הפחתת ריכוז הסידן החוץ תאי למחצית
תוצאות דומות נרשמו . 1.68 - ל1.35 -בניסוי זה ניתן לראות עליה בשיפוע העקום מ. מהביקורת
ר הן הפיכות באופן ניסויים אלה מאשרים שאכן התופעות שנצפו בגירוי ישי. משני טרמינלים נוספים
.סימטרי

50 השפעות שינויים בכניסת הסידן לטרמינל על התגובה לגירוי בתדר_________________________________________________________________________________
Figure 11 : שינויים גירוי ישיר והשפעת
השחרור חוץ תאי עלסידן בריכוז הסינפטי
A :לשינוי , ן השינוי היחסי בשחרורסיכום הקשר בי
על העקום מופיעים . שחרור-היחסי בשיפוע העקום תדר
ניסויי ההפחתה , )ריבועים מלאים(ניסויי הגירוי הישיר
וכמו כן ההבדל , )ריבועים פתוחים(בריכוז סידן חוץ תאי
הממוצע בין הטרמינלים הקרובים לאקסון למרוחקים
מציין דעיכה הקו המקווקו. )ריבוע מוצלב(ממנו
. (r20.99), 1.53 -אקספוננציאלית עם קבוע השווה ל
B : 80 גירויים בתדר 50סדרה של (השפעת הגירוי הישיר על הפאזה העולה של השחרור בטרמינל מרכזיHz ,
1.3µA;0.7ms .( לשם השוואה מנורמלת התגובה לגירוי עצב למקסימום התגובה בגירוי ישיר)ניתן לראות). קו מקווקו
: C). קו מקווקו אופקי( בלבד 8 - גירויים ל12 -שעם הכפלת השחרור מתקצר משך ההגעה למחצית ערך הרוויה מ
ניתן לראות אפקט . )גירוי אקסון (ובריכוז סידן חוץ תאי המופחת למחצית, התגובה לתדר של טרמינל מרכזי בביקורת
.המלווה בעליה בשיפוע העקום) מצוין באחוזים(ר ירידה בשחרו: fig.10C - המופיע בהפוך לזה של הגירוי הישיר
הראתה שקיים יחס אקספוננציאלי ) גירוי ישיר והפחתת ריכוז סידן(הצגה גרפית של כלל הניסויים
-לשינוי בשיפוע העקום תדר, טי הנגרם עקב השינוי בכניסת הסידןהפוך בין השינוי בשחרור הסינפ
מתואר fig.11A -ב). למשל עליה בשחרור הסינפטי עם ירידה בשיפוע העקום(שחרור של הטרמינל
ניתן . השינוי היחסי בשיפוע העקום כפונקציה של השינוי היחסי בשחרור הסינפטי בתדרים הגבוהים
ביחס ( שיפוע העקוםבחלה ירידה חדה יותר , יר חזק יותר באופן יחסילראות שככל שהגירוי היש
נקודה בודדת המציינת את היחס בין fig.11A -כמו כן מופיעות ב). הפוך לעליה בשחרור הסינפטי
לטרמינלים הקרובים תכולה קואנטלית (הטרמינלים המרכזיים הקרובים לאקסון למרוחקים ממנו
1 10 1000
2
4
6
C.
B.A.
stim. frequency Hz
quan
tal c
onte
nt
37%slope 1.68
slope 1.35control50% calcium
0 1 2 3 4 5
0.6
0.8
1.0
1.2
1.4
quantal content at 80Hz ratio
slope
ratio
direct stim.reduced Calciumclose/ remote from axon
0 5 10 15 20 25 300
1
2
3
4
5
ampl
itude
nA
stimulus. No.
nerve stim.direct stim.nerve, normalized

51 השפעות שינויים בכניסת הסידן לטרמינל על התגובה לגירוי בתדר_________________________________________________________________________________
הנקודות המייצגות את הטרמינלים מכל סדרות ניסויים אלה ).יותר וךנמ שיפועותר עם גבוהה י
. 1.53 -בעלת קבוע השווה ל, נופלות על עקומת דעיכה אקספוננציאלית
מלווים בשינוי , ניסויים אלה מראים שעליה או ירידה בכניסת הסידן לטרמינל בתגובה לגירוי
שיפוע העקום . הבולט בעיקר בתדרים הגבוהים, הרוויה של השחרור הסינפטימשמעותי ברמות
. שחרור משתנה ביחס אקספוננציאלי הפוך לשינוי החל בשחרור הסינפטי בתדרים הגבוהים-תדר
_______________________
סיכום .4.4הסידן תרומתם העיקרית של ניסויי הגירוי הישיר באה לביטוי בהשפעה הבו זמנית של כניסת
הגברה . וכך קושרת אותם יחדיו, המוגברת על שלושה מאפיינים שונים של השחרור הסינפטי
, בכניסת הסידן לטרמינל גוררת עלייה מהירה יותר של השחרור הסינפטי לרמות רוויה גבוהות יותר
אפקט סימטרי ניסויי ההפחתה בריכוז הסידן מראים. שחרור- העקום תדרשיפועבהמלווים בירידה
ל מפשט את ההסתכלות הכוללת על השחרור הסינפטי "הקשר בין שלושת המאפיינים הנ. הפוךו
מאידך אין בכוחם של ניסויים אלה להסביר חלק מההבדלים העקרוניים בין . בתגובה לגירוי בתדר
למרות העלייה הניכרת בשחרור . עקומי התגובה לתדר של טרמינלים פרוקסימליים ומרכזיים
בניגוד לרוויה , לרוויה בטרמינלים המרכזיים להגעההסינפטי תחת גירוי ישיר לא נרשמה נטייה שלו
כמו כן העלייה בשחרור . 50Hz -המתפתחת בטרמינלים הפרוקסימליים בתדרי גירוי של מעל ל
,ה לזה של גירוי האקסוןהבעוצמה חזקה מזו הגורמת לשחרור ז, הסינפטי תחת גירוי ישיר
ולכן מתקשה להסביר את היתרון , תרחשה בסדר גודל דומה על פני כל ספקטרום תדרי הגירויה
.נמוכיםגירוי תדרי לבתגובההגדול של הטרמינלים הפרוקסימליים בשחרור
דיון .4.5
יציבות הרישום .4.5.1
גע תוך גירוי האקסון בתדרים גבוהים עשויה להיפ יציבות הרישום מטרמינלים אקסיטטוריים
- שינויים בערך העלול לגרור, כווץ אפשרי של תאי השריר בתנאים אלה) 1. משני טעמים עיקריים
RS ,מאותן סיבות תתכן פגיעה פיזית בשלמות ממברנת ) 2. הטיה במדידת התגובותובעקבות זאת
רה יותר בסיבים הבעיה חמו. כדומהעליה בריכוז הסידן התוך תאי ו, ובהמשך דפולריזציההטרמינל

52 השפעות שינויים בכניסת הסידן לטרמינל על התגובה לגירוי בתדר_________________________________________________________________________________
מספר הגירויים בכל סידרה ל"הנ בעק. המגיבים לגירוי בכווץ חזק יותר מהמרכזיים, הפרוקסימליים
בתוך סדרת גירויים ולראיה מספר תימוכין לכך שבתנאים אלה הרישום נותר יציב, 50 -הוגבל ל
וכמו כן נותר יציב , גודל הקואנטום נותר זהה בכל התגובות במהלך סדרת הגירויים) 1 :וביניהן
, המשמש כפולס קליברציה טבעי לצורך העניין, AP/ENTC -משרעת ה) 2. במהלך הניסוי כולו
תוך שימוש גירוי ישיר ) 3. (fig.6A, fig.10A)נותרה קבועה בכל התגובות במהלך סדרת הגירויים
ולכן לא גורם לכווץ , מביא להפעלה מקומית של טרמינלים ספציפיים על גבי תא השריר TTX -ב
הניבה , אקסוןהבעוצמה שגררה שחרור סינפטי זהה לזה של גירוי , העובדה שגירוי זה. כללי שלו
אקסוןהגירוי ) א: מספר דבריםדה על מעי, (fig.10B) עקום תגובה לתדר הזהה לזה של גירוי העצב
גרם לדפולריזציה רבה יותר של עוצמות אלה לאבהגירוי הישיר ) ב .לא פגם ביציבות הרישום
.בהשוואה לגירוי האקסון שהוא הגירוי הפיזיולוגיהטרמינל
עשויים להעיד על כמו כן יש לציין שבסדרות ניסויים אלה לא הייתה שכיחות רבה יותר של סימנים ה
בדומה לניסויים שאינם כוללים גירוי , הרישום נותר יציב במשך שעות: פגיעה בשלמות הטרמינל
התגובה לתדר באותו טרמינל הניבו עקום שחרור זההעקום בדיקות חוזרות של ,בתדר גבוה
(fig.4) ,0.1 שלא עלה על, לאורך כל הניסוימנוחה קבוע הופיעו בתדר יםיאירועים ספונטנHz.
.ל אכן מייצג נאמנה את השחרור הסינפטי בתדרים גבוהים"ת אלה מראות שהרישום הנתצפיו
ובחלק מהפרפרטים אף לא ניתן היה לבדוק את , בסיבים הפרוקסימליים הבעיה הייתה קשה יותר
הפרפרטים בהם כן נערכו . עקב כווץ חזק מדי, התגובה לתדר של טרמינלים מאזור זה של השריר
והטרמינלים עמדו מינוריהכווץ היה , או לחילופין, לא ניצפה כווץשבהם הניסויים היו אלה
הסיבה לשונות הרבה בנטיית השריר להתכווץ בין פרפרטים . בקריטריונים המעידים על רישום יציב
, קרוב לודאי שמדובר בתכונה המבוקרת על ידי הסרטן בהתאם לצרכים שלו. שונים לא ברורה
. וכיוצא בזה, קצב המטבוליזם שלו, ה בה נמצאהמוכתבים בהתאם לסביב
התמיינות הטרמינלים והומוגניות האוכלוסיות השונות .4.5.2
מדידות פיזיולוגיות הביאו לחלוקת הטרמינלים השונים לקבוצות נפרדות בעלות תכונות
ראה מבוא (ומאוחר יותר הודגמה מורפולוגיה אופיינית לכל קבוצה שתמכה בחלוקה זו , אופייניות
שונות גבוהה בתכולה הקואנטלית של טרמינלים מרכזיים בתלות בדברקיימות עדויות ). 1.2.2
מגוון רחב של מראות אכן והתוצאות עד כה , )Cooper et al., 1996a(במיקומם ביחס לאקסון
ל תוארה כאן תת קבוצה של "בקצה הספקטרום הנ :טרמינלים המרכזיים בעלי תכונות שונות

53 השפעות שינויים בכניסת הסידן לטרמינל על התגובה לגירוי בתדר_________________________________________________________________________________
& Atwood( שקיומם היה ידוע בעיקר על סמך עדויות מורפולוגיות (silent)" שקטים"טרמינלים
Wojtowicz, 1999( . טרמינלים אלה)האקסון בכדוגמת הטרמינל המרוחק מ- (fig.8A,C,
בקצה השני . בתדרים גבוהים) עשרות גירויים(משחררים נוירוטרנסמיטור רק בתגובה לגירוי ממושך
הדומים יותר , של ספקטרום הטרמינלים המרכזיים ניצבים הטרמינלים הסמוכים לאקסון
ותפות לכל למרות הנאמר לעיל קיימות תכונות מש. בתכונותיהם לטרמינלים הפרוקסימליים
. הפרוקסימלייםאוכלוסיית הטרמינלים אותם כקבוצה מ מפרידותה, הטרמינלים המרכזיים שנבדקו
עליה אבסולוטית קטנה , )50%כ מעל "בד(בין תכונות אלה ריבוי יחסי של כישלונות בתדרים נמוכים
ללא הופעת הגבוהים הגירויריתדשחרור ב עם עלייה חדה ב,10Hz של י גירוי עד לתדרשחרורב
.רוויה
אולי , פיזיולוגיתאוכלוסיית הטרמינלים הפרוקסימליים אחידה יותר , בניגוד לטרמינלים האחרונים
בעבודה זו נדגמה אוכלוסייה אחת . המרכזיהם מקובצים באשכולות בסמוך לאקסון הודות לאופן בו
על סמך הומוגניות זו של .השונות הפיזיולוגית ביניהם הייתה נמוכה באופן יחסיו, של טרמינלים אלה
ממצא . מהקודמתפיזיולוגית יותר אחידה אכןוסייה זו ניתן להניח שאוכל,הטרמינלים שנבדקמדגם
פרוקסימליים מחזקים את ל ,בין הטרמינלים המרכזיים הסמוכים לאקסון הפיזיולוגי דמיוןוה, זה
ההשערה שחלק גדול מההבדלים הפיזיולוגיים בין הטרמינלים כן ניתן לייחס להבדלים המורפולוגיים
.הםביני
שיפוע עקום התגובה לגירוי בתדר .4.5.3
או כפי שנעשה בעבודה , תוך תאי, ניתן לתאר את השחרור הסינפטי כפונקציה של סידן חוץ תאי
ים רמת הסידן התוך תאי בטרמינלמספר עבודות הראו באופן עקבי ש. כפונקציה של תדר הגירוי, זו
,.Cooper et al., 1995a; Tank et al( עולה ביחס ישר לעליית תדר הגירוי מרכזיים בפרפרט זהה
1995; Msghina et al., 1999; Ravin et al., 1997( השחרור תלות ולכן יש בסיס להשוות בין
שיפוע למאוד דומה שחרור-תדרהעקום שיפוע השוואה זו מראה ש. הסינפטי בשני גורמים אלה
.Ravin et alי "תואר עהאחרון כפי ש, ) בהתאמה1.6 מול 1.68(שחרור -ממוצעהעקום ריכוז סידן
.Parnas et alי "עשתוארה לתלות טובה כמו כן קיימת התאמה . באותם טרמינלים) 1999(
)1982a(בשני המקרים השיפוע . באוכלוסיית הטרמינלים המרכזיים בין ריכוז סידן חוץ תאי לשחרור
;Dudel, 1981(ערך דומה נמדד גם בעבודות נוספות . 1.6 -ושווה ל, הלוגריתמי המתקבל הוא זהה

54 השפעות שינויים בכניסת הסידן לטרמינל על התגובה לגירוי בתדר_________________________________________________________________________________
Parnas et al., 1982a( . י " עו שנמדדלערכיםקיימת התאמה טובה גםWright et al. )Wright et
al., 1996a(בקירוב2 -שווה ליה שיפוע ההשם ערך , בקיבוע מתח באותם טרמינלים .
-סידןריכוז שחרור ועקומת -גם עבור הטרמינלים הפרוקסימליים קיים דמיון רב בין עקומת תדר
טרמינלים אלה . )Parnas et al., 1982c( .Parnas et alי "פי שזו מתוארת עכ, )חוץ תאי (שחרור
ההסתברות . של המרכזייםה השווה לכמחצית מזשיפועעם , המשחררים יותר בכל טווח העקומ
ולמעשה אין סף תדר להגברת השחרור הסינפטי כפי שקיים בטרמינלים , לשחרור בהם גבוהה
. המרכזיים
ראשית היא מאפשרת ללמוד . ל הוא תוצאה חשובה של עבודה זו"מיון בין שתי העקומות הנהד
, )Y (גירוי הסידן הנכנס לטרמינל בעקבות ה לשחרור שלתרומהעל מידת העקום באמצעות שיפוע
Ravin) (ss[+Ca2]) לזו של ריכוז הסידן הממוצע המתפתח בטרמינלים במהלך גירוי בתדרים גבוהים
et al.,1997,1999) . בין אוכלוסיות הטרמינלים השונות שחרור-תדרההבדלים בין שיפועי עקומי
יותרונמוכה, לשחרור בטרמינלים הפרוקסימלייםYשל יחסית מצביעים על תרומה גבוהה
.הולכת ופוחתת ככל שמרחק הטרמינל מהאקסון המרכזי עולהה, בטרמינלים המרכזיים
הגדלת -ל"מלמדת שלשני הגורמים הנשחרור -ותדרשחרור - סידן:מעבר לכך הדמיון בין העקומות
השחרור יש אפקט זהה על , והגברת תדר הגירוי מאידך, מחדהממוצע תאי תוךהריכוז הסידן
עד הגעת התגובה (ניתן להסיק שתדר הגירוי ל "הנבין העקומות הרב על סמך הדמיון . הסינפטי
תואמות תוצאות אלה . ללא ערך מוסף פרט לכך, י ליניארי להגברת ריכוז הסידןמשמש כל, )לרוויה
המראות שהשחרור תלוי בריכוז הסידן התוך תאי ) fig.6Aראה , Felmy et al. )2003 תוצאות של
,Stanley( ממצאים קודמים מסקנה זו תואמת גם. ללא תלות בהסטוריה של גירוי קודם, בלבד
1986; Liu & Stanley, 1995( על בתדר והגדלת ריכוז סידן גירוי גורמיםה שהראו שפעולת שני
מפחיתה בהתאם ) חוץ תאי(כלומר הגברת ריכוז הסידן , מנגנון השחרור באה אחד על חשבון השני
.ולהיפך, יכולת הפסיליטציה של הטרמינלאת
שונותטרמינלים וכלוסיותמאשחרור גורמים אפשריים להבדלים בין ה .4.5.4
רוב החוקרים סבורים שההבדלים המורפולוגיים בין הטרמינלים מסבירים רק , כפי שצוין במבוא
מאחר ושינויים בכניסת הסידן . ולכן דרושים הסברים נוספים, ביניהםחלק מההבדל הפיזיולוגי הניכר
, לתדר של הטרמינליםה שונים בין התגובלטרמינל משפיעים ישירות על חלק מהמאפיינים שנמצאו
.האחראי להבדלים אלההרי שסידן הוא בחירה טבעית כגורם

55 השפעות שינויים בכניסת הסידן לטרמינל על התגובה לגירוי בתדר_________________________________________________________________________________
Parnas :השחרור בין הטרמינליםמנגנון הבדלים בריכוז הסידן במנוחה או בקואופרטיביות
)Parnas et al., 1982c(שניתן , רור הסינפטי מהטרמינלעל סמך מודל המתאר את השח, הראו
בין הטרמינלים ברמות סידן גבוהות יותר במנוחה בטרמינלים השחרורהבדלילתלות את
ללא לעת עתה אין עדויות לכך ש. של מנגנון השחרוראו קואופרטיביות נמוכה יותר/ו, הפרוקסימליים
תאי בין הטרמינלים השונים אכן קיימים הבדלים מהותיים ברמות המנוחה של סידן תוךמתן גירוי
)Cooper et al., 1995a( . קואופרטיביות נמוכה יותר של הטרמינלים הפרוקסימליים עשויה
טרם נמצאו עדויות המצביעות על אולם , להתאים למאפייני התגובה לתדר של טרמינלים אלה
השיפוע המתון יותר של העקום דאי שקרוב לולכן .הבדלים בין מנגנוני השחרור ברמה המולקולרית
להלןבדלים בכניסת הסידן לטרמינלהלמשל ,טרמינלים אלה נובע מגורמים אחריםבשחרור -תדר
)Parnas et al., 1982c; Stanley, 1986; Cooper et al., 1995a(, הבדלים ולא משקפים
. בקואופרטיביות המנגנון עצמומולקולריים
בכניסת סידן הבדליםמצביעות על המספר תוצאות להלן :הבדלים בכניסת סידן לטרמינלים
. השונותלהבדלים בשחרור הסינפטי בין אוכלוסיות הטרמינליםכגורם התורם לטרמינל
כניסת הסידן באמצעות מתן גירוי חזק מהותית מזה הישיר הדגימו בבירור שהגברתניסויי הגירוי) 1
Parnasבהתאמה לממצאים קודמים של , מביא לשחרור מוגבר ,ת הביקורתגובתאת המניב
)Parnas et al., 1982c(, זו הפיכה עם הפחתה תופעה . שחרור-תדר ולירידה בשיפוע העקום
של לשחרור כתרומה דומיננטית יותר בשיפועניתן לפרש ירידה זו. צמת הגירוי הישירחוזרת של עו
ניסויי ההפחתה תוצאות .ביחס לתרומת ריכוז הסידן הממוצע בטרמינל, נכנס בתגובה לגירוי הסידןה
התוצאות ניסויים אלה עולים בקנה אחד עם. זו אכן הפיכהופעהתבריכוז הסידן החוץ תאי הדגימו ש
לאחר חרור ש-בשיפוע התלות ריכוז סידן ממוצעדומה המדגימות עליה , )Ravin et al.) 1999 של
, )Zucker )1974נצפתה גם בעבודה של תופעה דומה .הפחתת ריכוז הסידן החוץ תאי לכדי רבע
ניסת בדק בטרמינלים מהשריר הפותח את הצבת הגדולה באותו פרפרט את האפקט של ירידה בכש
-מ(בעבודה זו הקטנת ריכוז הסידן החוץ תאי . סידן על שחרור סינפטי בתגובה לגירוי בתדר
13.5mM1 - לmM (בהתאמה , שחרור-הביאה לירידה בשחרור עם עליה בשיפוע העקום תדר
.לתוצאה שהתקבלה בעבודה הנוכחית
סדרת הלךמ בשחרורקצב התפתחות ההשפעת כניסת הסידן על טמונה בתוצאה נוספת ) 2
3.14הדגימו עליה ממוצעת של פי ניסויי הגירוי הישיר תוצאות .ועל רמת הרוויה שלו, הגירויים
בקצב התפתחות 1.73ה של פי המלווה בהאצ, )80Hzבתדר גירוי של (בשחרור המקסימלי

56 השפעות שינויים בכניסת הסידן לטרמינל על התגובה לגירוי בתדר_________________________________________________________________________________
כלומר לא רק שהשחרור מתפתח לערכים גבוהים , ) גירויים7.3 - ל12.6 - ירד מ½St.n (שחרורה
תוצאה זו עשויה להסביר את ההבדל הניכר בקצב התפתחות . הוא גם מגיע לשם מהר יותר, תריו
-גבוה פיברוויה באחרונים השחרור : השחרור בין אוכלוסיית הטרמינלים המרכזיים לפרוקסימליים
נתונים אלה מצביעים על כך שכניסת סידן מוגברת עשויה להסביר . 2.1 -ומואץ פי, בממוצע2.8
נרמול ההאצה בקצב התפתחות , מעבר לכך. טרמינליםאוכלוסיות הכר מההבדלים בין חלק ני
-מראה שקצב התפתחות השחרור הואץ פי, ליחידות של קואנטהבניסויי הגירוי הישיר השחרור
מתוך כלל הפרמטרים המתארים את דפוס השחרור בתגובה לגירוי זה מצביע על כך שנתון. 5.4
.פרמטר המושפע ביותר משינויים בכניסת הסידן לטרמינלהור הוא קצב התפתחות השחר, בתדר
אור עדויות אלה ניתן לייחס להבדלים בכניסת סידן לטרמינל שלושה מההבדלים הברורים ל, לסיכום
) 2 .התגובה לגירוי בתדרעקום בשיפוע הבדלים )1 :שחרור הסינפטי בין הטרמינליםדפוסי הב
הבדלים ברמות ) 3 ).הבדלים בשיפוע העקוםמה ם ישירותהנגזרי( הבדלים בסף התדר לשחרור
מהלך בשחרורהבדלים בקצב התפתחות ה) 4. הרוויה של השחרור הסינפטי בתדרי גירוי גבוהים
.סדרת הגירויים
קיימות . תמיכות פיזיולוגיותלהבדלים בכניסת הסידן בין אוכלוסיות הטרמינלים השונות מספר
ציאל הפעולה לטרמינלים הפרוקסימליים חזקה יותר מאשר עדויות רבות לכך שפלישת פוטנ
וניתן להניח שהדפולריזציה החזקה יותר גוררת כניסת סידן רבה יותר בהתאם , במרכזיים
)Atwood & Bittner, 1971; Parnas et al., 1982c; Dudel, 1983( . בעבודה זו לא נבדקה
כמו כן . ת העבודה הנוכחית לא מספקות תמיכה ישירה לכךמשרעת פוטנציאל הפעולה כך שתוצאו
, קיימות עדויות ברורות להצטברות מהירה יותר ומוגברת של סידן בטרמינלים הפרוקסימליים
הסיבות . )Cooper et al., 1995a(כבר בטווח התדרים הנמוכים , בתגובה לגירויים חוזרים
,טרם ברורים במלואם) ו הצפויה הודות לכניסת הסידן המוגברתמעבר לז(להצטברות מוגברת זו
אין עדויות המצביעות על הבדלים בסוג . ותורמים לה בין השאר ריכוז גבוה יותר של תעלות סידן
.או ברגישותן למתח, )P/Qתעלות מסוג (תעלות הסידן בין שתי אוכלוסיות הטרמינלים
ת להסביר את השחרור הסינפטי המוגבר בטרמינלים לא ברור אם די בכניסת סידן מוגברת על מנ
פער השחרור הסינפטי בתגובה לגירוי בתדרים , בגירוי ישיר. נמוכים גירויהפרוקסימליים בתדרי
. נמוכים לא הצטמצם מהותית כפי שהצטמצם פער השחרור בין הטרמינלים בתדרי גירוי הגבוהים
ספקת הסבר מלא להבדל בשחרור הסינפטי בין אי לכך יש מקום לטעון שכניסת סידן מוגברת לא מ
מכוונים ההבדלים המורפולוגיים בין הטרמינלים . ביניהםמבדילוקיים גורם נוסף ה, הטרמינלים

