
BAB II TINJAUAN PUSTAKA 2.1 Melasma 2.1.1 Definisi ... - UNUD
28
BAB II TINJAUAN PUSTAKA 2.1 Melasma 2.1.1 Definisi Melasma Istilah melasma berasal dari bahasa Yunani yaitu “melas” yang memiliki arti hitam. Melasma merupakan hipermelanosis umum akuisita yang terdapat pada area kulit yang sering terpapar sinar matahari. Keluhan melasma biasanya terjadi secara simetris. Area predileksi melasma yaitu paling sering pada pipi, bagian atas bibir, dagu dan dahi, namun area lainnya juga dapat terkena. Istilah kloasma (berasal dari bahasa yunani “chloazein” artinya kehijauan) sering digunakan untuk menggambarkan keluhan melasma saat kehamilan. Bagaimanapun juga pigmentasi tersebut tidak pernah berwarna kehijauan sehingga istilah melasma lebih tepat digunakan (Debabrata, 2009; Handel dkk., 2014). 2.1.2 Epidemiologi Melasma Melasma pada dasarnya dapat mengenai semua ras terutama penduduk yang terpajan sinar matahari dengan intensitas cukup tinggi (daerah tropis) (Soepardiman, 2009). Balkrishnan dkk. (2003) menyatakan bahwa salah satu kondisi yang paling umum terjadinya melasma terutama pada individu keturunan Hispanik dan Asia. Sedangkan Sachdeva (2006) dan Dogra dkk. (2006) menjelaskan tipe kulit Fitzpatrick IV-V merupakan individu yang umum terkena penyakit ini. Melasma terutama dijumpai pada wanita usia reproduksi dengan riwayat langsung terpajan sinar matahari, meskipun didapatkan pula pada pria (± 10%). Di Indonesia, perbandingan kasus wanita dan pria yaitu 24 : 1. Insiden terbanyak pada
Transcript of BAB II TINJAUAN PUSTAKA 2.1 Melasma 2.1.1 Definisi ... - UNUD

BAB II
TINJAUAN PUSTAKA
2.1 Melasma
2.1.1 Definisi Melasma
Istilah melasma berasal dari bahasa Yunani yaitu “melas” yang memiliki arti hitam.
Melasma merupakan hipermelanosis umum akuisita yang terdapat pada area kulit
yang sering terpapar sinar matahari. Keluhan melasma biasanya terjadi secara
simetris. Area predileksi melasma yaitu paling sering pada pipi, bagian atas bibir,
dagu dan dahi, namun area lainnya juga dapat terkena. Istilah kloasma (berasal dari
bahasa yunani “chloazein” artinya kehijauan) sering digunakan untuk
menggambarkan keluhan melasma saat kehamilan. Bagaimanapun juga pigmentasi
tersebut tidak pernah berwarna kehijauan sehingga istilah melasma lebih tepat
digunakan (Debabrata, 2009; Handel dkk., 2014).
2.1.2 Epidemiologi Melasma
Melasma pada dasarnya dapat mengenai semua ras terutama penduduk yang terpajan
sinar matahari dengan intensitas cukup tinggi (daerah tropis) (Soepardiman, 2009).
Balkrishnan dkk. (2003) menyatakan bahwa salah satu kondisi yang paling umum
terjadinya melasma terutama pada individu keturunan Hispanik dan Asia. Sedangkan
Sachdeva (2006) dan Dogra dkk. (2006) menjelaskan tipe kulit Fitzpatrick IV-V
merupakan individu yang umum terkena penyakit ini.
Melasma terutama dijumpai pada wanita usia reproduksi dengan riwayat
langsung terpajan sinar matahari, meskipun didapatkan pula pada pria (± 10%). Di
Indonesia, perbandingan kasus wanita dan pria yaitu 24 : 1. Insiden terbanyak pada

