
Biologi populasi Rajungan
19
Oseanologi dan Limnologi di Indonesia (2010) 36(3): 443-461 ISSN 0125-9830 443 BIOLOGI POPULASI KEPITING BAKAU (Scylla serrata F.) DI HABITAT MANGROVE TAMAN NASIONAL KUTAI KABUPATEN KUTAI TIMUR oleh NIRMALASARI IDHA WIJAYA 1) , FREDINAN YULIANDA 2) , MENNOFATRIA BOER 2) dan SRI JUWANA 3) 1) Sekolah Tinggi Ilmu Pertanian Kutai Timur 2) Departemen Manajemen Sumberdaya Perikanan Institut Pertanian Bogor 3) Pusat Penelitian Oseanografi – LIPI, Jakarta Received 27 July 2010, Accepted 16 November 2010 ABSTRAK Taman Nasional Kutai memiliki 5 277,79 ha hutan bakau di sepanjang pesisir pantainya. Kepiting bakau merupakan salah satu sumberdaya yang terdapat dalam ekosistem bakau yang dapat dimanfaatkan. Penelitian biologi populasi Scylla serrata di hutan bakau Taman Nasional Kutai (TNK) bertujuan untuk mengetahui parameter pertumbuhan, distribusi ukuran, laju mortalitas dan laju eksploitasi penangkapan. Hasil pengukuran diharapkan dapat digunakan sebagai bahan pertimbangan untuk pengelolaan kepiting bakau di TNK, agar dapat dimanfaatkan secara optimal dan berkelanjutan. Pengumpulan data biologi S. serrata diperoleh dari pengamatan yang dilakukan selama 4 bulan di musim hujan dan 4 bulan di musim kemarau (yaitu November 2008 – Juni 2009) di tiga stasiun yang telah ditetapkan berdasarkan karakteristik habitat bakaunya, yaitu di Muara Sangatta, Teluk Perancis dan Muara Sangkima. Data yang terkumpul dianalisis dengan metode analitik menggunakan instrument FISAT-II (FAO-ICLARM Stock Assesment Tool II). Hasil penelitian menunjukkan bahwa pola pertumbuhan S. serrata jantan di habitat bakau TNK bersifat allometrik positif, sedangkan S. serrata betina bersifat allometrik negatif. Koefisien pertumbuhan (K) berkisar antara 0,45- 1,50. K di Muara Sangatta lebih tinggi (1,2-1,5) dibanding di lokasi Teluk Perancis (0,8-1,1) dan Muara Sangkima (0,45-0,69). Laju penangkapan S. serrata di TNK sudah berada di atas laju eksploitasi maksimal, hanya di Muara Sangkima masih dibawah laju eksploitasi yang diperbolehkan. Distribusi lebar karapas kepiting bakau (S. serrata) di zona hutan bakau mempunyai frekuensi tertinggi pada interval 109,5- 129,5 mm. Secara umum ukuran ini relatif lebih besar dibandingkan ukuran lebar karapas kepiting di zona depan hutan bakau dengan frekuensi tertinggi pada interval 78-89 mm dan di zona perairan pantai dengan frekuensi tertinggi pada interval 89,5- 121,5 mm. Kelimpahan individu betina matang gonad mencapai puncak pada bulan
-
Upload
arisandy08 -
Category
Education
-
view
84 -
download
1
Transcript of Biologi populasi Rajungan

Oseanologi dan Limnologi di Indonesia (2010) 36(3): 443-461 ISSN 0125-9830
443
BIOLOGI POPULASI KEPITING BAKAU (Scylla serrata F.)DI HABITAT MANGROVE TAMAN NASIONAL KUTAI
KABUPATEN KUTAI TIMUR
oleh
NIRMALASARI IDHA WIJAYA1), FREDINAN YULIANDA2),
MENNOFATRIA BOER2) dan SRI JUWANA3)
1)Sekolah Tinggi Ilmu Pertanian Kutai Timur2)Departemen Manajemen Sumberdaya Perikanan
Institut Pertanian Bogor3)Pusat Penelitian Oseanografi – LIPI, Jakarta
Received 27 July 2010, Accepted 16 November 2010
ABSTRAK
Taman Nasional Kutai memiliki 5 277,79 ha hutan bakau di sepanjang pesisir pantainya. Kepiting bakau merupakan salah satu sumberdaya yang terdapat dalam ekosistem bakau yang dapat dimanfaatkan. Penelitian biologi populasi Scylla serrata di hutan bakau Taman Nasional Kutai (TNK) bertujuan untuk mengetahui parameter pertumbuhan, distribusi ukuran, laju mortalitas dan laju eksploitasi penangkapan. Hasil pengukuran diharapkan dapat digunakan sebagai bahan pertimbangan untuk pengelolaan kepiting bakau di TNK, agar dapat dimanfaatkan secara optimal dan berkelanjutan. Pengumpulan data biologi S. serrata diperoleh dari pengamatan yang dilakukan selama 4 bulan di musim hujan dan 4 bulan di musim kemarau (yaitu November 2008 – Juni 2009) di tiga stasiun yang telah ditetapkan berdasarkan karakteristik habitat bakaunya, yaitu di Muara Sangatta, Teluk Perancis dan Muara Sangkima. Data yang terkumpul dianalisis dengan metode analitik menggunakan instrument FISAT-II (FAO-ICLARM Stock Assesment Tool II). Hasil penelitian menunjukkan bahwa pola pertumbuhan S. serrata jantan di habitat bakau TNK bersifat allometrik positif, sedangkan S. serratabetina bersifat allometrik negatif. Koefisien pertumbuhan (K) berkisar antara 0,45-1,50. K di Muara Sangatta lebih tinggi (1,2-1,5) dibanding di lokasi Teluk Perancis (0,8-1,1) dan Muara Sangkima (0,45-0,69). Laju penangkapan S. serrata di TNK sudah berada di atas laju eksploitasi maksimal, hanya di Muara Sangkima masih dibawah laju eksploitasi yang diperbolehkan. Distribusi lebar karapas kepiting bakau (S. serrata) di zona hutan bakau mempunyai frekuensi tertinggi pada interval 109,5-129,5 mm. Secara umum ukuran ini relatif lebih besar dibandingkan ukuran lebar karapas kepiting di zona depan hutan bakau dengan frekuensi tertinggi pada interval 78-89 mm dan di zona perairan pantai dengan frekuensi tertinggi pada interval 89,5-121,5 mm. Kelimpahan individu betina matang gonad mencapai puncak pada bulan

WIJAYA, YULIANDA, BOER & JUWANA
444
Januari, Februari dan Maret. Diduga terjadi puncak kelimpahan yang kedua pada bulan Agustus dan September.
Kata kunci: Kepiting bakau, Scylla serrata, Taman Nasional Kutai, sebaran ukuran, parameter pertumbuhan, laju penangkapan.
ABSTRACT
BIOLOGY POPULATION OF MUD CRAB (Scylla serrata) AT MANGROVE HABITAT OF KUTAI NATIONAL PARK, EAST KUTAI REGENCY. Kutai National Park possess 5 277.79 ha mangrove forest along its coastal area. Mangrove crabs is one of its natural resources that can be exploited. Biology population research on mangrove forests of Kutai National Park (KNP) aims to identify parameters of growth, size distribution, rate of mortality and rate of exploitation of Scylla serrata. The results could be used as consideration on optimal and sustainable management of mangrove crabs in KNP. Collection of biological data of the mangrove crab was done 4 months during the rainy season and 4 months during the dry season (that is November 2008 to June 2009), at three station that can be selected according to characteristic of its mangrove habitat, that is at Muara Sangatta, Teluk Perancis and Muara Sangkima. Data collections were analyzed with analytical methods using FISAT-II (FAO-ICLARM Stock Assessment Toool II) instruments. The results showed that the growth pattern of the males crab at mangrove forest of KNP was allometric positive, wereas the females were allometric negative. Growth coefficient index (K) of the mangrove crab ranged from 0.45 to 1.50. K at Muara Sangatta higher (1.2 to 1.5) than at location Teluk Perancis (0.8 to 1.1) and Muara Sangkima (0.45 to 0.69). The fishing rate of the mangrove crab in KNP was already on the maximum exploitation rate, only at Muara Sangkima fishing rate exploitation rate is slightly below the maximum exploitation. The distribution of carapace width of S. serrata in the mangrove forest zones has the highest frequency at interval 109.5 to 129.5 mm. In general, that size relatively bigger than those in the middle zone of mangrove forests that was in the range of 78 to 79 mm, those in the inshore zone in the range of 89.5 to 121.5 mm. Peak of the abundance of the mature females was during January, February and March. The second peak was assumed in August to September.
Key words: Mangrove crabs, Scylla serrata, Kutai National Park , size distribution, growth parameters, fishing rate.

