
in Vitro - fp.unmas.ac.idfp.unmas.ac.id/wp-content/uploads/2018/11/vol.-13-no.6.pdfISSN: 0215-7950...
51
ISSN 0215-7950 E-ISSN 2339-2479 Volume 13, Nomor 6, November 2017 191 199 207 216 229 Aplikasi Gelombang Mikro sebagai Pengendali Cendawan Patogen Terbawa Benih Kedelai Raden Sujayadi, Supyani, Edi Purwanto Pengaruh Elektroterapi dan Termoterapi secara in Vitro terhadap Eliminasi Onion yellow dwarf virus Siti Shofiya Nasution, Diny Dinarti, Sri Hendrastuti Hidayat Asam Salisilat sebagai Penginduksi Ketahanan Tanaman Padi terhadap Penyakit Hawar Daun Bakteri Christoffol Leiwakabessy, Meity Suradji Sinaga, Kikin Hamzah Mutaqin, Trikoesoemaningtyas, Giyanto Keanekaragaman dan Patogenisitas Fusarium spp. Asal Beberapa Kultivar Pisang Widya Sari, Suryo Wiyono, Ali Nurmansyah, Abdul Munif, Roedhy Poerwanto Kisaran Inang Cowpea mild mottle virus dan Respons Varietas Kedelai Mimi Sutrawati, Sri Hendrastuti Hidayat, Bonny Purnomo Wahyu Soekarno, Ali Nurmansyah, Gede Suastika
Transcript of in Vitro - fp.unmas.ac.idfp.unmas.ac.id/wp-content/uploads/2018/11/vol.-13-no.6.pdfISSN: 0215-7950...

ISSN 0215-7950E-ISSN 2339-2479
Volume 13, Nomor 6, November 2017
191
199
207
216
229
Aplikasi Gelombang Mikro sebagai Pengendali Cendawan Patogen Terbawa Benih Kedelai Raden Sujayadi, Supyani, Edi Purwanto
Pengaruh Elektroterapi dan Termoterapi secara in Vitro terhadap Eliminasi Onion yellow dwarf virusSiti Shofiya Nasution, Diny Dinarti, Sri Hendrastuti Hidayat
Asam Salisilat sebagai Penginduksi Ketahanan Tanaman Padi terhadap Penyakit Hawar Daun BakteriChristoffol Leiwakabessy, Meity Suradji Sinaga, Kikin Hamzah Mutaqin, Trikoesoemaningtyas, Giyanto
Keanekaragaman dan Patogenisitas Fusarium spp. Asal Beberapa Kultivar PisangWidya Sari, Suryo Wiyono, Ali Nurmansyah, Abdul Munif, Roedhy Poerwanto
Kisaran Inang Cowpea mild mottle virus dan Respons Varietas KedelaiMimi Sutrawati, Sri Hendrastuti Hidayat, Bonny Purnomo Wahyu Soekarno, Ali Nurmansyah, Gede Suastika

ISSN: 0215-7950E-ISSN: 2339-2479
Volume 13, Nomor 6, November 2017
Terakreditasi B, SK DIKTI Nomor : 12/M/Kp/II/2015
Penanggung JawabAchmadi Priyatmodjo Muhammad Taufik
Ketua Dewan PenyuntingSri Hendrastuti Hidayat
Anggota Dewan PenyuntingAbdul Gafur Gayuh Rahayu Tarkus Suganda Agus Purwantara Radix Suharjo Tri Asmira Damayanti Ali Nurmansyah Supramana Tri Puji Priyatno Baharuddin Supriadi Wiwiek Sri Wahyuni
Penyunting PelaksanaAgustin Wydia GunawanDono Wahyuno
Business ManagerIfa ManzilaWidodo
SekretariatAnik NurhayatiDepartemen Proteksi Tanaman, Fakultas Pertanian Institut Pertanian Bogor Jalan Kamper, Kampus IPB Darmaga Bogor 16680 Tel/Faks: 0251-8621267 Surel: [email protected] Homepage: http://jurnal.ipb.ac.id/index.php/jfiti
PenerbitPerhimpunan Fitopatologi IndonesiaJurnal Fitopatologi Indonesia diterbitkan pada Januari, Maret, Mei, Juli, September, dan November
Harga berlangganan (sudah termasuk ongkos kirim) untuk satu tahun:Pribadi Rp350 000.-Institusi/ Perpustakaan Rp450 000.-
BankBank Mandiri KCP Sholeh Iskandar Bogor no rek. 900 000 364 6123 a/n Ifa Manzila dan Sri Hendrastuti Hidayat



ISSN: 0215-7950
191
Volume 13, Nomor 6, November 2017Halaman 191–198
DOI: 10.14692/jfi.13.6.191
*Alamat penulis korespondensi: Program Studi Agroteknologi, Fakultas Pertanian, Universitas Sebelas Maret. Jalan Ir. Sutami No. 36-A, Kentingan, Surakarta 57126. Tel: 0271-637457, Surel: [email protected].
Aplikasi Gelombang Mikro sebagai Pengendali Cendawan Patogen Terbawa Benih Kedelai
Micro wave Application to Control Seed-borne Fungal Pathogens of Soybean
Raden Sujayadi, Supyani*, Edi Purwanto Universitas Sebelas Maret, Surakarta 57126
ABSTRAK
Salah satu kendala yang dihadapi dalam penyediaan benih kedelai (Glycine max) bermutu ialah infeksi cendawan patogen terbawa benih. Perlakuan gelombang mikro merupakan salah satu alternatif metode pengendalian cendawan terbawa benih yang cukup efektif sehingga perlu dikembangkan. Penelitian ini terdiri atas dua tahap. Tahap ke-1 bertujuan menetapkan kadar air benih kedelai yang baik untuk memertahankan mutu fisiologi benih menggunakan durasi gelombang mikro. Tahap ke-2 bertujuan menentukan durasi perlakuan gelombang mikro yang paling efektif mengurangi tingkat infeksi cendawan patogen terbawa benih menggunakan kadar air terbaik dari hasil percobaan tahap ke-1. Hasil penelitian menunjukkan bahwa kadar air 9.20% paling baik memertahankan mutu fisiologi benih setelah perlakuan gelombang mikro. Perlakuan gelombang mikro dengan durasi 60 detik mengendalikan Aspergillus flavus dengan efektif, sedangkan durasi 80 detik mengendalikan Fusarium sp., Curvularia sp., dan A. niger.
Kata kunci: Glycine max, kadar air benih, mutu fisiologi, tingkat infeksi
ABSTRACT
One of the obstacles in providing high quality soybean (Glycine max) seed is the infection of seed-borne fungal pathogens. Micro wave treatment is one of the alternative methods to control the seed-borne pathogens effectively but it needs to be further developed. This research consisted of two stages. The first stage was aimed to determine the best soybean seed moisture content in maintaining seed physiological quality after exposing to micro wave. The second stage was aimed to determine the most effective duration of micro wave exposure to reduce the rate of infection of seed-borne fungal pathogens using the best moisture content resulted from the first stage. The experiment on seed moisture indicated that 9.20% was the best seed moisture content in maintaining the physiological quality when the seed was exposed to micro wave. Furthermore, micro wave exposure of 60 seconds was able to control Aspergillus flavus, while the exposure of 80 seconds was able to control Fusarium sp., Curvularia sp., and A. niger.
Key words: Glycine max, infection rate, physiological quality, seed moisture content

J Fitopatol Indones Sujayadi et al.
192
PENDAHULUAN
Kedelai merupakan sumber protein nabati yang penting dan murah. Sebanyak 67.28% (1.96 juta ton) kebutuhan nasional kedelai dipenuhi melalui impor (Pusdatin 2015). Salah satu faktor penyebabnya ialah produktivitas kedelai nasional yang rendah, yakni 1.57 ton ha-1
pada tahun 2015 (BPS 2016). Benih bermutu dan bersertifikat
merupakan salah satu komponen utama untuk meningkatkan produksi kedelai. Salah satu aspek mutu benih ialah mutu patologi yang tinggi, artinya benih bebas dari infeksi patogen terbawa benih, baik cendawan, bakteri, virus, dan nematoda (Ilyas 2012).
Cendawan patogen terbawa benih dapat berada di permukaan, di dalam jaringan atau tercampur bebas bersama benih. Sumber inokulum cendawan berasal dari lapangan, kontaminasi saat panen, pengolahan, pengemasan, penyimpanan atau selama pendistribusian benih (Agarwal dan Sinclair 1996). Mathur dan Kongsdal (2003) melaporkan bahwa cendawan patogen yang terbawa benih kedelai diantaranya, yaitu Cercospora kikuchii, C. sojina, dan Fusarium spp. Menurut Ahmed et al.(2016), cendawan yang ditemukan pada benih kedelai antara lain Aspergillus spp., Curvularia spp., Fusarium spp., Penicillium spp., dan Phomopsis spp. Sedangkan jenis cendawan pada hasil penelitian Rao et al. (2015), antara lain M. phaseolina, C. dematium, Aspergillus flavus, A. niger, Rhizopus sp., Curvularia sp., Alternaria, Cladosporium, dan Fusarium sp.
Salah satu alternatif pengendalian cendawan patogen terbawa benih ialah perlakuan gelombang mikro, yang telah diujikan pada beberapa komoditas, antara lain kedelai (Reddy et al. 1995), aster cina (Han 2010), kacang buncis (Friesen et al. 2014), barley (Iuliana et al. 2013), dan cabai (Najah et al. 2016). Friesen et al. (2014) melaporkan paparan gelombang mikro kurang dari 50 detik dapat menurunkan tingkat infeksi patogen Colletotrichum lindemuthianum (0.14% per detik paparan), namun tetap menjaga penurunan daya berkecambah sebesar <10%. Sementara
Reddy et al. (1995) menyatakan, semakin lama durasi perlakuan gelombang mikro pada benih kedelai yang diinokulasi buatan dengan patogen Diaphorte phaseolorum, tingkat infeksi patogen semakin menurun, serta diikuti dengan penurunan daya berkecambah dan vigor benih. Penelitian lain yang dilakukan oleh Motallebi (2016) terhadap benih kanola, kedelai dan kesumba yang diinokulasi buatan dengan A. niger menunjukkan bahwa semakin tinggi daya gelombang mikro, menyebabkan viabilitas benih dan tingkat infeksi patogen A. niger semakin menurun. Pada penelitian sebelumnya, kadar air benih yang digunakan jauh lebih tinggi dari persyaratan kadar air maksimum benih kedelai bersertifikat (11%), selain itu cendawan patogen yang digunakan untuk inokulasi buatan hanya satu spesies.
Penelitian ini bertujuan untuk menentukan durasi perlakuan gelombang mikro dan kadar air yang paling efektif mengendalikan cendawan patogen terbawa benih kedelai, namun tetap dapat mempertahankan mutu fisiologis benih.
BAHAN DAN METODE
Alat yang digunakan antara lain oven micro wave komersial (LG, model MH6042D, 220V-50Hz, 2,450MHz, 450W), termometer, germinator cabinet tipe IPB 73 2A/B, oven listrik (Memmert), cawan porselen, saringan 40 mesh, lemari pendingin (Kirsch), mikroskop stereo dan majemuk, serta alat penunjang lainnya. Benih kedelai yang digunakan adalah varietas Grobogan dari Kabupaten Grobogan, Jawa Tengah, dipanen bulan Oktober 2016. Penelitian terdiri atas dua tahap.
Tahap I. Penentuan Kadar Air Benih dengan Mutu Fisiologis Terbaik Setelah Perlakuan Gelombang Mikro
Tahap I bertujuan menetapkan kadar air benih kedelai yang paling baik mempertahankan mutu fisiologis benih setelah perlakuan gelombang mikro. Kadar air benih kedelai ditetapkan menjadi tiga taraf perlakuan, yaitu kadar air rendah (9.20%), sedang (11.25%), dan tinggi (13.73%), yang

J Fitopatol Indones Sujayadi et al.
193
dihitung menggunakan angka bobot benih awal dan bobot benih akhir yang ekuivalen dengan kadar air (KA) benih yang diinginkan dengan rumus berikut (ISTA 2014):Bobot benih yang diinginkan
100 – KA awal100 – KA akhir × = bobot awal
benih
Bobot benih diturunkan dengan cara benih dijemur di bawah sinar matahari pada pukul 07.00–10.00 pagi pada wadah plastik dengan suhu maksimal 40 °C, selama 2–4 hari sampai diperoleh bobot benih yang ekuivalen dengan tiga taraf kadar air yang diinginkan. Kadar air maksimum untuk benih kedelai berdasarkan standar pengujian laboratorium ialah sebesar 11% (Kepmentan 2015).
Penetapan kadar air benih kedelai menggunakan metode oven suhu konstan tinggi (Kepmentan 2015). Sebanyak 10 g benihkedelai digerus menggunakan penggiling, disaring menggunakan saringan 40 mesh hingga diperoleh sebanyak 4.5–5 g butiran kasar, selanjutnya dimasukkan ke dalam cawan porselen untuk ditimbang beserta tutupnya. Cawan beserta tutupnya yang berisi butiran kasar dimasukkan ke dalam oven listrik dengan suhu konstan tinggi 130 ± 2 °C selama 1 jam. Setelah itu, cawan didinginkan dalam desikator selama 30 menit dan ditimbang berat kering benih beserta cawan dan tutupnya yang dihitung dengan rumus berikut:
KA = M2 – M3M2 – M1 × 100%, dengan
KA, kadar air (%); M1, berat cawan kosong; M2, berat awal (benih dan cawan sebelum dipanaskan dalam oven); M3, berat akhir (benih dan cawan setelah dipanaskan dalam oven).
Benih selanjutnya diberi perlakuan gelombang mikro menggunakan oven gelombang mikro frekuensi 2.450 MHz/daya 450 W (Aladjadjiyan 2010). Dimasukkan pula beaker glass berisi air sebanyak 200 mLuntuk mencegah kerusakan benih (Tylkowska et al. 2010). Durasi perlakuan gelombang mikro ialah 0, 20, 40, 60, 80, 100 detik. Benih selanjutnya diuji mutu fisiologisnya menggunakan metode uji antar kertas digulung (Kepmentan 2015). Sebanyak 400 butirbenih kedelai untuk setiap perlakuan
(4 ulangan, masing-masing ulangan 100 butir)ditanam pada kertas buram yang telah dilembapkan dengan air steril, lalu digulung dan ditempatkan dalam germinator cabinet tipe IPB 73 2A/B pada suhu kamar.
Parameter mutu fisiologis yang diamati ialah viabilitas dan vigor benih. Viabilitas dengan tolok ukur daya berkecambah (DB) dan potensi tumbuh maksimum (PTM) dihitung menggunakan rumus berikut (ISTA 2014):
∑ benih yang ditanamDB = × 100%∑ KN I+ ∑ KN II
∑ benih yang ditanamPTM = × 100%, dengan∑ benih yang tumbuh
DB, daya berkecambah; Σ KN I, jumlah kecambah normal pada 5 hari setelah tanam (HST); Σ KN II, jumlah kecambah normal pada 8 HST; dan PTM, potensi tumbuh maksimum.
Parameter vigor benih diamati dengan tolok ukur indeks vigor (IV) dan kecepatan tumbuh (KCT). Pengamatan KCT dilakukan setiap hari sampai hari ke-8. Perhitungan IV dan KCT menggunakan rumus berikut.
∑ benih yang ditanamIV = × 100%∑ KN I
KCT etmalKN( )% = ∑ tn
0 tN , dengan
IV, indeks vigor; Σ KN I, jumlah kecambah normal pada 5 HST; KCT, kecepatan tumbuh; N, persentase kecambah normal setiap waktu pengamatan; dan tn, waktu akhir pengamatan (hari ke-8).
Percobaan ini disusun dalam rancangan acak lengkap (RAL) faktorial. Setiap perlakuan diulang sebanyak 4 kali. Kadar air benih yang paling baik dalam memertahankan mutu fisiologisnya setelah diberi perlakuan gelombang mikro digunakan sebagai kadar air benih acuan untuk tahap II.
Tahap II. Pengaruh Durasi Perlakuan Gelombang Mikro terhadap Tingkat Infeksi Cendawan Patogen Terbawa Benih Kedelai
Tahap ini bertujuan menentukan durasi perlakuan gelombang mikro yang paling baik