wanita usia 30-44 tahun (Soepardiman, 2009). Berdasarkan penelitian Febrianti,
Aryani Sudharmono, Irma Bernadette (2006) di Departemen Ilmu Kesehatan Kulit
dan Kelamin FK Universitas Indonesia/RS. Dr. Cipto Mangunkusumo Jakarta,
menunjukkan hasil bahwa epidemiologi melasma 97,93% wanita dan 2,07% pria.
Sebuah studi cross sectional dilakukan oleh Nkwo (2011), dengan pemeriksaan
klinis dan kuesioner pada 400 wanita hamil. Dengan hasil prevalensi melasma adalah
15,8%. Pola malar terlihat pada 65,9% kasus, sedangkan 33,8% dari pasien memiliki
pola sentrofasial. Tidak ditemukan kasus dengan pola mandibula. Pada 54,7% kasus
dipengaruhi juga oleh riwayat keluarga atau genetik. Sedangkan menurut Achar Arun
dan Rathi SK (2011), pola melasma yang tersering adalah pola sentrofasial (54,44%).
Dengan pemeriksaan dermoskopi didapatkan tipe dermal merupakan tipe tersering
(54,48 %), sedangkan tipe epidermal sebanyak 21,4 % dan tipe campuran sebanyak
24,03 %.
2.1.3 Etiopatogenesis Melasma
2.1.3.1 Biologi Melanosit
Jumlah seluruh sel pigmen dalam tubuh yaitu sebanyak 1,5 gram. Sebagian besar
melanosit ini terletak di dalam epidermis. Proses produksi melanin di dalam melanosit
melibatkan tidak hanya produksi melanosom di dalam melanosit yang dikenal dengan
melanogenesis, namun juga merekrut dan mentransfer granul pigmen ini melalui
dendrit bercabang menuju ke keratinosit epidermis. Tiap melanosit epidermis bersama
dengan sel-sel epidermis membentuk suatu unit melanin epidermis. Walaupun jumlah
unit melanin epidermis aktif berbeda di tiap area tubuh (gambar 2.1), namun jumlah
keratinosit yang dilayani oleh melanosit tetap sama. Diperkirakan satu melanosit
mengirimkan melanosom ke 30-40 keratinosit viabel. Hubungan antara melanosit
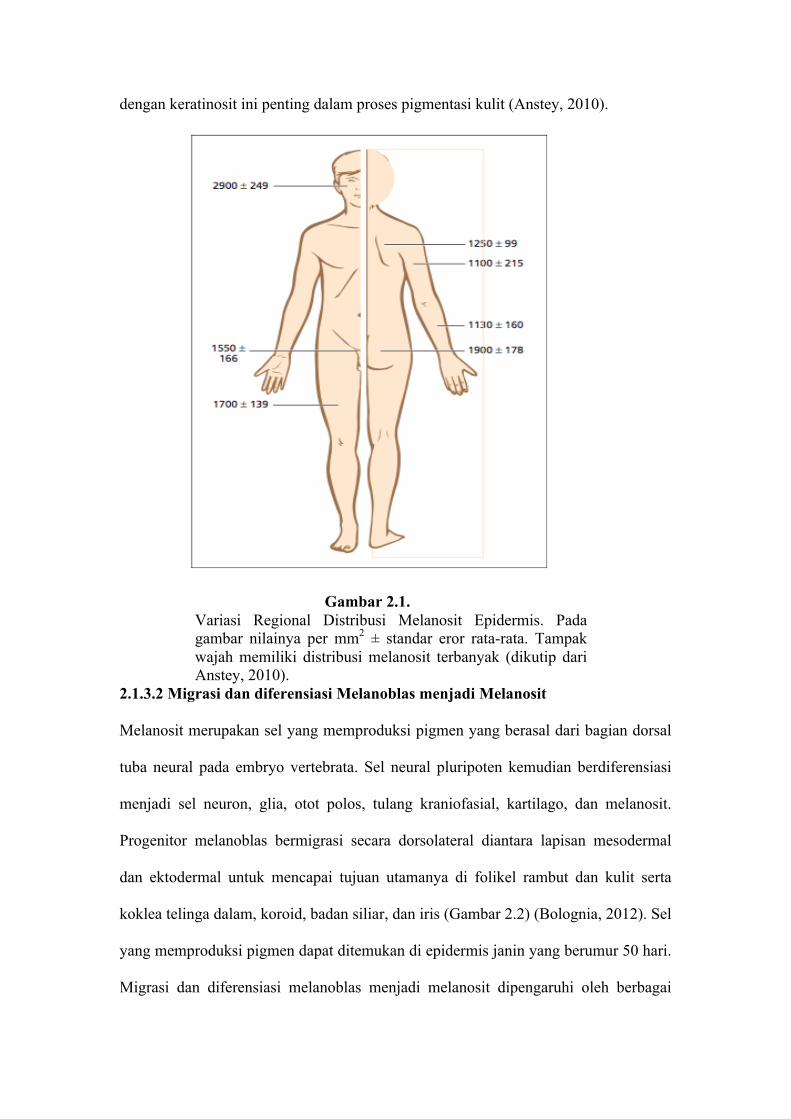
dengan keratinosit ini penting dalam proses pigmentasi kulit (Anstey, 2010).
Gambar 2.1. Variasi Regional Distribusi Melanosit Epidermis. Pada gambar nilainya per mm2 ± standar eror rata-rata. Tampak wajah memiliki distribusi melanosit terbanyak (dikutip dari Anstey, 2010).
2.1.3.2 Migrasi dan diferensiasi Melanoblas menjadi Melanosit
Melanosit merupakan sel yang memproduksi pigmen yang berasal dari bagian dorsal
tuba neural pada embryo vertebrata. Sel neural pluripoten kemudian berdiferensiasi
menjadi sel neuron, glia, otot polos, tulang kraniofasial, kartilago, dan melanosit.
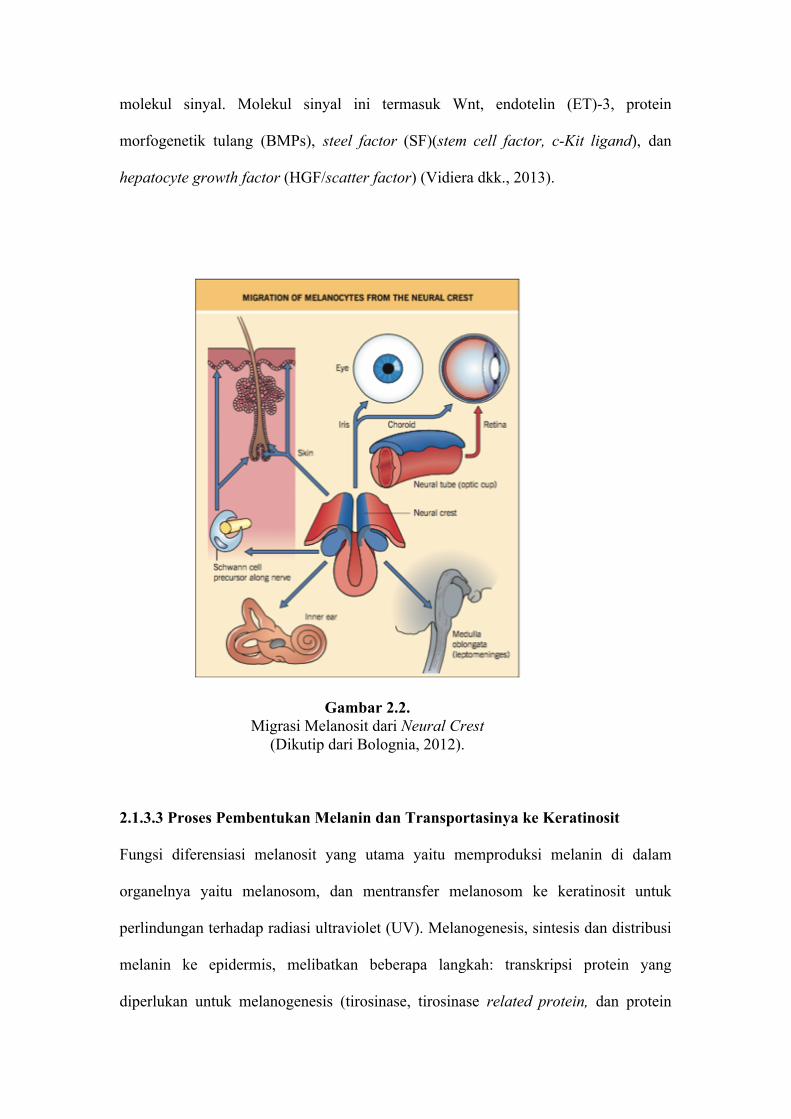
Progenitor melanoblas bermigrasi secara dorsolateral diantara lapisan mesodermal
dan ektodermal untuk mencapai tujuan utamanya di folikel rambut dan kulit serta
koklea telinga dalam, koroid, badan siliar, dan iris (Gambar 2.2) (Bolognia, 2012). Sel
yang memproduksi pigmen dapat ditemukan di epidermis janin yang berumur 50 hari.
Migrasi dan diferensiasi melanoblas menjadi melanosit dipengaruhi oleh berbagai
molekul sinyal. Molekul sinyal ini termasuk Wnt, endotelin (ET)-3, protein
morfogenetik tulang (BMPs), steel factor (SF)(stem cell factor, c-Kit ligand), dan
hepatocyte growth factor (HGF/scatter factor) (Vidiera dkk., 2013).
Gambar 2.2. Migrasi Melanosit dari Neural Crest
(Dikutip dari Bolognia, 2012).
2.1.3.3 Proses Pembentukan Melanin dan Transportasinya ke Keratinosit
Fungsi diferensiasi melanosit yang utama yaitu memproduksi melanin di dalam
organelnya yaitu melanosom, dan mentransfer melanosom ke keratinosit untuk
perlindungan terhadap radiasi ultraviolet (UV). Melanogenesis, sintesis dan distribusi
melanin ke epidermis, melibatkan beberapa langkah: transkripsi protein yang
diperlukan untuk melanogenesis (tirosinase, tirosinase related protein, dan protein

