
MAKALAH NUTRISI TANAMAN - blog.ub.ac.id · 2008c.). Selenite selanjutnya dikurangi menjadi selenide...
14
MAKALAH NUTRISI TANAMAN “ASSIMILASI DAN FUNGSI UNSUR HARA MIKRO MENGUNTUNGKAN Se, Ni, Al ” Disusun oleh: Kelas : F Kelompok : 3 Muhammad Hafizh B 125040200111232 Zahra Fitria 125040201111034 Miftah Nur Rokhmat 125040201111040 Alfiyah Kurniawati 125040201111078 Wirda Dayanti Br Ginting 125040201111089 Aryantana Hendarko P 125040201111114 Mar’atus Eski Rinata 125040201111161 UNIVERSITAS BRAWIJAYA FAKULTAS PERTANIAN PROGRAM STUDI AGROEKOTEKNOLOGI MALANG 2015
Transcript of MAKALAH NUTRISI TANAMAN - blog.ub.ac.id · 2008c.). Selenite selanjutnya dikurangi menjadi selenide...

MAKALAH NUTRISI TANAMAN
“ASSIMILASI DAN FUNGSI UNSUR HARA MIKRO
MENGUNTUNGKAN Se, Ni, Al ”
Disusun oleh:
Kelas : F
Kelompok : 3
Muhammad Hafizh B 125040200111232
Zahra Fitria 125040201111034
Miftah Nur Rokhmat 125040201111040
Alfiyah Kurniawati 125040201111078
Wirda Dayanti Br Ginting 125040201111089
Aryantana Hendarko P 125040201111114
Mar’atus Eski Rinata 125040201111161
UNIVERSITAS BRAWIJAYA
FAKULTAS PERTANIAN
PROGRAM STUDI AGROEKOTEKNOLOGI
MALANG
2015

1. PENDAHULUAN
1.1 Latar Belakang
Unsur hara merupakan suatu komponen yang dibutuhkan oleh tanaman dalam jumlah yang
tidak sedikit untuk membantu mendukung pertumbuhan tanaman yang optimal. Tumbuhan
memerlukan asupan unsur hara baik yang tersedia dialam (tanah) maupun yang diaplikasikan
atau diberikan oleh manusia untuk hidup, tumbuh dan menyelesaikan siklus hidupnya, sama
dengan manusia memerlukan makan untuk hidup. Unsur hara harus diberikan secara seimbang
untuk mendapatkan suatu hasil produksi tanaman yang optimal. Pemupukan seimbang yaitu
pupuk yang diberikan harus sesuai dengan apa yang dibutuhkan pada tanaman itu sendiri. Jumlah
kebutuhan akan unsur hara untuk jenis tanaman memiliki perbedaan. Unsur hara esensial
merupakan suatu kebutuhan tanaman yang sangat penting dan yang tidak bisa digantikan oleh
apapun dari semua jenis unsur hara. Unsur hara esensial terdiri dari menjadi dua unsur yaitu
unsur hara mikro (Mo, Cu, Zn, Mn, Fe, Bo, dan Cl) dan unsur hara makro (N, P, K, Ca, Mg, S).
Kelebihan dan kekuranagan unsur hara bagi tanaman dapat menyebabkan terhalangnya
pertumbuhan sehingga tidak optimal. Gejala kelebihan unsur hara pada tanaman dapat dilahat
dari gejala fisik pada bagian-bagian tanaman seperti gejala yang terdapat pada daun, batang,
bungan dan buah selain itu tanaman juga akan menunjukkan gejala seperti daun yag terhambat
sehingga pertumbuhan tanaman menjadi kerdil dan perubahan warna pada daun sering disebut
sebagai klorosi. Defisiensi unsur hara pada tanaman dapat terlihat pada daun-daun pada
umumnya. Salah satu contoh kekurangan unsur hara misalnya yaitu kekurangan unsur hara S,
tanaman yang kekurangan unsur S menunjukkan suatu defisiensi seperti terlihat gejala pada daun
tanaman mengunig, dan terkadang disertai dengan berubahnya warna daun mejadi kemerahan.
Sedangkan untuk gejala jika kelebihan unsur hara mikro yaitu dimana tanaman dapat terjadi
keracunan sehingga menyebabkan tanaman tidak tumbuh dengan baik. Pentingnya pengetahuan
dalam defisiensi tanaman akibat kekurangan unsur hara dapat dijadikan suatu pedoman maupun
petunjuk yang dapat digunakan oleh peani yang sedang berbudidaya tanaman dalam melakukan
untuk menentukan pemupukan yang tepat, optimal, dan jenis pupuk yang harus digunakan. Oleh
karena itu perlu adanya pengetahuan tentang defisiensi dan kelebihan unsur hara mikro pada
tanaman.
Pertumbuhan, perkembangan dan produksi suatu tanaman ditentukan oleh dua faktor utama
yaitu faktor genetik dan faktor lingkungan.Salah satu faktor lingkungan yang sangat menentukan
lajunya pertumbuhan, perkembangan dan produksi suatu tanaman adalah tersedianya unsur-unsur
hara yang cukup di dalam tanah.Diantaranya 105 unsur yang ada di atas permukaan bumi,
ternyata baru 16 unsur yang mutlak diperlukan oleh suatu tanaman untuk dapat menyelesaikan
siklushidupnya dengan sempurna.