J Fitopatol Indones Sujayadi et al.
194
menurunkan tingkat infeksi cendawan terbawa benih. Sebanyak 200 butir benih kedelai (4 ulangan, masing-masing sebanyak 50 butir) diberi perlakuan gelombang mikro selama 0, 20, 40, 60, 80, 100 detik. Selanjutnya dilakukan uji kesehatan benih menggunakan metode blotter test (Mathur dan Kongsdal 2003). Benih disterilisasi permukaan dengan cara direndam dalam larutan NaOCl 1%selama 1 menit, kemudian dibilas air steril 3 kali, dikeringanginkan di atas kertas saring steril pada kondisi aseptik. Selanjutnya, sebanyak 25 butir benih ditanam pada cawan petri berisi 3 lembar kertas saring yang telah dilembapkan dengan air steril. Cawan petri tersebut ditutup dan disegel plastik wrap, lalu diinkubasi dalam inkubator pada suhu 20–25 °Cselama 24 jam dengan penyinaran near ultra violet (NUV) 12 jam terang dan 12 jam gelapsecara bergantian. Benih kemudian diletakkan pada lemari pendingin pada suhu -20 °C selama 24 jam dan diinkubasi kembali dalam inkubator pada suhu 20-25 °C selama 5 hari dengan penyinaran NUV selama 12 jam terang dan 12 jam gelap secara bergantian. Pada hari ke-8 setelah tanam dilakukan pengamatan menggunakan mikroskop stereo dan majemuk untuk deteksi dan identifikasi cendawan yang tumbuh pada benih kedelai serta dihitung jumlah benih yang terinfeksi masing-masing cendawan.
Identifikasi spesies cendawan terbawa benih berdasarkan tipe pertumbuhan dan morfologi struktur cendawan seperti hifa, tubuh buah, spora, konidium dan koloni menggunakan acuan yang dibuat oleh Mathur dan Kongsdal (2003) dan Mathur dan Manandhar (2003).
Tingkat infeksi cendawan pada benih dihitung menggunakan rumus berikut:
∑ benih yang ditanam × 100%∑ benih terinfeksi
Tingkat infeksi =
Rancangan yang digunakan pada percobaan ini RAL satu faktor, yaitu durasi perlakuan gelombang mikro dengan 4 ulangan untuk setiap perlakuan.
Analisis DataData dianalisis menggunakan uji F untuk
mengetahui pengaruh tiap perlakuan. Apabila hasil analisis menunjukkan adanya pengaruh nyata dari perlakuan, akan diuji lanjut dengan DMRT pada α 5%.
HASIL
Kadar Air Benih Kedelai dengan Mutu Fisiologis Terbaik Setelah Perlakuan Gelombang Mikro
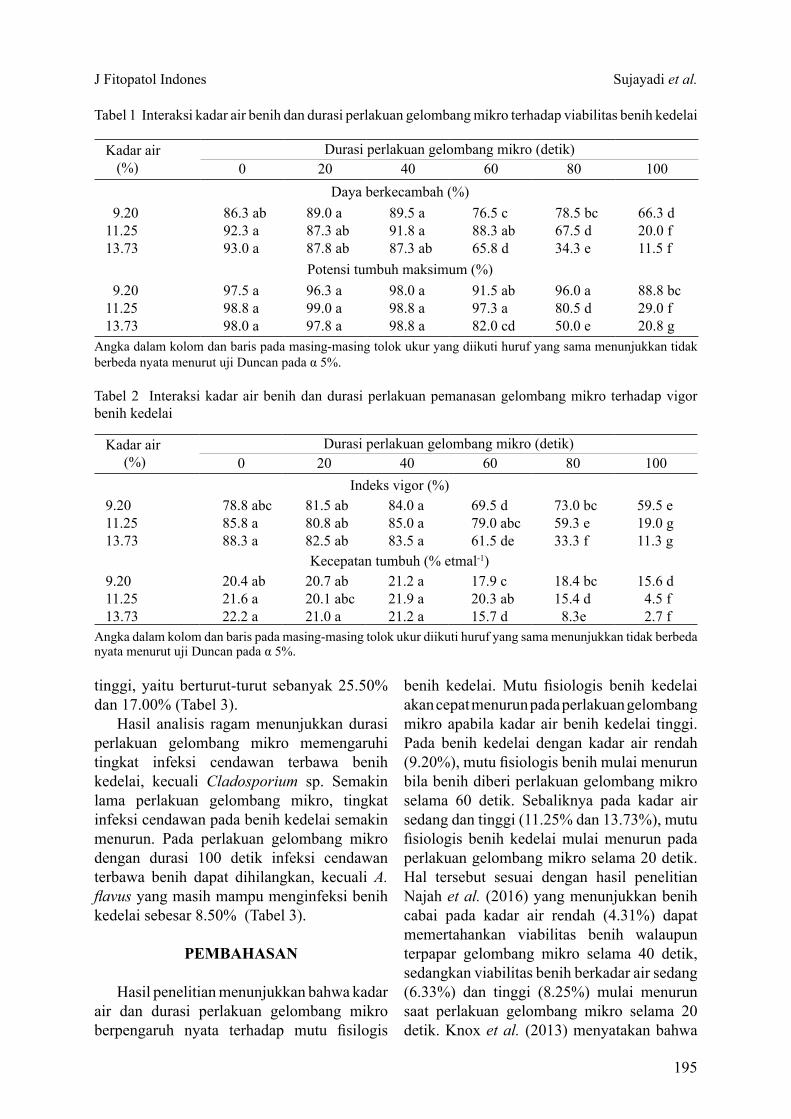
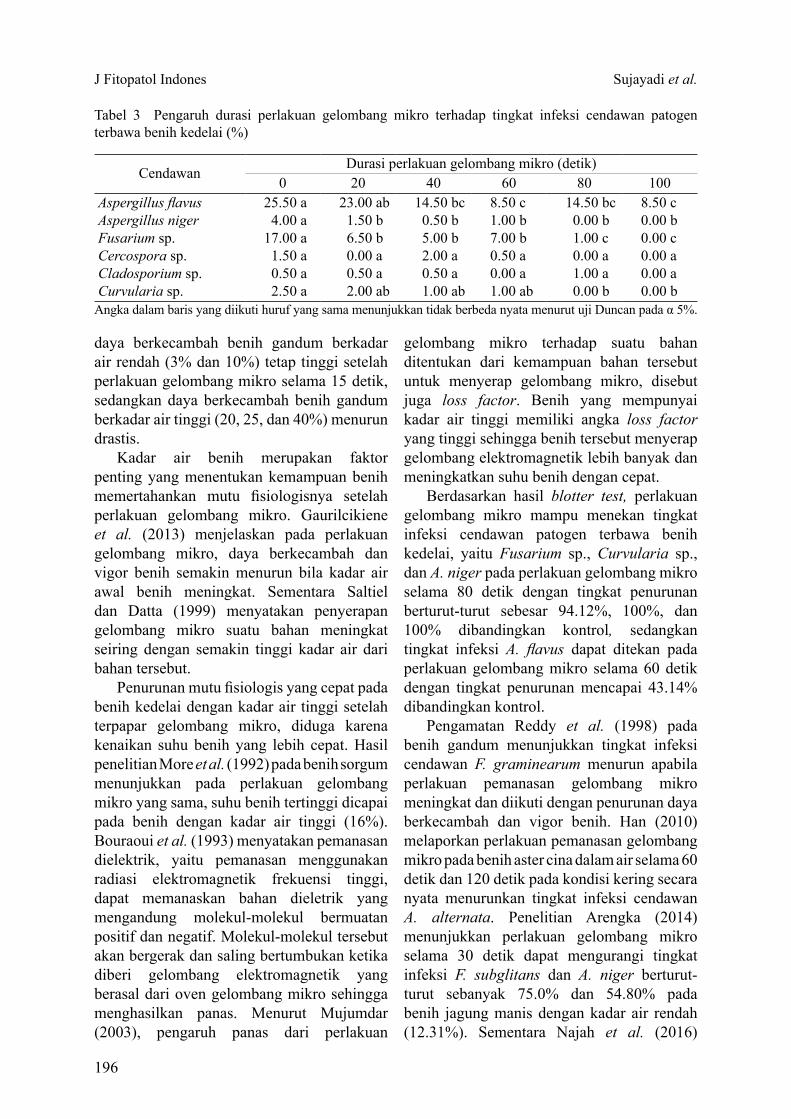
Hasil analisis ragam interaksi pengaruh kadar air dan durasi perlakuan gelombang mikro menunjukkan pengaruh nyata terhadap mutu fisiologis benih kedelai. Semakin lama durasi gelombang mikro, maka daya berkecambah benih semakin menurun. Pada kadar air yang rendah, benih mampu berkecambah lebih baik apabila durasi perlakuan gelombang mikro meningkat (Tabel 1).Durasi paparan 100 detik merupakan perlakuan gelombang mikro yang mematikan sehingga menurunkan daya berkecambah (DB) dan potensi tumbuh maksimum (PTM) benih dengan kadar air rendah (9.20%), sedangkan pada kadar air benih sedang (11.25%) dan kadar air benih tinggi (13.73%) durasi perlakuan gelombang mikro yang mematikan terjadi pada 80 detik dan 60 detik (Tabel 1). Pengaruh durasi perlakuan gelombang mikro tersebut, terjadi pula terhadap vigor benih kedelai, yaitu indeks vigor (IV) dan kecepatan tumbuh benih kedelai (Tabel 2).
Pengaruh Durasi Perlakuan Gelombang Mikro terhadap Tingkat Infeksi Cendawan Patogen Terbawa Benih Kedelai
Hasil uji kesehatan benih dengan blotter test menunjukkan sebanyak 6 spesies cendawan patogen terbawa benih kedelai terdeteksi dan teridentifikasi, yaitu Fusarium sp., Cladosporium sp., Curvularia sp., Cercospora sp., A. flavus dan A. niger. Dari semua cendawan yang ditemukan Fusarium sp. dan A. flavus merupakan cendawan terbawa benih kedelai dengan tingkat infeksi
J Fitopatol Indones Sujayadi et al.
195
tinggi, yaitu berturut-turut sebanyak 25.50% dan 17.00% (Tabel 3).
Hasil analisis ragam menunjukkan durasi perlakuan gelombang mikro memengaruhi tingkat infeksi cendawan terbawa benih kedelai, kecuali Cladosporium sp. Semakin lama perlakuan gelombang mikro, tingkat infeksi cendawan pada benih kedelai semakin menurun. Pada perlakuan gelombang mikro dengan durasi 100 detik infeksi cendawan terbawa benih dapat dihilangkan, kecuali A. flavus yang masih mampu menginfeksi benih kedelai sebesar 8.50% (Tabel 3).
PEMBAHASAN
Hasil penelitian menunjukkan bahwa kadar air dan durasi perlakuan gelombang mikro berpengaruh nyata terhadap mutu fisilogis
benih kedelai. Mutu fisiologis benih kedelai akan cepat menurun pada perlakuan gelombang mikro apabila kadar air benih kedelai tinggi. Pada benih kedelai dengan kadar air rendah (9.20%), mutu fisiologis benih mulai menurun bila benih diberi perlakuan gelombang mikro selama 60 detik. Sebaliknya pada kadar air sedang dan tinggi (11.25% dan 13.73%), mutu fisiologis benih kedelai mulai menurun pada perlakuan gelombang mikro selama 20 detik. Hal tersebut sesuai dengan hasil penelitian Najah et al. (2016) yang menunjukkan benih cabai pada kadar air rendah (4.31%) dapat memertahankan viabilitas benih walaupun terpapar gelombang mikro selama 40 detik, sedangkan viabilitas benih berkadar air sedang (6.33%) dan tinggi (8.25%) mulai menurun saat perlakuan gelombang mikro selama 20 detik. Knox et al. (2013) menyatakan bahwa
Tabel 1 Interaksi kadar air benih dan durasi perlakuan gelombang mikro terhadap viabilitas benih kedelai
Angka dalam kolom dan baris pada masing-masing tolok ukur yang diikuti huruf yang sama menunjukkan tidak berbeda nyata menurut uji Duncan pada α 5%.
Kadar air (%)
Durasi perlakuan gelombang mikro (detik)0 20 40 60 80 100
Daya berkecambah (%) 9.20 86.3 ab 89.0 a 89.5 a 76.5 c 78.5 bc 66.3 d11.25 92.3 a 87.3 ab 91.8 a 88.3 ab 67.5 d 20.0 f13.73 93.0 a 87.8 ab 87.3 ab 65.8 d 34.3 e 11.5 f
Potensi tumbuh maksimum (%) 9.20 97.5 a 96.3 a 98.0 a 91.5 ab 96.0 a 88.8 bc11.25 98.8 a 99.0 a 98.8 a 97.3 a 80.5 d 29.0 f13.73 98.0 a 97.8 a 98.8 a 82.0 cd 50.0 e 20.8 g
Tabel 2 Interaksi kadar air benih dan durasi perlakuan pemanasan gelombang mikro terhadap vigor benih kedelai
Angka dalam kolom dan baris pada masing-masing tolok ukur diikuti huruf yang sama menunjukkan tidak berbeda nyata menurut uji Duncan pada α 5%.
Kadar air (%)
Durasi perlakuan gelombang mikro (detik)0 20 40 60 80 100
Indeks vigor (%)9.20 78.8 abc 81.5 ab 84.0 a 69.5 d 73.0 bc 59.5 e11.25 85.8 a 80.8 ab 85.0 a 79.0 abc 59.3 e 19.0 g13.73 88.3 a 82.5 ab 83.5 a 61.5 de 33.3 f 11.3 g
Kecepatan tumbuh (% etmal-1)9.20 20.4 ab 20.7 ab 21.2 a 17.9 c 18.4 bc 15.6 d11.25 21.6 a 20.1 abc 21.9 a 20.3 ab 15.4 d 4.5 f13.73 22.2 a 21.0 a 21.2 a 15.7 d 8.3e 2.7 f
J Fitopatol Indones Sujayadi et al.
196
Tabel 3 Pengaruh durasi perlakuan gelombang mikro terhadap tingkat infeksi cendawan patogen terbawa benih kedelai (%)
Angka dalam baris yang diikuti huruf yang sama menunjukkan tidak berbeda nyata menurut uji Duncan pada α 5%.
CendawanDurasi perlakuan gelombang mikro (detik)
0 20 40 60 80 100Aspergillus flavus 25.50 a 23.00 ab 14.50 bc 8.50 c 14.50 bc 8.50 cAspergillus niger 4.00 a 1.50 b 0.50 b 1.00 b 0.00 b 0.00 bFusarium sp. 17.00 a 6.50 b 5.00 b 7.00 b 1.00 c 0.00 cCercospora sp. 1.50 a 0.00 a 2.00 a 0.50 a 0.00 a 0.00 aCladosporium sp. 0.50 a 0.50 a 0.50 a 0.00 a 1.00 a 0.00 aCurvularia sp. 2.50 a 2.00 ab 1.00 ab 1.00 ab 0.00 b 0.00 b
daya berkecambah benih gandum berkadar air rendah (3% dan 10%) tetap tinggi setelah perlakuan gelombang mikro selama 15 detik, sedangkan daya berkecambah benih gandum berkadar air tinggi (20, 25, dan 40%) menurun drastis.
Kadar air benih merupakan faktor penting yang menentukan kemampuan benih memertahankan mutu fisiologisnya setelah perlakuan gelombang mikro. Gaurilcikiene et al. (2013) menjelaskan pada perlakuan gelombang mikro, daya berkecambah dan vigor benih semakin menurun bila kadar air awal benih meningkat. Sementara Saltiel dan Datta (1999) menyatakan penyerapan gelombang mikro suatu bahan meningkat seiring dengan semakin tinggi kadar air dari bahan tersebut.
Penurunan mutu fisiologis yang cepat pada benih kedelai dengan kadar air tinggi setelah terpapar gelombang mikro, diduga karena kenaikan suhu benih yang lebih cepat. Hasil penelitian More et al. (1992) pada benih sorgum menunjukkan pada perlakuan gelombang mikro yang sama, suhu benih tertinggi dicapai pada benih dengan kadar air tinggi (16%). Bouraoui et al. (1993) menyatakan pemanasan dielektrik, yaitu pemanasan menggunakan radiasi elektromagnetik frekuensi tinggi, dapat memanaskan bahan dieletrik yang mengandung molekul-molekul bermuatan positif dan negatif. Molekul-molekul tersebut akan bergerak dan saling bertumbukan ketika diberi gelombang elektromagnetik yang berasal dari oven gelombang mikro sehingga menghasilkan panas. Menurut Mujumdar (2003), pengaruh panas dari perlakuan
gelombang mikro terhadap suatu bahan ditentukan dari kemampuan bahan tersebut untuk menyerap gelombang mikro, disebut juga loss factor. Benih yang mempunyai kadar air tinggi memiliki angka loss factor yang tinggi sehingga benih tersebut menyerap gelombang elektromagnetik lebih banyak dan meningkatkan suhu benih dengan cepat.
Berdasarkan hasil blotter test, perlakuan gelombang mikro mampu menekan tingkat infeksi cendawan patogen terbawa benih kedelai, yaitu Fusarium sp., Curvularia sp., dan A. niger pada perlakuan gelombang mikro selama 80 detik dengan tingkat penurunan berturut-turut sebesar 94.12%, 100%, dan 100% dibandingkan kontrol, sedangkan tingkat infeksi A. flavus dapat ditekan pada perlakuan gelombang mikro selama 60 detik dengan tingkat penurunan mencapai 43.14% dibandingkan kontrol.
Pengamatan Reddy et al. (1998) pada benih gandum menunjukkan tingkat infeksi cendawan F. graminearum menurun apabila perlakuan pemanasan gelombang mikro meningkat dan diikuti dengan penurunan daya berkecambah dan vigor benih. Han (2010) melaporkan perlakuan pemanasan gelombang mikro pada benih aster cina dalam air selama 60 detik dan 120 detik pada kondisi kering secara nyata menurunkan tingkat infeksi cendawan A. alternata. Penelitian Arengka (2014) menunjukkan perlakuan gelombang mikro selama 30 detik dapat mengurangi tingkat infeksi F. subglitans dan A. niger berturut-turut sebanyak 75.0% dan 54.80% pada benih jagung manis dengan kadar air rendah (12.31%). Sementara Najah et al. (2016)

J Fitopatol Indones Sujayadi et al.
197
melaporkan perlakuan gelombang mikro selama 40 detik pada benih cabai dengan kadar air rendah efektif menurunkan tingkat infeksi C. acutatum dengan tingkat efikasi mencapai 64.3%.
Perlakuan gelombang mikro dapat digunakan sebagai alternatif dalam mengendalikan cendawan terbawa benih kedelai. Kadar air benih rendah (9.20%) paling baik memertahankan mutu fisiologis benih kedelai setelah diberi perlakuan gelombang mikro. Durasi perlakuan gelombang mikro selama 80 detik paling efektif mengendalikan Fusarium sp., Curvularia sp., dan A. niger, sedangkan perlakuan 60 detik dapat menurunkan tingkat infeksi A. flavus.
UCAPAN TERIMA KASIH
Penulis menyampaikan terima kasih kepada Badan Penyuluhan dan Pengembangan SDM Pertanian, Kementrian Pertanian yang telah memberikan beasiswa pendidikan dan Balai Besar Pengembangan Pengujian Mutu Benih Tanaman Pangan dan Hortikultura (BB-PPMBTPH) atas segala dukungan dalam pelaksanaan penelitian.
DAFTAR PUSTAKA
Agarwal VK, Sinclair JB. 1996. Principles of Seed Pathology. Ed ke-2. Florida (US):CRC Press.
Ahmed O, Balogun OS, Fawole OB, Fabiyi OA, Hussein AT, Kassoum KO. 2016. Seed-borne fungi of soybeans (Glycine max [l.] Merr) in the guinea savannah agroecology of nigeria. J Agric Sci. 61(1): 57–68. DOI: http://dx.doi.org/10.2298/JAS1601057A.
Aladjadjiyan A. 2010. Effect of microwave irradiation on seeds of lentils (Lens culinaris, Med.). Rom J Biophys. 20(3):213–221.
Arengka D. 2014. Pemanfaatan gelombang mikro untuk mengendalikan patogen terbawa benih jagung manis (Zea mays saccharata Sturt.) [tesis]. Bogor (ID): Institut Pertanian Bogor.
Bouraoui M, Richard P, Fichtali J. 1993. A review of moisture content determination in foods using microwave oven drying. Food Res Int. 26: 49–57. DOI: http://dx.doi.org/10.1016/0963-9969(93)90105-R.
[BPS] Badan Pusat Statistik. 2016. Produktivitas kedelai menurut provinsi (kuintal/ha), 1993–2015. http://www.bps.go.id [diakses 3 Sep 2016].
Friesen AP, Conner RL, Robinson DE, Barton WR, Gillard CL. 2014. Effect of microwave radiation on dry bean seed infected with Colletotrichum lindemuthianum with and without the use of chemical seed treatment. Can J Plant Sci. 94(8):1373–1384. DOI: http://dx.doi.org/10.4141/cjps-2014-035.
Gaurilcikiene I, Ramanauskiene J, Dagys M,Simniskis R, Dabkevicius Z, Suproniene S. 2013. The effect of strong micro wave electric field radiation on: (2) wheat (Triticum aestivum L.) seed germination and sanitation. Zemdirb Agric. 100(2):185–190. DOI: http://dx.doi.org/10.13080/z-a.2013.100.024.
Han F. 2010. The effect of microwave treatment on germination, vigour and health of china aster (Callistephus chinensis Nees.) seeds. J Agric Sci. 2(4):201–210. DOI: http://dx.doi.org/10.5539/jas.v2n4p201.
Ilyas S. 2012. Ilmu dan Teknologi Benih (Teori dan Hasil-hasil Penelitian). Bogor (ID): IPB Press.
Iuliana C, Giancarla V, Sorina R. 2013. The effect of microwave irradiation on the germination of barley seeds (Hordeum vulgare L.). Di dalam: 48th Croatian & 8th International Symposium on Agriculture; 2013 Feb 17–22; Dubrovnik (HR): Josip Juraj Strossmayer University of Osijek. hlm 254–258.
[ISTA] International Seed Testing Association. 2014. International Rules for Seed Testing. 2014 edition. Zürich (CH): ISTA.
[Kepmentan] Keputusan Menteri Pertanian RI. 2015. Keputusan Menteri Pertanian RI No. 635/HK.150/C/07/2015 tentang Pedoman Teknis Pengambilan Contoh Benih dan Pengujian/Analisis Mutu Benih Tanaman Pangan.

J Fitopatol Indones Sujayadi et al.
198
[Pusdatin] Pusat Data dan Sistem Informasi Pertanian Kementerian Pertanian. 2015. Outlook komoditas pertanian subsektor tanaman pangan: kedelai. http://epublikasi.setjen.pertanian.go.id/arsip-outlook/81-outlook-tanaman-pangan [diakses 5 Sep 2016].
Rao TV, Rajeswari B, Keshavulu K, Varma VS. 2015. Studies on seedborne fungi of soybean. SSRG J Agric Env Sci. 2(1):16–24.
Reddy MVB, Kushalappa AC, Raghavan GSV, Stephenson MMP. 1995. Use of microwave energy for the eradication of seedborne Diaporthe phaseolorum in soybean and its effect on seed quality. J Microw Power Electromagn Energy. 30(4):199–204.
Reddy MVB, Raghavan GSV, Kushalappa AC, Paulitz TC. 1998. Effect of microwave treatment on quality of wheat seeds infected with Fusarium graminearum. J agric Engng Res. 71:113–117. DOI: http://dx.doi.org/10.1006/jaer.1998.0305.
Saltiel C, Datta AK. 1999. Heat and mass transfer in microwave processing. Adv heat transfer. 33:1–94. DOI: http://dx.doi.org/10.1016/S0065-2717(08)70303-4.
Knox OGG, McHugh MJ, Fountaine JM, Havis ND. 2013. Effects of microwaves on fungal pathogens of wheat seed. Cropping Systems Research Group. Scotland's Rural College (SAC). United Kingdom. Crop Protection. 50:12–16. DOI: http://dx.doi.org/10.1016/j.cropro.2013.03.009.
Mathur SB, Kongsdal O. 2003. Common Laboratory Seed Health Testing Methods for Detecting Fungi. Ed ke-1. Basserdorf (CH): ISTA.
Mathur SB, Manandhar HK. 2003. Fungi in Seeds. Ed ke-1. Copenhagen (DK): DGISP.
More HG, Magan N, Stenning BC. 1992. Effect of microwave heating on quality and mycoflora of sorghum grain. J Stored Prod Res. 28(4): 251–256. DOI: http://dx.doi.org/10.1016/0022-474X(92)90005-B.
Motallebi A. 2016. Effect of microwave radiation on seed viability, survival of Aspergillus niger van tieghem and oil quality of oilseeds crops canola, soybean and safflower. Acta Agric Slov. 107(1):73–80. DOI: http://dx.doi.org/10.14720/aas.2016.107.1.08.
Mujumdar AS. 2003. Drying Technology in Agriculture and Food Sciences. Enfield(US): Science Publishers, Inc.
Najah LN, Suhartanto MR, Widodo. 2016. Pengendalian Colletotrichum spp. terbawa benih cabai dengan paparan gelombang mikro. J Fitopatol Indones. 12(4):115–123. DOI: http://dx.doi.org/10.14692/jfi.12.4.115.

ISSN: 0215-7950
199
Volume 13, Nomor 6, November 2017Halaman 199–206
DOI: 10.14692/jfi.13.6.199
*Alamat penulis korespondensi: Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor, Jalan Kamper, Kampus Darmaga IPB, Bogor 16680Tel: 0251-8629364, Faks: 0251-8629362, Surel: [email protected]
Pengaruh Elektroterapi dan Termoterapi secara in Vitro terhadap Eliminasi Onion yellow dwarf virus
The Effect of in Vitro Electrotherapy and Thermotherapy on
Elimination of Onion yellow dwarf virus
Siti Shofiya Nasution, Diny Dinarti, Sri Hendrastuti Hidayat* Institut Pertanian Bogor, Bogor 16680
ABSTRAK
Onion yellow dwarf virus (OYDV) pada tanaman bawang putih telah dilaporkan dapat menyebabkan masalah pada produksi bawang putih. Penggunaan benih bawang putih bebas virus diharapkan dapat menekan insidensi penyakit di lapangan. Tujuan penelitian ini ialah mengembangkan metode eliminasi OYDV pada bahan perbanyakan umbi bawang putih melalui kombinasi perlakuan elektroterapi (0, 5, 10, 15, dan 20 mA selama 10 menit) dan termoterapi (23, 28, 33, dan 38 °C selama 4 minggu). Bawang putih kultivar Sangga Sembalun dan Lumbu Hijau digunakan sebagai bahan tanaman untuk eliminasi OYDV. Konfirmasi adanya infeksi virus dilakukan melalui metode reverse transcription-polymerase chain reaction (RT-PCR). Hasil penelitian menunjukkan bahwa termoterapi pada suhu 33 °C merupakan perlakuan terbaik untuk mengeliminasi OYDV pada umbi bawang putih meskipun tingkat efisiensi berbeda pada setiap kultivar. Efisiensi eliminasi OYDV mencapai 60% pada kultivar Lumbu Hijau, dan 40% pada kultivar Sangga Sembalun. Perlakuan elektroterapi dan kombinasinya dengan termoterapi belum dapat menghasilkan planlet bebas OYDV.
Kata kunci: Lumbu Hijau, Sangga Sembalun, planlet bebas OYDV, RT-PCR
ABSTRACT
Infection of Onion yellow dwarf virus (OYDV) are reported causing problems in garlic production. Planting virus-free bulbs might help reduce viral disease incidence in the field. This research was aimed to develop method for eliminating OYDV from garlic bulbs using combination of electrotherapy (0, 5, 10, 15, and 20 mA each for 10 minutes) and thermotherapy (23, 28, 33, 38 °C each for 4 weeks). Two garlic cultivars, i.e. Sangga Sembalun and Lumbu Hijau were used as seed bulbs for OYDV elimination tests. Virus infection was confirmed using transcription-polymerase chain reaction (RT-PCR). The result showed that thermotherapy at 33 °C was the best method to eliminate OYDV in garlic although the efficiency was not the same for all cultivars. The efficiency reached up to 60% for cv. Lumbu Hijau, whereas for cv. Sangga Sembalun only reached up to 40%. Electrotherapy alone or in combination with thermotherapy were not able to produce OYDV-free plantlets.
Key words: Lumbu Hijau, Sangga Sembalun, OYDV-free plantlets, RT-PCR

J Fitopatol Indones Nasution et al.
200
PENDAHULUAN
Bawang putih (Allium sativum) merupakan salah satu komoditas hortikultura yang digunakan sebagai bumbu di sebagian besar masakan Indonesia. Bawang putih diperbanyak secara vegetatif sehingga diduga tidak ada klon atau benih yang bebas dari penyakit terbawa benih. Beberapa jenis patogen diketahui dapat menular melalui perbanyakan tanaman vegetatif, termasuk virus. Menurut Diekmann (1997), kelompok virus yang umum menginfeksi tanaman bawang putih berasal dari genus Carlavirus, Potyvirus, dan Allexivirus. Beberapa jenis virus yang menginfeksi bawang putih dapat menyebabkan kehilangan hasil, khususnya Onion yellow dwarf virus (OYDV).
Infeksi OYDV pada bawang putih menurunkan kualitas dan hasil panen di Meksiko (Moreno et al. 2014). Infeksi OYDV pada tanaman bawang putih dapat menyebabkan kehilangan hasil sebesar 60% (Lot et al. 1998). Elnagar et al. (2009) melaporkan infeksi OYDV menurunkan bobot umbi bawang putih di Mesir hingga 58.78%. Sampai saat ini belum pernah dilaporkan pengaruh infeksi virus terhadap kehilangan hasil panen pada bawang putih di Indonesia.
Umbi bebas virus dapat diperoleh melalui penerapan teknik kultur jaringan dan efisiensi bebas virus dapat ditingkatkan melalui kombinasi beberapa metode eliminasi virus seperti elektroterapi dan termoterapi. Soliman et al. (2012) mengeliminasi OYDV dari umbi bawang putih melalui elektroterapi dengan perlakuan aliran listrik 15 mA selama 10 menit dan tidak menyebabkan gangguan pertumbuhan planlet. Torres et al. (2000) berhasil mengeliminasi virus dari umbi bawang putih menggunakan udara panas bersuhu 37 °C selama 35 hari dan tidak menyebabkan gangguan pertumbuhan planlet. Pengaruh elektroterapi dan kombinasi antara elektroterapi dan termoterapi terhadap eliminasi virus belum pernah dilakukan di Indonesia. Penelitian ini bertujuan untuk mendapatkan metode eliminasi OYDV pada bahan perbanyakan bawang putih yang efektif
melalui aplikasi kombinasi elektroterapi dan termoterapi secara in vitro.
BAHAN DAN METODE
Sampel umbi bawang putih diperoleh dari penangkar umbi bawang putih. Bawang putih kultivar Sangga Sembalun berasal dari Nusa Tenggara Barat dan kultivar Lumbu Hijau berasal dari Jawa Timur. Pemilihan sampel umbi bawang putih berdasarkan pengujian menggunakan metode DIBA pada penelitian sebelumnya (Nasution et al. 2016).
Eliminasi Virus dari Umbi Bawang PutihElektroterapi. Umbi bawang putih
yang digunakan ialah umbi yang positif terinfeksi OYDV. Umbi dipecah menjadi siung individual kemudian direndam dengan larutan fungisida dan bakterisida 2 g L-1. Siung dibelah dan diambil bagian tunas adventif yang digunakan sebagai bahan perlakuan. Tunas adventif direndam dalam chamber elektroforesis yang berisi tris borate EDTA (TBE) 0.5×. Tunas tersebut diberi perlakuan arus elektroterapi pada 5 taraf perlakuan, yaitu 0, 5, 10, 15, dan 20 mA masing-masing selama 10 menit. Setelah perlakuan elektroterapi, tunas adventif digunakan sebagai bahan tanaman untuk kultur jaringan.
Kultur Jaringan dan Termoterapi. Tunas adventif yang telah diberi perlakuan elektroterapi disterilisasi menggunakan NaOCL 1% selama 10 menit, kemudian dipindah ke dalam NaOCl 0.5% selama 5 menit,setelah itu dibilas air steril. Tunas pada bagian basal dipotong sekitar 2 mm yang digunakan sebagai eksplan untuk dikulturkan pada medium murashige-skoog (MS) yang me-ngandung 2ip (2 mg L-1) dan GA3 (0.3 mg L-1).Eksplan diinkubasi pada ruang kultur dan diberikan termoterapi dengan 4 taraf perlakuan, yaitu 23, 28 , 33 , dan 38 °C masing-masing selama 4 minggu. Setelah 4 minggu, tunas dipindahkan ke medium ½ MS yang mengandung NAA (1 mg L-1) untuk menginisiasi akar selama 2 minggu. Planlet yang terbentuk dan sudah berakar siap diaklimatisasi. Aklimatisasi dilakukan dengan

