Anatomi Dan Fisisologi Pendengaran
39
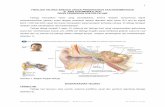
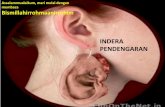
ANATOMI DAN FISIOLOGI PENDENGARAN Bab ini membahas sejumlah anatomi dan fisiologi alat-alat dalam telinga yang paling mendasar. Telinga dibagi menjadi telinga luar dan telinga tengah, koklea, dan sistem syaraf pusat. Fokus pembahasan dari bab ini adalah mengenai anatomi dan fisiologi pendengaran yang tergantung pada alat fungsionalnya. Anatomi pembedahan, vasculature dan fungsi dari tuba eustachius tidak ikut dibahas dalam bab ini. TELINGA LUAR Telinga luar terdiri dari daun telinga dan liang telinga, mulai dari meatus sampai ke membran timpani. Daun telinga pada manusia sebagian besar terdiri dari kartilago dan tidak memiliki otot sama sekali. Pada bagian tengah daun telinga, yaitu konka, terdapat saluran telinga luar yang memiliki panjang 2,5 cm. Sepertiga lateral saluran telinga luar dibentuk dari jaringan kartilago, yang juga mengandung kelenjar penghasil serumen dan folikel rambut. Duapertiga medial saluran telinga luar dibentuk dari tulang, yang dibatasi oleh sel epitel membran timpani. Telinga luar dan kepala bersifat pasif namun memiliki peranan penting pada sistem pendengaran karena memiliki alat akustik. Konka atau mangkuk aurikula
-
Upload
mulyati-sari -
Category
Documents
-
view
181 -
download
10
Transcript of Anatomi Dan Fisisologi Pendengaran

ANATOMI DAN FISIOLOGI PENDENGARAN
Bab ini membahas sejumlah anatomi dan fisiologi alat-alat dalam telinga
yang paling mendasar. Telinga dibagi menjadi telinga luar dan telinga tengah,
koklea, dan sistem syaraf pusat. Fokus pembahasan dari bab ini adalah mengenai
anatomi dan fisiologi pendengaran yang tergantung pada alat fungsionalnya.
Anatomi pembedahan, vasculature dan fungsi dari tuba eustachius tidak ikut
dibahas dalam bab ini.
TELINGA LUAR
Telinga luar terdiri dari daun telinga dan liang telinga, mulai dari meatus
sampai ke membran timpani. Daun telinga pada manusia sebagian besar terdiri
dari kartilago dan tidak memiliki otot sama sekali. Pada bagian tengah daun
telinga, yaitu konka, terdapat saluran telinga luar yang memiliki panjang 2,5 cm.
Sepertiga lateral saluran telinga luar dibentuk dari jaringan kartilago, yang juga
mengandung kelenjar penghasil serumen dan folikel rambut. Duapertiga medial
saluran telinga luar dibentuk dari tulang, yang dibatasi oleh sel epitel membran
timpani.
Telinga luar dan kepala bersifat pasif namun memiliki peranan penting
pada sistem pendengaran karena memiliki alat akustik. Konka atau mangkuk
aurikula memiliki kemampuan resonansi sebesar 5 kHz, dan permukaan aurikula
yang tidak teratur memilki kegunaan sebagai resonansi dan antiresonansi. Alat-
alat akustik berfungsi untuk menentukan sumber suara yang baik yang berada di
depan maupun di belakang pendengar.
Saluran telinga luar sangat penting, berupa tabung yang terbuka di satu sisi
dan tertutup di sisi lainnya, dan memiliki kemampuan resonansi gelombang suara
seperempatnya. Frekuensi resonansi (f0) ditentukan oleh panjang tuba dan
berbanding lurus dengan kurva. Jika panjang tuba 2,5 cm, maka frekuensi
resonansi berkisar 3,5 kHz.
F0 = velocity suara @ 350 m/dtk/(4x2,5 cm)

Gabungan dari berbagai suara pada lapangan pendengaran dapat berubah-
ubah sesuai dengan alat akustik seperti kepala dan telinga luar. Pada Gambar
129.2 ditunjukkan sekitar 15 dB suara terjadi dalam rata-rata 3 kHz pada manusia,
kucing dan tikus, dan sekitar 10 dB antara 2-5 kHz. Gangguan alat-alat akustik
pada telinga luar merupakan salah satu penyebab tuli akibat indera penciuman dan
terlihat sangat menonjol pada daerah frekuensi 4 kHz.
Alat-alat akustik seperti kepala dan telinga luar memiliki peranan penting
dalam fungsi pendengaran; pada tuli yang disebabkan karena indera penciuman
pada frekuensi 4 kHz. Dalam menentukan letak sumber suara, kepala bekerja
sebagai atenuasi pada frekuensi dimana lebar kepala lebih besar daripada panjang
gelombang suara. Jika frekuensi lebih besar dari 2 kHz, kepala akan memberikan
efek bayang-bayang, dimana perbedaan intensitas dalam telinga berkisar antara 5
hingga 15 dB digunakan untuk menentukan letak sumber suara. Pada frekuensi
yang lebih rendah, dimana panjang gelombang suara lebih besar daripada lebar
kepala, sedikit atenuasi akan dibutuhkan oleh kepala. Perbedaan waktu dalam
telinga (~0,6 mdetik bagi suara untuk melewati kepala) adalah pedoman penting
untuk menentukan letak sumber suara. Efek bayang-bayang kepala merupakan
alasan pemburu tangan kanan untuk menggunakan senapan dan senjata pada
telinga kiri yang tuli dari pada telinga kanannya dan sebaliknya. Moncong senjata,
dimana energi akustiknya sangat besar, akan menutup telinga kiri dan telinga
kanannya dilindungi dengan efek bayangan kepala.
Suara 10-15 dB yang diterima oleh telinga luar pada daerah 3-5 kHz
sangat berguna untuk mendeteksi dan membedakan energi rendah, suara frekuensi
tinggi seperti suara desahan. Pentingnya alat-alat akustik pada telinga luar dan
kepala direfleksikan pada susunan tambahan pendengaran dan evaluasi. Resonansi
pada saluran luar hampir mendekati 8 kHz pada bayi dan menurun pada dewasa
setelah berusia 2,5 tahun. Perkembangan alat-alat ini memiliki beberapa implikasi
klinis, terutama untuk pemeriksaan lapangan pendengaran dan mengatur alat
pendengar tambahan dan pemeriksaan pada bayi.