Layaknya manusia, tumbuhan juga membutuhkan makanan untuk pertumbuhan dan
perkembangannya. Unsur makro dan unsur mikro merupakan makanan bagi tanaman. Bedanya
hanya pada takaran yang dipakai oleh tanaman tersebut. Jika tanaman kekurangan satu unsur

hara saja (makro/mikro), walaupun unsur hara yang lain cukup banyak, maka produktivitas
pertumbuhan tanaman akan terganggu. Kunci nya adalah, pengelompokan kandungan unsur hara
makro dan mikro dalam tanah dapat kita gunakan untuk memperkirakan kebutuhan unsur hara
tanaman. Dengan itu kita dapat memberikan unsur hara (pupuk) dalam jumlah yang lengkap dan
seimbang sehingga kebutuhan sumber hara pada tanah akan optimal dan terjaga.
1.2 Tujuan
Untuk mengetahui assimilasi dan fungsi unsur hara mikro menguntungkan Se, Ni dan Al
yang terkandung didalam tanah serta gejala yang ditimbulkan akibat defisiensi (kekurangan)
unsur hara.

2. PEMBAHASAN
2.1 selenium
2.1.1 Umum
Kimia selenium (Se) memiliki fitur yang sama dengan sulfur. Selenium, seperti sulfur, bisa
eksis di -2 (selenide SE2), O (unsur selenium), 4 (selenite Se032) dan +6 (selenate Se042)
oksidasi. Selenium hadir dalam tanah dalam jumlah kecil (biasanya mulai dari 0,01 sampai
2mgkg); konsentrasi tinggi (> Smgkg ') yang ditemukan di tanah seleniferous (Mayland et al.,
1989). PH tanah dan Eh mempengaruhi spesies kimia Se hadir di tanah. Perhitungan
termodinamika menunjukkan bahwa bentuk dominan dari selenium adalah selenate di alkalin
dan tanah baik-teroksidasi (pe + pH> 15), Selenite tanah mineral di baik dikeringkan dengan pH
dari asam ke netral (7,5 <pe + pH <15), dan selenide dalam kondisi tanah berkurang (pe + pH
<7,5) (Elrashidi et al., 1987). Selenium merupakan mikronutrien penting untuk hewan, tapi
hakikat belum didirikan untuk tanaman yang lebih tinggi (Terry dkk, 2000;. Sors et al, 2005b.).
Defisiensi Se pada manusia adalah umum; telah diperkirakan bahwa antara 0,5 dan 1 miliar
orang di seluruh dunia mungkin memiliki cukup asupan Se (Combs, 2001). Karena makanan
nabati merupakan sumber penting dari Se bagi manusia dan hewan domestik, penting untuk
memahami bagaimana tanaman mengambil dan metabolisme Se.
2.1.2 Serapan
Selenate adalah analog kimia sulfat; mereka bersaing untuk transporter yang sama selama
serapan akar dan, dengan demikian, penyerapan selenate dapat sangat dikurangi dengan pasokan
sulfat tinggi (Mikkelson dan Wan, 1990; Zayed dan Terry, 1992). Konstanta afinitas (Km) untuk
sulfat dan penyerapan selenate ke akar barley ditemukan untuk menjadi serupa, 19 dan 151iM,
masing-masing (Leggett dan Epstein, 1956). Selenate juga kompetitif menghambat penyerapan
sulfat dari larutan hara; tapi penghambatan ini tidak mungkin nifikan sig dalam tanah untuk
tumbuh tanaman karena konsentrasi selenate dalam larutan tanah lebih rendah daripada sulfat.
Penelitian terbaru telah mengidentifikasi sejumlah mutan tahan selenite dari Arabidopsis
thaliana; fenotip disebabkan oleh mutasi pada afinitas tinggi sulfat transporter Sultrl; 2
mengakibatkan penyerapan menurun dari kedua sulfat dan selenate (Shibagaki et al, 2002;. El
Kassis et al, 2007.). Sultrl; 2 terlokalisir di ujung akar, korteks akar dan akar lateral, dan
ekspresinya ditingkatkan oleh defisiensi S. Transporter ini merupakan rute utama untuk masuk
sulfat dan selenate ke akar Arabidopsis thaliana (Shibagaki et al, 2002;.. El Kassis et al, 2007;
Barberon et al, 2008.). Pengaruh pasokan sulfat selenate penyerapan tidak hanya melalui
kompetisi langsung untuk transporter membran, tetapi juga melalui regulasi ekspresi gen
transporter sulfat. Tanaman Sulphur-kekurangan up-mengatur ekspresi gen transporter sulfat,
yang mengarah ke peningkatan tajam dalam kapasitas untuk penyerapan selenate (Li et al, 2008;.
Shinmachi et al, 2010.).
Spesies tanaman berbeda kuat dalam pengambilan Se dan akumulasi dalam tunas dan juga
dalam kapasitas mereka untuk mentolerir konsentrasi Se tinggi dalam jangka menengah dan /
atau dalam jaringan menembak rooting. Contoh perbedaan antara spesies tanaman akumulasi Se
ditunjukkan pada Tabel 8.12. Berdasarkan perbedaan-perbedaan ini tanaman dapat
diklasifikasikan menjadi Se-akumulator dan non-akumulator, dan orang-orang antara kedua jenis
sebagai Se-indikator. Beberapa spesies dari genus Astragalus, Xylorrhiza dan Stanleyea khas Se-
akumulator, dan mampu tumbuh di tanah Se tinggi (tanah seleniferous) tanpa efek yang
merugikan pada pertumbuhan dan mencapai konsentrasi Se menembak setinggi 20-30 mg Se g
dw (Rosenfeld dan beath, 1964). Namun, dalam Astragalus genus ada perbedaan besar antara

spesies dan ekotipe dalam kapasitas mereka untuk mengumpulkan Se, konsentrasi Se dalam
akumulator menjadi 100-200 kali lipat lebih tinggi daripada di non-akumulator (sedikit
perhatian, 1969; Davis, 1986). Anggota Brassicaceae seperti mustard hitam (Brassica nigra L.)
