







Bahasa
Halaman
Hukum


ORIGINALARTICLE
Vegetation and fire history of a Chinesesite in southern tropical Xishuangbannaderived from phytolith and charcoalrecords from Holocene sediments
Yansheng Gu1,2*, Deborah M. Pearsall3, Shucheng Xie4 and Jianxin Yu1
1Key Laboratory of Biogeology and
Environmental Geology (China University of
Geosciences), Ministry of Education, Wuhan
430074, China, 2Institute of Ecology and
Environment, China University of Geosciences,
Wuhan 430074, China, 3Department of
Anthropology, University of Missouri,
Columbia, MO 65211, USA and 4State Key
Laboratory of Geological Processes and Mineral
Resources, China University of Geosciences,
Wuhan 430074, China
*Correspondence: Yansheng Gu, Key Laboratory
of Biogeology and Environmental Geology
(China University of Geosciences), Ministry of
Education, Wuhan 430074, China.
E-mail: [email protected]
ABSTRACT
Aim The aims of this paper are to reconstruct the vegetation and fire history over
the past 2000 years in a well-preserved rain-forest area, to understand
interactions between climate, fire, and vegetation, and to predict how rain
forest responds to global warming and increased intensity of human activity.
Location Xishuangbanna, south-west China, 21–22� N, 101–102� E.
Methods Phytolith (plant opal silica bodies) morphotypes, assemblages, and
indices were used to reconstruct palaeovegetation and palaeoclimate changes in
detail. Micro-charcoal particles found in phytolith slides, together with burnt
phytoliths and highly weathered bulliform cells, were employed to reconstruct a
record of past fire occurrence. A survey of field sediments, lithology, and 14C
dating were also employed.
Results Phytoliths were divided into 11 groups and classified into 33 well-
described morphotypes according to their shape under light microscopy and their
presumed anatomical origins and ecological significance. The phytolith
assemblages were divided into six significant zones that reveal a complete
history of vegetation changes corresponding to climate variation and fire
occurrence. Phytolith assemblages and indices show that the palaeoclimate in the
study area is characterized by the alternation of warm–wet and cool–dry
conditions. Phytolith and charcoal records reveal that 12 fire episodes occurred.
Comparison of burnt phytoliths with an aridity index (Iph) shows that fire
episodes have a strong relationship with drought events.
Main conclusions Our results indicate that fire occurrence in the tropical rain
forest of Xishuangbanna is predominantly under the control of natural climate
variability (drought events). Nearly every fire episode is coupled with a climatic
event and has triggered vegetation composition changes marked by a pronounced
expansion of grasses. This indicates that drought interacts with fire to exert a
strong influence on the ecological dynamics of the rain forest. However, the
impact of human activity in recent centuries is also significant. Our results are
important for understanding the interactions between climate, fire, and
vegetation, and for predicting how rain forest responds to global warming and
increased human activity.
Keywords
Charcoal, China, climate change, fire history, Holocene sediments, phytoliths,
tropical rain forest, Xishuangbanna.
Journal of Biogeography (J. Biogeogr.) (2008) 35, 325–341
ª 2007 The Authors www.blackwellpublishing.com/jbi 325Journal compilation ª 2007 Blackwell Publishing Ltd doi:10.1111/j.1365-2699.2007.01763.x

INTRODUCTION
A given rain forest will experience variations in rainfall pattern
and in the frequency and intensity of droughts. The increase in
research into droughts in tropical forests has resulted in
increased attention being paid to forest–climate relationships
and rain-forest deforestation and fragmentation (Walsh, 1996;
Walsh & Newbery, 1999; Potts, 2003). These issues are
especially relevant in tropical monsoon areas, where climate
is characterized by a distinctive distribution of precipitation.
Severe droughts can lead to the destruction of habitat and a
decrease in biodiversity in the Tropics, especially if accom-
panied by fires, which accelerate these changes (Woods, 1989;
Kellman & Meave, 1997; Harrison, 2000, 2001; Laurance et al.,
2000; Williamson et al., 2000; Potts, 2003; van Nieuwstadt &
Sheil, 2005). Global warming could also exacerbate habitat
destruction if it promotes a drier climate or strong El Nino-
Southern Oscillation (ENSO) events in the Tropics (Laurance
& Williamson, 2001). An understanding of drought and fire
history is thus important in evaluating interactions among
natural climate variability (drought events), vegetation, and
human disturbance, at least during the last several millennia,
and may help to predict how tropical ecosystems will respond
to future climate change (Marchant, 2005).