TELINGA TENGAH
Perpindahan energi akustik telinga tengah terjadi dari saluran telinga luar
yang berisi udara ke koklea yang berisi cairan. Fungsinya sebagai alat penyesuai
impedansi karena sebagai pasangannya pada udara dengan impedansi rendah ke
koklea yang berisi cairan dengan impedansi tinggi. Penyesuaian impedansi ini
meliputi tiga faktor. Yang pertama dan faktor terpenting sebagai daerah penggetar
yang paling efektif pada membran timpani hampir mendekati 17 hingga 20 kali
lebih besar dari pada daerah penggetar di footplate. Faktor kedua sebagai tuas
rangkaian osikular. Incus lebih pendek bila dibandingkan dengan panjang
manubrium dan leher malleus. Faktor ketiga dan merupakan faktor terkecil,
adalah bentuk dari membran timphani. Gabungan ketiga faktor ini menekan
hampir sebesar 25-30 dB. Pengecualian pada penelitian impedansi akustik telinga,
data yang lebih banyak dari penelitian yang ditemukan pada mayat manusia,
dengan segala konsekuensinya, atau pada binatang, biasanya kucing. Membran
telinga berfungsi melindungi ruang telinga tengah dari benda asing pada saluran
telinga dan terutama mencegah insuflasi benda asing dari nasofaring melalui tuba
eustachius.
Getaran rangkaian dari tulang-tulang osikular ditunjukkan pada Gambar
129.3. Gerakan perpindahan dari membran timpani dan tulang-tulang rangkaian
osikular menyebabkan perpindahan kekuatan yang relatif efisien pada telinga
dalam dan keakuratan transmisi suara melewati telinga tegah sangat tinggi.
Distorsi sinyal suara tidak terjadi pada telinga tengah, bahkan untuk sinyal yang
datang dengan tingkat suara yang lebih besar dari 130 dB sound pressure level
(SPL).
Telinga tengah, meliputi membran timphani, rangkaian tulang-tulang
osikular ligamen-ligamen penyangga, dan ruang telinga tengah dapat dianggap
sebagai sistem gerakan pasif dengan kedua massa dan elemen-elemen pelengkap
dan alat-alat penguat resonansi. Sistem ini diteruskan hingga ke koklea, yang
berperan untuk memperbesar resistensi. Hasilnya adalah, telinga tengah lebih
lembab dan seimbang dan memilki frekuensi respon yang luas. Fungsi keluar-
masuk atau fungsi transfer telinga tengah ditunjukkan pada gambar 129.4a.

Perbandingan kecepatan volume stapes menjadi tekanan suara pada membran
timphani meningkat hingga 800-900 Hz pada manusia, dimana resonansi pada
telinga tengah ,dan penurunan pada frekuensi yang lebih tinggi. Fase perubahan
atau waktu kelambatan antara gerakan membran timphani dan stapes umumnya
meningkat sesuai dengan frekuensi (Gambar 129.4b). Meskipun telinga tengah
merupakan sistem yang luar biasa dalam merespon frekuensi, menyetarakan dan
dan sebagai alat transformasi, kurang dari setengah kekuatan suara yang
memasuki telinga tengah yang mencapai koklea karena energi suara diserap oleh
ligamen dan telinga tengah. Seperti yang ditunjukkan pada gambar 129.5, telinga
tengah manusia sebagian tidak efisien pada frekuensi yang lebih dari 2 kHz,
terutama bila dibandingkan dengan telinga kucing dan tikus. Hal ini juga penting
untuk mengingat bahwa hilangnya kekuatan sebanyak 50% berarti hanya
kehilangan sebanyak 3 dB. Fungsi pendengaran sangat dipengaruhi oleh
impedansi koklea, sama halnya seperti kombinasi efek akustik dari kepala, telinga
luar dan telinga tengah. Efek kombinasi dari alat-alat akustik seperti kepala,
telinga luar dan telinga tengah sama baiknya dengan impedansi yang masuk pada
koklea, memiliki efek yang sangat penting pada fungsi pendengaran. Sebagai
contoh, faktor-faktor berikut menentukan bentuk kurva audibility dan tingkat
frekuensi pendengaran manusia. (Gambar 129.6) Sebagai contoh, manusia tidak
dapat mendeteksi suara yang lebih besar atau mendekati 20 kHz karena suara
berfrekuensi tinggi seperti ini tidak dapat ditransmisikan secara efisien melalui
telinga tengah ke kolea. Contoh kedua dari transmisi suara ini ditunjukkan pada
gambar 129.7, dimana spektrum ukuran meriam pada lapangan suara
dibandingkan dengan meriam pada waktu ditransformasikan dan dibentuk oleh
alat-alat akustik seperti telinga luar, kepala, telinga tengah dan impedansi
masukan pada oklea. Energi berfrekuensi rendah tidak ditransmisikan ke dalam
koklea, dan daerah frekuensi tingkat energi terbesar antara 3 hingga 4 kHz. Alat-
alat akustik ini bertanggung jawab pada intensitas suara berfrekuensi rendah
(ukuran lapangan suara) untuk menghasilkan tuli berfrekuensi tinggi dan trauma
di daerah dasar koklea.

Dua otot lurik, musculus tensor timphani dan stapedius terletak pada
telinga tengah. Otot ini melekat pada malleus dan dipersyarafi oleh nervus
Trigeminus. Otot stapedius melekat pada stapes dan dipersyarafi oleh nervus
fasialis cabang stapedial. Otot stapedius dan tensor timphani merupakan otot lurik
paling kecil di tubuh manusia dan juga memiliki perbandingan inervasi yang
tinggi, karena satu serabut syaraf memperyarafi satu serabut otot. Kontraksi otot-
otot ini tidak mempengaruhi transmisi suara yang melewati telinga tengah,
pengaruh secara rinci dan tingkat pengaruhnya pada otot telinga tengah masih
belum sepenuhnya dimengerti. Sejumlah fungsi yang berbeda dikaitkan dengan
otot telinga tengah.
Salah satu fungsi otot telinga tengah adalah untuk melindungi koklea dari
suara yang keras. Bila suara lebih besar atau hampir 80 dB SPL didengar
monoaural atau binaural, akan terjadi refleks kontraksi otot stapedius konsensual
(bilateral). Kontraksi ini meningkatkan kekakuan rangkaian tulang osikular dan
membran timphani, mengurangi suara hingga mencapai 2 kHz. Walaupun
kontraksi tensor timphani merupakan sebagian respon yang tiba-tiba, data reflek
akustik dari manusia sendiri dengan pengaruh lingkungan syaraf nervus cranialis
V dan VII tidak mempengaruhi respon normal terhadap rangsang akustik.
Penelitian di lapangan dan berdasarkan tes laboratorium, tuli akibat bising
ditunjukkan membuktikan bahwa reflek stapedial melindungi koklea, dari suara
berfrekunsi rendah (<2 kHz) yang melebihi 90 dB. Karena reflek akustik lebih
dari 10 ms yang terus menerus, koklea bisa saja tidak terlindungi selama durasi
yang singkat, tidak dapat mengantisipasi suara yang impulsif.
Fungsi-fungsi berikut ini dikaitkan dengan otot-otot pada telinga tengah.
Beberapa fungsinya antara lain adalah untuk memperkuat dan mengkakukan
rangkaian tulang-tulang ossikular, berperan dalam menyuplai darah ke rangkaian
tulang-tulang ossikular, mengurangi fisiologi kebisingan yang disebabkan oleh
gerakan menguyah dan bersuara, memperbaiki rasio sinyal bising menjadi sinyal
berfrekuensi tinggi, terutama pada suara pembicaraan frekuensi tinggi seperti
suara desahan, yang diartikan sebagai atenuasi tingkat tinggi, bising frekuensi
rendah, berfungsi sebagai kontrol tambahan otomatis dan meningkatkan tingkat