dan brokoli (Brassica oleracea botrytis L.) juga mengumpulkan jumlah yang relatif besar dari Se
dan mungkin berisi, dan mentolerir, beberapa ratus pgSeg 'shootdw (Zayed dan Terry, 1992). Di
sisi lain, spesies tanaman pertanian dan hortikultura sebagian besar non-akumulator (sedikit
perhatian, 1981) dan toksisitas Se dapat terjadi bahkan pada konsentrasi di bawah l00igSeg
'(Mikkelsen et al., 1990). Putih et al. (2007a) dibandingkan selenate dan serapan sulfat oleh 39
spesies tanaman tumbuh dalam budaya hidroponik di bawah kondisi yang sama. Mereka
menemukan bahwa, di antara 37 spesies Se non-akumulator, ada hubungan positif yang sangat
erat antara daun S dan daun konsentrasi Se (Gambar. 8.1 lA), menunjukkan bahwa selenite dan
akumulasi sulfat yang sangat terkait. Secara umum, spesies Brassicaceae mampu mengakumulasi
lebih Se karena mereka memiliki kemampuan yang lebih besar untuk mengumpulkan akumulator
S. Dua Se (Astragalus racemosus dan Stanleya pinnata) termasuk dalam penelitian ini
menyimpang dari hubungan ini dengan memiliki konsentrasi Se tinggi pada daun. Kemampuan
untuk mengumpulkan selenate relatif terhadap sulfat dapat diukur dengan selenate / index
diskriminasi sulfat, yang merupakan rasio molar [daun Se / daun Sj / [solusi selenate / solusi
sulfat]. Sebagian besar spesies tanaman diuji oleh Putih et al. (2007a) memiliki indeks
diskriminasi sekitar 1, menunjukkan tidak ada diskriminasi yang jelas antara dua anion (Gambar.
8.llB). Gambar 8.11B juga termasuk data dari studi Bell et al. (1989) menguji dua jenis tanaman
dengan differentratios dari selenate / suiphate dalam larutan nutrisi. Beberapa spesies tanaman
(misalnya, glycyphyllos Astragalus, vulgaris Beta dan Medicago sativa) memiliki indeks di
bawah 1, menunjukkan bahwa transporter (s) mungkin memiliki afinitas yang lebih tinggi untuk
sulfat daripada selenate. Sebaliknya, Se-akumulator (Astragalus racemosus, Stanleya pinnata dan
Astra ga / kami bisulcatus) memiliki indeks diskriminasi antara 2 dan 10 yang merupakan bukti
kuat bahwa beberapa transporter di spesies ini memiliki selektivitas tinggi untuk selenate.
Selenite juga dapat hadir dalam tanah (Stroud et al., 2010), meskipun ketersediaan untuk
tanaman adalah lebih rendah dari selenate karena adsorpsi kuat dengan besi oksida / hidroksida
dalam tanah (Barrow dan Whelan, 1989). Mekanisme penyerapan selenite oleh tanaman tidak
dipahami dengan baik. Penelitian sebelumnya menyarankan bahwa Selenite dapat memasuki sel
akar pasif dengan difusi (Terry dkk., 2000). Namun, serapan Selenite adalah, setidaknya
sebagian, aktif (Arvy, 1993; Li et al, 2008c.). Serapan Selenite dihambat oleh adanya fosfat
dalam medium, tapi ditingkatkan oleh defisiensi P pada tanaman (Hopper dan Parker, 1989; Li et
al, 2008c.), Menunjukkan keterlibatan yang mungkin dari transporter fosfat dalam penyerapan
selenite. Pada pH rendah (<4.0), proporsi yang signifikan dari selenite yang terdisosiasi sebagai
H2SeO3, dan molekul netral ini dapat menyerap melalui NIP2 beras; (. Zhao et al, 2010) 1
saluran aquaporin, yang merupakan transporter asam silikat (lihat Bagian 8.3).
Sebuah perbedaan yang jelas antara selenate dan selenite adalah bahwa mantan cepat
translokasi dari akar ke tunas, sedangkan yang terakhir ini siap berasimilasi ke dalam bentuk
organik di akar tanaman dengan terbatas translokasi akar-to-shoot (Asher et al, 1977;. De Souza
et al, 1998;. Li et al, 2008c).. Perbedaan ini, bersama-sama dengan adsorpsi kuat selenite dalam
tanah, menjelaskan mengapa aplikasi dari selenate lebih efektif daripada selenite dalam
meningkatkan konsentrasi Se pada tanaman. Asam amino Seleno-seperti selenomethiomne dapat
segera diambil oleh bibit gandum (Abrams et al., 1990). Namun, pentingnya penyerapan ini di
tanah untuk tumbuh tanaman tidak jelas.