BIOLOGI POPULASI KEPITING BAKAU
445
PENDAHULUAN
Kepiting bakau telah menjadi komoditas perikanan penting di Indonesia sejak awal tahun 1980-an. Perikanan kepiting bakau di Indonesia diperoleh dari penangkapan stok alam di perairan pesisir, khususnya di area mangrove atau estuaria dan dari hasil budidaya di tambak air payau. Akhir-akhir ini, dengan semakin meningkatnya nilai ekonomi perikanan kepiting, penangkapan kepiting bakau juga semakin meningkat. Namun bersamaan dengan itu, rata-rata pertumbuhan produksi kepiting bakau di beberapa provinsi penghasil utama kepiting bakau justru agak lambat dan cenderung menurun (CHOLIK 1999).
Kepiting bakau yang bernilai sebagai sumber makanan dan pendapatan di Kosrae, Negara Bagian Micronesia, juga mengalami deplesi kelimpahan dan ukuran, akibat tekanan penangkapan yang dipengaruhi oleh distribusi penduduk dan lokasi usaha perikanan komersial (BONINE et al. 2008). Penurunan populasi kepiting bakau di alam diduga disebabkan oleh degradasi ekosistem mangrove dan kelebihan tangkap (over exploitation)(SIAHAINENIA 2008).
Kawasan konservasi Taman Nasional Kutai (TNK) memiliki hutan bakau pada lokasi 1-2 km dari tepi pantai ke arah daratan yang didominasi oleh jenis Rhizophora dan Bruguiera (TAMAN NASIONAL KUTAI 2005). Hutan bakau di kawasan TNK merupakan salah satu habitat kepiting di Kabupaten Kutai Timur. Di dalam kawasan mangrove tersebut saat ini telah berdiri empat desa definitif. Masyarakat yang tinggal dalam kawasan tersebut memanfaatkan sumberdaya yang terdapat dalam ekosistem mangrove untuk memenuhi kebutuhan hidupnya, baik sumberdaya hutan (vegetasi mangrove) maupun sumberdaya perikanannya, termasuk di antaranya kepiting bakau (Scylla serrata). Pemanfaatan sumberdaya di kawasan taman nasional perlu dikelola dan dibatasi penggunaannya agar tidak sampai merusak sumberdaya dan menghilangkan fungsi taman nasional sebagai kawasan konservasi.
Populasi kepiting bakau secara khas berasosiasi dengan hutan bakau yang masih baik, sehingga hilangnya habitat akan memberikan dampak yang serius pada populasi kepiting. Status bioekologi kepiting bakau yang berhubungan dengan biologi populasi dan pengelolaannya perlu dipahami untuk mendukung pengembangan dari perikanan tangkap dan budidaya kepiting bakau yang berkelanjutan (LE VAY 2001). Pertumbuhan, mortalitas, rekruitmen dan laju eksploitasi kepiting bakau dapat digunakan untuk menentukan tingkat penangkapan optimum (MSY) yang merupakan landasan dalam kebijakan pengelolaan penangkapan kepiting bakau (SPARRE & VENEMA 1999).
Tujuan penelitian ini untuk memperoleh parameter pertumbuhan, distribusi ukuran, laju mortalitas dan laju eksploitasi penangkapan Scylla serrata di habitat mangrove TNK. Hasil kajian diharapkan dapat digunakan sebagai suatu bahan pertimbangan dalam pengelolaan Scylla serrata di TNK agar dapat dimanfaatkan secara optimal dan berkelanjutan.

WIJAYA, YULIANDA, BOER & JUWANA
446
METODE PENELITIAN
Penetapan Stasiun PenelitianPenelitian dilakukan antara bulan November 2008 - Juni 2009 di kawasan
mangrove Taman Nasional Kutai (TNK) seluas 5 277, 79 ha. Habitat mangrove TNK dibagi menjadi 3 lokasi pengamatan, berdasarkan karakteristik habitat mangrovenya, yaitu Muara Sungai Sangatta (Stasiun A), Teluk Perancis (Stasiun B) dan Muara Sungai Sangkima (Stasiun C). Kelompok substasiun 1 zona tengah hutan mangrove, 2 zona depan hutan mangrove dan 3 zona perairan laut. (Gambar 1).
Mangrove di Muara Sangatta merupakan hutan bekas tebangan yang telah kehilangan pohon berdiameter besar dan sebagian dari areal ini telah dijadikan tambak. Di dalam plot tidak dijumpai pohon-pohon berdiameter besar, sebagian besar merupakan trubusan dengan diameter < 10 cm. Minimum ada 4 jenis pohon yang dijumpai di Muara Sangatta. Jenis yang dominan adalah Aegiceras corniculatum (gedangan) kemudian Nypa fructicans (nipah) Ceriops tagal (soga tinggi) dan Ceriops decandra (bido-bido) tidak dominan. Muara Sangatta dicirikan oleh kerapatan vegetasi, tekstur substrat, BOD dan kelimpahan S. serrata yang tinggi.
Mangrove di Teluk Perancis relatif masih utuh dibandingkan dengan Muara Sangatta. Pohon dengan diameter besar masih banyak ditemukan. Penebangan pohon ditemukan pada beberapa lokasi yang dijadikan tambak. Sebagian besar tambak tidak produktif lagi dan menjadi lahan kritis. Ada 3 jenis pohon bakau yang dijumpai di Teluk Perancis. Jenis pohon yang dominan adalah Rhizophora apiculata(bakau minyak). Rhizophora mucronata (bakau hitam) dan Bruguiera gymnorrhiza(bakau daun besar) kurang domiman. Kelompok substasiun B1 zona tengah hutan dan B3 zona perairan Teluk Perancis, dicirikan oleh parameter kelimpahan makrozoobenthos dan salinitas air yang tinggi.
Mangrove di Muara Sangkima mempunyai 6 jenis pohon, yang didominasi oleh Rhizophora apiculata (bakau minyak), kemudian Bruguiera gymnorrhiza (bakau daun besar) dan Ceriops decandra (bido-bido). Tiga jenis pohon waru tidak dominan, yaitu Osbornia octodonta, Hibiscus tiliaceus dan Lumnitzera littorea.Kelompok substasiun C1 zona tengah hutan mangrove Muara Sangkima, juga dicirikan dengan adanya kerapatan vegetasi, tekstur substrat, BOD dan kelimpahan S. serrata yang tinggi.