dinamika telinga, dan melembutkan ketidakteraturan fungsi transfer telinga
tengah.
KOKLEA
Koklea manusia bergelung, panjang tabung tulangnya sekitar 35 mm, terbagi
menjadi skala vestibuli, skala media, dan skala timphani (gambar 129.8). skala
vestibuli dan skala timphani berisi cairan perilimphe, suatu cairan ekstraseluler
yang berisi Kalium dengan konsentrasi 4 mEq/L dan natrium dengan konsentrasi
139 mEq/L. Skala media dibatasi oleh membran Reissner, suatu membran basilar
dan lembaran tulang spiral, dan dinding lateral. Skala media berisi cairan
endolimphe, suatu cairan intraselular dengan konsetrasi Kalium 144 mEq/L dan
konsentrasi Natrium 13 mEq/L. Skala media memilki aliran langsung positif (DC)
yang pada potensial istirahatnya mencapai 80 mV yang sedikit berkurang dari
dasar hingga ke apeks. Potensial endokoklear ini dihasilkan oleh lipatan
vaskularisasi yang banyak sekali dari dinding lateral koklea. Pompa Natrium-
Kalim-Adenosine Triphosphatase (Na+-K+-ATPase) pada beberapa sel tertentu
dilipatan vaskularisasi berperan terhadap potensial ini.
Energi akustik masuk ke dalam koklea melalui gerakan seperti penghisap
pada footplate stapes pada foramen window dan diiringi dengan gerakan
perilimphe di dalam skala vestibuli. Cairan perilimphe di dalam skala vestibuli
berhubungan dengan cairan perilimphe di skala timphani melalui saluran kecil
pada apeks koklea yang disebut dengan helikotrema. Organ Corti terletak di
membran basilar dan di lapisan tulang spiral. (Gambar 129.9). Panjang dasar dari
membran basilar mencapai ukuran 0,12 mm dan panjang apeksnya mencapai
ukuran 0,5 mm. Komponen utama dari organ Corti ini adalah sel rambut dalam
dan sel rambut luar, sel penyangga (Deiters, Hensen, Claudius), membran tektoria
dan kompleks lamina cuticular-reticular (Gambar 129.10). Sel penyanga
memberikan bentuk dan membantu proses metabolisme organ Corti. Proses
penulangan sel Deiters menjadi pertemuan sel-sel yang rapat sari sel lamina
retikular.

Sel-sel rambut dalam dan sel-sel rambut luar dari organ Corti berperan
penting dalam proses transduksi energi gerak (keseimbangan) menjadi energi
listrik (syaraf). Sel rambut luar sangat berbeda dengan sel rambut dalam. Gambar
129.11 dan tabel 129.1 menunjukkan perbedaannya. Perbedaan morfologi antara
sel rambut luar dan sel rambut dalam terletak pada inervasi syaraf (Gambar
129.12). Ganglion spiral, badan sel saraf pendengaran, mengirimkan akson ke
nukeus koklea di batang otak, dimana dendrit-dendrit melewati lamina tulang
spiral. Dari 50.000 syaraf yang menginervasi koklea, 90%-95% bersinaps
langsung pada sel rambut dalam. Syaraf ini disebut neuron tipe I. Masing-masing
sel rambut dalam diinervasi oleh hampir 15-20 neuron tipe I. Sebaliknya, 5%-10%
dari 50.000 neuron menginervasi sel-sel rambut luar (neuron tipe II). Masing-
masing neuron tipe II bercabang untuk menginervasi sekitar 10 sel rambut luar.
Pola masuknya persyarafan ke dalam koklea yaitu, mencapai 1.800 serabut syaraf
yang keluar, berasal dari kompleks olivarius superior ipsilateral dan kolateral,
masuk ke dalam koklea.
Proses transduksi diinisiasi oleh pergerakan membran basilar sebagai
respon terhadap pergerakan stapes akibat proses akustik. Pola pergerakan
membran basilar seperti gelombang datang (Gambar 129.14). Membran basilar
bagian basal lebih kaku dibandingkan dengan bagian apeks. Gelombang datang
selalu bergerak dari arah basal menuju apeks. Amplitudo maksimal pada gerakan
membran basilar bervariasi sesuai fungsi frekuensi stimulus. Gelombang datang
dihasilkan oleh suara berfrekuensi tinggi (10 kHz) yang bergerak maksimal di
dekat dasar koklea, sedangkan gelombang suara berfrekuensi rendah (125 Hz)
memiliki gerakan maksimal di daerah apeks. Gelombang datang membangkitkan
suara berfrekunsi tinggi yang tidak dapat mencapai daerah apeks koklea,
sedangkan gelombang suara berfrekuensi rendah dapat sampai di sepanjang
membran basilar.
Dulu, gerakan gelombang datang dianggap sebagai respon penyesuaian,
dengan pengenalan penyesuaian yang baik oleh transduksi, syaraf pendengaran
dan CNS. Berdasarkan data yang diperoleh, ditunjukkan bahwa gelombang datang
memiliki respon penyesuaian yang sangat baik, (Gambar 129.15) dan memiliki

kemampuan memilih frekunsi yang layak bagi telinga yang dapat dijelaskan oleh
alat-alat di dalam koklea.
Mekanisme dimana puncak penyesuaian yang baik di bangkitkan oleh
perubahan gerakan gelombang datang yang disebut sebagai cochlear modifier. Ini
adalah sebuah aktivitas sel-sel rambut luar yang mengubah frekuensi gerakan
membran basilar mendekati lokasi frekuensi terbaik dari alat-alat dalam koklea.
Perubahan ini berfungsi menyesuaikan kemampuan memilih dan sensitiv terhadap
frekuensi yang layak bagi telinga dan mampu mendeteksi suara yang sangat
redup. Tujuan proses aktif di dalam koklea, amplifier koklea, didukung oleh
fenomena otoacoustic emissions. Itulah mengapa, jika sinyal berdurasi singkat
diberikan kepada telinga, keluar gema dari koklea yang dapat direkam di saluran
tellinga luar. Karena energi gema lebih besar daripada energi sinyal berdurasi
sigkat, terjadilah proses aktif, amplifier koklea mulai bekerja. Faktor-faktor yang
berperan terhadap amplifier koklea antara lain gerakan sel-sel rambut luar dan
alat-alat gerak stereosilia dan membran tektoria.
Kompleks sel rambut stereosilia masih diragukan peranannya dalam
proses transduksi. Stereosilia merupakan bundle filamen aktin yang berbentuk
saluran dan masuk ke dalam piringan kutikular. Stereosilia ini juga saling
bersilangan. Stereosilia sel rambut dalam tidak berhubungan dengan membran
tektoria, tetapi sel rambut luar berhubungan secara langsung. Defleksi stereosilia
oleh gerakan pembukaan gelombang datang dan menutupnya channel ion non-
spesifik pada ujung stereosilia, menghasilkan aliran arus (Kalium) ke dalam sel
sensorik. Aliran ion Kalium ke dalam sel sensori dimodulasi oleh terbukanya dan
tertutupnya chanel ion stereosilia. Aliran ion Kalium ini disebabkan oleh potensial
endokoklear +80 mV yang ditambahkan ke dalam potensial negatif intrasel
rambut. Depolarisasi intraseluler ini menyebabkan aliran enzim, termasuk
Kalsium. Hal ini menyebabkan pelepasan transmiter kimia dan karena aktivasi
masuknya serabut saraf.
Walaupun kekakuan koklea lebih aktif daripada organ aktif yang lain tidak
lagi diperdebatkan, tetapi amplifier kolea dan cara kerja dasar biologis masih
dalam penelitian. Salah satu peranan amplifier koklea adalah kemampuan sel