Primary rain forest is well preserved in tropical Xis-
huangbanna (Fig. 1). The climate history of the region,
including drought episodes, has been reconstructed in part
using pollen and documentary records from surrounding areas
(Liu et al., 1986; Sun et al., 1986; Walker, 1986; Zhu, 1997;
Zhu & Cai, 2005). The lack of stratigraphic palaeoecological
data from Xishuangbanna has prevented us from determining
the extent to which the rain forest of this region is sensitive to
climate change. Little is known about the relationship between
climate, vegetation and fire events in the tropical rain forests of
Xishuangbanna.
The aims of this paper are to reconstruct the local
palaeovegetation, palaeoclimate, and fire history of Xis-
huangbanna, and to understand their interrelationships using
phytolith and charcoal records from fluvial sediments in the
well-preserved rain forest area. Phytoliths are an important
and reliable tool for reconstructing local vegetation and
climate. Grass phytoliths, in particular, offer a promising
means to differentiate between grasses at the subfamily level
and to infer subtle changes in palaeoenvironmental conditions
(Twiss, 1987, 1992; Fredlund & Tieszen, 1994, 1997; Alexandre
et al., 1997; Lu & Liu, 2003b). Grass phytolith indices have
been successfully used to reconstruct humidity and aridity in
marine sediments and grasslands (Diester-Haass et al., 1973;
Twiss, 1987; Fredlund & Tieszen, 1994, 1997; Alexandre et al.,
1997; Barboni et al., 1999; Parker et al., 2004), although they
have not been widely used in tropical rain-forest areas. Burnt
phytoliths and highly weathered bulliform cells, together with
Figure 1 Map showing the distribution of
modern tropical rain forest in southern Xis-
huangbanna, the sampling site, and places
referred to in the text (Rain forest types are
after Zhu, 1997).
Y. Gu et al.
326 Journal of Biogeography 35, 325–341ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

charcoal particles, are used here as indicators of fire episodes in
association with changes in vegetation and climate.
MATERIALS AND METHODS
Sampling
The samples studied here were taken from a site in tropical
Xishuangbanna in south-west China (Fig. 1). The study area is
located on the southern margin (21–22� N, 101–102� E) of
Xishuangbannan Dai Autonomous Prefecture in Yunnan
Province. Altitude ranges from 477 to 2007 m. The tropical
rain forest, which is part of the tropical Asian flora, occurs
mostly in the lower hills, mountain valleys, and the basin
below 900 m altitude (Zhu, 1997). It is mainly composed of
mixed rain forest and dipterocarp rain forest, with the former
widely distributed in the study area (Fig. 1). The tropical rain
forest in Xishuangbanna has been conserved since 1958, and is
now part of a State Nature Reserve in which there is little
human activity (Zhu, 1997; Zhu et al., 1998). The rain forest
has a high biodiversity: over 3500 species of higher plants, 700
species of higher animals, and 1500 species of insects have been
recorded (Cao & Zhang, 1997; Pu et al., 2001). The regional
climate is dominated by the south-west monsoon (Indian
Ocean monsoon), but the south-east monsoon also affects the
region (Zhang, 1988). The migrating rain-belt of the south-
west monsoon has a strong influence on the rain-forest
ecosystem, resulting in a marked dry season (November to
April) and a wet season (May to October). At Menglun station,
a mixed rain-forest area, the annual mean precipitation is
1463.3 mm, of which 85% falls during the wet season. In
contrast to the precipitation pattern, the monthly mean
temperature remains stable (21.7�C), although it rises slightly
in the wet season (25.7�C) (Liu & Li, 1996).
Samples were collected from a 1.5-m-deep profile of well-
developed fluvial sediments (clay and silt) in a seasonal pond
inside a 3-km-long gully (Fig. 2). The stratigraphy is described
in Table 1. Phytolith samples were taken at 2-cm intervals
from the top to a depth of 50 cm, and at 5-cm intervals from
50 cm to the bottom of the profile. The macro-charcoal
remains (> 1 mm) were identified by their shape and size and
described in situ.
Dating
A sample for radiocarbon dating was collected at a depth of
95–100 cm from an area where macro-charcoals were abun-
dant (layer 4 in Fig. 2). This was wet-sieved to remove
particles larger than 0.3 mm. 14C dating was performed at the
State Key Laboratory of Earthquake Dynamics of the China
Earthquake Administration, by liquid scintillation counting of
Figure 2 Sampling profile and lithological
features of sediments.