struktural Pmel17 dan Melan A), biosintesis melanin di dalam melanosom,
transportasi melanosom ke ujung dendrit melanosit, dan transfer melanosom ke
keratinosit (Xufeng dkk., 2012).
Gambar 2.3 Biosintesis Melanin. DHI: 5,6 dihydroxyindole; DHICA: 5,6 dihydroxyindole-2-carboxylic acid; DOPA: 3,4 dihidroxy-phenylalanine; MW: Molecular Weight; TRIP: Tyrosinase Related proterin (Dikutip dari Park et Mina, 2012).
Pada biosintesis melanin terdapat dua tipe melanin yang disintesis di dalam
melanosom : (1) eumelanin dan (2) pheomelanin. Eumelanin merupakan pigmen
berwarna hitam atau kecoklatan, dan tidak larut, sedangkan pheomelanin merupakan
pigmen berwarna terang, merah kekuningan yang mengandung sulfur, dan bersifat
larut. Melanin merupakan devatif indole dari 3,4 dihidroxy-phenylalanine (DOPA)

dan terbentuk didalam melanosom melalui beberapa langkah oksidatif (Gambar 2.3).
Sintesis kedua tipe melanin ini melalui langkah katalitik dimana asam amino tyrosine
dioksidase oleh enzim tyrosinase menjadi DOPA, yang merupakan langkah pertama
yang dikenal sebagai jalur Raper-Mason. Kemudian DOPA dioksidasi menjadi
DOPAquinone, yang kemudian diubah menjadi DOPAchrome. DOPAchrome dapat
diubah menjadi 5,6 dihydroxyindole (DHI) atau 5,6 dihydroxyindole-2-carboxylic
acid (DHICA). Reaksi ini dikatalisis oleh enzim DOPAchrome tautomeras atau TRP-
2. Tingkat warna eumelanin apakah berwarna coklat atau hitam berhubungan dengan
rasio DHI/DHICA, dimana rasio yang lebih tinggi akan memproduksi eumelanin
hitam dibandingkan coklat. DOPAquinone juga dapat bergabung dengan glutathione
atau cysteine untuk membentuk cysteinylDOPA, yang membentuk pheomelanin.
(Park et Mina, 2012). Setelah terbentuk melanin, maka selanjutnya melanin di dalam
melanosom akan ditransfer ke keratinosit melalui dendrit melanosit. Melanosom yang
terakumulasi pada ujung dendrit akan digenerasikan oleh mekanisme transport
kooperatif yang memfasilitasi pemanjangan dendrit sehingga mencapai keratinosit.
(Xufeng, 2012)
Fungsi utama melanin yaitu melindungi sel dari kerusakan DNA yang
disebabkan paparan sinar UV(280-400 nm) dengan mengabsorbsi radiasi UV tersebut.
Melanin mengabsorbsi radiasi UV dan mengubahnya menjadi energi panas yang
bersifat kurang toksik (Orazio dkk., 2013). Secara fisiologis radiasi sinar UV
diketahui dapat merangsang sintesis melanin di melanosit. Beberapa studi melaporkan
bahwa radiasi sinar UV dapat meningkatkan produksi faktor parakrin seperti
Adrenocorticotropin hormone (ACTH), endothelin-1, Fibroblast Growth Factor β (β
FGF), Melanocyte Stimulating Hormone α (α MSH), yang memegang peranan
penting dalam patogenesis UV mencetuskan melasma. Melanocyte Stimulating

Hormone α dapat mengurangi pembentukan fotoproduk DNA yang distimulasi UV
dengan meningkatkan nucleotide excision repair (NER) dan mengurangi kerusakan
DNA oksidatif dengan mengurangi ROS. Selain menginduksi faktor parakrin, UV
dapat menginduksi faktor transkripsi Upstrain Transcription Factor 1 (USF-1),
Micropthalmia Transcription Factor (Mitf), Activating Transcription Factor 2 (ATF
2), Nuclear factor erythroid 2 related factor 2 (Nrf-2), p53, serta inhibisi Nuclear
Factor κβ (NFκB). Efek akut sinar UV yaitu via NER menyebabkan survival
mealnosit. Dinamika melanogenesis diinduksi oleh paparan berulang tergantung dosis
UV, interval dan emisi spektrum UVA atau UVB. Dikatakan bahwa peningkatan
dosis UV diatas dosis paparan kumulatif tidak singnifikan meningkatkan
melanogenesis yang diinduksi UV. Seperti yang terlihat di keratinosit, UV dapat
menginduksi cell cycle arrest, aktivasi antioksidan dan enzym perbaikan DNA, serta
regulasi jalur apoptotik melanosit untuk menjaga integritas genomik dan survival
melanosit. Proses regulasi ini dapat meningkatkan melanogenesis sebagai fotoproteksi
pada epidermis dalam melawan sinar UV (Gambar 2.4) (Lee dkk., 2013).

Gambar 2.4 Radiasi Ultraviolet menginduksi pembentukan melanin yang akan ditransportasi ke
keratinosit sebagai fotoproteksi (Dikutip dari Lee dkk., 2013).
2.1.3.4 Pengaruh Faktor Genetik dan Ras dalam Mencetuskan Melasma
Predisposisi genetik dikatakan sangat berperanan dalam patogenesis melasma
dikarenakan adanya laporan tingginya insiden melasma diantara anggota keluarga
pada kelompok ras tertentu. Insidennya berkisar antara 10-70 % di Iran (54,7 %),
Singapore (10,2 %), dan laki-laki latin/Brazil (56,3 %-61%) (Sonthalia & Sarkar,
2015). Negara bagian Asia Tenggara sendiri, memiliki prevalensi bervariasi antara 40
% pada perempuan, 20 % pada laki-laki. Survei global oleh Ortonne dkk. (2009) yang
meneliti perempuan dari 9 negara melaporkan bahwa suseptibilitas untuk menderita
melasma lebih tinggi terdapat pada individu dengan tipe kulit III dan IV dan juga
dilaporkan terdapat prevalensi tinggi dalam hal riwayat keluarga yang positif sebagai
pencetus melasma pada ras afrika amerika.

Hal yang mendasari faktor genetik dalam mencetuskan melasma belum jelas,
namun terdapat perbedaan yang signifikan pada predominansi tipe melasma pada ras
yang berbeda. Secara umum, pola sentrofasial merupakan pola yang paling sering
dilaporkan diikuti dengan pola malar. Namun pada penelitian di India, pasien
melasma dilaporkan lebih banyak mengalami melasma tipe malar dibandingkan
sentrofasial (61 % : 29.3 %) (Sarkar, 2010). Pada penelitian yang dilakukan di Korea,
didapatkan hubungan antara regulasi gen yang terlibat dalam jalur sinyal
melanogenesis dengan melasma, yaitu ekspresi gen ADIPOQ yang mengatur
adiponektin (terlibat dalam melanogenesis) dan gen yang berperanan dalam fungsi
barier kulit seperti S100A8(S100 calcium binding protein A8), SPRR2A (Small
Proline Rich Protein 2A), SPRR2B (Small Proline Rich Protein 2B) dan KLK6
(Kalikrein related Peptidase 6). (Chung, 2014). Kang dkk. (2011) melaporkan adanya
peningkatan ekspresi gen yang berperanan dalam melanogenesis seperti TYRP1
(tyrosinase related protein 1).
2.1.3.5 Pengaruh Faktor Hormonal dalam Mencetuskan Melasma
Melasma terjadi pada 10-15 % perempuan hamil dan 10-25 % pada perempuan yang
memakai kontrasepsi oral. Dalam beberapa dekade, melasma dikenal sebagai topeng
kehamilan, dengan asumsi bahwa penyebabnya adalah peningkatan hormon
perempuan yang berhubungan dengan kehamilan atau pil kontrasepsi. Pada penelitian
yang dilakukan di Korea, didapatkan peningkatan ekspresi reseptor estrogen pada lesi
melasma dermis, dan peningkatan ekspresi reseptor progesteron pada lesi melasma
epidermis. (Jang dkk., 2010). Pada penelitian yang dilakukan oleh Mahmood dkk.
(2011) menemukan hubungan yang signifikan antara peningkatan jumlah estrogen
dengan melasma. Diperkirakan estrogen dapat meningkatkan jumlah Mc1R, yang