2.1.3 Asimilasi dan Metabolisme
Selenium berasimilasi dalam tanaman melalui S asimilasi jalur (Gambar 8.12.) (Terry dkk,
2000;. Sors et al, 2005b.). Dalam jalur ini, selenate diaktifkan oleh ATP suiphurylase untuk
adenosine 5'-phosphoselenate (apse), yang kemudian dikurangi menjadi selenite oleh APS
reduktase. Aktivasi selenate tampaknya menjadi langkah tingkat-membatasi untuk pengurangan
selenite, dan dapat diatasi dalam tanaman transgenik lebih mengekspresikan ATP suiphurylase
(Pilon-Smits et al., 1999). Langkah tingkat-membatasi Ini juga menjelaskan mengapa selenite
jauh lebih mudah berasimilasi pada tanaman dari selenate (De Souza et al, 1998;. Li et al,
2008c.). Selenite selanjutnya dikurangi menjadi selenide mungkin melalui reaksi non-enzimatik
menggunakan glutation tereduksi (GSH) sebagai reduktan (sors et al., 2005b). Selenide yang
berasimilasi dengan selenocysteine asam amino dikatalisis oleh sintase kompleks sistein, dan
selanjutnya berasimilasi selenomethionine melalui jalur biosintesis metionin. Kedua
selenocysteine dan selenomethionine dapat segera dimasukkan ke dalam protein dalam tanaman
non-akumulator melalui substitusi non-spesifik sistein dan metionin, masing-masing, dan ini
terutama substitusi ini yang menyebabkan toksisitas untuk p1ant, karena protein menjadi tidak
berfungsi atau kurang fungsional enzim dari protein S-mengandung sesuai (Eustice et al, 1981;.
Brown dan sedikit perhatian, 1982). Penggabungan asam selenoamino diduga sangat penting
dalam enzim dengan kelompok sulphhydryl (-SH) sebagai situs katalitik.
Kedua selenocysteine dan selenomethionine dapat termetilasi, dan kemudian tidak lagi
mampu menggantikan sistein dan metionin dalam sintesis protein. Akumulator selenium berbeda
dari non-akumulator dalam memiliki kemampuan yang kuat untuk mengubah selenocysteine
menjadi berbagai asam selenoamino non-protein, seperti Se-methylselenocysteine dan '--
glutamyl-Se-methylselenocysteine. Metilasi selenocysteine merupakan mekanisme penting dari
Se detoksifikasi pada tanaman akumulator. Langkah metilasi dikatalisis oleh selenocysteine
methyltransferase (Neuhierl dan Bock, 1996), dan aktivitas enzim ditemukan berkorelasi erat
dengan kemampuan akumulasi Se dalam delapan spesies Astragalus (sors et al., 2005b). Dalam
Se akumulator Astragalus bisulcatus dan Stanleya pinnata, daun muda mengandung konsentrasi
tinggi Se, dengan Se-methylselenocysteine terhitung lebih dari 70% dari total Se (Pickering et al,
2003;.. Freeman et al, 2006). Se-methylselenocysteine dan y-glutamil-turunannya juga
ditemukan di beberapa tanaman yang dapat dimakan, termasuk bawang putih, bawang, brokoli
dan lain-lain dari Allium dan Brassica keluarga, terutama bila ditanam di lingkungan Se-

diperkaya (Whanger, 2002; Rayman et al. 2008). Sebaliknya, biji-bijian sereal mengandung
terutama selenomethionine (Whanger, 2002;. Rayman et al, 2008).
Kemiripan antara S dan metabolisme Se pada tanaman juga ada dalam produksi senyawa
volatil yang dirilis oleh bagian aerial tanaman (lihat juga Bab 4). Senyawa selenide utama
volatile dimethylselenide (DMSE), yang selenomethionine adalah prekursor (Gbr. 8.12).
Tanaman juga bisa menguap dimethyldiselenide (DMDSe) yang dihasilkan melalui metilasi dan
oksidasi berikutnya selenocysteine. Tingkat Se penguapan bervariasi antara spesies tanaman.
Dengan pasokan 2OjiM selenate, beras, brokoli dan kubis diuapkan 200-350pg5em2 daun daerah
hari 'dibandingkan dengan kurang dari l5igSem2 luas daun hari' di bit gula, selada dan bawang
(Terry dkk., 1992). Dalam brokoli, yang terakumulasi hingga beberapa ratus .igSegdw, tingkat
pelepasan senyawa volatil Se sekitar tujuh kali lebih tinggi pada pasokan S rendah dibandingkan
dengan pasokan S tinggi, karena penghambatan penyerapan selenate dan persaingan di dalam
pabrik di situs S asimilasi dengan yang terakhir (Zayed dan Terry, 1992). Dalam sawi (Brassica
juncea), lebih Se telah diuapkan saat tanaman telah tersedia dengan selenite daripada dengan
selenate (De Souza et al., 1998). Bakteri rizosfir juga tampaknya memainkan peran penting
dalam Se penguapan (Terry dkk., 2000). Kemampuan tanaman dan mikroorganisme rizosfir
mereka terkait dengan menguap Se, atau menumpuk Se dalam biomassa tanaman, dapat
dimanfaatkan sebagai strategi fitoremediasi untuk membersihkan tanah Se-terkontaminasi (Terry
dkk., 2000).
