
PERTEMUAN 2 NEUROENDOKRINOLOGI
49
BAB I PENDAHULUAN A. Latar Belakang Neuroendokrinologi adalah studi tentang hubungan sistem regulatorik antara sistem saraf dan endokrin. Pelepasan hormon yang berasal dari neuroendokrin berlangsung seperti pelepasan neurotransmiter pada sel saraf, hanya sekretnya diditribusikan melalui sirkulasi darah. Dua tempat kerja utama dalam otak yang penting dalam regulasi fungsi reproduktif hipotalamus dan kelenjar pituitary siklus menstruasi dikontrol oleh steroid seks dan peptida yang diproduksi dalam folikel yang nantinya akan mengalami ovulasi. Namun, perkembangan selama 2 dekade terakhir ini menujukkan bahwa sekuens kejadian yang kompleks tersebut yang dikenal sebagai siklus menstruasi dikontrol oleh steroid seks dan peptida yang diproduksi dalam folikel yang nantinya akan mengalami ovulasi. Hipotalamus dan pengarahannya, serta pituitari adalah penting untuk bekerjanya keseluruhan mekanisme, tetapi fungsi endokrin yang menyebabkan ovulasi ditimbulkan oleh umpan balik endoktrin pada pituitary anterior. Hipotalamus memproduksi estradiol 1
description
neuroendokrinologi
Transcript of PERTEMUAN 2 NEUROENDOKRINOLOGI

BAB I
PENDAHULUAN
A. Latar Belakang
Neuroendokrinologi adalah studi tentang hubungan sistem regulatorik
antara sistem saraf dan endokrin. Pelepasan hormon yang berasal dari
neuroendokrin berlangsung seperti pelepasan neurotransmiter pada sel saraf,
hanya sekretnya diditribusikan melalui sirkulasi darah. Dua tempat kerja utama
dalam otak yang penting dalam regulasi fungsi reproduktif hipotalamus dan
kelenjar pituitary siklus menstruasi dikontrol oleh steroid seks dan peptida
yang diproduksi dalam folikel yang nantinya akan mengalami ovulasi.
Namun, perkembangan selama 2 dekade terakhir ini menujukkan bahwa
sekuens kejadian yang kompleks tersebut yang dikenal sebagai siklus
menstruasi dikontrol oleh steroid seks dan peptida yang diproduksi dalam
folikel yang nantinya akan mengalami ovulasi.
Hipotalamus dan pengarahannya, serta pituitari adalah penting untuk
bekerjanya keseluruhan mekanisme, tetapi fungsi endokrin yang menyebabkan
ovulasi ditimbulkan oleh umpan balik endoktrin pada pituitary anterior.
Hipotalamus memproduksi estradiol yang dapat diproduksi di otak. Estradiol
adalah salah satu dari tiga hormon yang berperan dalam membentuk esterogen
dan merupakan hormon utama dalam reproduksi wanita yang tidak hanya
diproduksi di dalam alat reproduksi wanita saja. Estradiol dapat mempengaruhi
fungsi tubuh, terutama dalam pengaturan ingatan dan berat badan.
Hipotalamus selain memproduksi estradiol juga membantu pengendalian
gonadotrophin releasing hormone (GnRH). Hormon GnRH mengatur
pertumbuhan, perkembangan seksual, dan fungsi reproduksi. GnRH merupakan
hormon tropik utama dalam regulasi fungsi sel gonadotropin sehingga
memegang peranan penting dalam sistem reproduksi disamping TRH
(Thyrotropin Releasing Factor) dan PIF (Prolactine Inhibiting Factor).
Untuk lebih memahami kaitan antara sistem saraf dan hormon yang
mempengaruhi di sistem reproduksi maka makalah ini akan membahas lebih
1

dalam neuroendokrin dalam sistem reproduksi (neuroendocrinology of
reproduction).
B. Tujuan:
1. Mengetahui neuroendokrin dalam sistem reproduksi
2. Mengetahui mekanisme kerja hipotalamus hipofisis dalam reproduksi
3. Mengetahui konsep neurohormon
4. Mengetahui sekresi prolaktin
5. Mengetahui hipotalamus dan sekresi GnRH
2

BAB II
PEMBAHASAN
A. Neuroendokrin Dalam Sistem Reproduksi
Terdapat dua tempat kerja utama dalam otak yang penting dalam regulasi
fungsi reproduktif – hipotalamus dan kelenjar pituitari. Dahulu, kelenjar
pituitari dianggap sebagai kelenjar utama. Kemudian muncullah suatu konsep
baru dimana pituitari diberi peranan lebih rendah sebagai bagian dari suatu
orkestra, dengan hipotalamus sebagai konduktornya, dan merespon kepada
pesan-pesan dari sistem saraf perifer maupun pusat dan menunjukkan
pengaruhnya dengan melalui neurotransmiter yang dikirimkan menuju pituitari
oleh jaringan vasa porta. Tanpa melihat daerah mana yang dominan, tesis
konvesionalnya adalah bahwa kompleks sistem saraf pusat-pituitari
menentukan dan mengarahkan kronologi kejadian-kejadian perkembangan
dalam ovarium yang responsif.
Suatu pemahaman menyeluruh mengenai gambaran biologi reproduktif
ini akan memberi keuntungan bagi klinisi yang menghadapi permasalahan
dalam endokrinologi ginekologis. Dengan pemahaman ini, klinisi dapat
memahami efek misterius dari stres, diet, olah raga, dan pengaruh-pengaruh
lain pada aksis pituitari-gonad. Lebih lanjut, kita akan siap untuk
mempergunakan berbagai agen farmakologis yang merupakan bagian dari
penelitian neuroendokrin. Sampai saat ini, bab ini memberikan tinjauan
berorientasi klinis mengenai status neuroendokrinologi reproduktif saat ini.
B. Sirkulasi Porta Hipotalamus Hipofisis
Sistem porta adalah susunan pembuluh darah dimana darah vena mengalir
langsung dari satu anyaman kapiler melalui pembuluh penghubung keanyaman
kapiler lain. Sistem portal hipotalamus hipofisis menjadi penghubung penting
antar otak dan sengaian besar sistem endokrin. Sistem ini berawal di dasar
hipotalamus dengan sekelompok kapiler yang menyatu membentuk pembuluh-
pembuluh porta halus, yang mengalir melalui tangkai penghubung kedalam
hipofisis anterior. Hipotalamus terletak pada dasar otak tepat diatas junction
3

nervus optikus. Untuk mempengaruhi kelenjar pituitari anterior, otak
memerlukan suatu cara untuk mengadakan transmisi atau hubungan. Arteri
hipofisialis superior membentuk suatu jaringan kapiler padat dalam eminensia
median tersebut, yang kemudian mengalami drainase kedalam vasa porta yang
menurun sepanjang batang pituitari sampai ke pituitari anterior.
Arah aliran darah dalam sirkulasi porta hipofisialis ini adalah dari otak
menuju pituitari. Bagian batang neural, yang mengganggu sirkulasi porta ini,
menyebabkan inaktivitas dan atrofi gonad, bersama dengan penurunan aktivitas
adrenal dan tiroid mencapai tingkat basal. Dengan regenerasi vasa porta, fungsi
pituitari anterior dapat dikembalikan. Karena itu, kelenjar pituitari anterior
berada dibawah pengaruh hipotalamus melalui neurohormon-neurohormon
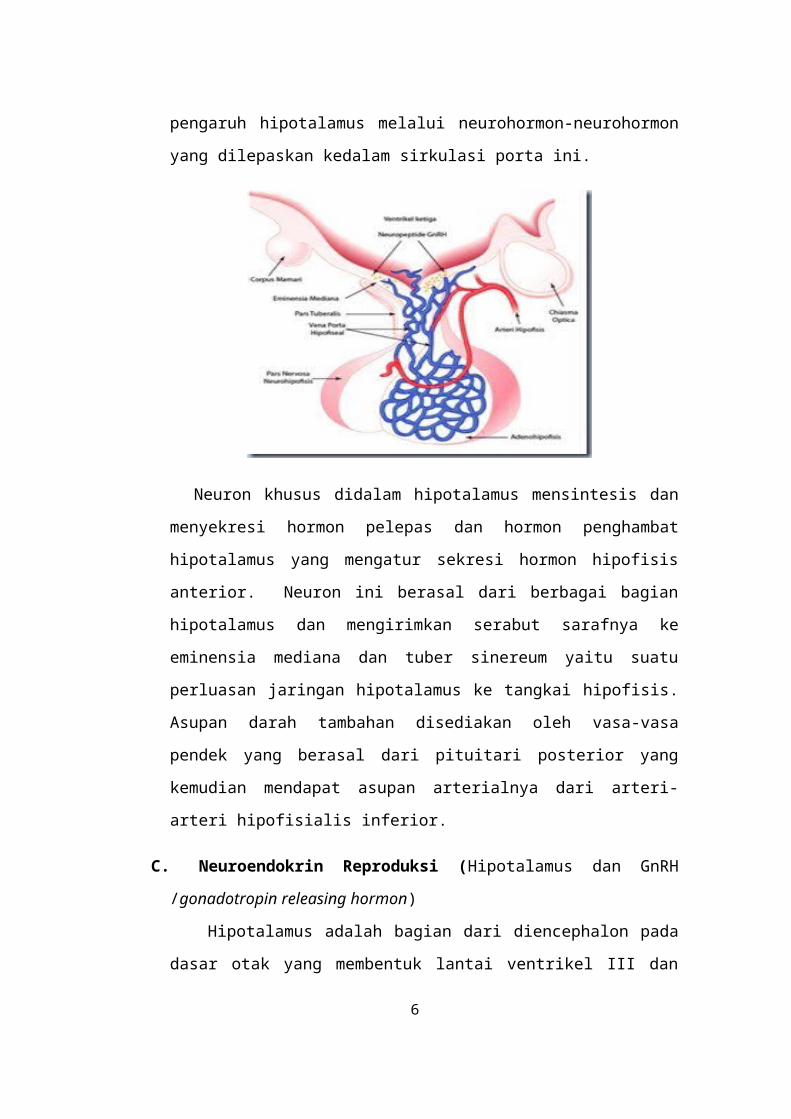
yang dilepaskan kedalam sirkulasi porta ini.
Neuron khusus didalam hipotalamus mensintesis dan menyekresi hormon
pelepas dan hormon penghambat hipotalamus yang mengatur sekresi hormon
hipofisis anterior. Neuron ini berasal dari berbagai bagian hipotalamus dan
mengirimkan serabut sarafnya ke eminensia mediana dan tuber sinereum yaitu
suatu perluasan jaringan hipotalamus ke tangkai hipofisis. Asupan darah
tambahan disediakan oleh vasa-vasa pendek yang berasal dari pituitari
posterior yang kemudian mendapat asupan arterialnya dari arteri-arteri
hipofisialis inferior.
4