J Fitopatol Indones Nasution et al.
201
merendam planlet dalam larutan fungisida dan bakterisida 2 g L-1 selama 5 menit, kemudian planlet ditiriskan di atas kertas serap steril. Planlet ditanam pada medium arang sekam steril selama 2 minggu di laboratorium. Deteksi virus dilakukan pada tanaman bawang putih hasil aklimatisasi menggunakan metode reverse-transcription polymerase chain reaction (RT-PCR).
Metode RT-PCRMetode RT-PCR diawali dengan
mengisolasi total RNA dari 0.1 g jaringan daun komposit 4 botol kultur dalam 1 ulangan menggunakan bufer ekstraksi ditambahkan 1% merkaptoetanol (Doyle dan Doyle 1987). Sintesis complementary DNA (cDNA) dilakukan dengan menyiapkan 10 µL reaksi yang mengandung 3 µL RNA, 1 µL primer Potyvirus, 2 µL 50 mM DTT, 2 µL 5x buffer RT, 1 µL 10 mM dNTP, 0.5 µL ribolock, 0.35 µL MMuLV, 0.15 µL water free nuclease. Reaksi transkripsi balik (RT) dilakukan dalam mesin automated thermal cycler (Gene Amp PCR System 9700; PE Applied Biosystem, USA) yang diprogram untuk satu siklus pada suhu 65 °C (5 menit), 42 °C (60 menit), dan 70 °C (10 menit). Primer yang digunakan untuk amplifikasi PCR merupakan primer spesifik OYDV dengan ukuran target 601 pb, OYDVF (5’-CGAAGCAAATTGCCAAGCAG-3’) dan OYDVR (5’-CGATTAGCTGCCCCTCT AAC-3’) (Mahmoud et al. 2008). Reaksi amplifikasi (25 µL) terdiri atas 9.5 µL dH2O, 12.5 µL GTG master mix, 1 µL masing-masing primer (forward-reverse) dan 1 µL cDNA. Amplifikasi cDNA menggunakan mesin GeneAmp PCR System 9700 dengan kondisi reaksi denaturasi awal pada suhu 94 °C selama 3 menit, sebanyak 35 siklus pada suhu denaturasi 94 °C selama 30 detik; penempelan primer 52 °C selama 1 menit; ekstensi 72 °C selama 1 menit; dan ekstensi akhir 72 °C selama 7 menit. Amplikon yang diperoleh dielektroforesis pada tegangan 50 volt selama 50 menit dalam 1% tris borate EDTA gel agarosa, selanjutnya gel agarosa direndam dalam etidium bromida (EtBr) lalu divisualisasi di bawah UV transilluminator.
Rancangan Percobaan dan Analisis DataPengujian disusun dalam rancangan
percobaan petak terbagi. Perlakuan termoterapi (23, 28, 33, dan 38 °C) sebagai petak utama dan perlakuan elektroterapi (0, 5, 10, 15, dan 20 mA) sebagai anak petak. Masing-masing percobaan dilakukan dengan 3 ulangan, tiap ulangan terdiri atas 4 botol kultur dan tiap botol kultur terdapat 1 eksplan. Peubah yang diamati ialah ersentase hidup eksplan, jumlah daun, dan tinggi planlet. Data penelitian dianalisis ragam (ANOVA) dan perlakuan yang berpengaruh nyata diuji lanjut dengan uji jarak berganda Duncan (DMRT) pada taraf 5%.
HASIL
Termoterapi dan Elektroterapi pada Persentase Hidup Eksplan, Jumlah Daun dan Tinggi Planlet
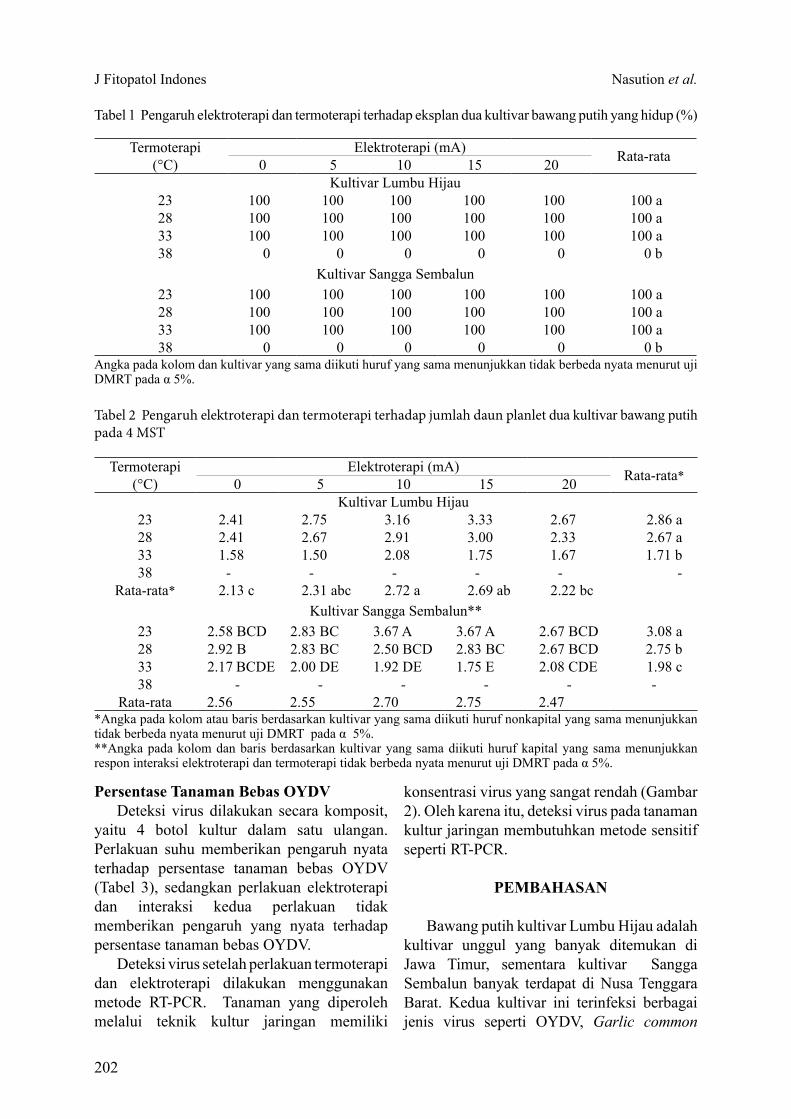
Pertumbuhan eksplan sampai dengan 4 minggu setelah tanam (MST) pada kultivar Lumbu Hijau dan Sangga Sembalun mencapai 100%, kecuali pada perlakuan suhu 38 °C (Tabel 1). Eksplan yang diberi perlakuan suhu 38 °C berubah warna menjadi putih dan mati. Hasil analisis menunjukkan bahwa termoterapi berpengaruh nyata terhadap persentase hidup eksplan bawang putih, sedangkan elektroterapi dan interaksi kedua perlakuan tidak berbeda nyata terhadap persentase hidup eksplan bawang putih.
Jumlah daun (Tabel 2) dan tinggi planlet (Gambar 1) pada kedua kultivar dipengaruhi oleh termoterapi dan elektroterapi. Peningkatan suhu menyebabkan penurunan jumlah daun dan tinggi planlet. Pada kultivar Lumbu Hijau elektroterapi memberikan pengaruh nyata terhadap jumlah daun dan tinggi planlet, sedangkan interaksi antara kedua faktor tidak berpengaruh nyata terhadap kedua parameter tersebut. Kondisi yang berbeda terjadi pada kultivar Sangga Sembalun. Elektroterapi tidak berpengaruh nyata terhadap jumlah daun dan tinggi planlet, tetapi interaksi kedua perlakuan memberikan pengaruh nyata terhadap jumlah daun dan tinggi planlet.
J Fitopatol Indones Nasution et al.
202
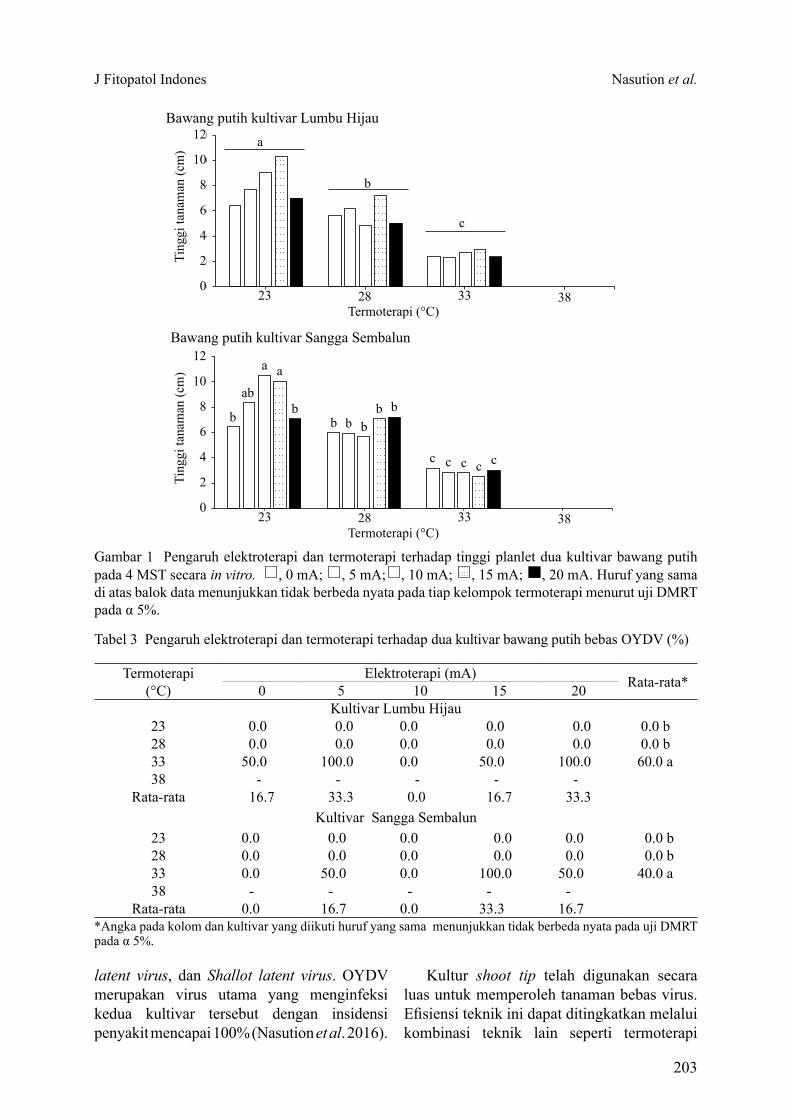
Persentase Tanaman Bebas OYDVDeteksi virus dilakukan secara komposit,
yaitu 4 botol kultur dalam satu ulangan. Perlakuan suhu memberikan pengaruh nyata terhadap persentase tanaman bebas OYDV (Tabel 3), sedangkan perlakuan elektroterapi dan interaksi kedua perlakuan tidak memberikan pengaruh yang nyata terhadap persentase tanaman bebas OYDV.
Deteksi virus setelah perlakuan termoterapi dan elektroterapi dilakukan menggunakan metode RT-PCR. Tanaman yang diperoleh melalui teknik kultur jaringan memiliki
konsentrasi virus yang sangat rendah (Gambar 2). Oleh karena itu, deteksi virus pada tanaman kultur jaringan membutuhkan metode sensitif seperti RT-PCR.
PEMBAHASAN
Bawang putih kultivar Lumbu Hijau adalah kultivar unggul yang banyak ditemukan di Jawa Timur, sementara kultivar Sangga Sembalun banyak terdapat di Nusa Tenggara Barat. Kedua kultivar ini terinfeksi berbagai jenis virus seperti OYDV, Garlic common
Tabel 2 Pengaruh elektroterapi dan termoterapi terhadap jumlah daun planlet dua kultivar bawang putih pada 4 MST
Termoterapi (°C)
Elektroterapi (mA) Rata-rata*0 5 10 15 20Kultivar Lumbu Hijau
23 2.41 2.75 3.16 3.33 2.67 2.86 a28 2.41 2.67 2.91 3.00 2.33 2.67 a33 1.58 1.50 2.08 1.75 1.67 1.71 b38 - - - - - -
Rata-rata* 2.13 c 2.31 abc 2.72 a 2.69 ab 2.22 bcKultivar Sangga Sembalun**
23 2.58 BCD 2.83 BC 3.67 A 3.67 A 2.67 BCD 3.08 a28 2.92 B 2.83 BC 2.50 BCD 2.83 BC 2.67 BCD 2.75 b33 2.17 BCDE 2.00 DE 1.92 DE 1.75 E 2.08 CDE 1.98 c38 - - - - - -
Rata-rata 2.56 2.55 2.70 2.75 2.47*Angka pada kolom atau baris berdasarkan kultivar yang sama diikuti huruf nonkapital yang sama menunjukkan tidak berbeda nyata menurut uji DMRT pada α 5%.**Angka pada kolom dan baris berdasarkan kultivar yang sama diikuti huruf kapital yang sama menunjukkan respon interaksi elektroterapi dan termoterapi tidak berbeda nyata menurut uji DMRT pada α 5%.
Tabel 1 Pengaruh elektroterapi dan termoterapi terhadap eksplan dua kultivar bawang putih yang hidup (%)
Termoterapi (°C)
Elektroterapi (mA) Rata-rata0 5 10 15 20Kultivar Lumbu Hijau
23 100 100 100 100 100 100 a28 100 100 100 100 100 100 a33 100 100 100 100 100 100 a38 0 0 0 0 0 0 b
Kultivar Sangga Sembalun23 100 100 100 100 100 100 a28 100 100 100 100 100 100 a33 100 100 100 100 100 100 a38 0 0 0 0 0 0 b
Angka pada kolom dan kultivar yang sama diikuti huruf yang sama menunjukkan tidak berbeda nyata menurut uji DMRT pada α 5%.
J Fitopatol Indones Nasution et al.
203
Tabel 3 Pengaruh elektroterapi dan termoterapi terhadap dua kultivar bawang putih bebas OYDV (%)
Termoterapi (°C)
Elektroterapi (mA) Rata-rata*0 5 10 15 20Kultivar Lumbu Hijau
23 0.0 0.0 0.0 0.0 0.0 0.0 b28 0.0 0.0 0.0 0.0 0.0 0.0 b33 50.0 100.0 0.0 50.0 100.0 60.0 a38 - - - - -
Rata-rata 16.7 33.3 0.0 16.7 33.3Kultivar Sangga Sembalun
23 0.0 0.0 0.0 0.0 0.0 0.0 b28 0.0 0.0 0.0 0.0 0.0 0.0 b33 0.0 50.0 0.0 100.0 50.0 40.0 a38 - - - - -
Rata-rata 0.0 16.7 0.0 33.3 16.7*Angka pada kolom dan kultivar yang diikuti huruf yang sama menunjukkan tidak berbeda nyata pada uji DMRT pada α 5%.
latent virus, dan Shallot latent virus. OYDV merupakan virus utama yang menginfeksi kedua kultivar tersebut dengan insidensi penyakit mencapai 100% (Nasution et al. 2016).
Kultur shoot tip telah digunakan secara luas untuk memperoleh tanaman bebas virus. Efisiensi teknik ini dapat ditingkatkan melalui kombinasi teknik lain seperti termoterapi
Gambar 1 Pengaruh elektroterapi dan termoterapi terhadap tinggi planlet dua kultivar bawang putih pada 4 MST secara in vitro. , 0 mA; , 5 mA; , 10 mA; , 15 mA; , 20 mA. Huruf yang sama di atas balok data menunjukkan tidak berbeda nyata pada tiap kelompok termoterapi menurut uji DMRT pada α 5%.
0,0
2,0
4,0
6,0
8,0
10,0
12,0
23 28 33 38
12
10
8
6
4
2
023 28 33 38
Termoterapi (°C)
Ting
gi ta
nam
an (c
m)
a
b
c
Bawang putih kultivar Lumbu Hijau
0,0
2,0
4,0
6,0
8,0
10,0
12,0
23 28 33 3823 28 33 38Termoterapi (°C)
12
10
8
6
4
2
0
Ting
gi ta
nam
an (c
m)
b
ab
a a
bb b b
b b
c c c c c
Bawang putih kultivar Sangga Sembalun

J Fitopatol Indones Nasution et al.
204
dan elektroterapi. Termoterapi memberikan pengaruh nyata terhadap persentase hidup eksplan. Perlakuan suhu terlalu tinggi berpengaruh negatif terhadap pertumbuhan eksplan. Suhu 38 °C dapat menghambat pertumbuhan eksplan. Hasil yang sama dilaporkan oleh Conci et al. (2005) yang melakukan penelitian pada tanaman bawang putih, eksplan meristem yang diberi perlakuan suhu 36 °C selama 30–40 hari dapat tumbuh sekitar 55–57% dibandingkan dengan eksplan yang tidak diberi perlakuan dapat tumbuh sekitar 80-90%. Mahmoud et al. (2009) melaporkan bahwa tingkat regenerasi semakin menurun pada tanaman kentang yang diberi perlakuan suhu 37 °C selama 40 hari secara in vitro.
Jumlah daun dan tinggi planlet dipengaruhi oleh termoterapi dan elektroterapi pada kedua kultivar. Suhu rendah memberikan pengaruh baik terhadap perkembangan planlet, sementara suhu tinggi dapat menghambat perkembangan planlet. Eksplan yang diberi perlakuan elektroterapi 10 mA pada suhu 23 °Cmemiliki jumlah daun dan tinggi tanaman lebih baik dibandingkan dengan eksplan yang tidak diberikan perlakuan elektroterapi. Hal ini sesuai dengan penelitian Soliman et al. (2012) bahwa elektroterapi pada 10 mA selama 10 menit dapat meningkatkan tingkat regenerasi planlet bawang putih. Tanaman kentang yang diberi perlakuan elektroterapi 10 mA selama 10 menit dapat meningkatkan regenerasi tanaman in vitro (Mahmoud et al. 2009). Aliran listrik dilaporkan dapat memacu diferensiasi tanaman in vitro (Goldsworthy 1987). Regenerasi eksplan in vitro oleh elektroterapi bergantung pada besarnya aliran listrik dan kultivar tanaman yang digunakan
(Badarua dan Chiru 2014). Menurut Dovas et al. (2001), RT-PCR merupakan metode sensitif dalam deteksi OYDV. Sensitivitas RT-PCR 102–104 kali lebih tinggi dibandingkan dengan metode enzyme-linked immunosorbent assay (ELISA).
Percobaan eliminasi menggunakan metode elektroterapi pada berbagai tingkat arus listrik memperlihatkan bahwa perlakuan elektroterapi tidak berpengaruh nyata terhadap persentase tanaman bebas virus. Hal ini mungkin disebabkan konsentrasi virus yang tinggi dan ukuran tunas adventif terlalu besar. Lassois et al. (2013) menyatakan bahwa ketersediaan tanaman bebas virus selain bergantung pada teknik yang dikembangkan juga perlu memperhatikan karakteristik virus, tipe jaringan yang diberikan perlakuan, dan jenis tanaman yang digunakan. Dewi dan Slack (1994) menambahkan bahwa efisiensi protokol untuk mengeliminasi virus akan menurun jika eksplan yang digunakan mengandung konsentrasi virus yang tinggi atau telah terinfeksi oleh berbagai jenis virus.
Menurut Conci et al. (2005), efisiensi untuk mendapatkan tanaman bebas virus bergantung dari jenis virus target dan kultivar bawang putih yang digunakan. Ghaemizadeh et al. (2014) berhasil memperoleh tanaman bebas OYDV dengan efisiensi 77% melalui perlakuan suhu 36 oC dikombinasikan dengan kultur stem-disc. Suhu yang tinggi dapat menekan multiplikasi virus, yaitu dengan meningkatkan degradasi RNA virus (Wang dan Valkonen 2008). Perlakuan suhu tinggi dapat mengakibatkan pecahnya ikatan hidrogen dan disulfida protein penyelubung virus, diikuti dengan ikatan kovalen fosfodiester asam nukleat yang dapat mengakibatkan
Gambar 2 Hasil amplifikasi Onion yellow dwarf virus dari tanaman bawang putih setelah perlakuan elektroterapi dan termoterapi. M, Penanda DNA 1 kb; 1, Kontrol positif; 2, Kontrol negatif; 3–7, Suhu 23 °C dan 0, 5, 10, 15, 20 mA; 8–12, Suhu 28 °C: 0, 5, 10, 15, 20 mA dan 13–17, Suhu 33 °C dan 0, 5, 10, 15, 20 mA.
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
601 bp

J Fitopatol Indones Nasution et al.
205
kemunduran infektivitas dan penghambatan replikasi virus (Panattoni et al. 2013).
Perlakuan terbaik untuk mendapatkan tanaman bebas OYDV ialah perlakuan suhu 33 °C karena mampu menghasilkan tanaman bebas OYDV hingga 60% pada kultivar Lumbu Hijau dan 40% pada kultivar Sangga Sembalun.
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada ACIAR atas dukungannya melalui kegiatan penelitian: Increasing Productivity of Allium and Solanaceous Vegetable Crops in Indonesia and Sub-Tropical Australia (Hort/2009/056).
DAFTAR PUSTAKA
Badarua CL, Chiru N. 2014. Effect of some therapies on potato plantlets infected with Potato Virus X (PVX). In: Proceeding of Bioatlas 2014 Conference; 2014 May 15–17; Brasov (RO): BIOATLAS Conference. Hlm 11–17.
Conci VC, Perotto MC, Cafrne E, Lunello P. 2005. Program for intensive production of virus-free garlic plants. In: Proceeding IVth IS on Edible Alliaceae; 2004 April 21-26; Beijing (CN): Acta Hort. DOI: https://doi.org/10.17660/ActaHortic.2005.688.25.
Dewi IS, Slack A. 1994. Therapy cycling to eliminate high-titered, multiple virus infection in vitro potato plantlets. Bul Agron. 22(2):35–43.
Diekmann M. 1997. FAO/IPGRI Technical Guidelines for the Safe Movement of Germplasm. No. 18. Allium spp. Food and Agriculture Organization of the United Nations. Rome (IT): International Plant Genetik Resources Institute.
Dovas CI, Hatziloukas E, Salomon R, Barg E, Shiboleth Y, Katis NI. 2001. Comparison of methods for virus detection in Allium spp. J Phytopathol. 149(11–12):731–737. DOI: https://doi.org/10.1046/j.1439-0434.2001.00705.x.
Doyle JJ, Doyle JJ. 1987. A rapid DNA isolation of procedure for small quantities of fresh leaf tissue. Phytochem Bull. 19:11–19.
Elnagar S, El-Sheikh MAK, Wahab ASA. 2009. Effect of natural infection with Onion yellow dwarf virus (OYDV) on yield of onion and garlic crops in Egypt. In: 4th Conference on Recent Technologies in Agriculture; 2009 November 3-5; Cairo (EG): Faculty of Agriculture Cairo University. hlm 34–39.
Ghaemizadeh F, Dashti F, Khodakaramian G, Sarikhani H. 2014. Combination of stem-disc dome culture and thermotherapy to eliminate Allexiviruses and Onion yellow dwarf virus from garlic (Allium sativum cv. Hamedan). Arc Phytol Plant Protection. 4(47):499–507. DOI: https://doi.org/10.1080/03235408.2013.813123.
Goldsworthy A. 1987. Electrical stimulation of tissue culture growth and morphogenesis. Agricell Repory. 8:14–14.
Lassois L, Lepoivre P, Swennen R, Houwe IVD, Panis P. 2013. Thermotherapy, chemotherapy, and meristem culture in banana. In: Lambardi M, Ozudogru EA, Jain SM, editor. Protocols for Micropropagation of Selected Economically-Important Horticultural Plants. Methods in Molecular Biology. New York (US): Humana Press. Hlm 419-433.
Lot H, Chovelon V, Souche S, Delecolle B. 1998. Effects of Onion yellow dwarf and Leek yellow stripe viruses on symptomatology and yield loss of three french garlic cultivars. Plant Dis. 82(12):1381–1385. DOI: https://doi.org/10.1094/PDIS.1998.82.12.1381.
Mahmoud SYM, Hosseny MH, Abdel-Ghaffar MH. 2009. Evaluation of some therapies to eliminate Potato Y Potyvirus from potato plants. Inter J Virol. 5(2):64–76. DOI: https://doi.org/10.3923/ijv.2009.64.76.
Moreno PL, Jaramill SLI, Celedon MB, Malagon RR, Palenius NHG. 2014. Effect of natural virus infection on quality and yield of garlic elite lines. J Experimen Biol Agric Sci 2(2S):243–250.
Nasution SS, Hidayat SH, Dinarti D. 2016. Deteksi dan identifikasi virus pada bawang putih di Indonesia. Di dalam: Prosiding Pengendalian Penyakit pada Tanaman

J Fitopatol Indones Nasution et al.
206
Ramah Lingkungan II; 2016 Agustus 27; Yogyakarta (ID): Perhimpunan Fitopatologi Indonesia Komda Joglosemar. Hlm 315
Panattoni A, Luvisi A, Triolo E. 2013. Elimination of viruses in plants: twenty years progress. Spanish J Agric Res. 11(1):173–188. DOI: https://doi.org/10.5424/sjar/2013111-3201.
Soliman AM, Mahmoud SYM, Dawood RA. 2012. Molecular characterization of Onion yellow dwarf virus (garlic isolate) with production of virus-free plantlets.
Int J Virol. 8(1):61–70. DOI: https://doi.org/10.3923/ijv.2012.61.70.
Torres AC, Fajardo TV, Dusi AN, Resende RO, Buso JA. 2000. Shoot tip culture and thermotherapy for recovering virus-free plants of garlic. Hort Bras. 18(3):192–195. DOI: https://doi.org/10.1590/S0102-05362000000300010.
Wang Q, Valkonen JPT. 2008. Cryoterapy of shoot tips: novel pathogen eradication method. Trends Plant Science. 14(3):119–122. DOI: https://doi.org/10.1016/j.tplants.2008.11.010.