BIOLOGI POPULASI KEPITING BAKAU
447
Gambar 1. Peta lokasi dan stasiun penelitian di ekosistem mangrove Taman Nasional Kutai, 2008-2009.
Figure 1. Map of the studi area and research station in mangrove ecosystemKutai National Park, 2008-2009.
Pengumpulan Data BiologiJenis kepiting Scylla yang digunakan dalam penelitian ini diidentifikasi
berdasarkan ciri-ciri yang dibuat oleh Keenan et al. (1998), yaitu chela dan kaki-kakinya memiliki pola poligon yang sempurna untuk kedua jenis kelamin dan pada abdomen betina. Warna bervariasi dari unggu hijau sampai hitam kecoklatan. Duri pada karapas diantara dua mata tinggi, rata dan agak tumpul dengan tepian yang cenderung cekung dan membulat. Duri pada bagian luar cheliped berupa dua duri tajam pada propodus dan sepasang duri tajam pada carpus (Gambar 2).
Biologi Scylla serrata dikaji dengan melihat parameter pertumbuhan, pola distribusi, laju mortalitas dan laju eksploitasi. Pengumpulan data biologi Scylla serrata diperoleh dari observasi terhadap kepiting bakau yang dilakukan selama 8 bulan (Nopember 2008 - Juni 2009), pada lokasi 3 stasiun pengamatan yang diplih sesuai karakteristik habitat mangrove seperti tersebut di atas. Data primer dan sekunder (15 responden) yang dikumpulkan berupa musim, lokasi tangkapan, sebaran ukuran, jenis kelamin, lama upaya menangkap dan hasil tangkapan harian yang ditangkap dengan alat tangkap kait, rakkang maupun rengge.

WIJAYA, YULIANDA, BOER & JUWANA
448
Duri diantara dua mata Pola poligonal pada kaki-kaki renang
Duri pada propodus (lengan bawah) dan carpus (siku)
Pola poligonal pada abdomen
Karapas
Gambar 2: Ciri-ciri khas kepiting bakau, Scylla serrata.Figure 2: Characteristic features of mangrove crab, Scylla serrata.
Analisis DataBerdasarkan data tersebut dilakukan beberapa analisis yaitu: hubungan
panjang-bobot, yang digambarkan dalam dua bentuk yaitu isometrik dan allometrik berdasarkan HILE yang diacu oleh EFFENDIE (1979). Pendugaan kelompok ukuran dilakukan dengan menganalisis data frekuensi lebar karapas yang dianalisis menggunakan program Bhattacharya’s Method yang dikemas dalam paket program FISAT II (FAO-ICLARM Stock Assesment Tool). Analisis pertumbuhan, Plot Ford-Walford untuk menduga parameter pertumbuhan L∞ dan K dari persamaan von Bertalanfly. Pendugaan laju eksploitasi S. serrata dilakukan dengan pendugaan laju mortalitas alami (Z) berdasarkan persamaan Beverton dan Holt. Nilai Z dan pendugaan laju mortalitas alami (M) digunakan untuk menduga kematian kepiting bakau akibat penangkapan (F) = Z-M. Selanjutnya laju eksploitasi kepiting bakau

BIOLOGI POPULASI KEPITING BAKAU
449
(E) dapat diduga dengan menggunakan persamaan: E = F : Z, nilai Yield per Rekrut (Y/R) untuk menentukan laju eksploitasi maksimal yang diijinkan.
HASIL DAN PEMBAHASAN
Sebaran Spasial Ukuran Kepiting Bakau (Scylla serrata)Kepiting bakau (Scylla serrata) tidak memiliki bagian tubuh keras yang
permanen sebagai indikator pelacak umur, sehingga metode interpretasi ukuran tubuh yang digunakan adalah lebar karapas. Hasil tangkapan S. serrata di kawasan mangrove TNK (Stasiun ABC) memiliki struktur ukuran lebar karapas yang bervariasi berdasarkan lokasi penangkapannya. Lokasi penangkapan dalam penelitian ini meliputi tiga kondisi habitat yang berbeda, yaitu kawasan bagian tengah hutan mangrove (substasiun 1), kawasan pinggiran hutan mangrove (garis pantai) (substasiun 2), dan kawasan perairan pantai (inshore) (substasiun 3). Alat tangkap yang digunakan pada setiap lokasi berbeda-beda, tergantung pada spesifikasi dan kemampuan alat tangkap. Pada bagian tengah hutan mangrove digunakan pengait, pada bagian pinggir pantai digunakan rakkang, dan di perairan pantai digunakan alat tangkap rengge dengan ukuran diameter jaring 2 inchi. Jumlah individu S. serrata hasil tangkapan ketiga jenis alat tangkap pada tiga lokasi selama 8 bulan disajikan pada Tabel 1.
Tabel 1. Jumlah individu S. serrata hasil tangkapan alat pengait, rakkang dan rengge, November 2008 – Juni 2009.
Table 1. The number of S. serrata captured by the fishing gear hook, trap and gillnet, November 2008 to June 2009.
Total Individu Size Max-Min (mm)Fishing gear
Male FemaleSum of Total
(ind.) Male FemaleHook (pengait) 669 311 980 68-154 65-171Trap (rakkang) 526 449 975 50-143 45-155Gillnet (rengge) 59 141 200 70-142 73-135
Tabel 1 menunjukkan bahwa ukuran lebar karapas terkecil kepiting yang tertangkap adalah 45 mm dan ukuran lebar karapas terbesar adalah 171 mm. Ukuran kepiting yang terkecil menunjukkan bahwa kepiting yang mulai dapat tertangkap oleh alat tangkap adalah kepiting yang berukuran 45 mm. Grafik histogram sebaran ukuran dan sebaran jenis kelamin S. serrata berdasarkan hasil tangkapan dari bulan November 2008 - Juni 2009 dengan menggunakan alat tangkap rakkang, rengge, dan pengait dapat dilihat pada Gambar 3.