57 השפעות שינויים בכניסת הסידן לטרמינל על התגובה לגירוי בתדר_________________________________________________________________________________
הבדלים אפשריים אלה . בפיזור המרחקים בין תעלות הסידן לאתרי השחרור השוניםלהבדל אפשרי
ועשויים לשנות , סידן שונים בעקבות גירויריכוזי טווח רחב של חושפים את אתרי השחרור השונים ל
אפשרי . (Meinrenken et al., 2002)את ההסתברות לשחרור מהם בטווח של שני סדרי גודל
אתרי השחרור נחשפים חלק ניכר מוהסידור המורפולוגי של הטרמינלים הפרוקסימליים בנוי כך ש
זו עשויה להסביר גם את תופעת הרוויה סברה . כבר בתדרי גירוי נמוכיםלריכוזי סידן גבוהים יחסית
. 50Hz -לבתדרי גירוי של מעל בטרמינלים אלההמתפתחת
פינוי מנגנוני כאן המקום לציין את יעילות :הטרמינלהבדלים ביעילות מנגנוני פינוי הסידן מתוך
הסידן מציטופלסמת הטרמינל כגורם נוסף העשוי להסביר חלק מהבדלי התגובה לגירוי בתדר בין
שמקורם (הבדלים אלה תוארו בין טרמינלים פאזיים לטוניים . הטרמינלים הפרוקסימליים למרכזיים
יש לסייג ). 1.2.4ראה מבוא (ר את הצבת הגדולה בשריר הסוג) בשני מוטורנוירונים נפרדים
ראשית אין עדויות ברורות להבדלים ביעילות מנגנוני פינוי הסידן . אפשרות זו משתי סיבות עיקריות
אולם לאור ההבדלים בשחרור הסינפטי ביניהם לא ניתן לשלול , בין טרמינלים שמקורם באקסון יחיד
יה היא ההבדלים הניכרים בשחרור יהסיבה השנ. אלהקיום של הבדלים גם ביעילות מנגנונים
כיוון שאלה באים לידי , שאינם מוסברים על ידי הבדל אפשרי במנגנוני הפינוי, הבודד בין הטרמינלים
. ביטוי בעיקר במהלך גירוי מתמשך
, מלבד כניסת סידן מוגברת, תוצאות העבודה מצביעות על כך שאכן קיים גורם נוסף, לסיכום
על סמך . לשחרור הסינפטי המוגבר מהטרמינלים הפרוקסימליים בתדרי הגירוי הנמוכיםהאחראי
שוניל או , ים שונםיי מורפולוגים לפיזיולוגיה שונה הנגזרת ממבנ זהשייך הבדללהתוצאות לא ניתן
יעילות מנגנוני פינוי הסידן, רגישות מנגנון השחרור לסידן, ריכוז סידן במנוחהכמובגורם אחר
.וכדומה
לטרמינל שלפוטנציאציה ליכולת הכניסת סידן בעקבות גירוי יחס הפוך בין .4.5.5
י בתכולה הקואנטלית של התגובה כל מניפולציה שהביאה לשינו, בניסויים המתוארים בפרק זה
הביאה לשינוי ביחס הפוך בשיפוע העקום , כפי שזו נמדדה בתדרים הגבוהים, שעברה פוטנציאציה
לכניסת הסידן אליו יכולת הפוטנציאציה של הטרמינל בין , יחס הפוך זה. (fig.11A) שחרור-תדר
י "וכמו כן ע, בקיבוע מתח של הטרמינל)Wright et al. )Wright et al., 1996aי "הודגם גם ע
Stanley )Stanley, 1986; Liu & Stanley, 1995( ,בריכוזי תוך גירוי בתדר של סינפסת דיונון

58 השפעות שינויים בכניסת הסידן לטרמינל על התגובה לגירוי בתדר_________________________________________________________________________________
ומאששות את , ל" תוצאות העבודה הנוכחית תואמות היטב את התצפיות הנ.סידן חוץ תאי שונים
.מידת כניסת הסידן אליוביטוי ללמעשה היא של הטרמינל פוטנציאציה /הפסיליטציהיכולת ההנחה ש
הפוך בין מידת הגברת אקספוננציאלי התקבל יחס בתנאי כניסת סידן שונים בעבודה הנוכחית
ההפוך המשמעות הכמותית של היחס . (fig.11A) שחרור- לשיפוע עקום תדר המקסימליהשחרור
מראה שעליה Xל על גבי ציר "אקסטרפולציה של היחס הנ. ל טרם ברורה"בין שני המאפיינים הנ
ים בטרמינל) עקב גירוי ישיר חזק או עליה משמעותית בריכוז סידן חוץ תאי(ניכרת בשחרור הסינפטי
מעניין לציין שערך זה נמדד . 1 -המרכזיים תוריד את שיפוע העקום לערך אסימפטוטי המתקרב ל
בדומה לכך ירידה ניכרת בשחרור הסינפטי . (fig.7A)בתנאים פיזיולוגיים בטרמינלים הפרוקסימליים
השיפוע ערך הצפוי לציין את , 2.4 -צפויה להעלות את שיפוע העקומה לערך מקסימלי השווה ל
. )Ravin et al., 1999: ראהapparent Hill Coefficient (המקסימלי של העקום בתנאי ניסוי אלה
. של הטרמינל נגזרת ממידת כניסת הסידן אליוהפוטנציאציה יכולת ל תומכים בכך ש"כל הנ
בעקום התגובה לגירוי בתדר" מדרגות"נוכחות .4.5.6
האופן בו הוא , אחת התופעות המעניינות בתגובה לתדר של הטרמינלים המרכזיים בעיקר
Stanleyי "תופעה זהה תוארה ע. ולא באופן הדרגתי רציף, "מדרגות"התגובה מתפתחת בשלוש
)Stanley, 1986; Liu & Stanley, 1995(עבודות אלה הניחו בסיס להנחה . סינפסת דיונון ב
. לעליית ריכוז הסידן הממוצע בטרמינלבדבר התפתחות הפוטנציאציה כביטוי , שהוזכרה לעיל
המתרחשת באופן , שלושת המדרגות עשויות לייצג קשירה של שלושה יוני סידן למנגנון השחרור
קשירת יון הסידן הרביעי . ועליית ריכוז הסידן בטרמינל בהתאם, הדרגתי עם הגברת תדר הגירוי
Bertram et al. )Bertram etי "ל ע"ות התופעה הנהמודל שפותח בעקב. מביאה לשחרור עצמו
al., 1996(עם קבועי קישור , מניח זיקה שונה לכל אחד מאתרי קשירת הסידן באתר השחרור
באופן זה הקשירה מתחילה מהאתר בעל הזיקה הגבוהה ביותר ומסתיימת באתר . והתנתקות שונים
כל קשירה כזו מלווה בעליית מדרגה בשחרור . שחרור הוזיקולההמביא ל, בעל הזיקה הנמוכה ביותר
. הסינפטי
והיה שכיח יותר משמעותית , מהטרמינלים50% -בעבודה זו השחרור הסינפטי במדרגות נצפה בכ
בקירוב 4,10,30Hzהמדרגות הופיעו בתדרים . בטרמינלים המרכזיים בהשוואה לפרוקסימליים
בהנחה שהתופעה אכן . בקירוב במרכזיים10,30,60Hzים ובתדר, בטרמינלים הפרוקסימליים

59 השפעות שינויים בכניסת הסידן לטרמינל על התגובה לגירוי בתדר_________________________________________________________________________________
של ריכוז הסידן , על ציר תדר הגירוי, היא תומכת בממצאים בדבר התפתחות מאוחרת יותר, עקבית
. )Cooper et al., 1995a; Msghina et al., 1999(בטרמינלים המרכזיים בהשוואה לפרוקסימליים
ניתן . ל בטרמינלים הפרוקסימליים"טשטוש היחסי של הופעת המדרגות הניתכנו מספר סיבות ל
לחלק גדול יכותסמהממוצע בריכוז הסידן , להניח שעל מנת להבחין במדרגות בשחרור הסינפטי
ריכוז הסידן כאשר רוני סינכשחרוראפשר יתשכך , מאתרי השחרור הפעילים צריך להיות זהה
, הגדולים יותר בטרמינלים הפרוקסימליים. ביחס לקשירתו לאתר השחרורעובר סף מסויםהממוצע
מדויק פוחתת ולכן ההסתברות לסנכרון , מספר הסינפסות ואתרי השחרור רב יותר מאשר במרכזיים
בנוסף לכך השחרור מהטרמינלים הפרוקסימליים נשען . רי השחרורעם עליית מספר הסינפסות ואת
מצב . המתפתח בזמן הגירוי, בוופחות על ריכוז הסידן הממוצע , יותר על סידן הנכנס בעקבות הגירוי
כיוון שכניסת , שחרור ביחס לריכוז הסידן הממוצע המתפתח בטרמינלזה גורם לחוסר אחידות ב
מאידך בטרמינלים . לשחרורתורמת יותר והיא זו ה, אתר לאתרשונה בין עשויה להיות הסידן
ריכוז הסידן הממוצע אילו ו, לצורך השחרור ופחות דומיננטיתהמרכזיים כניסת הסידן נמוכה יותר
באופן . לשחרוריחסבבנקודה דומה כך שרבים מהם נמצאים , הוא זה הדוחף את האתרים לשחרור
.זה עשויות להתקבל מדרגות בשחרור
יחסית גבוהיבה אפשרית נוספת קשורה בעובדה שהשחרור הסינפטי מהטרמינלים הפרוקסימליים ס
שווה 1Hz -לשם השוואה השחרור מטרמינלים אלה בתגובה לגירוי ב. כבר בתדרי גירוי נמוכים
לושת על מנת להבחין בש. 20Hzבערכו לשחרור הסינפטי מהטרמינלים המרכזיים בתדר גירוי של
נמוכה מאודיש להניח שהשחרור הסינפטי מהטרמינל צריך להתחיל מרמה , ל"הנ" המדרגות"
השחרור , לפיכך יתכן שבתדרים נמוכים. כפי שקורה בטרמינלים המרכזיים, ולעלות בהדרגה
ולכן לא , או שתייםהסינפטי בחלק גדול מהטרמינלים הפרוקסימליים כבר עבר מדרגת שחרור אחת
הנחה זו עולה בקנה אחד עם העלייה המוקדמת של השחרור . ניתן להבחין בהם בספקטרום הנבדק
. והופעה מוקדמת של רוויה, שיפועבהירידה , הסינפטי כבר בתדרים נמוכים בטרמינלים אלה
אין , מינלים שנבדקומכיוון שתופעת השחרור הסינפטי במדרגות לא התרחשה באופן עקבי בטר
ולהסיק ממנה מסקנות מרחיקות לכת בנוגע לתהליך , כרגע מקום להתייחס אליה כאל תופעה כללית
כמו כן קיימות עדויות רבות לכך שעם עליית תדר הגירוי מגויסים אתרי שחרור . השחרור עצמו
הופעה של שלוש לא ברור האם גיוס זה עולה בקנה אחד עם ו, נוספים שלא היו פעילים קודם לכן
על מנת להבין את התופעה וההשלכות שלה יש צורך בתיאור . ברורות בשחרור הסינפטי" מדרגות"
והמבדיל אותם מהטרמינלים בהם אין מדרגות ברורות , מפורט יותר של הטרמינלים בהם מתרחשת

60 השפעות שינויים בכניסת הסידן לטרמינל על התגובה לגירוי בתדר_________________________________________________________________________________
טה בדיקה אפקטיבית של נוכחות מדרגות אלה צריכה להתבצע בתנאים בהם מושגת שלי. בשחרור
.voltage clampלמשל בתנאי , אופטימלית והדרגתית בכניסת הסידן לטרמינל
אספקטים פיזיולוגיים של עקום התגובה לגירוי בתדר .4.5.7
ההבדלים שנמצאו בין הטרמינלים השונים מעידים על יכולת שחרור סינפטי גבוהה של
מיוחד בשני הקצוות יתרון זה משמעותי ב. הטרמינלים הפרוקסימליים בכל טווח התדרים הנבדק
במקרה האחרון . ובסדרות גירויים קצרות בתדרים גבוהים, בתדרים נמוכים: הפיזיולוגיים של הטווח
, ערך הרוויה הגבוה אליו מטפס השחרור: השחרור הסינפטי משמעותי הודות לשני מאפיינים
רמינלים בסדרת גירויים כזו הט. ביחס לטרמינלים המרכזיים, והמהירות הכפולה בה מתפתח
הטרמינלים המרכזיים תורמים לשחרור . לפתח שחרור סינפטי משמעותי" מספיקים"המרכזיים לא
וסדרת הגירויים , 10Hzהנע בממוצע סביב , "סף לשחרור"הסינפטי במידה ותדר הגירוי גבוה מעל ה
נטיות ניתן להבחין במספר תופעות רלווEMGברישומי . גירויים10 -כלומר מעל ל, ארוכה מספיק
שנית סדרות הירי . 10Hzראשית תדר הירי בזמן שהשריר פעיל לא יורד מתחת ל . בהקשר זה
מנתונים אלה ניתן להסיק . הקצרות בתדר גבוה נשענות על ירי ממושך יחסית בטווח תדרי הביניים
כך , שיתכן ותפקידם של הטרמינלים המרכזיים לקרב את השריר אל סף ההתכווצות המכאני שלו
אישוש ההשערה . מינימלי של ירי בתדר גבוה יביא לתגובה אופטימלית ומהירה של השרירשפרץ
מלווה ברישום תנועות , מדויק ומקיף יותרEMGובין השאר רישום , ל מחייב ניסויים נוספים"הנ
. הצבת

61
השפעת אינהיביציה פרסינפטית על התגובה לגירוי .5 בתדר
___________________________________________________________________
Crayfish -אינהיביציה פרסינפטית ב: מבוא .5.1
פיזיולוגיה ומורפולוגיה .5.1.1
אינהיביציה פרסינפטית פועלת ישירות על טרמינל כך ששחרור הנוירוטרנסמיטור ממנו בעקבות
& Dudelי "ורי זה עבסרטן תואר לראשונה מנגנון אינהיביט. פלישת פוטנציאל הפעולה אליו פוחת
Kuffler 1961 בשנת )Dudel & Kuffler, 1961( . 2גירוי האקסון האינהיביטוריms לפני הגירוי
צירוף קלאסי המצביע , האקסיטטורי הביא לירידה בתכולה הקואנטלית ללא שינוי בגודל הקואנטום
הגדלת מוליכות ממברנת האקסון או הטרמינל המנגנון שהוצע הוא. על מנגנון אינהיביטורי פרסינפטי
עקב כך קטנה הדפולריזציה . ובהתאם ירידה בהתנגדות הכניסה של הטרמינל, האקסיטטורי
וכניסת הסידן תלויית המתח מופחתת בהתאם , בטרמינל בעקבות פלישת פוטנציאל פעולה
(shunt) .י "בעבודות עוקבות נחסמה האינהיביציה עPicrotoxin , שאיששה את ההשערה תוצאה
& GABAA )Takeuchiוהצביעה על מעורבות מוליכות כלור המתווכת דרך רצפטורי , ל"הנ
Takeuchi, 1969; Blundon & Bittner, 1992; Parnas et al., 2000b( . המנגנון הפרסינפטי
ריים ולכן דורש תשתית ל מחייב הימצאות סינפסות אינהיביטוריות על גבי טרמינלים אקסיטטו"הנ
שהמפורט בהם , פורסמו מספר מחקרים מורפולוגיים התומכים בקיום תשתית זו. מבנית מתאימה
& Govind et al. )Atwood & Morin, 1970; Govind et al., 1995; Jahromiי "נערך ע
Atwood, 1974; Lang et al., 1972( . מכל הסינפסות 22%ממחקר זה עולה שבממוצע
באזור . והן אלה המשרות את האינהיביציה הפרסינפטית, Axoaxonalהאינהיביטוריות הן מסוג
על חלק . הדל יותר בסינפסות אלה, באזור המרכזי15% מול 30%: הפרוקסימלי מספרן היחסי כפול
י אזורי בשנ. מהטרמינלים האקסיטטוריים באזור המרכזי לא נמצאו כלל סינפסות אינהיביטוריות
ומכילות לרוב אתר שחרור אחד או , השריר הסינפסות האינהיביטוריות הללו דומות מורפולוגית
ממוקמות על האקסון ) 8%(מיעוטן : מיקומן ביחס לטרמינלים האקסיטטוריים משתנה. שניים
ועל , בקטע המחבר בין טרמינלים סמוכים, בכניסה: ורובן נמצאות בסמיכות לטרמינל, הפרטרמינלי
. )Govind et al., 1995(גבי הטרמינל עצמו

62 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
מודלים חשמליים .5.1.2
והעלו שאלות נוספות , ל"עדויות מורפולוגיות אלה סיפקו תשתית מבנית לנתונים הפיזיולוגיים הנ
רישום תוך . בנוגע למשמעות המיקום של הסינפסה האינהיביטורית על גבי הטרמינל האקסיטטורי
,Atwood, Segev )Atwood et al., 1984; Segevולכן, הוא מסובך טכניתתאי מהטרמינל
במודל זה שולבה מוליכות . ניסו לענות על שאלה זו באמצעות בניית מודל חשמלי)1990
ונמדדה השפעתה על , אינהיביטורית במקומות שונים על גבי האקסון או הטרמינל האקסיטטורי
מודלים אלה מנבאים . ממברנת הטרמינל עם פלישת פוטנציאל הפעולה אליוהמתח המתפתח על
כאשר הסינפסה האינהיביטורית ממוקמת על גבי ביחס לשחרור יעילות מקסימלית של האינהיביציה
מיקום זה נמצא אפקטיבי . בסמיכות מקסימלית לאתר השחרור עצמו, הטרמינל האקסיטטורי עצמו
יש לציין שבמצב זה לא .ארוך ופסיבי חשמלית" צוואר"י "מהאקסון עבמיוחד בטרמינלים המופרדים
.תתרחש ירידה במשרעת פוטנציאל הפעולה הנרשם בסמוך לטרמינל
אנליזה קואנטלית .5.1.3
במטרה להבין האם האינהיביציה משרה ירידה בהסתברות לשחרור או ירידה במספר אתרי
ותוך גירוי של האקסון , רי בביקורתהשחרור נעשתה אנליזה קואנטלית של השחרור האקסיטטו
התאמת התפלגות השחרור . )Atwood & Tse, 1988; Golan et al., 1994(האינהיביטורי
כפי הנראה עקב הוספה של מרכיב , הסינפטי תחת אינהיביציה למודל בינומי הייתה חלקית בלבד
נמצאה , הושגה התאמה סטטיסטית טובהבטרמינלים בהם. י האינהיביציה"שונות לא הומוגני ע
. תחת אינהיביציהn, pירידה בערכי שני הפרמטרים
עדות למנגנונים אינהיביטוריים נוספים .5.1.4
קיימים בסרטנים מנגנונים , המתווכת דרך מוליכות כלור שתוארה לעילshunt inhibitionמלבד
הודגמו מוליכויות אשלגן וכלור תלויות בסרטן. י רצפטורים שונים"אינהיביטוריים אחרים המתווכים ע
GABA ,י ריכוזים גבוהים של "שאינן נחסמות ביעילות עpicrotoxin )Marder & Paupardin-
Tritsch, 1978( .Miwa et al. )Miwa et al., 1990(תארו מוליכות אשלגן פרסינפטית בלובסטר ,
תפקידה . GTPומצומדת לחלבון קושר , )Baclofen -רגיש ל( GABAB י רצפטור"המתווכת ע
לא השפיעה כמעט Pertussis toxinי "שכן חסימתה ע, הפיזיולוגי של מוליכות זו לא לגמרי ברור
מנגנון נוסף של אינהיביציה . GABA -כלל על האינהיביציה המעוררת באמצעות חשיפה ל

63 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
שרשמה )Barry )Barry, 1984י " הודגם לראשונה עCrayfish -פרסינפטית בטרמינלים של ה
אגוניסט ( Baclofenירידה בתכולה קואנטלית ללא שינויי מוליכות פוסטסינפטית בתגובה למתן
- רשמו בתגובה ל)Blundon & Bittner, 1992( Blundon & Bittnerבהמשך . GABAB)לרצפטור
Baclofenבקרבת , ה קלה אך מובהקת במשרעת פוטנציאל הפעולה באקסון האקסיטטורי יריד
. הטרמינל
Fischer & Parnasי " ניתנה עGABABעדות ברורה יותר למעורבות אינהיביציה מתווכת
)Fischer & Parnas, 1996a(חלקית , שגירו ישירות ובפולס זרם קבוע את הטרמינל ובכך נטרלו
עם חשיפת הטרמינל לבקלופן נרשמה ירידה משמעותית . shunt inhibitionאפקטים של , תלפחו
GABABי אנטגוניסט ספציפי לרצפטור "ירידה זו נחסמה ע. בתגובות האקסיטטוריות לגירוי ישיר
(2OH-Saclofen) .תוצאות דומות התקבלו גם ע" Parnas et al. )Parnas et al., 1999( . כמו
הראו שגירוי האקסון האינהיביטורי בתדרים )Fischer & Parnas, 1996b(בעבודה נוספת , כן
י "המתווכת ע, שאיננה נוכחת בתדרי גירוי נמוכים, מפעיל אינהיביציה נוספת100Hz)(גבוהים
. ובעלת זמן מחצית ארוך יחסיתGABABרצפטור
שהראו אינהיביציה )Parnas et al. )Parnas et al., 2000bי "עמנגנון אפשרי נוסף הודגם
שאינה פועלת על מנגנון השחרור דרך הפחתת כניסת הסידן , פרסינפטית פרדוכסלית לכאורה
אינהיביציה זו הייתה אפקטיבית גם כאשר גירו את האקסון האינהיביטורי בו זמנית לגירוי . לטרמינל
וכאמור לא , picrotoxinי "האינהיביציה נחסמת לחלוטין ע. (2ms)ואף אחריו , טטוריהאקסון האקסי
.כפי שהשתקף ממדידות המבוססות על הדמיית סידן, הפחיתה את כניסת הסידן לטרמינל
אינהיביציה טונית .5.1.5
Parnas et al. )Parnas. בנוסף לזו המעוררת, קיימות מספר עדויות לקיום אינהיביציה טונית
et al., 1982b(לאחר ניוון של , הראו האטה בקבוע דעיכה של תגובה פוסטסינפטית אקסיטטורית
.(common inhibitor)האקסון האינהיביטורי בלובסטר
& Golan & Grossman )Golanי " ניתנה עCrayfish -עדות לאינהיביציה פרסינפטית טונית ב
Grossman, 1996( , שתיארו הגברה בשחרור האקסיטטורי לאחר טיפול ממושך במעכבי האנזים
י "הגברה זו נובעת מירידה באינהיביציה המתווכת ע. Picrotoxin -וב, GABA (GAD)המסנטז
.כשחלקו היחסי של כל אחד מהם טרם ברור, GABAA ,GABABהרצפטורים