ISSN: 0215-7950
207
Volume 13, Nomor 6, November 2017Halaman 207–215
DOI: 10.14692/jfi.13.6.207
*Alamat penulis korespondensi: Departemen Proteksi Tanaman, Fakultas Pertanian Institut Pertanian Bogor, Jalan Kamper, Kampus Darmaga IPB, Bogor 16680 Tel: 0251-8629364, Faks: 0251-8629362, surel: [email protected]
Asam Salisilat sebagai Penginduksi Ketahanan Tanaman Padi terhadap Penyakit Hawar Daun Bakteri
Salicylic Acid as Inducers for Rice Resistance Against Bacterial Leaf Blight Disease
Christoffol Leiwakabessy, Meity Suradji Sinaga, Kikin Hamzah Mutaqin, Trikoesoemaningtyas, Giyanto*
Institut Pertanian Bogor, Bogor 1660
ABSTRAK
Penggunaan asam salisilat sebagai penginduksi ketahanan beberapa jenis tanaman saat ini mulai mendapat banyak perhatian. Asam salisilat diyakini berperan penting dalam menginduksi ketahanan tanaman terhadap patogen. Penelitian ini bertujuan menentukan peran dan keefektifan asam salisilat sebagai penginduksi ketahanan tanaman padi (Oryza sativa) terhadap penyakit hawar daun bakteri yang disebabkan oleh Xanthomonas oryzae pv. oryzae. Percobaan disusun dalam rancangan acak lengkap faktorial yang terdiri atas 3 faktor (varietas, asam salisilat, dan patotipe), tiap perlakuan diulang 3 kali. Hasil penelitian menunjukkan bahwa asam salisilat mampu memperpanjang periode laten X. oryzae pv. oryzae patotipe IV dan VIII pada padi varietas Conde dan Ciherang, dapat menginduksi ketahanan varietas padi, dan mampu menekan laju infeksi kedua patotipe bakteri tersebut. Asam salisilat juga mampu menginduksi ketahanan padi varietas Conde dan Ciherang dalam menekan area di bawah kurva perkembangan penyakit (AUDPC) X. oryzae pv. oryzae patotipe IV dan VIII.
Kata kunci: AUDPC, Oryaza sativa, patotipe, Xanthomonas oryzae pv. oryzae
ABSTRACT
The use of salicylic acid as resistance inducers agents in several plants species was well known. Salicylic acid has been believed to play an important role in inducing resistance against some pathogen. This research was aimed to study the role and effectiveness of salicylic acid as inducers for the resistance of some rice variety against bacterial leaf blight caused by Xanthomonas oryzae pv. oryzae. The experiment was arranged in completely randomized factorial design with three factors i.e. rice variety, salicylic acid, and pathotypes, each treatment was replicated three times. The results showed that salicylic acid could prolong the latent period of X. oryzae pv. oryzae pathotypes IV and VIII in paddy rice varieties Conde and Ciherang, induce the resistance of rice varieties as well as reducing the rate of infection of the two mentioned pathotypes. Salicylic acid was also able to induce the resistance of Conde and Ciherang varieties in order to suppress area under disease progress curve (AUDPC) of X. oryzae pv. oryzae pathotypes IV and VIII.
Key words: AUDPC, Oryaza sativa, pathotypes, Xanthomonas oryzae pv. oryzae

J Fitopatol Indones Leiwakabessy et al.
208
PENDAHULUAN
Penyakit hawar daun bakteri (HDB) merupakan salah satu penyakit penting pada pertanaman padi. Penyakit HDB yang disebabkan oleh bakteri patogen X. oryzae pv. oryzae menjadi salah satu penyebab kehilangan hasil di negara-negara penghasil padi khususnya di Asia (Ezuka dan Kaku 2000). Berbagai upaya pengendalian penyakit ini telah dilakukan baik secara biotik (Putra dan Giyanto 2014; Krishanti et al. 2015) maupun secara abiotik (Mustafa et al. 2013; Kim et al. 2015), tetapi hingga saat ini penyakit HDB masih menjadi kendala utama produksi padi di daerah tropis maupun subtropis. Berdasarkan hasil survei Sudir dan Yuliani (2016) pada 10 provinsi di Indonesia ditemukan 3 patotipe yang dominan, yaitu patotipe III (30%), IV (36%), dan VIII (34%).
Asam salisilat adalah salah satu agens penginduksi ketahanan yang dilaporkan dapat digunakan untuk pengendalian patogen tanaman. Menurut Hayat et al. (2010), asam salisilat merupakan senyawa fenol sederhana yang berperan penting dalam mengatur proses fisiologi dan respons imunisasi tanaman. Pemanfaatan asam salisilat sebagai sinyal transduksi dalam jaringan pertahanan tanaman telah diamati dan dikarakterisasi pada sejumlah gen yang berfungsi dalam biosintesis asam salisilat. Rangkaian dari proses ini meliputi konjugasi, akumulasi, dan crosstalk hormon tanaman seperti asam jasmonat, etilen, asam absisi, auksin, giberrelin, sitokinin, dan brassinosteroid (An dan Mou 2011).
Menurut Mohan-Babu et al. (2003), induksi ketahanan tanaman padi terhadap penyakit HDB dengan konsentrasi asam salisilat 1000 μmol menyebabkan peningkatan kandungan senyawa fenol sehingga dapat menekan perkembangan penyakit HDB. Kombinasi perlakuan asam salisilat dan sodium nitroprusside dapat meningkatkan pigmen fotosintesis dan aktivitas beberapa enzim pertahanan tanaman pada tanaman kacang tanah yang terinfeksi Peanut mottle virus (Kobeasy et al. 2011). Mandal et al. (2009) melaporkan bahwa aplikasi asam alisilat
200 µM pada daun dan akar dapat menekan perkembangan penyakit layu Fusarium (Fusarium oxysporum f.sp. lycopersicum) pada tanaman tomat dan terjadi peningkatan enzim fenilalaninamonia liase dan peroksidase. Asam salisilat secara eksogen dapat memengaruhi pertumbuhan tanaman, fotosintesis, hubungan air dan tanaman, dan aktivitas beberapa jenis enzim yang berperan terhadap cekaman biotik dan abiotik (Zamaninejad et al. 2013). Berdasarkan hal tersebut maka penelitian ini bertujuan memperoleh konsentrasi asam salisilat yang optimum sebagai penginduksi ketahanan padi varietas IR64, Ciherang, dan Conde terhadap X. oryzae pv. oryzae patotipe IV dan VIII.
BAHAN DAN METODE
Persiapan Tanam dan Xanthomonas oryzae pv. oryzae
Tiga varietas benih padi (IR64, Ciherang, dan Conde) koleksi BB Padi Sukamandi didesinfeksi dengan natrium hipoklorit selama 2 menit dan dibilas 3 kali dengan air steril. Benih padi dikeringanginkan dan disterilisasi dengan metode hot water treatment pada suhu 55 °C selama 20 menit. Selanjutnya benih padi direndam dalam air steril selama 24 jam dan disemai pada wadah plastik berisi medium campuran tanah dan pupuk kandang (1:1, b/b). Setelah berumur 14 hari, bibit padi dipindahkan ke pot plastik berdiameter 30 cm × 40 cm.
Inokulum X. oryzae pv. oryzae patotipe IV dan VIII (koleksi BB Padi Sukamandi) diperbanyak pada medium Wakimoto (OEPP 2007) dan diamati pertumbuhannya selama 72 jam, selanjutnya disimpan pada medium agar-agar miring sebelum digunakan untuk pengujian patogenisitas. Patotipe IV dan VIII digunakan dalam penelitian ini karena lebih dominan dibandingkan dengan patotipe III. Konsentrasi X. oryzae pv. oryzae yang digunakan untuk pengujian patogenisitas ialah 107 cfu mL-1. Uji patogenisitas bakteri X. oryzae pv. oryzae dilakukan pada padi varietas IR64 berumur 30 hari untuk memastikan bahwa galur ini benar-benar X. oryzae pv oryzae patotipe IV dan VIII.

J Fitopatol Indones Leiwakabessy et al.
209
Rancangan PenelitianPenelitian disusun dalam rancangan
acak lengkap faktorial dengan tiga faktor, yaitu asam salisilat (A, A0=0, A1=5, A2=10, A3=15 mM); patotipe X. oryzae pv. oryzae (B, B0=kontrol, B1=Patotipe Xoo IV, B2= Patotipe Xoo VIII; dan varietas padi (C, C1=IR64, C2=Ciherang, C3=Conde). Setiap perlakuan diulang tiga kali. Setiap satuan percobaan diambil 4 sampel tanaman untuk diamati perkembangan penyakit HDB.
Aplikasi Asam Salisilat dan Inokulasi Xanthomonas oryzae pv. oryzae
Penentuan konsentrasi asam salisilat (HOC9H8COOH, Merck KGaA cas No. 66-72-7) mengacu pada hasil uji pendahuluan dengan berbagai konsentrasi asam salisilat (SA) (0, 5, 10, 15, dan 20 mM). Asam salisilat disiapkan dengan melarutkan dalam alkohol 70% dan dibuat larutan stok sebanyak 200 mLuntuk setiap perlakuan uji. Asam salisilat disemprotkan pada tanaman padi berumur 40 hari setelah tanam (HST) dengan konsentrasi 5, 10, 15 mM sebanyak 20 mL per tanaman.
Inokulasi X. oryzae pv. oryzae (patotipe IV dan VIII) dilakukan pada tanaman padi berumur 43 HST dengan konsentrasi 107 cfu mL-1 menggunakan metode clipping, yaitu memotong ujung daun, tempat ujung ini dicelupkan ke dalam suspensi X. oryzae pv. oryzae (Ou 1985). Tanaman yang telah diinokulasi disungkup dengan plastik transparan selama 3–4 hari supaya kelembapan tetap tersedia bagi patogen.
PengamatanPengamatan penyakit dilakukan
terhadap periode laten, laju infeksi (Van der Planck 1963), dan area di bawah kurva perkembangan penyakit (AUDPC) (Madden et al. 2007). Pengamatan ini dilakukan dengan menggunakan kriteria skor kerusakan untuk menghitung keparahan penyakit HDB, yaitu 0, 0 < x ≤ 1% daun terserang HDB; 1, 1 < x ≤ 5% daun terserang HDB; 2, 5 < x ≤ 15% daun terserang HDB; 3, 15 < x ≤ 25% daun terserang HDB; 4, 25 < x ≤ 50% daun terserang HDB; dan 5, > 50 % daun terserang HDB (modifikasi
SES IRRI, 2014). Pengamatan penyakit dilakukan dengan interval waktu 5 hari.
Analisis Aktivitas Enzim-Enzim yang Terlibat dalam Pertahanan Tanaman
Analisis aktivitas enzim-enzim pertahanant a n a m a n m e n c a k u p p e r o k s i d a s e (Hammerschmidt et al. 1982), polifenol-oksidase (Malick dan Singh 1980), dan β-1-3 glukanase (Pan et al. 1991) dilakukan sebelum dan setelah inokulasi X. oryzae pv. oryzae. Data hasil pengamatan dianalisis menggunakan perangkat lunak Microsoft Excel 2013 dan SAS versi 9.2. Apabila terdapat perbedaan nyata dilanjutkan dengan uji Duncan pada α 5% dan 1%.
HASIL
Patogenisitas X. oryzae pv. oryzaeHasil uji patogenisitas X. oryzae pv.
oryzae patotipe IV dan VIII terhadap varietas IR64 menunjukkan rata-rata gejala penyakit HDB mulai muncul pada hari ke-5 sampai ke-8 disertai perubahan warna daun dari hijau menjadi keabu-abuan. Patotipe IV lebih dominan dibandingkan dengan patotipe VIII pada padi varietas IR64. Hal ini menunjukkan bahwa dengan periode laten yang lebih singkat patotipe IV lebih cepat berkembang pada jaringan tanaman daripada patotipe VIII.
Penyakit HDB pada Varietas Padi yang Diinokulasi dengan Dua Patotipe Xanthomonas oryzae pv. oryzae dan Pemberian Asam Salisilat
Analisis ragam menunjukkan bahwa asam salisilat mampu menekan perkembangan penyakit HDB dan memperpanjang periode laten patogen ini pada varietas padi. Hal ini diketahui dari nilai AUDPC yang rendah. Asam salisilat mampu meningkatkan induksi ketahanan varietas padi untuk menekan laju infeksi penyakit HDB dan memperpanjang periode laten varietas padi, tetapi tidak dapat menekan AUDPC. Asam salisilat mampu memengaruhi induksi ketahanan varietas padi dalam menekan laju infeksi dan AUDPC patotipe X. oryzae pv. oryzae, tetapi tidak
J Fitopatol Indones Leiwakabessy et al.
210
berpengaruh nyata terhadap periode laten. Pengaruh asam salisilat, patotipe, dan varietas tidak berbeda nyata terhadap AUDPC (Tabel 1).
Periode laten pada padi varietas Conde tanpa asam salisilat lebih panjang dan laju infeksinya lebih rendah, sedangkan pemberian asam salisilat dengan konsentrasi 10 dan 15 mM tanpa inokulasi patogen memiliki laju infeksi yang tinggi, dan periode laten panjang. Periode laten yang lebih panjang dan laju infeksi yang lebih rendah ditunjukkan oleh pemberian asam salisilat 10 mM pada ketiga varietas dibandingkan dengan tanpa pemberian asam salisilat dengan konsentrasi 5 dan 15 mM setelah diinokulasi dengan X. oryzae pv. oryzae patotipe IV dan VIII (Tabel 2).
Perlakuan asam salisilat 10 mM pada padi varietas IR64 mampu menekan AUDPC patotipe IV dan VIII, tetapi tidak berbeda dengan perlakuan IR64-SA0. Nilai AUDPC pada perlakuan IR64-SA10 diperoleh lebih rendah dibandingkan dengan perlakuan IR64-SA5 dan IR64-SA15, tetapi tidak berbeda
dengan IR64-SA0 dan IR64-SA15 pada patotipe yang sama. Perlakuan IR64-SA10 terhadap patotipe IV diperoleh AUDPC lebih rendah dibandingkan dengan perlakuan Conde-SA5 dan Conde-SA15. Perlakuan yang sama tidak berbeda nyata dengan Ciherang-SA0, Ciherang-SA5, Ciherang-SA10, dan Ciherang-SA15, serta Conde-SA0 dan Conde-SA10 (Tabel 3).
Nilai AUDPC perlakuan SA10 pada padi varietas Ciherang terhadap patotipe IV tidak berbeda nyata dengan perlakuan Ciherang-SA0, Ciherang-SA5, dan Ciherang-SA15, tetapi berbeda nyata dengan Ciherang-SA15 terhadap patotipe VIII. Perlakuan yang sama juga tidak berbeda nyata terhadap perlakuan IR64-SA10 dan IR64-SA5 dalam menekan patotipe VIII serta IR64-SA0 terhadap patotipe IV (Tabel 3).
Perlakuan tanpa asam salisilat (SA0) pada padi varietas Conde menunjukkan nilai AUDPC berbeda nyata dibandingkan dengan perlakuan Conde-SA15, Conde-SA10, dan
Pengamatan SA Pato Var SA x Pato SA x Var Pato x Var SA x Pato x VarPeriode laten 0.00** 0.00** 0.00** 0.00** 0.00** 0.19tn 0.42tn
Laju infeksi 0.00** 0.02* 0.00** 0.08tn 0.00** 0.00** 0.11tn
AUDPC 0.00** 0.00** 0.11tn 0.00** 0.86tn 0.02* 0.39tn
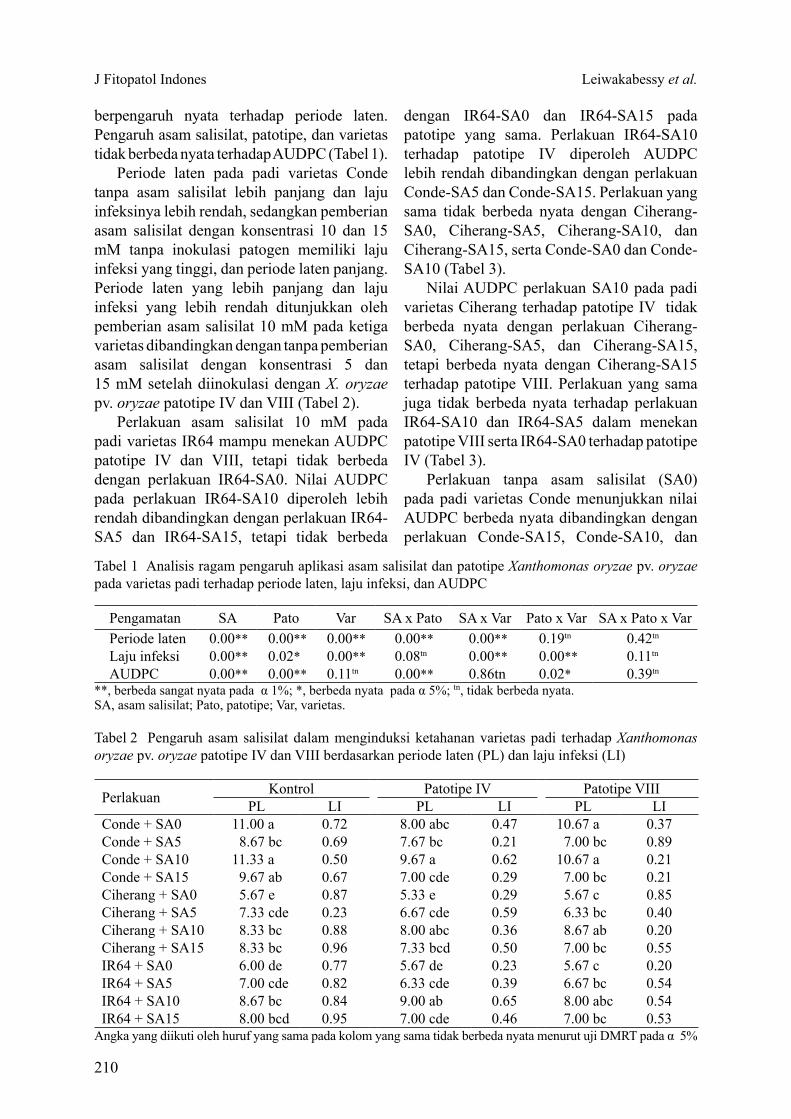
Tabel 1 Analisis ragam pengaruh aplikasi asam salisilat dan patotipe Xanthomonas oryzae pv. oryzae pada varietas padi terhadap periode laten, laju infeksi, dan AUDPC
**, berbeda sangat nyata pada α 1%; *, berbeda nyata pada α 5%; tn, tidak berbeda nyata.SA, asam salisilat; Pato, patotipe; Var, varietas.
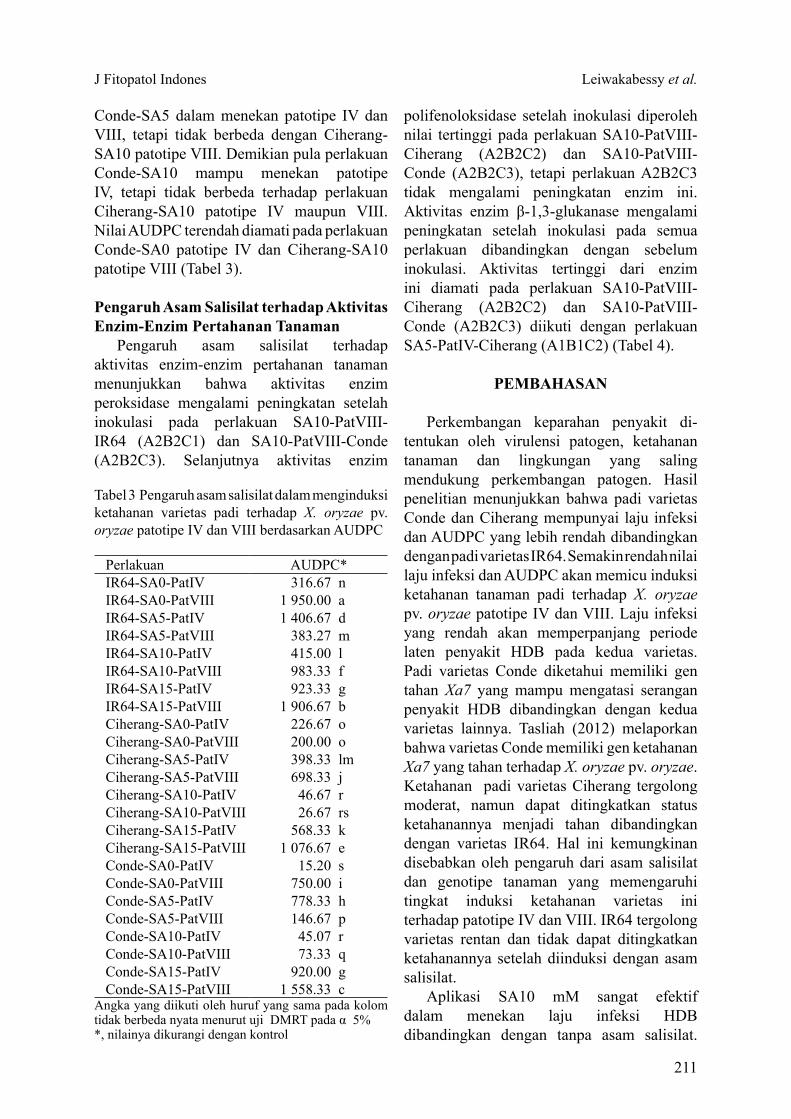
Tabel 2 Pengaruh asam salisilat dalam menginduksi ketahanan varietas padi terhadap Xanthomonas oryzae pv. oryzae patotipe IV dan VIII berdasarkan periode laten (PL) dan laju infeksi (LI)
Angka yang diikuti oleh huruf yang sama pada kolom yang sama tidak berbeda nyata menurut uji DMRT pada α 5%
Perlakuan Kontrol Patotipe IV Patotipe VIIIPL LI PL LI PL LI
Conde + SA0 11.00 a 0.72 8.00 abc 0.47 10.67 a 0.37Conde + SA5 8.67 bc 0.69 7.67 bc 0.21 7.00 bc 0.89Conde + SA10 11.33 a 0.50 9.67 a 0.62 10.67 a 0.21Conde + SA15 9.67 ab 0.67 7.00 cde 0.29 7.00 bc 0.21Ciherang + SA0 5.67 e 0.87 5.33 e 0.29 5.67 c 0.85Ciherang + SA5 7.33 cde 0.23 6.67 cde 0.59 6.33 bc 0.40Ciherang + SA10 8.33 bc 0.88 8.00 abc 0.36 8.67 ab 0.20Ciherang + SA15 8.33 bc 0.96 7.33 bcd 0.50 7.00 bc 0.55IR64 + SA0 6.00 de 0.77 5.67 de 0.23 5.67 c 0.20IR64 + SA5 7.00 cde 0.82 6.33 cde 0.39 6.67 bc 0.54IR64 + SA10 8.67 bc 0.84 9.00 ab 0.65 8.00 abc 0.54IR64 + SA15 8.00 bcd 0.95 7.00 cde 0.46 7.00 bc 0.53
J Fitopatol Indones Leiwakabessy et al.
211
Conde-SA5 dalam menekan patotipe IV dan VIII, tetapi tidak berbeda dengan Ciherang-SA10 patotipe VIII. Demikian pula perlakuan Conde-SA10 mampu menekan patotipe IV, tetapi tidak berbeda terhadap perlakuan Ciherang-SA10 patotipe IV maupun VIII. Nilai AUDPC terendah diamati pada perlakuan Conde-SA0 patotipe IV dan Ciherang-SA10 patotipe VIII (Tabel 3).
Pengaruh Asam Salisilat terhadap Aktivitas Enzim-Enzim Pertahanan Tanaman
Pengaruh asam salisilat terhadap aktivitas enzim-enzim pertahanan tanaman menunjukkan bahwa aktivitas enzim peroksidase mengalami peningkatan setelah inokulasi pada perlakuan SA10-PatVIII-IR64 (A2B2C1) dan SA10-PatVIII-Conde (A2B2C3). Selanjutnya aktivitas enzim
polifenoloksidase setelah inokulasi diperoleh nilai tertinggi pada perlakuan SA10-PatVIII-Ciherang (A2B2C2) dan SA10-PatVIII-Conde (A2B2C3), tetapi perlakuan A2B2C3 tidak mengalami peningkatan enzim ini. Aktivitas enzim β-1,3-glukanase mengalami peningkatan setelah inokulasi pada semua perlakuan dibandingkan dengan sebelum inokulasi. Aktivitas tertinggi dari enzim ini diamati pada perlakuan SA10-PatVIII-Ciherang (A2B2C2) dan SA10-PatVIII-Conde (A2B2C3) diikuti dengan perlakuan SA5-PatIV-Ciherang (A1B1C2) (Tabel 4).
PEMBAHASAN
Perkembangan keparahan penyakit di-tentukan oleh virulensi patogen, ketahanan tanaman dan lingkungan yang saling mendukung perkembangan patogen. Hasil penelitian menunjukkan bahwa padi varietas Conde dan Ciherang mempunyai laju infeksi dan AUDPC yang lebih rendah dibandingkan dengan padi varietas IR64. Semakin rendah nilai laju infeksi dan AUDPC akan memicu induksi ketahanan tanaman padi terhadap X. oryzae pv. oryzae patotipe IV dan VIII. Laju infeksi yang rendah akan memperpanjang periode laten penyakit HDB pada kedua varietas. Padi varietas Conde diketahui memiliki gen tahan Xa7 yang mampu mengatasi serangan penyakit HDB dibandingkan dengan kedua varietas lainnya. Tasliah (2012) melaporkan bahwa varietas Conde memiliki gen ketahanan Xa7 yang tahan terhadap X. oryzae pv. oryzae. Ketahanan padi varietas Ciherang tergolong moderat, namun dapat ditingkatkan status ketahanannya menjadi tahan dibandingkan dengan varietas IR64. Hal ini kemungkinan disebabkan oleh pengaruh dari asam salisilat dan genotipe tanaman yang memengaruhi tingkat induksi ketahanan varietas ini terhadap patotipe IV dan VIII. IR64 tergolong varietas rentan dan tidak dapat ditingkatkan ketahanannya setelah diinduksi dengan asam salisilat.
Aplikasi SA10 mM sangat efektif dalam menekan laju infeksi HDB dibandingkan dengan tanpa asam salisilat.
Perlakuan AUDPC*IR64-SA0-PatIV 316.67 n IR64-SA0-PatVIII 1 950.00 aIR64-SA5-PatIV 1 406.67 dIR64-SA5-PatVIII 383.27 mIR64-SA10-PatIV 415.00 lIR64-SA10-PatVIII 983.33 f IR64-SA15-PatIV 923.33 gIR64-SA15-PatVIII 1 906.67 bCiherang-SA0-PatIV 226.67 oCiherang-SA0-PatVIII 200.00 oCiherang-SA5-PatIV 398.33 lmCiherang-SA5-PatVIII 698.33 jCiherang-SA10-PatIV 46.67 rCiherang-SA10-PatVIII 26.67 rsCiherang-SA15-PatIV 568.33 kCiherang-SA15-PatVIII 1 076.67 eConde-SA0-PatIV 15.20 sConde-SA0-PatVIII 750.00 iConde-SA5-PatIV 778.33 hConde-SA5-PatVIII 146.67 pConde-SA10-PatIV 45.07 rConde-SA10-PatVIII 73.33 qConde-SA15-PatIV 920.00 gConde-SA15-PatVIII 1 558.33 c
Tabel 3 Pengaruh asam salisilat dalam menginduksi ketahanan varietas padi terhadap X. oryzae pv. oryzae patotipe IV dan VIII berdasarkan AUDPC
Angka yang diikuti oleh huruf yang sama pada kolom tidak berbeda nyata menurut uji DMRT pada α 5% *, nilainya dikurangi dengan kontrol