2.1.4 Efek Menguntungkan di Pertumbuhan Tanaman
Selenium sangat penting bagi manusia dan hewan karena kebutuhan Se di sejumlah enzim,
seperti glutation peroksidase, di mana selenocysteine berfungsi sebagai situs katalitik. Meskipun
ada glutathione peroksidase enzim-seperti pada tanaman yang lebih tinggi, mereka muncul untuk
mengandung sistein, tidak selenocysteine, di situs aktif enzim (Terry dkk., 2000). Meskipun
kurangnya bukti-bukti definitif Se esensial pada tumbuhan tingkat tinggi, ada laporan bahwa
dosis kecil Se meningkatkan pertumbuhan tanaman atau reproduksi. Hartikainen dan rekan kerja
melaporkan bahwa sejumlah kecil Se ditambahkan ke tanah peningkatan pertumbuhan ryegrass,
tertunda penuaan selada, dan meningkatkan resistensi selada dan ryegrass untuk Uy iradiasi
(Hartikainen, 2005). Efek ini tampaknya terkait dengan aktivitas ditingkatkan glutathione
peroxidase dan mengurangi peroksidasi lipid. Lyons et al. (2009) menunjukkan bahwa
penambahan 20-50 nM Na selenite untuk larutan nutrisi peningkatan produksi benih Brassica
rapa sebesar 43%, sementara tidak memiliki efek pada biomassa total tanaman. Tanaman
Selenite diperlakukan memiliki aktivitas total pernafasan lebih tinggi pada daun dan bunga, serta
peningkatan konsentrasi protein sitokrom oksidase di bunga. Penelitian ini dilakukan dengan
pengobatan + Se dan kontrol yang bertempat di dua kamar dikendalikan lingkungan yang
terpisah untuk mencegah transfer volatil Se dari perlakuan dengan kontrol. Konsentrasi Se tinggi
pada tanaman akumulator dapat menawarkan perlindungan terhadap herbivora (misalnya, Galeas
et al., 2008), sehingga pemberian keuntungan adaptif terhadap tanaman. Pengamatan ini
mendukung hipotesis pertahanan elemental diajukan untuk menjelaskan evolusi dari logam atau
metalloid hiperakumulasi sifat (Boyd, 2007). Perbedaan besar dalam konsentrasi Se pada
tanaman pertama menarik perhatian di l930s ketika menyadari bahwa toksisitas Se bertanggung
jawab untuk gangguan tertentu pada hewan merumput di vegetasi asli dari tanah seleniferous
(Brown dan sedikit perhatian, 1982; Miller et al, 1991.).

2.1.5 biofortifikasi
Saat ini sudah ada kesadaran yang lebih besar tentang pentingnya Se bagi kesehatan
manusia daripada di masa lalu, seperti kesadaran bahwa persentase besar penduduk di banyak
negara memiliki intake yang tidak memadai Se (Combs, 2001; Rayman, 2008). Selenium
memasuki rantai makanan terutama melalui serapan tanaman dari tanah. Konsentrasi selenium
dalam tanaman pangan sangat bervariasi sebagai akibat dari variasi geologi dan tanah kondisi
yang mendasari; asupan manusia Se juga bervariasi antara negara dan wilayah negara,
mencerminkan variasi dalam konsentrasi Se dalam makanan (Combs, 2001; Rayman, 2008).
Konsentrasi Se minimum untuk hewan dan manusia adalah sekitar 50- 100 pg Se kg dw di pakan
ternak / makanan (gissel-Nielsen et al., 1984). Strategi untuk meningkatkan asupan manusia Se
adalah tanaman biofortifikasi, baik melalui penggunaan pupuk Se (biofortifikasi agronomi) atau
dengan perbaikan genetik pada tanaman akumulasi Se. Biofortifikasi agronomi telah
dipraktekkan di Finlandia sejak pertengahan 1980-an dengan penambahan wajib sejumlah kecil
Se Na sebagai selenate semua pupuk multi-hara (6-l 6mg Se kg 'pupuk). Praktek ini telah
meningkatkan konsentrasi Se dalam sereal, sayuran dan produk hewani dan lebih dari dua kali
lipat asupan Se oleh penduduk Finlandia (Hartikainen, 2005). Dibandingkan dengan
mengarahkan suplementasi Se, biofortifikasi agronomi dianggap menguntungkan dalam
anorganik Se berasimilasi dengan tanaman ke dalam bentuk organik, yang lebih bioavailable
untuk manusia. Selain itu, tanaman bertindak sebagai penyangga yang efektif yang dapat
mencegah kecelakaan asupan Se berlebihan oleh manusia yang mungkin terjadi dengan
suplementasi langsung (Hartikainen, 2005). Tidak seperti mikronutrien lain seperti Fe dan Zn,