64 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
Parnas et al. )Parnas et al., 1999( בשחרור האקסיטטורי 60% -מתארים הגברה של עד כ
המעידה על נוכחות אינהיביציה פרסינפטית טונית , GABABלאחר חסימה טונית של רצפטור
. י רצפטור זה"המתווכת ע
___________________________
סיכום .5.1.6
והאקסונים רשת של טרמינלים אינהיביטוריים הממוקמים על גבי הטרמינלים האקסיטטוריים
אחראית להפעלת האינהיביציה הפרסינפטית , בעיקר באזור הפרוקסימלי, המובילים אליהם
של shunt -וגורמת ל, Picrotoxinי "בזמן אינהיביציה נפתחת מוליכות כלור הנחסמת ע. במערכת זו
קיימות עדויות על מנגנונים נוספים התורמים לאפקט . פוטנציאל הפעולה הפולש לטרמינל
הבאה לביטוי עם גירוי האקסון האינהיביטורי GABABינהיביטורי כמו מוליכות מתווכת רצפטור הא
המדכאת את השחרור האקסיטטורי מבלי להפחית GABAAואף מוליכות מתווכת , בתדרים גבוהים
.את כניסת הסידן לטרמינל
וקינטיקת ההשפעהGABA -רגישות הטרמינלים ל .5.2ן השפעת אינהיביציה פרסינפטית על התגובה לתדר של טרם אפיו: GABA -רגישות ל
בדיקה זו . וקבועי הזמן המאפיינים אותה, GABAנבדקה רגישותם לריכוזים עולים של , הטרמינלים
, כך שהאפקט על השחרור הסינפטי יהיה ברור מחדGABAאפשרה מאוחר יותר בחירת ריכוזי
.כל הניסויים בפרק זה נערכו בגירוי האקסון. אולם מבלי שהתגובה תתקרב לערכים אפסיים מאידך
. GABA -אחד הממצאים הבולטים הראשונים הוא ההבדל ברגישות שתי אוכלוסיות הטרמינלים ל
השחרור 40µM -ובריכוזים הגבוהים מ, בכל המקרים הטרמינלים המרכזיים היו רגישים יותר
בות מטרמינל מרכזי אופייני דוגמא לרישום תגו. הסינפטי ירד ברוב הטרמינלים לרמה אפסית
40µM של GABAתחת ריכוז , (50Hz)למרות תדר הגירוי הגבוה באופן יחסי . fig.12A -מופיעים ב
המציינים תכולה Y - כל צירי הfig.12B,C -יש לשים לב שב. ( אינן כשלון12 תגובות מתוך 3רק
fig.12D -ב). וין גם באחוזים מהביקורת מצGABAולכן השחרור תחת , קואנטלית הם לוגריתמיים
כתלות , טרמינלים מרכזיים6 -מופיע עקום המתאר את הירידה הממוצעת בשחרור הסינפטי ב
. GABAבריכוז
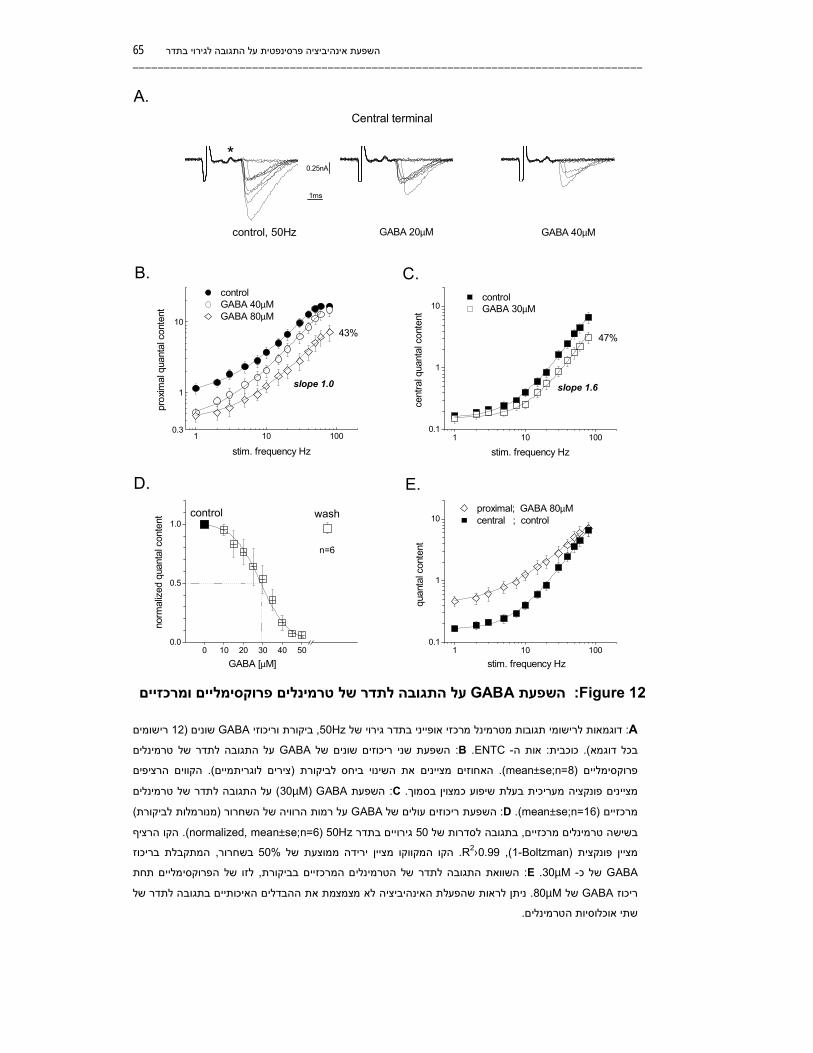
65 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
Figure 12: השפעת GABAעל התגובה לתדר של טרמינלים פרוקסימליים ומרכזיים
A : 50דוגמאות לרישומי תגובות מטרמינל מרכזי אופייני בתדר גירוי שלHz , ביקורת וריכוזיGABA רישומים 12( שונים
ובה לתדר של טרמינלים על התגGABAהשפעת שני ריכוזים שונים של : ENTC .B -אות ה: כוכבית). בכל דוגמא
הקווים הרציפים ). צירים לוגריתמיים(האחוזים מציינים את השינוי ביחס לביקורת . (mean±se;n=8)פרוקסימליים
על התגובה לתדר של טרמינלים GABA (30µM)השפעת : C. מציינים פונקציה מעריכית בעלת שיפוע כמצוין בסמוך
) מנורמלות לביקורת( על רמות הרוויה של השחרור GABAזים עולים של השפעת ריכו: D. (mean±se;n=16)מרכזיים
הקו הרציף . 50Hz (normalized, mean±se;n=6) גירויים בתדר 50בתגובה לסדרות של , בשישה טרמינלים מרכזיים
וז המתקבלת בריכ, בשחרור50%הקו המקווקו מציין ירידה ממוצעת של . R20.99, (Boltzman-1)מציין פונקצית
GABA30 - של כµM .E :לזו של הפרוקסימליים תחת , השוואת התגובה לתדר של הטרמינלים המרכזיים בביקורת
ניתן לראות שהפעלת האינהיביציה לא מצמצמת את ההבדלים האיכותיים בתגובה לתדר של . 80µM של GABAריכוז
.שתי אוכלוסיות הטרמינלים
1 10 1000.3
1
10
GABA 40µMGABA 20µMcontrol, 50Hz
E.
43%
D.
C.
slope 1.6slope 1.0
B.
A.
controlGABA 40µMGABA 80µM
prox
imal
qua
ntal
con
tent
stim. frequency Hz
0 10 20 30 40 500.0
0.5
1.0
n=6
control wash
norm
aliz
ed q
uant
al c
onte
nt
GABA [µM]
1 10 1000.1
1
10
47%
cent
ral q
uant
al c
onte
nt
controlGABA 30µM
stim. frequency Hz
1 10 1000.1
1
10proximal; GABA 80µMcentral ; control
quan
tal c
onte
nt
stim. frequency Hz
Central terminal
*
1ms
0.25nA

66 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
והאפקט נבדק , 50Hz גירויים בתדר של 50דרות של גורו במספר ס(n=6) הטרמינלים בכל ריכוז
העקום מראה .Boltzmannלעקום הותאמה פונקצית . הסינפטיעל רמת הרוויה של השחרור
. השחרור הסינפטי יורד למחצית מערך הביקורת30µM - של כGABAבריכוז ש
וב נדרשו ריכוזים כפולים ולכן לר, ריכוזים דומים בטרמינלים הפרוקסימליים גרמו לאפקט קל בלבד
מסיבה זו הניסויים בטרמינלים הפרוקסימליים נערכו בשני ריכוזי . על מנת להגיע לאפקט משמעותי
GABA שונים (80 ,40µM) , 30דומה יותר לזה של נמצא כשהאפקט של הריכוז הגבוה יותרµM
. טרמינלים המרכזייםב
, טרמינלים מרכזיים9 - נבדק בGABAקבוע הזמן של השפעת : GABAקינטיקת השפעת
בניסויים אלה גורו הטרמינלים בסדרה קצרה של חמישה גירויים בתדר של . הסמוכים לאקסון
100Hzנוהל גירוי זה הוכיח את עצמו כאחד הרגישים לצורך זיהוי השפעה של . אחת לשתי שניות
ת ההזרמה של תמיסה בכל הניסויים תוך מספר שניות לאחר התחל. אינהיביציה פרסינפטית
GABAבניסויים אלה ריכוזי . ניתן היה לזהות בעין ירידה ברורה בשחרור הסינפטיGABAהכוללת
מדידת הזמן בניסויים אלה החלה מרגע הזלפת . על מנת להבטיח אפקט ברור50µMנעו סביב
סויים בני. שנעשתה בשניות הראשונות באופן מואץ יותר, לתוך האמבטGABAהתמיסה הכוללת
GABAזמן מחצית האפקט הנמדד של . שניות10 -אלה זמן החלפת התמיסה באמבט לקח פחות מ
, כמעט בכל המקרים הייתה מלווה באחלמה מלאהGABAהשטיפה של . שניות בממוצע40היה
בכל המקרים ניתן היה לזהות בעין התחלת אחלמה של . והתרחשה אף היא במהירות דומה
ניסויים בהם . שניות בממוצע33עם זמן מחצית של , פורות מתחילת השטיפההתגובות תוך שניות ס
.השטיפה לא הייתה מלאה ומהירה לא נכללו בתוצאות
ובמקרים חריגים זמן מחצית , על הטרמינלים הפרוקסימליים הייתה איטית יותרGABAהשפעת
. המרכזייםבדומה לזו של הטרמינלים , השטיפה לרוב הייתה מהירה. האפקט ארך דקות
על מופע הרוויה של התגובה לתדרGABAהשפעת .5.3
- על עקומי התגובה לתדר של טרמינלים פרוקסימליים ומרכזיים מתוארת בGABAהשפעת
fig.12B,C .והרגישות הנמוכה יחסית של הטרמינלים , הודות לתכולה הקואנטלית הגבוהה
שונים GABAויים בטרמינלים אלה בשני ריכוזי נערכו הניס, כאמור לעיל, GABA -הפרוקסימליים ל
(40, 80µM) .30 -בטרמינלים המרכזיים עלייה בריכוז מעבר לµM הביאה במקרים רבים לירידה

67 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
מסיבה זו רק בחלק מטרמינלים אלה נעשו ניסויים בריכוז נוסף של . דרסטית בשחרור הסינפטי
GABA .
גובה לגירוי בתדרהפעלת אינהיביציה לא משפיעה על שיפוע עקום הת .5.3.1
הממצא הבולט בניסויים אלה הוא היעדר שינוי משמעותי בשיפוע עקומי התגובה לתדר של כלל
-שיפועי העקומים נותרים זהים למרות ירידת השחרור הסינפטי ל. הטרמינלים בזמן אינהיביציה
של ממצא זה לא תואם את זה . בטרמינלים הפרוקסימליים והמרכזיים בהתאמה47% - ו43%
הגירוי הישיר הביא לעליה בהם , ובטרמינלים המרכזיים בפרט, הניסויים שתוארו בפרק הקודם
. וההפחתה בריכוז הסידן החוץ תאי לאפקט הפוך, בשחרור הסינפטי המלווה בירידת שיפוע העקום
מכיל מרכיב נוסף מלבד עומד מנגנון הבבסיס האינהיביציה שעשוי להצביע על כך חוסר התאמה זה
ההשפעה . מופיע בדיון בסוף הפרקעל כך נוסף פירוט. (shunt)פחתה בכניסת הסידן לטרמינל הה
על השחרור הסינפטי ) בריכוז סידןגירוי ישיר והפחתה , GABA(היחסית של שלושת הטיפולים
. בסוף העבודה1-ובאופן איכותי בטבלא, fig.13D -המקסימלי ושיפוע העקום מוצגים באופן גרפי ב
מעניין לציין שהרוויה האופיינית , למרות היעדר השפעת האינהיביציה על שיפוע עקום התגובה לתדר
ימינה על ציר התדר בעקבות האינהיביציה לטרמינלים הפרוקסימליים בתדרים הגבוהים מוסטת
הסטה זו . בדומה יותר לשחרור הסינפטי האופייני לטרמינלים המרכזיים, ) במסגרתfig.13Aראה (
כמעט את זה של " להדביק "40µM של GABAשל הרוויה מאפשרת לשחרור הסינפטי תחת ריכוז
ולכן לפתח למעשה , )80Hz בתדר 90% לעומת 1Hz מהביקורת בתדר 45%(הביקורת
.פירוט תופעה זו מופיע בהמשך. פוטנציאציה מקסימלית גבוהה יותר באופן משמעותי
הפעלת אינהיביציה לא מקטינה את ההבדלים האיכותיים בין אוכלוסיות הטרמינלים .5.3.2
, למרות ההסטה ימינה של רווית התגובה לתדר של הטרמינלים הפרוקסימליים תחת אינהיביציה
לצורך . גיש שהשחרור הסינפטי מטרמינלים אלה עדיין שונה מהותית מזה של המרכזייםחשוב להד
לזה של , (80µM)העניין ניתן להשוות את השחרור הסינפטי הפרוקסימלי תחת אינהיביציה
שני עקומי . מאחר ושניהם מגיעים לערכי שחרור מקסימליים דומים, הטרמינלים המרכזיים בביקורת
ניתן לראות שהאינהיביציה לא השפיעה באופן מהותי על . fig.12E -מופיעים בל "התגובה לתדר הנ
ובפרט ההבדלים ברמות השחרור , אותם הבדלים איכותיים ברורים הקיימים בין העקומים בביקורת
. ובשיפועי העקומים, הסינפטי בתדרים הנמוכים

68 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
Figure 13 : השפעתGABAהטרמינלים על הפוטנציאציה המקסימלית של
A : השפעת שני הריכוזים השונים שלGABAהריכוז . על עקום הפוטנציאציה המקסימלית בטרמינלים פרוקסימליים
ואילו הגבוה יותר מסיט את , וממתן את הופעת הרוויה) שמאלה(הנמוך יותר מסיט את העקום לתדרים נמוכים יותר
. ם אין השפעה מהותית על קינטיקת התפתחות הפוטנציאציהבשני המקרי). ימינה(התפתחותה לתדרים גבוהים יותר
. עם סקלה ליניארית של ציר הפוטנציאציה) בלבדGABA 40µM -ביקורת ו(במסגרת הקטנה ניתן לראות גרף זהה
נמוך בתדרי הגירוי GABAגרף זה מדגיש את היעלמות הרוויה והתגברות הפוטנציאציה המתרחשים תחת ריכוז
75% - מנמיכה את הפוטנציאציה המקסימלית בטרמינלים מרכזיים ל30µM בריכוז GABA -ול בטיפ: B. הגבוהים
השוואת הפוטנציאציה המקסימלית הגבוהה : C. ללא השפעה מהותית על קינטיקת ההתפתחות, מערך הביקורת
. (30µM GABA)לנמוכה ביותר בטרמינלים המרכזיים , (40µM GABA)ביותר בטרמינלים הפרוקסימליים
השפעת : D. ואילך20Hzהפוטנציאציה בטרמינלים המרכזיים מתפתחת לערכים גבוהים יותר החל מתדרי גירוי של
+Ca2הפחתת out ,ומתן , גירוי ישיר חזקGABA ציר ( על השינויים היחסיים בשחרור המקסימליX( , ושיפוע עקום
+Ca2ניתן לראות שהשפעת הפחתת . Y(; mean±seציר (התגובה לתדר outהפוכה וסימטרית למתן גירוי ישיר חזק ,
הקו המקווקו מציין . מפחית משמעותית את השחרור מבלי להשפיע כמעט על שיפוע העקוםGABA -אולם הטיפול ב
את היחס האקספוננציאלי המתאר את השינויים היחסיים המתרחשים בשחרור המקסימלי ובשיפוע עקום התגובה
.Fig 9A -כפי שמופיע גם ב, לתדר
1 10 1000.1
1
10
50
1 10 100 0
10
20
30
D.
200%
C.
B.A.controlGABA 40µMGABA 80µM
prox
imal
max
. pot
entia
tion
stim. frequency Hz1 10 100
0.1
1
10
10075%control
GABA 30µM
cent
ral m
ax. p
oten
tiatio
n
stim. frequency Hz
1 10 20 100
0.1
1
6.510
100
max
. pot
entia
tion
proximal; GABA 40µMcentral; GABA 30µM
stim. frequency Hz
0.3 0.5 1 5
0.7
0.8
0.9
1.0
1.1
1.2
1.3
slo
p ra
tio
central terminalsreduced Ca2+
direct stim.GABA 30µM
m ratio

69 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
על ערכי הפוטנציאציה המקסימליתGABAהשפעת .5.3.3
, יש לציין שקשה לקבוע במדויק את הערכים המוחלטים של הפוטנציאציה בטרמינלים המרכזיים
הקושי נובע מהצורך במדידה אמינה של התכולה הקואנטלית בתדרי . תוך שימוש בנוהל גירוי זה
זו נכון יותר להתייחס רק לשינויים היחסיים מסיבה. הנמוכה ממילא בטרמינלים אלה, גירוי נמוכים
.תדר גירוי-ולשינויים יחסיים בשיפוע העקום פוטנציאציה, המתרחשים בפוטנציאציה
, בטרמינלים הפרוקסימליים אפקט האינהיביציה על ערכי הפוטנציאציה המקסימלית איננו מונוטוני
, הפוטנציאציה המקסימלית באופן משמעותי שלגורר הגברה GABA (40µM) מתן .כפי שצוין לעיל
, גורמת לאינהיביציה חזקה יותרGABAהגדלת ריכוז . (fig.13A)ובממוצע כפליים ביחס לביקורת
התנהגות זו מזכירה את עקום הפעמון . המפחיתה את הפוטנציאציה אל מתחת לערכי הביקורת
עם ירידת ריכוז הסידן השחרור הסינפטי יורד : בריכוז סידן חוץ תאיהמתאר את תלות הפסיליטציה
ורק לאחר ירידה נוספת בריכוז הסידן הפסיליטציה , אולם הפסיליטציה בשלב ראשון עולה, מונוטונית
. )Parnas et al., 1982c(יורדת
ואף , תחות האינהיביציהבטרמינלים המרכזיים התקבל רושם שהפוטנציאציה לא עולה עם התפ
. 30µM (fig.13B) של GABA עבור ריכוז 25% -יורדת ב
השוואת עקומות הפוטנציאציה בין שתי אוכלוסיות הטרמינלים מראה שוב שהפעלת אינהיביציה לא
מוצגים שני עקומי הפוטנציאציה fig.13C -ב. מצמצמת את ההבדלים האיכותיים הקיימים ביניהם
40µMכלומר זה של הפרוקסימליים בנוכחות , מלית הקרובים ביותר בשני סוגי הטרמינליםהמקסי
GABA , 30וזה של המרכזיים בנוכחותµM GABA . הפוטנציאציה של הטרמינלים הפרוקסימליים
ומעבר לכך הטרמינלים המרכזיים מפתחים פוטנציאציה חזקה , 20Hzגבוהה יותר עד לתדר של
נטול (20Hz)יש לציין שתדר זה . 10Hzבביקורת מעבר לתדר של גירוי של בדומה למתרחש , יותר
פרט לשינוי יחסי זה הפעלת . משמעות נקודתית ומשמש לצורך השוואה וקנה מידה בלבד
.האינהיביציה לא צמצמה את ההבדלים האיכותיים בין עקומי הפוטנציאציה של שני סוגי הטרמינלים
_______________________
םסיכו .5.3.4
מסיטה כלפי מטה את עקום התגובה לתדר של הטרמינלים GABAי "אינהיביציה המושרית ע
בטרמינלים . לטרמינלשלא בדומה לירידה בכניסת סידן, מבלי לשנות את השיפוע שלו
ולכן , נמוכים יחסית קיים מצב בו הפוטנציאציה המקסימלית מתגברתGABAהפרוקסימליים בריכוזי

70 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
. על רמת הרוויה של השחרור הסינפטי בתדרים גבוהים פוחתת באופן יחסיהשפעת האינהיביציה
הפעלת האינהיביציה לא מקטינה את ההבדלים העקרוניים הקיימים בין עקומי התגובה לתדר
.והפוטנציאציה של קבוצות הטרמינלים השונות
על מופע העלייה של השחרור הסינפטי GABAהשפעת .5.4פתחות השחרור הסינפטי נבחנה תוך שימוש בשני נוהלי גירוי על קינטיקת התGABAהשפעת
והשני סדרות של חמישה , 40Hz-80Hz גירויים בתדרים של 50הראשון כלל סדרות בנות . שונים
בדומה למוזכר בהקשר (וזאת על מנת לאפשר מיצוע מספר רב של תגובות , 100Hzגירויים בתדר
).זה בפרק השלישי
בתגובה לסדרת גירויים ארוכהקצב התפתחות השחרור על GABAהשפעת .5.4.1
ארוך כפליים בטרמינלים המרכזיים בהשוואה קצב התפתחות השחרור כזכור מהפרק הקודם
מאיץ את , כמו כן ניסויי הגירוי הישיר מראים שגירוי חזק יותר של טרמינלים מרכזיים. לפרוקסימליים
על פוטנציאל הפעולה shuntמאחר והאינהיביציה מפעילה . באופן מובהקקצב התפתחות השחרור
מעניין לבדוק גם , )Parnas et al., 2000b(ובכך מפחיתה את כניסת הסידן אליו , הפולש לטרמינל
ניתן לראות בטרמינלים fig.14A,B -ב. ות השחרורקצב התפתח על GABAאת השפעת
על סדרת התגובות לתדר גירוי של GABAאת השפעת ) n=12(ומרכזיים ) =6n(פרוקסימליים
80Hz .ניתן לראות . ערכי הרוויה של סדרות הגירויים בזמן אינהיביציה נורמלו לערך הביקורת
בממוצע את מספר הגירויים הדרושים להגיע שבטרמינלים הפרוקסימליים האינהיביציה מכפילה
ערך האופייני יותר לתגובות המתקבלות , גירויים9.2 - גירויים ל4.3 -מ: למחצית מערך הרוויה
קצב התפתחות ניתן לומר אם כן שבטרמינלים אלה האינהיביציה מאיטה את . מטרמינלים מרכזיים
מעניין לציין בטרמינלים . רבים יותר עד להגעת הפוטנציאציה לרוויהכך שנדרשים גירויים, השחרור
לא גרם להאטה משמעותית נוספת של GABAהריכוז הגבוה יותר של . אלה שני ממצאים נוספים
כמו כן האטה זו התרחשה בעיקר בתדרי גירוי , מעבר לזה שצוין לעיל, קצב התפתחות השחרור
). 30Hzעל מ(גבוהים

71 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
Figure 14 : השפעתGABA והפוטנציאציהשחרורהקצב התפתחות על
A,B : השפעתGABAבתגובה לסדרת גירויי, על משך התפתחות הפוטנציאציה בטרמינלים פרוקסימליים ומרכזיים
שבהשפעת A -ניתן לראות ב. לאחר המיצוע1 -ערכי הרוויה נורמלו ל80Hz (mean±se;n=8;n=12) בתדר אקסון
GABAמשך התפתחות הפוטנציאציה בטרמינלים הפרוקסימליים מוכפל .C,D : סיכום השפעתGABA קצב על
בטרמינלים פרוקסימליים ומרכזיים , 40-80Hz בטווח התדרים אקסון בתגובה לסדרות גירוייהתפתחות השחרור
)St.n½ :ב). ערך הרוויה בתדר גירוי נתוןמספר הגירויים הדרוש לעירור תגובה השווה למחצית- C מופיעה עמודה
תכולה : p<0.001 .E,F,G: כוכביות(mean±se;n=8;n=12)המציינת את השפעת גירוי בו זמני של האינהיביטור
לאותן ) Yציר (GABA תחת טיפול 100Hz)בתדר ( קצרה אקסוןקואנטלית של חמשת התגובות לסדרת גירויי
ומרכזיים , מרכזיים הסמוכים לאקסון, התוצאות מוצגות עבור טרמינלים פרוקסימליים). Xציר (תגובות בביקורת
השיפוע מצוין על גבי (הקו המקווקו מציין רגרסיה ליניארית . (mean±se;n=8;n=18;n=12)המרוחקים מהאקסון
, נותרו קבועיםהסמיכות הרבה של קו הרגרסיה לראשית הצירים מעידה על כך שהיחסים בין התגובות ). הגרף
. בשחרור50% -למרות ירידה של כ, כלומר לא חל שינוי משמעותי בפוטנציאציה של התגובות
0 10 20 30 400.0
0.5
1.0
stimulus No.
prox
imal
nor
mal
ized
rele
ase
controlGABA 40µM
0 10 20 30 400.0
0.5
1.0
cent
ral n
orm
aliz
ed re
leas
e
stimulus No.
controlGABA 30µM
0 2 4 6 8 10 120
2
4
6
proximal
slope 0.53
control quantal content
GAB
A qu
anta
l con
tent
0 2 4 6 8 100
2
4 "close" central
slope 0.42
control quantal content0.0 0.2 0.4 0.6 0.8 1.0 1.2
0.0
0.2
0.4
0.6
"remote" central
slope 0.55
B.A.
*
control quantal content
control GABA 40 GABA 80 inhibitor0
2
4
6
8
10
12
µ µ
proximal
G.
St.n
½
F.E.
D.C.
control GABA 300
2
4
6
8
10
12
µ
central
St.n
½
*