Table 1 Stratigraphic description of sediments from the tropical
rain forest in Xishuangbanna.
Depth
(cm)
Layer
no. Lithological description
0–15 (1) Predominantly black clay with roots and organics;
locally with abundant macro-charcoal particles;
macro-charcoal size up to 0.8 cm in diameter with
mostly angular shapes
15–65 (2) Black silt clay with locally abundant macro-charcoal
particles; macro-charcoal size up to 0.5 cm in
diameter with mostly angular shapes; locally with
circular quartz particles of size up to 0.2 cm in
diameter
65–85 (3) Light yellow silt locally with elliptic pebbles of size
up to 4 cm in diameter; circular quartz particles of
size up to 0.5 cm in diameter; abundant macro-
charcoal particles up to 0.3 cm in diameter with
mostly angular shapes
85–100 (4) Black silt clay with abundant macro-charcoal
particles of size up to 0.8 cm in diameter; locally
with elliptic pebbles up to 2 cm in diameter
Vegetation and fire history from phytolith and charcoal records
Journal of Biogeography 35, 325–341 327ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

benzene samples after washing with 5% NaOH and 10% HCl
solutions. Age was calculated using software from the LKB
Company according to the Chinese sugar standard; calibra-
tion was performed using calib v4.1.2 and a conventional
half-life of 5568 years (Stuiver & Reimer, 1993). Dating
showed that the sediments in the profile we studied were
deposited from before 1780 ± 110 yr bp until the present.
Tang (1992) reported a 14C date of 1660 ± 150 yr bp at a
depth of 1 m in fluvial sediments deposited near the study
area, which indicates a stable sedimentation rate in the area
(Tang, 1992).
Phytolith extraction, identification and counting
Phytolith extraction was carried out at the Key Laboratory of
Biogeology and Environmental Geology at the Chinese
University of Geosciences. The extraction method followed
Wang & Lu (1993). Air-dried samples of 10 g were
deflocculated in NaHCO3, and this was followed by carbon-
ate removal using 15% HCl and oxidation of organic matter
using a 30% solution of H2O2. After removal of sand grains
and clay minerals by sieving and gravity sedimentation,
respectively, phytoliths were transferred into a ZnI2 solution
(density 2.30 g cm)3), centrifuged at 2000 rpm for 10 min,
and washed with 1% acetic acid. After drying, phytoliths
were embedded in Canada balsam to make slides for
microscopic observation. Identification and counting for 35
phytolith samples was carried out at the Phytolith Laboratory
of the Department of Anthropology, University of Missouri-
Columbia. Extracts were scanned at 400 · magnification
using a Zeiss light microscope. For each sample (each slide),
phytoliths were identified and counted in 10 fields; counts
ranged from 126 to 720 phytoliths (with the majority around
400).
The identification of phytoliths in geological sediments is
based on morphological comparison with modern specimens,
as geological reference specimens are not available. In this
study, phytoliths were described and compared with the
phytolith reference collections at the University of Missouri
(see MU phytolith website: http://www.missouri.edu/~phyto)
and with published sources (Twiss et al., 1969; Brown, 1984;
Mulholland, 1989; Twiss, 1992; Bozarth, 1993; Wang & Lu,
1993; Carnelli et al., 2004; Stromberg, 2004; Lu et al., 2006;
Piperno, 2006). All phytolith morphotypes identified here
were described using International Code for Phytolith
Nomenclature (ICPN) 1.0 protocols (Madella et al., 2005)
whenever possible.
An aridity index (Iph) and a climate index (Ic) were
calculated from phytolith count data using the approaches of
Diester-Haass (Diester-Haass et al., 1973; Alexandre et al.,
1997) and Twiss (1987, 1992). The aridity index, Iph, is the
ratio of Chloridoideae to total Chloridoideae and Panicoideae
phytoliths. The climate index, Ic, is the ratio of Pooideae to
total Pooideae, Panicoideae, and Chloridoideae phytoliths,
with high values corresponding to cool temperatures (Twiss,
1992).