C. Neuroendokrin Reproduksi (Hipotalamus dan GnRH /gonadotropin
releasing hormon)
Hipotalamus adalah bagian dari diencephalon pada dasar otak yang
membentuk lantai ventrikel III dan sebagian dinding lateralnya. Bagian dalam
hipotalamus merupakan sel neural peptidergik yang mensekresi releasing
hormone dan inhibiting hormone. GnRH merupakan salah satu hormon yang
dikeluarkan oleh hipotalamus yang dapat merangsang pengeluaran hormon
gonadotropin yaitu FSH dan LH dari hipofisis anterior. FSH dan LH
merangsang ovarium pada wanita dan testis pada pria dan mempengaruhi
fungsi reproduksi.
Sel-sel yang memproduksi GnRH berasal dari area olfaktoria, melalui
migrasi selama embriogenesis, sel-sel bergerak ke nervus kranialis yang
menghubungkan hidung dan otak bagian depan menuju lokasi utamanya yaitu
nukleus arkuata hipotalamus dimana dapat ditemukan 1000-3000 sel-sel yang
memproduksi GnRH. Neuron khusus didalam hipotalamus mensintesis dan
menyekresi hormon pelepas dan hormon penghambat hipotalamus yang
mengatur sekresi hormon hipofisis anterior. Neuron ini berasal dari berbagai
bagian hipotalamus dan mengirimkan serabut sarafnya ke eminensia mediana
dan tuber sinereum yaitu suatu perluasan jaringan hipotalamus ke tangkai
hipofisis.
Hormon yang dihasilkan oleh hipotalamus adalah :
Growth Hormone-Releasing Hormone (GHRH) berfungsi meningkatkan
sekresi Growth Hormone (GH) atau hormon pertumbuhan dari kelenjar
pituitari anterior.
Growth Hormone-Inhibiting Hormone (GHIH) atau disebut juga
somatostatin adalah hormon yang menghambat sekresi hormon
pertumbuhan (Growth Hormone) dari kelenjar pituitari anterior.
Thyroid-releasing hormone (TRH) berfungsi untuk merangsang sekresi
TSH (Thyroid-stimulating hormone)
Corticotropin-releasing hormone (CRH) berfungsi merangsang sekresi
hormon ardenokortikotropik (ACTH)
5
Gonadotropin-releasing hormone (GnRH) berfungsi merangsang sekresi
LH (Luteinizing Hormone) dan FSH (Follicle-stimulating hormone).
Prolactin-releasing hormone (PRH) merangsang sekresi prolaktin
Prolactin-inhibiting hormone (PIH) menghambat sekresi prolaktin.
D. Kelenjar Hipofise
Hipofise merupakan kelenjar endokrin yang mempunyai kemampuan
untuk mempengaruhi kelenjar-kelenjar endokrin yang lain dengan perantaraan
hormon-hormon yang dihasilkannya.
Terdiri atas 2 bagian yaitu :
1. Neurohipofisis (pituitari posterior) mensekresikan hormon antidiuretik dan
hormon oksitosin
2. Adenohipofisis (hipofise anterior)
Hormon yang disekresikan dari hipofisis anterior ini disebut tropik.
Hormon yang mempengaruhi sekresi hormon dari kelenjar endokrin lain.
Hormon tersebut yaitu :
• Hormon pertumbuhan (growth hormone, GH, somatotropin),
hormone primer yang bertanggung jawab mengatur pertumbuhan tubuh
keseluruhan, juga penting dalam metabolisme intermediate.
• Thyroid-stimulating hormone (TSH, tiritropin) merangsang sekresi
hormone tiroid dan pertumbuhan kelenjar tiroid.
• Hormon adrenokortikotropik (adrenocorticotropic hormone,
ACTH, adrenokortikotropin) merangsang sekresi kortisol oleh korteks
adrenal dan mendorong pertumbuhan korteks adrenal.
• Follicle-stimulating hormone (FSH) memiliki fungsi berbeda pada
wanita dan pria.
• Luteinizing hormone (LH) juga berfungsi berbeda pada wanita dan
pria.
• Prolaktin (PRL) meningkatkan perkembangan payudara dan
produksi susu pada wanita. Fungsinya pada pria belum jelas, meskipun
bukti menunjukkan bahwa hormon ini mungkin merangsang produksi
reseptor LH di testis.
6

E. Mekanisme Kerja Hipotalamus Hipofisis Dalam Reproduksi
1. Pengontrolan aktivitas kelenjar hipofisis sebagian besar dilakukan oleh
hipotalamus dengan suatu modulasi langsung yanng sangat penting melalui
mekanisme umpan balik.
2. Nukleus hipotalamus yang berhubungan dengan proses reproduksi antara
lain adalah nukleus supraoptik, paraventrikular, arkuata, ventromedial dan
suprakiasma
3. Beberapa sinyal saraf instriksik yang berhubungan dengan sistem
reproduksi dibentuk di hipotalamus. Sinyal ini berasal dari suatu generator
denyut (pulse generator) untuk GnRH (gonadotropin releasing hormon) dan
dari hormon dopaminergik yang proyeksinya menuju eminensia mediana
hipotalamus.
4. Sinyal neuroendokrin yang dibentuk dalam hipotalamus diperantarai oleh
faktor pelepas peptida yang berjalan sepanjang sistem portal hipotalamus-
hipofisis ke tempat kerja mereka di kelenjar hipofisis
5. Kemampuan reproduksi bergantung pada hubungan antara hipotalamus,
hipofisis anterior, organ reproduksi dan sel sasaran hormon seks.
6. Hubungan ini menggunakan banyak mekanisme regulatorik yang digunakan
oleh sistem tubuh lain untuk mempertahankan homeostatis misalnya kontrol
umpan balik negatif.
F. Konsep Neurohormon
Sejumlah besar bukti menunjukkan bahwa pengaruh hipotalamus pada
pituitari dicapai melalui bahan-bahan yang disekresikan dalam sel-sel dari
hipotalamus dan dikirimkan ke pituitari oleh sistem vasa porta. Memang,
proliferasi sel dan ekspresi gen pituitari dikontrol oleh peptida-peptida
hipotalamik dan reseptornya, ada sesuatu yang sangat khusus mengenai darah
yang mendrainase hipotalamus basal. Sebuah perkecualian terhadap pola
umum pengaruh positif ini adalah kontrol sekresi prolaktin.
7