relatif mudah untuk meningkatkan konsentrasi Se pada tanaman pangan dengan pemupukan,
karena selenate sangat bioavailable untuk tanaman dan mudah diangkut dari akar ke tunas, di
mana ia berasimilasi ke dalam bentuk organik yang berbeda. Hal ini ditunjukkan dalam studi
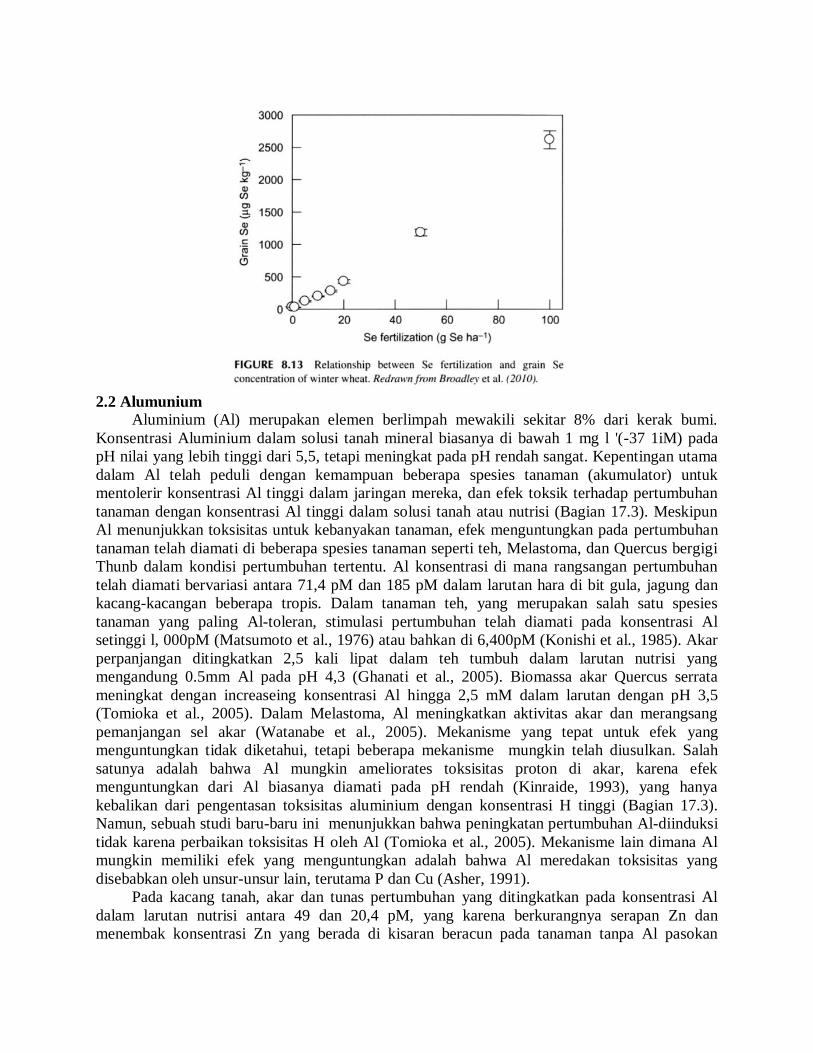
lapangan dengan gandum musim dingin, menunjukkan bahwa konsentrasi Se dalam gandum
meningkat secara linear dari 30jigkg 'dalam kontrol untuk 2.600 jigkg dalam pengobatan
menerima l00gSeha tambahan' dalam bentuk Na selenate (Broadley et al., 2010) (Gbr. 8.13).
Selain itu, tepung terigu Se-diperkaya berisi didominasi selenomethionine (-80%), dengan
selenocysteine, Se metil selenocysteine dan anorganik Se juga hadir dalam proporsi kecil (Hart et
al., 2011).
Jumlah pemulihan diterapkan Se oleh tanaman gandum adalah 20-35%; sisanya itu
mungkin kehabisan keluar dari kedalaman perakaran meninggalkan sedikit efek residu pada
tanaman berikutnya (Broadley et al, 2010;. Stroud et al, 2010.). Secara umum, aplikasi dari
selenate lebih efektif dalam meningkatkan hasil panen Se konten dari selenite (Mikkelsen et al,
1990;. Hawkesford dan Zhao, 2007).
Biofortifikasi Se pada tanaman melalui perbaikan genetik memerlukan variasi genetik
dalam penyerapan dan / atau asimilasi Se. Dalam genotipe roti gandum diuji, variasi genetik
dalam konsentrasi Se butir kecil dibandingkan dengan variasi lingkungan (Lyons et al, 2005;.
Zhao et al, 2009.). Variasi antar-spesies jauh lebih luas, dengan Se akumulator mewakili fenotip
ekstrim (Gbr. 8.11). Sejumlah studi telah melaporkan peningkatan beberapa kali lipat dalam
penyerapan Se oleh tanaman transgenik lebih-mengekspresikan berbagai gen yang terlibat dalam
S! Se asimilasi, tetapi studi ini umumnya bertujuan untuk meningkatkan Se serapan untuk tujuan
fitoremediasi tanah Se-terkontaminasi (Pilon Smits 2009). Apakah tanaman transgenik dapat
mengakumulasi lebih Se dari tanah dengan ketersediaan Se terbatas masih belum jelas.
2.2 Alumunium
Aluminium (Al) merupakan elemen berlimpah mewakili sekitar 8% dari kerak bumi.
Konsentrasi Aluminium dalam solusi tanah mineral biasanya di bawah 1 mg l '(-37 1iM) pada
pH nilai yang lebih tinggi dari 5,5, tetapi meningkat pada pH rendah sangat. Kepentingan utama
dalam Al telah peduli dengan kemampuan beberapa spesies tanaman (akumulator) untuk
mentolerir konsentrasi Al tinggi dalam jaringan mereka, dan efek toksik terhadap pertumbuhan
tanaman dengan konsentrasi Al tinggi dalam solusi tanah atau nutrisi (Bagian 17.3). Meskipun
Al menunjukkan toksisitas untuk kebanyakan tanaman, efek menguntungkan pada pertumbuhan
tanaman telah diamati di beberapa spesies tanaman seperti teh, Melastoma, dan Quercus bergigi
Thunb dalam kondisi pertumbuhan tertentu. Al konsentrasi di mana rangsangan pertumbuhan
telah diamati bervariasi antara 71,4 pM dan 185 pM dalam larutan hara di bit gula, jagung dan
kacang-kacangan beberapa tropis. Dalam tanaman teh, yang merupakan salah satu spesies
tanaman yang paling Al-toleran, stimulasi pertumbuhan telah diamati pada konsentrasi Al
setinggi l, 000pM (Matsumoto et al., 1976) atau bahkan di 6,400pM (Konishi et al., 1985). Akar
perpanjangan ditingkatkan 2,5 kali lipat dalam teh tumbuh dalam larutan nutrisi yang
mengandung 0.5mm Al pada pH 4,3 (Ghanati et al., 2005). Biomassa akar Quercus serrata
meningkat dengan increaseing konsentrasi Al hingga 2,5 mM dalam larutan dengan pH 3,5
(Tomioka et al., 2005). Dalam Melastoma, Al meningkatkan aktivitas akar dan merangsang
pemanjangan sel akar (Watanabe et al., 2005). Mekanisme yang tepat untuk efek yang
menguntungkan tidak diketahui, tetapi beberapa mekanisme mungkin telah diusulkan. Salah
satunya adalah bahwa Al mungkin ameliorates toksisitas proton di akar, karena efek
menguntungkan dari Al biasanya diamati pada pH rendah (Kinraide, 1993), yang hanya
kebalikan dari pengentasan toksisitas aluminium dengan konsentrasi H tinggi (Bagian 17.3).