Phytolith morphotypes and classification
Phytolith morphotypes are described in detail (see Table 2)
and illustrated in Fig. 3. The major plant groups documented
in the profile samples are Poaceae (Gramineae), Cyperaceae,
Asteraceae (Compositae), ferns, gymnosperms, and broad-
leaved trees (Fig. 3). Poaceae phytoliths include short cells,
long cells, bulliform cells, and hair cells (grass-type). Short cells
can be classified as Pooideae (rondel and trapeziform),
Panicoideae (bilobate, cross, and polylobate), Chloridoideae
(square saddle), Bambusoideae (oblong concave saddle or
collapsed saddle) or Arundinoideae (trapeziform saddle or
plateaued saddle), based on their micro-morphological char-
acteristics and typical descriptions (Twiss et al., 1969; Brown,
1984; Twiss, 1987, 1992; Mulholland, 1989; Wang & Lu, 1993;
Fredlund & Tieszen, 1994; Piperno & Pearsall, 1998) (Fig. 3).
Sponge spicules and diatoms were also present in samples in
association with phytoliths.
It is important to mention that classification based on short
cells does not correspond completely to the grass subfamilies
designated above. For example, rondel short cells can occur in
Oryzoideae and Panicoideae; crosses can occur in Bambuso-
ideae, Oryzoideae and Arundinoideae; and bilobates can occur
in Bambusoideae, Oryzoideae and Arundinoideae (Twiss et al.,
1969; Twiss, 1992; Barboni et al., 1999). It should also be noted
that a small number of Pooideae phytoliths are produced in
Chloridoieae and Panicoideae (Brown, 1984; Mulholland,
1989; Barboni et al., 1999). These overlaps should be taken
into account during phytolith interpretation.
The correct classification of saddles is of great significance
for interpreting a climate signal on the basis of the climate (Ic)
and aridity (Iph) indices. Bambusoideae contribute large
numbers of oblong concave saddles (collapsed saddles), and
Arundinoideae produce many distinctive phytoliths called
trapeziform saddles (plateaued saddles). Compared with
saddles from the above two grass subfamilies, saddles from
Chloridoideae are either shorter and thicker or have different
three-dimensional structures. In particular, square saddles are
mostly ‘squat’, meaning that the axis of the phytolith
exhibiting the double-edge, saddle-like outline is wider than
the other axes, or the two axes are of equal dimensions
(Piperno, 2006). These distinctive morphological characteris-
tics of saddles permit reliable subfamily-level classification,
enabling Bambusoideae to be distinguished from Chlorido-
ideae and Arundinoideae (Wang & Lu, 1993; Piperno &
Pearsall, 1998; Lu et al., 2006; Piperno, 2006).
Opaque platelets with systematic perforations are produced
in the inflorescences of selected species of Asteraceae (Piperno,
1988; Bozarth, 1992; Mercader et al., 2000). The Cyperaceae
are characterized by polyhedrons with conical projections
(Piperno, 1988, 1989, 2006; Bozarth, 1992; Ollendorf, 1992;
Wang & Lu, 1993; Lu et al., 2006; Pearsall et al., 2006) (Fig. 3).
Ferns are rich in elongates with two undulating ridges, prisms
that are sinuate or cavate, and oblong lacunose phytoliths
(Piperno, 1988; Wang & Lu, 1993; Carter, 2002). Gymnosperm
trees produce predominantly parallelepipedal contorteds and
Y. Gu et al.
328 Journal of Biogeography 35, 325–341ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

Tab
le2
Des
crip
tio
no
fp
hyt
oli
thm
orp
ho
typ
es,
anat
om
ical
ori
gin
,p
alae
oec
olo
gica
lsi
gnifi
can
cean
dav
aila
ble
lite
ratu
re.
Maj
or
pla
nt
gro
up
Nam
eO
ther
Nam
esF
ig.
3M
orp
ho
typ
ed
escr
ipti
on
Size
ran
ge
(lm
)
An
ato
mic
al
Ori
gin
Pal
aeo
eco
logi
cal
sign
ifica
nce
Ref
eren
ces
Pan
ico
idea
e
gras
ses
Bil
ob
ate*
Du
mb
bel
l(a
)–(d
)B
ilo
bat
e,h
avin
gtw
o
lob
es
<25
Ep
ider
mal
sho
rtce
ll
Mo
sto
fth
eP
anic
oid
eae
are
C4
pla
nts
flo
uri
shin
g
inw
arm
,tr
op
ical
to
sub
tro
pic
alre
gio
ns
wit
h
am
od
erat
eam
ou
nt
of
avai
lab
leso
ilm
ois
ture
.