72 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
והתגובות בביקורת ותחת אינהיביציה מתפתחות , בטרמינלים המרכזיים לא נרשמה תופעה דומה
סיכום תוצאות אלה עבור סדרות גירויים בטווח. (fig.14B)בקינטיקה זהה עם מתן הגירויים
.fig.14C,D - מופיע ב40-80Hzהתדרים
בין שתי אוכלוסיות קצב התפתחות השחרור על GABAטרם ברורה משמעות ההבדל בהשפעת
: כמו כן לא ידוע האם שתי התופעות האינהיביטוריות שנצפו בטרמינלים הפרוקסימליים. הטרמינלים
הן ביטוי לפעולתו של מנגנון יחיד, תפתחות השחרורהוההאטה בקצב , הירידה בשחרור הסינפטי
כך שלא ניתן , )בטרמינלים המרכזיים" חזק"הפוך בניסויי הגירוי הישיר ה-סימטריבדומה לאפקט ה(
בהמשך הפרק עדויות לכך . או שמא מקורן בהפעלת מסלולים אינהיביטוריים שונים, להפריד ביניהן
. ל"פריד בין שתי התופעות הנשקיימים מצבים בהם ניתן לה
על התפתחות עצמת הפוטנציאציה בתגובה לסדרת גירויים קצרהGABAהשפעת .5.4.2
בנות חמישה , על התגובה המופקת מסדרות גירויים קצרותGABAבסעיף זה נבדקה השפעת
ניסויים אלה משלימים את הקודמים היות ומאפשרים כימות מדויק של . 100Hzגירויים בתדר של
). 500מעל (הודות למיצוע של מאות רבות של תגובות , לים בהתפתחות עצמת הפוטנציאציההבד
, "חזקים"מרכזיים , פרוקסימליים: השפעת האינהיביציה נבדקה על שלוש קבוצות טרמינלים
ניתן לראות את fig.14E,F,G -ב). בדומה למוזכר בפרק הקודם(ומרכזיים המרוחקים מהאקסון
, )Xציר (כפונקציה של אותן תגובות בביקורת , )Yציר (ת בסדרה תחת אינהיביציה חמשת התגובו
הקו המקווקו מציין התאמה של סדרות התגובות לפונקציה . עבור כל קבוצת טרמינלים בנפרד
או בסמוך מאוד לראשית , ל דרך"מעבר הפונקציות הליניאריות הנ. ליניארית ללא הפעלת אילוצים
אין השפעה משמעותית לא על ערכי הפוטנציאציה ולא על קינטיקת GABA -הצירים מלמדת של
בשחרור 50% - הגורמים לירידה של עד לGABAיש לסייג ממצא זה לריכוזי (ההתפתחות שלה
בטרמינלים ). מאחר ובריכוזים גבוהים יותר כן נצפית ירידה בעצמת הפוטנציאציה, הסינפטי
, להתגבר עם הירידה בשחרור הסינפטי תחת אינהיביציההמרכזיים קיימת נטייה של הפוטנציאציה
ל באה לידי ביטוי בגרף כסטייה קלה של עקום ההתאמה "הנטייה הנ(אולם נטייה זו איננה מובהקת
). הליניארית מראשית הצירים
על מופע הרוויה של השחרור GABAראוי להדגיש ממצא נוסף המדגיש את ההבדל בהשפעת
האפקט (fig.14E)בטרמינלים הפרוקסימליים . על מופע העלייה שלומול ההשפעה, הסינפטי
בלבד ברמת הרוויה של השחרור 10%ריכוז זה גרם לירידה של . 40µM GABAהמוצג הוא של

73 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
בהשוואה לירידה של , (fig.12B) קצב התפתחות השחרורהסינפטי בתדרים גבוהים הודות להאטת
ממצא זה מדגיש את הרגישות . בשחרור הסינפטי בתגובה לסדרה קצרה בת חמישה גירויים50%
ואת ההשפעה הברורה של בחירת נוהל הגירוי , של סדרות גירויים קצרות בתדר גבוה לאינהיביציה
.על תוצאות הניסוי
_______________________
סיכום .5.4.3
ניתן ראשית לפרק את , יים בתדר גבוה על התגובה לסדרת גירוGABAלצורך תאור השפעת
. התגובה באופן מלאכותי לארבעה משתנים כך שניתן יהיה להתייחס לכל אחד מהם בנפרד
הפוטנציאציה של התגובה הבודדת עם התקדמות סדרת , (1Hz)המשתנים הם התגובה הבודדת
.רוויה עצמהורמת ה, עד לרוויהקצב התפתחות השחרור, הגירויים
אך ההשפעה על , חלה ירידה משמעותית בתגובה הבודדתGABAבטרמינלים המרכזיים עם מתן
במצב זה הפוטנציאציה המקסימלית נשארת דומה . הפוטנציאציה וקצב התפתחותה היא מינימלית
ינפטי יורדות באופן פרופורציוני לירידה ורמות הרוויה אליהם מגיע השחרור הס, לזו שבביקורת
. בתגובה הבודדת
קצב התפתחות והיא האטת , בטרמינלים הפרוקסימליים מתרחשת תופעה אחרת תחת אינהיביציה
במצב זה הפוטנציאציה מתחילה להתפתח בקינטיקה זהה . בעיקר בתדרי גירוי גבוהים, השחרור
עקב זאת . לביקורת היא ממשיכה להתפתח למשך מספר כפול של גירוייםאך בניגוד , לביקורת
במקרה זה הערכים המחושבים של הפוטנציאציה . רמת הרוויה כמעט ולא יורדת בתדרים הגבוהים
בו ניתן לזהות GABA -ממצא זה חשוב היות ומצביע על חלון בריכוזי ה. המקסימלית עולים
מבלי לזהות בהכרח את , ו בסדרות קצרות של גירוייםאינהיביציה ברורה בתגובות בודדות א
הסיבות האפשריות לתלות האפקט האינהיביטורי . ל בסדרות גירויים ארוכות יותר"האפקט הנ
. בתדר הגירוי נדונה בסוף הפרק
ולכן העובדה שמהלך , גבוהים יותר עצמת הפוטנציאציה של הטרמינלים יורדתGABAבריכוזי
. ותר אינה מפצה עוד על הירידה בשחרור הסינפטיהפוטנציאציה ממושך י

74 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
י גירוי האינהיביטור על התגובה לתדר"השפעת אינהיביציה המושרית ע .5.5
הקדמה .5.5.1
לפני השחרור מהטרמינל האקסיטטורי 2ms -פוטנציאל פעולה הפולש לטרמינל האינהיביטורי כ
,.Dudel & Kuffler, 1961; Parnas et al(מפעיל את האינהיביציה הפרסינפטית באופן אופטימלי
2000b(י הגדלת מוליכות כלור " אינהיביציה זו מתווכת בעיקר ע)רגישה ל- Picrotoxin( , ומכאן
.חשיבות התזמון המדויק של הפעלת הטרמינל האינהיביטורי
האקסון לאחרונה הצטברו עדויות המצביעות על כך שאינהיביציה המעוררת באמצעות גירוי
בעבודה זו הודגם אפקט אינהיביטורי . )Parnas et al., 2000b(האינהיביטורי מערבת מנגנון נוסף
בניגוד לגירוי המסורתי של הטרמינל האינהיביטורי לפני , חלקי תוך גירוי שני הטרמינלים בו זמנית
יר לתלות את פעילות אינהיביציה זו הודות לגירוי הבו זמני של הטרמינלים פחות סב. האקסיטטורי
חיזוק . שהאפקטיביות שלו מותנית בהקדמת הגירוי האינהיביטורי, במנגנון מוליכות הכלור שלהלן
ירידה האופיינית , נוסף לכך הוא שבזמן האינהיביציה לא נרשמה ירידה בכניסת הסידן לטרמינל
.C12°ה זו נעשה בטמפרטורה של יש להדגיש שהרישום בעבוד. GABAי "לאינהיביציה המושרית ע
של פוטנציאל shuntממצא זה מספק דרך פיזיולוגית לבודד מנגנון אינהיביטורי שלא נשען על
.ולבדוק את השפעתו על התגובה לתדר של הטרמינל, הפעולה האקסיטטורי
השפעת גירוי האינהיביטור על עקום התגובה לתדר של טרמינלים פרוקסימליים .5.5.2
הגירוי האינהיביטורי ניתן בו זמנית עם . ל בוצעה בטרמינלים פרוקסימליים"סדרת הניסויים הנ
. כאשר פיפטת הגירוי הונחה על האקסונים בסמוך ככל הניתן לאתר הרישום, הגירוי האקסיטטורי
ראה (האקסיטטורי והאינהיביטורי יחדיו: בשני האקסוניםAPמת הגירוי נקבעה כך שתעורר עוצ
במצב זה נרשם סיגנל פרסינפטי המורכב מסכום שני הסיגנלים הנפרדים לכל . ) גירוישיטות
הרבה בין הרביהק. fig.15A -בדוגמא לסיגנל זה ולרישומי תגובות אופייניות ניתן לראות . אקסון
זמן הקצר בין ארטיפקט הגירוי לסיגנל מרווח ה באה לידי ביטוי בםרישווהת הגירוי ודרוטאלק
האינהיביטורי על AP -עם גירוי האינהיביטור ניתן לזהות בבירור את התווספות אות ה. הפרסינפטי
של גירוי בו והאפקט , C19° בטמפרטורה של נערכוהניסויים ). כוכביות(גבי האות האקסיטטורי
. באותם טרמינליםGABA - טיפול בלזה שלהושווה על השחרור האינהיביטור זמני של

75 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
Figure 15 :השפעת גירוי האינהיביטור על התגובה לתדר בטרמינלים הפרוקסימליים
A : 80בתדר של אקסון תגובות מטרמינל פרוקסימלי אופייני לגירוי 15רישומיHz . משמאל תוך גירוי האקסון האקסיטטורי
את מימין ניתן לראות , פרסינפטיאות הה: כוכבית. אקסיטטורי ואינהיביטורי זמני של שני האקסונים ומימין גירוי בו, בלבד
ההשפעה של גירוי בו זמני של האינהיביטור מול מתן : B. הגידול הברור באות החל עם גירוי האקסון האינהיביטורי
GABA על התגובה לתדר (mean±se;n=5) , הגירוי האינהיביטורי משפיע באופן ). פרוקסימליים(באותם טרמינלים
על עקום GABAהשוואת ההשפעה של גירוי האינהיביטור מול : C. הומוגני על השחרור בכל ספקטרום תדרי הגירוי
דוגמא : D,E. גירוי האינהיביטור משמר היטב את דפוסי עקום הביקורת). ליניאריYציר (הפוטנציאציה המקסימלית
התפתחות : D. בניסוי בודד, 80Hzבתגובה לגירוי בתדר ור על משך התפתחות הפוטנציאציה להשפעת גירוי האינהיביט
ערכי הרוויה של התגובות : 80Hz (mean±Sd;n=8) .Eהתגובות בביקורת ותוך גירוי האינהיביטור לסדרות גירויים בתדר
GABAטנציאציה בניגוד להשפעת ניתן לראות שגירוי האינהיביטור לא מאריך את משך התפתחות הפו, 1 -מנורמלים ל
(40,80µM)באותו טרמינל .
1 10 100
1
10
E.D.
C.
slope 1.05
B.
A.
controlGABA 40 µMcontrol+inhibitor
prox
imal
qua
ntal
con
tent
stim. frequency Hz1 10 100
0
5
10
15
20
25
30 control GABA 40 µM control+inhibitor
prox
imal
pot
entia
tion
stim. frequency Hz
0 10 20 30 40 500
5
10
15
quan
tal c
onte
nt
controlcontrol+inhibitor
stimulus No.0 10 20 30
0.0
0.5
1.0
controlcontrol+inhibitor GABA 40µM GABA 80µMno
rmal
ized
qua
ntal
con
tent
stimulus No.
**
Excitor+Inhibitor 80HzExcitor 80Hz
1ms
0.25nA

76 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
ניתן . (fig.15B) בממוצע 22% - הביאה להנחתת עקום התגובה לתדר בזההפעלת נוהל גירוי
GABAבניגוד להשפעה האינהיביטורית של , לראות שההנחתה היא אחידה בכל תדרי הגירוי
(40µM)ההבדל בין גירוי האינהיביטור . ר הגירויהולכת ופוחתת עם עליית תד, באותם טרמינלים
הגירוי האינהיביטורי מפחית . (fig.15C) בולט בעקומי הפוטנציאציה המקסימלית GABAלבין מתן
. את השחרור הסינפטי מבלי להשפיע כלל על ערכי הפוטנציאציה המקסימלית
ן מובהק את דפוס בהם גירוי האינהיביטור משמר באופ, הבדל זה בולט במיוחד בתדרים הגבוהים
בניגוד להתמתנות הופעת הרוויה בתדרים גבוהים , הרוויה אליו מגיע השחרור הסינפטי בביקורת
ולמעשה , ביחס לביקורת214% -במקרה האחרון הפוטנציאציה המקסימלית עולה ל. GABAתחת
, םלסיכו . זהות לאלה שבביקורתGABA רמות הרוויה של השחרור הסינפטי תחת 80Hzבתדר של
י גירוי בו זמני של "תוצאות אלה מראות שאכן ניתן לגרום לאינהיביציה של השחרור הסינפטי ע
אינהיביציה זו לא גורמת לשינויים כלשהם בדפוס עקום הפוטנציאציה . הטרמינל האינהיביטורי
ומחזקת את הסברה שגירוי בו זמני של האינהיביטור פועל על השחרור הסינפטי , המקסימלית
. GABAנלים האקסיטטוריים באופן שונה ממתן מהטרמי
קצב התפתחות השחרורהשפעת גירוי האינהיביטור על .5.5.3
של סדרת תגובות לגירוי בתדר קצב התפתחות השחרורגירוי האינהיביטור לא גרם להאטת
דוגמא fig.15D -ניתן לראות ב. GABAבניגוד להארכה המובהקת הנגרמת בהשפעת , גבוה
למרות ירידת . בטרמינל פרוקסימלי, 80Hzלהשפעת האינהיביטור על סדרת תגובות בתדר גירוי של
. קצב התפתחות השחרור מערך הביקורת לא נרשמה כל האטה ב64%השחרור הסינפטי לערך של
גירויים 7 - ל4 -פר הגירויים למחצית ערך הרוויה מ עולה באותו טרמינל מסGABAבהשפעת
(fig.15E) , וזאת למרות ההשפעה הקלה יותר שלGABAמכאן . על רמת הרוויה של השחרור
גבוה GABAבריכוז . קצב התפתחות השחרורשהירידה בשחרור הסינפטי לא מחייבת גם האטה ב
תוצאות דומות התקבלו . גירויים20 - הסינפטי מגיע לרוויה רק כעבור כיותר ניתן לראות שהשחרור
כשמספר הגירויים למחצית ערך הרוויה , עבור כל חמשת הטרמינלים שנבדקו בסדרת ניסויים זו
-מופיע ב( בביקורת 5 - בהשוואה ל(mean±se;n=5) 0.57 ± 5.25 -בגירוי האינהיביטור שווה ל
fig.14C .(
_______________________

77 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
סיכום .5.5.4
גירוי בו זמני של האינהיביטור עם האקסון האקסיטטורי בתדרים שונים גורם לאינהיביציה
של פוטנציאל הפעולה האקסיטטוריshuntתוך ניצול מנגנון נוסף מלבד ליצירת פרסינפטית
)Parnas et al., 2000b(. עקום התגובה לתדר של של ברורה הנחתהלאינהיביציה זו הביאה
ולא השפיעה איכותית על , ירידה זו הייתה אחידה בכל תדרי הגירוי. הטרמינלים הפרוקסימליים
על הטרמינלים GABAלהשפעת לא זהההשפעה זו . עקום הפוטנציאציה של הטרמינל
האטת קצב התפתחות תוך , שהתאפיינה בהגברת הפוטנציאציה המקסימלית,הפרוקסימליים
.בתגובה לגירוי בתדר גבוה שחרורה
דיון .5.6
GABA -הבדל ברגישות טרמינלים מרכזיים ופרוקסימליים ל .5.6.1
על השחרור GABAא תוארה כאן עקומה מובנית המתארת את השפעת ריכוז למרות של
הבדלי הרגישות בינם לבין הטרמינלים המרכזיים היה בולט , הסינפטי מהטרמינלים הפרוקסימליים
על מנת להנחית את השחרור GABAלרוב נדרשו ריכוזים כפולים ויותר של . בכל הניסויים
ממצא זה לא מפתיע אם סוקרים את . מה לזו שבמרכזייםמהטרמינלים הפרוקסימליים לרמה דו
הטרמינלים . ההבדלים המורפולוגיים והפיזיולוגיים הידועים בין הטרמינלים מאזורי השריר השונים
וקיימות עדויות רבות לכך ,הפרוקסימליים הם טרמינלים גדולים הנמצאים בסמיכות לאקסון המרכזי
, י לטרמינלים אלה או עד לנקודה הסמוכה מאוד אליהםשפוטנציאל הפעולה פולש באופן אקטיב
פיזיולוגי זה צפוי -מבנה מורפולוגי. עובדה התורמת לשחרור הסינפטי המוגבר מטרמינלים אלה
ובמיוחד עם זו פועלת באמצעות , להקשות באופן ניכר על הנחתת השחרור הסינפטי מטרמינל כזה
סברה זו זוכה לחיזוק בדמות הממצא המורפולוגי . )Segev, 1990(הגדלת מוליכות הממברנה שלו
המעיד על שכיחות כפולה ויותר של טרמינלים אינהיביטוריים על גבי טרמינלים אקסיטטוריים
(Axoaxonal) באזור הפרוקסימלי של השריר )Govind et al., 1995( . שכיחות גבוהה זו עשויה
ואכן גירוי האינהיביטור נמצא אפקטיבי , ל בהנחתת השחרור הסינפטי"היחסי הנלפצות על הקושי
למרות ריבוי הטרמינלים האינהיביטוריים באזור זה מאידך . באוכלוסיית הטרמינלים הפרוקסימליים
על מנת להתגבר על מנגנוני הדפולריזציה GABAנדרשו ריכוזים גבוהים יותר של של השריר עדיין
, לאזור הטרמינלים האינהיביטוריים טובהGABAבמידה ומניחים שהחדירות של . להאבטרמינלים

78 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
שקרוב לודאי , יםהאינהיביטוריהטרמינלים על היעילות הגבוהה של סתירה לכאורה זו מצביעה
ה יעילה של מאפשרים הנחתבאופן זה ו, בלבד גבוהים בנקודות ספציפיותGABAמפרישים ריכוזי
הסמיכות בין בהקשר זה נשאלת השאלה מה מידת . באמבט GABA שלןבהשוואה למת, השחרור
במידה ובטרמינלים . בטרמינלים הפרוקסימליים ביחס למרכזייםאתרי השחרורלתעלות הסידן
הסתברות (הפרוקסימליים תעלות הסידו אכן סמוכות יותר לאתר השחרור כפי שהוצע קודם לכן
יש צורך בפתיחת מוליכות גבוהה יותר על מנת , )Meinrenken et al., 2002: ראה, גבוהה לשחרור
מאידך הטרמינלים האינהיביטוריים . גבוהים יותרGABAכלומר ריכוזי , ליצור אינהיביציה אפקטיבית
כך שיכולתם להפעיל אינהיביציה (Govind et al., 1995) ממוקמים בסמיכות רבה לאתרי השחרור
. יחסיתאפקטיבית גבוהה
על פני varicositiesל הטרמינלים המרכזיים קטנים יותר ובנויים מורפולוגית בצורה של "בניגוד להנ
לטרמינלים אלה הוא בעל אופי שטף המטען המגיע. שלוחות ארוכות של האקסון האקסיטטורי
" צוואר"ה יאזוראף באחד מת הטרמינל או ממברנולכן די בהגדלה מעטה יחסית במוליכות , פסיבי
ובשחרור , על מנת לפגום משמעותית ברמת הדפולריזציה בטרמינלאליו של האקסון המוליך
(Axoaxonal)טרמינלים האינהיביטוריים בהתאם לכך צפיפות ה. )Segev, 1990(הסינפטי ממנו
ואף קיימות עדויות לכך שהשחרור הסינפטי מחלק , )Govind et al., 1995(באזור זה נמוכה יותר
בהנחה שאכן עבר דיפוזיה ( GABAמהטרמינלים לא הושפע כלל ממתן ריכוזים גבוהים של
עדויות אלה מצביעות על כך שחלק . )Fischer & Parnas, 1996a() לממברנת הטרמינל
קיימת .GABA -סינפטיים ל-מהטרמינלים המרכזיים לא מכילים רצפטורים סינפטיים או חוץ
מהמדיום החוץ GABA נמוכה של דיפוזיהמ סובלת טרמינליםשתת קבוצה קטנה זו שלאפשרות
סיבה ניראית לעין להנחה אולם אין , נעדרתאי לאזור ממברנת הטרמינל ולכן האפקט האינהיביטורי
שאינם נתונים באזור זה מתארות טרמינלים אכן מה גם שהעדויות המורפולוגיות לעיל , זו
ניסויים שלושהבקרוב לודאי שמסיבות אלה .)Govind et al., 1995( לאינהיביציה פרסינפטית
עם האקסון האקסיטטוריתזמני-ובהאינהיביטור גורה םבמהלכו , אלהמרכזייםבטרמינלים שנערכו
.)תוצאות ניסויים אלה לא הוצגו (בשחרור הסינפטיירידה עקבית נצפתהלא
? מה עומד בבסיס האינהיביציה אם לא הפחתה בכניסת הסידן לטרמינל .5.6.2
כלומר , המאפיינים העיקריים של עקום התגובה לתדר של טרמינל הם נקודת ההתחלה שלו
תוצאות הפרק הקודם הצביעו . ושיפוע העקום, ) לצורך העניין1Hzבתדר גירוי של (השחרור הבודד

79 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
להשפיע ביחס הפוך ותנוטיים כניסת הסידן לטרמינלים המרכזשמניפולציות המשפיעות על על כך
לפיכך . ולהפך, כלומר ירידה בשחרור הבודד עם עליה בשיפוע העקום, על שני מאפיינים אלה
י הפחתה בכניסת סידן לטרמינל הרי שניתן היה "עבעיקר במידה והאפקט האינהיביטורי מתווך
תוצאות הניסויים מראות . ירידה בשחרור הבודדלצפות לעליה בשיפוע העקום בד בבד עם ה
הנחיתה את השחרור הסינפטי מבלי בטרמינלים המרכזיים GABAי "שהשריית אינהיביציה ע
.שלא בדומה להפחתת ריכוז הסידן החוץ תאי, להשפיע על שיפוע העקום
לווה ל יש מקום לעמוד על משמעות ההבדל בין הנחתת השחרור הסינפטי המ"לאור התוצאות הנ
במקרה . להנחתת השחרור שאיננה מלווה בשינוי שיפוע העקום ,שחרור-בעליית שיפוע העקום תדר
משמעות העלייה בשיפוע העקום היא שהשחרור כעת הוא פונקציה , של הפחתת ריכוז הסידן
ss[+Ca2]ה יותר של גבוה יחסיתקרוב לודאי עקב תרומה (מעריכית גבוהה יותר של תדר הגירוי
שחרור הסינפטי בהשוואה בעליית תדר הגירוי גוררת עלייה חדה יותר ,במילים אחרות. )רורלשח
מסיבה זו קיים באופן תיאורטי תדר גירוי גבוה מספיק כך שהשחרור הסינפטי המונחת . לביקורת
. )בהנחה שלא מתפתחת רוויה (דת הפתיחה הנמוכה יחסיתלמרות נקו, ידביק את זה של הביקורת
לשם דוגמא נניח תהליך המנחית את . יעילות ההנחתה הולכת ופוחתת עם עליית תדר הגירויכלומר
במקרה . 1.6 - ל1.3 -תוך עליית שיפוע העקום מ, מהביקורת25%לכדי 1Hz -השחרור הבודד ב
בזכות העלייה , הנחתה ידביק וישתווה לזה של הביקורת השחרור שעבר100Hzזה בתדר של
. בשיפוע העקום
ועשוי להסביר גם את ההבדלים בין , תהליך זה אופייני כאמור לירידה בכניסת הסידן לטרמינל
יתכן ותהליך ). סמוכים או מרוחקים מהאקסון המרכזי(הקבוצות השונות של הטרמינלים המרכזיים
. GABAי ריכוזים נמוכים של "נלים הפרוקסימליים בזמן אינהיביציה עדומה לזה מתרחש גם בטרמי
.בהמשך פירוט נוסף בעניין זה
תוצאות פרק זה מראות שהאינהיביציה ככלל אינה משנה את שיפוע עקום התגובה לתדר ולפיכך
,מכאן שהמנגנון שבבסיסה שונה. פועלת על השחרור הסינפטי באופן אחיד גם בתדרי גירוי גבוהים
המהווה כלי מלאכותי בלבד , ים המרכזייםויעיל יותר בהשוואה להפחתת כניסת הסידן לטרמינל
? נשאלת השאלה אילו מנגנונים עומדים בבסיס פעולת אינהיביציה זו". אינהיביציה"להשריית
ניתן , בהשוואה לגירוי האינהיביטור, על השחרור הסינפטיGABAאת ההשפעה השונה של מתן
סינפטיים על גבי ממברנת הטרמינלים -מניחים שקיימים רצפטורים אינהיביטוריים חוץלהסביר אם
בחלק מהטרמינלים המרכזיים כפי שצוין לעיל ניתן לשלול קיום רצפטורים (או האקסון האקסיטטורי