J Fitopatol Indones Leiwakabessy et al.
212
Hal ini menunjukkan bahwa secara endogen kandungan asam salisilat pada tanaman padi cukup tinggi daripada tanaman lainnya sehingga jika konsentrasi ditingkatkan maka akan terjadi peningkatan reaksi kerentanan tanaman ini terhadap penyakit HDB. Secara endogen kandungan asam salisilat pada tanaman padi 10 kali lebih tinggi daripada tanaman lainnya, yaitu sebanyak 30-40 μg per tanaman (Silverman et al. 1995; Vicente dan Plasencia 2011). Ibrahim (2012) juga menyatakan bahwa aplikasi asam salisilat 10 mM pada tanaman wortel dapat menekan Xanthomonas vesicatoria.
Setiap patogen, demikian juga dengan X. oryzae pv. oryzae membutuhkan energi yang cukup dalam proses infeksi dan kolonisasi. Pada saat awal proses infeksi, energi yang tersedia tidak cukup bagi patogen tersebut sehingga membutuhkan waktu yang panjang untuk melakukan proses infeksi ke tanaman. Keragaman respons antarpatotipe kemungkinan disebabkan oleh pengaruh dari genotipe tanaman dan patogen X. oryzae pv. oryzae dalam memberikan respons terhadap penyakit HDB. Dalam patosistem penyakit HDB interaksi antara tanaman inang dan patogen bersifat dinamis dan memiliki kesamaan dengan teori gen for gen ketika
gejala penyakit terjadi akibat adanya interaksi inkompatibel antara genavirulen dan gen resisten dari patogen dan tanaman (Nayak 2008).
Saat X. oryzae pv. oryzae menginfeksi tanaman diduga akan mengubah keseimbangan hormon asam salisilat sehingga memicu pengaktifan senyawa-senyawa pertahanan tanaman. Saat patogen menyerang tanaman terjadi manipulasi terhadap hormon tanaman yang dapat berdampak positif maupun negatif baik melalui peningkatan ketahanan tanaman maupun pengaruhnya terhadap pertumbuhan tanaman seperti kekerdilan, jantan mandul, dan penurunan produksi (Holeski et al. 2012). Selanjutnya Ankenbauer dan Cox (1988) menyatakan bahwa beberapa genus Xanthomonas dapat menyintesis asam salisilat selain berperan dalam ketahanan tanaman terhadap infeksi patogen, juga dapat memediasi tanaman terhadap cekaman abiotik.
Hasil penelitian menunjukkan terdapat peningkatan aktivitas enzim pertahanan tanaman pada perlakuan asam salisilat. Dari ketiga enzim yang dianalisis, enzim peroksidase merupakan enzim yang kurang terlihat aktivitasnya baik sebelum dan setelah inokulasi dibandingkan kedua enzim yang lainnya. Pemberian asam salisilat 10 mM
PerlakuanSebelum inokulasi Sesudah inokulasi
Peroksidase Polifenoksidase β-1.3-glukanase Peroksidase Polifenoksidase β-1.3-
glukanaseA0B1C1 0.000 0.000 0.002 0.000 0.003 0.337A1B1C1 0.000 0.013 0.002 0.000 0.007 0.323A2B2C1 0.000 0.000 0.003 0.020 0.023 0.328A3B1C1 0.000 0.003 0.003 0.000 0.013 0.326A0B1C2 0.020 0.023 0.003 0.000 0.023 0.335A1B1C2 0.016 0.007 0.003 0.000 0.013 0.417A2B2C2 0.000 0.027 0.003 0.000 0.027 0.470A3B1C2 0.000 0.007 0.003 0.000 0.013 0.362A0B1C3 0.000 0.027 0.004 0.000 0.017 0.023A1B1C3 0.000 0.007 0.003 0.000 0.023 0.055A2B2C3 0.000 0.002 0.003 0.018 0.027 0.467A3B2C3 0.000 0.000 0.003 0.000 0.000 0.330
Tabel 4 Aktivitas enzim pertahanan tanaman (unit µg-1) pada tiga varietas padi yang terinduksi ketahanannya oleh asam salisilat sebelum dan setelah inokulasi X. oryzae pv. oryzae
A (asam salisilat: A0=0, A1=5, A2=10, A3=15 mM), B (patotipe X. oryzae, pv. oryzae: B0=kontrol, B1=patotipe IV, B2=patotipe VIII), C1 (varietas: C1=IR64, C2=Ciherang, C3=Conde).

J Fitopatol Indones Leiwakabessy et al.
213
setelah inokulasi mampu meningkatkan aktivitas enzim peroksidase pada varietas Conde. Kedua jenis enzim yang lain seperti polifenoloksidase dan β-1,3-glukanase aktivitasnya lebih terlihat. Resistensi varietas kentang berkorelasi dengan aktivitas enzim polifenoloksidase, PAL, asam klorogenat, dan fenol yang berperan penting dalam menghambat infeksi penyakit busuk lunak pada kentang (Pectobacterium atrosepticum, P. carotovorum subsp. brasiliensis, dan Dickeya spp.) (Ngadze et al. 2012). Asam salisilat memengaruhi aktivitas enzim-enzim antioksidan seperti hidrogen peroksida yang bekerja sebagai second messenger dalam mengatur respons pertahanan tanaman (Harfouche et al. 2008).
Hasil penelitian membuktikan bahwa asam salisilat berperan penting dalam meningkatkan induksi ketahanan tanaman padi terhadap patotipe X. oryzae pv. oryzae. Agens penginduksi asam salisilat berpotensi untuk dikembangkan dalam pengendalian penyakit HDB.
Perlindungan tanaman terhadap patogen melalui teknologi induksi ketahanan dapat dilakukan secara eksogen. Induksi ketahanan tanaman dapat digunakan sebagai agens penginduksi kimia atau biologi yang menjadi komponen penting dalam program pengelolaan penyakit tanaman.
Perlakuan konsentrasi SA10 mM secara tunggal terbukti efektif dapat menginduksi ketahanan varietas Conde dan Ciherang untuk menekan X. oryzae pv. oryzae patotipe IV dan VIII. Perlakuan asam salisilat mampu meningkatkan aktivitas enzim peroksidase dan polifenoloksidase pada varietas IR64 dan Conde serta β-1,3-glukanase pada ketiga varietas. Mekanisme aplikasi asam salisilat dalam menekan X. oryzae pv. oryzae masih perlu dikaji lebih dalam pada ekosistem padi yang berbeda.
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Badan Litbang Pertanian, Kementerian Departemen Pertanian yang telah mendanai
sebagian kegiatan penelitian ini melalui program KKP3N dengan No. kontrak: 691/LB.620/I.1/2/2013.
DAFTAR PUSTAKA
An C, Mou Z. 2011. Salicylic Acid and its function in plant immunity. J Integrative Plant Biol. 53(6):412–428. DOI: https://doi.org/10.1111/j.1744-7909.2011.01043.x.
Ankenbauer RG, Cox CD. 1988. Isolation and characterization of Pseudomonas aeruginosa mutants requiring salicylic acid for pyochelin biosynthesis. J Bacteriol. 170:5364–5367. DOI: ht tps: / /doi .org/10.1128/jb.170.11.5364-5367.1988.
Ezuka A, Kaku H 2000. A historical review of bacterial blight of rice. Bull Nat Inst Agrobiol Res. 15(2000):1–207.
Hammerschmidt R, Nuckles M, Kuc J. 1982. Association of enhanced peroxidase activity with induced systemic resistance o f c u c u m b e r t o C o l l e t o t r i c h u m lagenarium. Physiol Plant Pathol. 20:73–82. DOI: https://doi.org/10.1016/0048-4059(82)90025-X.
Harfouche A, Rugini E, Mencarelli F, Botondi R, Muleo R. 2008. Salicylic acid induces H2O2 production and endochitinase gene expression but not ethylene biosynthesis in Castanea sativa in vitro model system. J Plant Physiol. 165:734–744. DOI: https://doi.org/10.1016/j.jplph.2007.03.010.
Hayat Q, Hayat S, Irfan M, Ahmad A. 2010. Effect of exogenous salicylic acid under changing environment: a review. Environ Exp Bot. 68(1):14–25.
D O I : h t t p s : / / d o i . o r g / 1 0 . 1 0 1 6 / j .envexpbot.2009.08.0055.
Holeski L, Jander G, Agrawal A. 2012. Transgenerational defense induction and epigenetic inheritance in plants. Trends Ecol Evol. 27:618–626. DOI: https://doi.org/10.1016/j.tree.2012.07.011.
Ibrahim YE. 2012. Activities of antioxidants enzymes in salicylic acid treated tomato against Xanthomonas vesicatoria. Afr J Microbiol Res. 6(27):5678–5682.

J Fitopatol Indones Leiwakabessy et al.
214
[IRRI] International Rice Research Institute. 2014. Standard evaluation system for rice. Manila (PH): IRRI.
Kim SI, Song JT, Jeong JY, Seo HS. 2015. Niclosamide inhibits leaf blight caused by Xanthomonas oryzae in rice. Nature. 6:21209. DOI: https://doi.org/10.1038/srep21209.
Krishanti NPRA, Wahyudi AT, Nawangsih AA. 2015. Non-pathogenic phyllosphere bacteria producing bioactive compounds as biological control of Xanthomonas oryzae pv oryzae. Int J Pharma Bio Sci. 6(1):801–810.
Kobeasy MI, El-Beltagi HS, El-Shazly MA, Khattab EAH. 2011. Induction of resistance in Arachis hypogaea L. against Peanut mottle virus by nitric oxide and salicylic acid. Physiol Mol Plant Pathol. 76(2):112–118. DOI: https://doi.org/10.1016/j.pmpp.2011.07.005.
Mandal S, Mallick N, Mitra A. 2009. Salicylic acid-induced resistance to Fusarium oxysporum f. sp. lycopersici in tomato. Plant Physiol Biochem. 47(7):642–649. DOI: ttps://doi.org/10.1016/j.plaphy.2009.03.001.
Madden L, Hughes G, van den Bosch F. 2007. The Study of Plant Disease Epidemics. St. Paul (US): APS Press.
Malick C, Singh M. 1980. Phenolics. Plant enzymology. Histo Enzymol. 190:286.
Mohan-Babu R, Saajena A, Samundeesar V, Sreedhar A, Vidhyasekeran P, Reddy MS. 2003. Induction of bacterial blight (Xanthomonas oryzae pv. oryzae) resistance in rice by treatment with acibenzolar-S-methyl. Ann Appl Biol. 143(2003):333–340. DOI: https://doi.org/10.1111/j.1744-7348.2003.tb00302.x.
Mustafa A, Saleem IY, Mahmood S, Hannan A, Akhtar M. 2013. Field evaluation of new fungicides against rice (Oryza sativa) diseases. Pak J Phytopathol. 25(02):141–145.
Nayak D, Bose LK, Singh UD, Singh S, Nayak P. 2008. Measurement of genetic diversity of virulence in populations of
Xanthomonas oryzae pv. oryzae in India. Comm Biomet Crop Sci. 3(1):16–28.
Ngadze E, Icishahayo D, Coutinho T, van der Waals J. 2012. Role of polyphenol oxidase, peroxidase, phenylalanine ammonia lyase, chlorogenic acid, and total soluble phenols in resistance of potatoes to soft rot. Plant Dis. 96:186–192. DOI: https://doi.org/10.1094/PDIS-02-11-0149.
Ou SH. 1985. Rice Disease 2nd edition. London (UK): CMI. Hlm 30.
OEPP/EPPO 2007. Xanthomonas oryzae. 37(3): 543–553. DOI: https://doi.org/10.1111/j.1365-2338.2007.01162.x.
Pan S, Ye X, Kuc J. 1991. Association of β-1,3-glucanase activity and isoform pattern with systemic resistance to blue mold in tobacco induced by stem injection with Peronospora tabacina or leaf inoculation with tobacco mosaic virus. Physiol Mol Plant Pathol. 39(1):25–39. DOI: https://doi.org/10.1016/0885-5765(91)90029-H.
Putra C, Giyanto. 2014. Kompatibilitas Bacillus spp. dan aktinomiset sebagai agens hayati Xanthomonas oryzae pv. oryzae dan pemacu pertumbuhan Padi. J Fitopatol Indones. 10(5):160–169. DOI: https://doi.org/10.14692/jfi.10.5.160.
Silverman, PM, Seskar D, Kanter P, Schweizer JP, Mėtraux I, Raskin I. 1995. Salicylic acid in rice biosynthesis, conjugation and possible role. Plant Physiol. 108: 633–639. DOI: https://doi.org/10.1104/pp.108.2.633.
Sudir, Yuliani D. 2016. Composition and distribution of Xanthomonas oryzae pv. oryzae pathotypes, the pathogen of rice bacterial leaf blight in Indonesia. Agrivita. 38(2):174–185. DOI: https://doi.org/10.17503/agrivita.v38i2.588.
Tasliah. 2012. Gen ketahanan tanaman padi terhadap bakteri hawar daun (Xanthomonas oryzae pv. oryzae). J Litbang Pertanian. 31(3):103–112.
Van der Planck J. 1963. Plant Diseases: Epidemics and Control. New York (US): Academic Press.

J Fitopatol Indones Leiwakabessy et al.
215
Vicente R, Plasencia J. 2011. Salicylic acid beyond defence: its role in plant growth and development. J Exp Bot. 62(10):3321–3338. DOI: https://doi.org/10.1093/jxb/err031.
Zamaninejad M, Khorasani S, Moeini M, Heidarian A. 2013. Effect of salicylic acid
on morphological characteristics, yield and yield components of corn (Zea mays L.) under drought condition. Eur J Exp Biol. 3(2):153–161.

ISSN: 0215-7950
216
Volume 13, Nomor 6, November 2017Halaman 216–228
DOI: 10.14692/jfi.13.6.216
*Alamat penulis korespondensi: Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor. Jalan Kamper, Kampus IPB Darmaga, Bogor 16680. Tel: 0251-8629364, Faks: 0251-862362, Surel: [email protected].
Keanekaragaman dan Patogenisitas Fusarium spp. Asal Beberapa Kultivar Pisang
Diversity and Pathogenicity of Fusarium spp. Isolated from Several Banana Cultivars
Widya Sari, Suryo Wiyono*, Ali Nurmansyah, Abdul Munif, Roedhy PoerwantoInstitut Pertanian Bogor, Bogor 16680
ABSTRAK
Penyakit layu fusarium yang disebabkan oleh cendawan patogen Fusarium oxysporum f. sp. cubense merupakan penyakit penting pada tanaman pisang. Penelitian ini bertujuan mengidentifikasi spesies Fusarium yang diisolasi dari beberapa kultivar tanaman pisang bergejala layu secara morfologi dan molekuler. Isolat-isolat Fusarium yang diperoleh selanjutnya diuji tingkat patogenisitasnya, dan sebarannya pada berbagai bagian tanaman pisang. Berdasarkan pengamatan beberapa karakter morfologi (laju pertumbuhan, warna koloni, jumlah konidium, bentuk makrokonidium, mikrokonidium, dan klamidospora), F. oxysporum f. sp. cubense dapat dibedakan dengan spesies Fusarium yang lain akan tetapi tidak dapat membedakan ras atau galurnya. Hasil identifikasi molekuler menggunakan primer spesifik VCG 01213/16 dan Foc1/Foc2 diperoleh berturut-turut 13 isolat F. oxysporum f. sp. cubense TR4, dan 7 isolat F. oxysporum f. sp. cubense non TR4; sedangkan menggunakan primer universal ITS4/ITS5 diidentifikasi 7 isolat F. solani, 1 isolat F. verticillioides dan 2 isolat bukan Fusarium sp. Cendawan F. oxysporum f. sp. cubense TR4 dan non-TR4 banyak ditemukan di bagian batang semu tanaman pisang, sedangkan F. solani dan F. verticillioides dominan ditemukan di bonggol pisang. Uji patogenisitas menunjukkan bahwa isolat F. oxysporum f. sp. cubense non TR4 dan TR4 mampu menimbulkan nekrosis pada bonggol bibit pisang kultivar Ambon Kuning. Gejala yang sama ditemukan pada hasil inokulasi F. solani tetapi dengan keparahan yang lebih rendah.
Kata kunci: batang semu, Fusarium oxysporum f. sp. cubense, kultivar Ambon Kuning, nekrosis
ABSTRACT
Fusarium wilt disease caused by pathogenic fungus F. oxysporum f. sp. cubense is an important disease in banana plants. This research was aimed to identifiy Fusarium spp. collected from banana plants showing wilt disease based on morphology and molecular characters. The fungi isolates obtained was further examined for its pathogenicity and distribution in various parts of banana plants. Based on morphological characters, i.e. growth rate, total conidia, colony color, macroconidium, microconidium, and chlamydospores, F. oxysporum f. sp. cubense can be differentiated from other species of Fusarium, but not the strain. Molecular analysis using specific primers VCG 01213/16 and Foc1/Foc 2 successfully identified 13 and 7 isolates of F. oxysporum f. sp. cubense TR4 and F. oxysporum f. sp. cubense non TR4, respectively. Analysis using universal primer ITS4/ITS5 identified 7 isolates of F. solani, 1 isolate of F. verticillioides, and 2 isolates non Fusarium. Infection of F. oxysporum f. sp. cubense TR4 and non-TR4 were found mostly in pseudo stems, whereas F. solani and F. verticillioides dominantly colonized

J Fitopatol Indones Sari et al.
217
banana corm. Pathogenicity test showed that F. oxysporum f. sp. cubense TR4 and non-TR4 caused necrosis on corm of cv. Ambon Kuning. Similar necrosis symptom was also observed on infection of F. solani but with less severity.
Key words: Ambon Yellow, F. oxysporum f. sp. cubense, necrosis, pseudo stem
PENDAHULUAN
Beberapa provinsi di Indonesia, di antaranya Sumatera Utara, Sumatera Barat dan Jawa Barat tercatat sebagai sentra produksi pisang (Kementan 2015). Jawa Barat berada di peringkat pertama sebagai produsen pisang pada tahun 2011. Sayangnya, produksi pisang dari Jawa Barat semakin menurun karena penyakit layu fusarium di beberapa kabupaten sentra produksi pisang, yaitu Cianjur, Majalengka, dan Bandung Barat (Pusdatin 2014).
Banyak faktor yang menyebabkan rendahnya produksi pisang di Indonesia salah satunya ialah penyakit layu yang disebabkan oleh cendawan F. oxyporum Schlecht f. sp cubense. Williams et al. (2010) melaporkan bahwa penyakit layu fusarium merupakan faktor utama penyebab menurunnya produksi pisang di Indonesia. Cendawan terdiri atas beberapa ras dan galur dengan tingkat virulensi yang berbeda, serta mempunyai kemampuan bertahan dalam tanah tanpa inang utama hingga 40 tahun (Ploetz 1990).
Penelitian penyakit layu fusarium pisang serta distribusi ras F. oxysporum f. sp. cubense pada berbagai varietas dan lokasi sudah banyak dilakukan, termasuk distribusi VCG F. oxysporum f. sp. cubense pada beberapa kultivar pisang di beberapa provinsi di Indonesia. Williams et al. (2010) telah berhasil mengidentifikasi 8 kelompok VCG F. oxysporum f. sp. cubense (0120/15, 0121, 0123, 0124/5, 0126, 01213/16, 01218 dan 01219) yang tersebar di Aceh, Sumatera Barat, Yogyakarta, Kalimantan Barat, Kalimantan Timur, Sulawesi Tenggara dan Sulawesi Utara. Semua VCG menyerang hampir semua varietas pisang, tapi varietas Barangan, Raja Bulu, dan Ambon Putih merupakan varietas dengan tingkat serangan tertinggi. F. oxysporum f. sp. cubense VCG 01213/16 yang dikenal sebagai F. oxysporum f. sp. cubense ras 4 tropika
(TR4) merupakan kelompok yang paling dominan ditemukan Ploetz (2006). Leslie dan Summerell (2006) melaporkan terdapat lebih dari 10 Fusarium spp. yang berasosiasi dengan pisang di dunia, termasuk F. camptoceras, F. concentricum, F. musarum, F. proliferatum, F. semitectum (F. pallidoroseum, F. incarnatum), F. compactum, F. thapsinum, F. verticillioides, dan F. subglutinans.
Penelitian tentang beberapa spesies Fusarium yang berasosiasi dengan tanaman pisang bergejala layu, sebarannya pada beberapa kultivar dan bagian tanaman pisang serta pengujian patogenisitas dari masing-masing spesies tersebut pada pisang belum pernah dilakukan di Indonesia. Tujuan penelitian ini: menentukan karakter morfologi dan molekuler cendawan Fusarium spp. yang terdapat pada berbagai kultivar dan bagian tanaman pisang yang menunjukkan gejala layu; menentukan ras F. oxysporum f. sp. cubense dari berbagai kultivar pisang yang bergejala layu fusarium dan distribusinya pada bagian tanaman pisang; serta menentukan tingkat patogenisitasnya secara in planta pada pisang kultivar Ambon Kuning.
BAHAN DAN METODE
Pengambilan Sampel di Lapangan Pengambilan sampel dilakukan di daerah
endemik penyakit layu fusarium pada pisang, yaitu di Jawa Barat dan Sumatera Barat. Metode pengambilan sampel dilakukan secara komposit 8 desa di Kabupaten Cianjur Jawa Barat dan 2 desa di Sungai Lantik Kabupaten Lima Puluh Kota, Sumatera Barat dengan minimal luas lahan 100 m2. Sampel diambil dari bagian batang semu, bonggol dan akar tanaman pisang yang menunjukkan gejala layu. Sampel tanah diambil secara komposit pada 5 titik di rizosfer tanaman pisang.