Sekresi dan transplantasi batang pituitari menyebabkan pelepasan
prolaktin dari pituitari anterior, ini mengimplikasikan kontrol inhibitorik
negatif oleh hipotalamus. Lebih lanjut, kultur jaringan pituitari anterior
melepaskan prolaktin pada tidak adanya jaringan hipotalamus atau ekstraknya.
Agen-agen neuroendokrin yang berasal dari dalam hipotalamus memiliki
efek stimulatorik positif pada hormon pertumbuhan, thyroid- stimulating
hormone (TSH), hormon adrenokortikotropin (ACTH), maupun gonadotropin,
dan merupakan neurohormon-neurohormon individual dari hipotalamus.
Neurohormon yang mengontrol gonadotropin disebut gonadotropin-releasing
hormone (GnRH).
Neurohormon yang mengontrol prolaktin disebut prolactin-inhibiting
hormone dan berupa dopamin. Corticotropin-Releasing Hormone (CRH)
manusia adalah suatu peptida asam amino 41 yang merupakan regulator utama
sekresi ACTH, dan yang juga mengaktivasi sistem saraf simpatis. Seperti yang
akan kita lihat, CRH dapat menekan sekresi gonadotropin, suatu aksi yang
sebagian dimediasi oleh inhibisi GnRH oleh endorfin. Disamping efeknya pada
pituitari, efek perilaku dalam otak telah dijumpai untuk beberapa diantara
hormon-hormon pelepas tersebut.
Thyrotropin-releasing hormone (TRH) mengantagonisasi kerja sedatif dari
sejumlah obat dan juga memiliki efek antidepresan direk pada manusia.
Awalnya, diyakini bahwa ada dua hormon pelepas yang berbeda, satu untuk
follicle-stimulating hormone (FSH) dan untuk hormon luteinisasi (LH). Saat ini
telah diterima bahwa terdapat satu neurohormon (GnRH) untuk kedua
gonadotropin. GnRH adalah peptida kecil dengan 10 asam amino dengan
sejumlah variasi dalam sekuens asam amino diantara berbagai mamalia.
Sekarang telah jelas bahwa GnRH memiliki fungsi autokrin-parakrin
diseluruh tubuh. GnRH terdapat jaringan neural maupun jaringan nonneural,
dan reseptor terdapat dalam banyak jaringan ekstrapituitari (misalnya folikel
ovarium dan plasenta). Walaupun GnRH pada semua mamalia adalah identik,
terdapat bentuk-bentuk non-mamalia lain, menunjukkan bahwa molekul GnRH
telah ada setidaknya selama 500 juta tahun.
8

Sebuah analisis mengenai evolusi GnRH menunjukkan 3 bentuk mayor:
GnRH yang terlokalisir pada hipotalamus (GnRH-I), terbentuk pada inti
pertengahan otak dan diluar otak (GnRH-II), dan terbentuk pada sejumlah
spesies ikan (GnRH-III), dan karenanya menunjukkan gambaran dari berbagai
bentuk GnRH sebelum munculnya vertebra.
Mungkin anggapan bahwa pituitari adalah kelenjar utama tidak boleh
dilupakan. Walaupun pituitari diatur oleh input dari daerah-daerah lain,
fungsinya penting untuk mempertahankan kehidupan. Perkembangan dan
aktivitas pituitari berada dibawah kontrol hipotalamus (dengan input dari
daerah-daerah sistem saraf pusat lainnya), dan respon pituitari diatur dengan
baik oleh pesan-pesan hormonal dari jaringan yang menjadi target hormon-
hormon trofik pituitari. Disamping itu, pituitari memiliki sistem autokrin-
parakrinnya sendiri untuk perbaikan dan supresi pertumbuhan dan fungsi.
Tetapi kelenjar pituitari merupakan fokus untuk semua aktivitas ini, dan peran
koordinasi sentral ini penting untuk kehidupan normal.
G. Sekresi Prolaktin
Ekspresi gen prolaktin terjadi dalam laktotrof dari kelenjar pituitari
anterior, dalam endometrium yang mengalami desidualisasi, dan dalam
miometrium. Prolaktin yang disekresikan dalam berbagai tempat ini bersifat
identik, tetapi terdapat perbedaan dalam mRNA yang menunjukkan perbedaan
dalam regulasi gen prolaktin. Transkripsi gen prolaktin diatur oleh suatu faktor
transkripsi (sebuah protein yang disebut Pit-1) yang berikatan dengan regio
promoter 5’ dan yang juga diperlukan untuk sekresi hormon pertumbuhan dan
TSH. Disamping itu, transkripsi gen prolaktin diatur oleh interaksi estrogen
dan reseptor-reseptor glukokortioid dengan sekuens 5’ flanking.
Fungsi utama prolaktin pada mamalia adalah laktogenesis, mungkin
berperan dalam supresi seksualitas segera setelah bangkitnya nafsu seksual dan
orgasme. Ekspresi gen prolaktin diregulasi lebih lanjut oleh faktor-faktor
spesies-spesifik lain. Transkripsi gen prolaktin dirangsang oleh estrogen dan
dimediasi oleh reseptor estrogen yang berikatan dengan elemen-elemen
estrogen-responsif.
9

Beberapa faktor menunjukkan efek stimulatorik pada sekresi prolaktin
(faktor-faktor pelepas prolaktin), terutama TRH, peptida usus vasoaktif (VIP),
epidermal growth factor, dan mungkin GnRH. Faktor- faktor ini saling
berinteraksi, mempengaruhi responsivitas umum laktotrof. Namun,
homeostasis prolaktin diregulasi terutama oleh prolaktin sendiri, dengan umpan
balik pada neuron-neuron pelepas dopamin. Mekanisme dopaminergik ini
sangat dipengaruhi oleh estrogen, baik secara langsung maupun melalui
neurotransmiter.
Sekresi prolaktin dapat dipahami dengan memandang dopamin yang
diterima melalui sistem porta sebagai hal yang bertanggung-jawab untuk
terjadinya inhibisi tonik, dan sistem dopaminergik dirangsang oleh prolaktin
(menurunkan sekresi) dan dihambat oleh estrogen (meningkatkan sekresi).
Pengaruh-pengaruh modulasi meliputi aktivitas inhibitorik opioid endogen dan
stimulasi oleh berbagai substansi, termasuk serotonin dan neuropeptida Y.
H. Hipotalamus Dan Sekresi GnRH
1. Hipotalamus
Hipotalamus adalah bagian dari diensefalon yang terletak pada dasar
otak yang membentuk lantai dan sebagian dari dinding lateral ventrikel
ketiga. Dalam hipotalamus terdapat sel-sel neural peptidergik yang
mensekresi hormon-hormon pelepas dan penghambat. Sel-sel ini memiliki
karakteristik neuron maupun sel-sel kelenjar endokrin. Sel-sel ini memberi
respon kepada sinyal-sinyal dalam aliran darah, maupun kepada
neurotransmiter dalam otak dalam suatu proses yang dikenal sebagai
neurosekresi. Dalam neurosekresi, suatu neurohormon atau neurotransmiter
disintesis pada ribosom dalam sitoplasma neuron, dikirimkan kedalam
sebuah granula dalam aparatus Golgi, dan kemudian oleh aliran akson aktif
dikirim ke ujung akhir neuron untuk disekresikan kedalam pembuluh darah
atau melewati sinaps.
Neuron GnRH terdapat dalam jaringan yang kompleks dan saling
berhubungan satu sama lain dan dengan banyak neuron lain. Pengaturan
fisik ini memungkinkan interaksi ganda dengan neurotransmiter, hormon,
dan growth factor untuk memodulasi pelepasan GnRH. Pengiriman GnRH
10

menuju sirkulasi porta dilakukan melalui jalur aksonal, yaitu traktus GnRH
tuberoinfundibuler.
2. Sekresi GnRH
Waktu paruh GnRH hanya 2-4 menit. Karena degradasi cepat ini,
dikombinasi dengan banyaknya dilusi pada saat masuk kedalam sirkulasi
perifer, jumlah GnRH yang aktif secara biologis tidak dapat lepas dari
sistem porta. Karena itu, kontrol siklus reproduktif bergantung pada
pelepasan GnRH secara konstan. Sebaliknya, fungsi ini bergantung pada
interaksi kompleks dan terkoordinasi antara hormon pelepas ini,
neurohormon-neurohormon lain, gonadotropin pituitari, dan steroid gonad.
Interaksi antara substansi-substansi ini diatur oleh efek umpan balik, baik
stimulatorik positif maupun inhibitorik negatif.
Loop umpan balik panjang mengacu pada efek umpan balik dari kadar
hormon kelenjar target dalam sirkulasi, dan ini terjadi dalam hipotalamus
maupun pituitari. Loop umpan balik pendek menunjukkan umpan balik
negatif dari hormon pituitari pada sekresinya sendiri, mungkin melalui efek
inhibitorik pada hormon- hormon pelepas dalam hipotalamus.
Umpan balik ultrapendek mengacu pada inhibisi oleh hormon-hormon
pelepas terhadap sintesisnya sendiri. Sinyal-sinyal ini maupun sinyal-sinyal
dari pusat- pusat yang lebih tinggi dalam sistem saraf pusat dapat
memodifikasi sekresi GnRH melalui sekelompok neurotransmiter, terutama
dopamin, norepinefrin, dan endofrin dan juga serotonin dan melatonin.
Dopamin dan norepinefrin disintesis dalam akhiran-akhiran saraf melalui
dekarboksilasi dihidrofenilalanin (DOPA), yang kemudian disintesis oleh
hidroksilasi tirosin. Dopamin adalah prekursor intermediet dari norepinefrin
dan epinefrin, tetapi dopamin sendiri berfungsi sebagai neurotran smitter
kunci dalam hipotalamus dan pituitari. Suatu konsep yang paling
bermanfaat adalah untuk melihat inti arkuata sebagai pusat aksi, dengan
melepaskan GnRH kedalam sirkulasi porta secara pulsatil.
Seperti GnRH, gonadotropin juga disekresikan secara pulsatil, dan
memang, pola pulsatil dari pelepasan gonadotropin mencerminkan pola
pulsatil GnRH. Sekresi GnRH dan gonadotropin selalu bersifat pulsatil,
11

