OSTEOINMUNOLOGIA: BASES CELULARES Y · PDF fileidentificado como una molécula de...
50
OSTEOINMUNOLOGIA: BASES CELULARES Y MOLECULARES DEL REMODELAMIENTO ÓSEO ROSAHYRA MILENA SANDOVAL COTE PONTIFICIA UNIVERSIDAD JAVERIANA CIENCIAS EXACTAS Y NATURALES ESPECIALIZACIÓN EN LABORATORIO DE INMUNOLOGIA CLINICA SANTA FÉ DE BOGOTA 2005
Transcript of OSTEOINMUNOLOGIA: BASES CELULARES Y · PDF fileidentificado como una molécula de...

OSTEOINMUNOLOGIA: BASES CELULARES Y MOLECULARES DEL REMODELAMIENTO ÓSEO
ROSAHYRA MILENA SANDOVAL COTE
PONTIFICIA UNIVERSIDAD JAVERIANA CIENCIAS EXACTAS Y NATURALES
ESPECIALIZACIÓN EN LABORATORIO DE INMUNOLOGIA CLINICA SANTA FÉ DE BOGOTA
2005

OSTEOINMUNOLOGIA: BASES CELULARES Y MOLECULARES DEL REMODELAMIENTO ÓSEO
ROSAHYRA MILENA SANDOVAL COTE
Monografía
Director, Dr. EDGAR GARAVITO Profesor Especialización en Laboratorio de Inmunología Clínica
PONTIFICIA UNIVERSIDAD JAVERIANA CIENCIAS EXACTAS Y NATURALES
ESPECIALIZACIÓN EN LABORATORIO DE INMUNOLOGIA CLINICA SANTA FÉ DE BOGOTA
2005

Nota de Aceptación: ______________________________ ______________________________ ______________________________ ______________________________
_____________________________ Director, Dr. EDGAR GARAVITO
Santa Fe de Bogota Noviembre 15 de 2005

CONTENIDO
Pág.
INTRODUCCIÓN 13
1. CELULAS ÓSEAS 15
1.1 OSTEOBLASTOS 15
1.2 OSTEOCLASTOS 17
2. REMODELAMIENTO ÓSEO 19
3. REGULACIÓN DE LA HOMEOSTASIS ÓSEA 20
4. VIAS DE SEÑALIZACIÓN CELULAR INVOLUCRADAS EN
OSTEOCLASTOGÉNESIS 21
4.1 TRANCE 22
4.2 TRANCE – R 25
4.3 OPG 27
4.4 PTH/VITAMINA D3/CALCITONINA 29
5. SISTEMA INMUNE EN OSTEOCLASTOGÉNESIS 31

6. UTILIDAD CLINICA DE LOS MARCADORES DE
REMODELAMIENTO ÓSEO 35
6.1 TIPOS DE MARCADORES 38
6.1.1 Marcadores de Formación Ósea 38
6.1.1.1 Fosfatasa Alcalina Total y Fosfatasa Alcalina Ósea 38
6.1.1.2 Osteocalcina 38
6.1.1.3 Propenopéptidos Amino y Carboxiterminal del Procolágeno I 38
6.1.2 Marcadores de Resorción Ósea 39
6.1.2.1 Hidroxiprolina 39
6.1.2.2 Puentes de Piridinolina y Deoxipiridinolina 39
6.1.2.3 Péptidos Terminales o Telopéptidos del Colágeno 39
6.1.2.4 Fosfatasa Ácida Resistente a Tartrato 40
5. CONCLUSIONES 41
BIBLIOGRAFIA 42

LISTA DE TABLAS
pág.
Tabla 1 Moléculas que regulan los niveles de TRANCE y OPG 24
Tabla 2 Marcadores de Remodelamiento Óseo 37

LISTA DE FIGURAS
pág.
Figura 1 Diferenciación de células óseas 16
Figura 2 Interacción RANK – RANKL 29
Figura 3 Papel de la célula T activada en la formación y diferenciación
de osteoclastos 32

RESUMEN
El hueso es un tejido dinámico que brinda un soporte mecánico, protección
física y la capacidad de movimiento. Además el hueso es el sitio de
almacenamiento para diferentes minerales, y es allí donde se producen las
células sanguíneas. La masa ósea depende del balance entre síntesis y
destrucción del hueso mediada por osteoblastos y osteoclastos. Los
osteoblastos se derivan de células primitivas mesenquimales por acción del
factor de crecimiento trasformante β (TGF-β), el factor de crecimiento de
fibroblastos (FGF) y proteínas morfogenéticas óseas (BMPs). Los
osteoclastos se originan de precursores hematopoyéticos y en su
diferenciación participan el GM-CSF (Factor estimulador de colonias
granulocito-monocito) y el M-CSF (factor estimulador de colonias manocito-
macrófago). Diferentes estudios han establecido la relación entre
osteoblastos y sus células precursoras estromales con los precursores
hematopoyeticos de los osteoclastos y los mediadores solubles que modulan
estas interacciones incluyendo el factor de diferenciación de los osteoclastos
ODF o TRANCE y la Osteoprotegerina (OPG). Factores solubles como la
hormona paratiroidea (PTH), la vitamina D3 y la calcitonina también son
críticos en el mantenimiento de este balance. El hueso es un tejido
metabolicamente muy activo; mediante la cuantificación en el suero y en la
orina de moléculas derivadas del hueso, se ha logrado detectar trastornos en
el recambio óseo, estas moléculas son productos que resultan de la actividad
enzimática de las células durante la formación o la resorción, o son también
componentes de la matriz ósea liberados durante la síntesis o la degradación
del hueso.

DEFINICIÓN DEL PROBLEMA
La morfogénesis de los huesos y su modelación dependen de la producción
de matriz ósea mediada por los osteoblastos y su resorción por los
osteoclastos. Los osteoblastos se derivan de una célula precursora
mesenquimal pluripotencial que se diferencia en unidades formadoras de
colonias fibroblastos (CFU-F). La diferenciación ocurre en un microambiente
donde participan hormonas, factores de crecimiento y citocinas (Baylink,
1993). Entre las citocinas las más importantes son el factor de crecimiento
trasformante beta (TGF-β), el cual estimula la quimiotaxis de precursores de
osteoblastos (Alliston, 2001), y el factor de crecimiento de fibroblastos (FGF),
el cual activa al osteoblasto para favorecer la formación de hueso (Yu, 2003).
Otras moléculas de importancia que modulan este proceso son las proteínas
morfogenéticas óseas (BMPs) (Canalis, 2003), las cuales participan en el
crecimiento esquelético durante la embriogénesis y la reparación de las
fracturas (Canalis, 2003). A medida que el hueso se mineraliza el osteoblasto
deja de tener actividad de síntesis de material osteoide y se convierte en
osteocito, el cual va quedando incluido en el hueso rígido y empieza a tener
funciones de receptor de fuerzas externas principalmente determinadas por
la actividad muscular (Mackie, 2003).
Los osteoclastos son las células más importantes en el proceso de resorción
ósea y se caracterizan por su gran tamaño y por sus múltiples núcleos
resultantes de la fusión de células precursoras (Boyle, 2003 y Miyamoto,
2003). Se originan a partir de precursores hamatopoyéticos de la línea que
da lugar a los monocitos/macrófagos. Al igual que los osteoblastos, el
reclutamiento y diferenciación de los osteoclastos está mediado por
hormonas, factores de crecimiento y citocinas. Dentro de los factores que
están involucrados en el reclutamiento de las células precursoras de los

osteoclastos figuran el GM-CSF (factor estimulante de colonias de
granulocitos y monocitos) (Sarma, 1996) y el M-CSF (factor estimulante de
colonias de macrófagos). El GM-CSF induce resorción ósea y la formación
de osteoclastos, mientras que el M-CSF actúa sobre la célula madre del
sistema hematopoyético para mantener la sobrevida de los monocitos
precursores de osteoclastos (Sarma, 1996).
En años recientes se ha avanzado significativamente en el entendimiento de
los mecanismos celulares y moleculares que regulan el remodelamiento
óseo. Estos estudios han permitido establecer la relación entre los
osteoblastos y sus células precursoras estromales, con los precursores
hematopoyéticos de los osteoclastos y los mediadores solubles que modulan
estas interacciones. Está relación fue establecida claramente en estudios
realizados in vitro e in vivo, los cuales demostraron que era necesaria la
presencia de osteoblstos/células estromales y su interacción con los
precursores de los osteoclastos, para que se produjera la diferenciación y
activación de estos últimos.
El remodelamiento óseo es un proceso dinámico que ocurre en diferentes
proporciones a lo largo de la vida. Este proceso se coloca de manifiesto en el
momento de reparar microfracturas u otros daños que afecten el esqueleto.
Además, como los huesos son el lugar de almacenamiento de calcio, este
proceso por consiguiente regula las concentraciones de este mineral en el
rango necesario para mantener su equilibrio. Este proceso se refleja en la
liberación de una serie de moléculas, las cuales pueden ser medidas en
suero o sangre, que sirven como marcadores bioquímicos de la formación y
resorción ósea. Entre estos están los telopéptidos del procolágeno tipo I; el
precursor del constituyente más abundante de la matriz ósea, el colágeno I
(Delmas, 2000).

JUSTIFICACIÓN
En los últimos años se ha demostrado la estrecha relación que existe entre el
Sistema Inmune y el Tejido Óseo, en particular como las células de ambos
tejidos se originan a partir de precursores comunes y a su vez como las
citocinas producidas por diferentes células tienen efectos regulatorios
importantes sobre células tanto del sistema inmunitario como óseas.
Por lo anterior es importante revisar y analizar los mecanismos celulares y
moleculares involucrados en el remodelamiento óseo. Con está información
se espera tener un mejor entendimiento de la patogénesis de las alteraciones
óseas, su relación con la Inmunología y valorar la utilidad potencial que
puedan tener los diferentes marcadores biológicos de la remodelación ósea
en el diagnostico y seguimiento de dichas alteraciones.