rambutya untuk berkontraksi dan memanjang sebagai respon terhadap sinyal
listrik yang disebut sebgai somatic electromotility. Suatu protein yang disebut
prestin telah ditemukan di dalam sel rambut luar dan dianggap sebagai protein
penggerak sel rambut luar dan sebagai pengendali gerakan listrik sel rambut.
Fokus bahasan lainnya adalah aksi channel ion Kalium dan Kalsium yang terjadi
di basal amplifier koklea serta regulasinya. Yang ketiga, menganggap bahwa
kumpulan protein-protein pengerak antara sel-sel rambut dapat membangkitkan
proses osilasi alat-alat elastis sel lainnya. Yang terakhir, mengenai aksi channel
Kalsium. Spesifikasi dasar biologi amplifier koklea sangat penting pada tuli akibat
gangguan pada amplifier koklea.
Neurotransmiter sistem afferent dan efferent merupakan pokok bahasan
dalam penelitian ini. Sebagai sistem afferent, analisa exitatori reseptor asam
amino diekspresikan oleh reaksi rantai polymerase-transkiptase, hibridisasi, dan
analisa immunokimiawi menyatakan bahwa glutamat adalah neurotransmitter
afferent. Glutamat ditemukan di sel ganglion spiral dan sel sensori. Substansi
transmitter utama serat efferent koklea adalah asetilkolin. Yang memungkinkan
organ Corti memodifikasi gerakan dengan mengubah gerakan sel rambut luar
dibawah pengaruh sistem efferent. Fungsi asetilkolin pada reseptor adalah untuk
menghasilkan hiperpolarisasi membran sel dan menggandakan konduksi input sel.
Reseptor asetilkolin terdiri dari muskarinik dan nikotinik. Neurotransmitter lain
pada sistem efferent yaitu asam γ-aminobutirat dan beberapa peptida neuroaktif
lainnya.
POTENSIAL BESAR KOKLEA
Empat potensial besar (ekstraseluler) yang terdapat di dalam koklea; potensial
endolimphatik (endokoklea), mikrofonik koklea, jumlah potensial, dan potensial
aksi syaraf (Gambar 129.16). Tidak seperti potensial koklea lainnya, potensial
endolimphatik tidak membangkitkan respon rangsang akustik, tetapi memiliki
potensial DC 80-100 mV pada skala media. Ini berasal dari striae vaskularis
dinding lateral koklea. Striae vascularis ini dianggap sebagai sumber energi atau
“baterai” koklea, merupakan bagian yang penting sekali dalam proses transduksi.

Sumber energi alami ini berhubungan erat dengan vaskulatur striae vaskularis dan
Na+-K+-adenosintrifosfat (ATPase). Pompa ini terbagi menjadi beberapa jenis sel
koklea, antara lain; sel kecil striae vascularis, sel sulkus luar, dan fibrosit yang
melekat pada membrana Reissner dan ligamnetum spiral. Masih dilakukan
mengenai asal sumber energi dan hal-hal khusus mengenai pertukaran ion,
sedangkan NA-K ATPase telah diketahui memiliki peranan yang penting dalam
transportasi ion pada koklea.
Tidak berfungsinya mekanisme yang terlibat dalam produksi endolimfe
dan potensial endolimfatik dapat menyebabkan penurunan pendengaran yang
disebut sebgai metabolic presbycusis. Ketika aliran endolimfe ke ductus reuniens
terhambat akan terjadi peningkatan tekanan dari endolimfatik dan terjadilah
hydrops.
Tabel 129.1
STRUKTUR DAN INERVASI SEL-SEL RAMBUT DALAM DAN SEL-SEL
RAMBUT LUAR
Karakteristik Sel-sel rambut dalam Sel-sel rambut luar
Jumlah 3.500 12.000
Bentuk Seperti botol/labu Silinder
Stereosilia
Jumlah sel rambut Sedikit Banyak
Susunan 3 atau 4 baris, barisan
berbentuk seperti kurva
landai
6 atau 7 baris, barisan
berbentuk seperti huruf V
atau W
Perlekatan dengan
membran tektorial
Tidak melekat sama
sekali
Melekat dengan erat
stereosilia yang paling
panjang
Ultrastruktur
Letak nukleus badan sel Di tengah Di dasar
Organ sitoplasmik Tersebar Berdekatan dengan

membran sel
Spesialisasi presipnatik Banyak Sedikit atau tidak ada
sama sekali
Kandungan glikogen Rendah Tinggi
Hubungan dengan sel
penyangga
Melekat erat Melekat hanya pada
bagian permukaan dan
dasar
Inervasi masuk
Sel ganglion Tipe I Tipe II
Jumlah sel ganglion 27.000 2.100
Rasio sel ganglion
dengan sel rambut
1,8:1 5,7:1
Inervasi keluar
Sumber Kompleks olivary
superior lateral
Kompleks olivary
superior medial
Target postsipnatik Masukan dendrit Basal sel rambut
Mikrophonic dari koklea berupa arus bolak-balik atau alternating current
yang biasanya direkam pada koklea atau disekitar foramen window. Hal tersebut
menunjukkan aliran dari ion potasium yang sebagian besar melalui sel rambut
luar; dimana resistensi elektrik dari sel rambut luar ditentukan oleh pergerakan
yang terjadi pada membrana basalis. Ketika stereocilia mengarah jauh dari
modiolus, resistensi dari sel rambut akan menurun. Akibatnya terjadi peningkatan
aliran dan sedikit penurunan pada potensial endolimfatik. Ketika stereocilia
mengarah ke modiolus maka resistensi dari sel rambut akan meningkat dan aliran
listrik akan menurun disertai denan penurunan dari potensial endolimfatik.
Fluktuasi dari arus yang bersesuaian, mikrophonic dari koklea,bergantung pada
ada tidaknya sel rambut luar. Tidak seperti potensial neural, gelombang dari
mikrophonik koklea menggambarkan pergerakan dari membrana basilar. Potensial
summating adalah potensial DC yang direkam pada koklea sebagai respon
terhadap suara yang diberikan. Rekaman dari potensial DC ini dapat dibuat pada

skala timpani, media maupun vestibuli dan dimana terdapat elektrode besar pada
liang telinga manusia. Potensial tersebut dapat bersifat negatif maupun positif dan
dapat berbelik menurut polaritasnya, tergantung pada lokasi elektroda atau tingkat
dan frekuensi dari rangsangan. Potensial summating mungkin memiliki beberapa
sumber namun potensial ini menggambarkan perubahan potensial DC yang
diakibatkan oleh rangsangan dari intrasel dari sel rambut luar. Sel rambut dalam
memberikan pengaruh yang lebih sedikit.
Seluruh bagian syaraf maupun gabungan potensial aksi dibangkitkan dari
serabut syaraf auditorius. Gabungan potensial aksi paling baik direkam dengan
menggunakan elektrode besar yang diletakkan disekitar foramen window atau
syaraf auditorius dengan sinyal frekensi yang tinggi dan dalam waktu yang cepat.
Ketika sinyal menghasilkan aktivitas syaraf yang sinkron, yang disummed
menjadi gabungan potensial aksi. Amplitudo dari gabungan potensial aksi dapat
meningkatkan rangsangan intensitas sampai 40 sampai 50 dB, dimana keadaan
laten menjadi menurun selama intensitas rangsangan ditingkatkan. Pada tingkat
yang tinggi, puncak yang kedua terkadang perlu diamati untuk melihat aktivitas
refleks dari nukleus koklea. Gabungan dari potensial aksi dapat direkam secara
klinis dengan menggunakan elektroda yang ditempelkan pada kulit kepala atau
liang telinga atau dengan melalui pendekatan trantimpanic dimana elektode
ditempatkan di sekitar foramen window. Perbandingan amplitudo dari potensial
summating dengan amplitudo gambungan potensial aksi digunakan sebagai
indikator adanya fistula perilimfe namun validitas dari indikator ini masih
diragukan.
FISIOLOGI NERVUS KE-DELAPAN
Nervus Auditorius mempunyai kurang lebih 30.000 serabut syaraf pada manusia
dan kurang lebih 50.000 serabut syaraf pada hewan (kucing). Salah satu hasil
penemuan yang paling penting akhir-akhir ini adalah mengenai ditemukannya 90-
95% serabut syaraf (Tipe I, berupa serabut syaraf radier), yang mempersyarafi sel
rambut dalam, sedangkan 5-10% (Tipe II, berupa serabut syaraf spiral)
mempersyarafi sel rambut luar (Gambar 129.12). Kebanyakan serabut syaraf