tetapi perbaikan pola pulsatil dari sekresi gonadotropin terjadi sesaat
sebelum pubertas disertai dengan peningkatan LH pada malam hari. setelah
pubertas, perbaikan sekresi pulsatil dipertahankan selama masa 24 jam,
tetapi amplitudo dan sekresinyabervariasi. Pada pubertas, aktivitas arkuata
dimulai dengan pelepasan GnRH berfrekuensi rendah dan berlanjut melalui
suatu siklus percepatan frekuensi, yang ditandai oleh pasase dari inaktivitas
relatif, menjadi aktivasi nokturnal, menjadi pola dewasa penuh. Perubahan
progresif dalam FSH dan LH mencerminkan aktivasi sekresi pulsatil GnRH.
Pelepasan steroid ovarium juga bersifat pulsatil, berkoordinasi dengan
pulsasi LH, stimulator utama steroidogenesis dalam ovarium. penggunaan
FSH untuk tujuan ini). Karakteristik pulsasi LH (dan mungkin juga
karakteristik pulsasi GnRH) selama siklus menstruasi adalah sebagai
berikut:
Amplitudo Rata-rata Pulsasi LH:
Fase folikuler awal 6,5 IU/L.
Pertengahan fase folikuler 5,0 IU/L
Fase folikuler lanjut 7,2 IU/L
Fase luteal awal 15,0 IU/L
Pertengahan fase luteal 12,2 IU/L
Fase luteal lanjut 8,0 IU/L
Frekuensi Rata-rata Pulsasi LH:
Fase folikuler awal 90 menit.
Fase folikuler lanjut 60-70 menit.
Fase luteal awal 100 menit.
Fase luteal lanjut 200 menit.
Sekresi pulsatil lebih sering terjadi selama fase folikuler tetapi
amplitudonya lebih rendah dibandingkan dengan fase luteal. Melambatnya
frekuensi pulsasi GnRH pada fase luteal lanjut merupakan suatu perubahan
yang penting, yang mendorong sintesis dan sekresi FSH dan karena itu,
memungkinkan peningkatan FSH yang penting untuk siklus berikutnya.
Harus ditekankan bahwa angka-angka ini bukan tidak dapat berubah.
12

Terdapat variabilitas yang cukup besar antar dan intra individu, dan
terdapat kisaran normal yang luas. Walaupun terdapat kerugian karena
waktu paruhnya yang panjang, telah dipastikan bahwa sekresi FSH
berkorelasi dengan sekresi LH. Perubahan dalam amplitudo relatif kecil;
karena itu, meningkat dan menurunnya kadar gonadotropin dalam sirkulasi
kebanyakan dipengaruhi oleh perubahan frekuensi pulsasi. Selama transisi
fase luteal-folikuler, frekuensi pulsasi meningkat kurang lebih 4-5 kali lipat.
Gonadotropin oleh pituitari anterior, karenanya efek ini dimediasi
melalui pelepasan GnRH dalam hipotalamus. Dopamin langsung
disekresikan kedalam darah porta, sehingga akan berperilaku seperti
neurohormon. Karena itu, dopamin dapat langsung menekan aktivitas
GnRH arkuata dan juga dapat dikirim melalui sistem porta untuk secara
langsung dan spesifik menekan sekresi prolaktin oleh pituitari. Jalur
dopamin tuberoinfundibuler hiporalamikus bukanlah satu- satunya jalur
dopamine dalam SSP, dan jalur ini hanyalah satu dari dua jalur dopamin
utama dalam hipotalamus. Tetapi jalur inilah yang langsung berperan dalam
reulasi sekresi prolaktin. Disamping itu, prolaktin yang dikirim ke lobus
intermediet pituitari menekan pelepasan melanocyte-stimulating hormone
Traktus Norepinefrin.
Kebanyakan badan sel yang mensintesis norepinefrin terletak dalam
mesensefalon dan batang otak bagian bawah. Sel-sel ini juga mensintesis
serotonin. Akson-akson untuk transpor amin naik kedalam jaringan
forebrain medial untuk berakhir dalam berbagai struktur otak termasuk
hipotalamus. Konsep yang ada saat ini adalah bahwa katekolamin biogenik
memodulasi pelepasn pulsatil GnRH. Norepinefrin dianggap memiliki efek
stimulatorik pada GnRH, sedangkan dopamin dan serotonin memiliki efek
inhibitorik. Untuk memahami permasalahan klinis, paling baik adalah
memandang dopamin sebagai inhibitor GnRH dan prolaktin. Namun, masih
sedikit yang diketahui mengenai peranan serotonin.
Kemungkinan kerja katekolamin adalah dengan mempengaruhi
frekuensi (dan mungkin amplitudo) pelepasan GnRH. Karena itu, faktor-
faktor farmakologis atau fisiologis yang mempengaruhi fungsi pituitari
13

mungkin mempengaruhi pituitary dengan mengubah sintesis atau
metabolisme katekolamin dan, karenanya, mengubah juga pelepasan GnRH
secara pulsatil.
B. Neuropeptida Y.
Sekresi dan ekspresi gen neuropeptida Y dalam neuron-neuron hipotalamus
diatur oleh steroid gonad. Neuropeptida Y merangsang pelepasan GnRH secara
pulsatil dan dalam pituitari akan mempotensiasi respon gonadotropin terhadap
GnRH. Karenanya, neuropeptida Y dapat memfasilitasi sekresi pulsatil GnRH
dan gonadotropin. Pada tidak adanya estrogen, neuropeptida Y menghambat
sekresi gonadotropin. Karena kekurangan nutrisi dikaitkan dengan peningkatan
neuropeptida dan peningkatan jumlah neuropeptida Y pernah dijumpai dalam
cairan serebrospinal wanita-wanita dengan anoreksia dan bulimia nervosa,
neuropeptida Y dipandang sebagai setidaknya suatu rantai penghubung antara
nutrisi dan fungsi reproduktif. Gen untuk subunit α dari gonadotropin
diekspresikan dalam pituitari maupun plasenta. Subunit β untuk human
chorionic gonadotropin (hCG) diekspresikan dalam plasenta tetapi hanya
diekspresikan secara minimal (dan dengan perubahan dalam strukturnya)
dalam pituitari, sedangkan subunit β LH, seperti yang diharapkan,
diekspresikan dalam pituitari tetapi tidak banyak diekspresikan dalam plasenta.
Reseptor-reseptor GnRH diatur oleh banyak agen, termasuk GnRH itu
sendiri, inhibin, aktivin, dan steroid seks. Penurunan respon gonadotropin
terhadap terus berlanjutnya stimulasi GnRH berlebihan dan berkepanjangan
tidak disebabkan oleh hilangnya reseptor-reseptor GnRH saja tetapi juga
meliputi desensitisasi dan uncoupling reseptor. Sintesis gonadotropin terjadi
pada retikulum endoplasma yang kasar. Hormon ini dikirim kedalam granula-
garanula sekretorik oleh sisterna Golgi dari aparat Golgi dan kemudian
disimpan sebagai granula- granula sekretorik. Sekresi memerlukan migrasi
(aktivasi) granula-garanula sekretorik matur menuju membran sel dimana
perubahan permeabilitas membran menyebabkan penonjolan granula-garanula
sekretorik sebagai respon terhadap GnRH. Langkah pembatasan kadar
14

gonadotropin dalam sintesis gonadotropin adalah ketersediaan subunit beta
dimana ketersediaan ini bergantung pada GnRH.
Pengikatan GnRH pada reseptornya dalam pituitari mengaktivasi berbagai
messenger dan respon. Hal yang segera terjadi adalah pelepasan sekretorik
gonadotropin, sedangkan respon lambat mengadakan persiapan untuk
pelepasan sekretorik berikutnya. Salah satu dari respon lambat ini adalah kerja
GnRH mematangkan diri sendiri yang menyebabkan timbulnya respon yang
lebih besar terhadap pulsasi GnRH selanjutnya karena suatu seri kejadian
biokimiawi dan biofisik intraseluler kompleks.
Kerja mematangkan diri sendiri ini penting untuk mencapai peningkatan
tinggi pada pertengahan siklus; ini memerlukan paparan estrogen, dan dapat
diperbaiki oleh progesteron. Kerja progesteron yang penting ini bergantung
pada paparan estrogen (untuk meningkatkan jumlah reseptor progesteron) dan
aktivasi reseptor progesteron oleh fosforilasi yang dirangsang oleh GnRH.
Kerja yang terakhir ini merupakan contoh komunikasi silang antara peptida dan
reseptor hormon steroid. Lima tipe sel sekretorik yang berbeda terdapat
bersama-sama dalam kelenjar pituitari anterior: gonadotrop, laktotrop, tirotrop,
somatotrop, dan kortikotrop. Interaksi autokrin dan parakrin berkombinasi
untuk menyebabkan kontrol yang lebih kompleks terhadap sekresi pituitari
daripada sekedar reaksi terhadap faktor-faktor pelepas dari hipotalamus dan
modulasi oleh sinyal-sinyal umpan balik.
Sistem GnRH merupakan mekanisme primer, peptida-peptida hipotalamus
lain dapat mempengaruhi sekresi GnRH. Peptida dapat berinteraksi dengan
GnRH pada pituitari; peptida dapat dikirim ke kelenjar pituitari dimana peptida
tersebut akan langsung mempengaruhi gonadotropin (misalnya oksitosin, CRF,
dan neuropeptida Y) atau secara tidak langsung memiliki efek pada sekresi
FSH dan LH dengan merangsang pelepasan subsansi-substansi aktif dalam
pituitari (misalnya glalanin, interleukin) dan aktivitas autokrin-parakrin
melibatkan peptida- peptida yang disintesis oleh sel-sel pituitari.
C. Sistem Autokrin-Parakrin Intrapituitari
Sitokin dan growth factor intrapituitari menyediakan suatu sistem autokrin-
parakrin untuk mengatur perkembangan dan replikasi sel pituitari maupun
15