merupakan reseptor yang akan berikatan dengan MSH yang selanjutnya dapat
merangsang melanogenesis. Estrogen juga dapat meningkatkan tirosinase, walaupun
mekanisme yang mendasarinya belum jelas. (Mahmood, 2011).
2.1.3.6 Pengaruh Sinar Ultraviolet dalam Mencetuskan Melasma
Radiasi ultraviolet merupakan faktor lingkungan utama yang berperanan dalam
patogenesis melasma. Hal ini berkaitan dengan sinar ultraviolet dalam menghasilkan
stress oksidatif (Miller & Tsao, 2011). Menurut Nishigori (2015), sinar ultraviolet
dapat mempengaruhi sel keratinosit yang nantinya akan berperanan dalam proses
terbentuknya ROS serta NO. Foton pada sinar UV dapat bereaksi dengan atom
oksigen membentuk derivatif radikal bebas seperti superoksid, hidrogen peroksida,
dan radikal hidroksil reaktif. Radikal bebas dapat menyerang makromolekul seperti
protein, lipid, DNA, dan RNA sehingga membentuk fotoproduk pirimidin atau
menyebabkan peroksidase lipid yang akhirnya menghasilkan NO, serta sitokin dan
enzim yang berperanan dalam proses inflamasi (Gambar 2.5). Selanjutnya radikal
bebas serta sitokin inflamasi ini dapat merangsang keratinosit untuk melepaskan
molekul sinyal seperti αMSH, prostaglandin E2 (PGE2), hormon adrenokortikotropin
(ACTH), dan endothelin-1 yang dapat meningkatkan aktivitas tirosinase. Aktivitas
tirosinase yang meningkat dapat menyebabkan peningkatan proses melanogenesis
yang berperanan dalam patogenesis terjadinya melasma. (Kadekaro, 2003).

Gambar 2.5 Sinar Ultraviolet menginduksi pembentukan ROS sebagai penanda stres oksidatif
dalam tubuh (Dikutip dari Nishigori, 2015).
2.1.3.7 Pengaruh Faktor lainnya dalam Mencetuskan Melasma
Faktor lainnya yang dapat menginduksi melasma yaitu paling banyak banyak dibahas
adalah inflamasi. Mediator inflamasi seperti nitric oxide synthase (iNOS), vascular
endothelial growth factor (VEGF), Granulocyte macrophage colony stimulating
factor (GM-CSF), endothelin 1, dan stem cell factor ditemukan meningkat pada lesi
kulit pasien melasma, dan menginduksi peningkatan angiogenesis dan sel inflamasi.

(Passeron, 2013).
Kosmetik dan perawatan seperti peeling serta laser dapat menginduksi
timbulnya melasma. (Duarte & Campos, 2007). Obat-obatan juga dapat menyebabkan
timbulnya melasma karena bersifat fotosensitisasi. Obat-obatan tersebut diantaranya
antikonvulsan, antimalaria, tetrasiklin, amiodarone, sulfonylurea, besi, tembaga,
emas, perak, arsenik, dan bismuth. Namun patogenesisnya masih menjadi perdebatan.
(Guarneri, 2014). Faktor pikologis dan neural juga diperkirakan berhubungan dengan
melasma. Menurut Bak dkk. (2009) yang membandingkan kulit dengan lesi melasma
dengan kulit tanpa lesi melasma pada 6 pasien perempuan dengan melasma,
menemukan terdapat peningkatan ekspresi nerve growth factor receptor (NGFR),
endopeptidase neural, dan serabut saraf pada dermis superfisial lesi melasma.
Saat ini beberapa penelitian juga mengemukakan kemungkinan peranan
gangguan metabolisme lipid dalam patogenesis melasma. Menurut Kang dkk. (2011)
mendapatkan bahwa terdapat penurunan ekspresi gen metabolisme lipid dengan
konsekuensi gangguan fungsi barier kulit pada penderita melasma.
2.1.4 Gambaran Klinis Melasma
Karakteristik melasma memiliki ciri khas berupa makula kecoklatan dengan bentuk
ireguler dan batas yang tegas. Lesi melasma timbul pada area yang sering terpapar
sinar matahari, khususnya pada wajah dan leher, dan kadang-kadang juga terdapat
pada lengan atas dan daerah sternal. (Miot dkk., 2009).
Onset melasma biasanya timbul pada perempuan usia reproduktif (20-40 tahun).
Gambaran pigmen meningkat saat musim panas. Setidaknya terdapat 3 distribusi
klinis pada melasma di wajah yaitu sentrofasial (64 %), malar (27 %), dan mandibular
(9 %). Perjalanan penyakitnya biasanya kronis, memudar saat paparan UV berkurang.

(Jennifer dkk., 2009). Pada pola sentrofasial, lesinya terutama terdapat pada bagian
tengah wajah seperti glabela, frontal, nasal, zigomatik, bibir atas dan dagu. Untuk
melasma tipe malar, lesinya sebagian besar di daerah kedua pipi dan batang hidung,
sedangkan pola mandibular lesinya sebagian besar terletak di daerah ramus
mandibula. Melasma ekstrafasial sangat jarang terjadi dan dapat bermanifestasi pada
lengan atas, lengan bawah, leher, sternum dan kadang-kadang pada punggung.
(Handel, 2014).
Berdasarkan pemeriksaan lampu wood, tipe melasma dibedakan menjadi tipe
epidermal, dermal, dan tipe campuran. Pada melasma tipe epidermal ditandai dengan
tepi yang tegas, lesinya berwarna coklat tua, tampak lebih jelas di bawah pemeriksaan
lampu wood, dan respon terhadap terapi cukup baik. Pada melasma tipe dermal,
ditandai dengan tepi yang tidak terlalu tegas, lesinya berwarna coklat terang atau
kebiruan. Pada pemeriksaan lampu wood tampak tidak berubah. Lesi melasma dermal
memberikan respon yang buruk terhadap terapi. Pada melasma tipe campuran,
merupakan tipe yang paling sering, kombinasi makula berwana coklat gelap, coklat
terang, dan kebiruan. Pada pemeriksaan lampu wood tampak pola yang bercampur.
Terdapat perbaikan sebagian dengan pengobatan (Sarkar dkk., 2014).
2.1.5 Derajat Keparahan Melasma
Derajat keparahan melasma dapat diukur dengan menggunakan alat ukur The
Melasma Area and Severity Index (MASI) (Gambar 2.6) (Pallavi dkk., 2015). Alat
ukur ini memberikan pengukuran derajat keparahan melasma yang lebih akurat secara
kuantitas. Skor MASI diukur dengan menggunakan pemeriksaan subjektif yang
meliputi 3 faktor: luas area keterlibatan (A), tingkat kegelapan (D), dan Homogenitas
(H). Luas keterlibatan diukur sesuai area yaitu area dahi (f) : 30 %, area malar kanan