WIJAYA, YULIANDA, BOER & JUWANA
450
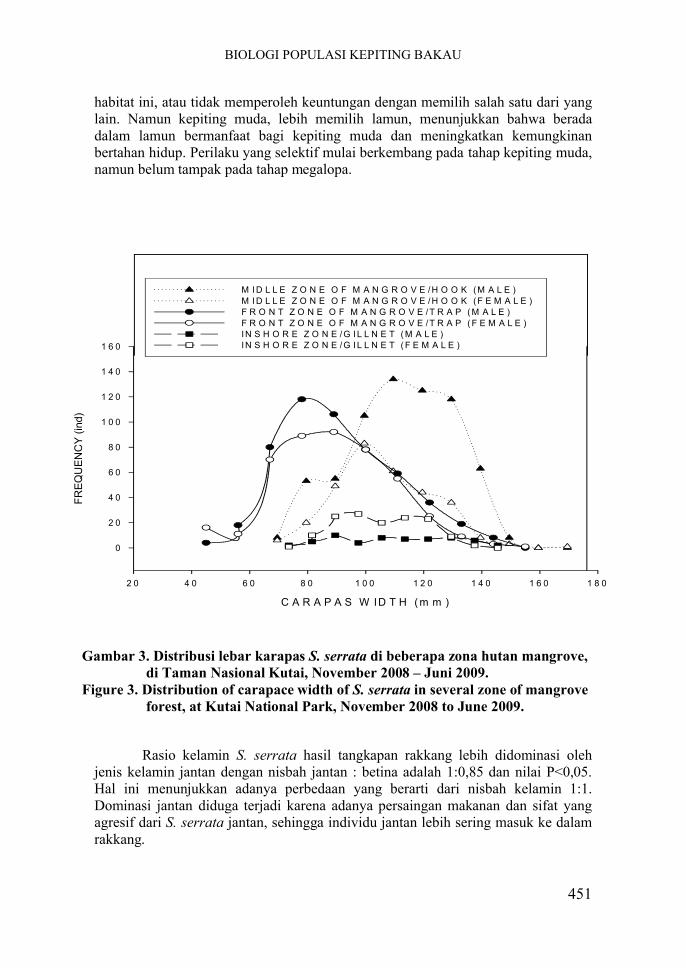
Lebar karapas S. serrata di tengah hutan mangrove, yang diperoleh dari hasil tangkapan alat pengait, umumnya berukuran lebih dari 100 mm, dengan frekuensi tangkapan tertinggi pada interval 109,5-129,5 mm. Persentase jumlah kepiting yang berukuran lebar karapas kurang dari 100 mm adalah 16,12%, artinya hanya 16% kepiting yang ditangkap dengan alat pengait yang belum dewasa kelamin, sedangkan sisanya, sekitar 83,88% merupakan kepiting yang diduga sudah dewasa kelamin. Hal ini menunjukkan bahwa di kawasan tengah hutan mangrove cenderung lebih banyak terdapat kepiting yang berukuran besar.
Rasio kelamin S. serrata di zona tengah hutan mangrove lebih didominasi jenis kelamin jantan dengan nisbah jantan:betina adalah 1:0,47 dan nilai P<0,05 menunjukkan adanya perbedaan yang berarti dari pergeseran nisbah kelamin 1:1. Dominasi jantan dapat terjadi karena adanya pola migrasi pada kepiting bakau. Kepiting S. serrata melangsungkan perkawinan di perairan mangrove dan secara berangsur-angsur sesuai dengan perkembangan telurnya, kepiting betina akan beruaya (berenang) ke laut dan memijah, sedangkan kepiting jantan tetap di perairan hutan bakau atau muara sungai (HILL 1975). Hasil ini bersesuaian dengan hasil penelitian LE VAY et al. (2007) yang menemukan bahwa hasil tangkapan kembali (recaptured) kepiting bakau S. paramamosain yang telah ditandai (marking) adalah 79% tertangkap pada malam hari di dataran lumpur pinggiran mangrove menuju ke laut, 14% yang tertangkap pada siang hari di dalam mangrove dengan pancingan dan 7% tertangkap gillnets ditetapkan setidaknya pada jarak 1 km lepas pantai dari pinggiran bakau.
Pada zona depan hutan mangrove diperoleh struktur ukuran lebar karapas Scylla serrata pada kelas ukuran kurang dari 100 mm mencapai 77,95%, dengan frekuensi tangkapan tertinggi pada interval 78-89 mm. Sisanya sebesar 22,05% merupakan kepiting berukuran lebih dari 100 mm. Hal ini menunjukkan bahwa lebih banyak kepiting muda yang tertangkap di lokasi zona depan mangrove dengan menggunakan alat tangkap rakkang. Alat rakkang umumnya dipasang di muara sungai, pinggiran sungai, pinggiran pantai yang berlumpur dan sering terendam air pasang. Banyaknya kepiting muda yang tertangkap dengan rakkang disebabkan pada tingkat megalopa kepiting mulai beruaya pada dasar perairan berlumpur menuju perairan pantai, dan biasanya pertama kali memasuki perairan muara sungai, kemudian ke perairan berhutan bakau untuk kembali melangsungkan perkawinan (AFRIANTO & LIVIAWATY 1993).
WEBLEY et al. (2009) menyatakan bahwa megalopa dari beberapa spesies kepiting menunjukkan seleksi habitat aktif ketika akan menetap. Megalopa ini biasanya memilih habitat yang kompleks secara struktural yang dapat memberikan perlindungan dan makanan. Kepiting lumpur yang portunid, S. serrata, umumnya ditemukan di muara yang berlumpur Indo-Pasifik Barat setelah mencapai lebar karapas > 40 mm. Meskipun telah dilakukan upaya besar, mekanisme perekruitan kepiting lumpur remaja ke muara tidak dipahami karena megalopa dan tahap awal crablets (lebar karapas < 30 mm) jarang ditemukan. Binatang ini ditempatkan di arena di mana mereka punya pilihan habitat: lamun, lumpur atau pasir dan arena di mana mereka tidak punya pilihan. Berlawanan dengan asosiasi yang ditunjukkan oleh megalopa kepiting portunid lain, megalopa S. serrata tidak selektif terhadap habitat muara ini, menunjukkan bahwa mereka cenderung tidak akan memilih
BIOLOGI POPULASI KEPITING BAKAU
451
habitat ini, atau tidak memperoleh keuntungan dengan memilih salah satu dari yang lain. Namun kepiting muda, lebih memilih lamun, menunjukkan bahwa berada dalam lamun bermanfaat bagi kepiting muda dan meningkatkan kemungkinan bertahan hidup. Perilaku yang selektif mulai berkembang pada tahap kepiting muda, namun belum tampak pada tahap megalopa.
Gambar 3. Distribusi lebar karapas S. serrata di beberapa zona hutan mangrove, di Taman Nasional Kutai, November 2008 – Juni 2009.
Figure 3. Distribution of carapace width of S. serrata in several zone of mangrove forest, at Kutai National Park, November 2008 to June 2009.
Rasio kelamin S. serrata hasil tangkapan rakkang lebih didominasi oleh jenis kelamin jantan dengan nisbah jantan : betina adalah 1:0,85 dan nilai P<0,05. Hal ini menunjukkan adanya perbedaan yang berarti dari nisbah kelamin 1:1. Dominasi jantan diduga terjadi karena adanya persaingan makanan dan sifat yang agresif dari S. serrata jantan, sehingga individu jantan lebih sering masuk ke dalam rakkang.
2 D G r a p h 1
C A R A P A S W ID T H ( m m )
2 0 4 0 6 0 8 0 1 0 0 1 2 0 1 4 0 1 6 0 1 8 0
FR
EQ
UE
NC
Y (
ind)
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
1 4 0
1 6 0
M ID L L E Z O N E O F M A N G R O V E /H O O K (M A L E ) M ID L L E Z O N E O F M A N G R O V E /H O O K (F E M A L E ) F R O N T Z O N E O F M A N G R O V E /T R A P ( M A L E ) F R O N T Z O N E O F M A N G R O V E /T R A P ( F E M A L E ) IN S H O R E Z O N E /G IL L N E T ( M A L E ) IN S H O R E Z O N E /G IL L N E T ( F E M A L E )