sintesis dan sekresi hormon pituitari. Seperti pada kebanyakan jaringan,
interaksi antar substansi-substansi ini merupakan sesuatu yang kompleks, tetapi
mekanisme aktivin-inhibin perlu mendapat perhatian khusus.
D. Aktivin, Inhibin, Dan Folistatin
Aktivin dan inhibin adalah anggota-anggota peptida dari keluarga
transforming growth factor-β. Inhibin terdiri dari dua peptida yang berbeda
(dikenal sebagai subunit α dan β) yang dikaitkan oleh ikatan disulfida.
Karena itu,terdapat tiga subunit untuk inhibin: alfa, beta-A, dan beta-B.
Tiap subunit adalah produk dari RNA messenger yang berbeda; karena itu,
masing-masing subunit merupakan derivat dari molekul prekursornya masing-
masing yang berukuran besar. Inhibin disekresikan oleh sel-sel granulosa,
tetapi RNA messenger untuk rantai alfa dan beta juga telah dijumpai dalam
gonadotrop pituitari. Inhibin secara selektif menghambat sekresi FSH namun
tidak menghambat sekresi LH. Sel-sel yang aktif mensintesis LH akan
merespon kepada inhibin dengan meningkatkan jumlah reseptor GnRH, sel-sel
FSH dominan akan ditekan oleh inhibin. Inhibin memiliki efek kecil atau
bahkan tidak memiliki efek pada produksi hormon pertumbuhan, ACTH, dan
prolaktin.
Aktivin, juga berasal dari sel-sel granulosa, tetapi juga terdapat dalam
gonadotrop pituitari, mengandung dua subunit yang identik dengan subunit
beta A dan B inhibin. Aktivin memperbaiki sekresi FSH dan menghambat
respon prolaktin, ACTH, dan hormon pertumbuhan. Aktivin meningkatkan
respon pituitari terhadap GnRH dengan memperbaiki pembentukan reseptor
GnRH. Efek aktivin dapat diblokade oleh inhibin dan folistatin. Berlawanan
dengan peranan utama inhibin dalam menekan sekresi FSH, aktivin memiliki
berbagai aktivitas, yang melibatkan tulang, neuron, penyembuhan luka, dan
fungsi autokrin-parakrin pada banyak organ.
Folistatin adalah suatu peptida yang disekresi oleh berbagai sel pituitari,
termasuk gonadotrop. Peptida ini juga telah disebut sebagai protein penekan
FSH karena kerja utamanya: inhibisi sintesis dan sekresi FSH serta respon FSH
16

terhadap GnRH. Ringkasnya, GnRH merangsang sintesis dan sekresi
gonadotropin maupun aktivin, inhibin, dan folistatin. Aktivin memperbaiki
aktivitas GnRH sedangkan folistatin menekan aktivitas GnRH.
Dalam plasenta dan medulla adrenalis, pemrosesan POMC menghasilkan
peptida-peptida yang menyerupai α -MSH dan peptida-peptida β -endorfin. Β -
Endorfin juga pernah terdeteksi dalam ovarium dan testis. Dalam otak, produk
utamanya adalah opiat, dengan sedikit ACTH. Dalam hipotalamus produk
utamannya adalah β -endorfin dan α –MSH pada daerah inti arkuata dan inti
ventromedial. β-Endorfin mempengaruhi berbagai fungsi hipotalamus,
termasuk regulasi reproduksi, suhu.
Peptida Opioid dan Siklus Menstruasi Tonus opioid merupakan bagian
penting dari fungsi dan sifat siklik menstruasi. Walaupun estradiol saja sudah
meningkatkan sekresi endorfin, kadar tertinggi endorfin terjadi pada terapi
sekuensial menggunakan estradiol dan progesteron (pada monyet-monyet yang
telah menjalani ovariektomi). Karena itu, kadar endorfin endogen mengalami
peningkatan selama siklus dari kadar nadir selama menstruasi sampai kadar
tertinggi selama fase luteal. Karenanya, sifat siklik normal memerlukan
periode sekuensial aktivitas opioid hipotalamus yang tinggi (fase luteal) dan
rendah (selamamenstruasi).
Penurunan frekuensi pulsasi LH dikaitkan dengan peningkatan pelepasan
endorfin. Nalokson meningkatkan frekuensi maupun amplitudo pulsasi LH.
Karenanya, opioid endogen menghambat sekresi gonadotropin dengan
menekan pelepasan GnRH oleh hipotalamus.
Opiat tidak memiliki efek pada respon pituitari terhadap GnRH. Steroid
dari gonad memodifikasi aktivitas opioid endogen, dan umpan balik negatif
dari steroid pada gonadotropin tampaknya dimediasi oleh opiat endogen.
Karena berfluktuasinya kadar opiat endogen dalam siklus menstruasi berkaitan
dengan perubahan kadar estradiol dan progesteron, maka menarik untuk
menganggap bahwa steroid seks secara langsung merangsang aktivitas reseptor
opioid endogen. Tidak ada efek opioid pada kadar gonadotropin
postmenopause atau yang telah menjalani ooforektomisasi, dan respon
terhadap opiat dapat dikembalikan dengan pemberian estrogen, progesteron,
17

atau keduanya. Baik estrogen maupun progesteron saja dapat meningkatkan
opiat endogen, tetapi estrogen memperbaiki kerja progesteron, hal ini dapat
menjelaskan supresi maksimal terhadap GnRH dan frekuensi pulsasi
gonadotropin selama fase luteal, bukti umum menunjukkan bahwa opiat
endogen memiliki pengaruh inhibitorik pada sekresi GnRH.
Umpan balik negatif dari progesteron pada sekresi GnRH (mekanisme
utama untuk inhibisi ovulasi yang dikaitkan dengan kontrasepsi progestin) jelas
sebagian dimediasi oleh opiat endogen namun juga dimediasi oleh mekanisme
neural lain yang belum diketahui. Tonus inhibitorik opiat endogen akan
berkurang pada saat terjadinya peningkatan ovulatorik, sehingga
memungkinkan dihentikannya supresi. Ini mungkin merupakan respon
terhadap estrogen, secara spesifik terhadap penurunan pengikatan reseptor
opioid dan pelepasan opioid yang diinduksi oleh estrogen.
E. Otak Dan Ovulasi
Studi-studi klasik dalam berbagai rodent menunjukkan adanya pusat
umpan balik dalam hipotalamus yang memberi respon kepada steroid dengan
pelepasan GnRH. Pelepasan GnRH adalah akibat dari hubungan kompleks
namun terkoordinasi antara neurohormon, gonadotropin pituitari, dan steroid
gonad yang ditunjukkan oleh istilah umpan balik positif dan negatif. Kadar
FSH dianggap sangat diregulasi oleh hubungan umpan balik inhibitorik negatif
dengan estradiol. Untuk LH, baik hubungan umpan balik inhibitorik negatif
dengan estradiol maupun umpan balik stimulatorik positif dengan tingginya
kadar estradiol telah dibuktikan.
Pusat-pusat umpan balik terletak dalam hipotalamus dan disebut sebagai
pusat tonik dan siklik. Pusat tonik mengatur kadar basal harian gonadotropin
dan responsif terhadap efek umpan balik negatif steroid. Pusat siklik pada otak
wanita bertanggung-jawab untuk peningkatan gonadotropin pada pertengahan
siklus, suatu respon yang dimediasi oleh umpan balik positif estrogen. Secara
spesifik, peningkatan gonadotropin pada pertengahan siklus dianggap
disebabkan oleh berlebihannya GnRH sebagai respon kepada kerja umpan
balik positif estradiol pada pusat sikluk hipotalamus.
18