(rm) : 30 % , area malar kiri (lm) : 30 %, dan area dagu (c) : 10 %. Skor luas
keterlibatan dikategorikan menjadi 0-6. Skor 0 = tidak ada keterlibatan, 1 = <10%, 2 =
10-29 %, 3 = 30-49 %, 4= 50-69 %, 5 = 70-89 %, dan 6 = 90-100 %. Tingkat
kegelapan dan homogenitas diukur dengan skala 0-4 (0 = tidak ada; 1 = ringan, 2 =
sedang, 3 = jelas terlihat, 4 = maksimal/berat). Homogenitas juga dapat diukur dengan
skala 0-4, yaitu 0 = tidak ada hiperpigmentasi, 1 = terdapat titik-titik hiperpigmentasi
(specks), 2 = area kecil hiperpigmentasi dengan diameter < 1,5 cm, 3 = diameter area
yang terlibat > 2cm, 4 = tidak ada area yang jernih dalam area yang terlibat (Bhor &
Pande, 2006). Skor MASI dihitung dengan menjumlahkan skala keparahan untuk
tingkat kegelapan dan homogenitas, lalu dikalikan dengan nilai area keterlibatan
setiap area wajah. Skor Total MASI = 0,3 A(f) [D(f) + H(f)] + 0,3 A(lm) [D(lm) +
H(lm)] + 0,3A(rm) [D(rm) + H(rm)] + 0,1A(c) [D(c) + H(c)]. Skor total adalah 0-48
(Kimborough dkk., 1994). Penentuan derajat keparahan melasma menurut Ali, dkk
(2013), yaitu derajat ringan 0-16,9 ; derajat sedang 17-32,9 ; dan derajat berat 33-48.
Gambar 2.6.

Melasma Area and Severity Index (MASI) (dikutip dari Pallavi dkk., 2015).
Karena sulitnya untuk menilai tingkat homogenitas lesi melasma, maka
dikembangkan skor MASI modifikasi (tanpa menghitung homogenitas). Skor MASI
modifikasi ini ternyata cukup valid dan reliabel dibandingkan skor MASI
konvensional, bahkan lebih efisien. Skor total MASI modifikasi = 0,3 A(f) D(f) + 0,3
A(lm) D(lm) + 0,3A(rm) D(rm) + 0,1A(c) D(c). Jumlah skor total adalah 0-24.
(Pandya dkk., 2011).
2.1.6 Pemeriksaan Penunjang
Pemeriksaan penunjang dalam menegakkan diagnosis melasma dapat
dilakukan dengan pemeriksaan lampu wood, dermoskopi, serta pemeriksaan
histopatologi. Pemeriksaan lampu wood merupakan pemeriksaan paling
sederhana yang dapat membantu dalam klasifikasi melasma. Pada pemeriksaan
lampu wood akan menujukkan aksentuasi yang semakin jelas pada melasma tipe
epidermal. Sedangkan pada melasma tipe dermal, tidak terdapat aksentuasi pada
penyinaran menggunakan lampu wood. Pada melasma tipe campuran dapat dijumpai
aksentuasi yang semakin jelas dan juga lesi yang tidak terdapat aksentuasi. Beberapa
ahli juga menambahkan tipe 4 yaitu pada individu dengan tipe kulit fitzpatricks V-VI,
dengan kulit yang gelap, lampu wood tidak dapat dievaluasi, karena pada pasien ini,
memiliki melanosit yang banyak dan merata, dimana sebagian besar sinar lampu
wood terserap oleh melanin yang banyak, sehingga tidak menimbulkan aksentuasi
(Tamler dkk., 2009).
Pemeriksaan dermoskopi merupakan teknik noninvasif dimana prinsip kerjanya
menggunakan alat optikal yang memperbesar obyek dari 6-400x. Alat ini terbukti
sebagai alat yang reliabel untuk visualisasi langsung pada kelainan pigmentasi kulit.