auditorius berupa serabut syaraf tipe I dimana banyak terdapat sel rambut dalam.
Serabut syaraf radier ini memiliki badan sel bipolar pada ganglion spiral. Serabut
syaraf luar yang berupa spiral bersifat monopolar dan tidak memiliki selaput
myelin. Bagian dari serabut syaraf auditorius kebanyakan didapat dari
menyisipkan mikro elektroda pada serabut syaraf yang keluar dari meatus
auditorius internal. Pengukuran dari fungsi nervus auditorius dapat dilakukan
dengan menggunakan penghitungan spontan kurva tuning, dan fungsi intensitas
(tingkat-level).
Nervus auditorius pada hewan tingkat mamalia kebanyakan mengalami
kemnduruan oleh karena tidak adanya stimulasi akustik pada hewan ini. Serabut
syaraf ini telah diklasifikasikan menjadi 3 kategori berdasarkan tingkat aktivitas
spontannya menjadi level tinggi (18-120 per detik), tingkat sedang (0.5-18 per
detik) dan tingkat rendah (0-0,5 per detik). Serabut syaraf dengan tingkat aktivitas
spontan yang tinggi memiliki respon terhadap sinyal yang dihasilkan oleh
auditorius pada jumlah yang kecil dibandingkan dengan serabut syaraf dengan
tingkat aktivitas spontan yang sedang dan rendah. Serabut syaraf dengan aktivitas
spontan yang tinggi memilki dendrit tebal yang cenderung berakhir pada sisi sel
rambut dalam yang berhadapan dengan sel rambut luar. Serabut syaraf dengan
aktivitas spontan yang sedang dan rendah memiliki dendrit tipis yang berakhir
pada sisi sel rambut dalam yang berhadapan dengan modiolus. Penelitian yang
berlangsung saat ini memperkirakan bahwa serabut syaraf dengan aktivitas
spontan yang tinggi mempunyai ujung yang berbeda pada nukleus koklearis
auditorius dengan serabut syaraf yang memiliki aktivitas spontan yang rendah.
Dengan kata lain, aktivitas spontan dari serabut syaraf yang terjadi tidak secara
acak tetapi terbukti berbeda secara anatomis dan fungsional (12-15). Pengukuran
dengan kurva tunning dari masing-masing serabut syaraf auditorius merupakan
pengukuran yang paling mendasar dari fungsi nervus auditorius. Terdapat
pengkontrolan dari peningkatan tinggi frekuensi dan tingkat nada. Penambahan
tingkat nada dilakukan sampai terdapat perubahan kriteria pada tingkat firing
berupa 1-2 spike per detik. Digunakan peningkatan tinggi dari nada pada frekuensi
yang luas dan tingkat nada terendah direkam untuk menentukan frekuensi yang

menyebabkan adanya suatu kelainan. Hasil dari kurva isorespon ini dinamakan
tunning curve.
Gambar 129.17 menunjukkan tunning curve dari 6 serabut syaraf yang
berbeda. Peningkatan tajam dari tunning curve menunjukkan karakteristik dan
frekuensi yang paling baik dari serabut syaraf. Bagian dengan frekuensi dan
karakteristik rendah yang mempersyarafi sel rambut dalam di bagian apikal dari
koklea, serabut syaraf dengan karakteristik frekuensi yang tinggi mempersyarafi
sel rambut dalam mulai dari bagian basal dan seterusnya. Tunning curve
dideskripsikan menurut frekuensi akhir atau karakteristik frekuensi, bagian
frekuensi yang tinggi dan rendah dan bagian belakang. Serabut syraf dengan
karakteristik frekuensi kurang dari 1 kHz berbentuk huruf V yang tajam. Serabut
syaraf dengan karakteristik frekuensi yang lebih tinggi memiliki ujung pada
karakteristik frekuensi dan tail yang memanjang pada frekuensi yang lebih
rendah. Bagian yang tinggi dari tunning curve merupakan daerah frekuensi yang
lebih besar dari karakteristik frekuensi. Jika terjadi peningkatan karakteristik
frekuensi, bagian tinggi dari tunning curve akan lebih curam dengan adanya slope
yang dapat melebihi 500 dB per oktaf. Karakteristik dari tunning curve serabut
syaraf auditorius sama dengan isoampitude curve dari gelombang mekanik
(Gambar 129.15)
Kerusakan pada sel-sel sensoris, termasuk dengan stereosilia, berpengaruh
besar dalam bentuk tunning curve (Gambar 129.18). Bagian kanan bawah dari
gambar tersebut menunjukkan bahwa ketika terjadi kerusakan pada sel rambut
dalam, tunning curve dari serabut syaraf auditorius dari bentuk normal sel rambut
dalam akan berubah menjadi beberapa bentuk lain. Tidak terdapat daerah puncak
yang sensitif; batas ambang dari serabut syaraf akan meningkat menjadi sekitar
40-45 Db. Bagian dengan frekuensi tinggi tidak memiliki slope yang curam dan
bagian dengan frekuensi yang rendah menjadi lebih sensitif atau hipersensitif.
Karakteristik frekuensi dari serabut syaraf akan lebih tampak pada frekuensi yang
rendah dan lebar dari serabut syaraf akan tampak menjadi lebih besar. Bagian kiri
atas dari gambar 129.18 menunjukkan konsekuensi dari kerusakan pada sebagian
stereocilia dari sel rambut luar. Ambang batasyang terjadi sekitar 30 Db namun