Konsep klasik ini tidak tepat. Masalahnya adalah bahwa konsep ini
secara tidak akurat menggambarkan kejadian pada rodent, tetapi
mekanismenya pada primata berbeda. Pada primata, “pusat” peningkatan
gonadotropin pada pertengahan siklus bergeser dari hipotalamus ke pituitari.
Percobaan-percobaan pada monyet menunjukkan bahwa GnRH, yang berasal
dari hipotalamus, memgang peranan permisif dan suportif. Sekresi pulsatilnya
merupakan syarat penting bagi fungsi pituitari normal, tetapi respon umpan
balik yang mengatur kadar gonadotropin dikontrol oleh umpan balik steroid
ovarium pada sel-sel pituitari anterior.
Pemberian GnRH secara pulsatil melalui pompa intravena
mengembalikan sekresi LH. Pemberian estradiol kemudian dapat menyebabkan
respon umpan balik negatif dan positif, suatu mekanisme kerja yang jelas harus
langsung terjadi pada pituitari anterior karena hipotalamus tidak ada dan GnRH
diberikan dalam frekuensi dan dosis yang tetap dan tidak berubah. Pemberian
GnRH intravena sebagai bolus menyebabkan peningkatan kadar LH dan FSH
dalam darah dalam waktu 5 menit, mencapai puncak dalam waktu sekitar 20-
30 menit untuk LH dan 45 menit untuk FSH.
Kadar LH dan FSH kembali pada kadar praterapi setelah beberapa jam.
Jika diberikan melalui infus konstan pada dosis submaksimal, pertama-tama
terjadi peningkatan cepat dan mencapai pucak dalam waktu 30 menit, diikuti
oleh pendataran atau penurunan antara 45 dan 90 menit, dan kemudian terjadi
peningkatan kedua yang dapat dipertahankan setelah 225-240 menit. Respon
bifasik ini menunjukkan adanya dua pool fungsional dari gonadotropin
pituitari.
Pool yang siap dilepaskan (sekresi) menimbulkan respon awal, dan
respon yang lebih lambat bergantung pada pool simpanan gonadotropin yang
kedua. Terdapat tiga kerja positif utama GnRH pada elaborasi gonadotropin:
1. Sintesis dan penyimpanan ( pool penyimpanan) gonadotropin.
2. Aktivasi – pergerakan gonadotropin dari pool penyimpanan ke pool yang
siap untuk sekresi langsung, mekanisme kerja pematangan diri sendiri.
3. Pelepasan segera (sekresi langsung) gonadotropin.
19

Sekresi, sintesis, dan penyimpanan mengalami perubahan selama siklus.
Pada awal siklus, saat kadar estrogen rendah, baik kadar sekresi maupun
penyimpanan juga rendah. Dengan meningkatnya kadar estradiol, terjadi
peningkatan dalam penyimpanan, disertai dengan sedikit perubahan pada
sekresi. Karena itu, pada fase folikuler awal, estrogen memiliki efek positif
pada respon sintesis dan penyimpanan, sehingga menciptakan asupan
gonadotropin yang memenuhi syarat untuk terjadinya peningkatan pada
pertengahansiklus.
Pelepasan gonadotropin yang prematur dicegah dengan kerja negatif
(inhibitorik) estradiol pada respon sekretorik pituitari terhadap GnRH. Semakin
mendekati pertengahan siklus, respon selanjutnya terhadap GnRH lebih besar
daripada respon awal, menunjukkan bahwa tiap respon tidak hanya
menginduksi pelepasan gonadotropin tetapi juga mengaktivasi pool
penyimpanan untuk respon berikutnya. Kerja sensitisasi atau pematangan oleh
GnRH ini juga melibatkan peningkatan jumlah reseptornya sendiri dan
memerluan adanya estrogen. Estrogen sendiri mampu meningkatkan jumlah
reseptor GnRH. Peningkatan estrogen pada pertengahan siklus mempersiapkan
gonadotrop untuk merespon lebih lanjut terhadap GnRH.
Peningkatan LH pada masa ovulatorik diyakini merupakan respon
terhadap kerja umpan balik positif estradiol pada pituitari anterior. Jika kadar
estradiol dalam sirkulasi mencapai konsentrasi kriitis dan konsentrasi ini
dipertahankan selama jangka waktu kritis, kerja inhibitorik pada sekresi LH
akan berubah menjadi kerja stimulatorik. Mekanisme kerja steroid ini tidak
diketahui pasti, tetapi bukti eksperimental menunjukkan bahwa kerja umpan
balik positif melibatkan banyak mekanisme, termasuk peningkatan konsentrasi
reseptor GnRH dan peningkatan sensitivitas pituitari terhadap GnRH.
Umpan balik negatif reseptor estrogen bekerja melalui sistem-sistem
yang berbeda pada tingkat pituitari, inhibisi sekresi FSH oleh estrogen
dikaitkan dengan penurunan ekspresi aktivin oleh pituitari. Disamping itu,
estradiol secara langsung menghambat gen subunit beta FSH dengan
mempengaruhi protein korepreson (protein adapter) untuk berikatan dengan
gen dan menekan transkripsi. Peningkatan pada pertengahan siklus harus
20

terjadi pada waktu yang tepat dalam siklus untuk menyebabkan ovulasi folikel
matur yang telah siap.
Adanya GnRH jelas perlu; pemberian antagnois GnRH pada wanita-
wanita pada pertengahan siklus akan mencegah peningkatan LH. GnRH
mengalami peningkatan dalam darah perifer wanita dan darah isoform FSH
dengan aktivitas biologis yang lebih besar juga meningkat selama fase luteal
lanjut, suatu perubahan yang jelas ditujukan kearah pendorongan pertumbuhan
folikel ovarium baru untuk siklus berikutnya. Peningkatan FSH pada
pertengahan siklus memiliki tujuan klinis penting. Korpus luteum normal
memerlukan induksi jumlah reseptor LH yang adekuat pada sel-sel granulosa,
suatu kerja FSH yang spesifik. Disamping itu, FSH menimbulkan perubahan-
perubahan intrafolikuler penitng yang diperlukan untuk ekspulsi fisik ovum.
Karena itu, peningkatan FSH pada pertengahan siklus memegang
peranan penting dalam memastikan ovulasi dan korpus luteum normal. Sekresi
progesteron yang mulai terjadi, segera sebelum ovulasi, adalah kuncinya.
Progesteron, pada kadar rendah dan pada adanya estrogen, memperbaiki
sekresi LH dari pituitari dan bertanggung-jawab untuk peningkatan FSH
sebagai respon terhadap GnRH. Seiring dengan terjadinya perubahan
morfologis luteinisasi dalam folikel yang berovulasi yang ditimbulkan oleh
peningkatan LH, lapisan granulosa mulai mensekresi progesteron langsung
kedalam aliran darah. Proses luteinisasi dihambat oleh adanya oosit; karena itu,
sekresi progesteron relatif tertekan, sehingga memastkan bahwa hanya sedikit
progesteron yang mencapai otak. Setelah ovulasi, luteinisasi cepat dan penuh
disertai oleh peningkatan nyata kadar progesteron, yang, pada adanya estrogen,
akan menyebabkan kerja umpan balik negatif besar untuk menekan sekresi
gonadotropin.
Kerja progesteron ini terjadi pada dua tempat, hipotalamus dan pituitari.
Jelas ada kerja pusat untuk menurunkan GnRH. Sebuah peranan penting utnuk
progesteron adalah untuk memediasi perlambatan pulsasi GnRH pada fase
luteal lanjut, dan mendorong peningkatan FSH yang diperlukan untuk memulai
siklus berikutnya. Progesteron gagal memblokade pelepasan gonadotropin
21

yang diinduksi oleh estradiol pada monyet dengan lesi hipotalamus jika
diberikan penggantian GnRH pulsatil. Karena itu, kadar progesteron yang
tinggi akan menghambat ovulasi pada tingkat hipotalamus. Sebaliknya, kerja
fasilitatorik progesteron kadar rendah hanya bekerja pada pituitary sebagai
respon terhadap GnRH.
F. Kelenjar Pineal
Walaupun tidak memiliki peran fisiologis pada manusia, fungsi
reproduktif dari hipotalamus mungkin juga dibawah kontrol penghambatan
otak melalui kelenjar pineal. Pineal muncul sebagai hasil pertumbuhan atap
ventrikel ketiga, tetapi segera setelah bayi lahir, ia kehilangan semua koneksi
neural efferent dan afferent dengan otak. Malahan sel-sel parenkim menerima
inervasi simpatetik yang luar biasa dan baru yang membiarkan kelenjar pineal
menjadi organ neuroendokrin aktif yang memberi respon tehadap stimuli
hormonal dan photic dan memperlihatkan ritme sirkardian.
Pineal adalah sumber substansi yang menghambat gonad.
Bagaimanapun, mekanisme pineal tidak essensial sama sekali untuk fungsi
gonad. Fungsi reproduksi normal kembali lagi pada tikus yang telah
dipinealektomi beberapa minggu setelah pinealektomi dilakukan; wanita buta
memiliki fertilitas normal, dan pinealektomi pada primata tidak mempengaruhi
perkembangan pubertas.
Pada manusia, sekresi melatonin meningkat setelah gelap dan
memuncak pada tengah malam, dan kemudian menurun. Ritme ini bersifat
endogen, berasal dari nucleus suprachiasmatik. Pencahayaan tidak
menyebabkan ritme, tetapi mempengaruhi waktunya. Peran yang
memungkinkan pada manusia adalah memberi ritme sirkardian pada fungsi
lainnya seperti temperatur dan tidur. Pada semua vertebrata yang diuji sejauh
ini, terdapat ritme musiman dan harian pada sekresi melatonin: nilai tinggi
selama gelap dan rendah selama terang, sekresi lebih besar pada musim dingin
dibandingkan musim panas. Desinkronisasi selama perjalanan melalui wilayah
waktu yang berbeda bisa menyebabkan gejala kompleks yang dikenal sebagai
22