Namun, sebuah studi baru-baru ini menunjukkan bahwa peningkatan pertumbuhan Al-diinduksi
tidak karena perbaikan toksisitas H oleh Al (Tomioka et al., 2005). Mekanisme lain dimana Al
mungkin memiliki efek yang menguntungkan adalah bahwa Al meredakan toksisitas yang
disebabkan oleh unsur-unsur lain, terutama P dan Cu (Asher, 1991).
Pada kacang tanah, akar dan tunas pertumbuhan yang ditingkatkan pada konsentrasi Al
dalam larutan nutrisi antara 49 dan 20,4 pM, yang karena berkurangnya serapan Zn dan
menembak konsentrasi Zn yang berada di kisaran beracun pada tanaman tanpa Al pasokan

(Asher, 1991). Baru-baru ini, mekanisme lain yang mungkin untuk efek menguntungkan Al-
diinduksi telah dilaporkan. Aluminium meningkatkan aktivitas superoksida dismutase (SOD),
katalase (CAT) dan askorbat peroksidase (APX) di akar kedua tanaman teh utuh dan sel kultur
(Ghanati et aL., 2005). Peningkatan Al-diinduksi dalam aktivitas enzim antioksidan dapat
menyebabkan peningkatan integritas membran, lignifikasi tertunda dan penuaan, mengakibatkan
stimulasi pertumbuhan. Dalam Melastoma, alasan utama untuk peningkatan pertumbuhan Al-
diinduksi diusulkan untuk menjadi pengentasan keracunan Fe oleh Al (Watanabe et al., 2006).
Pertumbuhan ditingkatkan oleh Al lebih kuat di bawah lebih dari Fe, dan konsentrasi Fe
menurun Al baik di akar dan tunas. Kelebihan Fe, seperti yang terjadi pada tanah asam,
menginduksi produksi spesies oksigen reaktif, yang mengarah ke gangguan berbagai fungsi sel.
Oleh karena itu, pengentasan Al-diinduksi toksisitas Fe mungkin penting dalam tanah (Watanabe
et al., 2006). Kesimpulannya, konsentrasi rendah dari Al mungkin memiliki efek menguntungkan
pada pertumbuhan dalam kondisi tertentu, dan efek yang menguntungkan ini mungkin fenomena
yang lebih umum pada spesies tanaman dengan Al toleransi yang tinggi dan kapasitas tinggi Al
serapan (akumulator). Namun, dalam non-akumulator, efek negatif dari Al pada pertumbuhan
tanaman di tanah pH rendah adalah aturan (Bagian 17.3).
2.3 Nikel
2.3.1 Fungsi nikel (Ni) dalam sintesis tanaman
Nikel mempunyai fungsi penting dalam sintesis protein diantaranya adalah Ni-glyoxylase,
NiFe-hydrogenase Pada sintesis protein urease, nikel berperan dalam pembentukan struktur
katalitik pada enzyme. Pada proses fotosintesis, nikel akan bekerjasama dengan enzim
hidrogenase dan struktur Fe-S, sehingga berkaitan dengan sintesis urease, maka pada tanaman
yang kekurangan nikel, aplikasi urea lewat daun (foliar aplication) akan menyebabkan akumulasi
dari urea pada daun yang kelamaan dapat mengakibatkan nekrotik (Tabel 7.30). Nikel
mengaktifkan beberapa enzim yang berfungsi pada proses perkecambahan. Ketersediaan nikel
pada tanaman tergantung dari spesies tanaman dan juga transport di dalam jaringan akar.
2.3.2 Asimilasi unsur hara mikro Nikel (Ni)
Unsur hara mikro nikel (Ni) tersedia dalam bentuk Ni (II) dan merupakan struktur komplek
yang stabil dengan histidin, cystein dan citrate. Konsentrasi nikel dalam tanaman antara 0.01ug
g-1 dw > 10 ug g dw-1, dan jumlah yang sedikit tersebut sangat penting pada proses
perkecambahan. Sehingga ketersediaan air dalam tanaman sangat diperlukan untuk mendukung
kapasitas tukar kation pada saat proses perkecambahan.. Gambar 7.22 Korelasi antara aplikasi
nikel dan prosentase perkecambahan

Akumulasi urea (sampai 3% dari bobot kering) mengarah ke ujung helaian daun tanaman yang
defisien-Ni (Tabel 7.31) Pada kedelai konsentrasi ureide (termasuk purine bebas dan uric acid)
rendah dan tidak terpengaruh oleh suply Ni.
Berbagai jalur biosintesis urea dalam tanaman telah diketahui (Gambar 7.23). Siklus ornithine
protein, pada daun dewasa, pada pertumbuhan reproduktif, dan dalam perkecambahan benih
legume.