tidak terlalu pendek, masih ada gambaran daerah puncak yang tajam, dan ujung
dari frekuensi yang rendah bersifat hipersensitif. Ketidakteraturan dari tunning
curve ini dapat menjelaskan kelainan yang ditemui pada monoaural diplacusis;
dimana nada pada satu telinga (800 Hz) memiliki 2 puncak, sebagai contoh, satu
nada pada 800 Hz dan nada lainnya sekitar 2.8 kHz.
Bagian atas kiri dari Gambar 129.18 menunjukkan tunning curve pada
stereocilia sel rambut dalam yang mengalami kerusakan atau kekacauan, dimana
kebanyakan dari stereocilia dari sel rambut luar bersifat normal atau mendekati
normal. Ambang batas akan meningkat sekitar 30 dB namun bentuk dari tunning
curve masih dalam batas normal. Bagian bawah kiri dari gambar menunjukkan
respon terhadap sinyal pada frekuensi nada yang berjarak sempit yaitu pada nada
yang lebih besar dari 90 dB SPL. Pada kasus ini, sel-sel sensoris masih ada namun
stereosilia dari sel rambut dalam sudah hancur dan bagian sel rambut luar sudah
mengalami kekacauan yang besar. Aktivitas syaraf yang normal tersebut;
termasuk kepada sensitifitas (deteksi dari suara yang lemah) dan kekuatan
frekuensi, bergantung pada bentuk sel rambut luar dan stereosilia yang masih
normal.
Walaupun ambang batas dari serabut syaraf auditorius berhubungan
dengan tingkat pemberhentian yang spontan, kebanyakan serabut syaraf afferen
(60%) memiliki tingkat spontanitas yang tinggi dengan ambang batas sekitar 20
dB yang lebih besar dari ambang batas pada hewan. Serabut syaraf dengan
akitivitas spontan yang rendah memiliki ambang batas sekitar 60 dB. Jjarak relatif
dinamik pada kebanyak serabut syaraf auditorius adalah sekitar 30 dB terhadap
saturasinya (Gambar 129.19), sedangkan beberapa serabut dengan aktivitas
spontan yang rendah memiliki jarak relatif dinamik yang lebih luas. Besarnya Jark
relatif dinamik pada manusia (0 Db SPL sampai dengan lebih dari 100 dB SPL),
menyebabkan sistem auditorius harus memiliki serabut syaraf yang ambang
batasnya memiliki jarak yang luas dan memilkiki tingkat firing yang dapat
menutupi intensitas yang tinggi. Kemampuan telinga manusia untuk memberikan
respon pada suara yang lebih besar dari 120-dB Range (10,12) merupakan suatu
hal yang luar biasa. Salah satu caranya adalah dengan menggunakan serabut

syaraf dengan aktivitas spontan yang rendah; dan penarikan karakteristik
frekuensi dari serabut syaraf.
Salah satu tuli sensorineural yang paling sering adalah akibat pajanan
terhadap bising yang berlebihan. Gambar 129.20 memberikan penjelasan.
Kebisingan tergantung pada aktifitas total saraf pendengaran. Seperti yang
ditunjukkan pada Gambar 129.20A, jumlah serabut yang aktif meningkat
perlahan-lahan sesuai dengan peningkatan intensitas, dan hanya ujung curve
tunning yang aktif. Sejalan dengan peningkatan intensitas lebih lanjut, ekor curve
tunning juga ikut terlibat, dan sejumlah serat aktif bertambah secara cepat. Pada
kasus tuli sensorineural, ujung kurva tuning menghilang, dan serabut tidak aktif
sampai tingkat sinyal cukup mencapai ekor kurva tuning. Secara kasar, beberapa
serabut akhirnya aktif secara bersamaan.
ORGAN PENDENGARAN NONLINEAR
Kelebihan utama dari telinga tengah adalah organ linearnya, sedangkan koklea
dan saraf pendengaran adalah bentuk nonlinear. Kemungkinan mengenai
nonlinear yang paling banyak dipelajari adalah nada campuran, yang digambarkan
dalam hubungannya dengan emisi koklea, dan supresi dua nada, pada saat
direkam di serabut saraf pendengaran.
Supresi dua nada adalah pengurangan angka yang dihasilkan oleh sebuah
nada ketika nada kedua diberikan. Gambar 129.21 memperlihatkan curve tunning
yang disertai dengan batas daerah supresi di atas karakteristik frekuensi serabut
saraf dan di bawah karakteristik frekuensi serabut. Nada yang terdapat pada
daerah titik atau supresi pada gambar mengurangi nilai hasil yang disebabkan oleh
pemeriksaan nada. Nada eksitator dan supresor dilakukan secara bersamaan, dan
oleh karena sedikit atau tidak adanya keterlambatan waktu yang berhubungan
dengan fenomena ini, tidak ada bukti yang sesuai bahwa hal tersebut merupakan
hasil kerja dari saraf, efek ini lebih sering disebut supresi daripada inhibisi.
Supresi dua nada dalam unit tunggal tergambar dalam potensial aksi. Gambar
129.21 (kanan) memperlihatkan curve tunning potensial aksi disertai dengan
daerah supresi pada daerah titik. Pada kasus ini, amplitudo potensial aksi diubah

oleh sinyal supresi, walaupun pada unit tunggal (kiri), angka hasil saraf berkurang
dalam jumlah yang berubah-ubah (20%). Unit tunggal dan daerah supresi
potensial aksi memiliki sifat yang serupa. Sama seperti supresi dua nada dapat
diteliti pada respon interseluler DC sel rambut dalam, terdapat kemungkinan
supresi dua nada yang berawal di mekanisme cochlear alami dan sebelum sel
rambut dalam.
Pada keadaan tuli sensorineural yang disebabkan karena adanya paparan
terhadap bising ataupun penggunaan obat-obat yang bersifat ototoksik, akan
terdapat pengaruh yang besar pada penekanan tingkat dua nada jika dapat diukur.
Penekanan tingkat dua nada terlihat normal atau mendekati normal pada tuli
koklea dimana sel-sel sensoris termasuk stereosilia normal atau mendekati
normal, namun terdapat pengaruh pada vaskularisasi dari stria tersebut. Hal inilah
yang dapat menyebabkan terjadinya prebiskus.
Emisi otoakustik adalah suara yang dapat ditangkap oleh liang telinga
ketika membrane timpani menerima getara yang ditransmisikan ke telinga tengah
dari koklea. Pemeriksaan emisi otoakustik menunjukkan bahwa koklea tidak
hanya merupakan penerima energi akustik yang aktif namun dapat juga
membangkitkan atau mengamplifikasikan suara. Terdapat dua jenis emisi
otoakustik. Emisi otoakustik spontan dihasilkan oleh koklea tanpa adanya
stimulus dari luar dan terdapat pada nada murni yang stabil pada -10 sampai 30
dB, dimana ditemukan pada 30-40% telinga sehat (18,19). Frekuensi dari emisi
otoakustik spontan tidak dapat menunjukkan bagian pasti dari koklea yang
mengalami kerusakan, namun hanya merefleksikan daerah pada koklea dengan
aktivitas sel rambut luar yang terganggu. Emisi otoakustik spontan dapat direkam
pada jangka panjang namun dengan variasi sistematis pada frekuensi dan
amplitudo.
Emisi otoakustik yaitu transient-evoked dihasilkan setelah adanya stimulus
akustik yang berasal dari luar. Emisi otoakustik transient-evoked dihasilkan oleh
suatu probe yang dipasang pada bagian telinga luar. Gelombang yang terbentuk,
akibat adanya gerakan osilasi dapat dilihat pada emisi otoakustik transient evoked,
terjadi akibat adanya hubungan antara gerakan dari membran timpani, yang