WIJAYA, YULIANDA, BOER & JUWANA
452
Scylla serrata hasil tangkapan pada zona perairan pantai dengan menggunakan alat rengge, menunjukkan sebesar 42% merupakan kepiting yang berukuran kurang dari 100 mm, dan sisanya (58%) merupakan kepiting yang berukuran lebih dari 100 mm dan diduga sudah dewasa kelamin. Sebaran lebar karapas S. serrata lebih bervariasi dibandingkan kedua alat sebelumnya, dengan frekuensi tangkapan tertinggi pada interval 89,5-121,5 mm.
Rasio kelamin S. serrata hasil tangkapan rengge lebih didominasi jenis kelamin betina dengan nisbah jantan:betina adalah 1:2,5 (P<0,05). Lebih banyaknya kepiting betina yang tertangkap karena pola migrasi reproduksi kepiting betina yang memijah di laut, sehingga mereka berenang ke laut dan tertangkap oleh alat rengge. Rengge (gillnet) digunakan di perairan dangkal di pesisir. Nelayan umumnya tidak secara khusus menggunakan rengge untuk menangkap kepiting, namun hanya merupakan hasil sampingan selain ikan yang menjadi tujuan utama tangkapan. Waktu penggunaan rengge dapat siang atau malam hari. Kepiting betina yang tertangkap sebagian adalah kepiting yang matang gonad dan akan memijah, atau sebagian lagi adalah kepiting betina yang salin (selesai memijah). Variasi pada ukuran lebar karapas kepiting yang tertangkap oleh alat rengge terjadi karena sebagian adalah kepiting betina matang gonad yang bermigrasi ke laut untuk memijah dan sebagian lagi adalah kepiting muda (crablets) yang bermigrasi ke mangrove untuk mencari makan dan kawin.
Sebaran Temporal Induk Betina Matang Gonade (TKG IV)Pengamatan terhadap induk betina matang gonad dilakukan secara
morfologi pada semua sampel kepiting. Tingkat Kematangan gonad (TKG) yang diamati adalah TKG IV, yang secara morfologi dapat diamati dengan jelas secara visual. Grafik sebaran frekuensi induk betina TKG IV pada masing-masing stasiun disajikan pada Gambar 4.
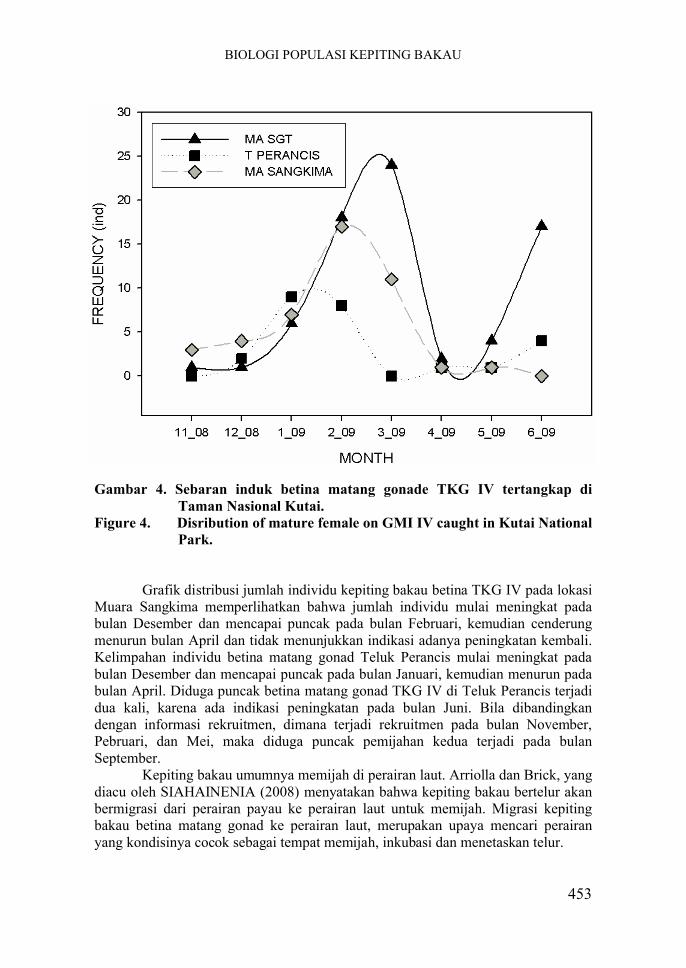
Induk betina matang gonad TKG IV yang tertangkap di habitat mangrove TNK mempunyai sebaran ukuran lebar karapas antara 91 -171 mm, sedangkan ukuran berat tubuhnya berkisar antara 170-870 gram. Kelimpahan individu betina matang gonad terbanyak di Muara Sangatta dibanding di lokasi lain. Hal ini terjadi karena Sungai Sangatta merupakan sungai terbesar di kawasan hutan mangrove TNK, sehingga menjadi pintu masuk utama kepiting bakau yang beruaya kembali ke hutan mangrove.
Grafik distribusi jumlah individu kepiting bakau betina TKG IV pada lokasi Muara Sangatta memperlihatkan bahwa jumlah individu mulai mengalami peningkatan pada bulan Januari dan mencapai puncak pada bulan Maret, kemudian cenderung menurun bulan April dan ada indikasi mulai meningkat kembali pada bulan Juni. Diduga di lokasi Muara Sangatta puncak frekuensi betina matang gonad yang kedua terjadi pada bulan Agustus, berdasarkan perkiraan adanya rekruitmen yang terjadi pada bulan Oktober.
BIOLOGI POPULASI KEPITING BAKAU
453
Gambar 4. Sebaran induk betina matang gonade TKG IV tertangkap di Taman Nasional Kutai.
Figure 4. Disribution of mature female on GMI IV caught in Kutai National Park.
Grafik distribusi jumlah individu kepiting bakau betina TKG IV pada lokasi Muara Sangkima memperlihatkan bahwa jumlah individu mulai meningkat pada bulan Desember dan mencapai puncak pada bulan Februari, kemudian cenderung menurun bulan April dan tidak menunjukkan indikasi adanya peningkatan kembali. Kelimpahan individu betina matang gonad Teluk Perancis mulai meningkat pada bulan Desember dan mencapai puncak pada bulan Januari, kemudian menurun pada bulan April. Diduga puncak betina matang gonad TKG IV di Teluk Perancis terjadi dua kali, karena ada indikasi peningkatan pada bulan Juni. Bila dibandingkan dengan informasi rekruitmen, dimana terjadi rekruitmen pada bulan November, Pebruari, dan Mei, maka diduga puncak pemijahan kedua terjadi pada bulan September.
Kepiting bakau umumnya memijah di perairan laut. Arriolla dan Brick, yang diacu oleh SIAHAINENIA (2008) menyatakan bahwa kepiting bakau bertelur akan bermigrasi dari perairan payau ke perairan laut untuk memijah. Migrasi kepiting bakau betina matang gonad ke perairan laut, merupakan upaya mencari perairan yang kondisinya cocok sebagai tempat memijah, inkubasi dan menetaskan telur.