J Fitopatol Indones Sari et al.
218
Isolasi Fusarium spp. Jaringan batang semu dipotong dengan
ukuran 0.5–1 cm sedangkan bonggol dan akar pisang sakit dipotong dengan ukuran 1 × 1 cmkemudian disterilisasi permukaan dengan alkohol 70% dan dibilas akuades steril. Potongan sampel diletakkan pada medium agar-agar air (AA) dan diinkubasi selama 3–5 hari pada suhu kamar.
Isolasi Fusarium spp. dari tanah rizosfer dilakukan dengan membuat suspensi tanah dalam air steril dengan perbandingan 1:50 (b/v). Sebanyak 0.1 mL suspensi diteteskan di atas medium martin agar (Martin 1950), diratakan dengan gelas penyebar, dan diinkubasi pada suhu 27 °C selama 3–5 hari. Koloni diamati menggunakan mikroskop. Koloni yang memiliki ciri spesifik Fusarium, yaitu mempunyai makrokonidium, mikrokonidium, dan klamidospora dimurnikan pada medium agar-agar dekstrosa kentang (ADK) komposisi ¼ resep untuk menjaga virulensinya dan disimpan untuk pengujian selanjutnya.
Identifikasi Morfologi dan Molekuler Fusarium spp.
Biakan Fusarium spp. diremajakan pada medium ADK komposisi 1/4 resep (Dita et al. 2010)dan diinkubasi pada suhu ruang. Pengamatan dilakukan pada hari ke-3, 5, dan 7 yang meliputi laju pertumbuhan koloni, warna koloni, kepadatan konidium (dihitung menggunakan haemositometer), bentuk makrokonidium, mikrokonida, dan klamidospora. Penghitungan laju pertumbuhan koloni menggunakan rumus laju pertumbuhan eksponensial, yaitu
r = 1t ln( (Pt
Po , dengan
r, laju pertumbuhan; t, waktu pengamatan; Pt, diameter koloni cendawan pada pengamatan terakhir; Po, diameter koloni cendawan pada pengamatan awal (Leslie dan Summerell 2006).
Idetifikasi molekuler semua isolat Fusarium spp. dilakukan dalam beberapa tahapan. Tahap awal ialah ekstraksi total DNA menggunakan sodium dodecyl sulfate (SDS) (Aamir et al. 2015) yang sudah dimodifikasi, dilanjutkan dengan amplifikasi DNA dengan
teknik PCR. Bahan yang digunakan pada teknik PCR, yaitu DreamTaq PCR Master Mix (ThermoscientificTM, US), 2 primer spesifik Foc1/Foc2, 242 pb (Lin et al. 2009), dan F. oxysporum f. sp. cubense TR4-R, 462 pb (Dita et al. 2010) serta 1 primer universal ITS4 dan ITS5, 550 pb (White et al. 1990). Hasil amplifikasi di elektroforesis menggunakan gel agarosa 2% dengan pewarnaan etidium bromida (0.5 µg mL-1) pada tegangan 100 voltselama 20 menit. Hasil visualisasi didokumentasi menggunakan geldoc digital imaging system (Gel imaging G:Box Chemi XX6, AlphaMetricBiotech, Germany (DE)).
Produk amplifikasi dengan primer universal ITS4 dan ITS5 selanjutnya disikuensing di PT Genetica Science Indonesia kemudian dibandingkan urutan DNA dengan yang lainnya menggunakan program BLAST dari National Center for Biotechnology Information (NCBI). Hasil sikuensing selanjutnya dilakukan penyejajaran menggunakan program Chromaspro versi 1.7.5. Analisis filogenetika menggunakan piranti lunak MEGA versi 6.0 untuk windows (Tamura et al. 2013).
Patogenisitas pada Bibit PisangUji patogenisitas semua isolat dilakukan
pada bibit pisang kultivar Ambon Kuning hasil kultur jaringan berumur 1.5 bulan setelah aklimatisasi. Pengujian ini menggunakan Rancangan Acak Lengkap (RAL) dengan 30 perlakuan dan 4 ulangan, perlakuan tersebut adalah 13 isolat Foc TR4, 7 isolat Foc non TR4, 1 isolat F. verticilloides, 7 isolat F. solani dan 2 isolat non Fusarium sebagai kontrol, yaitu isolat T. polyzona (isolat asal tanah rizosfer tanaman pisang kultivar Nangka) dan Clonostachys rosea (isolat asal batang tanaman pisang kultivar Ambon Kuning). Bibit pisang uji ditanam pada medium tanam campuran tanah dan kompos dengan perbandingan 2:1 (b/b). Medium tanam disterilisasi dan diinokulasi masing-masing 10 g biakan 30 isolat uji dalam medium beras dengan kerapatan konidia 107 cfu mL-1.
Pengamatan diskolorasi pada bonggol dilakukan dengan memotong bonggol bibit

J Fitopatol Indones Sari et al.
219
umur 60 hari setelah inokulasi. Pengamatan keparahan penyakit diukur menggunakan skor sebagai berikut: 0, tidak ada diskolorasi; 1, diskolorasi 0%<V≤10%; 2, diskolorasi 10%<V≤ 33%; 3, diskolorasi 33%<V≤66%; 4 , d i s k o l o r a s i 6 6 % < V ≤ 1 0 0 % ; d a n 5, diskolorasi 100% (Corderio 1994).
HASIL
Penelitian ini menghasilkan 30 isolat yang berasal dari batang semu, bonggol, akar, dan tanah rizosfer. Sebanyak 21 isolat berasal dari kultivar Ambon Kuning, 5 isolat dari pisang Nangka, serta masing-masing 2 isolat berasal dari kultivar Raja Sereh dan Raja Bulu. Isolat terbanyak didapat dari batang semu pisang, yaitu sebanyak 14 isolat, sementara 10 isolat didapat dari bonggol, 2 isolat dari akar, dan 4 isolat dari tanah.
Berdasarkan pengamatan laju per-tumbuhan, warna koloni, jumlah konidium, bentuk konidia dan klamidospora didapatkan Fusarium yang beragam. Beberapa memiliki ciri khas F. oxysporum (Isolat BG2, BT3, BG6, BT11, BT14A, BT14B, BT14C, BT15, BT17, BT18, BT18C, TA23A, A24, BG10B, BT12, BT16A, BT16B, BT16C, BT28, BT29); dan F. solani (isolat BG1, BG4, BG10A, A19, A20, A21, TA23) serta 1 isolat BG7 mempunyai ciri makro dan mikro konidium yang berbeda dengan F. oxysporum dan F. solani (Tabel 1).
Kisaran laju pertumbuhan semua isolat adalah 0.17–0.38 cm per hari (Gambar 1).Beberapa islat dari F. oxysporum f. sp. cubense (BG2, BT3A, BT12, BT14A, BT14B, BT16A, BT16B), dan beberapa isolat F. solani (BG1, BG4, BG10A) serta isolat BG7 (F. verticilloides) mempunyai laju pertumbuhan yang lebih tinggi dibanding dari isolat lainnya.
Warna koloni untuk setiap kelompok Fusarium spp. didominasi oleh warna putih, tetapi beberapa isolat F. oxysporum f. sp. cubense non TR4 dan TR4 juga memiliki warna koloni merah muda, ungu muda, dan krem. Tipe koloni didominasi oleh tipe seperti kapas dan tipis. Pada masing-masing isolat Fusarium spp. terdapat sporodokium.
Cendawan F. oxysporum mempunyai makrokonidium dan mikrokonidium dengan karakter yang berbeda dengan isolat F. solani. Makrokonidium F. oxysporum berbentuk seperti bulan sabit dengan beberapa sekat, mikrokonidium berbentuk oval atau seperti ginjal, serta fialid ada yang tunggal atau bercabang. Mikrokonidium terbentuk pada fialid. Ada yang tunggal, berkelompok pada ujung fialid dan tersusun seperti rantai. Klamidospora berbentuk bulat, berdinding tebal, terbentuk di ujung hifa, di tengah hifa atau makrokonidium dan bisa tersusun seperti rantai.
Makrokonidium F. oxysporum f. sp. cubense memiliki ukuran 27–55 × 3.3–5.5 μm,sel berbentuk lurus atau sedikit bengkok dengan 4–8 sekat dengan ujung atas runcing dan tujung bawah datar. Mikrokonidium berbentuk bulat atau menyerupai ginjal, berukuran 5–16 × 2.4–3.5 μm, memiliki 1 atau 2 sekat. Fialid dan mikrokonidium berkelompok dengan ujung miring ke kiri. Klamidospora berdiameter 7–11 μm, berbentuk bulat secara tunggal atau berantai. F. solani dibedakan dengan F. oxysporum berdasarkan karakteristik bentuk makrokonidium juga bentuk mikrokonidium yang relatif lebih besar untuk F. solani dan mikrokonidium umumnya berbentuk elips (Gambar 2).
Secara umum kepadatan kondium berkisar 7.71–9.4 cm-2 (data dikonversi menggunakan log10). Analisis molekuler pada 30 isolat menggunakan primer Foc1/Foc2 berhasil mengamplifikasi 7 isolat F. oxysporum f. sp. cubense nonTR4 dengan ukuran ~242 pb, sedangkan amplifikasi menggunakan primer FocTR4 menghasilkan 13 isolat positif F. oxysporum f. sp. cubense TR4 berukuran ~462 pb (Gambar 3). Penentuan ras F. oxysporum f. sp. cubense nonTR4 dilakukan dengan cara membandingkan hasil amplifikasi kedua primer spesifik F. oxysporum f. sp. cubense yang digunakan. Apabila dapat diamplifikasi oleh kedua primer F. oc1/Foc 2 dan TR4, berarti isolat merupakan F. oxysporum f. sp. cubense TR4, begitu juga bila hanya teramplifikasi pada primer TR4 saja. Tetapi apabila hanya teramplifikasi oleh
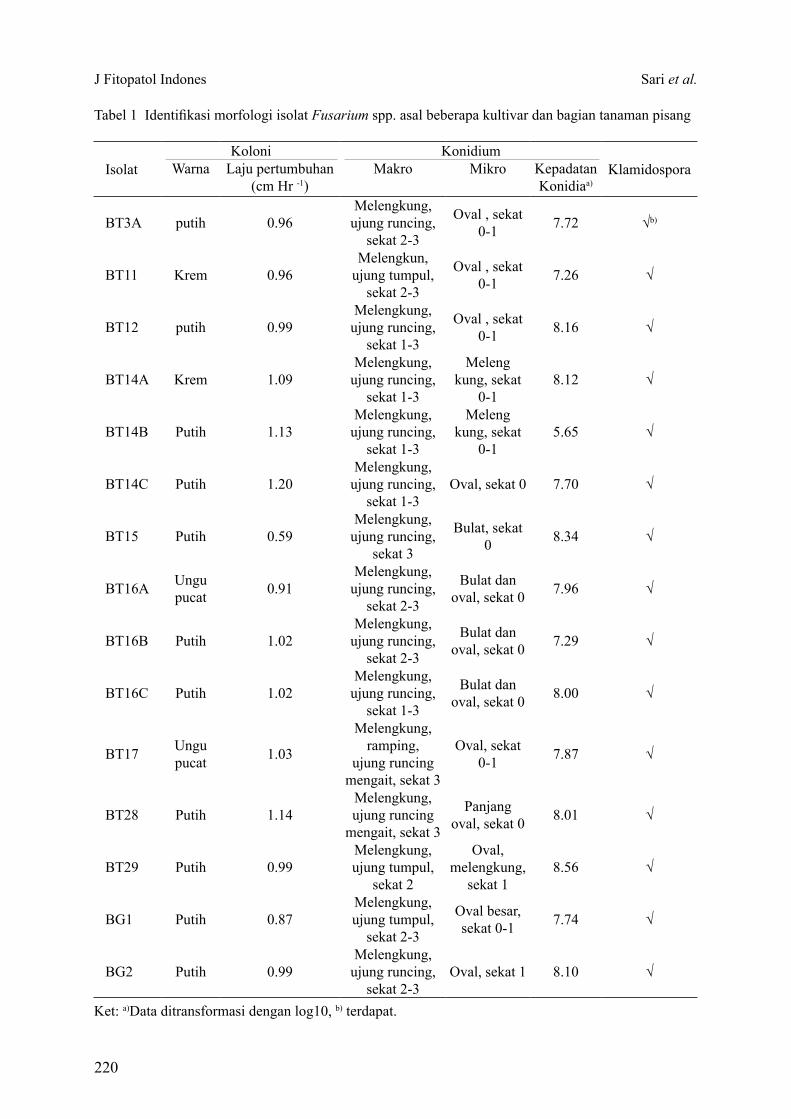
J Fitopatol Indones Sari et al.
220
Tabel 1 Identifikasi morfologi isolat Fusarium spp. asal beberapa kultivar dan bagian tanaman pisang
Ket: a)Data ditransformasi dengan log10, b) terdapat.
IsolatKoloni Konidium
KlamidosporaWarna Laju pertumbuhan(cm Hr -1)
Makro Mikro Kepadatan Konidiaa)
BT3A putih 0.96Melengkung,
ujung runcing, sekat 2-3
Oval , sekat 0-1 7.72 √b)
BT11 Krem 0.96Melengkun,
ujung tumpul, sekat 2-3
Oval , sekat 0-1 7.26 √
BT12 putih 0.99Melengkung,
ujung runcing, sekat 1-3
Oval , sekat 0-1 8.16 √
BT14A Krem 1.09Melengkung,
ujung runcing, sekat 1-3
Meleng kung, sekat
0-18.12 √
BT14B Putih 1.13Melengkung,
ujung runcing, sekat 1-3
Meleng kung, sekat
0-15.65 √
BT14C Putih 1.20Melengkung,
ujung runcing, sekat 1-3
Oval, sekat 0 7.70 √
BT15 Putih 0.59Melengkung,
ujung runcing, sekat 3
Bulat, sekat 0 8.34 √
BT16A Ungu pucat 0.91
Melengkung, ujung runcing,
sekat 2-3
Bulat dan oval, sekat 0 7.96 √
BT16B Putih 1.02Melengkung,
ujung runcing, sekat 2-3
Bulat dan oval, sekat 0 7.29 √
BT16C Putih 1.02Melengkung,
ujung runcing, sekat 1-3
Bulat dan oval, sekat 0 8.00 √
BT17 Ungu pucat 1.03
Melengkung, ramping,
ujung runcing mengait, sekat 3
Oval, sekat 0-1 7.87 √
BT28 Putih 1.14Melengkung, ujung runcing
mengait, sekat 3
Panjang oval, sekat 0 8.01 √
BT29 Putih 0.99Melengkung, ujung tumpul,
sekat 2
Oval, melengkung,
sekat 18.56 √
BG1 Putih 0.87Melengkung, ujung tumpul,
sekat 2-3
Oval besar, sekat 0-1 7.74 √
BG2 Putih 0.99Melengkung,
ujung runcing, sekat 2-3
Oval, sekat 1 8.10 √

J Fitopatol Indones Sari et al.
221
...Lanjutan Tabel 1
IsolatKoloni Konidium
KlamidosporaWarna Laju pertumbuhan(cm Hr -1)
Makro Mikro Kepadatan Konidiaa)
BG4 Putih 0.73Melengkung, ujung tumpul,
sekat 2-3
Oval besar, sekat 0-1 7.98 √
BG6 Putih 1.00Melengkung, ujung runcing, sekat 2-3
Berbentuk ginjal, sekat 1
7.85 √
BG7 Merah muda 0.90
Panjang, sedikit melengkung, ujung Runcing, sekat 3
Memanjang, sekat 1 7.91 -
BG10A Putih 1.02Melengkung, ujung tumpul, sekat 2
Oval, melengkung, sekat 1
7.01 √
BG10B Ungu pucat 1.14
Melengkung, ujung runcing, sekat 2-3
Memanjang, sekat 0 7.60 √
BG18 Putih 1.10Melengkung, ujung tumpul, sekat 2
Oval, melengkung, sekat 1
7.59 √
BG18C Krem 1.14Melengkung, ujung runcing, sekat 3
Panjang oval, sekat 0 7.07 √
A19 Putih 1.14Melengkung, ujung tumpul, sekat 2-3
Oval besar, sekat 0-1 7.47 √
A20 Putih 1.14Melengkung, ujung tumpul, sekat 3
Oval, sekat 0 7.73 √
TA21 Putih 1.14Melengkung, ujung tumpul, sekat 2-3
Oval besar, sekat 0-1 8.05 √
TA23 Krem 1.12Melengkung, ujung runcing, sekat 3
Oval, sekat 1 7.74 √
TA23A Putih 1.10Melengkung, ujung runcing, sekat 3
Oval, sekat 1 7.84 √
TA24 Putih 1.10Melengkung, ujung runcing, sekat 3
Oval, sekat 1 7.84 √
Ket: a)Data ditransformasi dengan log10, b) terdapat.
primer Foc1/Foc2, dapat diasumsikan bahwa isolat-isolat tersebut positif F. oxysporum f. sp. cubense ras 1 atau ras 2. Pada penelitian ini tidak dilakukan analisis VCG sehingga tidak didapatkan VCG spesifik yang merujuk pada ras-ras F. oxysporum f. sp. cubense lainnya.
Hasil BLAST-N dari sikuen produk PCR dengan primer ITS4 dan ITS5 yang teramplifikasi pada ~550 pb menghasilkan 7 isolat yang identik dengan F. solani, satu isolat F. verticillioides sedangkan dua isolat terakhir, yaitu isolat TA25 dan BT26 tidak
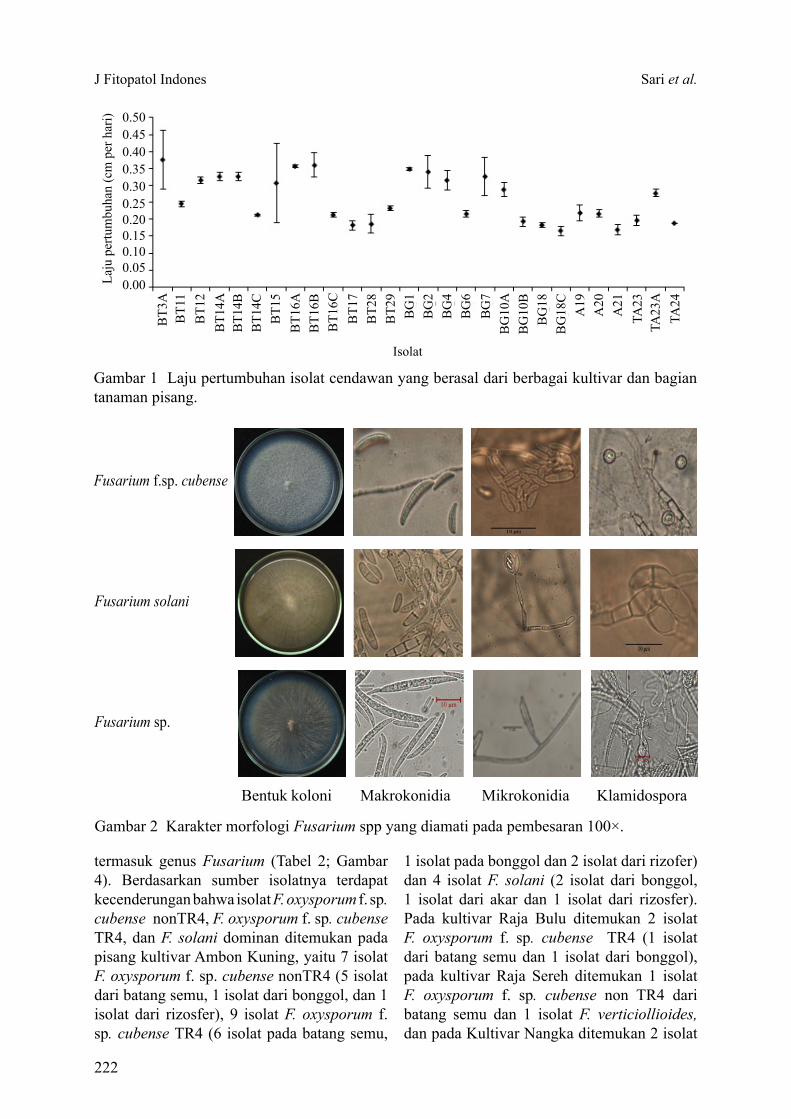
J Fitopatol Indones Sari et al.
222
Gambar 2 Karakter morfologi Fusarium spp yang diamati pada pembesaran 100×.
Bentuk koloni Makrokonidia Mikrokonidia Klamidospora
Fusarium f.sp. cubense
Fusarium solani
Fusarium sp.
Gambar 1 Laju pertumbuhan isolat cendawan yang berasal dari berbagai kultivar dan bagian tanaman pisang.
Isolat
0.500.450.400.350.300.250.200.150.100.050.00La
ju p
ertu
mbu
han
(cm
per
har
i)
BT14
CB
T15
BT1
6AB
T16B
BT1
6C
BT3
AB
T11
BT1
2BT
14A
BT14
B
BT1
7B
T28
BT2
9B
G1
BG
2B
G4
BG
6B
G7
BG
10A
BG
10B
BG
18B
G18
CA
19A
20A
21TA
23TA
23A
TA24
termasuk genus Fusarium (Tabel 2; Gambar 4). Berdasarkan sumber isolatnya terdapat kecenderungan bahwa isolat F. oxysporum f. sp. cubense nonTR4, F. oxysporum f. sp. cubense TR4, dan F. solani dominan ditemukan pada pisang kultivar Ambon Kuning, yaitu 7 isolat F. oxysporum f. sp. cubense nonTR4 (5 isolat dari batang semu, 1 isolat dari bonggol, dan 1 isolat dari rizosfer), 9 isolat F. oxysporum f. sp. cubense TR4 (6 isolat pada batang semu,
1 isolat pada bonggol dan 2 isolat dari rizofer) dan 4 isolat F. solani (2 isolat dari bonggol, 1 isolat dari akar dan 1 isolat dari rizosfer). Pada kultivar Raja Bulu ditemukan 2 isolat F. oxysporum f. sp. cubense TR4 (1 isolat dari batang semu dan 1 isolat dari bonggol), pada kultivar Raja Sereh ditemukan 1 isolat F. oxysporum f. sp. cubense non TR4 dari batang semu dan 1 isolat F. verticiollioides, dan pada Kultivar Nangka ditemukan 2 isolat

J Fitopatol Indones Sari et al.
223
Gambar 3 Hasil amplifikasi isolat Fusarium spp. A, F. oxysporum f. sp. cubense TR4 menggunakan primer FocTR4 (Isolat BG2, BT3, BG6, BT11, BT14A, BT14B, BT14C, BT15, BT17, BG18, BG18C, TA23A, TA24); B, F. oxysporum f. sp. cubense non TR4 menggunakan primer Foc1/Foc2 (Isolat BG10B, BGT12, BT16A, BT16B, BT16C, BT28, BT29); C, Fusarium spp. dan non Fusarium menggunakan primer ITS4 dan ITS5 (ISOLAT BG1,BG 4, BT7, BGT10A, A19, A20, A21, TA23, TA25, BT26); M, Penanda 1 kb (Thermo scientific).
BG2 BT3 BG6 BT11 BT14A BT14 BT14C BT15 BT17 BG18 BG18C TA23A TA24M
462 pbA
242 pbB
BG10B BT12 BT16A BT16B BT16C BT28 BT29M
550 pbC
BG1 BG4 BT7 BG10A A19 A20 A21 TA23 TA25 BT26M
F. oxysporum f. sp. cubense TR4 dan 2 isolat F. solani.
Hasil uji patogenisitas isolat Fusarium spp. yang sudah dikarakteriasi secara morfologi dan molekuler pada bibit pisang Ambon Kuning menunjukkan bahwa beberapa isolat F. oxysporum f. sp. cubense nonTR4 dan TR4 mampu menimbulkan nekrosis pada bonggol pisang Ambon kuning dengan skala keparahan yang bervariasi. Beberapa isolat F. oxysporum
f. sp. cubense nonTR4 (isolat BT12, BT29) mempunyai nilai skala keparahan nekrosis yang lebih tinggi dibandingkan dengan isolat F. oxysporum f. sp. cubense TR4. Isolat BG2 merupakan isolat F. oxysporum f. sp. cubense TR4 yang memiliki nilai skala keparahan yang lebih tinggi dibanding dengan F. oxysporum f. sp. cubense TR4 lainnya.
Isolat F. solani (BG1 dan BG4) mampu menimbulkan gejala nekrosis yang sama
Tabel 2 Tingkat homologi sikuen nukleotida isolat non Fusarium oxysporum f. sp. cubense berdasarkan ITS yang dibandingkan data GenBank NCBI
Isolat Deskripsi Query Cover (%)
Identitas(%) No Aksesi
BG1 Fusarium solani BG1 99 100 FJ224382.1BG4 Fusarium solani BG4 99 100 FJ224382.1BG10A Fusarium solani BG10A 99 100 FJ224382.1A19 Fusarium sp. A19 100 99 EU750681.1A20 Fusarium sp. A20 100 99 EU750681.1A21 Fusarium sp. A21 100 99 EU750681.1TA23 Fusarium solani TA23 99 100 JQ277276.1BG7 Fusarium verticillioides BG7 100 100 KM434131.1TA25 Trametes polyzona TA25 100 99 JN164978.1BT26 Clonostachys rosea BT26 100 99 HQ607798.1