Dengan pemeriksaan ini, warna melanin tergantung dari kuantitas, kepadatan, dan
lokasinya. Hitam dengan pigment network yang teratur apabila terletak di stratum
korneum, coklat tua terletak di lapisan epidermis yang lebih bawah, dan biru atau
keabuan dengan pigment network yang tidak teratur apabila terletak di dermis. Tipe
campuran, memiliki gambaran campuran dari kedua tipe di atas (Braun dkk., 2005).
Pemeriksaan histopatologi merupakan pemeriksaan yang paling sensitif
dibandingkan pemeriksaan lainnya, namun kurang disukai, karena bersifat invasif.
Pemeriksaan ini jarang dilakukan karena sebagian besar lesi melasma di wajah,
sedangkan pemeriksaan invasif pada wajah akan menimbulkan banyak
ketidaknyamanan bagi pasien. Pada pemeriksaan histopatologi lesi melasma tipe
epidermal akan tampak deposit pigmen melanin terbatas pada lapisan stratum basal
dan suprabasal, kadang-kadang tersebar sampai ke stratum korneum. Tipe dermal,
akan menunjukkan pigmentasi terdapat di bagian atas dan bawah dermis dengan
infiltrat limfositik perivaskular pada dermis atas (Sarkar dkk., 2014).
2.1.7 Penatalaksanaan Melasma
Penatalaksanaan pada melasma terdiri dari perlindungan terhadap sinar UV,
menghilangkan faktor predisposisi seperti kontrasepsi oral, dan kombinasi antara
terapi topikal dan prosedural (Gambar 2.7). Pada fase intensif, krim kombinasi tripel,
baik formulasi Kligman atau formulasi Kligman modifikasi, digunakan setiap malam.
Apabila perbaikan pada melasma berjalan lambat, dapat dikombinasi dengan terapi
prosedural seperti pengelupasan kimiawi, intense pulse light, dan mikrodermabrasi.
Apabila terjadi perbaikan, maka dilanjutkan dengan terapi pemeliharaan dengan
hidrokuinon (maksimal 6 bulan), asam kojik, niasinamid, dan arbutin. Pada lesi
melasma yang resisten dapat dilakukan terapi laser menggunakan laser ablatif, namun
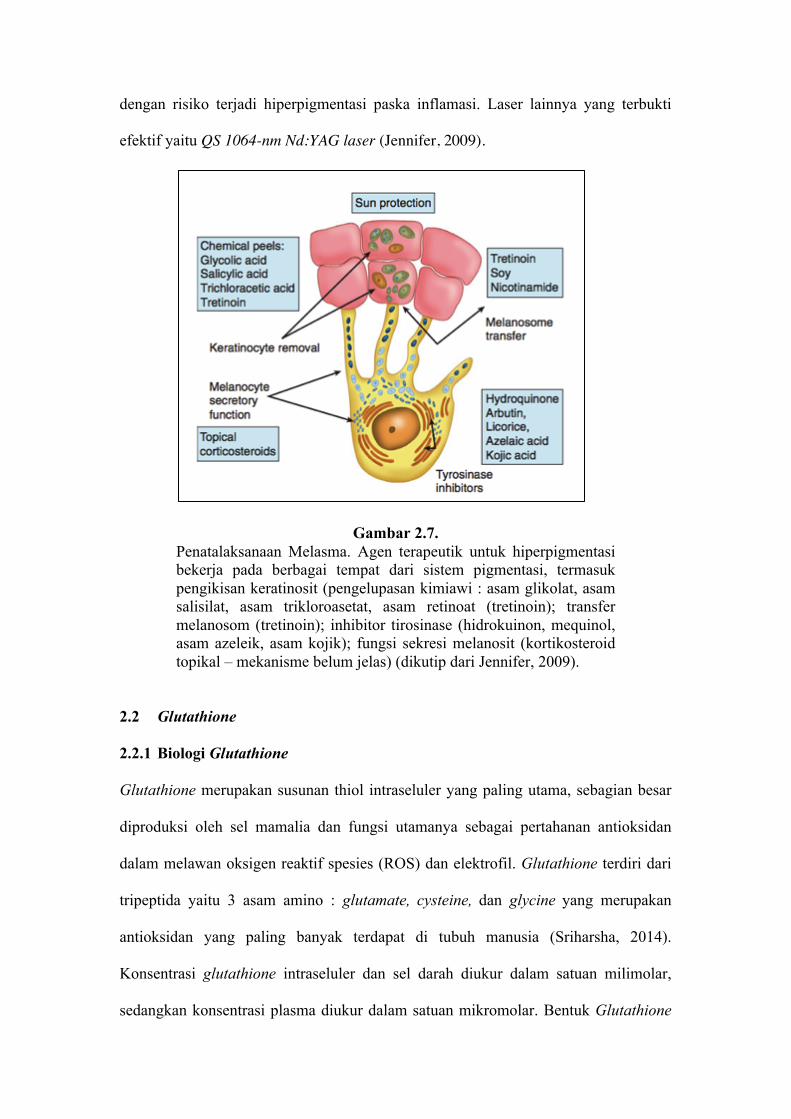
dengan risiko terjadi hiperpigmentasi paska inflamasi. Laser lainnya yang terbukti
efektif yaitu QS 1064-nm Nd:YAG laser (Jennifer, 2009).
Gambar 2.7. Penatalaksanaan Melasma. Agen terapeutik untuk hiperpigmentasi bekerja pada berbagai tempat dari sistem pigmentasi, termasuk pengikisan keratinosit (pengelupasan kimiawi : asam glikolat, asam salisilat, asam trikloroasetat, asam retinoat (tretinoin); transfer melanosom (tretinoin); inhibitor tirosinase (hidrokuinon, mequinol, asam azeleik, asam kojik); fungsi sekresi melanosit (kortikosteroid topikal – mekanisme belum jelas) (dikutip dari Jennifer, 2009).
2.2 Glutathione
2.2.1 Biologi Glutathione
Glutathione merupakan susunan thiol intraseluler yang paling utama, sebagian besar
diproduksi oleh sel mamalia dan fungsi utamanya sebagai pertahanan antioksidan
dalam melawan oksigen reaktif spesies (ROS) dan elektrofil. Glutathione terdiri dari
tripeptida yaitu 3 asam amino : glutamate, cysteine, dan glycine yang merupakan
antioksidan yang paling banyak terdapat di tubuh manusia (Sriharsha, 2014).
Konsentrasi glutathione intraseluler dan sel darah diukur dalam satuan milimolar,
sedangkan konsentrasi plasma diukur dalam satuan mikromolar. Bentuk Glutathione

tripeptida dapat berupa glutathione bebas (GSHt), tereduksi (GSH), teroksidasi
(GSSG), ataupun berikatan dengan protein. Kadar Glutathione sebagian besar dalam
bentuk tereduksi yaitu 98 % dari total glutathione. Konsentrasi normal glutathione
(GSH) pada sel darah orang sehat yaitu sebesar 849 ± 163 µ mol/L untuk individu
berusia 18-73 tahun tanpa memandang jenis kelamin. Sedangkan konsentrasi GSH
plasma yaitu sebesar 3,39 ± 1,04 µ mol/L untuk kelompok umur yang sama (Michelet
dkk., 1995). Penelitian lainnya menunjukkan kadar normal GSH yang berbeda apabila
dilakukan pemeriksaan di darah (whole blood) yaitu 0.684 - 2.525 mmo/L, dan 0.002
- 0.1136 mmol/L. Konsentrasi GSH di dalam plasma ataupun whole blood
dipengaruhi oleh gaya hidup seseorang, yaitu merokok (>10 batang rokok/ hari) dan
konsumsi alkohol (1-50 mg/hari). Kebiasaan merokok akan meningkatkan kadar GSH
sebanyak 26 % dibandingkan individu yang tidak merokok. Kebiasaan minum alkohol
juga akan meningkatkan kadar GSH. Hal ini dikarenakan peningkatan produksi
radikal bebas oleh gaya hidup ini, menyebabkan peningkatan kebutuhan sintesis GSH
oleh tubuh untuk menetralisir radikal bebas ini, sehingga tidak sampai terjadi
kelainan. (Michelet dkk., 1995). Beberapa penyakit kronis juga dapat menyebabkan
penurunan kadar glutathione di dalam tubuh. Penelitian berkelanjutan menunjukkan
beberapa penyakit berhubungan dengan rendahnya kadar glutathione yaitu penyakit
paru obstruktif kronis (terutama emfisema), asma, penyakit hati kronis, sindroma
metabolik, kanker, autisme, cystic fibrosis, psoriasis, vitiligo, alopesia areata,
polymorphic light eruption, akne vulgaris (Sonthalia dkk., 2016). Menurut Perricone
dkk. (2009), kadar GSH juga menurun pada berbagai penyakit degeneratif kronis
lainnya terutama pada penyakit inflamasi dan penyakit yang dimediasi imunologi,
misalnya penyakit alzheimer, parkinson, depresi, artritis rematoid, aterosklerosis,
angina pektoris, hipertensi, stroke, gangguan fungsi tiroid dan pankreas.