WIJAYA, YULIANDA, BOER & JUWANA
454
Dengan demikian merupakan juga upaya penjamin kelangsungan hidup embrio serta bagi larva yang dihasilkan. Kecocokan tersebut menurut KASRY (1996), terutama terhadap parameter suhu dan salinitas lingkungan. intensitas pemijahan tertinggi atau puncak musim pemijahan kepiting bakau terjadi pada bulan Februari sampai April. Hal tersebut berarti puncak musim pemijahan kepiting bakau terjadi pada akhir musim hujan sampai menjelang awal musim panas. SIAHAINENIA (2008) menduga hal ini dimaksudkan untuk menjamin ketersediaan pakan alami bagi larva yang akan ditetaskannya. Pada musim hujan sejumlah besar zat hara dari daratan terangkut ke laut melalui aliran sungai maupun aliran air tawar lainnya, sehingga produktifitas perairan menjadi lebih tinggi. Kondisi ini ditunjang dengan intensitas cahaya matahari yang tinggi pada musim panas, yang menyebabkan terjadinya fotosintesa fitoplankon. Kelimpahan fitoplankton selanjutnya akan berdampak terhadap kehadiran zooplankton yang merupakan makanan alami larva kepiting bakau. HASTUTI (1998), menyatakan bahwa telur tingkat akhir, embrio, dan larva kepiting bakau merupakan penghuni laut dengan media bersalinitas tinggi (polihaline). Pada stadia ini kepiting bakau berada dalam lingkungan media dengan osmolaritas yang mantap yang mendekati isoosmotik dengan cairan internal tubuhnya. Hal tersebut di atas berarti, mulai awal pembuahan sel telur, kepiting bakau sudah membutuhkan perairan dengan salinitas yang relatif tinggi.
Pola PertumbuhanPola pertumbuhan kepiting bakau dianalisa menggunakan metode regresi dengan melihat hubungan antara lebar karapas kepiting bakau dengan bobot tubuhnya. Hubungan panjang dan bobot S. serrata (a dan b) disajikan pada Tabel 2. Hasil uji t nilai b untuk kepiting jantan maupun betina menunjukkan bahwa thit lebih besar dibanding ttabel, sehingga dapat dikatakan hubungan lebar karapas dengan bobot S. serrata di habitat mangrove TNK tidak isometrik. Perbedaan ini cukup signifikan dengan nilai P<0,05.
Nilai b akan menjadi indikator yang mendeskripsikan pola pertumbuhan kepiting bakau, sedangkan dari nilai koefisien korelasi (r2) dapat diketahui keeratan hubungan antara lebar karapas kepiting bakau dan bobot tubuhnya, sehingga dapat ditentukan apakah individu dalam suatu populasi dapat diduga bobot tubuhnya dengan mengetahui ukuran tubuhnya atau tidak. Nilai koefisien korelasi (r2) 0,886-0,924 menunjukkan bahwa terdapat hubungan yang cukup erat antara ukuran lebar karapas dengan bobot tubuhnya, sehingga biomass populasi kepiting bakau dapatdiduga dengan mengetahui ukuran lebar karapasnya. Penelitian ALI et al. (2004) menunjukkan hubungan lebar karapas-bobot untuk S. serrata jantan di ekosistem mangrove di Khulna Bangladesh adalah W = 0,0078 CW3,06. sedangkan pada S. serrata betina W = 0,0078 CW1,8928. Hubungan lebar karapas dengan bobot pada induk betina S. serrata matang gonade di Estuari Umlalazy Afrika Selatan adalah Y = 0,0014 X2,56 (DAVIS et al. 2004).
BIOLOGI POPULASI KEPITING BAKAU
455
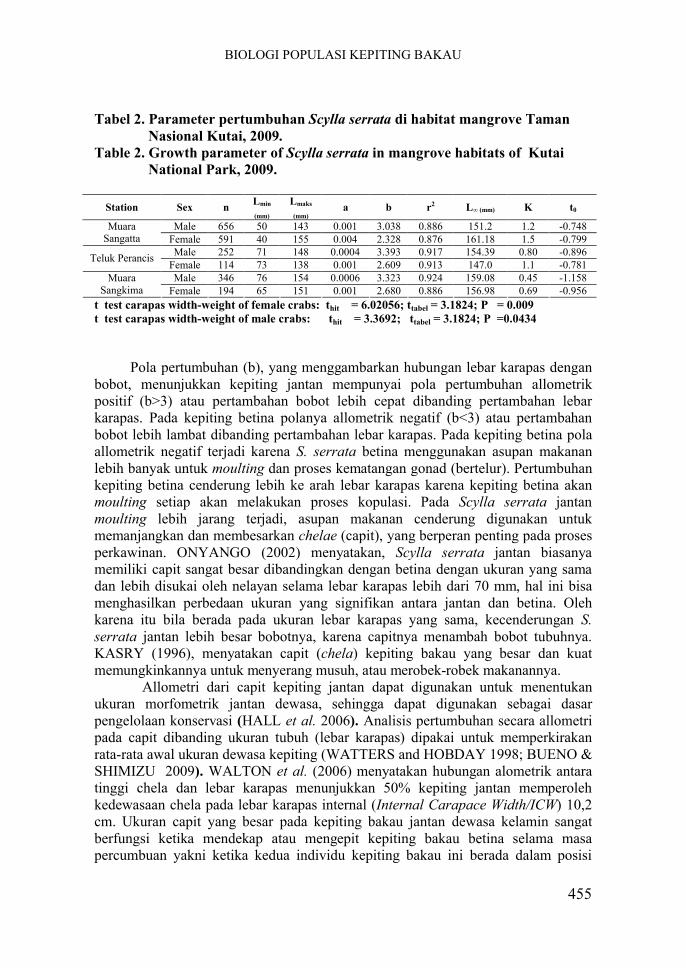
Tabel 2. Parameter pertumbuhan Scylla serrata di habitat mangrove Taman Nasional Kutai, 2009.
Table 2. Growth parameter of Scylla serrata in mangrove habitats of Kutai National Park, 2009.
Station Sex nLmin
(mm)
Lmaks
(mm)a b r2 L∞ (mm) K t0
Male 656 50 143 0.001 3.038 0.886 151.2 1.2 -0.748Muara Sangatta Female 591 40 155 0.004 2.328 0.876 161.18 1.5 -0.799
Male 252 71 148 0.0004 3.393 0.917 154.39 0.80 -0.896Teluk Perancis
Female 114 73 138 0.001 2.609 0.913 147.0 1.1 -0.781Male 346 76 154 0.0006 3.323 0.924 159.08 0.45 -1.158Muara
Sangkima Female 194 65 151 0.001 2.680 0.886 156.98 0.69 -0.956t test carapas width-weight of female crabs: thit = 6.02056; ttabel = 3.1824; P = 0.009t test carapas width-weight of male crabs: thit = 3.3692; ttabel = 3.1824; P =0.0434
Pola pertumbuhan (b), yang menggambarkan hubungan lebar karapas dengan bobot, menunjukkan kepiting jantan mempunyai pola pertumbuhan allometrik positif (b>3) atau pertambahan bobot lebih cepat dibanding pertambahan lebar karapas. Pada kepiting betina polanya allometrik negatif (b<3) atau pertambahan bobot lebih lambat dibanding pertambahan lebar karapas. Pada kepiting betina pola allometrik negatif terjadi karena S. serrata betina menggunakan asupan makanan lebih banyak untuk moulting dan proses kematangan gonad (bertelur). Pertumbuhan kepiting betina cenderung lebih ke arah lebar karapas karena kepiting betina akan moulting setiap akan melakukan proses kopulasi. Pada Scylla serrata jantan moulting lebih jarang terjadi, asupan makanan cenderung digunakan untuk memanjangkan dan membesarkan chelae (capit), yang berperan penting pada proses perkawinan. ONYANGO (2002) menyatakan, Scylla serrata jantan biasanya memiliki capit sangat besar dibandingkan dengan betina dengan ukuran yang sama dan lebih disukai oleh nelayan selama lebar karapas lebih dari 70 mm, hal ini bisa menghasilkan perbedaan ukuran yang signifikan antara jantan dan betina. Oleh karena itu bila berada pada ukuran lebar karapas yang sama, kecenderungan S. serrata jantan lebih besar bobotnya, karena capitnya menambah bobot tubuhnya. KASRY (1996), menyatakan capit (chela) kepiting bakau yang besar dan kuat memungkinkannya untuk menyerang musuh, atau merobek-robek makanannya.
Allometri dari capit kepiting jantan dapat digunakan untuk menentukan ukuran morfometrik jantan dewasa, sehingga dapat digunakan sebagai dasar pengelolaan konservasi (HALL et al. 2006). Analisis pertumbuhan secara allometri pada capit dibanding ukuran tubuh (lebar karapas) dipakai untuk memperkirakan rata-rata awal ukuran dewasa kepiting (WATTERS and HOBDAY 1998; BUENO & SHIMIZU 2009). WALTON et al. (2006) menyatakan hubungan alometrik antara tinggi chela dan lebar karapas menunjukkan 50% kepiting jantan memperoleh kedewasaan chela pada lebar karapas internal (Internal Carapace Width/ICW) 10,2 cm. Ukuran capit yang besar pada kepiting bakau jantan dewasa kelamin sangat berfungsi ketika mendekap atau mengepit kepiting bakau betina selama masa percumbuan yakni ketika kedua individu kepiting bakau ini berada dalam posisi