OBJETIVO
Analizar los mecanismos celulares y moleculares involucrados en el
remodelamiento óseo, haciendo énfasis en los marcadores con potencial
utilidad clínica.

INTRODUCCIÓN El termino Osteoinmunología ha sido propuesto para describir la
interdisciplinariedad que existe entre el sistema Inmune y el Sistema
Esquelético (Arron, 2000). Recientemente se ha observado que tanto el
sistema inmune y esquelético se regulan el uno al otro en mucho mas grado
que lo que previamente se pensaba (Rho, 2004). En particular varias
condiciones patológicas las cuales se acompañan de una excesiva perdida
ósea, tales como artritis reumatoidea, enfermedad periodontal y algunas
anormalidades óseas asociadas a tumores muestras estar influenciadas por
componentes celulares (por ejemplo linfocitos T) y por factores solubles
(interferón) (Rho, 2004).
El papel de las células T en la biología ósea ha sido ampliamente aceptado
desde el descubrimiento de miembros de la familia del factor de necrosis
tumoral (TNF), como la citocina de activación relacionada con el TNF
(TRANCE), también conocida como ligando para el receptor de activación de
NF-κB (RANKL), ligando para osteoprotegerina (OPGL) y factor de
diferenciación de osteoclastos (ODF) (Rho, 2004). TRANCE fue inicialmente
identificado como una molécula de expresión en células T activadas (Wong
1997). Posteriormente se demostró que TRANCE es expresado sobre
osteoblastos activos y que es esencial para la osteoclastogenesis (Wong,
1999). Los receptores y ligandos en esta superfamilia tienen una estructura
única que les permite acoplarse directamente a vías de señalización para
proliferación celular, sobrevida y diferenciación (Theill, 2002).
Los miembros de la familia del TNF son dos proteínas trasmembrana que se
ensamblan en trímeros funcionales. El receptor de TNF (TNFR) es una
13

glicoproteína trasmembrana tipo 1 que presenta dominios extracelulares ricos
en cisteína. Existen dos subgrupos de TNFR homólogos, el primero contiene
dominios extracelulares de muerte que se une a TRADD o FADD los cuales
permiten la activación de la caspasa 8 e inducen apoptosis. El otro grupo se
une a factores asociados al receptor de TNF (TRAFs) los cuales son
moléculas adaptadoras que permiten la señalización corriente abajo,
permitiendo la activación de JNK y NF-κB promoviendo el crecimiento y
sobrevida celular (Theill, 2002).
De otra parte las células del sistema inmune son derivadas a partir de
células progenitoras de la medula ósea. Las citocinas producidas por estas
células regulan el metabolismo óseo. Por consiguiente el papel del sistema
inmune en la homeostasis ósea ha generado gran interés.
14

1. CELULAS ÓSEAS
La formación de los huesos y su modelación dependen de la producción de
matriz ósea mediada por osteoblastos y su resorción por los osteoclastos.
1.1 OSTEOBLASTOS
Los osteoblastos son células de origen mesodérmico cuya función primaria
es la formación de hueso (Figura 1). Los osteoblastos se derivan de una
célula precursora mesenquimal pluripotencial que se diferencia en unidades
formadoras de colonias de fibroblastos (CFU-F). La diferenciación ocurre en
un microambiente donde participan hormonas, factores de crecimiento y
citocinas (BaylinK, 1993). Entre las citocinas las más importantes son el
factor de crecimiento trasformante beta (TGF-β), el cual estimula la
quimiotaxis de precursores de los osteoblastos (Alliston, 2001) y el factor de
crecimiento de fibroblastos (FGF) el cual activa el osteoblasto para favorecer
la formación de hueso (Yu, 2003). Otras moléculas de importancia que
modulan este proceso son las proteínas morfogenéticas óseas (BMPs)
(Canalis, 2003), las cuales participan en el crecimiento esquelético durante la
embriogénesis y la reparación de fracturas (Canalis, 2003). A medida que el
hueso se mineraliza el osteoblasto deja de tener actividad de síntesis de
material osteoide, se rodea de matriz ósea y se convierte en osteocito que
adquiere la capacidad para responder a fuerzas externas principalmente
determinadas por la actividad muscular (Mackie, 2003). Una vez los
osteocitos son atrapados pierden su habilidad para producir matriz ósea. Sin
embargo los osteocitos pueden sintetizar ciertas moléculas de la matriz ósea
como la osteocalcina, y depositar minerales en la matriz ósea, además
forman una red con los osteocitos adyacentes y los osteoblastos de la
superficie ósea a través de prolongamientos celulares que viajan en el
15
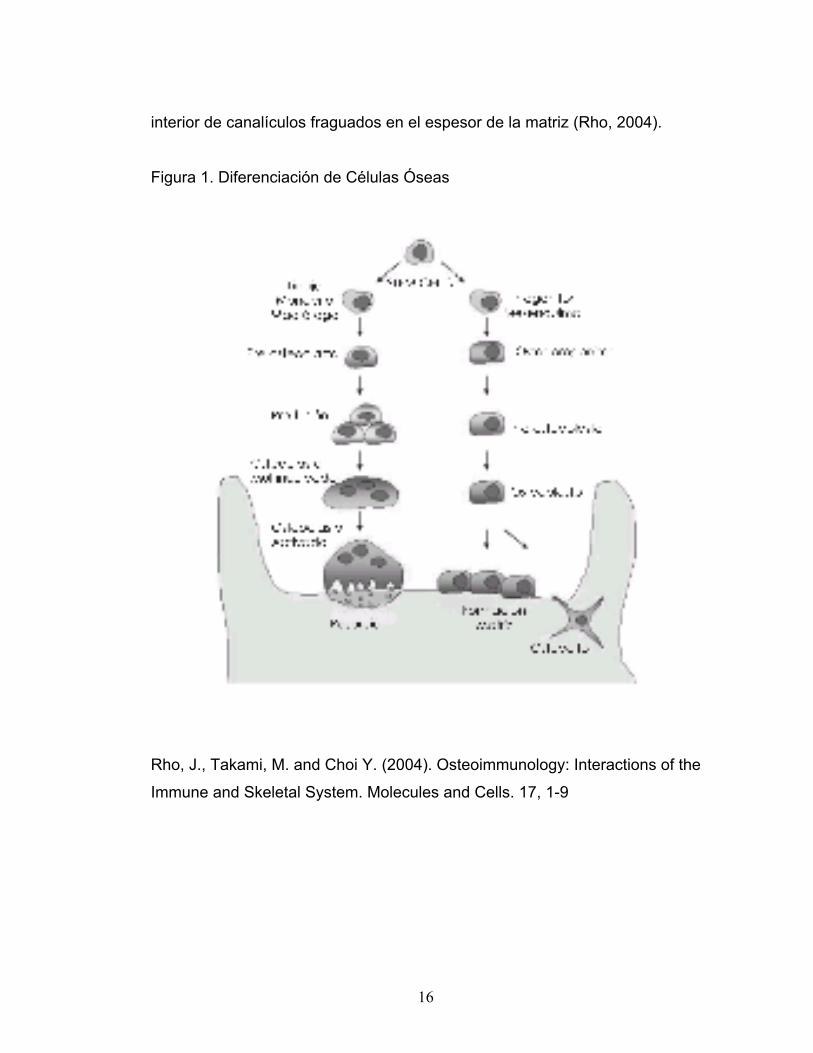
interior de canalículos fraguados en el espesor de la matriz (Rho, 2004).
Figura 1. Diferenciación de Células Óseas
Rho, J., Takami, M. and Choi Y. (2004). Osteoimmunology: Interactions of the
Immune and Skeletal System. Molecules and Cells. 17, 1-9
16

1.1 OSTEOCLASTOS
Los osteoclastos difieren dramáticamente de los osteoblastos, se
caracterizan por su gran tamaño y por sus múltiples núcleos resultantes de la
fusión de las células precursoras (Boyle, 2003). Se originan a partir de
precursores hematopoyeticos de la línea monocito/macrófago (Figura 1). Al
igual que los osteoblastos, el reclutamiento y diferenciación de los
osteoclastos esta mediado por hormonas, factores de crecimiento y citocinas.
Dentro de los factores que regulan el proceso de diferenciación de los
osteoclastos figuran el factor estimulante de colonias granulocito/monocito
(GM-CSF) (Sarma, 1996) y el factor estimulante de colonias de macrófagos
(M-CSF). El GM-CSF induce resorción ósea y la formación de osteoclastos,
mientras que el M-CSF actúa sobre la célula madre del sistema
hematopoyético para mantener la sobrevida de los monocitos precursores de
osteoclastos (Sarma, 1996).
Los osteoclastos resorben matriz ósea mineralizada modulando la formación
ósea y el crecimiento. La cantidad de osteoclastos en la matriz ósea es
relativamente pobre, de alrededor de 2 a 3 por µm3, con una vida media de 8
a 7 semanas (Suda, 1997). Se localizan sobre la superficie ósea, alojados en
lagunas de resorción del hueso trabecular o en túneles cilíndricos del hueso
cortical. Las lagunas de resorción y los túneles de estas áreas son creadas
por la propia actividad del osteoclasto (Suda, 1997).
Los osteoclastos poseen diferentes características que lo hacen células
particulares como son su forma discoide con borde irregular y zonas claras.
El borde rugoso contiene elementos que resorben componentes orgánicos e
inorgánicos del hueso. Los osteoclastos secretan enzimas, tales como la
fosfatasa ácida resistente a tartrato (TRAP) por consiguiente decalcifican el
hueso, y otras enzimas tales como catepsina K, que degradan proteínas de
17

la matriz ósea (Roodman, 1996). Los osteoclastos pueden liberar protones
directamente vía H+ATPasa localizada en el borde irregular. El resultado de
la acidificación favorece la disolución del mineral óseo en las áreas de
resorción (Zaide, 2003). Varias enzimas proteolíticas tales como enzimas
lisosomales y proteasas de cisteína, son también liberadas a través del borde
irregular en las zonas de resorción (Rho, 2004).
Los osteoclastos expresan además altos niveles de αvβ3 integrina, receptor
para la vitronectina (Helfrich, 1999), que le permite a los osteoclastos
adherirse a una variedad de constituyentes de la matriz ósea in vitro, pero su
papel en la función de los osteoclastos in vivo aun no es claro. Se ha
observado altos niveles de expresión de αvβ3 integrina en el borde rugoso.
Este receptor es necesario para la adhesión firme y progresiva a la matriz en
la medida en que el osteoclasto excava la matriz ósea durante la resorción
(Helfrich, 1999).
18