80 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
) תא השריר(קיומם של רצפטורים אינהיביטוריים כאלה תואר בממברנת התא הפוסטסינפטי ). אלה
גרמה GABAהפעלת רצפטורים אלה באמצעות מתן . )Gunzel & Galler, 2000(רט של אותו פרפ
תופעה שלא נרשמה עם , ) דרך התעלות-HCO3מעבר של ( של התא pH -לירידה מובהקת ב
תופעה דומה תוארה גם בנוגע . הפעלת הרצפטורים הסינפטיים באמצעות גירוי האינהיביטור
,Onodera & Takeuchi(ורים האקסיטטוריים הפוססינפטיים באותו פרפרט לאוכלוסיית הרצפט
י הרצפטורים הסינפטיים " גרם לירידה בשחרור שתווכה עKainateבמקרה האחרון מתן . )1980
ממצאים אלה מניחים יסוד . סינפטיים-י הרצפטורים החוץ"בהשוואה לעליה בשחרור שתווכה ע
GABAכלומר מתן דיפוזי של , דומה מתרחשת גם בממברנת התא הפרסינפטילאפשרות שתופעה
תשובה טובה לסוגיה זו ניתן . עשוי להשפיע על השחרור הסינפטי באופן שונה מגירוי האינהיביטור
תוך שימוש (רישום הזמן באלקטרודה חלל נפח ה ישירות לGABAלקבל באמצעות אפליקציה של
. כפי שנעשה כאןופן דיפוזי ולא בא,)במערכת מתאימה לכך
הניסיון להסביר את ההשפעה השונה של גירוי , GABAאם נתעלם לצורך העניין מהשפעת
shunt -מתוך הנחה שמנגנון ה, האינהיביטור והפחתה בריכוז סידן חוץ תאי על השחרור הסינפטי
ת שטף הסידן מפחיתה אshuntהפעלת . נתקל בקשיים מהותיים, הוא האחראי היחידי לפעולה זו
ואילו הפחתת ריכוז הסידן החוץ תאי , החודר לטרמינל הודות להנחתת גל הדפולריזציה הפולש אליו
ל הירידה הנצפית "בשני המקרים הנ). VCa) Nernst -מפחיתה את שטף הסידן עקב ירידה ב
וספק אם הסיבות השונות , י הפחתת שטף הסידן החודר לטרמינל"בשחרור הסינפטי מתווכת ע
.להפחתה זו עשויות להסביר את ההבדלים המהותיים שתוארו בירידה בשחרור הסינפטי
קיים מנגנון בטרמינלים המרכזיים מכאן שתוצאות העבודה הנוכחית מחזקות את ההנחה ש
בנוסף (אינהיביטורי המנחית את השחרור הסינפטי מבלי להפחית את כניסת הסידן לטרמינל
,.Parnas et al( .Parnas et alי "וכחות מנגנון כזה הוצעה ענ). הקלאסיshunt -למנגנון ה
2000b( , מתוך ניסויים בהם נעשה שימוש בנוהל בו גורו שני האקסונים בו זמנית) האקסיטטורי
,.Slutsky et al( .Slutsky et alהצעה זו התבססה בין השאר על ממצאים של ). והאינהיביטורי
, י רצפטור מוסקריני"שהדגימו אינהיביציה פרסינפטית המתווכת ע, בטרמינלים של צפרדע)1999
. ללא הנחתה של זרמי הסידן בטרמינל
יתכן ומנגנון דומה עומד בבסיס ההנחתה של השחרור הסינפטי בכל ספקטרום תדרי הגירוי
. של הטרמינלים לא משתנהכך שתחת אינהיביציה שיפוע עקום התגובה לתדר, בטרמינלים אלה

81 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
הקלאסי shunt -חיזוק נוסף לנוכחות מנגנון אינהיביטורי מתווך מוליכות שפעולתו שונה מזו של ה
.מובאת בפרק הבא
?ממה נובעים ההבדלים בין התגובה לתדר של טרמינלים מרכזיים ופרוקסימליים .5.6.3
לתדר של הטרמינלים כפי שנדון בפרק הקודם קיימים הבדלים ברורים בין עקום התגובה
העובדה שאינהיביציה . כשאחד הבולטים בהם הוא שיפוע העקום, הפרוקסימליים למרכזיים
מראה שאין לה השפעה , פרסינפטית לא משפיעה על שיפוע עקום התגובה לתדר של הטרמינלים
ק כפי שצוין בפר. משמעותית על חלק מאותם גורמים פיזיולוגיים העומדים בבסיס ההבדל ביניהם
ולכן , גם שינויים בכניסת הסידן לטרמינל לא מגשרים על מכלול ההבדלים בין הטרמינלים, הקודם
. נראה שמקור ההבדלים בין הטרמינלים נעוץ בגורמים נוספים
לקבוע אילו מההבדלים הפיזיולוגיים שאלה שההתייחסות המדעית אליה טרם מוצתה היא הניסיון
ובמיוחד , הברורים ביניהםורפולוגייםם באופן ישיר מההבדלים המבין הטרמינלים השונים נובעי
עקב הקשיים הטכניים הכרוכים .כפי שהוזכר לעיל, הסמיכות בין תעלות הסידן ואתרי השחרור
ת ריכוז סידן יהדמייש מקום לתקוף שאלה זו באמצעות , ברישום ישיר מהטרמינלים מרכזיים
.ותו פרפרט מחקר מורפולוגי של אמשולבת עם
קצב התפתחות השחרורהקשר בין הפעלת אינהיביציה ל .5.6.4
של השחרורקצב התפתחות אחת התוצאות המעניינות בפרק זה היא השפעת האינהיביציה על
בטרמינלים הפרוקסימליים מתקבלת הפרדה בין . התגובות בתוך סדרת הגירויים בתדר גבוה
GABAעם מתן . למספר הגירויים שהתפתחות זו נמשכת, הפוטנציאציה מגירוי לגירויהתפתחות
אולם בדיקה של סדרת , לא נמדד שינוי מובהק בהתפתחות עצמת הפוטנציאציה מגירוי לגירוי
ובות ארוכה יותר מראה שהפוטנציאציה תחת אינהיביציה ממשיכה להתפתח למשך מספר תג
מכאן שהפוטנציאציה המקסימלית בטרמינלים . בהשוואה לביקורת, גירויים כפול בקירוב
. GABAהפרוקסימליים עולה תחת ריכוזים נמוכים יחסית של
עם זאת . קצב התפתחות השחרורבבטרמינלים המרכזיים בזמן אינהיביציה לא נרשם שינוי דומה
גירוי ישיר בעוצמה : כפי שמוזכר בפרק הקודם, ניתן לעורר בטרמינלים אלה את התופעה ההפוכה
מאיץ באופן , המעוררת בטרמינלים אלה שחרור הסינפטי חזק יותר מזה שמעורר הגירוי הפיזיולוגי
). מקצר את מספר הגירויים להגעה למחצית ערך הרוויה ( התפתחות השחרורקצבמובהק את

82 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
ניתן , )Cooper et al. )Cooper et al., 1995aובהסתמך על תוצאות עבודתם של , מהאמור לעיל
, ידן בטרמינל הוא פונקציה של הקינטיקה בה מתפתח ריכוז הסקצב התפתחות השחרורלשער ש
כאןכיוון שריכוז הסידן בטרמינל לא נמדד נת לכך עדות ישירהלא נית (עם מתן הגירויים החוזרים
, שחרוריותר של ה) איטית(ירידה בכניסת הסידן לטרמינל תביא להתפתחות ממושכת . )באופן ישיר
בה )Winslow et al. )Winslow et al., 1994חיזוק למסקנה זו מתקבל מעבודתם של . ולהפך
בניסוי זה למרות הירידה הניכרת בשחרור הסינפטי . BAPTA -נבדקה הפוטנציאציה לאחר טיפול ב
אולם גירוי , )100Hz, חמישה גירויים(לא חל שינוי בפוטנציאציה בתגובה לסדרת גירויים קצרה
להתפתח עד לפיצוי מלא של התגובה ךממשישחרור הראה שה) 20Hz, גירויים600(ממושך
. כלומר הפוטנציאציה המקסימלית עלתה באופן ניכר, בהשוואה לביקורת
בטרמינלים הפרוקסימליים אינהיביציה בריכוזי . ל על פעולת האינהיביציה"כעת ניתן להשליך את הנ
GABA 40שלµM מבלי לפגוע כמעט ברמות , קצב התפתחות השחרורמאיטה באופן מובהק את
של פוטנציאל הפעולה shuntי "אפשר ואפקט זה מתווך בעיקרו ע. הרוויה של השחרור הסינפטי
בריכוזים . )Parnas et al., 2000b(המביא להתפתחות איטית יותר של ריכוז הסידן בטרמינל
אולם לא בפרופורציה , קצב התפתחות השחרורספת ב ניכרת האטה נוGABAגבוהים יותר של
גבוהים GABAריכוזי ) 1. לממצא זה שני הסברים אפשריים. לירידה הניכרת בשחרור הסינפטי
גורמים להפחתת כניסת הסידן לטרמינל לרמה הגורמת לפגיעה משמעותית יותר בשחרור הסינפטי
אלה מפעילים מנגנון אינהיביטורי נוסף GABAריכוזי ) 2. קצב התפתחות השחרורופחות להאטת
.המנחית בעיקר את השחרור הסינפטי, shunt -מלבד ל
קצב לא נרשם בזמן המתן כל שינוי ב, GABA - רגישותם הרבה ללמרות, בטרמינלים המרכזיים
shunt -יתכן ובטרמינלים אלה מנגנון ה. מעלה מספר אפשרויותתוצאה זו . התפתחות השחרור
י עבודות הגירוי "אפשרות הנתמכת ע, הקלאסי הוא לאו דווקא המנגנון האינהיביטורי הדומיננטי
.Parnas et al ועבודתם של )Fischer & Parnas)Fischer & Parnas, 1996b, a הישיר של
)Parnas et al., 2000b( ,בה גורו שני האקסונים בו זמנית .
פחות רגיש לשינויים בקינטיקת קצב התפתחות השחרוראפשרות נוספת היא שבטרמינלים אלה
בטרמינלים המרכזיים ריכוז הסידן מתפתח לרמות . התפתחות הסידן בהשוואה לשחרור הסינפטי
ויתכן ושינויים בקינטיקה זו עשויים , )Cooper et al., 1995a( איטית יותר נמוכות יותר ובצורה
. קצב התפתחות השחרורהשחרור הסינפטי ופחות על מידת להשפיע יותר על

83 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
יי היבטים נוספים של הקשר בין הפעלת אינהיביציה להאטת קצב הפוטנציאציה מתקבלים מניסו
קצב התפתחות בניסויים אלה גירוי בו זמני של האינהיביטור לא האט את . גירוי האינהיביטור
תוצאה זו עולה בקנה אחד עם . למרות הירידה המובהקת שנצפתה בשחרור הסינפטיהשחרור
של פוטנציאל לבד ב shunt -האמור לעיל מאחר וגירוי בו זמני של האינהיביטור לא צפוי לגרום ל
,.Parnas et al(ולפיכך לא אמור להשפיע על קינטיקת התפתחות הסידן בטרמינל , הפעולה
2000b( .
קיים הסבר קצב התפתחות השחרוריש לציין שלחוסר ההשפעה של גירוי האינהיביטור על
י האינהיביטור בתדר גבוה גורם לפוטנציאציה של הסבר זה נעוץ בעובדה שגירו. אלטרנטיבי
פוטנציאציה אינהיביטורית זו מתפתחת באופן מקביל . האינהיביציה עם התקדמות סדרת הגירויים
ועקב כך עשויה להפסיק את התפתחות השחרור הסינפטי , לפוטנציאציה של התגובה האקסיטטורית
מאחר ומידת GABA - לא מתרחש בניסויי הנטרול זה. ולהביא לרוויה מוקדמת ונמוכה יותר שלו
קרוב לודאי . בלבדGABAותלויה בריכוז , האינהיביציה המופעלת בניסויים אלה קבועה
אולם לא סביר , קצב התפתחות השחרורשלפוטנציאציה האינהיביטורית תפקיד מסוים בשימור
.GABA - האופיינית לניסויי הקצב התפתחות השחרורשהיא מקזזת באופן מדויק את ההאטה ב
נגזר מקינטיקת התפתחות קצב התפתחות השחרורש התוצאות מספקות עדות עקיפה לכך ,לסיכום
קצב התפתחות השחרורבטרמינלים הפרוקסימליים . )שלא נמדד ישירות כאן (ריכוז הסידן בטרמינל
,.Cooper et al(בהתאם להתפתחות המהירה יותר של ריכוז הסידן בטרמינלים אלה , מהיר יותר
1995a( . בהשפעתGABAמואטקצב התפתחות השחרור חל עיכוב בהתפתחות ריכוז הסידן ו .
,.Parnas et al(יתה את כניסת הסידן לטרמינל הפעלה בו זמנית של האינהיביטור לא מפח
2000b( , קצב התפתחות השחרורולפיכך לא מאיטה את .
, קצב התפתחות השחרור לא מאיטה את )GABAמתן (בטרמינלים המרכזיים האינהיביציה
ממצא זה . מפחיתה באופן משמעותי את כניסת הסידן לטרמינלובהתאם לכך ניתן להניח שאינה
מחזק את טענת חלק מהחוקרים בדבר קיום מנגנון אינהיביטורי נוסף המפחית את השחרור
,.Parnas et al(מבלי להשפיע מהותית על שטף הסידן לתוך הטרמינלים המרכזיים , הסינפטי
2000b( .

84 השפעת אינהיביציה פרסינפטית על התגובה לגירוי בתדר _________________________________________________________________________________
שחרור הבודדעמידות הפוטנציאציה לאינהיביציה בהשוואה להעמידות היחסית של ה .5.6.5
תוך גירוי בסדרות של , סעיף זה מתייחס לפוטנציאציה כפי שהופיעה בניסויי גירוי האקסון
מתן (בו הפעלה של אינהיביציה פרסינפטית " חלון אינהיביטורי"קיים . 100Hzחמישה גירויים בתדר
GABA (מבלי לפגוע בעצמת , מהביקורת50%כים סביב מפחיתה את השחרור הסינפטי לער
הפעלת , השונה בין טרמינל אחד לשני, יש להדגיש שמעבר לסף מסוים. הפוטנציאציה של השחרור
עמידות יחסית של . אינהיביציה חזקה יותר גרמה ברוב הטרמינלים לירידה הדרגתית בפוטנציאציה
למרות תכונות , רמינלים שנבדקו בפרק זההפוטנציאציה לאינהיביציה הייתה תוצאה עקבית בכל הט
. השחרור השונות שלהם ולמרות ההשפעה המובהקת של אותה אינהיביציה על השחרור הסינפטי
עמידות יחסית של . עקביות ממצא זה מעידה על תכונה בסיסית של מנגנון השחרור הסינפטי
;Zucker, 1974(פרט פוטנציאציה לירידה בריכוז הסידן בטרמינל תוארה זה מכבר באותו פר
Parnas et al., 1982c; Winslow et al., 1994( ,במידה , ל"ועשויה להסביר את הממצא הנ
במקרה זה תוצאות הפרק מחזקות את . י ירידה בכניסת הסידן לטרמינל"והאינהיביציה אכן תווכה ע
הם בעלי ) פסיליטציה(ציאציה הגורסת שאתרי הקישור האחראים לפוטנ, אחת הסברות המקובלות
ומכאן ההבדל בין , זיקה גבוהה יותר לסידן בהשוואה לאלה האחראים על השחרור הסינפטי
יש לציין שטרם קיים קונצנזוס בנוגע לסוגיה . רגישויות שני התהליכים לירידה בריכוז הסידן בטרמינל
המראות רגישות גבוההבשנים האחרונות התפרסמו עבודות רבותכפי שצויין קודם לכן ו, זו
(~10µM)לסקירה ראה. של מנגנון השחרור לסידן - Augustine) 2001(, Meinrenken et al.
, ישנן עבודות השוללות שינוי בזיקה של מנגנון השחרור לסידן במהלך פסיליטציה כמו כן ).2003(
. בפרפרט ממערכת עצבים מרכזית)Felmy et al., )Felmy et al., 2003דוגמת העבודה של
התוך הסידן של ריכוזהפסיליטציה לסיכום סופרלינארי ם חלק ניכר מבעבודה זו הכותבים מייחסי
או מניפולציה , אופן בו אינהיביציה פרסינפטיתטרם קיימת הסכמה בנוגע לל "לאור הנ, לסיכום. תאי
השפעה על הפוטנציאציה של פרסינפטית אחרת עשויה לגרור ירידה מובהקת בשחרור ללא
ם מורפולוגיים ואחרים במאפייני כמו כן נראה שטרמינלים מפרפרטים שונים נבדלים .התגובה
מנגנון והשפעתו עלהסידן התוך תאי ריכוז , תעלות הסידןבין שווי המשקל הקובעים את אופי
.השחרור

85
י "היבטים מנגנוניים של האינהיביציה המתווכת ע .6GABA
___________________________________________________________________
מבוא .6.1ונים על גבי הפרקים הקודמים בעבודה זו תארו את מאפייני התגובה לתדר של הטרמינלים הש
פרק זה ממשיך בבחינת . ואת השפעת האינהיביציה הפרסינפטית על תגובה זו, opener -שריר ה
תוך ניסיון לבודד בכלים פיזיולוגיים את , המסקנות העיקריות הנובעות מהפרקים הקודמים
הניסויים בפרק זה בוצעו. המנגנונים העומדים בבסיס האינהיביציה הפרסינפטית שתוארה לעיל
: לרוב תוך הפרדה ברורה בין שתי תת האוכלוסיות, בעיקר באוכלוסיית הטרמינלים המרכזיים
.והמרוחקות ממנו, הסמוכות לאקסון המרכזי
אחת הסוגיות המרכזיות העולה מהפרקים הקודמים היא שאלת נוכחות מנגנון אינהיביטורי מלבד
ש להראות אפקט אינהיביטורי המושתת בטרמינל יshuntעל מנת לאתר . הקלאסיshunt -מנגנון ה
נניח לצורך ). דפולריזציה בפרט(הפועלת להפחתת שינויים במתח הממברנה , על שינוי מוליכות
באופן חוץ , כך שניתן לשלוט בקלות, )פסיביות(העניין שהממברנה הופכת לבעלת תכונות ליניאריות
בתנאים אלה הפעלת . תח הפיפטהעל מפל המתח על פני פיסת הממברנה הנמצאת בצמוד לפ, תאי
על עקרון . בזמן מתן גירוי קצר לא יגרום לשינוי במפל המתח על פני פיסת הממברנהshuntמנגנון
, הופך את הממברנה לפסיבית TTX (0.5µM)מתן . זה מבוסס החלק הראשון של הפרק הנוכחי
דה ובתנאים אלה האפקט במי. י שימוש בגירוי ישיר"והשליטה על מתח ממברנת הטרמינל מושגת ע
shunt -הרי שניתן לשייך אינהיביציה זו ל) בהשוואה לביקורת בגירוי האקסון(האינהיביטורי נעלם
.של גל המתח הפולש לטרמינל
בחלק השני של הפרק מרוכזות תוצאות הניסויים בהם נעשה שימוש בפרמקולוגיה על מנת לסווג
מעורבות של שני . סינפטית בטרמינלים המרכזייםאת המוליכות המתווכת את האינהיביציה הפר
, 5.1.1ראה מבוא ( בתווך האינהיביציה ידועה כבר מזה שנים GABAA GABABהרצפטורים
. ומאששות מחקרים קודמים מובאות בקצרה, ומסיבה זו תוצאות הנוגעות לרצפטורים אלה) 5.1.4
נוהל הגירוי ששימש . GABAC ל נבדקה גם מעורבותו של רצפטור מסוג"בנוסף לרצפטורים הנ
נוהל זה . אחת לשתי שניות, 100Hzלניסויי הפרמקולוגיה כלל סדרות של חמישה גירויים בתדר
: נבחר כיוון שהתגובה שמעורר רגישה לשלושה מופעים אפשריים ושונים לכאורה של אינהיביציה

GABA 86י "היבטים מנגנוניים של האינהיביציה המתווכת ע _________________________________________________________________________________
קצב ושינויים ב) יה או ירידהעל(של התגובה פוטנציאציה בשינויים , ירידה בתכולה הקואנטלית
ומתואר באופן , האפקט נמדד על חמש התגובות בסדרה). התפתחות מאוחרת (התפתחות השחרור
בחלק מהמקרים בוצעו הניסויים תוך ). באחוזים ביחס לביקורת(כמותי על התגובה החמישית
.ירגירוי האקסון וגירוי יש: שימוש בשתי שיטות הגירוי
על התגובות לגירוי ישיר מול גירוי האקסון GABAהשפעת .6.2 על טרמינלים מרכזיים תחת GABAבחלק זה מפורטות תוצאות הניסויים בהם נבחנה השפעת
בסדרה הראשונה נרשמו . בהשוואה להשפעה הנרשמת בגירוי האקסון, )TTXעם ובלי (גירוי ישיר
וממוקמים בסיבי השריר , מאד לאקסון המרכזיהתגובות משישה טרמינלים מרכזיים הסמוכים
טרמינלים אלה מראים תכונות האופייניות יותר . הפרוקסימליים יותר מבין הסיבים המרכזיים
נוהל הגירוי . כמו הסתברות גבוהה מאד לשחרור ותכולה קואנטלית גבוהה, לטרמינלים פרוקסימליים
מאחר והשחרור הסינפטי 40µM נקבע על GABA -ריכוז ה. 100Hzכלל חמישה גירויים בתדר
והמטרה הייתה לעורר אינהיביציה חזקה , מטרמינלים אלה דומה יותר לזה של הפרוקסימליים
. (100Hz)ת התגובות בסדרהאפקט האינהיביטורי נבדק על התגובה החמישית . יחסית
פן דומה גם על הטרמינלים על התגובה המעוררת בגירוי ישיר נבחנה באוGABAהשפעת
עבור טרמינלים אלה כבר קיימות עדויות המצביעות על ירידה . המרכזיים המרוחקים מהאקסון
י "הנחסמת ע, )10µM -פחות מ (GABAברורה בתגובות לגירוי ישיר עם מתן ריכוזים נמוכים של
& GABAB .)Fischerאנטגוניסט לרצפטור , 2OH-saclofen - וpicrotoxin -טיפול משולב ב
Parnas, 1996a(.
אינהיביציה בגירוי ישיר מול גירוי האקסון בטרמינלים מרכזיים הסמוכים לאקסון .6.2.1
. fig.16A - מוצגות בGABA -דוגמאות לתגובות בגירוי אקסון וגירוי ישיר בביקורת ותחת טיפול ב
יר מעורר תגובת ביקורת בעלת ערך דומה מאוד לזה שהגירוי הישfig.16B -ראשית ניתן לראות ב
תגובת הביקורת צונחת באופן TTX -ואילו לאחר טיפול ב, של תגובת הביקורת לגירוי האקסון
ומעיד , בנוגע לטרמינלים הפרוקסימליים)Dudel )Dudel, 1983י "ממצא זהה תואר ע. משמעותי
מעורר למעשה פוטנציאל פעולה באקסון ) ובר סף מסויםבמידה וע(על כך שהגירוי הישיר
תכונת הסף TTX -לאחר טיפול ב. הפולש לטרמינל בדומה למתרחש בגירוי האקסון, הפרטרמינלי
במצב זה גירוי ישיר על ספי שהיה מביא לתגובה . של הממברנה נעלמת והיא הופכת ליניארית