WIJAYA, YULIANDA, BOER & JUWANA
456
berpasangan (doublers), serta untuk membalik tubuh kepiting bakau betina ketika proses kopulasi akan berlangsung (SIAHAINENIA 2008). Capit yang besar juga dibutuhkan kepiting bakau jantan dewasa kelamin untuk bertarung dengan jantan lainnya dalam upaya mempertahankan wilayah kawin (matting territory), mempertahankan dirinya sendiri serta melindungi dan mempertahankan betina yang menjadi pasangan kawinnya, mengingat menjelang kopulasi kepiting bakau betina melakukan pergantian kulit (moulting) sehingga bertubuh lunak dan sangat rentan terhadap serangan atau bahkan pemangsaan dari kepiting bakau lainnya, karena sifat kanibalisme yang dimilikinya (KASRY 1996).
Dengan menggunakan bantuan program Elefan dari FISAT-II diperoleh nilai dugaan kurva pertumbuhan von Bertalanffy yang meliputi panjang infiniti (L∞) dan kecepatan pertumbuhan (K). Parameter pertumbuhan S. serrata di habitat mangrove TNK dapat dilihat pada Tabel 2. Hasil analisis program Elefan memperlihatkan lebar karapas maksimum yang dapat dicapai berkisar antara 143-155 mm dengan kecepatan pertumbuhan (K) berkisar antara 0,45-1,50. Kecepatan pertumbuhan kepiting betina lebih besar dibandingkan kepiting jantan, karena kepiting betina lebih sering melakukan moulting dibanding kepiting jantan, sehingga cenderung lebih cepat mencapai L∞. LE VAY et al. (2007) menemukan nilai K dari 170 ekor kepiting S. paramamosain yang tertangkap kembali sebesar 2.39 (r2 = 0,63) dengan t0 pada usia - 0,0095 tahun
Kecepatan pertumbuhan S. serrata di Muara Sangatta lebih tinggi dibanding lokasi lainnya. Umumnya kepiting yang ditangkap di Muara Sangatta berukuran belum dewasa kelamin (lebar karapas kurang dari 110 mm). WIJAYA (2010) menduga, perbedaan nilai K S. serrata di tiga lokasi pengamatan tersebut disebabkan oleh perbedaan ekologi dan ciri habitat mangrovenya.
Menurut SIAHAINENIA (2008) kepiting yang berukuran kecil memberikan garis regresi ke arah slope yang lebih tajam, karena modus tertinggi yang dilalui garis pertumbuhan lebih banyak pada kelompok kepiting kecil, sehingga nilai K menjadi besar. Kecepatan pertumbuhan S. serrata di Muara Sangkima menunjukkan kecenderungan yang relatif lebih kecil dibanding pada kedua lokasi lainnya. Hal ini berkaitan dengan kondisi ukuran lebar karapas kepiting S. serrata yang ditemukan di wilayah tersebut umumnya berukuran lebih dari dewasa kelamin, sehingga kecepatan pertumbuhannya menjadi lebih lambat. Kepiting betina dewasa lebih banyak menggunakan energinya untuk pertumbuhan dan perkembangan gonad(LAVINA yang diacu oleh SIAHAINENIA 2008). ONYANGO (2002) menyatakan S. serrata betina yang lebih besar tertangkap selama aktif makan, sehinggamemungkinkan mereka menyimpan energi yang cukup untuk migrasi dan bertelur.
Pola pertumbuhan S. serrata jantan di habitat mangrove TNK bersifat allometrik positif, sedangkan S. serrata betina bersifat allometrik negatif. Kepiting S. serrata mempunyai koefisien pertumbuhan (K) berkisar antara 0,45–1,50. Koefisien pertumbuhan S. serrata di Muara Sangatta lebih tinggi dibanding lokasi lainnya. Tekanan penangkapan kepiting bakau S. serrata sudah berada di atas laju eksploitasi maksimal, hanya di lokasi Muara Sangkima tekanan penangkapan masih sedikit di bawah eksploitasi maksimal. Distribusi lebar karapas S. serrata di zona tengah hutan mangrove, yang diperoleh dari hasil tangkapan alat pengait, umumnya berukuran lebih dari 100 mm, dengan frekuensi tangkapan tertinggi pada interval

BIOLOGI POPULASI KEPITING BAKAU
457
109,5-129,5 mm. Ukuran lebar ini relatif lebih besar dibandingkan ukuran lebar karapas S. serrata di zona depan hutan mangrove, dimana frekuensi tangkapan tertinggi pada interval 78-89 mm, dan di zona perairan pantai, dimana frekuensi tangkapan tertinggi pada interval 89,5-121,5 mm. Kelimpahan individu betina matang gonad mencapai puncak pada bulan Januari, Februari, dan Maret. Puncak kelimpahan yang kedua diduga terjadi pada bulan Agustus dan September.
Laju MortalitasMortalitas adalah angka kematian dalam populasi. Laju mortalitas adalah
laju kematian, yang didefinisikan sebagai jumlah individu yang mati dalam satu satuan waktu. Laju mortalitas total dapat disebabkan karena adanya laju mortalitas alami dan atau laju mortalitas penangkapan. Laju mortalitas alami pada kepiting bakau disebabkan karena kepiting bakau tidak pernah tertangkap sehingga mati alami karena umur tua, atau karena daya dukung lingkungan yang rendah, misalnya akibat perubahan lingkungan yang ekstrim atau tidak tercukupinya makanan alami/kelaparan (SPARRE & VENEMA 1999).
Analisis laju mortalitas kepiting bakau dilakukan dengan menggunakan estimasi mortalitas dari FISAT-II, yang didasarkan pada data lebar karapas kepiting bakau yang tertangkap. Laju mortalitas total (Z) digambarkan sebagai nilai numerik dari kemiringan (slope) garis regresi antara logaritma N/dt terhadap umur relatif kepiting yang tertangkap, dan dihitung dari persamaan pertumbuhan VON BERTALANFFY yang dikenal dengan metode kurva hasil tangkapan. Nilai laju mortalitas total, mortalitas alami, dan mortalitas penangkapan disajikan pada Tabel 3.
Hasil analisis menunjukkan di ketiga lokasi umumnya tekanan penangkapan (eksploitasi faktual) kepiting bakau S. serrata sudah berada di atas laju eksploitasi maksimal, hanya kepiting jantan di lokasi Muara Sangkima yang laju eksploitasi faktualnya masih sedikit di bawah eksploitasi maksimal (Tabel 3).
Tabel 3. Laju mortalitas dan laju eksploitasi S. serrata.Table 3. The rate of mortality and the rate of exploitation of S. serrata.
STATION SEX Z M FE
faktualE
maxMale 2.89 1.2584 1.6316 0.5645 0.457
Muara SangattaFemale 2.41 1.0744 1.3356 0.554 0.407Male 2.87 0.9430 1.9270 0.671 0.606
Teluk PerancisFemale 3.40 1.1774 2.2226 0.654 0.555Male 1.36 0.64177 0.71823 0.5281 0.555
Muara SangkimaFemale 1.79 0.85202 0.93798 0.5240 0.516

WIJAYA, YULIANDA, BOER & JUWANA
458
Tingginya kematian karena penangkapan di Muara Sangatta diduga karena adanya pemukiman nelayan, sehingga aktifitas penangkapan cukup tinggi. Selain itu, kondisi ekosistem mangrove Muara Sangatta juga telah terdegradasi akibat tingginya pembukaan mangrove untuk tambak. Areal lahan kritis di mangrove Muara Sangatta mencapai ± 440,3 ha. SIAHAINENIA (2008) pada penelitiannya di Kabupaten Subang juga menemukan bahwa kelimpahan kepiting bakau terendah umumnya dijumpai pada zona belakang hutan yang memiliki tingkat kerapatan vegetasi mangrove rendah, serta berada di sekitar areal pemukiman penduduk atau mendapat tekanan akibat tingginya aktifitas masyarakat.
Tingginya angka mortalitas penangkapan di Teluk Perancis diduga karena penangkapan kepiting bakau di lokasi ini lebih banyak menggunakan alat tangkap pancing/pengait. Teluk Perancis memiliki hutan mangrove yang masih cukup rapat, sehingga alat tangkap yang sesuai digunakan adalah pengait. Alat tangkap pengait cenderung hanya menangkap kepiting yang berukuran besar saja, akibatnya hasil perhitungan konstanta pertumbuhan (K) menjadi kecil, karena semakin besar kepiting semakin lambat pertumbuhan lebar karapasnya. Nilai K merupakan salah satu variabel yang dipakai dalam formula untuk menghitung mortalitas alami. Kecilnya nilai K akan mempengaruhi nilai mortalitas alami (M) menjadi lebih kecil(PAULI yang diacu oleh SPARRE & VENEMA 1999), dan akibatnya nilai mortalitas penangkapan (F) cenderung menjadi lebih besar. Selain itu, di Dusun Teluk Lombok yang berdekatan dengan Teluk Perancis juga cukup banyak penduduk, sehingga aktifitas penangkapan juga menjadi lebih besar. Laju penangkapan S. serrata di lokasi Muara Sangkima cenderung lebih rendah dibanding kedua lokasi lainnya. Rendahnya tekanan penangkapan diduga karena lokasi ini jauh dari pemukiman penduduk. WALTON (yang diacu oleh EWEL 2008) menyatakan bahwa populasi Scylla serrata dapat mempunyai sebaran ukuran yang berbeda karena perbedaan kondisi lingkungan dan pola penangkapan. Maka EWEL (2008) menyarankan peraturan lokal (local regulations) sebagai tambahan larangan daerah (regional restrictions) mungkin layak (appropriate) untuk banyak wilayah di Indo-Pacific. Pemantauan populasi secara teratur dapat meningkatkan komunitas kecil terpisah mengelola sumberdaya penting secara berkelanjutan.
KESIMPULAN
Pola pertumbuhan Scylla serrata jantan di habitat mangrove TNK bersifat allometrik positif, sedangkan S. serrata betina bersifat allometrik negatif. Koefisien pertumbuhan (K) S. serrata berkisar antara 0,45-1,50. Koefisien pertumbuhan S. serrata di Muara Sangatta lebih tinggi disbanding lokasi lainnya. Tekanan penangkapan kepiting bakau S. serrata sudah berada di atas laju eksploitasi maksimal, hanya di lokasi Muara Sangkima tekanan penangkapan masih sedikit di bawah eksploitasi maksimal. Distribusi lebar karapas S. serrata di zona tengah hutan