2.3.3 Diagnosis kekurangan / keracunan nikel (Ni)
Diagnosis kekurangan maupun kelebihan unsur Ni sangat penting. Mengetahui diagnosis
gejala kekurangan maupun kelebihan unsur Ni, akan mampu melakukan tindakan penyelamatan
terhadap tanaman tersebut. Defisiensi Ni mengurangi aktivitas urease sehingga terjadi keracunan
urea necrosis pada ujung daun. Gejala kekurangan nikel biasanya Nampak seperti daun bulai
yang menguning sepanjang daun yakni mulai dari pangkal hingga ujung daun. Pada daun bagian
bawah akan mengalami klorosis (intervena klorosis).
Diagnosis kekurangan maupun kelebihan unsur Ni sangat penting. Mengetahui diagnosis
gejala kekurangan maupun kelebihan unsur Ni, akan mampu melakukan tindakan penyelamatan
terhadap tanaman tersebut. Defisiensi Ni mengurangi aktivitas urease sehingga terjadi keracunan
urea necrosis pada ujung daun. Gejala kekurangan nikel biasanya Nampak seperti daun bulai
yang menguning sepanjang daun yakni mulai dari pangkal hingga ujung daun. pada daun bagian
bawah akan mengalami klorosis (intervena klorosis)
2.4 UNSUR LAIN
Persyaratan untuk elemen seperti yodium (I) dan vanadium (V) adalah cukup mapan untuk
jenis tanaman yang lebih rendah tertentu, termasuk ganggang laut (I) dan ganggang air tawar (V)
dan jamur (V). Laporan pada stimulasi pertumbuhan tanaman yang lebih tinggi oleh unsur-unsur
lain yang langka dan tidak jelas. Contoh ini adalah efek dari V pada pertumbuhan tomat
(Basiouny, 1984), atau efek dari titanium (Ti) pada pertumbuhan (Pais, 1983), aktivitas enzim
dan fotosintesis (Dumon dan Ernst, 1988) dari berbagai tanaman spesies. Untuk informasi lebih
lanjut tentang V dan saya melihat Bollard (1983), dan Ti melihat Dumon dan Ernst (1988). Baru-
baru ini bunga telah meningkat dalam unsur-unsur tanah jarang lanthanium (La) dan cerium (Ce)
untuk peningkatan pertumbuhan tanaman. Campuran dari kedua elemen yang digunakan pada
skala besar di Cina sebagai semprotan daun atau perlakuan benih spesies tanaman pertanian dan
hortikultura. Jumlah yang diberikan adalah dalam kisaran khas untuk mikro. Ada laporan dari
peningkatan substansial dalam pertumbuhan tanaman dan hasil di bawah kondisi lapangan yang,
bagaimanapun, memerlukan dokumentasi lebih berhati-hati dan reproduksi dalam kondisi yang
terkendali. Untuk informasi lebih lanjut lihat Asher (1991). Ada sejumlah besar laporan tentang
adanya logam berat, seperti kadmium, kromium, timbal dan merkuri, pada tumbuhan tingkat
tinggi. Sebagian besar laporan ini terutama berkaitan dengan pencemaran lingkungan, kehadiran
logam berat dalam rantai makanan, dan perbedaan genotypical dalam konsentrasi racun kritis
logam berat pada tanaman (Ernst dan Joose-van Damme, 1983). Bukti yang meyakinkan dari
efek menguntungkan dari logam berat pada pertumbuhan tanaman yang lebih tinggi kurang.

3. PENUTUP
Kimia selenium (Se) memiliki fitur yang sama dengan sulfur. Selenium, seperti sulfur,
bisa eksis di -2 (selenide SE2), O (unsur selenium), 4 (selenite Se032) dan +6 (selenate Se042)
oksidasi. Selenium hadir dalam tanah dalam jumlah kecil (biasanya mulai dari 0,01 sampai
2mgkg); konsentrasi tinggi (> Smgkg ') yang ditemukan di tanah seleniferous (Mayland et al.,
1989). Aluminium (Al) merupakan elemen berlimpah mewakili sekitar 8% dari kerak bumi.
Konsentrasi Aluminium dalam solusi tanah mineral biasanya di bawah 1 mg l '(-37 1iM) pada
pH nilai yang lebih tinggi dari 5,5, tetapi meningkat pada pH rendah sangat. Kepentingan utama
dalam Al telah peduli dengan kemampuan beberapa spesies tanaman (akumulator) untuk
mentolerir konsentrasi Al tinggi dalam jaringan mereka, dan efek toksik terhadap pertumbuhan
tanaman dengan konsentrasi Al tinggi dalam solusi tanah atau nutrisi. Nikel mempunyai fungsi
penting dalam sintesis protein diantaranya adalah Ni-glyoxylase, NiFe-hydrogenase Pada sintesis
protein urease, nikel berperan dalam pembentukan struktur katalitik pada enzyme. Pada proses
fotosintesis, nikel akan bekerjasama dengan enzim hidrogenase dan struktur Fe-S, sehingga
berkaitan dengan sintesis urease, maka pada tanaman yang kekurangan nikel, aplikasi urea lewat
daun (foliar aplication) akan menyebabkan akumulasi dari urea pada daun yang kelamaan dapat
mengakibatkan nekrotik.

DAFTAR PUSTAKA
Marschner, P. 2012. Marschner’s Mineral Nutrition of Higher Plants. p. 135-185




