2. REMODELAMIENTO ÓSEO
La morfogénesis y remodelamiento óseo implican la síntesis de matriz ósea
por parte del osteoblasto y la coordinada resorción ósea por el osteoclasto.
Se estima que alrededor del 10% del total de masa ósea en humanos es
recambiada anualmente. Un imbalance entre la actividad de los osteoclastos
y osteoblastos puede resultar de cambios hormonales o perturbaciones
inflamatorias, resultando en anormalidades esqueléticas caracterizadas por
disminución (osteoporosis) o incremento (osteopetrosis) de la masa ósea
(Theill, 2002).
El incremento de la actividad osteoclastica es observado en muchos
desordenes osteopenicos, incluyendo osteoporosis posmenopáusica,
enfermedad de Paget, metástasis ósea lítica o artritis reumatoidea; en los
cuales hay un incremento en la resorción ósea. Además existen varios
factores como M-CSF, IL-1, TGF-β, TGF-α, TNF-β, IL-6 Vitamina D3,
calcitonina, prostaglandina E2 (PGE2), hormona paratiroidea (PTH), los
cuales pueden afectar la osteoclastogenesis a diferente nivel. Sin embargo
observaciones experimentales han mostrado que estos factores no son
esenciales para el desarrollo de osteoclastos in vivo (Theill, 2002).
19

3. REGULACIÓN DE LA HOMEOSTASIS ÓSEA La homeostasis ósea es regulada por el balance de la función de
osteoblastos y osteoclastos. La formación ósea por osteoblastos y la
resorción ósea por osteoclastos están firmemente acopladas para el
manteniemiento de la salud ósea. La actividad de formación ósea por
osteoblastos incrementa la masa ósea, mientras la actividad de resorción
ósea de los osteoclastos reduce la masa ósea (Suda, 2001). La acción de
estas células esta regulada por factores de crecimiento y citocinas. La
alteración de este balance generalmente lleva a enfermedades óseas, tales
como osteoporosis y osteopetrosis (Helfrich, 2003).
La osteoporosis es una enfermedad metabólica en la cual la masa ósea y la
densidad mineral ósea continuamente disminuye debido a una incrementada
actividad de los osteoclastos. La osteoporosis guía a un alto riesgo de
fracturas óseas. De forma diferente, la osteopetrosis hace referencia a una
condición heterogénea generalmente caracterizada por incremento de la
densidad ósea a causa de un daño en la resorción ósea. En condiciones
normales, la resorción ósea por los osteoclastos, en sitios específicos, activa
los osteoblastos para remplazar el hueso perdido; cuando este balance se
altera se presenta alguna de las enfermedades mencionadas anteriormente.
Este disbalance puede resultar de cambios relacionados con las hormonas,
citocinas, nutrición, metabolismo, factores mecánicos, o una combinación de
cualquiera o de todos estos factores (Rho, 2004).
20

4. VIAS DE SEÑALIZACIÓN CELULAR INVOLUCRADAS EN
OSTEOCLASTOGENESIS
Los osteoblastos que regulan el metabolismo óseo están relacionados con
las hormonas osteotrópicas, con las citocinas que regulan la formación ósea
y además, con el desarrollo de osteoclastos y la activación de la resorción
ósea (Rho, 2004). Los osteoclastos son derivados del linaje monocito-
macrófago en respuesta a las acciones coordinadas de diferentes hormonas
osteotropicas incluyendo la vitamina D3 (1,25(OH)2D3) y hormona
paratiroidea (PTH). Estas hormonas han sido identificadas en estudios
genéticos de enfermedades que causan anormalidades esqueléticas en
ratones, ratas y humanos (Zelzer, 2003).
Ratones con una disrupción en el gen del M-CSF (ratones op/op) exhiben un
fenotipo osteopetropico (Yoshida, 1990). Estudios en estos ratones han
claramente mostrado que el M-CSF juega un importante papel en
osteoclastogenesis. M-CSF es requerido para diferenciación de los
osteoclastos. Los osteoblastos derivados de ratones op/op no favorecen la
formación de osteoclastos in vitro (Suda, 1997). Adicionalmente se ha
observado que el M-CSF participa en el mantenimiento de osteoclastos
maduros. Esto sugiere que el M-CSF participa en los estados tempranos de
diferenciación de los osteoclastos así como en la movilidad y sobrevida de
osteoclastos maduros. Ahora bien, el factor de crecimiento endotelial
vascular recombinante humano (VEGF) puede inducir formación de
osteoclasto en ratones op/op (Kaku, 2001). El ligando para FLT3 también
sustituye al M-CSF para inducir el desarrollo de osteoclastos bajo
condiciones especificas (Lean, 2001).
21

4.1 TRANCE En 1997, varios grupos de investigadores estudiando moléculas de unión en
una línea de células estromales identificaron una molécula que favorecía la
osteoclastogenesis, por lo cual recibió el nombre de ODF (del ingles factor de
diferenciación de osteoclastos) (Tsuda, 1997). Posteriormente en una librería
de cADN de una línea celular mielomonocitica de ratón 32D se aisló una
molécula que se unía a la osteoprotegerina (OPG) y se le denomino ligando
de la OPG (OPGL), la cual resulto ser idéntica al ODF (Suda, 1999). Mas
aun, otro factor de la familia del ligando del TNF fue clonado en forma casi
simultanea durante la búsqueda de genes que regulan la apoptosis en
hibridomas de células T de ratón (Tsuda, 1997); este factor se denomina
citocina inducida por activación relacionada con el TRANCE (Tsuda, 1997).
Otros investigadores identificaron un factor implicado en la diferenciación de
los monocitos y células dendríticas por clonación de una librería de cADN de
células dendríticas humanas, y lo denominaron RANKL (Tsuda, 1997). Por
consiguiente las moléculas ODF, OPGL, TRANCE resultaron ser la misma
que el RANKL.
TRANCE es expresado en la superficie de osteoblastos y células epiteliales
de glándulas mamarias y en células T activadas (Rho, 2004). La expresión
de TRANCE es inducida en respuesta a citocinas osteotrópicas secretadas
por células inmunes, incluyendo interleucina – 1 (IL-1), IL-6 e IL-11 (Walsh,
2003). Ratones deficientes en TRANCE exhiben un fenotipo similar a
osteopetrosis y no desarrollan nódulos linfoides y glándulas mamarias (Theill,
2002).
Una combinación de TRANCE y M-CSF induce formación de osteoclastos a
partir de monocitos derivados de bazo y medula ósea en ausencia de
osteoblastos (Kim, 2002). Aunque el M-CSF ha sido identificado como un
22