2.2.2 Homeostasis Glutathione
Glutathione disintesis secara de novo dalam dua tahap reaksi enzimatik yang
memerlukan ATP. Pada langkah pertama, cysteine dan glutamate terhubung melalui
reaksi yang dikatalisis oleh γ-glutamylcysteine synthase (γ-GCS) membentuk γ-
glutamylcysteine. Reaksi pertama ini merupakan sintesis GSH yang bersifat terbatas
karena tergantung dari ketersediaan cysteine. Sintesis GSH selanjutnya dikatalisis
oleh glutathione synthetase (GS), dalam reaksi dimana γ-glutamyl-cysteine secara
kovalen berikatan dengan glycine. Fungsi antioksidan GSH ditentukan oleh
redoxactive thiol (-SH) dari cysteine yang teroksidasi saat GSH mereduksi molekul
target. Saat terjadi reaksi dengan ROS atau elektrofil, maka GSH akan teroksidasi
membentuk GSSG, yang dapat kembali menjadi bentuk GSH karena tereduksi oleh
GSSG reductase (GR) (Gambar 2.9) (Luschak, 2012). Keseimbangan siklus redoks
daripada glutathione dapat digambarkan pada gambar 2.8 (Sonthalia dkk., 2016).

Gambar 2.8. Homeostasis Glutathione meliputi baik mekanisme intra maupun ekstraseluler. Glutathione disintesis baik secara de novo maupun jalur sintesis alternatif. Sintesis de novo membutuhkan 3 asam amino dan energi dalam bentuk ATP. Glutamate mungkin tersedia dalam bentuk konversi dari γ-glutamyl amino acid menjadi 5-oxoproline, yang kemudian diubah menjadi glutamate. Dua molekul ATP digunakan untuk biosintesis satu molekul GSH. Sintesis alternatif meliputi baik reduksi dari GSSG atau penggunaan prekursor yang dibentuk dari hidrolisis GSH atau konjugatnya oleh γ-L-glutamyl transpeptidase pada permukaan eksternan plasma membran yang dikirim kembali ke sel dalam bentuk asam amino atau dipeptida. GSH terdegradasi dalam berbagai proses. Dalam hal detoksifikasi reaktif spesies dan elektrofil seperti methyglyoxal, GSH terlibat dalam glutathionylation protein dan beberapa proses lainnya, seperti biosintesis leukotrien dan prostaglandin, dan reduksi ribonukleotida (dikutip dari Lushchak, 2012).

Gambar 2.9. Siklus redoks Glutathione menggambarkan perubahan glutathione teroksidasi dan
tereduksi (Sonthalia dkk., 2016).
2.2.3 Fungsi Glutathione
Glutathione merupakan protein yang secara alami diproduksi di dalam tubuh dan
memiliki 3 fungsi perlindungan yang esensial yaitu : sebagai antioksidan,
meningkatkan sistem imunitas, dan sebagai detoksifikasi. Selain itu glutathione
memiliki fungsi tambahan pada sel yaitu : sebagai cadangan cysteine, menyimpan dan
menghantarkan nitric oxide, berpartisipasi dalam metabolisme estrogen, leukotrien,
dan prostaglandin, serta mereduksi ribonukleotida menjadi deoksiribonukleotida,
maturasi kelompok iron-sulfur dari protein yang berbeda, terlibat dalam proses faktor
transkripsi tertentu (khususnya yang terlibat dalam sinyal redoks), dan detoksifikasi
berbagai susunan endogenus dan xenobiotik (Zhang & Forman, 2009).
2.2.4 Glutathione dalam Eliminasi Oksigen Reaktif dan Nitrogen Spesies.

Glutathione merupakan antioksidan yang sangat penting, yang bereaksi secara
langsung dengan ROS, RNS, dan spesies reaktif lainnya khususnya HO�, HOCl, RO�,
RO2�, 1O2, dan ONOO- (Gambar 2.10). Glutathione juga berperan dalam detoksifikasi
produk dari oksidasi lipid oleh ROS seperti Malonic dialdehyde (MDA) dan 4-
hydroxy 2-nonenal, dan mungkin juga produk interaksi ROS dengan komponen
seluler lainnya (Luschack, 2012).
Gambar 2.10. Keterlibatan Glutathione dalam eliminasi ROS dan RNS. Radikal hidroksil dan nitric oxide (setelah oksidasi menjadi bentuk NO+) atau peroxynitrite dapat berinteraksi secara langsung dengan GSH membentuk GSSG. Hidrogen peroksida dapat dipecah oleh katalase atau oleh glutathione peroxidase (Gpx). Selanjutnya akan membutuhkan GSH untuk mengurangi peroksida (Luschack, 2012). 2.2.5 Peranan Glutathione dalam Patogenesis Melasma
Peran glutathione pada patogenesis melasma dikatakan melalui beberapa mekanisme
yaitu baik berpengaruh secara langsung maupun tidak langsung pada aktivitas enzim
tirosinase. Tirosinase secara langsung dihambat melalui chelation copper site oleh
grup thiol pada GSH. Kedua GSH menghambat transfer seluler tirosinase ke

premelanosome, yang merupakan tempat awal untuk sistesis melanin. Ketiga, inhibisi
tirosinase secara tidak langsung didapatkan melalui efek antioksidan dari GSH.
Glutathione mengubah melanogenesis dari sintesis eumelanin menjadi phaemelanin
melalui reaksi antara grup thiol dan dopaquinone membentuk konjugat sulfhydryl
dopa (Sonthalia dkk., 2016).
Pada proses biosintesis melanin, dikatakan bahwa pada saat terdapat GSH
ataupun cysteine, maka akan terjadi ikatan dengan DOPAquinone dan mengalihkan
sintesis pigmen dari eumelanin menuju ke pheomelanin melalui cysteinylDOPA
(Gambar 2.11 dan 2.12) (Ito & Wakamatsu, 2008 ; Sonthalia, 2016).
Gambar 2.11. Biosintesis Melanin. Pembentukan Pheomelanin
(dikutip dari Ito & Wakamatsu, 2008).
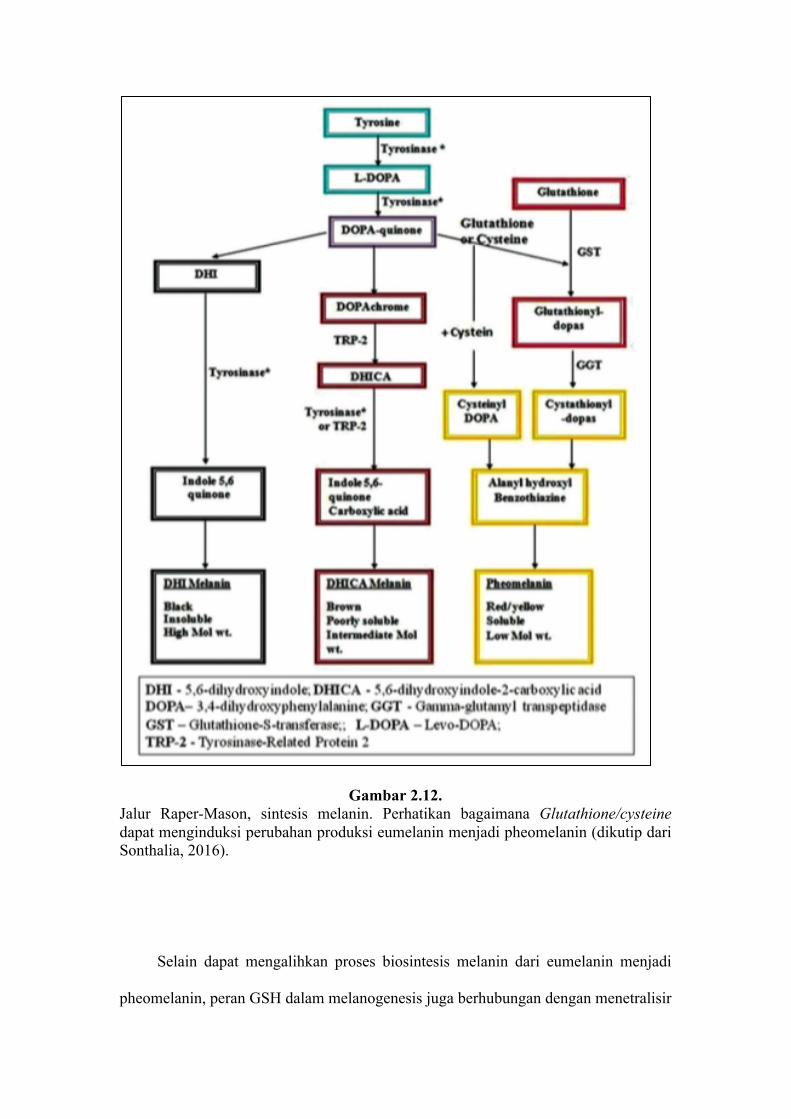
Gambar 2.12. Jalur Raper-Mason, sintesis melanin. Perhatikan bagaimana Glutathione/cysteine dapat menginduksi perubahan produksi eumelanin menjadi pheomelanin (dikutip dari Sonthalia, 2016).
Selain dapat mengalihkan proses biosintesis melanin dari eumelanin menjadi
pheomelanin, peran GSH dalam melanogenesis juga berhubungan dengan menetralisir