BIOLOGI POPULASI KEPITING BAKAU
459
mangrove, yang diperoleh dari hasil tangkapan alat pengait, umumnya berukuran lebih dari 100 mm, dengan frekuensi tangkapan tertinggi pada interval 109,5-129,5 mm. Ukuran lebar ini relatif lebih besar dibandingkan ukuran lebar karapas S. serrata di zona depan hutan mangrove, dimana frekuensi tangkapan tertinggi pada interval 78-89 mm; dan di zona perairan pantai, dimana frekuensi tangkapan tertinggi pada interval 89,5-121,5 mm. Kelimpahan individu betina matang gonad mencapai puncak pada bulan-bulan Januari, Februari dan Maret. Puncak kelimpahan yang kedua diduga terjadi pada bulan Agustus dan September.
DAFTAR PUSTAKA
AFRIANTO, E. dan E. LIVIAWATY 1993. Pemeliharaan kepiting. Kanisius. Yogyakarta: 78 hal.
ALI, M.Y., D. KAMAL, S.M.M. HOSSAIN, M.A. AZAM, W. SABBIR, A. MURSHIDA, B. AHMED and K. AZAM 2004. Biological studies of the mud crab, Scylla serrata (Forskal) of the sundarbans mangrove ecosystem in Khulna Region of Bangladesh. Pakistan Journal of Biological Sciences 7: 1981-1987.
BONINE, K.M., E.P. BJORKSTEDT, K.C. EWEL and M. PALIK 2008. Population characteristic of the mangrove crab S. serrata (Decapoda: Portunidae) in Kosrae, Federation States of Micronesia: Effect of harvest and implications for management. Jurnal Pacific Science 62: 1-19.
BUENO, S.L.S. and R.M. SHIMIZU 2009. Allometric growth, sexual maturity and adult male chelae dimorphism in Aegla franca (Decapoda: Anomura: Aeglidae). Journal of Crustacean Biology 29(3): 317-328.
CHOLIK, F. 1999. Review of mud crab culture research in Indonesia. ACIAR Proceedings No. 78. Proceedings of An International Scientific Forum Held In Darwin, Australia, 21–24 April 1997. Canberra. Australia: 14-20.
DAVIS, JA., G.J. CHURCHILL, T. HECHT and P. SORGELOSS 2004. Spawning characteristics of the South African mudcrab Scylla serrata (Forskall) in captivity. Journal of The World Aquaculture Society 35:121-133.
EFFENDIE, M.I. 1979. Metode biologi perikanan. Yayasan Dewi Sri. Bogor: 112 hal.
EWEL, K.C. 2008. Mangrove crab (Scylla serrata) populations may sometimes be best managed locally. Journal of Sea Research 59: 114 – 120.

WIJAYA, YULIANDA, BOER & JUWANA
460
HALL, N.G., K.D. SMITH, S. de LESTANG and I.C. POTTER 2006. Does the largest chela of the males of three crab species undergo an allometric change that can be used to determine morphometric maturity ?. ICES J. Mar. Sci.63 (1): 140-150.
HASTUTI, S. 1998. Pertumbuhan embrio kepiting bakau, Scylla serrata, pada beberapa tingkat salinitas media. (Thesis). Program Pascasarjana IPB. Bogor.
HILL B. J. 1975. Abundance, breeding and growth of the crab Scylla serrata in two South African estuaries. Marine Biology 32: 119–126.
KASRY, A. 1996. Budidaya kepiting bakau dan biologi ringkas. Penerbit Bharata. Jakarta. 93 hal.
KEENAN, CP., PJF. DAVIE, DL. MANN. 1998. A Revision of the genus Scylla De Haan, 1983 (Crustacea: Decapoda: Brachyura: Portunidae). The Raffles Bulletin of Zoology 46 (1): 217-245.
LE VAY, L. 2001. Ecology and management of mud crab Scylla spp. Asian Fisheries Science 14:101-111.
LE VAY L., V. N. UT and M.E. WALTON 2007. Population ecology of the mud crab Scylla paramamosain (Estampador) in an estuarine mangrove system; a mark-recapture study. Marine Biology 151:1127–1135.
ONYANGO, S. D. 2002. The breeding cycle of Scylla serrata (Forskål, 1755) at Ramisi River estuary, Kenya. Wetlands Ecology and Management 10: 257–263.
SPARRE, P. and S.C. VENEMA 1999. Introduksi pengkajian stok ikan tropis buku-1 manual (Edisi Terjemahan). Kerjasama Organisasi Pangan, Perserikatan Bangsa-Bangsa dengan Pusat Penelitian dan Pengembangan Perikanan, Badan Penelitian dan Pengembangan Pertanian. Jakarta: 435 hal.
SIAHAINENIA, L. 2008. Bioekologi kepiting bakau (Scylla spp.) di ekosistem mangrove Kabupaten Subang Jawa Barat. Disertasi S3. Sekolah Pascasarjana IPB. Bogor.
[TNK] TAMAN NASIONAL KUTAI. 2005. Data dasar Taman Nasional Kutai. Balai Taman Nasional Kutai. Bontang, Kalimantan Timur.
WATTERS, G. and A.J. HOBDAY 1998. A new method for estimating the morphometric size at maturity of crabs. Can. J. Fish. Aquat. Sci. 55(3): 704-714.

BIOLOGI POPULASI KEPITING BAKAU
461
WALTON, MARK E., L. LE VAY, L.M. TRUONG, and V.N. UT 2006. Significance of mangrove–mudflat boundaries as nursery grounds for the mud crab, Scylla paramamosain. Marine Biology 149: 1199–1207.
WEBLEY, J.A.C., R.M. CONNOLLY and R.A. YOUNG 2009. Habitat selectivity of megalopae and juvenile mud crabs (Scylla serrata): implications for recruitment mechanism. Marine Biology 156: 891-899.
WIJAYA, N.I. 2010. Pengelolaan zona pemanfaatan ekosistem mangrove melalui optimasi pemanfaatan sumberdaya kepiting bakau (Scylla serrata) di Taman Nasional Kutai Provinsi Kalimantan Timur. Disertasi S3. Mayor Pengelolaan Sumberdaya Pesisir dan Lautan. Sekolah Pascasarjana, Institut Pertanian Bogor. Manuscript.




