factor crucial para el desarrollo de osteoclastos no lo es de forma única. Esto
sugiere que TRANCE debe estar participando en la diferenciación y
formación de los osteoclastos además de su intervención en la formación de
lagunas de resorción ósea (Rho, 2004).
TRANCE regula la comunicación entre célula T y célula dendrítica, la
sobrevida de la célula dendrítica y organogénesis de nodos linfoides (Theill,
2002). La producción de TRANCE por células T activadas puede
directamente regular, como ya se menciono, la osteoclastogénesis y el
remodelamiento óseo. Esto explica porque las enfermedades autoinmunes,
el cáncer, leucemias, asma, infecciones virales crónicas y enfermedades
periodontales resultan en una perdida ósea local y sistémica (Theill, 2002).
La expresión de TRANCE puede ser aumentada por factores de resorción
ósea tales como glucorticoides, vitamina D3, IL-1, IL-6, IL11, IL17, TNF-α,
PGE2 y PTH (Tabla 1). Usando sistemas de cultivos in vitro se ha observado
que TRANCE puede activar osteoclastos maduros y mediar
osteoclastogénesis en presencia de M-CSF. Ratones trance-/- presentan
osteopetrosis severa, crecimiento atrofiado y defectos en la erupción de los
dientes, además sus osteoblastos no participan en la osteoclastogénesis. Sin
embargo estos ratones contienen precursores hematopoyéticos que pueden
diferenciarse en osteoclastos maduros in vitro en presencia de TRANCE
recombinante y M-CSF (Theill, 2002).
23
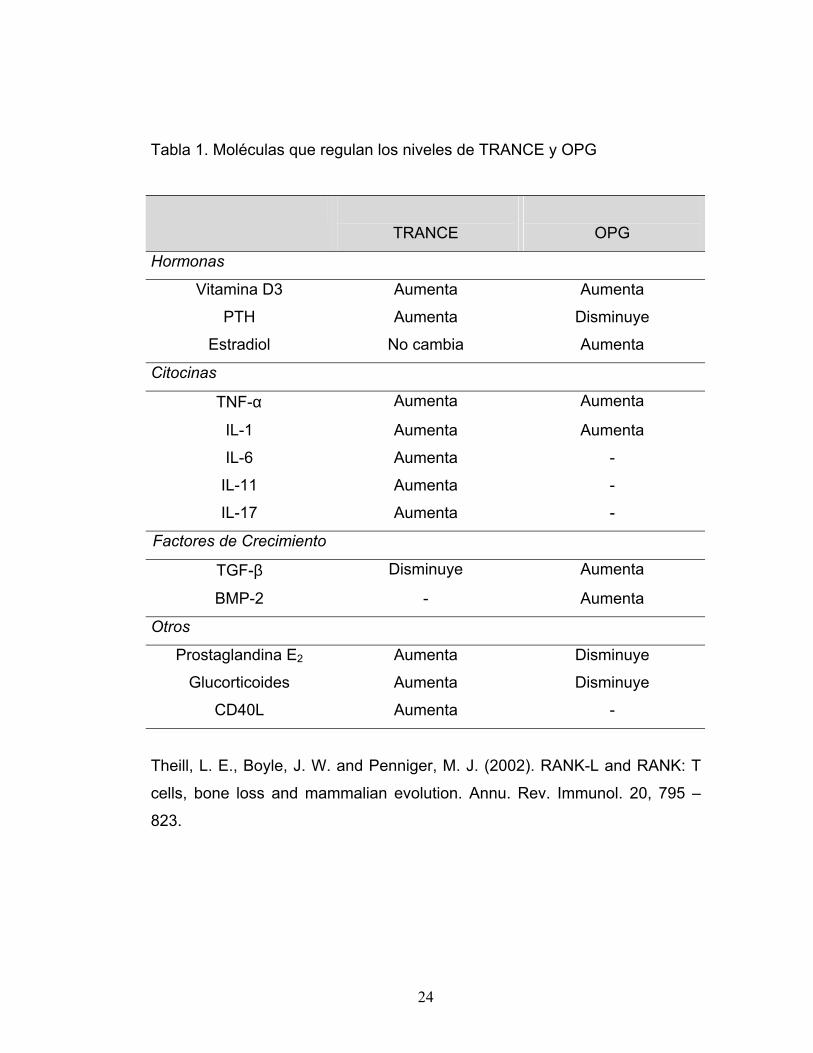
Tabla 1. Moléculas que regulan los niveles de TRANCE y OPG
TRANCE
OPG
Hormonas
Vitamina D3 Aumenta Aumenta
PTH Aumenta Disminuye
Estradiol No cambia Aumenta
Citocinas
TNF-α Aumenta Aumenta
IL-1 Aumenta Aumenta
IL-6 Aumenta -
IL-11 Aumenta -
IL-17 Aumenta -
Factores de Crecimiento
TGF-β Disminuye Aumenta
BMP-2 - Aumenta
Otros
Prostaglandina E2 Aumenta Disminuye
Glucorticoides Aumenta Disminuye
CD40L Aumenta -
Theill, L. E., Boyle, J. W. and Penniger, M. J. (2002). RANK-L and RANK: T
cells, bone loss and mammalian evolution. Annu. Rev. Immunol. 20, 795 –
823.
24

4.2 TRANCE – R El receptor de TRANCE, también llamado TRANCE-R, RANK, OPGLR,
ODFR, es expresado como un heterotrímero trasmembrana en progenitores
de osteoclastos, células B, células dendríticas y células epiteliales de
glándula mamaria. TRANCE-R es un miembro de la superfamilia de
receptores del TNF y presenta alta homología con CD40. TRANCE-R es una
proteína trasmembrana con una cola citoplasmática de alrededor de 383
aminoácidos que contienen tres dominios de unión (I, II, III) (Walsh, 2003).
In vitro la unión de TRANCE – TRANCE-R resulta en osteoclastogénesis de
células progenitoras y la activación de osteoclastos maduros. La activación
de TRANCE-R depende de la señal dada por TRANCE la cual es mediada
por una molécula adaptadora denominada factor asociado al receptor para
TNF (TRAFs) de las cuales se conocen alrededor de seis (TRAF 1-6). TRAF
1, 2, 3, 5 y 6 interactúan con la cola citoplasmática de TRANCE en regiones
específicas (Darnay 1999). Aunque cinco diferentes TRAFs interactúan con
la cola citoplasmática de TRANCE-R, la señalización mediada por TRAF6 en
particular parece ser crucial para la formación de osteoclastos. Ratones
deficientes en TRAF6 presentan defectos en la formación de osteoclastos y
muestran osteopetrosis (Lomaga, 1999).
NFκ-B es una molécula de señalización y un factor de activación que actúa
corriente abajo de TRAF6. La señal de activación de NFκ-B está
completamente bloqueada en ratones deficientes de TRAF6. Sin embargo se
ha observado que la activación de c-Jun kinasa solo esta parcialmente
disminuida en ratones deficientes de TRAF6 ya que TRAF2 puede mediar
esta señalización (Lomaga, 1999). Estas observaciones son muy similares a
las vistas en ratones deficientes de TRANCE-R (Dougall, 1999). Ratones
25

deficientes de NFκ-B muestran un fenotipo de osteopetrosis y presentan
defectos en la formación de osteoclastos (Iotsova, 1997).
Otros estudios han mostrado que TRANCE activa la serin-treonin-kinasa
antiapoptotica Akt/PKB a través del complejo de señalización c-Src y TRAF6
e induce la cascada de señalización a través de TAB2/Tak1 (Lee, 2002). c-
Src y TRAF6 interactúan el uno con el otro y con TRANCE-R, deficiencias en
c-Src o la adición de inhibidores de kinasas de la familia Src bloquean la
activación de Akt/PKB en osteoclastos. TRAF6 aumenta la actividad de c-Src
guiando a una fosforilación corriente abajo de moléculas de señalización
tales como c-Cb1 (Theill, 2002).
Con base en los datos presentados se puede afirmar que TRAF6 juega un
papel importante en la señalización dada por la interacción de TRANCE-
TRANCE-R ya que activa corriente abajo otras moléculas como Src/Cb1/Akt
y TAB2/TAK1. De esta forma TRAF6 es un factor crítico en la activación de
osteoclastos maduros, pero otros TRAFs pueden compensar parcialmente la
perdida de TRAF6 durante el desarrollo de osteoclastos.
Ratones con una mutación genética a nivel de TRANCE-R presentan un
bloqueo en el desarrollo de los osteoclastos que se ve reestablecido una vez
se introduce TRANCE-R recombinante en células progenitoras de medula
ósea, similar a lo que se observa en ratones trance-/- . Así la interacción entre
TRANCE expresado por células estromales/osteoblastos y su receptor
TRANCE-R expresado sobre precursores de osteoclastos es esencial para la
osteoclastogénesis Ratones con defectos en TRAF6 tienen un fenotipo óseo
similar al de ratones trance-/- y trance-r-/- por un bloqueo parcial de la
osteoclastogénesis y defectos en la activación de osteoclastos maduros
(Theill, 2002).
26

4.3 OPG En 1997, varios grupos de investigadores identificaron de forma simultánea
una proteína que interfería con la diferenciación de osteoclastos. El grupo de
Simonet y colaboradores identificaron una proteína que en cultivos de células
hematopoyéticas y estromales inhibió la formación de los osteoclastos de
forma dependiente de la dosis, y su administración subcutánea en ratones
indujo un aumento de masa ósea y previno la perdida ósea tras ooforectomia
(Simonet, 1997). Debido a esa habilidad de proteger el hueso, los
investigadores denominaron a esta molécula osteoprotegerina (OPG) y en
ratones trasgénicos para OPG, la sobreexpresión de la OPG indujo
osteopetrosis asociada a reducción significativa del numero de osteoclastos,
sin reducción concomitante del numero de macrófagos (Simonet, 1997).
Coincidencialmente otros grupos aislaron un factor similar en medio de
cultivo de fibroblastos humanos, el cual era capaz de inhibir la formación de
los osteoclastos; un grupo la denomino factor inhibidor de la
osteoclastogénesis (OCIF), mientras otro grupo la denomino molécula-1
similar al TNF (TR1) (Tan, 1997). Posteriormente se demostró que la
secuencia del ácido dexosirribonucleico (ADN) de la OPG era idéntica a la
OCIF y se estableció que OCIF, TR1 Y OPG eran la misma molécula (Tsuda,
1998). Él termino que hoy en día se emplea para designar al ligando soluble
que inhibe la diferenciación de los osteoclastos es la OPG.
La OPG es un miembro de la superfamilia del TNFR que regula
negativamente la formación y activación de osteoclastos. La OPG es
producida primariamente por osteoblastos y/o células estromales y sus
precursores, pero también puede ser expresada por células B y células
dendríticas. La OPG es un receptor señuelo, el cual interrumpe la interacción
entre TRANCE y TRANCE - R ya que se une a este ultimo. Por lo tanto la
27