jet lag. Proses pencernaan melatonin meningkatkan durasi dan kualitas tidur,
tetapi waktu optimal pemberian belum diketahui.
Pineal bekerja sebagai penghubung antara lingkungan dan fungsi
pituitary-hipotalamus. Untuk menafsirkan dengan benar lamanya hari, hewan
perlu ritme harian dalam hal sekresi melatonin. Koordinasi antara temporal dan
informasi lingkungan ini penting terutama pada peternak musiman. Ritme
pineal ini tampaknya memerlukan suprachiasmatic nucleus, mungkin pada sisi
dimana fungsi pineal dan perubahan cahaya dikoordinasikan.
Melatonin disintesa dan disekresi oleh kelenjar pineal dan bersirkulasi
dalam darah seperti hormon klasik. Ia mempengaruhi target organ yang jauh,
khususnya pusat neuroendokrin sistem saraf sentral. Apakah melatonin
disekresikan secara primer kedalam CSF atau darah, masih diperdebatkan,
tetapi bukti terbanyak adalah darah. Melatonin dari CSF bisa mencapai
hipotalamus melalui transportasi tanycyte. Perubahan gonad sehubungan
dengan melatonin diperantarai oleh hipotalamus, dan menunjukkan efek
penekanan umum terhadap sekresi pulsatile GnRH dan fungsi reproduktif.
Pada manusia, level melatonin darah tertinggi pada tahun pertama kehidupan
(dengan level tertinggi pada malam hari), kemudian menurun sesuai umur,
akhirnya hilang, beberapa menyatakan, penekanan GnRH sebelum pubertas.
Hipotesis ini ditantang oleh asosiasi wanita buta dengan usia menarche lebih
awal dari normal. Lagi pula, pinealektomi pada monyet tidak mempengaruhi
pubertas.
Aktifitas pineal dapat dipandang sebagai jaring keseimbangan antara
hormon dan pengaruh yang diperantarai neuron. Pineal mengandung reseptor
untuk hormon seks yang aktif, estradiol, testosteron, dihidrotestosteron,
progesteron, dan prolaktin. Selanjutnya, pineal mengubah tertosteron dan
progesteron menjadi metabolit 5-reduced yang aktif, dan androgen
diaromatisasi menjadi estrogen. Pineal juga terlihat unik karena
neurotransmiter katekolamin (norepinefrin) yang berinteraksi dengan reseptor
membran sel, merangsang sintesa seluler reseptor androgen dan estrogen. Pada
umumnya, aktifitas simpatetik menghasilkan ritme sirkardian yang lebih
diutamakan daripada efek hormonal. Meskipun muncul berbagai petunjuk,
23

tidak ada fakta pasti mengenai peran pineal pada manusia. Namun hubungan
penting antara pencahayaan dan ritme sirkardian berlanjut ke focus perhatian
pada kelenjar pineal sebagai koordinator. Terdapat distribusi musiman pada
konsepsi manusia di negara-negara utara dengan penurunan aktifitas ovarium
dan laju konsepsi selama musim dingin yang gelap. Lagi pula, pineal dapat
mengacaukan fungsi gonad normal.
Seorang laki-laki dengan penundaan masa pubertas yang diakibatkan
hipogonadotropin, dilaporkan memiliki kelenjar pineal yang membesar dan
hiperfungsi. Lama kelamaan level melatoninnya menurun secara spontan dan
terjadi perkembangan fungsi gonadal pituitari yang normal. Level melatonin
yang tinggi di malam hari dilaporkan pada pasien dengan amenorrhea
hipotalamus dan wanita dengan anoreksia nervosa. Pengaruh kelenjar pineal
yang tepat kemungkinan adalah sinkronisasi siklus menstruasi diantara wanita
yang menghabiskan waktu bersama. Peningkatan signifikan dari sinkronisasi
siklus diantara teman sekamar dan antara teman dekat terjadi pada 4 bulan
pertama dalam lingkungan asrama mahasiswa wanita. Peningkatan yang serupa
dalam hal sikronisasi telah diamati pada wanita teman sekerja, ditandai oleh
level ketergantungan yang sama atau lebih besar dibanding level menghadapi
tekanan pekerjaan. Bagaimanapun, usaha-usaha untuk mereplikasi hasil-hasil
ini tidak selalu berhasil.
Melatonin tersedia dalam dosis 1 – 5 mg yang menghasilkan level
darah 10-100 kali lebih tinggi dibanding puncak normal di waktu malam.
Efeknya antara lain meningkatkan rasa mengantuk dan menurunkan
kewaspadaan. Tidak tersedia data mengenai konsekuensi jangka panjang
terhadap fungsi reproduksi. Sejumlah indole lain (juga turunan tryptophan)
telah diidentifikasi pada kelenjar pineal. Peran biologis indole ini masih sukar
untuk dipahami, tetapi sebagian telah diamati. Arginine vasotocin dibedakan
dari oksitosin oleh asam amino tunggal pada posisi 8, dan dari vasopresin oleh
asam amino tunggal pada posisi 3. Pada umumnya, Arginine vasotocin
memiliki aksi penghambatan terhadap gonad dan sekresi pituitari untuk
prolaktin dan LH. Namun peran yang tepat masih belum ditemukan.
24

G. Sekresi Gonadotropin Sepanjang Kehidupan Janin, Anak-Anak, Dan Masa Pubertas
Kita sering mempertimbangkan peristiwa endokrin selama masa pubertas
sebagai suatu kesadaran, suatu awal. Bagaimanapun, secara endokrinologis,
masa pubertas bukanlah awal, tetapi hanya tahapan lain dalam perkembangan
awal suatu konsepsi. Perkembangan pituitari anterior pada manusia dimulai
antara minggu keempat dan kelima kehidupan janin, dan pada minggu ke-12
masa kehamilan hubungan vaskuler antara hipotalamus dan pituitari mulai
berfungsi. Terdapat produksi gonadotropin sepanjang kehidupan janin, selama
masa kanak-kanak, dan sampai kehidupan dewasa. Level FSH dan LH yang
luar biasa, serupa dengan level postmenopause, dapat diukur pada janin. GnRH
terdeteksi pada hipotalamus pada kehamilan 10 minggu, dan pada 10-13
minggu ketika hubungan vaskuler telah lengkap, FSH dan LH diproduksi pada
pituitari. Puncak konsentrasi pituitari untuk FSH dan LH terjadi sekitar 20-23
minggu kehidupan intrauterine, puncak level sirkulasi pada usia 28 minggu.
Peningkatan laju produksi gonadotropin sampai pertengahan kehamilan
menggambarkan kemampuan pertumbuhan poros hipotalamus-pituitari untuk
mencapai kapasitasnya secara penuh. Terdapat peningkatan sensitifitas
penghambatan oleh steroid dan penurunan sekresi gonadotropin yang dimulai
pada saat pertengahan kehamilan. Sensitifitas penuh terhadap steroid tidak
tercapai sampai akhir masa bayi. Kemunculan gonadotropin setelah melahirkan
menggambarkan hilangnya steroid plasenta dalam level yang tinggi. Jadi, pada
tahun pertama kehidupan terdapat aktifitas folikel pada ovarium yang berbeda
dengan akhir masa kanak-kanak ketika sekresi gonadotropin ditekan.
Selanjutnya, kemunculan gonadotropin postnatal lebih besar dibanding pada
bayi yang lahir prematur.
Fungsi testis pada janin dapat dihubungkan dengan pola hormon janin.
Produksi awal testosteron dan diferensiasi seksual merupakan respon terhadap
level HCG janin, mengingat produksi testosteron selanjutnya dan diferensiasi
maskulin tampak diatur oleh gonadotropin pituitari janin. Penurunan level
testosteron pada akhir masa kehamilan mungkin menggambarkan penurunan
level gonadotropin. Pembentukan janin dari sel-sel leydig entah bagaimana
25

menghindari down-regulation dan merespon terhadap level tinggi HCG dan LH
dengan meningkatkan steroidogenesis dan multiplikasi sel. Generasi sel-sel ini
digantikan oleh generasi dewasa yang menjadi fungsional pada masa pubertas
dan merespon level tinggi HCG dan LH dengan down-regulation dan
menurunkan steroidogenesis.
Terdapat perbedaan seks pada level gonadotropin janin. Terdapat
pituitari dan sirkulasi FSH serta level LH pituitari yang lebih tinggi pada fetus
perempuan. Level lebih rendah pada laki-laki mungkin akibat testosteron testis
dan produksi inhibin. Pada bayi, kemunculan FSH postnatal lebih
memperlihatkan tanda dan lebih lama pada wanita, sedangkan nilai LH tidak
setinggi itu. Aktifitas awal ini disertai oleh level inhibin yang sebanding
dengan batas rendah yang diamati selama fase folikuler pada siklus menstruasi.
Setelah postnatal, level gonadotropin mencapai titik terendah selama awal
masa kanak-kanak (sekitar usia 6 bulan pada laki-laki dan 1-2 tahun pada
wanita) dan kemudian berkembang sedikit antara 4-10 tahun. Masa kanak-
kanak ini ditandai oleh gonadotropin level rendah pada pituitari dan darah,
respon pituitari terhadap GnRH rendah, dan penekanan maksimal hipotalamus.
Signal tepat yang mengawali peristiwa pubertas tidak diketahui. Pada
perempuan, steroid pertama yang muncul dalam darah adalah
dehydroepiandrosteron (DHA) dan sulfatnya (DHAS), dimulai pada umur 6-8
tahun, sesaat sebelum FSH mulai meningkat. Level estrogen, sama dengan LH,
tidak muncul sampai usia 10-12 tahun. Jika permulaan masa pubertas dipicu
oleh hormon pertama untuk ditingkatkan dalam sirkulasi, maka peran steroid
adrenal harus dipertimbangkan. Bagaimanapun, tidak ada bukti untuk
menunjukkan bahwa steroid adrenal diperlukan untuk ketepatan waktu
pubertas, dan adrenarche terlihat tidak tergantung, tidak dikontrol oleh
mekanisme yang sama yang mengatur gonad. Juga tidak terdapat hubungan
nyata yang ditunjukkan antara sekresi melatonin dan masa pubertas. Karena
studi lebih difokuskan pada jumlah sekresi melatonin dibanding ritme sekresi,
maka pertanyaan ini masih belum terjawab.
Sebelum pubertas, level gonadotropin rendah tetapi masih berhubungan
dengan denyutan (walaupun agak tak teratur). Permulaan klinis masa pubertas
26