Tw
iss
etal
.(1
969)
,B
row
n
(198
4),
Pip
ern
o(1
988)
,T
wis
s
(199
2),
Mu
lho
llan
d&
Rap
p
(199
2),
Wan
g&
Lu
(199
3),
Pea
rsal
l(2
000)
,M
adel
laet
al.
(200
5)
Cro
ss*
(e),
(g)–
(h)
Qu
adra
-lo
bat
e,h
avin
g
fou
rlo
bes
<15
Tw
iss
etal
.(1
969)
,B
row
n
(198
4),
Pip
ern
o(1
988)
,T
wis
s
(199
2),
Mu
lho
llan
d&
Rap
p
(199
2),
Pea
rsal
l(2
000)
,M
adel
la
etal
.(2
005)
Cyl
ind
rica
l
po
lylo
bat
e*
Po
lylo
bat
eo
r
com
ple
x
du
mb
bel
l
(am
),(a
p)–
(aq
)P
oly
lob
ates
hav
ing
mo
re
than
two
lob
esli
nea
rly
arra
nge
d
<35
Tw
iss
etal
.(1
969)
,T
wis
s(1
992)
,
Mu
lho
llan
d&
Rap
p(1
992)
,
Pea
rsal
l(2
000)
,M
adel
laet
al.
(200
5)
Bam
bu
soid
eae
gras
ses
Ob
lon
g
con
cave
sad
dle
*
Lo
ng
sad
dle
or
coll
apse
d
sad
dle
(f),
(i)–
(k)
Co
llap
sed
sad
dle
or
Ch
usq
uea
-typ
esa
dd
le,
usu
ally
wid
th<
hei
ght
<20
Bam
bu
soid
eae
gras
ses
are
C3
pla
nts
flo
uri
shin
gin
war
mo
rh
ot,
hu
mid
tro
pic
alto
sub
tro
pic
al
regi
on
s.
Pip
ern
o(1
988)
,T
wis
s(1
992)
,
Wan
g&
Lu
(199
3),
Pip
ern
o&
Pea
rsal
l(1
998)
,Sa
se&
Ho
son
o
(200
1),
Li
etal
.(2
005)
,
Mad
ella
etal
.(2
005)
,L
uet
al.
(200
6)
Aru
nd
ino
idea
e
gras
ses
Tra
pez
ifo
rm
sad
dle
*
(m)–
(n)
Pla
teau
edsa
dd
leo
r
Ph
ragm
ites
-typ
eo
r
Ari
stid
a-ty
pe
<15
Mo
sto
fth
e
Aru
nd
ino
idea
ear
eC
3
pla
nts
flo
uri
shin
gin
war
m,
sem
iari
dto
mo
ist
regi
on
s.
Wan
g&
Lu
(199
3),
Pip
ern
o&
Pea
rsal
l(1
998)
,L
u&
Liu
(200
3a),
Pip
ern
o(2
006)
Od
dSi
de
view
like
acu
pw
ith
a
rou
nd
bas
ean
dca
vate
sid
e
<10
Ch
lori
do
idea
e
gras
ses
Squ
are
sad
dle
*
Sho
rtsa
dd
le(o
)–(s
)C
hlo
rid
oid
eae
sad
dle
,
usu
ally
wid
th‡
hei
ght
<15
Ch
lori
do
idea
egr
asse
sar
e
typ
ical
C4
gras
ses
gro
win
gin
war
m,
arid
to
sem
iari
dre
gio
ns
wh
ere
soil
mo
istu
reis
very
low
.
Tw
iss
etal
.(1
969)
,B
row
n
(198
4),
Mu
lho
llan
d(1
989)
,
Tw
iss
(199
2),
Mu
lho
llan
d&
Rap
p(1
992)
,W
ang
&L
u
(199
3),
Pip
ern
o&
Pea
rsal
l
(199
8),
Bar
bo
ni
etal
.(1
999)
,G
ob
etz
&B
oza
rth
(200
1),
Lu
&L
iu
(200
3b),
Mad
ella
etal
.(2
005)
Po
oid
eae
gras
ses
Ro
nd
el*
Co
nic
al,
hat
-sh
aped
,
circ
ula
r,o
val
(t)–
(u)
Co
nic
al<
15P
oo
idea
egr
asse
sar
ety
pic
al
C3
pla
nts
gro
win
gin
coo
l
and
wet
regi
on
sw
ith
hig
h
lati
tud
eso
rel
evat
ion
s.