OPG es un inhibidor efectivo de la maduración de osteoclastos y su
activación in vitro (Theill, 2002).
OPG se produce en respuesta a proteína morfogénica ósea 2 (BMP-2), IL-1,
IL-6, IL-11, TGFβ y estrógenos (Ahlen, 2002). La inducción de OPG es
inhibida por PTH, glucocorticoides y prostaglandina E2 (PGE2). La
sobreexpresión de OPG en ratones trasgénicos resulta en osteopetrosis,
ratones deficientes en OPG muestran una marcada osteoporosis como
resultado de un incremento en él numero de osteoclastos funcionales,
indicando que los niveles de masa ósea correlacionan con los niveles de
OPG en ratones (Ahlen, 2002)
Estos hallazgos sugieren que la OPG regula negativamente la formación y
función de los osteoclastos. Así la señalización de TRANCE/TRANCE-
R/OPG junto con el M-CSF juega un papel importante en la homeostasis
ósea.
La relación entre osteoblastos y osteoclastos en la regulación del
remodelamiento óseo podría entonces resumirse así: TRANCE, que se
expresa en la superficie del osteoblasto/célula estromal, se une al TRANCE-
R expresado en la superficie de la célula precursora del osteoclasto e induce
la diferenciación y activación del osteoclasto. La OPG que se encuentra en
forma soluble o que es secretada por los osteoblastos, puede unirse al
TRANCE bloqueando la interacción entre el osteoblasto y los precursores de
osteoclasto e inhibiendo la formación de osteoclastos. TRANCE tiene la
capacidad de activar los osteoclastos y estimular osteoclastos inactivos para
iniciar la resorción ósea e inhibir su apoptosis (Figura 2).
28

Figura 2. Interacción RANK – RANKL, permite la diferenciación a
osteoclastos maduros.
Teitelbaum, S. L. And Ross, F. P. (2003). Genetic Regulation of Osteoclast
Development and Function. Nature. Vol.4. 638 – 649.
4.4 PTH/ Vitamina D3/ Calcitonina
Además de estas citocinas implicadas en el mantenimiento del equilibrio
óseo, existen otros factores solubles cuya actividad modula
significativamente la masa ósea; los más importantes de estos son la
hormona PTH, la vitamina D3 y la calcitonina (Bakker, 2003; Suda, 2003 y
Zaidi, 2002). Sin embargo, la acción de las mismas en el remodelamiento
29

óseo parece ser secundaria al mantenimiento del equilibrio del calcio y el
fósforo plasmático. De todas formas, el papel de la PTH y de la vitamina D3
en la patogénesis de la osteoporosis es interesante ya que ambas hormonas
son potentes moduladores de la actividad osteoclastica, a la vez que regulan
la absorción intestinal de calcio y su excreción renal (Ahmad, 2003).
El aumento en la concentración de PTH en la circulación, promueve la
resorción ósea al incrementar el número de osteoblastos y la tasa de
actividades entre las células osteoclasticas individuales, aunque el efecto de
la PTH parece ser multifactorial, ya que estimula los osteoblastos, aumenta la
producción de proteasas y disminuye la actividad de los inhibidores de
osteoclastos. Tiene acción directa sobre los osteoclastos mediada por el
AMP cíclico y promueve la fusión de células mononucleares favoreciendo la
formación de células gigantes multinucleadas (Bakker, 2003).
La principal función de la vitamina D3 es la de aumentar la actividad resortiva
de los osteoclastos presentes sin aumentar su numero (Suda, 2003).
El papel fisiológico de la calcitonina es menos conocido aunque se sabe que
participa en la embriogénesis del esqueleto (etapa de formación y
crecimiento acelerado del hueso) ya que es un agente con gran potencia
anti-resortiva (antiosteoclastica). La calcitonina produce inhibición de la
actividad osteoclastica e induce la apoptosis de los osteoclastos (Zaidi,
2002).
30

5. SISTEMA INMUNE EN OSTEOCLASTOGÉNESIS
Muchos estudios han investigado la relación entre el sistema inmune y el
sistema óseo en varios modelos animales de enfermedades humanas. Sin
embargo las bases moleculares y celulares de la unión entre estos dos
sistemas es aún desconocida. Aunque actualmente se tiene conocimiento de
que los osteoclastos, los cuales son mediadores importantes del balance
óseo, se originan a partir de precursores hematopoyéticos que también dan
origen a células inmunes (Rho, 2004).
La osteoclastogénesis es positivamente regulada por la interacción de
TRANCE – TRANCE-R y negativamente regulada por OPG (Walsh, 2003).
Muchos estudios muestran que células T activadas pueden inducir
osteoclastogénesis a través de la expresión de TRANCE. De otra parte
células T activadas que producen interferón - γ (INF-γ) pueden inhibir la
osteoclastogénesis por interferir con la vía de señalización TRANCE-
TRANCE-R (Figura 3). Esto sugiere que células T activadas regulan, tanto
positiva como negativamente, la osteoclastogénesis (Rho, 2004). El papel
de las células T activadas en la osteoclastogénesis puede depender de la
acción de TRANCE y la acción negativa del INF-γ. Este balance además
puede verse afectado por factores sistémicos y locales, incluyendo
infecciones de origen microbiano, nutrición, metabolismo, factores
mecánicos, hormonas y citocinas tales como TNFs e Interleucinas (Rho,
2004).
31
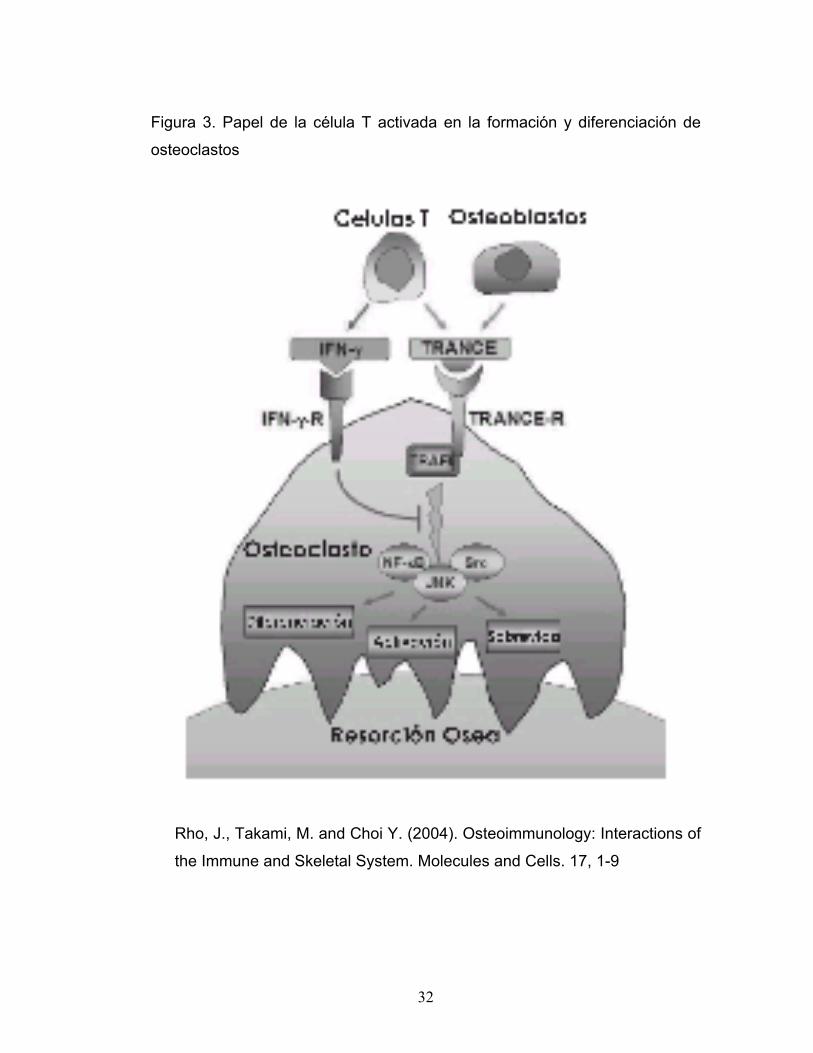
Figura 3. Papel de la célula T activada en la formación y diferenciación de
osteoclastos
Rho, J., Takami, M. and Choi Y. (2004). Osteoimmunology: Interactions of
the Immune and Skeletal System. Molecules and Cells. 17, 1-9
32

Varias citocinas inflamatorias han mostrado jugar un papel importante en la
osteoclastogénesis, incluyendo INF-α, INF-β e INF-γ. El efecto negativo de
esta ultima citocina en la osteoclastogénesis ha sido mostrado y bien
caracterizado. Ratones inyectados con INF-γ muestran defectos en la
formación de osteoclastos multinucleados. Estos hallazgos fueron
observados por Takayanagi quien mostró que INF-γ producido por células T
bloquea la osteoclastogénesis al interferir con la señalización
TRANCE/TRANCE-R (Figura 3). INF-γ además induce degradación de
TRAF6, una molécula como ya se menciono critica en la vía de señalización
TRANCE/TRANCE-R, bloqueando corriente abajo NF-κB y JNK. Se ha
observado que ratones que carecen de INFGR1, un componente del receptor
del INF-γ, muestran perdida de masa ósea debido a un aumento en la
formación de osteoclastos y de la resorción ósea (Takanayanagi, 2002). INF-
α pueden también funcionar como un modulador negativo de la señalización
de TRANCE/TRANCE-R al bloquear la activación de NF-κB y JNK
(Takayanagi, 2000). Otros estudios han mostrado que INF-β también inhibe
la osteoclastogénesis por interferir con la expresión de c-Fos inducida por
TRANCE, un factor de trascripción que es esencial para la formación de
osteoclastos (Takayanagi, 2002). Ratones que carecen de IFNAR, un
componente del receptor de INFα/β, presentan osteoclastos con una mayor
capacidad funcional resultando en un fenotipo como el de osteoporosis
(Takayanagi, 2002).
La inflamación incrementa la osteoclastogénesis. Citocinas inflamatorias
secretadas por células inmunes tales como TNF-α o IL-1, pueden inducir la
expresión de TRANCE en osteoblastos jugando un papel central en perdida
ósea por inflamación (Walsh, 2003). La IL-7 pueden actuar como un potente
factor osteoclastogénico en condiciones de perdida ósea asociada a
inflamación (Toraldo, 2003). Se ha observado que la concentración de
33