GABA 87י "היבטים מנגנוניים של האינהיביציה המתווכת ע _________________________________________________________________________________
Figure 16 : השפעתGABAגובה לגירוי עצב מול התגובה לגירוי ישיר על הת
A: דוגמאות לרישומי תגובות מטרמינל מרכזי מרוחק בגירוי עצב וישיר (0.7ms;1µA) ,80תדר בHzביקורת ולאחר מתן ב
GABA (15µM) . ניתן לראות את ההשפעה החזקה על התגובה לגירוי ישירB : השוואת השפעתGABA על השחרור
השחרור בתגובה לגירוי החמישי בסדרת גירויים (TTXוישיר עם , ישיר, ים הסמוכים לאקסון בגירוי עצבמטרמינלים מרכזי
TTXעם על השחרור בגירוי ישיר picrotoxin (Ptx. 50µM) - וGABAהשפעת : 100Hz ,mean±se;n=6.( Cבתדר
יתן לראות ששלוש עוצמות הגירוי מעוררות נ. (mean±se;n=5)צמות גירוי שונות ובשלוש ע, באותה אוכלוסיית טרמינלים
על השחרור GABA ,picrotoxin, 2OH-saclofenהשפעת : A .D -שחרור המכסה את כל ספקטרום השחרור המופיע ב
כל נקודה מייצגת ממוצע עם סטיית תקן של חמש . TTX (1Hz)מטרמינל מרכזי מרוחק בתגובה לגירוי ישיר בנוכחות
בשישה ) Yציר (לזה שבגירוי ישיר ) Xציר ( על השחרור בגירוי עצב GABAשוואה בין השפעת ה: E. תגובות סמוכות
מיקום המשושים מציין את הירידה יחסית לביקורת ברמות הרוויה המקסימליות של השחרור . טרמינלים מרכזיים מרוחקים
הכוכביות . תקן של ששת הטרמינליםהמחומש המלא מציין ממוצע ושגיאת . בכל סוג גירוי60-80Hz)בתדרי גירוי של (
החץ מסמן את הטרמינל ממנו נלקחו . מציינות את הטרמינלים בהם השחרור בשני סוגי הגירוי היה זהה בביקורת
.1ניתן לראות בבירור שכל הנקודות נמצאות מתחת לקו המקווקו המציין שיפוע של . A -הרישומים המופיעים ב
0
1
2
3
4
5
6
7
C.
direct+TTXdirectaxon
E.D.
B.
A.
control GABA (40µM) wash
quan
tal c
onte
nt
stimulation method
0
1
2
3
4
5
6
7
0.7 1.0 1.3
control+TTX GABA (50µM) Ptx. WASH
quan
tal c
onte
nt, d
irect
stim
.stimulation amplitude (µA)
0 500 1000 15000
2
4
6
8
direct stim.
axon stim.
+2OH-saclofen+picrotoxinGABA
quan
tal c
onte
nt, d
irect
stim
.
t sec0.0 0.2 0.4 0.6 0.8 1.0
0.0
0.2
0.4
0.6
0.8
1.0
#
*
**
normalized GABA inhibited release
mean +/- se
dire
ct s
timul
atio
n
nerve stimulation
1ms
0.25nA
GABA (15µM)control
X Axis Title

GABA 88י "היבטים מנגנוניים של האינהיביציה המתווכת ע _________________________________________________________________________________
תופעה זו תוארה . גובה התלויה ישירות בעוצמת הגירוי בלבדלתכעת מביא , מקסימלית קודם לכן
.עצמת גירוי בטרמינלים הפרוקסימליים- באמצעות בניית עקומת שחרור)Dudel )Dudel, 1983י "ע
הממצא העיקרי מניסויים אלה הוא היעדר כמעט מוחלט של אינהיביציה בגירוי ישיר לאחר מתן
TTX . י "על כך שהאינהיביציה הניכרת שנרשמה בביקורת מתווכת קרוב לודאי עממצא זה מעיד
shuntמנגנון זה מאבד חלק ניכר מיעילותו לאחר טיפול ב. של פוטנציאל הפעולה הפולש לטרמינל-
TTX וההשפעה של GABAהופכת זניחה .
באופן יורדTTX -ל מחייבת הסתייגות מאחר והשחרור הסינפטי לאחר טיפול ב"התוצאה הנ
מסיבה זו יתכן לכאורה שבגלל הירידה בשחרור האינהיביציה . TTXמשמעותי בהשוואה לגירוי ללא
על מנת לשלול . או שהשפעתה על השחרור הסינפטי פחות בולטת לעין, הופכת פחות אפקטיבית
אפשרות זו בוצעה סדרת ניסויים משלימה מתוך כוונה לבדוק האם האפקט האינהיביטורי פועל
בניסויים אלה נבדקה השפעת האינהיביציה . פן שונה על תגובות המעוררות בעצמות גירוי שונותבאו
בחמישה טרמינלים נוספים מאותה , בשלוש עצמות גירוי שונותTTXעל השחרור הסינפטי תחת
. על מנת להבטיח אינהיביציה חזקה, 50µM שנבחר לניסויים אלה היה GABA -ריכוז ה. אוכלוסייה
שהגירוי החלש יותר מניב תגובה בעלת ערך דומה לזו של השחרור fig.16C - לראות בניתן
הביאה להכפלת 1.6, 1.25 -חיזוק עצמת הגירוי פי. בסדרת הניסויים הקודמתTTXהסינפטי תחת
כ מזה של תגובת הביקורת לגירוי האקסון "ולערך גבוה יותר סה, בהתאמה3.4, 2 -התגובה פי
. (fig.16C)סויים הקודמת בסדרת הני
הראשונה היא שהאינהיביציה לא מראה מגמת . סדרת ניסויים זו הניבה שתי תוצאות עיקריות
אמנם האינהיביציה חזקה יותר מאשר בסדרת . (fig.16C)התחזקות עם העלייה בשחרור הסינפטי
שם , זו הנרשמת בגירוי האקסוןאולם עדיין נמוכה משמעותית מ, )5% מול 22%(הניסויים הקודמת
תוצאה זו תומכת ). fig.12Dראה (50µM של GABAיורדת התגובה לערכים אפסיים בריכוזי
במנגנון האינהיביציה shunt -בדבר תפקידו המרכזי של ה, בממצאי סדרת הניסויים הקודמת
.הפרסינפטית בטרמינלים אלה
חסימה זו . Picrotoxin (fig.16C)י " האינהיביציה עתוצאה שנייה מסדרת ניסויים זו היא חסימת
ניתן לייחס עלייה זו לחסימת . בממוצע מעל לערך הביקורת31%מביאה לעליה הפיכה של
קרוב לוודאי עקב גירוי ישיר בו זמני , אינהיביציה הפועלת כבר על השחרור הסינפטי בתנאי ביקורת
.פירוט מופיע בדיון. ציה טוניתאו אינהיבי/של הטרמינל האינהיביטורי ו

GABA 89י "היבטים מנגנוניים של האינהיביציה המתווכת ע _________________________________________________________________________________
______________________
בטרמינלים המרכזיים GABAי "התוצאות מראות שעיקר האינהיביציה המושרית ע: סיכום
על פוטנציאל הפעולה הפולש shuntי "מתווכת ע, הממוקמים בסמיכות לאקסון המרכזי, הגדולים
י גירוי בו זמני של הטרמינל "טונית או המופעלת ע, תוכן אינהיביציה נוספ, אינהיביציה זו. לטרמינל
.Picrotoxinי "נחסמות ע, האינהיביטורי
אינהיביציה בגירוי ישיר מול גירוי אקסון בטרמינלים מרכזיים המרוחקים מהאקסון .6.2.2
פועלת באופן GABAי "בטרמינלים המרוחקים קיימות עדויות לכך שהאינהיביציה המושרית ע
& Fischer & Parnas )Fischer. י גירוי ישיר"עיל על תגובה המעוררת עשונה מזה שתואר ל
Parnas, 1996a( תיארו ירידה ברורה בשחרור הסינפטי בתגובה לריכוזים נמוכים של GABA ,
,Dudel( לאמבט לא גרמה לעליה בשחרור הסינפטי מעבר לרמת הביקורת picrotoxinוהוספה של
. בניגוד למתואר למעלה לגבי הטרמינלים הסמוכים לאקסון, )1983
דוגמא לאחד מהניסויים . ל"חזרה על ניסויים אלה בעבודה הנוכחית איששה את הממצאים הנ
ואת החסימה ) TTXבנוכחות (GABA ניתן לראות את האפקט הברור של fig.16D -מופיעה ב
לא ) GABABאנטגוניסט ספציפי לרצפטור (2OH-saclofen הוספה של. Picrotoxinי "המלאה ע
בכל חמשת הניסויים שבוצעו לפי נוהל . גררה עלייה נוספת בשחרור הסינפטי מעבר לרמת הביקורת
. זה התקבלו תוצאות זהות
ניסויים אלה מראים שאכן הפיכת ממברנת הטרמינלים המרוחקים לפסיבית לא מחסנת אותם בפני
, לא רק שהטרמינלים לא הראו חסינות לאינהיביציה, יותר מכך. GABAי "רית עאינהיביציה המוש
אלא אף התקבל הרושם שבתנאי גירוי ישיר הטרמינלים באופן פרדוקסלי הופכים רגישים יותר
. נמוכים יותרGABAכלומר מגיבים לריכוזי , לאינהיביציה בהשוואה לגירוי האקסון
נערכה סדרת , ר ועבודה זו עוסקת בעיקר בתגובה לתדרומאח, על מנת לבדוק את הרושם הזה
על התגובה לתדר המעוררת באמצעות GABAניסויים נוספת בה נבדק האפקט האינהיביטורי של
הניסויים . באותם טרמינלים, י גירוי האקסון"והושווה לזה שעל התגובה המעוררת ע, גירוי ישיר
.20-40µM של GABAוזי בריכ, בוצעו בשישה טרמינלים מרכזיים מרוחקים
. נורמל לביקורת לפי סוג הגירוי) גירוי אקסון וישיר(בכל ניסוי השחרור הסינפטי תחת אינהיביציה
בטרמינלים המרוחקים הגברת עצמת הגירוי הישיר מביאה להתפתחות מונוטונית הדרגתית של
מסיבות אלה בסדרת . )Dudel, 1983( לא מקטינה את השחרור TTXכשהוספת , השחרור הסינפטי

GABA 90י "היבטים מנגנוניים של האינהיביציה המתווכת ע _________________________________________________________________________________
והטרמינלים סווגו קודם לכן כפסיביים באמצעות עקומת , TTX-ניסויים זו לא היה צורך בשימוש ב
). משמאלfig.3 -ראה דוגמא ב(שחרור רציפה -עצמת גירוי ישיר
של (p=0.003)באופן מובהק ואכן מצביעות על רגישות מוגברת , התוצאות המתקבלות הן עקביות
בממוצע ירד . (fig.16E)הטרמינלים המרוחקים לאינהיביציה בגירוי ישיר בהשוואה לגירוי האקסון
ואילו זה , (n=6;76±12) מערך הביקורת 76% -השחרור הסינפטי המעורר בגירוי האקסון ל
ת עצמ) מסומנים בכוכביות(בשלושה מהטרמינלים . (n=6;27±9) 27% -המעורר בגירוי ישיר ירד ל
הגירוי הישיר נקבעה כך שיתקבל שחרור סינפטי בעל ערך הדומה בממוצע לזה המתקבל מגירוי
ובשלושת הטרמינלים הנותרים השחרור הסינפטי , 80Hz (98±6%;n=3:בתדר גירוי של (האקסון
ניתן לראות שגם בטרמינלים בהם . (n=3;284±88%)בגירוי ישיר היה גבוה יותר משמעותית
הרי , )כוכביות(י המעורר בגירוי האקסון ובגירוי ישיר היו בעלי ערכים כמעט זהים השחרור הסינפט
. GABA -שהשני היה רגיש יותר באופן מובהק ל
כמו כן , זההGABAל בשני סוגי הגירוי השונים האינהיביציה מושרית תוך שימוש בריכוז "בניסוי הנ
הישיר לא מגרה גם את האקסונים העדויות מצביעות על כך שבטרמינלים המרוחקים הגירוי
ולכן מקור ההבדל בהשפעתה , מכאן שהפעלת האינהיביציה בשני המקרים זהה. האינהיביטוריים
. נעוץ קרוב לודאי בהבדל שבין שתי שיטות הגירוי
על מאפייני התגובה לתדר בגירוי ישיר GABAהשפעת
הביא לירידה חדה GABAמתן , בשלושה טרמינלים מתוך ששת הטרמינלים בסדרת ניסויים זו
כך שלא ניתן היה להתאים בצורה מהימנה , )10%עד לכדי (בשחרור הסינפטי בתגובה לגירוי ישיר
בשלושת הטרמינלים הנותרים ההשפעה הייתה זהה לזו שעל . שיפוע לעקום התגובה לתדר
קצב התפתחות ולא האטה ב, בפרט לא נרשמה עליה בשיפוע עקום. התגובה לתדר בגירוי האקסון
. בתדרי הגירוי הגבוהיםהשחרור
______________________
התוצאות מאששות ממצאים קודמים בדבר רגישות טרמינלים מרכזיים המרוחקים :סיכום
הגירוי הישיר מגביר באופן פרדוכסלי , יותר מכך. מהאקסון המרכזי לאינהיביציה בתנאי גירוי ישיר
עקום התגובה לתדר בגירוי ישיר מושפע באופן . GABA -רה את רגישות טרמינלים אלה ללכאו
כלומר ללא השפעה על שיפוע העקום וללא , דומה לזה שמושפע עקום התגובה לתדר בגירוי האקסון
. קצב התפתחות השחרורהאטה ב

GABA 91י "היבטים מנגנוניים של האינהיביציה המתווכת ע _________________________________________________________________________________
אפיון פרמקולוגי של המוליכות האינהיביטורית .6.3
6.3.1. GABAA
muscimolבחלק זה מתוארות תוצאות סדרות ניסויים בהם נעשה שימוש באגוניסט
. picrotoxinובאנטגוניסט
Muscimol: תוך הזרמה של , בטרמינלים מרכזיים בסדרה זו נערכו ארבעה ניסוייםMuscimol
(10-20µM)בכל הניסויים התוצאה הייתה עקבית ודומה מאוד לזו של מתן . לאמבטGABA .
, גירויים5 (בתגובה החמישית בסדרה (mean±se;n=4) 42±8%בממוצע נגרמה ירידה של
100Hz( , שהתרחשה בקבוע זמן ממוצע(t1/2) שניות 23 מהיר יחסית של )שניות 40 -בהשוואה ל
. שניות30עם קבוע זמן של , השטיפה אף היא התרחשה במהירות). GABAעבור
Picrotoxin: האפקט של טיפול ב- Picrotoxin (50µM)טרמינלים מרכזיים20 - נבדק ב ,
. במחצית מהם התגובה עוררה תוך שימוש בנוהל גירוי האקסון ובמחצית השניה בנוהל גירוי ישיר
picrotoxinלאחר הוספה של , GABA (30µM)לאחר מתן , השחרור הסינפטי נמדד בביקורת
GABAי "בכל הניסויים נחסמה האינהיביציה המושרית ע .טיפהולבסוף לאחר ש, לתמיסת האמבט
.להלן פירוט תוצאות הניסויים בגירוי האקסון ובגירוי ישיר. באופן מלא
מתן ) סמוכים ומרוחקים מהאקסון( בגירוי האקסון של כלל הטרמינלים המרכזיים :גירוי האקסון
GABA (30µM) טי בשחרור הסינפ47% הביא לירידה ממוצעת של(fig.17A) , עם חסימה
קבוע הזמן הממוצע של . Picrotoxin (96.5±4.2;n=10) -מלאה של ההנחתה לאחר טיפול ב
). שניות בשטיפה30 -בהשוואה ל( שניות 23.3החסימה עמד על
על טרמינלים הסמוכים לאקסון GABAהשפעת , כפי שצוין לעיל, TTX עם בגירוי ישיר:גירוי ישיר
Picrotoxin -בטרמינלים הסמוכים לאקסון הטיפול ב. ה המרוחקים ממנו הייתה שונהלעומת אל
בשחרור הסינפטי ביחס לביקורת 31%הביא לחסימת האינהיביציה ולהגברה הפיכה של
(131±8;n=5) (fig.16C) . הגברה זו מבטאת כאמור חסימה של אינהיביציה טונית או זו
.רוי ישיר בו זמני של הטרמינלים האינהיביטורייםי גי"המופעלת קרוב לודאי ע

GABA 92י "היבטים מנגנוניים של האינהיביציה המתווכת ע _________________________________________________________________________________
Figure 17 : השפעותpicrotoxin ,TPMPA ,baclofenעל השחרור הסינפטי
A : חסימת האפקט שלGABA (30µM)י " על התגובה לגירוי עצב עpicrotoxin (mean±se;n=10) .י "התגובה עוררה ע
- וTPMPA (50µM)השפעת : B. בטרמינלים מרכזיים, שניותאחת לשתי , 100Hzסדרות של חמישה גירויים בתדר
picrotoxin (Ptx.)על התגובה לגירוי עצב לאחר טיפול ב - GABA (40µM)ההשפעה המוצגת . בטרמינלים מרכזיים
האפקט הטוני של : C. (mean±se;n=6)המנורמלת לערך הביקורת שלה , היא על התגובה לגירוי החמישי בסדרה
baclofen (100µM) על השחרור מטרמינלים מרכזיים (mean±se;n=7) .תיתכן ירידה בלתי מובהקת סטטיסטית של כ-
על השחרור מטרמינל מרכזי מרוחק בגירוי אקסון וגירוי baclofenדוגמא לניסוי בו נבדקה השפעת : D. בשחרור10%
.(mean±sd)יסטית איננה מובהקת סטטbaclofenהירידה בשחרור לאחר מתן . TTXעם וללא , ישיר
בגירוי ישיר היה דומה מאוד לזה Picrotoxinבטרמינלים המרוחקים מהאקסון האפקט של
- הנחית את השחרור הסינפטי לGABA (40µM)מתן . המתרחש בגירוי האקסון באותם טרמינלים
החזיר את השחרור לערכי הביקורת Picrotoxin -וטיפול ב, מערך הביקורת בממוצע23%
(93±5;n=5) .דוגמא לאחד הניסויים מובאת ב- fig.16D .בטרמינלים אלה חשיפה ל- Picrotoxin
עדות לכך שהגירוי הישיר באזור זה של , לא גרמה לעלייה בשחרור הסינפטי מעבר לרמת הביקורת
1 2 3 4 50
2
4
6
B.A. control GABA Ptx. wash
cent
ral q
uant
al c
onte
nt
stimulus No.control GABA TPMPA Ptx. wash
0.0
0.5
1.0
norm
aliz
ed re
leas
e
1 2 3 4 50
2
4
6
cent
ral q
uant
al c
onte
nt
controlbaclofen (100µM)wash
stimulus No.
0.0
2.0
4.0
6.0
8.0
10.0
D.C.
axon direct direct+TTX stimulation method
control +baclofen
cent
ral q
uant
al c
onte
nt

GABA 93י "היבטים מנגנוניים של האינהיביציה המתווכת ע _________________________________________________________________________________
,Dudel )Dudelממצא המאשש את זה של , השריר לא עורר גם את הטרמינלים האינהיביטוריים
. באותם טרמינלים)1983
י "תוצאות אלה מאששות תוצאות קודמות בדבר חסימה מלאה של האינהיביציה ע, לסיכום
Picrotoxin )Takeuchi & Takeuchi, 1969; Blundon & Bittner, 1992; Parnas et al.,
2000b( מלבד לתוצאות של Fischer & Parnas )Fischer & Parnas, 1996b, a( , שם חסימת
י טיפול בחסם ספציפי של "והושלמה ע, הייתה חלקית בלבדPicrotoxinי " עGABAההשפעה של
. GABAB (2OH-Saclofen)רצפטור מסוג
6.3.2. GABAC
אולם בעל מספר תכונות המבדילות אותו , GABAA -בדומה ל, רצפטור זה מתווך מוליכות כלור
- בהשוואה לGABA) EC50=4µM -בין תכונות אלה זיקה גבוהה יותר משמעותית ל. וןמהאחר
27µM עבור GABAA( , תווך מוליכות כלור בעלת מוליכות נמוכה יותר)7.9pSבהשוואה ל -
29.6pS עבור GABAA ( עם זמן פתיחה ארוך יותר)>100ms ( והיעדר דסנסיטיזציה משמעותית
)Feigenspan et al., 1993; Feigenspan & Bormann, 1994( . בנוסף לכך הוא נחסם ביעילות
. )Biccuculine )Johnston, 1996; Chebib & Johnston, 1999י " אולם לא עPicrotoxinי "ע
ולכן , ל מתאימות למוליכות המתווכת את האינהיביציה הפרסינפטית במערכת זו"התכונות הנ
בשנים האחרונות פותחה לרצפטור . ות את בדיקת מידת מעורבות רצפטור זה לאטרקטיביתהופכ
. עובדה המאפשרת לבחון בפועל מעורבות זו, זה פרמקולוגיה ספציפית
TPMPA: TPMPA ((1,2,5,6-tetrahydropyridine-4-yl) methylphosphinic acid) הוא
עבור רצפטור 320µM - בהשוואה לGABAC) Kb=2µMאנטגוניסט תחרותי ספציפי לרצפטור
GABAA ()Ragozzino et al., 1996( .תוך שמירה על , על מנת להבטיח יעילות של האנטגוניסט
האינהיביציה הושרתה . )50µM )Ragozzino et al., 1996נעשה שימוש בריכוז של , ספציפיות
20-30 -כTPMPA ולאחר התייצבות התגובה נבדקה השפעת , GABA (40µM)באמצעות מתן
על מנת לוודא שהאינהיביציה Picrotoxinי "בהמשך נחסמה האינהיביציה ע. דקות מרגע המתן
. שהושרתה אכן הפיכה
סדרות של חמישה ( סדרת ניסויים זו בוצעה על שישה טרמינלים מרכזיים בנוהל גירוי האקסון
לא חסם כלל את TPMPA - ניתן לראות שfig.17B -מהתבוננות ב). 100Hzגירויים בתדר

GABA 94י "היבטים מנגנוניים של האינהיביציה המתווכת ע _________________________________________________________________________________
לאמבט גרמה לחסימה Picrotoxinהוספה של . GABAי "שהושרתה ע, ההנחתה של התגובה
. מערך הביקורת83% - ל35% -מהירה של האינהיביציה ולשיקום השחרור הסינפטי מ
י רצפטור מסוג "מערכת זו לא מתווכת ע בGABAי "ניסויים אלה מראים שהאינהיביציה המושרית ע
GABAC .
6.3.3. GABAB
בדומה לתוצאות , Picrotoxinי "בניסויים שתוארו עד כה נחסמה האינהיביציה באופן מלא ע
;Takeuchi & Takeuchi, 1969; Blundon & Bittner, 1992(שפורסמו בעבודות קודמות
Parnas et al., 2000b( ,נה מעבודות קודמות של אולם בשוFischer & Parnas )Fischer &
Parnas, 1996b, a( . התוצאות הלא עקביות בנוגע לתפקידו ומידת מעורבותו של רצפטור מסוג
GABABהודות לתוצאות . ל גם בעבודה הנוכחית" במערכת זו הותירו מקום לבחון את הסוגיה הנ
שאינו ידוע כחסם של , Picrotoxinי "מלאה של האינהיביציה עשהוצגו עד כה ותארו חסימה ה
ל במערכת "ההערכה הייתה שמידת מעורבות הרצפטור הנ ,GABABי רצפטור "מוליכות המתווכת ע
. או לחילופין באה לידי ביטוי בתנאי גירוי ורישום אחרים, זו מועטה
Baclofen: צפטור המתווך את במטרה להשלים את בירור הפרופיל הפרמקולוגי של הר
בריכוז Baclofenהאינהיביציה הפרסינפטית נערכו שתי סדרות ניסויים בהם נבדקה ההשפעה של
100µMב. על השחרור הסינפטי מטרמינלים מרכזיים-fig.17C מתוארת ההשפעה הממוצעת של
Baclofenי האקסון שעורר בתגובה לגירו, על השחרור האקסיטטורי משבעה טרמינלים מרכזיים
השחרור הסינפטי נידגם כל Baclofenמרגע המתן של . 100Hzבסדרות של חמישה גירויים בתדר
התוצאות . מספר דקות והושווה לביקורת על מנת להבטיח זיהוי של השפעה בעלת אופי חולף
בלתי ניתן לראות ירידה fig.17C-ב. דקות מרגע המתן40-60המוצגות נרשמו לאחר פרק זמן של
נעשתה בכל שבעת הטרמינלים , לאחר השטיפה). בממוצע10%(מובהקת בשחרור הסינפטי
, לאמבט שגרמה בכל המקרים לירידה משמעותית בשחרור הסינפטיGABAהזרמה קצרה של
בחתך ונמצא פעיל הוא נבדק Baclofen -כמו כן מתוך חשש לתקינות ה. fig.17A -בדומה למוצג ב
מתאים בודדים patch-clamp - תוך רישום ב,)מסלולים תוך קורטיקליים( בעכבריקליקורט-טלמו
המבוסס על משמעותי אינהיביטורי מרכיבנוכחותלא הדגימו ניסויים אלה . )Gil et al., 1997ראה (
. בתנאי ניסוי אלה GABABרצפטור