stres oksidatif, seperti yang digambarkan pada gambar 2.9. Produksi melanin, karena
adanya radiasi ultraviolet menyebabkan terbentuknya ROS yaitu H2O2, radikal
hidroksil dan radikal superoksida (O2�−) pada melanosit. Oksidatif ini sebagai
perantara quinones reaktif, yang bersifat sitotoksik terhadap protein dan DNA di
dalam sel. Akumulasi H2O2 pada melanosit normal dapat mempengaruhi sintesis
melanin. Sementara melanin dapat memberikan efek fotoprotektif, situasi ini menjadi
kompleks karena status antioksidan dan prooksidan tampak terdapat pada status
redoks di melanosit. Saat produksi melanin itu sendiri dapat menjadi sumber utama
pembentukan ROS, di pihak lain, keberadaan ROS dapat menyebabkan
melanogenesis yang abnormal termasuk overproduksi melanin. Penelitian-penelitian
sebelumnya telah menunjukkan bahwa peningkatan pembentukan ROS dapat terjadi
akibat overproduksi prooksidan di dalam sel, dan atau penurunan kapasitas
antioksidan seluler. Pembentukan ROS dapat diproduksi oleh mitokondria dan
peroksisom ketika proses metabolik seluler normal. Produksi ROS akan meningkat
saat terjadi kondisi patologis seperti inflamasi dan kanker, serta paparan terhadap
faktor lingkungan seperti bahan kimia dan radiasi UV (Gambar 2.13) (Denat dkk.,
2014). Kulit merupakan organ terluas yang berhadapan langsung dengan lingkungan,
sehingga merupakan sumber utama dari ROS yang diinduksi radiasi UV.
Pembentukan ROS/RNS yang diinduksi radiasi UV berhubungan dengan peningkatan
melanogenesis melalui peningkatan aktivitas tirosinase, protein dan mRNA dalam
melanosit. Pembentukan H2O2 atau NO dan kerusakan oksidatif juga dapat
meningkatkan produksi melanin dalam melanosit melalui aktivasi α MSH/Mc1r atau
jalur sinyal MITF, yang penting dalam proses melanogenesis (Dong dkk., 2010).

Gambar 2.13. Sumber pembentukan ROS dari eksogenus dan endogenus. Adanya pertahanan antioksidan baik itu yang bersifat nonenzimatik (diantaranya glutathione) atau enzimatik dapat menjaga kondisi melanosit tetap dalam status normal (dikutip dari Denat, 2014).
Antioksidan endogenus kulit termasuk GSH, memiliki potensi dalam menjaga
keseimbangan redoks dan juga sebagai sumber substrat untuk Gpx atau glutathione S-
transferase (GST), serta antioksidan enzimatik seperti katalase (CAT) dan Gpx
berperanan dalam netralisasi H2O2. Apabila terjadi penurunan antioksidan GSH dapat
menyebabkan terjadinya ketidakseimbangan status redoks yang nantinya akan
menyebabkan kerusakan oksidatif pada DNA, lipid dan protein, produksi produk
fotoreaksi yang menyebabkan fotooksidasi melanin dan gangguan sinyal sel (jalur
melanogenesis). Kesemuanya ini dapat menyebabkan gangguan pigmentasi (Gambar
2.14) (Panich, 2011).

Gambar 2.14. Radiasi UV memicu stres oksidatif atau penurunan antioksidan dapat memicu ketidakseimbangan status redoks yang akan menyebabkan gangguan pigmentasi (dikutip dari Panich, 2011).
Peran GSH dalam patogenesis melasma telah diteliti oleh beberapa peneliti.
Shuja dkk. (2005) mendapatkan penurunan jumlah GSH darah yang signifikan pada
penderita melasma dibandingkan dengan kontrol. Pada penelitian yang dilakukan oleh
Hamadi dkk. (2009), yang membandingkan peranan melatonin pada terapi melasma,
bahwa didapatkan GSH serum pada penderita melasma lebih rendah dibandingkan
kontrol dan kadar ini meningkat setelah diberikan perlakuan berupa melatonin baik
topikal maupun oral. Hal serupa juga didapatkan oleh Katiyar (2014) yang meneliti
efikasi terapi kombinasi pada pasien melasma dengan menggunakan kadar
antioksidan sebagai markernya. Pada penelitiannya, dijumpai kadar GSH darah yang
menurun secara signifikan pada penderita melasma apabila dibandingkan dengan
kontrol sebelum diberikannya terapi kombinasi.
Menurut Villarama & Maibach (2005) Glutathione merupakan senyawa yang
poten sebagai agen pemutih; dimana cara kerjanya dengan inaktivasi langsung

daripada enzim tirosinase; melalui mekanisme peralihan dari produksi eumelanin
menjadi pheomelanin; melalui reduksi dari radikal bebas dan peroksida; dan dengan
memodulasi kemampuan depigmentasi dari agen melanositotoksik. Hal serupa juga
didukung oleh Arjinpathana & Asawanoda (2012).
Kesimpulan yang dapat diambil dari penelitian-penelitian tersebut adalah GSH
merupakan antioksidan poten nonenzimatik yang sangat berperan dalam proses
patogenesis melasma. Adanya penurunan kadar GSH pada penderita melasma,
membuka kemungkinan dikembangkannya lebih banyak lagi penelitian mengenai
efektivitas terapi dengan menggunakan glutathione pada penderita melasma.




