TRANCE soluble secretado por células T se encuentra elevado en respuesta
a IL-7 (Toraldo, 2003).
Se ha demostrado que numerosas citocinas y hormonas osteotrópicas
regulan la formación y función de osteoclastos in vitro, pero su relevancia
funcional in vivo no ha sido establecida. Por ejemplo citocinas derivadas de
células T o macrófagos, tales como, IL-4, IL-6, IL-10, IL-12, IL-17, IL-18,
CD40L factor estimulante de colonias granulocito-macrófago (GM-CSF) y
proteína inflamatoria del macrófago (MIP-1), han mostrado regular la
formación y función de osteoclastos in vitro (Toraldo, 2003). El papel de
estas citocinas in vivo no ha sido claramente diferenciado. Ahora bien, IL-6
potencialmente estimula la formación de osteoclastos a través de la
inducción de TNF-α e IL-1 las cuales son secretadas en mieloma múltiple y
llevan a un incremento de la expresión de TRANCE (Toraldo, 2003)
La hipercalcemia es una de las manifestaciones clínicas más frecuentes y
serias asociadas con la acumulación de osteoclastos y marcadores de
resorción ósea (Stewart, 2002). IL-4 suprime el desarrollo de los osteoclastos
y la función de osteoclastos maduros por un mecanismo dependiente de
STAT6, que inhibe de manera irreversible la diferenciación programada de
osteoclatos activados vía TRANCE (Wei, 2002).
IL-18 e IL-12 pueden también sinergicamente inhibir la actividad de resorción
ósea pero el mecanismo no ha sido identificado in vivo. En conclusión existe
evidencia que sugiere que las células T juegan un papel crítico en el
desarrollo y función de los osteoclastos, a través de la señalización cruzada
entre INF y TRANCE en remodelamiento óseo (Horwood, 1998)
34

6. UTILIDAD CLINICA DE LOS MARCADORES DE REMODELAMIENTO
ÓSEO
Los marcadores de recambio óseo son sustancias presentes en la sangre y
la orina, que son producidas o liberadas durante la renovación ósea.
Básicamente son de dos tipos enzimas o proteínas secretadas por los
osteoblastos o los osteoclastos y sustancias producidas durante la formación
o destrucción del colágeno tipo I, la principal proteína de la matriz ósea
(Srivastava, 2005).
El remodelamiento óseo depende de los procesos de resorción y formación
ósea, acoplados en una secuencia orquestada, iniciada por los osteoclastos
y seguida por los osteoblastos. Grupos de osteoblastos y osteoclastos,
trabajando sincrónicamente se organizan formando las llamadas unidades
remodeladoras del hueso (BMU), las cuales se encargan de recambiar una
cantidad determinada de hueso en un sitio especifico del esqueleto. Una
BMU mantiene su integridad funcional por 4 a 8 meses, con un rango de 3
meses a 2 años. Aproximadamente un millón de BMU están activas en el
esqueleto de un adulto sano, en un momento dado, con una tasa de
recambio óseo anual de alrededor del 8-10%. En el calcáneo, las BMU crean
lagunas y ellas se mueven a través de la superficie; en hueso cortical, BMU
se mueven a través de la matriz ósea incrementando la porosidad
(Srivastava, 2005).
Durante la resorción ósea, los osteoclastos crean lagunas de resorción ósea
con microambientes de pH bajos, donde se disuelve la matriz inorgánica
exponiendo la matriz orgánica (mas del 90% de la matriz orgánica esta
compuesta de colágeno tipo I). A medida que la resorción ósea avanza las
enzimas de los osteoclastos digieren la matriz ósea orgánica liberando
35

productos de colágeno tipo I, los cuales incluyen fragmentos de péptidos de
ambos extremos, es decir telopéptidos N-terminales del colágeno tipo I
llamados NTx y telopéptidos C-terminales del colágeno tipo I llamados CTx
y anillos estructurales llamados crosslink de piridinolina (Bonde, 1994).
Normalmente la laguna de resorción creada por los osteoclastos es
completamente llena por material osteoide secretado por los osteoblastos.
Las células mesenquimales son atraídas por las BMU a los sitios de
resorción y estas finalmente se diferencian a osteoblastos. Los osteoblastos
activos secretan osteocalcina (OC), péptidos del pro-colágeno tipo I (N-
terminal – PINP y C – terminal – PICP) y fosfatasa alcalina especifica de
hueso (ALP); la concentración de OC, PINP, PICP y ALP en suero se
correlaciona con la tasa de formación ósea (Miller, 1999).
La formación ósea se completa con la mineralización, la cual ocurre en dos
fases. La primera fase ocurre inmediatamente después de la formación del
osteoide, cuando los cristales de hidroxiapatita son depositados entre la
matriz orgánica. La segunda fase incluye un proceso de mineralización más
lento que se extiende por meses a medida que más mineral es adicionado.
Secundario a la mineralización incrementa la densidad mineral ósea, pero no
el volumen de hueso nuevo (Agerback, 1991).
La formación y resorción ósea se encuentra en equilibrio en un individuo
sano. El pico máximo de masa ósea se alcanza a los 30 años tanto en
mujeres como en hombres y aproximadamente 0.4% de hueso es perdido
por año después de esta edad. Adicionalmente del 1% al 2% de hueso es
perdido por año en los primeros 5 años de la menopausia (Manolagas,
2000).
36
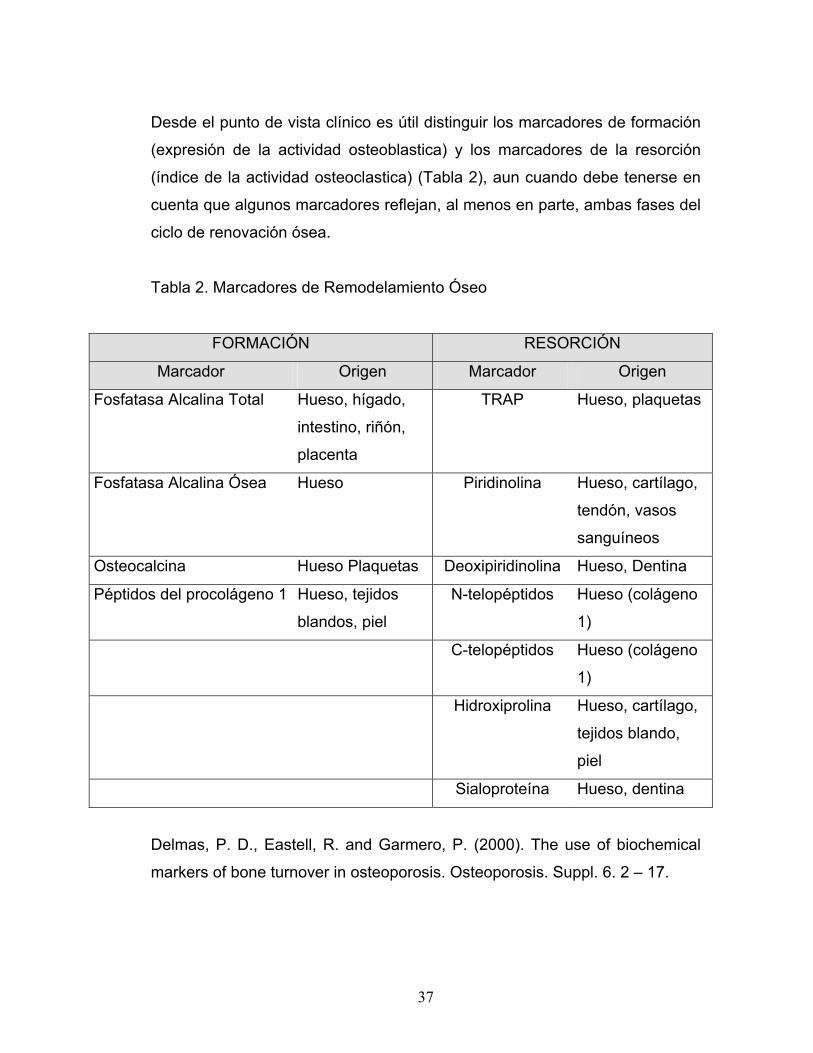
Desde el punto de vista clínico es útil distinguir los marcadores de formación
(expresión de la actividad osteoblastica) y los marcadores de la resorción
(índice de la actividad osteoclastica) (Tabla 2), aun cuando debe tenerse en
cuenta que algunos marcadores reflejan, al menos en parte, ambas fases del
ciclo de renovación ósea.
Tabla 2. Marcadores de Remodelamiento Óseo
FORMACIÓN RESORCIÓN
Marcador Origen Marcador Origen
Fosfatasa Alcalina Total Hueso, hígado,
intestino, riñón,
placenta
TRAP Hueso, plaquetas
Fosfatasa Alcalina Ósea Hueso Piridinolina Hueso, cartílago,
tendón, vasos
sanguíneos
Osteocalcina Hueso Plaquetas Deoxipiridinolina Hueso, Dentina
Péptidos del procolágeno 1 Hueso, tejidos
blandos, piel
N-telopéptidos Hueso (colágeno
1)
C-telopéptidos Hueso (colágeno
1)
Hidroxiprolina Hueso, cartílago,
tejidos blando,
piel
Sialoproteína Hueso, dentina
Delmas, P. D., Eastell, R. and Garmero, P. (2000). The use of biochemical
markers of bone turnover in osteoporosis. Osteoporosis. Suppl. 6. 2 – 17.
37