GABA 95י "היבטים מנגנוניים של האינהיביציה המתווכת ע _________________________________________________________________________________
Baclofen -בשלושה ניסויים נוספים נבדקה רגישות טרמינלים מרכזיים המרוחקים מהאקסון ל
. fig.17D -תוצאות אחד מהניסויים מוצגות ב. TTXללא ועם , בתנאי גירוי אקסון וגירוי ישיר
והדגימו שוב ירידה קלה אך בלתי מובהקת סטטיסטית בשחרור הסינפטי , קבייםהממצאים היו ע
עצמת הגירוי הישיר נקבעה כך שהוא עורר תגובה חזקה יותר מזה שעורר . בתנאי הגירוי הישיר
. בצורה האופיינית לטרמינלים אלה, TTXללא השפעה ניכרת של , גירוי האקסון
אולם , במערכת זוGABABמעורבות של רצפטור מסוג אין בכוחן של התוצאות האחרונות לשלול
. על תהליך השחרור הסינפטיניתן לומר בביטחון שבתנאים אלה לא הודגמה השפעה מהותית שלו
______________________
בתווך GABAA -התוצאות מאששות מעורבות ברורה של רצפטור הדומה פרמקולוגית ל: סיכום
חסם באופן מלא את האינהיביציה כפי Picrotoxinשימוש באנטגוניסט . יציה הפרסינפטיתהאינהיב
Takeuchi & Takeuchi, 1969; Blundon & Bittner, 1992; Fischer(שתואר בעבודות קודמות
& Parnas, 1996b, a; Parnas et al., 2000b( ,]אך ראה :Fischer & Parnas )Fischer &
Parnas, 1996b, a( .[ שימוש בחסם ספציפי לרצפטורGABAC , ובאגוניסט לרצפטורGABAB לא
. בתנאי רישום אלההביאו לתוצאות המצביעות על מעורבות רצפטורים אלה במערכת זו
דיון .6.4. אקסוןהשימוש בגירוי ישיר מאפשר לזהות שתי תופעות עיקריות שאין להן ביטוי ברור בגירוי ה
הטרמינלים הקרובים לאקסון המרכזי TTXראשית ניתן לראות באופן ברור שבתנאי גירוי ישיר עם
למרות שבשני המקרים , בניגוד לאלה המרוחקים ממנו, GABAכמעט לא מושפעים ממתן
תוצאה מעניינת נוספת היא . Picrotoxinנחסמת על ידי הי מוליכות "האינהיביציה מתווכת ע
בתנאי הגירוי הישיר בהשוואה GABA - המוגברת של הטרמינלים המרוחקים מהאקסון להרגישות
. לגירוי האקסון
ל מופיע בנפרד עבור כל אחת משתי אוכלוסיות הטרמינלים עקב ההבדלים "הדיון בתופעות הנ
.המורפולוגיים והפיזיולוגיים ביניהם

GABA 96י "היבטים מנגנוניים של האינהיביציה המתווכת ע _________________________________________________________________________________
האינהיביציה בטרמינלים מרכזיים הסמוכים לאקסון .6.4.1
ולקרבתם , התנגדות הכניסה של טרמינלים אלה נמוכה באופן יחסי הודות לקוטרם הגדול יחסית
מכאן שכדי . )Cooper et al., 1996a; Wright et al., 1996b(המורפולוגית והחשמלית לאקסון
פוטנציאל הפעולה . להשפיע על מתח הממברנה שלהם יש צורך בשטפים חשמליים חזקים יחסית
מספק את השטף החשמלי האקסיטטורי , או לנקודה הסמוכה מאד חשמלית אליהם, הפולש אליהם
תהיה ) כלור(אינהיביציה המסתמכת על שטף בעל כוח מניע חלש במתח המנוחה . הדרוש לשם כך
תיאור זה עשוי להסביר את הצורך . אפקטיבית רק במידה ותתאפיין במוליכות גבוהה יחסית
ובצפיפות רבה יותר של טרמינלים אינהיביטוריים בסביבת GABAריכוזים גבוהים יחסית של ב
יש להניח שהודות לצפיפות זו גירוי ישיר של הטרמינלים . טרמינלים גדולים הסמוכים לאקסון
. (fig.16C) המרכזיים הסמוכים לאקסון שיפעל גם את הטרמינלים האינהיביטוריים הסמוכים להם
טרמינלים אלה תפקידה העיקרי של האינהיביציה הוא להרחיק את פוטנציאל הפעולה יתכן וב
באופן זה פתיחת . המחבר את הטרמינל לאקסון" צוואר"כך שיעבור באופן פסיבי דרך ה, מהטרמינל
. )Segev, 1990(המוליכות תנחית ביעילות את משרעת גל המתח הפולש לטרמינל
מעורר פוטנציאל פעולה באקסון ) בעוצמה העוברת סף מסוים (TTXל הטרמינל ללא גירוי ישיר ש
ומעורר שחרור סינפטי בערך הדומה לזה המתקבל מגירוי האקסון , הפולש לטרמינלהפרטרמינלי
)Dudel, 1983( . במקרה זה מתןGABAבדומה , ינחית את השחרור הסינפטי בצורה אפקטיבית
. בגירוי האקסוןלמתרחש
הופך את ממברנת הטרמינל לפסיבית כך שגירוי ישיר לא יעורר TTX - בניגוד לאמור לעיל טיפול ב
העוקף , במקרה זה הגירוי הישיר הוא הגורם המעורר היחידי. פוטנציאל פעולה באקסון הפרטרמינלי
יה להיות מושפעת בצורה ולכן התגובה לגירוי זה לא צפו, הטרמינל" צוואר"מטבעו את ההולכה דרך
.GABAמשמעותית ממתן
ארגון זה של האינהיביציה עשוי להסביר מדוע היא מאבדת מיעילותה בטרמינלים אלה בתנאי גירוי
ל ביעילות האינהיביציה על כך שפרט למנגנון "בנוסף לכך מעידה הירידה החדה הנ. TTXישיר עם
למשל מנגנון הפועל , נון אינהיביטורי אפקטיבי אחר אין בטרמינלים אלה ביטוי לפעולת מנגshunt -ה
. ישירות על מוליכות הסידן בטרמינל או על מנגנון השחרור הסינפטי עצמו
אין בו התייחסות מפורשת לטרמינלים , מאחר והפרק הנוכחי מתמקד בטרמינלים המרכזיים
ופעילות , הסינפטילאור הדמיון של חלק ניכר ממאפייני השחרור, למרות זאת. הפרוקסימליים

GABA 97י "היבטים מנגנוניים של האינהיביציה המתווכת ע _________________________________________________________________________________
ניתן לשער , האינהיביציה בין הטרמינלים המרכזיים הסמוכים לאקסון לטרמינלים הפרוקסימליים
.שמנגנוני האינהיביציה בטרמינלים האחרונים דומים לאלו שנדונו לעיל
האינהיביציה בטרמינלים מרכזיים המרוחקים מהאקסון .6.4.2
ומתאפיינים במבנה varicosities -ם אלה בנויים כטרמינלי, בניגוד לאוכלוסייה הקודמת
ובפלישה פסיבית של פוטנציאל הפעולה , במיעוט סינפסות ואתרי השחרור, קומפקטי יותר
)Atwood, 1967; Bittner, 1968; Atwood & Bittner, 1971; Sherman & Atwood, 1972;
Dudel, 1982(ולכן ניתן להניח שהם , רוך מורפולוגית וחשמלית לטרמינלים אלה צוואר דק וא
התנגדות הכניסה הגבוהה יותר וקיבול הממברנה הנמוך . מבודדים יותר חשמלית מהאקסון המרכזי
, מאפשרים לשטף חשמלי נמוך יחסית לעורר בטרמינלים אלה שינויי מתח משמעותיים יותר
. בהשוואה לטרמינלים הגדולים
נותרת GABAי "לים אלה בתנאי גירוי ישיר האינהיביציה המושרית ענשאלת השאלה מדוע בטרמינ
. ואף יעילה יותר בהשוואה לזו המושרית בגירוי האקסון, יעילה
שלא דרך מפל (תשובה אפשרית אחת היא להניח שבטרמינלים אלה קיים מנגנון המקשר ישירות
או אף , מוליכות תעלות הסידן לGABAAבין מוליכות אינהיביטורית מתווכת ) המתחים על הממברנה
עדות ישירה למנגנון זה עשויה להתקבל מניסויי . )Parnas et al., 2000b(למנגנון השחרור עצמו
voltage clampתוך רישום והשוואה של זרמי הסידן והשחרור הסינפטי בביקורת , של הטרמינל
. )Wright et al. )Wright et al., 1996a, b בדומה לעבודות שנעשו על ידי, ותחת אינהיביציה
אולם בהחלט , תוצאות המחקר הנוכחי אמנם לא מאפשרות להוכיח ישירות קיום של מנגנון כזה
שכן אחרת קשה להסביר חלק מהמאפיינים הבולטים של האינהיביציה בטרמינלים , תומכות בקיומו
. אלה
ון אינהיביטורי ישיר אינו מסביר מדוע האינהיביציה בתנאי גירוי ישיר קיום מנגנ, למרות האמור לעיל
ובמיוחד כאשר השחרור הסינפטי בתגובה לשני סוגי , יעילה יותר משמעותית מאשר בגירוי אקסון
מסיבה זו יש מקום לחפש את הסיבה ליעילות ). משושים עם כוכביות, fig.16E(הגירוי זהה
אלא בגורם הקשור להבדל בין ,יביציה לא במנגנון האינהיביטורי עצמול של האינה"המוגברת הנ
במהלך הגירוי הישיר נוצר מפל מתחים בין תוך :השפעת שני סוגי הגירוי השונים על הטרמינל
כך שהמתח בתוך הפפיטה מוסט לערכים שליליים ומתקרב למתח התוך , הפיפטה לתווך החוץ תאי
הרי ששטף יוני כלור , ) באמבטGABAנוכחות (כות הכלור פתוחה במידה ומולי. תאי של הטרמינל

GABA 98י "היבטים מנגנוניים של האינהיביציה המתווכת ע _________________________________________________________________________________
יחדור מהפיפטה לתוך הטרמינל בעקבות הכוח האלקטרו המניע המשמעותי שיוצר פולס הגירוי
קרוב לוודאי ששטף זה זניח בהשוואה לשטף העובר דרך כלל ההתנגדות הנוצרת בין . השלילי
ת הכניסה הגבוהה וקיבול הממברנה הנמוך של אולם בהתחשב בהתנגדו, (RS)הפיפטה לממברנה
ובאזור , הטרמינלים המרוחקים יתכן שדי בשטף זה על מנת להוריד את המתח התוך תאי בטרמינל
במצב זה משך הדפולריזציה שיוצר הגירוי הישיר יתקצר בעקבות השטף . אתרי השחרור בפרט
הראה )Dudel )Dudel, 1983.ציהובסיום הגירוי אף צפוי להופיע שלב של היפרפולריז, השלילי
ששינויים במשך הגירוי הישיר הניתן לטרמינל עשויים להגדיל או להקטין את השחרור הסינפטי
פסיליטציה "קיצור משך הגירוי עשוי לבטל או להקטין תופעה המכונה . מהטרמינל במספר סדרי גודל
תופעה הגורמת להגברה משמעותית בשחרור הסינפטי )Katz & Miledi, 1965, 1971(" מוקדמת
עשויה picrotoxin -רגישה לתיאור זה מסביר כיצד מוליכות כלור .ומתרחשת במהלך הגירוי עצמו
על . ה בהנחתת השחרור הסינפטי מטרמינלים קומפקטיים גם בתנאי גירוי ישירלשמור על יעילות
לאחר בדיקת ,יוחלפו יוני הכלור בפיפטת הרישום בו בגירוי ישירניסוי לבצע ישסברה זו מנת לאשש
.ת בביקורGABAהאפקט האינהיביטורי של
גורמת לירידה ) גירוי אקסון (פתיחת מוליכות הכלור תוך גירוי האקסון האקסיטטורי, ל"בשונה מהנ
ירידה . )Blundon & Bittner, 1992; Golan, 1994(במשרעת פוטנציאל הפעולה הפולש לטרמינל
זה במהלך גירוי האקסון עשויה להפחית את השחרור הסינפטי בצורה פחות אפקטיבית בהשוואה
תופעות , עוקבתהופעת היפרפולריזציהווה בהעשויה אף להיות מללקיצור משך הדפולריזציה
, )APהנחתת (הבדל זה בין הפחתת משרעת הדפולריזציה. להתרחש בתנאי גירוי ישירותהעשוי
עשוי להסביר את הרגישות המוגברת של והופעה אפשרית של היפרפולריזציה לקיצור המשך שלה
. השחרור הסינפטי לאינהיביציה בתנאי הגירוי הישיר
רצפטור המתווך את האינהיביציהאפיון ה .6.4.3
בתווך GABAAניסויי הפרמקולוגיה מצביעים על מעורבות דומיננטית של רצפטור מסוג
רצפטור זה שונה מהרצפטור הקלאסי הודות למספר תכונות האופייניות יותר . האינהיביציה
-ניסויי ה. לית ודסנסיטיזציה מינימGABA -ביניהן רגישות גבוהה יחסית ל, GABACלרצפטור מסוג
GABA הצביעו על רגישות של EC50=29µM (fig.12D) , רגישות הנמצאת בטווח העליון של
מדדו )GABAA .Fischer & Parnas )Fischer & Parnas, 1996aהרגישות האופיינית לרצפטור
, GABAכוזים עולים של את העלייה במוליכות האקסון בסמוך לאותם טרמינלים בתגובה למתן רי

GABA 99י "היבטים מנגנוניים של האינהיביציה המתווכת ע _________________________________________________________________________________
ל מראה שהשפעת "הקרבה בין התוצאות הנ. EC50=20µM: וקיבלו תוצאה מעט נמוכה יותר
ומעידה על , הרצפטור על השחרור הסינפטי מפגרת אך במעט אחר העלייה במוליכות הממברנה
יטב רגישות זו מתיישבת ה. רגישות מנגנון השחרור בטרמינלים אלה לשינויים במוליכות הממברנה
הצופה שעליה במוליכות ממברנת הטרמינל , )Segev )Segev, 1990עם המודל החשמלי של
תגרור אינהיביציה יעילה בטרמינלים בעלי נתונים מורפולוגיים ופיזיולוגיים האופייניים לטרמינלים
. אלה
השיטות בהן , ממברנהי השפעה על מתח ה"באשר לפעילות נוספת של הרצפטור שאיננה מתווכת ע
נעשה שימוש בעבודה זו הניבו תוצאות התומכות בקיומה אולם לא מאפשרות להצביע ישירות על
. המנגנון המעורב בה
בתווך האינהיביציה בטרמינלים המרכזיים GABABלמרות עדויות ברורות למעורבות רצפטור מסוג
)Barry, 1984; Fischer & Parnas, 1996b, a( ,כמו גם , לא נמצא לכך חיזוק בעבודה הנוכחית
,Picrotoxin )Takeuchi & Takeuchiי "בעבודות אחרות בהן נחסמה האינהיביציה באופן מלא ע
1969; Blundon & Bittner, 1992; Parnas et al., 2000b( .ל "הסיבה לחוסר האחידות הנ
לאור העדויות המורפולוגיות . לא זכתה להתייחסות בעבודות אחרותואף, בתוצאות לא לגמרי ברורה
והפיזיולוגיות לחוסר ההומוגניות של השריית אינהיביציה פרסינפטית בטרמינלים המרכזיים בכלל
)Govind et al., 1995; Fischer & Parnas, 1996a( ,י " שהאינהיביציה המתווכת עיתכן בהחלט
. בפרט איננה מושרית באופן הומוגני על כלל אוכלוסיית הטרמינלים המרכזייםGABABרצפטור
תוך , ובחירה סובייקטיבית של טרמינלים בודדים מתוך אוכלוסייה המתאפיינת בשונות גבוהה, זאת
ל בין התוצאות של "שימוש בנהלי גירוי שונים עשויים להסביר חלק ניכר מחוסר ההתאמה הנ
. שונותהעבודות

GABA 100י "היבטים מנגנוניים של האינהיביציה המתווכת ע _________________________________________________________________________________
Table 1 :מתן , גירוי ישיר, השפעות הפחתת ריכוז סידן חוץ תאיGABA וגירוי האינהיביטור על מאפייני התגובה לתדר באוכלוסיות טרמינלים שונות
ים של השפעות הטיפולים השונים על המאפיינים העיקרי) אין השפעה, ירידה, עליה(בטבלא זו מצוינים באופן איכותי
.מציין היעדר השפעה משמעותית ≈, תכולה קואנטלית-m. עקום התגובה לתדר בטרמינלים מרכזיים ופרוקסימליים
סוג
:גירוי/הטיפול סוג הטרמינל :נבדק מאפיין
גירוי ישיר """" out[+Ca2] ביקורת גירוי GABAמתן "חזק"
האינהיביטור
- 0.076 מרכזי0.61 " $ "
שחרור בודד (m) " " " 1.16 פרוקסימלי
6.6 מרכזי(80Hz)
רמות השחרור " $ " הסינפטי ברוויה
(m) 16.5 פרוקסימלי (80Hz)
≈ " "
שיפוע עקום אין השפעה " $ 1.6 מרכזי התגובה לתדר
אין השפעה אין השפעה 1.05 פרוקסימלי )מעריך(
פוטנציאציה ≈ ≈ $ 80 מרכזי מקסימלית
מנורמל לתגובה ( אין השפעה " $ 14 פרוקסימלי )1Hz -ב
קצב התפתחות אין השפעה $ 9 מרכזי השחרור(St.n½)
אין השפעה " (9) 4-5 פרוקסימלי

101
וחשיבותםעיקרי הממצאים .7___________________________________________________________________
-עבודה זו מתארת לראשונה את השחרור הסינפטי מהטרמינלים האקסיטטורים השונים של שריר ה
openerתוצאות אלה מציגות למעשה באופן מלא . על פני ספקטרום פיזיולוגי רחב של תדרי גירוי
בניגוד לעבודות קודמות , את ספקטרום ההגברה הפרסינפטית קצרת הטווח המתרחשת בפרפרט
הודות . שנבחרו באופן סובייקטיבי, ו תופעות נקודתיות שהתבססו על נהלי גירוי מסוימיםבהן הוצג
לתוצאות אלה מתקבלת תמונה מלאה ומקיפה יותר של התכונות הפרסינפטיות המאפיינות
. 1' הממצאים העיקריים מסוכמים באופן איכותי בטבלא מס. טרמינלים שונים
המתארת (power)מרכזי בהקשר זה היא פונקצית החזקה הממצא ה:שחרורהתלות תדר גירוי
כשהחזקה בה מועלה תדר , את התלות בין תדר הגירוי לרמת הרוויה אליה מגיע השחרור הסינפטי
היא בעלת ערך ) עבור הטרמינלים המרכזיים1.68 - עבור הטרמינלים הפרוקסימליים ו1.05(הגירוי
ממצא . (Ravin et al., 1999) באותם טרמינליםהממוצע תאי התוךזהה לזו בה מועלה ריכוז הסידן
יסוד פיזיולוגי להשערה שהגירוי בתדר משמש כלי ליניארי להעלאת ריכוז הסידן התוך תאי זה מניח
את עשויים לשקףהשינוי שחל בתלות זו בעקבות שינויים בכניסת הסידן לטרמינל . בטרמינלים
בעקבותזו של כניסת הסידן של ריכו הסידן הממוצע בטרמינל מול לשחרור התרומה היחסית
. גירויה
על תרומתם גם המלמדים , בין עקומי התגובה לתדר של טרמינלים שונים קיימים הבדלים מהותיים
.כתלות בתדר הירי של המוטורנוירון האקסיטטורי המעצבב את השריר, השונה לשחרור הסינפטי
בעבודה זו נעשית לראשונה הבדלה ברורה בין : בתוך סדרת הגירוייםקצב התפתחות השחרור
פוטנציאציה ל) מספר הגירויים הדרוש להגעת התגובה לרוויה בתדר נתון (קצב התפתחות השחרור
ה בין תאוצה של אובייקט הבדל זה דומה להפרד). מידת הכפלת התגובה בתוך סדרת הגירויים(
הבדלה זו מאפשרת ). שאינו תלוי בהכרח בגודל התאוצה(למשך הזמן שתאוצה זו נמשכת , נתון
לרוב קיים מתאם הפוך בין שני מאפיינים . לקבל תמונה מקיפה ומלאה יותר של התהליך בכללותו
, ינית לטרמינלים הפרוקסימלייםהאופי, )הכפלה איטית של השחרור הסינפטי (חלשה קינטיקה: אלה
ומתאפיינת בערכי פוטנציאציה נמוכים יותר , דורשת מספר גירויים נמוך יותר להגעה לרוויה

102 וחשיבותםעיקרי הממצאים _________________________________________________________________________________
מצריכה גירויים רבים יותר עד להגעה , האופיינית לטרמינלים המרכזיים, קינטיקה חזקה. אבסולוטית
שחרורהניראה שעקום התפתחות . בוהים יותר אבסולוטיתומתאפיינת בערכי פוטנציאציה ג, לרוויה
כלומר השחרור הסינפטי מתחיל , שמאלה ביחס למרכזיים" מוסט"בטרמינלים הפרוקסימליים
בצעדים גדולים יותר ומגיע מוקדם " מתקדם", גבוהה יותר משמעותית" נקודה"בטרמינלים אלה מ
. יותר לרוויה
קצב התפתחות השחרורל, שחרור-נה קשר הפוך בין שיפוע העקום תדרכמו כן מאופיין לראשו
שחרור גבוה וקצב -לטרמינלים המרכזיים לדוגמא שיפוע עקום תדר: בתדרי הגירוי הגבוהים
של טרמינלים אלה מאיץ את התפתחות " חזק"גירוי ישיר . התפתחות פוטנציאציה נמוך
הפעלת אינהיביציה על הטרמינלים הפרוקסימליים , מנגד. שיפוע העקוםהפוטנציאציה ומקטין את
קצב התפתחות ממצאים אלה מעידים על כך ש. מאיטה את קצב ההתפתחות הפוטנציאציה
רגיש באופן יחסי לשינויים בכניסת ) הפוטנציאציהמובשונה (, בדומה לשחרור הסינפטי, השחרור
. הסידן לטרמינל
לא משפיע על קינטיקת התפתחות רמות GABA באופן מפתיע מתן : אינהיביציה פרסינפטית
, ולא על התפתחות עצמת הפוטנציאציה, )שחרור-שיפוע עקום תדר(הרוויה של השחרור הסינפטי
. פטיובטרמינלים המרכזיים אף לא על מספר הגירויים הדרוש על מנת להגיע לרוויה בשחרור הסינ
י "ומכאן שאפשרי והאינהיביציה מתווכת ע, הפחתה בכניסת הסידן לטרמינל לא מדמה ממצאים אלה
Parnas(כפי שהציעו חוקרים אחדים , הפועל ישירות על מנגנון השחרור, מנגנון אינהיביטורי שונה
et al., 2000b( .
שאינו עובר , GABA -רגישות גבוהה יחסית ל בעל GABAAי רצפטור "האינהיביציה ככלל מתווכת ע
, shuntי "בטרמינלים המרכזיים הסמוכים לאקסון עיקר האינהיביציה מתווכת ע. כמעט דסנסיטיזציה
. ואילו המנגנון האינהיביטורי הדומיננטי בטרמינלים המרכזיים המרוחקים מהאקסון עשוי להיות שונה

103
References
ARAQUE, A. & BUNO, W. (1995). Fast, persistent, Ca(2+)-dependent K+ current controls graded electrical activity in crayfish muscle. Pflugers Arch 430, 541-551.
ARAQUE, A., MARCHAND, A. & BUNO, W. (1998). Voltage-gated and Ca2+-activated conductances
mediating and controlling graded electrical activity in crayfish muscle. J Neurophysiol 79, 2338-2344.
ATWOOD, H. L. (1967). Variation in physiological properties of crustacean motor synapses. Nature 215,
57-58. ATWOOD, H. L. (1976). Organization and synaptic physiology of crustacean neuromuscular systems.
Prog Neurobiol 7, 291-391. ATWOOD, H. L. (1982). Synapses and neurotransmitters. In The biology of Crustacea, vol. 3. ed. C, A.
H. L. S. D., pp. 105-150. ATWOOD, H. L. & BITTNER, G. D. (1971). Matching of excitatory and inhibitory inputs to crustacean
muscle fibers. J Neurophysiol 34, 157-170. ATWOOD, H. L. & KWAN, I. (1976). Synaptic development in the crayfish opener muscle. J Neurobiol 7,
289-312. ATWOOD, H. L. & MORIN, W. A. (1970). Neuromuscular and axoaxonal synapses of the crayish opener
muscle. J Ultrastruct Res 32, 351-369. ATWOOD, H. L., SMYTH, T., JR. & JOHNSTON, H. S. (1969). Neuromuscular synapses in the cockroach
extensor tibiae muscle. J Insect Physiol 15, 529-535. ATWOOD, H. L., STEVENS, J. K. & MARIN, L. (1984). Axoaxonal synapse location and consequences for
presynaptic inhibition in crustacean motor axon terminals. J Comp Neurol 225, 64-74. ATWOOD, H. L. & TSE, F. W. (1988). Changes in binomial parameters of quantal release at crustacean
motor axon terminals during presynaptic inhibition. J Physiol (Lond) 402, 177-193. ATWOOD, H. L. & WOJTOWICZ, J. M. (1986). Short-term and long-term plasticity and physiological
differentiation of crustacean motor synapses. Int Rev Neurobiol 28, 275-362. ATWOOD, H. L. & WOJTOWICZ, J. M. (1999). Silent synapses in neural plasticity: current evidence. Learn
Mem 6, 542-571. AUGUSTINE ,G. J. (2001). How does calcium trigger neurotransmitter release ?Curr Opin Neurobiol 11,
320-326. BALNAVE, R. J. & GAGE, P. W. (1974). On facilitation of transmitter release at the toad neuromuscular
junction. J Physiol (Lond) 239, 657-675. BARRY, S. R. (1984). Baclofen has a presynaptic action at the crayfish neuromuscular junction. Brain
Res 311, 152-156. BERTRAM, R., SHERMAN, A. & STANLEY, E. F. (1996). Single-domain/bound calcium hypothesis of
transmitter release and facilitation. J Neurophysiol 75, 1919-1931. BITTNER, G. D. (1968). Differentiation of nerve terminals in the crayfish opener muscle and its functional
significance. J Gen Physiol 51, 731-758. BITTNER, G. D. & HARRISON, J. (1970). A reconsideration of the Poisson hypothesis for transmitter
release at the crayfish neuromuscular junction. J Physiol 206, 1-23.