J Fitopatol Indones Sari et al.
224
Gambar 4 Pohon filogenetika analisis sikuen nukleotida daerah ITS menggunakan metode neigbour-joining dan model jukes and cantor. Nilai Bootstrap 1000. Huruf yang dicetak tebal menunjukkan isolat yang diteliti.
BG7
TA23
BG1BG4BG10A
A19A20A21
BT26
TA25
dengan yang ditimbulkan F. oxysporum f. sp. cubense, walaupun dengan tingkat keparahan yang lebih rendah (Tabel 3; Gambar 5). Sementara itu pengujian dengan F. verticillioides (Isolat BG7) memperlihatkan sedikit perubahan warna pada bonggol bibit pisang tetapi belum sampai menyebabkan nekrosis pada bonggol bibit pisang.
PEMBAHASAN
Fusarium spp. merupakan cendawan yang mempunyai sebaran inang luas. F.
oxysporum f. sp. cubense merupakan 1 dari 6 patogen yang menyerang pembuluh tanaman dan sangat merugikan komoditi pertanian di dunia. Cendawan ini tersebar luas pada daerah penghasil pisang di Asia, Afrika, Australia, dan Amerika Tengah serta Selatan yang mampu mematikan tanaman pisang yang terserang (Hwang dan Ko 2014). Pada penelitian ini cendawan Fusarium spp. ditemukan pada bagian tanaman pisang yang sakit (bonggol, batang semu, akar, dan tanah rizosfer). Mikrokonidium Fusarium biasa berada di dalam jaringan pembuluh tanaman
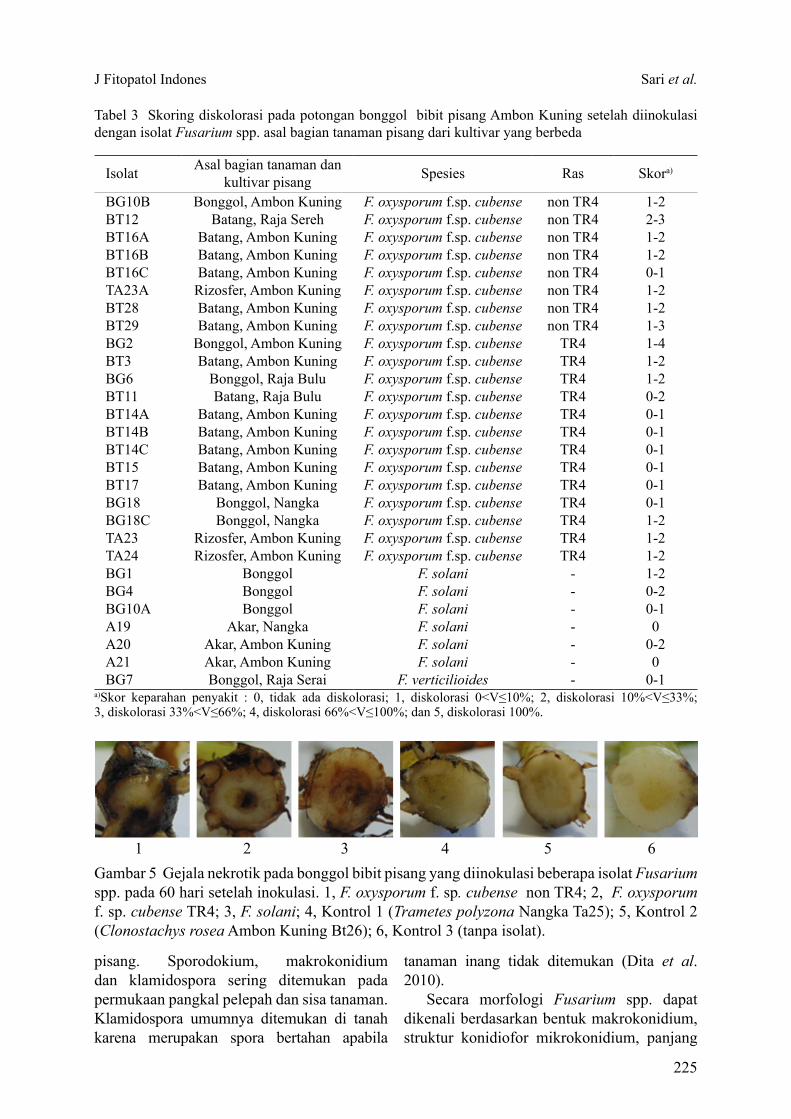
J Fitopatol Indones Sari et al.
225
pisang. Sporodokium, makrokonidium dan klamidospora sering ditemukan pada permukaan pangkal pelepah dan sisa tanaman. Klamidospora umumnya ditemukan di tanah karena merupakan spora bertahan apabila
tanaman inang tidak ditemukan (Dita et al. 2010).
Secara morfologi Fusarium spp. dapat dikenali berdasarkan bentuk makrokonidium, struktur konidiofor mikrokonidium, panjang
Tabel 3 Skoring diskolorasi pada potongan bonggol bibit pisang Ambon Kuning setelah diinokulasi dengan isolat Fusarium spp. asal bagian tanaman pisang dari kultivar yang berbeda
Isolat Asal bagian tanaman dan kultivar pisang Spesies Ras Skora)
BG10B Bonggol, Ambon Kuning F. oxysporum f.sp. cubense non TR4 1-2BT12 Batang, Raja Sereh F. oxysporum f.sp. cubense non TR4 2-3BT16A Batang, Ambon Kuning F. oxysporum f.sp. cubense non TR4 1-2BT16B Batang, Ambon Kuning F. oxysporum f.sp. cubense non TR4 1-2BT16C Batang, Ambon Kuning F. oxysporum f.sp. cubense non TR4 0-1TA23A Rizosfer, Ambon Kuning F. oxysporum f.sp. cubense non TR4 1-2BT28 Batang, Ambon Kuning F. oxysporum f.sp. cubense non TR4 1-2BT29 Batang, Ambon Kuning F. oxysporum f.sp. cubense non TR4 1-3BG2 Bonggol, Ambon Kuning F. oxysporum f.sp. cubense TR4 1-4BT3 Batang, Ambon Kuning F. oxysporum f.sp. cubense TR4 1-2BG6 Bonggol, Raja Bulu F. oxysporum f.sp. cubense TR4 1-2BT11 Batang, Raja Bulu F. oxysporum f.sp. cubense TR4 0-2BT14A Batang, Ambon Kuning F. oxysporum f.sp. cubense TR4 0-1BT14B Batang, Ambon Kuning F. oxysporum f.sp. cubense TR4 0-1BT14C Batang, Ambon Kuning F. oxysporum f.sp. cubense TR4 0-1BT15 Batang, Ambon Kuning F. oxysporum f.sp. cubense TR4 0-1BT17 Batang, Ambon Kuning F. oxysporum f.sp. cubense TR4 0-1BG18 Bonggol, Nangka F. oxysporum f.sp. cubense TR4 0-1BG18C Bonggol, Nangka F. oxysporum f.sp. cubense TR4 1-2TA23 Rizosfer, Ambon Kuning F. oxysporum f.sp. cubense TR4 1-2TA24 Rizosfer, Ambon Kuning F. oxysporum f.sp. cubense TR4 1-2BG1 Bonggol F. solani - 1-2BG4 Bonggol F. solani - 0-2BG10A Bonggol F. solani - 0-1A19 Akar, Nangka F. solani - 0A20 Akar, Ambon Kuning F. solani - 0-2A21 Akar, Ambon Kuning F. solani - 0BG7 Bonggol, Raja Serai F. verticilioides - 0-1
a)Skor keparahan penyakit : 0, tidak ada diskolorasi; 1, diskolorasi 0<V≤10%; 2, diskolorasi 10%<V≤33%; 3, diskolorasi 33%<V≤66%; 4, diskolorasi 66%<V≤100%; dan 5, diskolorasi 100%.
Gambar 5 Gejala nekrotik pada bonggol bibit pisang yang diinokulasi beberapa isolat Fusarium spp. pada 60 hari setelah inokulasi. 1, F. oxysporum f. sp. cubense non TR4; 2, F. oxysporum f. sp. cubense TR4; 3, F. solani; 4, Kontrol 1 (Trametes polyzona Nangka Ta25); 5, Kontrol 2 (Clonostachys rosea Ambon Kuning Bt26); 6, Kontrol 3 (tanpa isolat).
1 2 3 4 5 6

J Fitopatol Indones Sari et al.
226
fialid, pembentukan dan posisi klamidospora. Meskipun pengamatan morfologi tidak cukup untuk identifikasi lengkap, tapi banyak informasi yang dapat diperoleh saat biakan di medium kultur. Hasil pengamatan bentuk makrokonidium dan mikrokonidium menunjukkan adanya perbedaan spesifik antara F. oxysporum, Fusarium solani, F. verticillioides, T. polyzona dan C. rosea. T. polyzona merupakan cendawan tanah yang berasal dari rizosfer tanaman pisang kultivar Nangka dan C. rosea merupakan isolat yang berasal dari batang tanaman pisang kultivar Ambon Kuning. Kedua cendawan ini mempunyai karakter morfologi yang berbeda dengan Fusarium spp., dimana keduanya tidak memiliki makro dan mikrokonidia. T. polyzona mempunyai konidia dengan ukuran yang sangat kecil dibanding dengan mikrokonidia Fusarium spp. Ciri khas dari F. oxysporum mempunyai makrokonidium yang sedikit bengkok dengan ujung runcing, terdiri atas 3–4 sekat. Mikrokonidium yang berbentuk oval atau seperti ginjal, tanpa sekat dan terbentuk pada ujung fialid yang bertangkai pendek. Sedangkan F. solani mempunyai makrokonidium yang besar dan tumpul terdiri dari 3–4 sekat, dengan mikrokonidium yang berbentuk oval (dengan atau tanpa sekat) dan terbentuk pada ujung tangkai fialid yang panjang. F. oxysporum dan F. solani mempunyai klamidospora yang tersusun dalam satu rantai atau terdapat pada ujung dan tengah hifa. F. verticilliodes mempunyai makrokonidium yang lurus, panjang dan langsing, dengan sekat 3–5. Mikrokonidium berbentuk oval dan tanpa sekat. F. verticilliodes tidak mempunyai klamidospora.
Karakter morfologi laju pertumbuhan dan kerapatan konidium, warna dan tipe koloni yang didapatkan sangat bervariasi. Khususnya laju pertumbuhan dan kerapatan konidia tidak bisa dipakai sebagai acuan pembeda. Tetapi bentuk dan warna koloni memperlihatkan karakter yang berbeda diantara spesies. F. oxysporum umumnya mempunyai warna koloni putih, krem, dan ungu pucat, sedangkan F. solani didominasi oleh warna koloni putih. F. verticillioides berwarna merah muda dan
genus non fusarium mempunyai warna putih. Menurut Ploetz (1990) variasi warna pada F. oxysporum pada medium bisa disebabkan kondisi dan medium kultur yang digunakan.
Data karakter morfologi yang diperoleh belum bisa menunjukkan isolat F. oxysporum f. sp. cubense TR4 dan non TR4 karena secara umum bentuk makrokonidium dan mikrokonidium, warna koloni dan posisi klamidospora yang sama. Menurut O’Donnell et al. (2008) karakterisasi secara morfologi tidak bisa menggambarkan hubungan kekerabatan antarspesies bahkan antarras, juga tidak bisa digunakan untuk membedakan patogenisitas dari masing-masing spesies atau ras.
Identifikasi secara molekuler meng-gunakan primer spesifik dan universal primer dapat menyalin urutan basa DNA dari suatu organisme sehingga dapat digunakan untuk identifikasi sampai tingkat ras atau strain. Hasil karakterisasi secara molekuler terhadap 30 isolat Fusarium spp terlihat bahwa F. oxysporum f. sp. cubense TR 4 (13 isolat) paling banyak ditemukan, diikuti oleh F. oxysporum f. sp. cubense non TR4 (7 isolat ) dan F. solani (7 isolat) dan F. verticillioides. Hal ini menunjukkan bahwa sebaran F. oxysporum f. sp. cubense TR4 sudah meluas dan mendominasi di daerah pengambilan sampel. Hermanto et al. (2011) juga menemukan F. oxysporum f. sp. cubense TR4 sudah menyebar di hampir semua provinsi Indonesia dan menyerang hampir semua varietas pisang lokal yang bergenom AA (Rejang), AAA (Ambon Kuning, Ambon Hijau, Barangan), AAB (Ketan, Raja Bulu, Raja Sereh), dan ABB (Awak, Kepok).
Fusarium spp. mempunyai kisaran inang luas dan keragaman morfologi serta patogenisitas yang bervariasi. Sampai saat ini belum ada metode sederhana untuk membedakan antara strain patogenik dan nonpatogenik pada Fusarium spp (Alabouvette dan Couteaudier 1992). Hanya inokulasi pada tanaman inang yang dapat digunakan untuk menentukan sifat patogenik atau nonpatogenik Fusarium. Strain yang tidak dapat menyebabkan gejala pada tanaman dianggap sebagai nonpatogenik untuk tanaman tersebut.

J Fitopatol Indones Sari et al.
227
Secara alami Fusarium menyerang jaringan tanaman yang mengalami nekrosis sehingga tidak semua Fusarium yang ditemukan pada jaringan yang nekrosis merupakan patogen. Oleh karena itu untuk mengetahui patogen utama perlu dilakukan uji patogenisitas.
Semua isolat Fusarium yang diuji menimbulkan nekrosis dengan skala yang bervariasi pada bonggol bibit pisang. Patogenisitas dan virulensi cendawan F. oxysporum f. sp. cubense pada kerabat pisang berbeda-beda karena adanya perbedaan genotipe di dalam kerabat pisang, juga karena munculnya ras-ras baru F. oxysporum f. sp. cubense (Sahlan dan Nurhadi 1996). Terjadinya variasi virulensi di antara ras atau VCG F. oxysporum f. sp. cubense sudah banyak dilaporkan. Beberapa hal yang menyebabkan terjadinya variasi ini ialah adanya pengaruh interaksi inang dengan lingkungan yang bersifat kompleks serta adanya perbedaan karakter biologi, kimia, dan genetik yang dapat memengaruhi faktor-faktor penyebab virulensi seperti reproduksi spora dan produksi toksin (Groenewald 2005).
Hasil penelitian juga menemukan isolat F. solani yang mampu menimbulkan gejala nekrosis pada bonggol tetapi dengan nilai skala nekrosis yang rendah (skala: 1–2). F. solani mempunyai variasi genetik yang sangat tinggi sehingga mempunyai kisaran inang yang luas (Brasileiro et al. 2004). Hingga saat ini belum pernah dilaporkan F. solani yang diisolasi dari tanaman pisang bergejala layu dan dapat menimbulkan gejala nekrosis yang mirip dengan gejala yang ditimbulkan F. oxysporum f. sp. cubense pada bonggol bibit pisang. Penelitian ini merupakan laporan pertama yang menyatakan bahwa F. solani juga dapat menimbulkan gejala nekrosis pada bonggol bibit pisang. Namun, dikarenakan penelitian ini hanya dilakukan sampai 60 hsi (bibit pisang berumur 3.5 bulan) maka belum dapat dipastikan apakah isolat ini juga mampu menimbulkan gejala layu fusarium yang khas pada saat tanaman pisang memasuki fase generatif.
DAFTAR PUSTAKA
Aamir S, Sutar S, Singh SK, Baghela A. 2015. A rapid and efficient method of fungal genomic DNA extraction, suitable for PCR based molecular methods. Plant Pathol & Quarant. 5(2):74–81.
Alabouvette C, Counteaudier Y. 1992. Biological control of Fusarium wilt with nonpathogenic Fusarium. Di dalam: Tjamos EC, Cook RJ, Papavizas GC, editor. Biological Control of Plant Diseases. New York (US): Plenum Press. hlm 415–426..
Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ.1997. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25(17):3389–3402.
Brake VM, Pegg KG, Irwin JAG, Langdon PW. 1990. Vegetative compatibility groups within Australian populations of Fusarium oxysporum f. sp. cubense the cause of Fusarium wilt of bananas. Aust Agric Res. 41(5):863–870. DOI: http://dx.doi.org/ 10.1071/AR9900863.
Brasileiro BTRV, Coimbra MRM, de Morais Jr MAD, Oliveria NTD. 2004. Genetic variability within Fusarium solani species as revealed by PCR-fingerprinting based on PCR markers. Braz J Microbiol. 35(3):205–210. DOI: http://dx.doi.org/ 10.1590/S1517-83822004000200006.
Corderio M. 1994. Scala for Rating the Internal Corm Symtoms Caused by Fusarium Wilt. Honduras (US): INIBAB.
Dita MA, Waalwijk C, Buddenhaguen IW, Souza Jr MT, Kema GHJ. 2010. A molecular diagnosis for tropical race 4 of the banana Fusarium wilt pathogen. Plant Pathol. 59(2):348–357. DOI: http://dx.doi.org/ 10.1111/j.1365-3059.2009.02221.x.
Groenewald S. 2005. Biology, pathogenicity, and diversity of Fusarium oxysporum f. sp. cubense [tesis]. Pretoria (ZA): University of Pretoria.
Hermanto C, Sutanto A, Edison HS, Daniells JW, O’Neill WT, Sinohin VGO, Taylor

J Fitopatol Indones Sari et al.
228
P. 2011. Incidence and distribution of fusarium wilt disease of banana in Indonesia. Di dalam: The V International Symposium on Banana; 2009 September 14; ISHS-ProMusa Symposium on Global Perspectives on Asian Challenges 897. Guangzhou (CH). Hlm. 313–322.
Hwang SC, Ko WH. 2004. Cavendish banana cultivars resistant to Fusarium wilt acquired through somaclonal variation in taiwan. Plant Dis. 88(6):580–588. DOI: http://dx.doi.org/10.1094/PDIS.2004.88.6.580.
[KEMENTAN] Direktorat Jenderal Hortikultura 2015. Statistik produksi hortikultura 2014. https/hortikultura.p e r t a n i a n . g o . i d / w p - c o n t e n t /uploads/2016/02/Statistik-Produksi-2014.pdf [diakses 5 April 2017].
Kistler HC. 1997. Genetic diversity in the plant-pathogenic fungus Fusarium oxysporum. Phytopathol. 87:474–479. DOI: http://dx.doi.org/10.1094/PHYTO.1997.87.4.474.
Leslie JF, Summerell BA. 2006. The Fusarium Laboratory Manual. Ed ke-1. Oxford (GB): Blackwell. DOI: http://dx.doi.org/10.1002/9780470278376.
Lin YH, Chang JY, Liu ET, Huang JW and Chang PFL. 2009. Development of a molecular marker for specific detection of Fusarium oxysporum f. sp. cubense race 4. Eur J Plant Pathol. 123(3):353–365. DOI: http://dx.doi.org/10.1007/s10658-008-9372-4.
Martin JP. 1950. The use of acid, rose bengal and streptomycin in the plate method for estimating soil fungi. SOiZ Sci. 69: 215–232.
Moore NY, Pegg KG, Allen RN, Irwin JAG. 1993. Vegetative compatibility and distribution of Fusarium oxysporum f. sp. cubense in Australia. J Austr Expt Agric. 33(6):792–802. DOI: http://dx.doi.org/10.1071/ea9930797 .
O’Donnell K, Sutton DA, Fothergill A, Mc Carthy D, Rinaldi MG, Brandt ME, Zhang N, Geiser DM. 2008. Molecular phylogenetic diversity, multilocus haplotype nomenclature, and in vitro antifungal resistance within the Fusarium
solani species complex. J Clin Microbiol. 46(8):2477–2490. DOI: http://dx.doi.org/ 10.1128/JCM.02371-07.
Ploetz RC. 1990. Variability in Fusarium oxysporum f. sp. cubense. Can J Bot. 68(6):1357–1363. DOI: http://dx.doi.org/10.1139/b90-173.
Ploetz RC. 2006. Fusarium wilt of banana is caused by several pathogens referred to as Fusarium oxysporum f. sp. cubense. Phytopathol. 96(6):653–656. DOI: http://dx.doi.org/10.1094/PHYTO-96-0653.
[Pusdatin] Pusat Data dan Sistem Informasi Pertanian. 2014. Outlook Komoditi Pisang. http://epublikasi.setjen.pertanian.go.id/epublikasi/outlook/epublikasi/outlook/2014 [diakses10 Sep 2016].
Sahlan, Nurhadi, Hermanto C. 1996. Penyakit-penyakit utama tanaman pisang. Di dalam: Purnamo S, editor. Solok (ID): Balai Penelitian Tanaman Buah.
Stover RH, Buddenhagen IW. 1986. Banana breeding: polyploidy, disease resistance and productivity. Fruits 41(3):175–191.
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. 2013. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evolution. 30(12):2725–2729. DOI: http://dx.doi.org/10.1093/molbev/mst197 .
White TJ, Bruns T, Lee S, Taylor JW. 1990.Amplification and direct sequencing of fungal ribosomal rna genes for phylogenetics. Di dalam: Innes MA, Gelfand DH, Sninsky JJ, White TJ, editor. PCR Protocols: A Guide of Methods and Applications. New York (US): Academic Press. DOI: http://dx.doi.org/10.1016/B978-0-12-372180-8.50042-1.
Williams RC, Hermanto C, Suwanda, Komolong B, Kokoa P. 2010. Mitigating the threat of banana Fusarium wilt: understanding the agroecological distribution of pathogenic forms and developing disease management strategies. Final report. HORT/2005/136. Canberra (AU): ACIAR.

ISSN: 0215-7950
229
Volume 13, Nomor 6, November 2017Halaman 229–237
DOI: 10.14692/jfi.13.6.229
*Alamat Penulis Korespondensi: Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor. Jalan Kamper, Kampus IPB Dramaga, Bogor 16680. Tel: 0251629364, Faks: 0251629362, Surel: [email protected].
Kisaran Inang Cowpea mild mottle virus dan Respons Varietas Kedelai
Host Range of Cowpea mild mottle virus and Response of Soybean Varieties
Mimi Sutrawati, Sri Hendrastuti Hidayat*, Bonny Purnomo Wahyu Soekarno, Ali Nurmansyah, Gede Suastika
Institut Pertanian Bogor 16680
ABSTRAK
Cowpea mild mottle virus (CPMMV) merupakan salah satu virus yang menginfeksi kedelai dan menjadi penyakit endemik di Jawa dan Sumatera sejak pertama kali dilaporkan. Penelitian bertujuan menguji kisaran inang CPMMV dan mengevaluasi respons varietas kedelai baru terhadap isolat spesifik CPMMV. Pengujian kisaran inang CPMMV isolat CR16 dilakukan dengan cara menularkan virus secara mekanis pada11 spesies tanaman indikator, sedangkan tanaman kedelai yang dievaluasi terdiri atas 10 varietas. Respons ketahanan varietas kedelai ditentukan berdasarkan pada periode inkubasi, insidensi dan keparahan penyakit. Infeksi CPMMV pada tanaman uji dikonfirmasi dengan metode dot immunobinding assay (DIBA) menggunakan antiserum spesifik CPMMV. CPMMV isolat CR16 mampu menginfeksi 8 spesies tanaman Leguminoseae, dan Solanaceae secara sistemik; sedangkan pada Amaranthaceae hanya secara lokal. Semua tanaman indikator yang menunjukkan gejala sistemik dapat menjadi sumber inokulum CPMMV saat diinokulasi kembali pada tanaman kedelai. Kedelai varietas ‘Detam 1’, ‘Detam 2’, ‘Detam 3’, ‘Anjasmoro’, dan ‘Wilis’ tergolong rentan; sedangkan varietas ‘Detam 4’, ‘Malika’ dan ‘Dena 1’ tergolong moderat; serta varietas ‘Argomulyo’ dan ‘Grobogan’ tergolong tahan terhadap infeksi CPMMV isolat CR16.
Kata kunci : dot immunobinding assays, sistemik, sumber inokulum, tanaman indikator
ABSTRACT
Cowpea mild mottle virus (CPMMV) is one of important virus which infects soybean and become an endemic disease since the first time reported in Java and Sumatra. Research was conducted to study the host range of CPMMV and to evaluate response of new type soybean varieties to CPMMV specific isolate. Host range study of CPMMV CR16 isolate was conducted by sap transmission method to 11 species of indicator plants, whereas evaluation of soybean response involved 10 varieties. The resistance response of soybean variety was based on incubation period, and of either the disease incidence or severity. Virus infection on test plants were confirmed by dot immunobinding assay (DIBA) using specific CPMMV antisera. The CPMMV CR16 isolate was able to infect systemically 8 plant species belongs to Leguminoceae, and Solancaeae; while the virus caused local infection on Amaranthaceae. All plant species infected systemically proved to become sources of inoculum for CPMMV when they used in back inoculation to soybean. Response of soybean varieties to CPMMV CR16 isolate can be categorized into susceptible (Detam 1, Detam 2, Detam 3, Anjasmoro, Wilis), moderated (Detam 4, Malika, Dena 1) and resistant (Argomulyo and Grobogan).
Key words: dot immunobinding assays, indicator plant, source of inoculum, systemic

J Fitopatol Indones Sutrawati et al.
230
PENDAHULUAN
Cowpea mild mottle virus (CPMMV) yang tergolong dalam genus Carlavirus, famili Betaflexiviridae dapat ditularkan secara mekanis dan diduga bersifat tular benih. Berbeda dengan Carlavirus lainnya yang ditularkan oleh kutudaun, CPMMV ditularkan oleh Bemisia tabaci (Hemiptera: Aleyrodidae) secara nonpersisten (Jeyanandarajah dan Brunt (2008). Penyakit belang pada tanaman kedelai yang disebabkan oleh CPMMV pertama kali dilaporkan di Jawa dan Sumatera pada tahun 1984 (Iwaki et al. 1986). Gejala infeksi CPMMV ialah belang sistemik, pemucatan tulang daun, dan distorsi daun pada tanaman kedelai. Infeksi CPMMV menyebabkan penurunan bobot kering berangkasan tanaman kedelai hingga 15.5–53.4%, penurunan bobot biji kedelai 11.5–51.6%, dan penurunan kualitas biji 7.6–54.4% (Akin 2003).
Peneliti di berbagai negara melaporkan adanya perbedaan kisaran inang dan keragaman gejala infeksi CPMMV pada tanaman inang berbeda. Virus ini terdeteksi pada tanaman Solanaceae, yaitu Solanum melongena di Yordania dan Israel, tetapi isolat CPMMV tersebut tidak sama dengan isolat CPMMV yang menginfeksi Leguminosae (Mansour et al. 2008). Insidensi CPMMV pada tanaman kacang panjang berkisar 15-40% di Venezuela (Brito et al. (2012). Infeksi alami CPMMV pada kacang panjang dan kacang buncis juga dilaporkan di Taiwan (Chang et al. 2013). Hal tersebut menunjukkan infeksi CPMMV terus meluas pada berbagai tanaman inang di berbagai negara. Di Indonesia, selain pada kedelai infeksi CPMMV belum pernah dilaporkan.
Evaluasi ketahanan varietas kedelai yang dilakukan oleh Akin (2003) menunjukkan bahwa infeksi CPMMV dapat menurunkan bobot berangkasan tanaman dan bobot biji, serta meningkatkan jumlah biji tidak normal. Ia menyimpulkan kedelai varietas Taichung tergolong rentan dan galur B 3570 tergolong toleran terhadap infeksi CPMMV. Pengembangan varietas kedelai dengan
perbaikan genetika terus dilakukan untuk mendapatkan varietas unggul berdaya hasil tinggi, tetapi belum ada informasi respons ketahanan varietas terhadap infeksi CPMMV. Penelitian ini bertujuan menguji kisaran inang CPMMV dan respons ketahanan beberapa varietas kedelai terhadap infeksi CPMMV isolat CR16.
BAHAN DAN METODE
Pemurnian dan Propagasi Inokulum Isolat CR 16
Isolat CPMMV yang digunakan ialah isolat CR16 yang berasal dari daerah Ciranjang, Cianjur, Jawa Barat. Isolat tersebut dimurnikan terlebih dahulu sebelum digunakan sebagai sumber inokulum. Daun kedelai yang terinfeksi CPMMV isolat CR16 digerus dalam bufer fosfat 0.1 M pH 7.0 (1:5 b/v) sehingga diperoleh ekstrak tanaman sakit (sap). Daun primer tanaman kedelai ditaburi carborundum 600 mesh dan selanjutnya sap dioleskan pada daun tersebut. Tanaman yang telah diinokulasi dipelihara dalam kurungan kedap serangga. Tanaman yang menunjukkan gejala khas terinfeksi CPMMV berupa belang sistemik dan deformasi daun dideteksi dengan antiserum CPMMV dan CMV untuk memastikan infeksi tunggal CPMMV.
Pemurnian tahap ke-2 dilakukan menggunakan serangga vektor. Kutukebul sebanyak 10 ekor diletakkan pada tanaman kedelai sumber inokulum selama 24 jam untuk periode makan akuisisi. Satu hari kemudian, kutukebul dipindahkan ke tanaman kedelai sehat selama 48 jam untuk menularkan CPMMV. Tanaman dipelihara di dalam kurungan kedap serangga sampai muncul gejala. Konfirmasi infeksi CPMMV pada tanaman kedelai yang bergejala belang sistemik dan deformasi daun dilakukan pada 7 hari setelah inokulasi (HSI) dengan metode DAS-ELISA (Clark dan Adams (1977). Propagasi virus dilakukan pada tanaman kedelai varietas Wilis melalui penularan secara mekanis mengikuti metode yang diuraikan sebelumnya.