Por otra parte, la mayoría de los marcadores se encuentran no solo en el
hueso, si no en otros tejidos lo que les resta especificidad. Además, los
marcadores bioquímicos no son específicos de enfermedad pero reflejan
alteraciones en el metabolismo esquelético (Srivastava, 2005).
6.1 TIPOS DE MARCADORES 6.1.1 Marcadores de Formación Ósea 6.1.1.1 Fosfatasa Alcalina Total y Fosfatasa Alcalina Ósea. La fosfatasa
alcalina (FA) es una enzima que interviene en la formación ósea y en la
mineralización del osteoide. Aunque puede originarse en muchos tejidos, en
condiciones normales 50% de la FA sérica proviene del hígado y el otro 50%
es de procedencia ósea (Delmas, 2000).
6.1.1.2 Osteocalcina. La osteocalcina es una proteína con tres residuos de
ácido gamma-carboxiglutámico, que le proporcionan una característica de
apetencia por el calcio. Cuando se forma, parte de ella no se deposita en el
hueso, sino que pasa a la sangre, donde su determinación proporciona un
índice de la actividad osteoblástica (Woitge, 2001).
6.1.1.3 Propéptidos amino y carboxiterminal del procolágeno I. El
colágeno del hueso procede de una molécula de procolágeno sintetizada por
los osteoblastos. Al ser liberada sufre una escisión de sus extremos amino y
carboxiterminal, que resulta en la formación de la molécula de colágeno por
un lado y de los péptidos de los extremos amino y carboxilo por el otro. Estos
se conocen como “protelopéptidos amino y carboxiterminal del colágeno I”
(PINP – PICP respectivamente), y constituyen también un índice de actividad
osteoblástica (Delmas, 2000).
38

6.1.2 Marcadores de Resorción
La mayor parte de los mismos son productos de la degradación de la
molécula de colágeno.
6.1.2.1 Hidroxiprolina. La hidroxiprolina (HYP) es un aminoácido procedente
de la destrucción del colágeno presente en los diferentes tejidos, e incluso
del colágeno de la dieta. Lo primero le confiere una enorme inespecificidad, y
lo segundo obliga a hacer una dieta especial cuando se le quiere determinar.
La HYP se mide en orina; una alternativa a la realización de la dieta exenta
de colágeno, es su determinación en la orina de la segunda micción de la
mañana, con el sujeto en ayunas. En la actualidad, la medición de HYP es de
poca utilidad (Delmas, 2000).
6.1.2.2 Puentes de Piridinolina y Deoxipiridinolina (o puentes piridínicos). La piridinolina (Pid) y la deoxipiridinolina (Dpid) son puentes
formados entre las regiones terminales de las fibras de colágeno, durante su
maduración. La degradación del colágeno origina la liberación de los puentes
y de los distintos péptidos, constituye en consecuencia un índice de la
resorción ósea. Son marcadores mucho más específicos que la HYP, porque
son independientes de la dieta y también porque se encuentran en pocos
tejidos diferentes al hueso (la Pid se encuentra también en el cartílago, los
ligamentos y los vasos. La Dpid en la dentina). Los puentes piridínicos (Pid –
Dpid) se determinan en la orina (Woitge, 2001).
6.1.2.3 Péptidos Terminales o Telopéptidos del Colágeno. Los péptidos
terminales o telopéptidos se pueden medir en sangre o en orina. El
telopéptido carboxiterminal se puede determinar en dos formas, la primera el
octapéptido CTX (Carboxi – Terminal), también conocido como crosslaps; la
segunda, el ICTP (Colagen TeloPeptide I) o CTX-MMP (péptido
39

Carboxiterminal generado por metaloproteasas de la Matriz). El telopéptido
aminoterminal constituye el NXT u osteomark. Actualmente los N o C
telopéptidos se consideran los mejores índices de recambio óseo (Woitge,
2001).
6.1.2.4 Fosfatasa Ácida Resistente a Tartrato. Es una enzima implicada
en la actividad osteoclastica; se determina en la sangre. La complejidad de la
técnica y su relativa inespecificidad la hacen de poca utilidad (Delmas, 2000).
40

5. CONCLUSIONES
Los osteoclastos son componentes críticos en el mantenimiento de la
homeostasis ósea, y se originan a partir de precursores hematopoyéticos que
dan origen a células del Sistema Inmune. Existe una clara relación entre el
Sistema Inmune y el Sistema Óseo dependiente en ocasiones de diferentes
citocinas. De estas citocinas las cuales inicialmente se creía tenían un papel
único en la regulación del sistema inmune ahora se conoce que participan en
la diferenciación y función de los osteoclastos, de manera positiva o negativa
en la homeostasis ósea.
41

BIBLIOGRAFIA
1. Agerback, M. O., Eriksen, E. F., Kragstrup, J., Mosekilde, L. and
Melsen, F. (1991). A reconstruction of the remodeling cycle in normal
human cortical iliac bone. Bone Miner. 12, 101 – 112.
2. Ahlen, J., Andersson, S., Mukohyama, H. Roth, C., Backman, A.
Conoway, H. H. and Lerner, U. H. (2002). Characterization of the
bone-resorptive effect of interleukin-11 in cultured mouse calvarias
bones. Bone 31, 242 – 251.
3. Ahmad, A. M., Hopkins, M. T., Fraser, W. D., Ooi, C. G., Durham, B. H.
and Vora, J. P. (2003). Parathyroid hormone secretory pattern,
circulating activity, and effect on bone turnover in adult growth
hormone deficiency. Bone. 32, 170 – 179.
4. Alliston, T., Choy, L., Ducy, P., Karsenty, G. and DeryncK, R. (2001)
TGF-β induced repression of CBFA1 by Smad3 decreases cbfa1 and
osteocalcin expression and inhibits osteoblast differentiation. Embo
Journal. 20, 2254 – 2272.
5. Arron, J. R. and Choi, Y. (2000). Bone versus immune system. Nature
408, 535-536.
6. Bakker, A. D., Joldersma, M., Klein-Nulend, J. and Burger, E. H.
(2003). Interactive effects of parathyroid hormone and mechanical
stress on nitric oxide and prostaglandin production by primary mouse
osteoblastic cells. Am. J. Physiol. Endocrinol. Metab.
42

7. Baylink, D. J., Finkelman R. D. and Mohan, S. (1993). Growth factors
to stimulate bone formation. J. Bone Miner Res. 8 suppl 2, s565 –
s572.
8. Berenson, J. R. (2002). Treatment of hypercalcemia of malignancy
with bisphosphonates. Semin. Oncol. 29, 12 – 18.
9. Bonde, M., Qvist, P., Fledelius, C., Iris, R. J. and Christiansen, C.
(1994). Immunoassay for quantifying type I collagen degradation
products in urine evaluated. Clin Chem. 40, 2022 – 2025.
10. Boyle, W. J., Simonet, W. S. and Lacey, D. L. (2003) Osteoclast
differentiation and activation. Nature. 423. 337 – 342.
11. Canalis, E., Economides, A. N. and Gazzerro, E. (2003). Bone
morphogenetic proteins, their antagonist and the skeleton. 24, 218 –
235.
12. Chagraoui, H., Tulliez, M., Smayra, T., Komura, E., Giraudier, S., Yun,
T., Lassau, N., Vainchenker, W. And Wendling, F. (2003). Stimulation
of osteoprotegerin production is responsible for osteosclerosis in mice
overexpressing TPO. Blood. 101, 2983 – 2989.
13. Darnay, B. G., Ni, J., Moore, P. A. and Aggarwal, B. B. (1999).
Activation of NF-kappa B by RANK requires tumor necrosis factor
receptor associated factor (TRAF) 6 and NF-kappa B inducing kinase.
Identification of a novel TRAF6 interaction motif. J. Biol. Chem. 274,
7724 – 7773.
43

14. Delmas, P. D., Eastell, R. and Garmero. (2000). The use of
biochemical markers of bone turnover in osteoporosis. Osteoporosis.
Suppl. 6. 2 – 17.
15. Dougall, W. C., Glaccum, M., Charrier, K., Rohrbach, K., Brasel, K.,
De Smedt, T., Daro, E., Smith, J., Tometsko, M. E., Maliszewski, C. R.,
Armstrong, A., Shen, V., Bain, S., Cosman, D., Anderson, D.,
Morrissey, P. J., Peschon, J. J. and Schuh, J. (1999). RANK is
essential for osteoclast and lymph node development. Genes Dev. 13,
2412 – 2424.
16. Helfrinch, M. H., Hobson, R. P., Grabowski, P. S., Zurbriggen, A.,
Cosby, S. L., Dickson, G. R., Frase, W. D., Ooi, C. G., Selby, P. L.,
Crisp, A. J., Wallace, R. G., Kahn, S. and Ralston, S. H. (2000). A
negative search for a paramyxoviral etiology of Paget’s disease of
bone: molecular, immunogical and ultrastructural studies in UK
patients. J. Bone Miner Res. 15, 2315 – 2329.
17. Helfrinch, M. H. and Horton, M. A. (1999). Integrins and adhesion
molecules. In: Seibel, M. J., Robins, S. P., Bilezikan, J. P., editors.
Dynamic of bone and cartilage metabolism. San Diego: Academic
Press. 111 – 125.
18. Horwood, N. J., Udagawa, N., Elliott, J., Grail, D., Okamura, H.,
Kurimoto, M., Dunn, A. R., Martin, T. and Gillespie, M. T. (1998).
Interleukin 18 inhibits osteoclast formation via T cell production of
granulocyte macrophage colony – stimulating factor. J. Clin. Invest. vol
101. 595 – 603.
44