menyebabkan terjadinya fluktuasi pada koklea telinga (Gambar 129.22).
Walaupun stimulasi akustik yang diberikan hanya berupa click pada seluruh
koklea, respon stimulus dari emisi otoakustik transient-evoked dapat
menghasilkan frekuensi spesifik tentang koklea melalui pemisahan respon pada
frekuensi yang berbeda. Emisi otoakustik transient-evoked sangat sensitif
terhadap adanya kelainan pada koklea pada frekuensi tertentu. Frekuensi dimana
ambang dengar yang ada melebihi 20 sampai 30 dB tidak dapat dideteksi oleh
emisi otoakustik transient-evoked. Karena kesensitifannya terhadap kelainan dari
koklea, emisi otoakustik transient-evoked banyak digunakan pada program
screening untuk bayi baru lahir.
Emisi otoakustik distortion product juga telah banyak digunakan secara
klinis. Teknik pemeriksaan Emisi otoakustik transient evoked dan distortion
product bersifat saling melengkapi. Emisi otoakustik distortion product dapat
mendeteksi kelainan dengan frekuensi yang lebih luas dengan sensitivitas yang
rendah terhadap kondisi subklinis atau minor pada pasien dewasa. Ketika dua
nada murni, F1 dan F2 diberikan kepada koklea, akan terjadi emisi otoakustik
distortion product. Distortion product yang paling jelas adalah pangkat tiga dari
penyimpangan nada, 2F1-F2. Pengukuran emisi otoakustik distortion product
pada rangsangan yang berulang dapat menegakkan `growth rate’ dari emisi
otoakustik. Telinga yang sehat cenderung untuk memiliki growth rate 1 dB per
rangsangan 1 dB yang diberikan. Telinga dengan suatu kelainan menunjukkan
pertumbuhan yang lebih curam. Dapat terjadi kesalahan pada hasil emisi
otoakustik ini, oleh karena itu hasilnya harus dirata-ratakan dalam bentuk range.
Emisi otoakustik distortion product mudah direkam pada pasien dengan sistem
telinga tengah yang normal.
SISTEM SYARAF PUSAT AUDITORIUS
Jalur ascending dan descending dari sistem auditorius telah dijelaskan
sebelumnya secara singkat dalam hubungannya dengan aktivitas listrik yang
dihasilkan oleh nervus auditorius. Gambaran sistematis dari jalur ascending dan

descending tersebut digambarkan pada Gambar 129.23 dan 129.13. Diagram ini
merupakan gambaran sederhana dari sistem auditorius namun dapat memberikan
informasi mengenai kompleksitas dari sistem auditorius. Seluruh nervus delapan
afferen berhenti pada tingkat nukleus koklear. Lima tipe utama dari sel-sel
tersebut ditemukan pada nukleus koklea, masing-masing sel memiliki morfologi
dan fisiologi yang berbeda sesuai dengan respon terhadap stimulus yang
diberikan, respon offset dan modulasi frekuensi. Dari nukleus koklea, kebanyakan
serabut syaraf menyebrangi batang otak menuju ke kompleks olivarius superior
kontralateral; sementara itu sejumlah lainnya yang lebih sedikit menuju ke
kompleks olivarius superior ipsilateral.
Komplek olivarius superior merupakan pusat dari sistem auditorius
ascending, dimana input dari...... Nukleus auditorius yang berada di atas komplek
olivarius superior dapat bersifat eksitator dan inhibitor terhadap stimulus yang
diberikan pada telinga. Stimuls dari telinga kontralateral biasanya bersifat
eksitator terhadap badan sel sistem syaraf auditorius, sedangkan dari telinga yang
ipsilateral bersifat sebaliknya yaitu inhibitor. Seperti yang ditunjukkan pada
Gambar 129.13, kompleks olivarius superomedial merupakan asal dari serabut
eferen yang menyilang dan berakhir di sel rambut luar., sedangkan kompleks
olivarius laterosuperior merupakan asal dari serabut eferen yang tidak menyilang
dan berkahir pada sel rambut dalam. Walaupun sistem sayaraf efferent auditorius
memiliki beberapa fungsi khususnya dalam melindungi kklea dari pajanan bising
berlebihan, fungsi dari sistem ini masih belm diketahui dan sering menjadi
perdebatan (24).
Kolikulus inferior merupakan kompleks nukleus dengan 18 tipe sel dan
memiliki 5 daerah khusus. Kolikulus ini terlibat dalam seluruh peranan dan
perilaku dari auditorius termasuk dalam sensitivitas frekuensi, intensitas,
kenyaringan suara dan pendengaran binaural. Kolikulus inferior lebih jelas
dibandingkan dengan pusat relay. Badan geniculate medial dari thalamus
mengirimkan proyeksi kepada koreteks auditorius namun fungsi spesifiknya
masih belum diketahui.

Lokasi dari korteks auditorius adalah pada fissura sylvian dari lobus
temporalis; area skunder auditorius kebanyakan terletak berkelompok di sekitar
daerah primer. Pada masing-masing area sel tersusun secara tonotopikal dalam
bentuk kolumner, masing-masing kolumner memiliki karakteristik tertentu. Sel-
sel yang terdapat pada kolom bisa saja memiliki nada yang berbeda pada
karakteristik frekuensi yang sama; satu kolom tertentu dapat menghasilkan sandi
sementara kolom lain yang memiliki respon inhibitor terhadap rangsangan pada
satu telinga dan respon eksitator pada telinga yang lain, begitu juga sebaliknya.
Seperti hubungan antara thalamus dan korteks, nukleus dalam badan geniculate
medial yang mengirim serabut syaraf ke korteks auditorius juga menerima serabut
syaraf dari daerah yang sama pada korteks. Lesi bilateral pada lobus temporalis
dapat memberikan efek yang besar dan meluas (tuli kortikal, dimana berpengarh
berat pada organ-organ auditorius, termasuk kepada cara bicara, lokalisasi suara,
proses informasi pada lobus temporal dan deteksi adanya kerusakan-sinyal dalam
jangka pendek). Alat lain yang penting dalam sistem auditorius adalah tonotopic
nature. Mulai dari membran basiler sampai kepada korteks auditorius, sistem ini
terorganisir dengan frekuensi. Masing-masing daerah pada membran basilar
memberikan respon yang baik pada frekuensi tertentu-frekuensi tinggi berada
pada bagian dasar, dan frekuensi yang rendah berada pada bagian apeks.
Pengaturan tonotopic berada pada nukleus koklearis. Gambar 129.24
menunjukkan elektoda-elektroda yang masuk ke dalam nukleus koklea, serabut
syaraf dengan karakteristik frekuensi yang akan melakukan kontak, dan
karakteristik frekuensi yang akan membentuk kemajuan yang pesat. Data yang
sama terdapat pada seluruh nukleus dari sistem syaraf auditorius, termasuk dengan
korteks auditorius.
Pemeriksaan klinis yang memberikan informasi dasar mengenai sistem
sayaraf auditorius melibatkan interpretasi mengenai aktivitas listrik yang
dihasilkan oleh nervus kedelapan (evoked potensial). Pemeriksaan ABR (Auditory
Brainstem Response) merupakan salah satu komponen penilaian aktivitas listrik
pada nervus auditorius. Pemeriksaan ini pertama kali dilaporkan oleh Sohmer dan
Feinmesser pada tahun 1967 (26). ABR direkam melalui elektode permukaan