J Fitopatol Indones Sutrawati et al.
231
Pengujian Kisaran InangPengujian kisaran inang CPMMV
dilakukan pada 11 spesies tanaman dari famili Leguminosae (Vigna unguiculata subsp. sesquipedalis, V. radiata, Phaseolus vulgaris, Arachis hypogaea, dan Glycine max), Solanaceae (Lycopersicum esculentum, Solanum melongena, Nicotiana tabacum, dan N. benthamiana), Chenopodiaceae (Chenopodium amaranticolor), dan Amaranthaceae (Gomphrena globosa). Tanaman uji tersebut selanjutnya disebut tanaman indikator.
Percobaan disusun dalam rancangan acak kelompok dengan perlakuan 11 spesies tanaman indikator, masing-masing diulang sebanyak 10 tanaman. Inokulasi CPMMV dilakukan secara mekanis seperti yang telah diuraikan sebelumnya. Tanaman kontrol ialah tanaman yang diinokulasi hanya dengan bufer fosfat. Pengamatan meliputi tipe gejala dan masa inkubasi. Infeksi CPMMV dikonfirmasi secara serologi dengan metode Dot Immunobinding Assays (DIBA) pada 2 minggu setelah inokulasi (MSI).
Inokulasi balik CPMMV ke tanaman kedelai. Pengujian ini dilakukan untuk mengetahui potensi tanaman yang digunakan pada pengujian kisaran inang sebagai sumber inokulum CPMMV bagi tanaman kedelai. Tanaman indikator yang menunjukkan gejala sistemik dan dideteksi positif terinfeksi CPMMV digunakan sebagai sumber inokulum untuk penularan ke tanaman kedelai sehat menggunakan metode penularan mekanis seperti yang diuraikan sebelumnya. Gejala penyakit dan periode inkubasi diamati hingga 1 minggu setelah tanam (MST). Infeksi CPMMV dikonfirmasi secara serologi dengan metode DIBA.
Evaluasi Respons Varietas Kedelai terhadap Infeksi CPMMV
Varietas kedelai yang diuji terdiri atas 5 varietas kedelai kuning (Anjasmoro, Wilis, Argomulyo, Grobogan, Dena 1) dan 5 varietas kedelai hitam (Detam 1, Detam 2, Detam 3, Detam 4, Malika).
Penelitian disusun dalam rancangan acak kelompok dengan perlakuan 10 varietas kedelai, setiap perlakuan diulang 5 kali, masing-masing ulangan terdiri atas 4 tanaman. Benih kedelai ditanam pada medium campuran tanah, sekam, dan kompos dengan perbandingan 2:1:1 (b/b/b) di dalam pot plastik berukuran 30 cm × 30 cm dan dipelihara di rumah kasa.
Inokulasi CPMMV dilakukan secara mekanis pada daun tunggal tanaman kedelai berumur 7 hari setelah semai (HSS) mengikuti metode yang telah diuraikan sebelumnya. Peubah yang diamati meliputi tipe gejala, masa inkubasi, insidensi dan keparahan penyakit. Deteksi CPMMV dilakukan dengan metode DIBA pada 2 MSI. Insidensi dan keparahan penyakit dihitung dengan rumus:
IP = nN × 100%, dengan
IP, insidensi penyakit (%); n, jumlah tanaman terinfeksi CPMMV (berdasarkan hasil DIBA); N, total jumlah tanaman yang diamati.
N × VKP = × 100%, dengan(ni × vi)∑
i
i=0
KP, keparahan penyakit (%); n, jumlah sampel untuk kategori serangan tertentu; v, nilai skor untuk kategori serangan tertentu; Z, nilai skor tertinggi; N, jumlah sampel untuk skor tertinggi.
Penentuan skor gejala penyakit CPMMV pada kedelai mengikuti Akin (2003), yaitu 0, tidak ada gejala; 1, klorosis dan tulang daun memucat; 2, gejala mosaik dengan klorosis pada tulang daun dan permukaan daun; 3, mosaik berat, permukaan daun bergelombang, tepi daun melengkung ke atas; 4, malformasi daun (Gambar 1). Selanjutnya dibuat matriks skoring yang meliputi periode inkubasi (HSI), insidensi penyakit (%), dan keparahan penyakit (%) (Tabel 1).
Ketahanan varietas kedelai terhadap CPMMV ditentukan berdasarkan lama periode inkubasi (HSI), insidensi penyakit (%) dan keparahan penyakit (%) dikelompokkan sebagai berikut: tahan (total skor 3-4), moderat (total skor 6-9), dan rentan (total skor 10-12).

J Fitopatol Indones Sutrawati et al.
232
Deteksi CPMMVMetode DIBA untuk mendeteksi CPMMV
dilakukan mengikuti Mahmood et al. (1997). Daun sampel digerus dalam tris buffer saline (TBS: Tris-HCl 0.02 M dan NaCl 0.15M, pH 7.5) dengan perbandingan 1:10 (b/v). sebanyak 2 µL cairan perasan tanaman tersebut diteteskan ke membran nitroselulosa (Hybond
TM-P, Amersham Biosciences UK)
dan dikeringanginkan.
Selanjutnya membran direndam di dalam 10 mL larutan blocking non fat milk 2% dalam TBS yang mengandung Triton X-100 dengan konsentrasi akhir 2%. Membran kemudian diinkubasi pada suhu ruang sambil digoyang dengan kecepatan 50 rpm selama 2 jam menggunakan EYELA multi shaker MMS, dicuci sebanyak 5 kali dengan dH
2O, tiap
pencucian berlangsung 5 menit sambil digoyang dengan kecepatan 100 rpm. Selanjutnya
0 1 2
3 4
Gambar 1 Skor keparahan penyakit belang pada kedelai: 0, tidak bergejala; 1, tulang daun memucat dan daun melengkung ke bawah; 2, belang dengan klorosis ringan pada tulang daun; 3, belang dengan klorosis jelas pada tulang daun; dan 4, malformasi daun.
Tabel 1 Matriks skoring ketahanan varietas kedelai terhadap CPMMV berdasarkan periode inkubasi (HSI), insidensi penyakit (%), dan keparahan penyakit (%)
Parameter Kisaran nilai Skor
Periode inkubasi (HSI)
0-7 48-15 316-25 226-30 1
Insidensi penyakit (%)
0<25 125<50 250<75 375<100 4
Keparahan penyakit (%)
0<25 125<50 250<75 375<100 4

J Fitopatol Indones Sutrawati et al.
233
membran direndam dalam IgG-AP CPMMV (antiserum kedua CPMMV) (DSMZ, Jerman) dalam TBS yang mengandung 2% non fat milk dengan perbandingan 1:3000 (v/v), diinkubasi pada suhu 4 °C selama semalam. Membran kemudian dicuci sebanyak 5 kali dengan bufer TBS yang mengandung Tween 0.05% (TBST) dan dibilas dengan akuades. Selanjutnya, membran direndam selama 10-30 menit dalam 10 mL bufer substrat (Tris-HCl 0.1 M, NaCl 0.1 M, dan MgCl
2 5 mM) yang mengandung
45 µL nitro blue tetrazolium (NBT) dan 35 µL bromo chloro indolil phosphate (BCIP). Reaksi positif ditandai dengan perubahan warna putih menjadi ungu pada membran nitroselulosa yang telah ditetesi cairan perasan tanaman dan reaksi dihentikan dengan akuades.
HASIL
Kisaran Inang CPMMVInfeksi CPMMV pada tanaman indikator
dari famili Leguminoceae dan Solanaceae menunjukkan gejala belang sistemik, klorosis tulang daun, dan malformasi daun; sedangkan pada famili Chenopodiaceae dan Amaranthaceae menyebabkan bercak klorosis lokal pada daun yang diinokulasi (Gambar 2). Hasil deteksi dengan metode DIBA menunjukkan bahwa semua tanaman dengan gejala sistemik memberikan reaksi positif terhadap antiserum CPMMV, kecuali A. hypogaea. Demikian pula, semua tanaman yang menunjukkan gejala klorosis lokal menunjukkan reaksi positif terhadap antiserum CPMMV (Tabel 2).
Hasil inokulasi balik dari tanaman indikator yang menunjukkan gejala sistemik dan positif terinfeksi CPMMV berhasil menularkan CPMMV ke tanaman kedelai sehat (Tabel 3). Penularan CPMMV ke tanaman kedelai menghasilkan insidensi penyakit 10-80% dengan periode inkubasi 3-7 HSI. Dengan demikian, 8 spesies tanaman tersebut dapat menjadi inang CPMMV dan sekaligus sumber inokulum CPMMV untuk tanaman kedelai.
Ketahanan Varietas Kedelai terhadap CPMMV
Gejala infeksi CPMMV bervariasi bergantung pada varietas, antara lain belang ringan, belang parah, permukaan daun tidak rata dan mengerut, ukuran daun lebih kecil, serta perubahan bentuk daun (malformasi) (Tabel 4). Infeksi CPMMV terjadi pada semua varietas kedelai yang diuji walaupun dengan insidensi dan keparahan penyakit yang berbeda-beda. Insidensi penyakit paling rendah (27.27%) terjadi pada varietas Argomulyo, sedangkan paling tinggi (100%) ditunjukkan oleh varietas Detam 3 (Tabel 1). Periode inkubasi rata-rata yang ditunjukkan oleh varietas Detam 1, Detam 2’, Detam 3, Detam 4, Anjasmoro, Wilis ialah 6-7 hari setelah inokulasi (HSI). Periode inkubasi pada varietas Malika, Argomulyo, Grobogan, dan Dena 1 menunjukkan waktu yang lebih panjang, yaitu 7-28 HSI.
Pengelompokan respons ketahanan varietas kedelai terhadap CPMMV didasarkan pada periode inkubasi, insidensi penyakit, dan keparahan penyakit yang disusun dalam matriks skoring. Berdasarkan skor ini kedelai yang dikelompokkan rentan ialah Detam 1, Detam 2, Detam 3, Anjasmoro, dan Wilis; moderat ialah Detam 4, Malika dan Dena 1; dan tahan ialah varietas (Argomulyo dan Grobogan).
PEMBAHASAN
Gejala infeksi CPMMV pada kedelai di Indonesia berupa belang, pemucatan tulang daun, dan daun menggulung. CPMMV isolat CR16 yang digunakan dalam penelitian ini mampu menimbulkan gejala belang sistemik pada tanaman Leguminoseae dan Solanaceae. Hal serupa telah dilaporkan sebelumnya (Pardina et al. 2004), walaupun dalam penelitian tersebut isolat CPMMV yang menginfeksi Solanaceae berbeda dengan isolat yang menginfeksi Leguminoseae. Chang et al. (2013) melaporkan isolat CPMMV Taiwan dapat menginfeksi tanaman Leguminoseae,

J Fitopatol Indones Sutrawati et al.
234
a b c
fd e
g h i
j k l
Gambar 2 Gejala infeksi CPMMV isolat CR16 pada tanaman indikator. a, daun melepuh dan belang sistemik pada Vigna unguiculta subsp. sesquipedalis; b, pemucatan tulang daun dan distorsi daun pada Vigna radiata; c, pemucatan sistemik pada pucuk Arachis hypogaea; d, belang sistemik dan permukaan daun tidak rata pada Phaseolus vulgaris; e, pemucatan tulang daun pada Phaseolus vulgaris; f, belang sistemik dan melepuh pada Glycine max; g, klorosis lokal pada Chenopodium amaranticolor; h, klorosis lokal pada Gomphrena globosa; i, belang sistemik dan malformasi daun pada Nicotiana tabacum ; j, klorosis tulang daun dan belang sistemik pada N. benthamiana; k, belang sistemik dan malformasi daun pada Lycopersicum melongena; l, belang sistemik dan malformasi daun pada L. esculentum.

J Fitopatol Indones Sutrawati et al.
235
Tabel 3 Inokulasi balik CPMMV dari tanaman indikator ke tanaman kedelai
Tanaman indikator sebagai sumber inokulum
Gejala pada kedelai
Jumlah tanaman
bergejala*
Periode inkubasi (HSI)
Jumlah sampel terinfeksi
CPMMV **Vigna unguiculata belang sistemik 2 3 2Vigna radiata belang sistemik 1 6 1Phaseolus vulgaris belang sistemik 4 6 4Glycine max belang sistemik 8 7 8Nicotiana tabacum belang sistemik 4 7 4Solanum melongena belang sistemik 5 7 5Lycopersicum esculentum belang sistemik 2 7 2
*Jumlah tanaman uji untuk masing-masing tanaman indikator adalah 10 tanaman, kecuali N. benthamiana dan L.melongena 8 tanaman, L. esculentum7 tanaman; **Konfirmasi infeksi CPMMV dilakukan dengan metode DIBA.
Tabel 2 Gejala CPMMV isolat CR16 pada beberapa tanaman indikator
Tanaman indikator Gejala Jumlah tanaman bergejala (jumlah total tanaman)*
Periode Inkubasi
(HSI)
Jumlah sampel terinfeksi
CPMMV **LeguminoceaeVigna unguiculata Belang sistemik,
malformasi daun10(10) 33 9
Vigna radiata Klorosis tulang daun 1(10) 7 1Arachis hypogaea Klorosis tulang daun 1(10) 7 0Phaseolus vulgaris Belang, tulang daun
memutih4(10) 5 4
Glycine max Belang sistemik, daun melepuh
8(10) 7 8
AmaranthaceaeChenopodium amaranticolor Lesio local 1(10) 10 1Gomphrena globosa Lesio local 1(10) 7 1SolanaceaeNicotiana tabacum Belang sistemik,
malformasi daun7(10) 7 7
Nicotiana benthamiana Klorosis tulang daun 2(2) 14 2Lycopersicum melongena Belang sistemik,
malformasi daun7(10) 8 7
Lycopersicum esculentum Belang sistemik, malformasi daun
8(10) 8 8
*Jumlah tanaman bergejala (jumlah total tanaman), kecuali N. benthamiana 2 tanaman; **Konfirmasi infeksi CPMMV dilakukan dengan metode DIBA.
namun tidak menginfeksi Cucurbitaceae dan Solanaceae. Infeksi CPMMV isolat CR16 pada kacang panjang (V. unguiculata) menunjukkan gejala belang sistemik, pemucatan tulang daun, lamina daun berkerut, dan malformasi daun. Sedangkan Yadav (2013) melaporkan bahwa infeksi CPMMV pada kacang panjang di India menunjukkan gejala lesio klorosis yang berkembang menjadi bercak nekrosis dengan
belang lemah dan daun menggulung. Respons tanaman kacang buncis (P. vulgaris) terhadap CPMMV isolat CR16 menunjukkan hasil yang sama dengan penelitian Yadav (2013), yaitu gejala belang, klorosis pada tulang daun, dan daun keriting. Yadav (2013) melaporkan bahwa infeksi CPMMV pada N. tabacum tidak menunjukkan gejala, tetapi infeksi CPMMV isolat CR16 menyebabkan belang, pemucatan

J Fitopatol Indones Sutrawati et al.
236
kedelai. Oleh karena itu, penanaman kedelai disarankan tidak bersamaan dengan tanaman Leguminoseae dan Solanaceae karena berpotensi saling menularkan CPMMV.
Pengujian respons ketahanan sepuluh varietas kedelai menunjukkan sebagian besar varietas kedelai moderat dan rentan terhadap infeksi CPMMV isolat CR16. Dua varietas yang rentan terhadap CPMMV, ‘Anjasmoro’ dan ’Wilis’, merupakan yang paling banyak ditanam di Indonesia. Varietas ‘Anjasmoro’ paling banyak ditanam di Nusa Tenggara Barat, Nangro Aceh Darusssalam, dan Jawa Barat; sedangkan varietas ‘Wilis’ dan ‘Argomulyo’ mendominasi lahan kedelai di Jawa Timur (Krisdiana 2014). Varietas Grobogan merupakan kedelai yang tahan terhadap CPMMV dan paling banyak ditanam di Jawa Tengah. Penyakit belang CPMMV berpotensi menjadi penyakit yang endemik di sentra budi daya kedelai di Indonesia karena varietas yang paling banyak ditanam merupakan varietas rentan dan moderat terhadap CPMMV.
UCAPAN TERIMA KASIH
Penulis mengucapkan terimakasih atas bantuan dana Penelitian Disertasi Doktor kepada Direktorat Riset dan Pengabdian Masyarakat Direktorat Jenderal Penguatan Riset dan Pengembangan Kementerian Riset, Teknologi, dan Pendidikan Tinggi
tulang daun dan malformasi. Perbedaan kisaran inang dan respons tanaman indikator terhadap isolat CPMMV yang berbeda dapat disebabkan oleh perbedaan galur virus.
Uji inokulasi balik CPMMV dari tanaman indikator (Leguminoseae dan Solanaceae) ke tanaman kedelai menunjukkan potensi tanaman indikator sebagai sumber inokulum CPMMV bagi tanaman kedelai. Begitu pula sebaliknya, isolat CPMMV dari tanaman kedelai dapat menjadi inokulum bagi tanaman Leguminoseae dan Solanaceae lainnya. Dengan demikian insidensi penyakit belang akan cepat meningkat apabila tanaman dari kelompok Leguminoceae atau Solanaceae ditanam secara tumpang sari atau di lahan yang berdampingan dengan tanaman kedelai.
CPMMV isolat CR16 mempunyai kisaran inang yang berbeda dibandingkan dengan isolat CPMMV yang dilaporkan sebelumnya, yaitu isolat dari Taiwan dan India. Isolat CPMMV di Taiwan menginfeksi Leguminoceae, tetapi tidak menginfeksi Solanaceae (Chang et al. 2013). Pengujian kisaran inang CPMMV India menunjukkan bahwa CPMMV tidak menimbulkan gejala pada N. tabacum (Yadav et al. 2013). Namun, dalam penelitian ini terbukti isolat CPMMV CR16 dari tanaman kedelai mampu menginfeksi L. esculentum, S. melongena, dan N. tabacum. Tanaman inang CPMMV CR16 cukup luas dan berpotensi menjadi sumber infeksi untuk tanaman
Tabel 4 Pengelompokkan respons varietas kedelai terhadap CPMMV isolat CR16
VarietasPeriode inkubasi(HSI)
Gejala awal*Insidensi penyakit
(%)
Keparahan penyakit
(%)Total nilai Respons
‘Detam1’ 6-7 BR 75.00 57.5 11 Rentan‘Detam2’ 6-7 BR 72.73 55.0 10 Rentan‘Detam3’ 6-7 BR 100.00 57.5 11 Rentan‘Detam4’ 6-7 BR 55.56 27.5 9 Moderat‘Malika’ 6-28 BP, DM, DD 50.00 52.5 7 Moderat‘Anjasmoro’ 6-7 BP, DM, DD 57.14 75.0 11 Rentan‘Wilis’ 6-7 BP 70.00 70.0 10 Rentan‘Argomulyo’ 6-28 BR 27.27 22.5 4 Tahan‘Grobogan’ 7-28 BR 33.33 12.5 4 Tahan‘Dena1’ 7-22 BR 62.50 37.5 7 Moderat
*BR, belang ringan; BP, belang parah; DM, daun mengerut; DD, distorsi daun.

J Fitopatol Indones Sutrawati et al.
237
sesuai dengan Surat Perjanjian Penugasan Pelaksanaan Program Penelitian Batch I, nomor: 044/SP2H/LT/DRPM/II/2016, tanggal 17 Februari 2016.
DAFTAR PUSTAKA
Akin HM. 2003. Respon beberapa genotipe kedelai terhadap infeksi cowpea mild mottle virus. J HPT Trop. 3(2):40–43.
Brito M, Fernandez-Rodriguez T, Garrido MJ, Mejias A, Romano M, Marys E. 2012. First report of Cowpea mild mottle Carlavirus on yardlong bean (Vigna unguiculata subsp. sesquipedalis) in Venezuela. Viruses. 4(12):3804–11. DOI: http://dx.doi.org/10.3390/v4123804.
Chang C, Chien L, Tsai C, Lin Y, Cheng Y. 2013. First report of Cowpea mild mottle virus in cowpea and French bean in Taiwan. Plant Dis. 97(7):1001. DOI://doi. org/10.1094/PDIS-10-12-0935-PND.
Clark MF, Adams A. 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J Gen Virol. 34(3):475–483.
Iwaki M, Thongmeearkom P, Honda Y, Prommin M, Deema N, Hibi T, Iizuka N, Ong C, Saleh N. 1986. Cowpea mild mottle virus occurring on soybean and peanut in southeast Asian countries. Trop Agric Res Centre Tech. Bull. 21:106–120.
Jeyanandarajah P, Brunt AA. 1993. The natural occurrence, transmission, properties and possible affinities of
cowpea mild mottle virus. J Phytopathol. 137(2):148–156. DOI: http://dx.doi. org/10.1111/j.1439-0434.1993.tb01334.x.
Krisdiana R. 2014. Penyebaran varietas unggul kedelai dan dampaknya terhadap ekonomi perdesaan. J Pen Pert Tan Pangan. 33(1):61–69.
Mahmood T, Hein GL, French RC. 1997. Development of serological procedures for rapid and reliable detection of Wheat streak mosaic virus in a single wheat curl mite. Plant Dis. 81(3):250–253. DOI: https:// doi.org/10.1094/PDIS.1997.81.3.250.
Mansour A, Al-Musa A, Vetten H, Lesemann DE. 1998. Properties of a Cowpea mild mottle virus (CPMMV) isolate from eggplant in Jordan and evidence for biological and serological differences between CPMMV isolates from leguminons and solanaceous hosts. J Phytopathol. 146(11-12):539–547. DOI: http://dx.doi.org/10.1111/1439-0434. ep6401455.
Pardina PER, Arneodo J, Truol G, Herrera P, Laguna IG. 2004. First report of Cowpea mild mottle virus in bean crops in Argentina. Australas. Plant Pathol. 33(1):129–130. DOI: http://dx.doi.org/10.1590/S010041582006000300015.
Yadav MK, Biswas KK, Lal SK, Baranwal VK, Jain RK. 2013. A distinct strain of cowpea mild mottle virus infecting soybean in India. J Phytopathol. 161(10):739–744. DOI: http://dx.doi.org/10.1111/jph.12119.




