didahului oleh peningkatan frekuensi denyutan, amplitudo, dan keteraturan,
terutama selama malam hari. Pada saat penampakan karakteristik seks
sekunder, rata-rata level LH 2-4 kali lebih tinggi selama tidur daripada selama
terjaga. Pola ini tidak ada sebelum atau setelah masa pubertas dan merupakan
tanda awal perubahan yang terjadi di hipotalamus, dimana terdapat
peningkatan koordinasi neuron-neuron GnRH dengan meningkatkan sekresi
pulsatile GnRH. Pola ini dapat dideteksi pada individu yang mengalami
peningkatan dan penurunan derajat penekanan hipotalamus (seperti individu
dengan anoreksia nervosa yang bertambah buruk atau baik). Level FSH
distabilkan pada pertengahan pubertas, sementara level estradiol dan LH terus
meningkat sampai akhir masa pubertas. LH yang aktif secara biologis
ditemukan naik secara proporsional dibanding LH immunoreaktif pada
permulaan pubertas.
Maturasi pada masa pubertas harus melibatkan perubahan dalam
hipotalamus yang independen terhadap steroid ovarium. Perubahan maturitas
dalam hipotalamus diikuti oleh serangkaian kejadian yang teratur dan dapat
diprediksi. Peningkatan sekresi GnRH menimbulkan peningkatan responsifitas
pituitari terhadap GnRH (kombinasi steroid berpengaruh pada pituitary, dan
efek frekuensi denyutan GnRH pada jumlah reseptor GnRH), menyebabkan
peningkatan produksi dan sekresi gonadotropin. Peningkatan gonadotropin
bertanggung jawab terhadap pertumbuhan dan perkembangan folikuler dalam
ovarium serta peningkatan level steroid seks. Tingginya estrogen membantu
mencapai pola dewasa dari sekresi GnRH pulsatile, yang akhirnya
menimbulkan pola siklus menstruasi.
Kecenderungan kearah penurunan usia menarche dan periode
percepatan pertumbuhan telah berakhir. Pada studi prospektif selama 10 tahun
terhadap anak perempuan amerika yang sebaya dalam suatu kelas, rata-rata
usia menarche adalah 12,83 dengan rentang 9,14-17,70 tahun. Usia awal masa
pubertas bervariasi dan dipengaruhi oleh faktor genetik, kondisi sosial
ekonomi, dan kesehatan umum. Menarche yang lebih awal saat ini
dibandingkan dengan masa lalu, diakibatkan oleh peningkatan nutrisi dan
kesehatan yang lebih baik. Hal tersebut menunjukkan bahwa permulaan
27

pertumbuhan dan menarche terjadi pada berat badan khusus (48 kg) dan
persentase lemak tubuh (17%). Sehingga diduga bahwa hubungan ini
merefleksikan tahap metabolisme yang dibutuhkan. Walaupun hipotesa berat
kritis merupakan konsep yang berguna, variabilitas yang ekstrem pada
permulaan menarche menunjukkan bahwa tidak ada ukuran atau usia khusus
dimana seorang anak perempuan seharusnya diharapkan untuk mengalami
menarche.
Pada wanita, terjadi serangkaian kejadian khas yaitu permulaan
pertumbuhan, thelarche, pubarche, dan akhirnya menarche. Hal ini biasanya
dimulai antara usia 8 dan 14 tahun. Lamanya waktu untuk perkembangan ini
biasanya 2-4 tahun. Selama jangka waktu ini, dikatakan sebagai masa pubertas.
Tampak variasi individual yang besar pada rangkaian kejadian tersebut.
Sebagai contoh, pertumbuhan rambut pubis dan perkembangan payudara tidak
selalu berkorelasi.
Masa pubertas diakibatkan oleh reaktivasi poros hipotalamus-pituitari,
ketika sangat aktif selama kehidupan janin tetapi tertekan selama masa kanak-
kanak. Jika sistem sangat responsif, bagaimana mempertahankan fungsi
pengendalian sampai masa pubertas. Sistem gonad-pituitary-hipotalamus
bekerja sebelum masa pubertas tetapi sangat sensitive terhadap steroid, oleh
karena itu ditekan. Perubahan pada masa pubertas diakibatkan oleh
peningkatan sekresi gonadotropin secara berangsur-angsur yang terjadi karena
penurunan sensitifitas hypothalamic centers terhadap aksi inhibitori-negatif
steroid gonad. Hal ini dapat digambarkan sebagai kenaikan perlahan dari posisi
titik penurunan sensitifitas, menghasilkan peningkatan sekresi pulsatile GnRH,
yang menimbulkan peningkatan produksi gonadotropin dan stimulasi ovarium,
dan akhirnya untuk meningkatkan level estrogen. Alasan bahwa FSH
merupakan gonadotropin pertama yang dinaikkan pada masa pubertas adalah:
bahwa aktifitas arcuata dimulai dengan frekuensi rendah denyutan GnRH. Hal
ini berhubungan dengan kenaikan FSH dan sedikit perubahan pada LH.
Dengan penyesuaian frekuensi, FSH dan LH mencapai level dewasa.
Perkembangan respon umpan balik positif terhadap estrogen terjadi
belakangan. Penjelasan ini merupakan temuan terkenal dari anovulasi pada
28

bulan pertama (sepanjang 18 bulan) dari menstruasi. Namun sering ada
pengecualian dan ovulasi terjadi rata-rata pada saat menarche. Hasil perubahan
pada hipotalamus ini secara keseluruhan merupakan perkembangan
karakteristik seks sekunder, pencapaian set point level dewasa, dan
kemampuan bereproduksi.
BAB III
KESIMPULAN
Fungsi reproduksi normal memerlukan kerjasama hipotalamus hormon dan
organ target, siklus reproduksi normal membutuhkan sekresi pulsatil GnRH
dalam suatu kisaran frekuensi dan amplitudo tepat. Fisiologi dan patofisiologi
siklus tersebut setidaknya dalam kaitannya dengan kontrol pusat, dapat dijelaskan
melalui mekanisme yang mempengaruhi sekresi pulsatil GnRH. secara langsung
dengan memperngaruhi berbagai neurotransmiter.
1. Sekresi GnRH pulsatil harus berada dalam kisaran kritis untuk frekuensi dan
konsentrasi (amplitudo). Hal ini benar-benar perlu untuk fungsi reproduktif
normal.
2. GnRH hanya memiliki kerja positif pada pituitari anterior: sintesis dan
penyimpanan, aktivasi, dan sekresi gonadotropin. Gonadotropin disekresikan
dengan cara pulsatil sebagai respon terhadap pelepasan GnRH yang juga
bersifat pulsatil.
3. Frekuensi pulsasi GnRH yang lebih rendah mendorong sekresi FSH, dan
frekuensi pulsasi GnRH yang lebih tinggi mendorong sekresi LH.
4. Estrogen kadar rendah memperbaiki sintesis dan penyimpanan FSH dan LH,
memiliki sedikit efek pada sekresi LH, dan menghambat sekresi FSH.
5. Estrogen kadar tinggi menginduksi peningkatan LH pada pertengahan siklus,
dan estrogen dengan kadar tinggi dan teratur menyebabkan dipertahankannya
peningkatan sekresi LH.
6. Progesteron kadar rendah yang bekerjada pada tingkat kelenjar pituitari
memperbaiki respon LH terhadap GnRH dan bertanggung jawab untuk
peningkatan FSH pada pertengahan siklus.
29

7. Progesteron kadar tinggi menghambat sekresi gonadotropin dari pituitari
dengan menghambat pulsasi GnRH pada tingkat hipotalamus. Disamping itu,
progesteron kadar tinggi dapat mengantagonisasi respon pituitari terhadap
GnRH dengan mengganggu kerja estrogen.
DAFTAR PUSTAKA
Badziad A. 2003. Endokrinologi Ginekologi, Edisi ke-2. Jakarta
Cunningham FG, Mac Donald PC, Gant NF. 2014. Williams Obstetri. Edisi ke-23.
Jakarta. EGC
Guyton A. 2008. Fisiologi Kedokteran. Edisi ke-11. Jakarta. EGC
Heffner Linda J. 2006. At a Glance Sistem Reproduksi. Edisi ke-2. Jakarta. EGC
Jacob TZ, Baziad A. 1994. Endokrinologi Reproduksi. Edisi ke-1. Jakarta. KSERI
Sherwood L. 2012. Fisiologi Manusia, Edisi ke-6.
30