19. Iotsova, V., Caamano, J., Loy, J., Yang, Y., Lewin, A. and Bravo, R.
(1997). Osteopetrosis in mice lacking NF-kappa B1 and NF- kappa B2.
Nat. Med. 3, 1285 – 1289.
20. Kaku, M., Kohno, S., Kawata, T., Fujita, I., Tokimasa, C., Tsutsui, K.
and Tanne, K. (2001). Effects of vascular endothelial growth factor on
osteoclast induction during tooth movement in mice. J. Dent. Res. 80,
1880 – 1883.
21. Kim, N., Takami, M., Rho, J., Josei, R. and Choi, Y. (2002). A novel
member of the leukocyte receptor complex regulates osteoclast
differentiation, J. Exp. Med. 195, 201 – 209.
22. Lean, J. M., Fuller, K. and Chambers, T. J. (2001). FLT3 ligand can
substitute for macrophage colony – stimulating factor in support of
osteoclast differentiation and function. Blood. 98, 2707 – 2713.
23. Lee, S. W., Han, S. I., Kim, H. H. And Lee, Z. H. (2002). TAKI –
dependent activation of AP-1 and c-Jun N-terminal kinase by receptor
activator of NF-kappa B. J. Biochem. Mol. Biol. 35, 371 – 376.
24. Lomaga, M. A., Yeh, W. C., Sarosi, I., Duncan, G. S., Furlonger, C.,
Ho, A., Morony, S., Capparelli, C., Van, G., Kaufman, S. T., Cao, Z.,
Penninger, J. M., Paige, C. J., Lacey, D. L., Dunstan, C. R., Boyle, W.
J., Goeddel, D. V. and Mak, T. W. (1999). TRAF6 deficiency results in
osteopetrosis and defective interleukin – 1, CD40 and LPS signaling.
Genes Dev. 13, 1015 – 1024.
25. Mackie, E. J. (2003). Osteoblst: novel roles in orchestration of skeletal
architecture. Int. J. Biochem Cell Biol. 35, 1301 – 1305.
45

26. Manolagas S. C. (2000). Birht and death of bone cells: basic
regulatory mechanism and implications for the pathogenesis and
treatment of osteoporosis. Endocr Rev. 21, 115 – 137.
27. Miller, P. D., Baran, D. T. and Bilezekian, J. (1999). Practical clinical
application of biochemical markers of bone turnover. J. Clin.
Densitrometry. 2, 323 – 342.
28. Miyamoto, T. and Suda, T. (2003). Differentiation and function of
osteoclast. Keio J Med. 52, 1 – 7.
29. Rho, J., Takami, M. and Choi Y. (2004). Osteoimmunology:
Interactions of the Immune and Skeletal System. Molecules and Cells.
17, 1-9
30. Roodman, G. D. (1996). Advances in bone biology, the osteoclast.
Endocr. Rev. 17, 308 – 332.
31. Sarma, U. and Flanagan, A. M. (1996). Macrophage colony
stimulating factor induces substantial osteoclast generation and bone
resorption in human bone marrow cultures. Blood. 88, 2531 – 2540.
32. Simonet, W. S., Lacey, D. L., Dustan, C. R., Kelley, M., Chang, M. S.
and Luthy, R. (1997). Osteoprotegerin: a novel secreted protein
involved in the regulation of bone density. Cell. 89, 309 – 319.
33. Srivastava, A. K., Vliet, E. L., Lewiecki, M. E., Maricic, M.,
Abdelmalek, A., Gluck, O. and Baylink, D. J. (2005). Clinical use of
serum and urine bone markers in the management of osteoporosis.
Current medical research and opinion. 21, 1015 – 1026
46

34. Stewart, A. F. (2002). Hyperparathyroidism, humoral hypercalcemia of
malignancy, and the anabolic actions of parathyroid hormone and
parathyroid hormone-related protein on the skeleton. J. Bone Miner.
Res. 17, 758 – 762.
35. Suda, T., Kobayashi, K., Jimi, E., Udagawa, N. And Takahashi, N.
(2001). The molecular basis of osteoclast differentiation and activation.
Novartis Found. Symp. 232, 235 – 247.
36. Suda, T., Takahashi, N., Udagawa, N., Jimi, E., Gillespie, M. T. And
Martín, T. J. Modulation of osteoclast differentiation and function by the
new members of the tumor necrosis factor receptor and ligand families.
Endocr Rev. 20, 345 – 357.
37. Suda, T., Nakamura, I., Jimi, E. And Takahashi, N. (1997b).
Regulation of osteoclast function. J. Bone Miner. Res. 12, 869 – 879.
38. Suda, T., Ueno, Y., Fujii, K. And Shinki, T. (2003). Vitamin D and
bone. J. Cell Biochem. 88, 259 – 266.
39. Takayanagi, H., Kim, S., Matsuo, K., Suzuki, T., Sato, K., Yokochi, T.,
Oda, H., Nakamura, K., Ida, N., Wagner, E. F. and Taniguchi, T.
(2002). RANKL maintains bone homeostasis through c-Fos dependent
induction of interferon-beta. Nature. 416, 744 – 749.
40. Takayanagi, H., Ogasawara, K., Hida, S., Chiba, T., Murata, S., Sato,
K., Takaoka, A., Yokochi, T., Oda, H., Tanaka, K., Nakamura, K. And
Taniguchi, T. (2000). T cell mediated regulation of osteoclastogenesis
by signaling cross talk between RANKL and INF-gamma. Nature. 408,
600 – 605.
47

41. Tan K. B., Harrop, J., Reddy, M., Young, P., Terrett, J. and Emery, J.
(1997). Characterization of a novel TNF-like ligand and recently
described TNF ligand and recently described TNF ligand and TNF
receptor superfamily genes and their constitutive and inducible
expression in hematopoietic and non-hematopoietic cells. Gene. 204,
35 – 46.
42. Theill, L. E., Boyle, J. W. and Penniger, M. J. (2002). RANK-L and
RANK: T cells, bone loss and mammalian evolution. Annu. Rev.
Immunol. 20, 795 – 823.
43. Toraldo, G., Roggia, C., Qian, W. P., Pacifici, R. And Weitzmann, M.
N. (2003). IL-7 induces bone loss in vivo by induction of receptor
activator of nuclear factor kappa B ligand and tumor necrosis factor
alpha from T cells. Proc. Natl. Acad. Sci. USA. 100, 125 – 130.
44. Tsuda, E., Goto, M., Mochizuki, S., Yano, K., Kobayashi, F. And
Morinaga, T. (1997). Isolation of a novel cytokine from human
fibroblast that specifically inhibits osteoclastogenesis. Biochem
Biophys Res Commun. 234, 137 – 142.
45. Tsuda, E. and Higashio, K. (1998). Osteoclastogenesis inhibitory
factor (OCIF) /OPG. Nippon Rinsho. 56, 1435 – 1439.
46. Walsh, M. C. and Choi, Y. (2003). Biology of the TRANCE axis.
Cytokine Growth Factor Rev. 14, 251 – 263.
47. Wei, S., Wang, M. W., Teitelbaum, S. L. and Ross, F. P. (2002).
Interleukin – 4 reversibly inhibits osteoclastogenesis via inhibition of
48

NF-kappa B and mitogen – activated protein kinase signaling. J. Biol.
Chem. 277, 6622 – 6630.
48. Woitge, H. W. and Seibel, M. J. (2001). Biochemical markers to
survey bone turnover. Clin Rheum Dis. 27, 49 – 80.
49. Wong, B. R., Rho, J., Robinson, E., Orlinick, J., Chao, M., Kalachikov,
S., Cayani, E., Barlett, F. S., Frankel, W. N., Lee, S. Y., and Choi, Y.
(1997). TRANCE is a novel ligand of the tumor necrosis factor receptor
family that activates c-Jun N-terminal kinase in T cells. J. Biol. Chem.
272, 25190 - 25194.
50. Wong, B. R., Besser, D., Kim, N., Arron, J. R., Vologodskaia, M.,
Hanafusa, H. and Choi, Y. (1999) TRANCE, a TNF family member,
activates Akt/PKB through a signaling complex involving TRAF6 and c-
Src. Mol. Cell 4, 1041 – 1049.
51. Yoshida, H., Hayashi, S., Kunisada, T., Ogawa, M., Nishikawa, S.,
Okamura, H., Sudo, T. And Shultz, L. D. (1990). The murine mutation
osteopetrosis is in the coding region of the macrophage colony
stimulating factor gene. Nature. 345, 442 – 444.
52. Yu, K., Xu, J., Liu, Z., Sosic, D., Shao, J. and Olson, E. (2003).
Conditional inactivation of FGF receptor 2 reveals an essential role for
FGF signaling in the regulation of osteoblast function and bone growth.
Development. 130, 3064 – 3074.
53. Zaidi, M., Blair, H. C., Mooga, B. S., Abe, E., and Huang, C. L. (2003).
Osteoclastogenesis, bone resorption and osteoclast based
therapeutics. J. Bone Miner. Rev. 18, 599 – 609.
49

54. Zaidi, M., Moonga, B. S. and Abe, E. (2002). Calcitonin and bone
formation: a knockout full of suprises. J Clin Invest. 110, 1769 – 1771.
55. Zelzer, E. and Olsen, B. R. (2003). The genetics basis for skeletal
diseases. Nature. 423, 343 – 348.
50