yang ditempelkan pada berbagai posisi di kepala. ABR terdiri dari satu serial yang
terdiri atas 7 gelombang yang terjadi sekitar 10 milidetik stelah rangsangan
diberikan. Suatu ketentuan yang dipakai di Amerika Serikat yaitu menamakan
puncak gelombang tersebut dengan menggunakan angka romawi. Telah diketahui
bahwa ABR dibangkitkan oleh syaraf auditorius dan mengikuti jalur serabut
syaraf dan nukleus pada jaras auditorius pada batang otak. Masing-masing
gelombang dibangkitkan sebagai berikut; gelombang I dan II merupakan nervus
kedelapan; gelombang III merupakan nukleus koklearis; gelobang IV merupakan
lemniscus lateralis/olivarius superior, dan gelombang V merupakan lemniscus
lateralis/colliculus superior.
Pemeriksaan ABR dilakukan dengan pemberian rangsangan click karena
rangsangan tersebut memberikan hasil yang paling jelas. ABR digunakan secara
klinis untuk menentukan perkiraan sensitivitas auditorius dan penilaian
otoneurologis. Pemeriksaan ini mendeteksi lesi di sepanjang jaras auditorius dan
batang otak. Pemeriksaan dapat dilakukan dalam keadaan sadar dan hasilnya tidak
dipengaruhi oleh pemberian obat-obat untuk menurunkan kesadaran. Pemeriksaan
ini biasanya dilakukan pada anak-anak dalam keadaaan sedasi atau selama anak
sedang tidur.
Pemerikssan klinis audiometri untuk menilai evoked potesial saat ini telah
mengalami kemajuan dalam hal teknik. Auditory steady-state response (ASSR)
dapat memberikan hasil yang lebih jelas mengenai kelainan dan disfungsi dari
sistem auditorius. Tidak seperti ABR, yang hasilnya diperoleh dari rangsangan
yang sementara, ASSR menggunakan rangsangan yang terus-menerus. Nada pada
pemeriksaan ini bersifat spesifik karena nada yang terus menerus tidak memiliki
spektrum penyimpangan seperti yang disebabkan oleh pemberian rangsangan
berupa tone burst maupun click (27). Yang paling penting, ASSR juga dapat
dilakukan pada pasien sadar penuh.
Terdapat beberapa keuntungan pemeriksaan dengan ASSR jika
dibandingkan dengan pemeriksaan ABR. Keuntungan pertama, ASSR memiliki
teknik yang lebih baik dalam mengevaluasi telinga dengan alat bantu dengar
karena penggunaan alat bantu dengar dan implan koklea dapat menghasilkan

rangsangan yang terus menerus dengan distorsi daripada rangsangan sementara.
Selain itu, ASSR juga dapat memberikan informasi menganai frekuensi ambang
batas pada tingkat intensitas 120 dB atau lebih (28,29). Hal ini dapat mengkoreksi
penurunan pendengaran yang berat dan dalam yang tidak dapat dilakukan oleh
ABR. ASSR juga dapat digunakan pada pasien pediatri yang akan dilakukan
implan koklea (30). Yang terakhir, ASSR juga memiliki waktu yang lebih efisien
jika dibandingkan dengan ABR karena dapat menunjukkan ambang batas dalam
waktu yang singkat (31).
Neuroanatomis dari sistem pendengaran sangat rumit. Proses pengiriman
informasi dapat melibatkan proses pararel dan serial. Pembentuk sistem ini
merupakan satu syaraf dengan cabang-cabang tertentu pada area target. Proses
serial melibatkan sebuah serabut syaraf yang menuju pada satu target, yang
kemudian menuju target lainnya dan seterusnya. Pada sitem saraf pusat auditorius,
terdapat proses yang bersifat pararel dan serial. Karena sistem syaraf auditorius
sangat banyak, rumit dan merupakan sistem yang berkekuatan besar maka
interpretasi dari aktivitas listrik yang dihasilkan oleh nervus tersebut dan data-data
mengenai syaraf lain tidak mudah diperoleh.
RINGKASAN INTI
Alat pendengaran pada kepala dan telinga luar sangat penting karena dapat
melokalisasi sumber suara.
Telinga tengah berfungsi sebagai transformer antara air dan cairan yang
berada pada koklea dan menghasilkan suara sampai 25-30 dB. Kombinasi
alat-alat pendengaran yang terdapat di kepala, telinga luar, telinga tengah dan
impedansi yang masuk pada koklea menentukan tingkatan dari frekuensi
pendengaran pada manusia.
Koklea berbentuk seperti gulungan dari tulang rawan yang memiliki panjang
sekitar 35 mm dan terdiri atas 3 kompartemen- skala timpani, skala media dan
skala vestibuli. Skala timpani dan skala vestibuli berisi cairan perilimfe dan
dihubungkan dengan helikotrema yang berada pada puncak koklea. Skala
media berisi cairan endolimfe dan memiliki potensial DC yang positif sekitar

80 Mv yang dapat membangkitan kerja pompa Na+-K+-ATPase pada stria
vaskular.
Transduser pendengaran berupa organ Corti yang terdiri atas sel-sel sensoris
(tiga baris sel rambut luar dan satu baris sel rambut dalam). Defleksi dari
stereosilia (rambut-rambut) sel-sel sensoris terjadi karena adanya rangsang
mekanik yang menginisiasi terjadinya tranduksi.
Gelombang mekanik, yang berasal dari permukaan dasar sampai kepada
puncak dari koklea, dapat membangkitkan suatu pergerakan menyerupai
piston dari tulang pendengaran stapes. Gelombang mekanik ini memiliki
puncak yang tajam pada dasar suara dengan frekuensi yang tinggi yang akan
berlanjut kepada puncak yang tajam bersamaan dengan turunnya frekuensi
suara.
Defleksi dari stereosilia oleh gelombang mekanik akan membuka dan
menutup kanal ion; akibatnya akan ada aliran (ion potasium) ke sel-sel
sensoris. Aliran dari potasium sejumlah +80 mV menjadikan potensial
intraseluler pada sel rambut dalam dan luar negatif. Akibat adanya
depolarisasi akan terjadi cascade dari enzym yang melepaskan transmitter dan
mengaktifkan kerja serabut syaraf aferen.
Sekitar 90-95% serabut syaraf tipe radier (tipe I) mempersyarafi sel rambut
dalam. Sementara itu, sekitar 5-10% (tipe II, serabut syaraf tipe spiral)
mempersyarafi sel rambut luar. Masing-masing sel rambut dalam dipersyarafi
oleh 15-20 searabut syaraf tipe I. Masing-masing serabut syaraf tipe II
mempersayarafi sekitar 10 sel rambut luar. Sekitar 1.800 serabut syaraf eferen
berasal dari sel-sel sensoris dari nukleus olivarius superior ipsilateral dan
kontralateral.
Pengukuran fungsi dari nervus auditorius yang paling mendasar adalah dengan
menggunakan tunning curve dari masing-masing serabut syaraf auditorius.
Tunning curve dari masing-masing serabut syaraf hampir sama dengan
tunning curve dari gelombang mekanik. Kerusakan pada sel-sel sensoris dan
stereosilia dapat mengubah tampilan dari tunning curve, termasuk dengan
sensitifitas dan ketajaman dari tunning.

Sistem dari telinga bagian tengah bersifat pasif dan sejalan dengan respon
terhadap sinyal sebesar lebih dari 130 dB SPL, namun telinga bagian dalam
bersifat aktif dengan dan memiliki amplifikasinya sendiri dan bersifat -
nonliniar. Hal ini yang membuat telinga dalam dapat memberikan respon
terhadap intesitas suara yang luas dan menunjukkan dasar dari fenomena
supresi.
Walaupun sistem syaraf eferen dari auditorius berkembang dengan baik,
fungsi pastinya belum dapat dikenal dengan jelas. Sistem ini mungkin
berperan dalam tranduksi koklea dan melindungi koklea dari pajanan suara
yang berlebihan.