BITTNER, G. D. & SCHATZ, R. A. (1981). An examination of the residual calcium theory for facilitation of transmitter release. Brain Res 210, 431-436.
BLUNDON, J. A. & BITTNER, G. D. (1992). Effects of ethanol and other drugs on excitatory and inhibitory
neurotransmission in the crayfish. J Neurophysiol 67, 576-587. BLUNDON, J. A., WRIGHT, S. N., BRODWICK, M. S. & BITTNER, G. D. (1993). Residual free calcium is not
responsible for facilitation of neurotransmitter release. Proc Natl Acad Sci U S A 90, 9388-9392. BOLLMANN, J. H., SAKMANN, B. & BORST, J. G. (2000). Calcium sensitivity of glutamate release in a calyx-
type terminal. Science 289, 953-957. CHEBIB, M. & JOHNSTON, G. A. (1999). The 'ABC' of GABA receptors: a brief review. Clin Exp Pharmacol
Physiol 26, 937-940. COOPER, R. L., HARRINGTON, C. C., MARIN, L. & ATWOOD, H. L. (1996a). Quantal release at visualized
terminals of a crayfish motor axon: intraterminal and regional differences. J Comp Neurol 375, 583-600.
COOPER, R. L., MARIN, L. & ATWOOD, H. L. (1995a). Synaptic differentiation of a single motor neuron:
conjoint definition of transmitter release, presynaptic calcium signals, and ultrastructure. J Neurosci 15, 4209-4222.
COOPER, R. L., STEWART, B. A., WOJTOWICZ, J. M., WANG, S. & ATWOOD, H. L. (1995b). Quantal
measurement and analysis methods compared for crayfish and Drosophila neuromuscular junctions, and rat hippocampus. J Neurosci Methods 61, 67-78.
COOPER, R. L., WINSLOW, J. L., GOVIND, C. K. & ATWOOD, H. L. (1996b). Synaptic structural complexity
as a factor enhancing probability of calcium-mediated transmitter release. J Neurophysiol 75, 2451-2466.
CRIDER, M. E. & COOPER, R. L. (1999). Importance of stimulation paradigm in determining facilitation
and effects of neuromodulation. Brain Res 842, 324-331. CRIDER, M. E. & COOPER, R. L. (2000). Differential facilitation of high- and low-output nerve terminals
from a single motoneuron. J Appl Physiol 88, 987-996. DATYNER, N. B. & GAGE, P. W. (1980). Phasic secretion of acetylcholine at a mammalian neuromuscular
junction. J Physiol (Lond) 303, 299-314. DAVIS, G. W. & MURPHEY, R. K. (1994). Retrograde signaling and the development of transmitter
release properties in the invertebrate nervous system. J Neurobiol 25, 740-756. DEL CASTILO, J. & KATZ, B. (1954). J. Physiol. 124, 560. DELANEY, K. R. & TANK, D. W. (1991). Calcium-dependent and calcium-independent enhancement of
transmitter release at the crayfish neuromuscular junction studied with fura-2 imaging. Ann N Y Acad Sci 635, 452-454.
DELANEY, K. R. & TANK, D. W. (1994). A quantitative measurement of the dependence of short-term
synaptic enhancement on presynaptic residual calcium. J Neurosci 14, 5885-5902. DELANEY, K. R., ZUCKER, R. S. & TANK, D. W. (1989). Calcium in motor nerve terminals associated with
posttetanic potentiation. J Neurosci 9, 3558-3567. DODGE, F. A., JR. & RAHAMIMOFF, R. (1967). On the relationship between calcium concentration and the
amplitude of the end-plate potential. J Physiol (Lond) 189, 90P-92P. DUDEL, J. (1981). The effect of reduced calcium on quantal unit current and release at the crayfish
neuromuscular junction. Pflugers Arch 391, 35-40. DUDEL, J. (1982). Transmitter release by graded local depolarization of presynaptic nerve terminals at
the crayfish neuromuscular junction. Neurosci Lett 32, 181-186.

DUDEL, J. (1983). Graded or all-or-nothing release of transmitter quanta by local depolarizations of nerve terminals on crayfish muscle? Pflugers Arch 398, 155-164.
DUDEL, J. & KUFFLER, S. W. (1961). Presynaptic inhibition at the cryfish neuromuscular junction. J.
Physiol. 155, 543-562. DUDEL, J., PARNAS, I. & PARNAS, H. (1982). Neurotransmitter release and its facilitation in crayfish. III.
Amplitude of facilitation and inhibition of entry of calcium into the terminal by magnesium. Pflugers Arch 393, 237-242.
DUDEL, J., PARNAS, I. & PARNAS, H. (1983). Neurotransmitter release and its facilitation in crayfish
muscle. VI. Release determined by both, intracellular calcium concentration and depolarization of the nerve terminal. Pflugers Arch 399, 1-10.
FATT & KATZ, B. (1952). FEIGENSPAN, A. & BORMANN, J. (1994). Differential pharmacology of GABAA and GABAC receptors on
rat retinal bipolar cells. Eur J Pharmacol 288, 97-104. FEIGENSPAN, A., WASSLE, H. & BORMANN, J. (1993). Pharmacology of GABA receptor Cl- channels in rat
retinal bipolar cells. Nature 361, 159-162. FELMY, F., NEHER, E. & SCHNEGGENBURGER, R. (2003). Probing the intracellular calcium sensitivity of
transmitter release during synaptic facilitation. Neuron 37, 801-811. FELMY ,F., NEHER ,E. & SCHNEGGENBURGER ,R. (2003). The timing of phasic transmitter release is
Ca2+-dependent and lacks a direct influence of presynaptic membrane potential .PNAS 100, 15200-15205.
FENGLER, B. T. & LNENICKA, G. A. (2002). Activity-dependent plasticity of calcium clearance from
crayfish motor axons. J Neurophysiol 87, 1625-1628. FISCHER, Y. & PARNAS, I. (1996a). Activation of GABAB receptors at individual release boutons of the
crayfish opener neuromuscular junction produces presynaptic inhibition. J Neurophysiol 75, 1377-1385.
FISCHER, Y. & PARNAS, I. (1996b). Differential activation of two distinct mechanisms for presynaptic
inhibition by a single inhibitory axon. J Neurophysiol 76, 3807-3816. FISHER, S. A., FISCHER, T. M. & CAREW, T. J. (1997). Multiple overlapping processes underlying short-
term synaptic enhancement. Trends Neurosci 20, 170-177. FLOREY, E. & CAHILL, M. A. (1982). The innervation pattern of crustacean skeletal muscle. Morphometry
and ultrastructure of terminals and synapses. Cell Tissue Res 224, 527-541. GOLAN, H. (1994). In Physiology. Ben Gurion University. GOLAN, H. & GROSSMAN, Y. (1996). Block of glutamate decarboxylase decreases GABAergic inhibition
at the crayfish synapses: possible role of presynaptic metabotropic mechanisms. J Neurophysiol 75, 2089-2098.
GOLAN, H., MOORE, H. J. & GROSSMAN, Y. (1994). Quantal analysis of presynaptic inhibition, low
[Ca2+]0, and high pressure interactions at crustacean excitatory synapses. Synapse 18, 328-336. GOVIND, C. K. (1982). Synapses and neurotransmitters. In The biology of Crustacea, vol. 3. ed. C, A. H.
L. S. D., pp. 63-103. GOVIND, C. K., ATWOOD, H. L. & PEARCE, J. (1995). Inhibitory axoaxonal and neuromuscular synapses in
the crayfish opener muscle: membrane definition and ultrastructure. J Comp Neurol 351, 476-488. GROSSMAN, Y., PARNAS, I. & SPIRA, M. E. (1979). Differential conduction block in branches of a
bifurcating axon. J Physiol 295, 283-305.

GUNZEL, D. & GALLER, S. (2000). Intracellular pH of crab and crayfish muscle fibre types during GABA application and inhibitory nerve stimulation. J Comp Physiol [B] 170, 523-529.
HOCHNER, B., PARNAS, H. & PARNAS, I. (1991). Effects of intra-axonal injection of Ca2+ buffers on
evoked release and on facilitation in the crayfish neuromuscular junction. Neurosci Lett 125, 215-218.
JAHROMI, S. S. & ATWOOD, H. L. (1974). Three-dimensional ultrastructure of the crayfish neuromuscular
apparatus. J Cell Biol 63, 599-613. JOHNSON, E. W. & WERNIG, A. (1971). The binomial nature of transmitter release at the crayfish
neuromuscular junction. J Physiol 218, 757-767. JOHNSTON, G. A. (1996). GABAc receptors: relatively simple transmitter -gated ion channels? Trends
Pharmacol Sci 17, 319-323. KAMIYA, H. & ZUCKER, R. S. (1994). Residual Ca2+ and short-term synaptic plasticity. Nature 371, 603-
606. KATZ, B. & MILEDI, R. (1965). Release of acetylcholine from a nerve terminal by electric pulses of
variable strength and duration. Nature 207, 1097-1098. KATZ, B. & MILEDI, R. (1968). The role of calcium in neuromuscular facilitation. J Physiol (Lond) 195,
481-492. KATZ, B. & MILEDI, R. (1971). The effect of prolonged depolarization on synaptic transfer in the stellate
ganglion of the squid. J Physiol 216, 503-512. KORN, H., FABER, D. S. & TRILLER, A. (1986). Probabilistic determination of synaptic strength. J
Neurophysiol 55, 402-421. LANG, F., ATWOOD, H. L. & MORIN, W. A. (1972). Innervation and vascular supply of the crayfish opener
muscle: scanning and transmission electron microscopy. Z Zellforsch Mikrosk Anat 127, 189-200. LINDER, T. M. (1974). The accumulative properties of facilitation at crayfish neuromuscular synapses. J
Physiol 238, 223-234. LIU, Y. & STANLEY, E. F. (1995). Calcium binding sites of the transmitter release mechanism: clues from
short-term facilitation. J Physiol Paris 89, 163-166. MARDER, E. & PAUPARDIN-TRITSCH, D. (1978). The pharmacological properties of some crustacean
neuronal acetylcholine, gamma-aminobutyric acid, and L-glutamate responses. J Physiol (Lond) 280, 213-236.
MARKRAM, H., WANG, Y. & TSODYKS, M. (1998). Differential signaling via the same axon of neocortical
pyramidal neurons. Proc Natl Acad Sci U S A 95, 5323-5328. MATVEEV, V., SHERMAN, A. & ZUCKER, R. S. (2002). New and corrected simulations of synaptic
facilitation. Biophys J 83, 1368-1373. MEINRENKEN ,C. J., BORST ,J. G. G. & SAKMANN ,B. (2002). Calcium Secretion Coupling at Calyx of Held
Governed by Nonuniform Channel-Vesicle Topography .J. Neurosci . 22, 1648-1667. MEINRENKEN ,C. J., BORST ,J. G. G. & SAKMANN ,B. (2003). Local routes revisited: the space and time
dependence of the Ca2+ signal for phasic transmitter release at the rat calyx of Held .J Physiol (Lond) 547, 665-689.
MILEDI, R. & THIES, R. (1971). Tetanic and post-tetanic rise in frequency of miniature end-plate
potentials in low-calcium solutions. J Physiol 212, 245-257. MIWA, A., UI, M. & KAWAI, N. (1990). G protein is coupled to presynaptic glutamate and GABA receptors
in lobster neuromuscular synapse. J Neurophysiol 63, 173-180.

MSGHINA, M., MILLAR, A. G., CHARLTON, M. P., GOVIND, C. K. & ATWOOD, H. L. (1999). Calcium entry related to active zones and differences in transmitter release at phasic and tonic synapses. J Neurosci 19, 8419-8434.
ONODERA, K. & TAKEUCHI, A. (1980). Distribution and pharmacological properties of synaptic and
extrasynaptic glutamate receptors on crayfish muscle. J Physiol 306, 233-250. PARNAS, H., DUDEL, J. & PARNAS, I. (1982a). Neurotransmitter release and its facilitation in crayfish. I.
Saturation kinetics of release, and of entry and removal of calcium. Pflugers Arch 393, 1-14. PARNAS, H., DUDEL, J. & PARNAS, I. (1986). Neurotransmitter release and its facilitation in crayfish. VII.
Another voltage dependent process beside Ca entry controls the time course of phasic release. Pflugers Arch 406, 121-130.
PARNAS, H., SEGEL, L., DUDEL, J. & PARNAS, I. (2000a). Autoreceptors, membrane potential and the
regulation of transmitter release. Trends Neurosci 23, 60-68. PARNAS, H., VALLE-LISBOA, J. C. & SEGEL, L. A. (2002). Can the Ca2+ hypothesis and the Ca2+-voltage
hypothesis for neurotransmitter release be reconciled? Proc Natl Acad Sci U S A 99, 17149-17154. PARNAS, I., DUDEL, J. & GROSSMAN, Y. (1982b). Chronic removal of inhibitory axon alters excitatory
transmission in a crustacean muscle fiber. J Neurophysiol 47, 1-10. PARNAS, I. & PARNAS, H. (1986). Calcium is essential but insufficient for neurotransmitter release: the
calcium-voltage hypothesis. J Physiol (Paris) 81, 289-305. PARNAS, I., PARNAS, H. & DUDEL, J. (1982c). Neurotransmitter release and its facilitation in crayfish
muscle. V. Basis for synapse differentiation of the fast and slow type in one axon. Pflugers Arch 395, 261-270.
PARNAS, I., RASHKOVAN, G., ONG, J. & KERR, D. I. (1999). Tonic activation of presynaptic GABAB
receptors in the opener neuromuscular junction of crayfish. J Neurophysiol 81, 1184-1191. PARNAS, I., RASHKOVAN, G., RAVIN, R. & FISCHER, Y. (2000b). Novel mechanism for presynaptic
inhibition: GABA(A) receptors affect the release machinery. J Neurophysiol 84, 1240-1246. PIERCE, J. P. & MENDELL, L. M. (1993). Quantitative ultrastructure of Ia boutons in the ventral horn:
scaling and positional relationships. J Neurosci 13, 4748-4763. RAGOZZINO, D., WOODWARD, R. M., MURATA, Y., EUSEBI, F., OVERMAN, L. E. & MILEDI, R. (1996). Design
and in vitro pharmacology of a selective gamma-aminobutyric acidC receptor antagonist. Mol Pharmacol 50, 1024-1030.
RAVIN ,R., SPIRA , M., PARNAS ,H. & PARNAS ,I. (1997). Simultaneous measurement of intracellular Ca2+
and asynchronous transmitter release from the same crayfish bouton .J Physiol (Lond) 501, 251-262
RAVIN, R., PARNAS, H., SPIRA, M. E., VOLFOVSKY, N. & PARNAS, I. (1999). Simultaneous measurement of
evoked release and [Ca2+]i in a crayfish release bouton reveals high affinity of release to Ca2+. J Neurophysiol 81, 634-642.
ROSENTHAL, J. (1969). Post-tetanic potentiation at the neuromuscular junction of the frog. J Physiol 203,
121-133. SCHNEGGENBURGER, R. & NEHER, E. (2000). Intracellular calcium dependence of transmitter release
rates at a fast central synapse. Nature 406, 889-893. SEGEV, I. (1990). Computer study of presynaptic inhibition controlling the spread of action potentials
into axonal terminals. J Neurophysiol 63, 987-998. SHERMAN, R. G. & ATWOOD, H. L. (1972). Correlated electrophysiological and ultrastructural studies of a
crustacean motor unit. J Gen Physiol 59, 586-615.

SLUTSKY, I., PARNAS, H. & PARNAS, I. (1999). Presynaptic effects of muscarine on ACh release at the frog neuromuscular junction. J Physiol 514 ( Pt 3), 769-782.
SMITH, D. O. (1983a). Axon conduction failure under in vivo conditions in crayfish. J Physiol 344, 327-
333. SMITH, D. O. (1983b). Variable activation of synaptic release sites at the neuromuscular junction. Exp
Neurol 80, 520-528. STANLEY, E. F. (1986). Decline in calcium cooperativity as the basis of facilitation at the squid giant
synapse. J Neurosci 6, 782-789. STRAWN, J. R. & COOPER, R. L. (2002). The effects of ethanol on pre-synaptic components of synaptic
transmission in a model glutamatergic synapse: the crayfish neuromuscular junction. Comp Biochem Physiol C Toxicol Pharmacol 131, 395-404.
TAKEUCHI, A. & TAKEUCHI, N. (1969). A study of the action of picrotoxin on the inhibitory neuromuscular
junction of the crayfish. J Physiol (Lond) 205, 377-391. TANG, Y., SCHLUMPBERGER, T., KIM, T., LUEKER, M. & ZUCKER, R. S. (2000). Effects of mobile buffers on
facilitation: experimental and computational studies. Biophys J 78, 2735-2751. TANK, D. W., REGEHR, W. G. & DELANEY, K. R. (1995). A quantitative analysis of presynaptic calcium
dynamics that contribute to short-term enhancement. J Neurosci 15, 7940-7952. TARASKEVICH, P. S. (1971). Reversal potentials of L-glutamate and the excitatory transmitter at the
neuromuscular junction of the crayfish. Biochim Biophys Acta 241, 700-703. THOMPSON, C. S. & ATWOOD, H. L. (1984). Synaptic strength and horseradish peroxidase uptake in
crayfish nerve terminals. J Neurocytol 13, 267-280. THOMSON, A. M. (1997). Activity-dependent properties of synaptic transmission at two classes of
connections made by rat neocortical pyramidal axons in vitro. J Physiol 502 ( Pt 1), 131-147. THOMSON, A. M., DEUCHARS, J. & WEST, D. C. (1993). Single axon excitatory postsynaptic potentials in
neocortical interneurons exhibit pronounced paired pulse facilitation. Neuroscience 54, 347-360. VAN DER KLOOT, W. & MOLGO, J. (1993). Facilitation and delayed release at about 0 degree C at the frog
neuromuscular junction: effects of calcium chelators, calcium transport inhibitors, and okadaic acid. J Neurophysiol 69, 717-729.
VIELE, K., STROMBERG, A. J. & COOPER, R. L. (2003). Estimating the number of release sites and
probability of firing within the nerve terminal by statistical analysis of synaptic charge. Synapse 47, 15-25.
VYSHEDSKIY, A., ALLANA, T. & LIN, J. W. (2000). Analysis of presynaptic Ca2+ influx and transmitter
release kinetics during facilitation at the inhibitor of the crayfish neuromuscular junction. J Neurosci 20, 6326-6332.
VYSHEDSKIY, A. & LIN, J. W. (1997). Activation and detection of facilitation as studied by presynaptic
voltage control at the inhibitor of the crayfish opener muscle. J Neurophysiol 77, 2300-2315. WEINREICH, D. (1971). Ionic mechanism of post-tetanic potentiation at the neuromuscular junction of the
frog. J Physiol 212, 431-446. WERNIG, A. (1972). Changes in statistical parameters during facilitation at the crayfish neuromuscular
junction. J Physiol (Lond) 226, 751-759. WINSLOW, J. L., DUFFY, S. N. & CHARLTON, M. P. (1994). Homosynaptic facilitation of transmitter release
in crayfish is not affected by mobile calcium chelators: implications for the residual ionized calcium hypothesis from electrophysiological and computational analyses. J Neurophysiol 72, 1769-1793.

WOJTOWICZ, J. M. & ATWOOD, H. L. (1986). Long-term facilitation alters transmitter releasing properties at the crayfish neuromuscular junction. J Neurophysiol 55, 484-498.
WOJTOWICZ, J. M., MARIN, L. & ATWOOD, H. L. (1994). Activity-induced changes in synaptic release sites
at the crayfish neuromuscular junction. J Neurosci 14, 3688-3703. WOJTOWICZ, J. M., PARNAS, I., PARNAS, H. & ATWOOD, H. L. (1988). Long-term facilitation of synaptic
transmission demonstrated with macro-patch recording at the crayfish neuromuscular junction. Neurosci Lett 90, 152-158.
WRIGHT, S. N., BRODWICK, M. S. & BITTNER, G. D. (1996a). Calcium currents, transmitter release and
facilitation of release at voltage-clamped crayfish nerve terminals. J Physiol (Lond) 496, 363-378. WRIGHT, S. N., BRODWICK, M. S. & BITTNER, G. D. (1996b). Presynaptic calcium currents at voltage-
clamped excitor and inhibitor nerve terminals of crayfish. J Physiol (Lond) 496, 347-361. YOUNKIN, S. G. (1974). An analysis of the role of calcium in facilitation at the frog neuromuscular
junction. J Physiol (Lond) 237, 1-14. ZUCKER, R. S. (1973). Changes in the statistics of transmitter release during facilitation. J Physiol
(Lond) 229, 787-810. ZUCKER, R. S. (1974). Crayfish neuromuscular facilitation activated by constant presynaptic action
potentials and depolarizing pulses. J Physiol (Lond) 241, 69-89. ZUCKER, R. S. & REGEHR, W. G. (2002). Short-term synaptic plasticity. Annu Rev Physiol 64, 355-405.

ABSTRACT
One of the classic characteristics of chemical synapses is the ability to facilitate
release in response to high frequency stimulation. During the past decades many
researchers have described different aspects of Facilitation, Potentiation and other
delayed types of synaptic amplification, however, there are still some significant
gaps in our understanding of short term synaptic plasticity processes. The nature of
the relation between stimulation frequency and plateau levels of synaptic release
(frequency response curve FR curve) has not been characterized yet, nor has the
effect of stimulation frequency on the kinetics and duration of potentiation. The
differences between phasic and tonic synapses regarding these aspects are not
fully understood, so are the ways various presynaptic factors, such as extracellular
calcium concentration ([Ca+2]OUT) or presynaptic inhibition, affects the frequency
response curve of a given terminal.
All these topics were studied at the opener muscle NMJ of the first walking leg of the
Crayfish. Experiments were conducted on two different types of terminal
populations: proximal (phasic type) and central (tonic type), using both nerve and
direct stimulation methods.
Results show significant similarity between synaptic FR curves, and the classic
curve that describes release as a function of [Ca+2]OUT. Lowering [Ca+2]OUT
decreases release while increasing the FR curve slope, and applying a powerful
(stronger than the physiologic) direct stimulation to the terminal, results in the exact
opposite effect. These and some other evidence suggest that the FR curve slope
could represent the release cooperativity, supporting the hypothesis that views
cooperativity decrease as a marker for increasing release potentiation. Several
significant differences were measured between FR curves of proximal and central

terminals, mainly a higher slope and a longer duration of potentiation at the central
terminals. Activating presynaptic inhibition (by applying GABA) had no effect on the
FR curves slope nor on most of other differences mentioned above, suggesting that
presynaptic inhibition reduces release using some other mechanism, in addition to
reducing [Ca+2]Out. Although this inhibition is mediated via a GABAA-like receptor, the
exact physiologic mechanism by which it exerts its effect is still unclear, particulary
at the central terminals.

This work was carried out under the supervision of: Prof Yoram Grosman
In the Department: Physiology
Faculty: Health Sciences

The Effects of Presynaptic Inhibition on Frequency Response of Crayfish Neuromuscular Synapses
Thesis submitted in partial fulfillment
of the requirements for the degree of
DOCTOR OF PHILOSOPHY
by
Ben Alkahe
Submitted to the Senate of Ben-Gurion University
of the Negev
Approved by the advisor
Approved by the Dean of the Kreitman School of Advanced Graduate Studies
April 2005 Beer-Sheva

The Effects of Presynaptic Inhibition on Frequency Response of Crayfish Neuromuscular Synapses
Thesis submitted in partial fulfillment
of the requirements for the degree of
DOCTOR OF PHILOSOPHY
by
Ben Alkahe
Submitted to the Senate of Ben-Gurion University
of the Negev
April 2005 Beer-Sheva
Copyright © 2022 FDOKUMEN




