Mu
lho
llan
d(1
989)
,M
ulh
oll
and
&R
app
(199
2),
Wan
g&
Lu
(199
3),
Fre
dlu
nd
&T
iesz
en
(199
4),
Pea
rsal
l(2
000)
,M
adel
la
etal
.(2
005)
Vegetation and fire history from phytolith and charcoal records
Journal of Biogeography 35, 325–341 329ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd
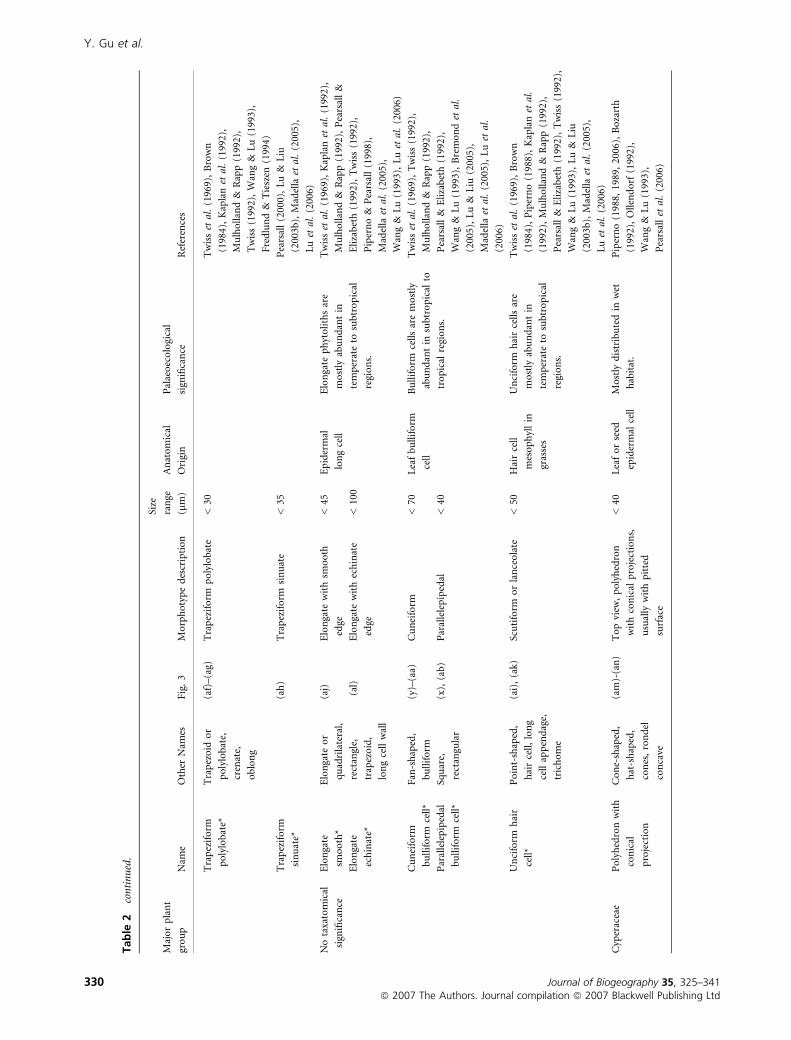
Tab
le2
con
tin
ued
.
Maj
or
pla
nt
gro
up
Nam
eO
ther
Nam
esF
ig.
3M
orp
ho
typ
ed
escr
ipti
on
Size
ran
ge
(lm
)
An
ato
mic
al
Ori
gin
Pal
aeo
eco
logi
cal
sign
ifica
nce
Ref
eren
ces
Tra
pez
ifo
rm
po
lylo
bat
e*
Tra
pez
oid
or
po
lylo
bat
e,
cren
ate,
ob
lon
g
(af)
–(a
g)T
rap
ezif
orm
po
lylo
bat
e<
30T
wis
set
al.
(196
9),
Bro
wn
(198
4),
Kap
lan
etal
.(1
992)
,
Mu
lho
llan
d&
Rap
p(1
992)
,
Tw
iss
(199
2),
Wan
g&
Lu
(199
3),
Fre
dlu
nd
&T
iesz
en(1
994)
Tra
pez
ifo
rm
sin
uat
e*
(ah
)T
rap
ezif
orm
sin
uat
e<
35P
ears
all
(200
0),
Lu
&L
iu
(200
3b),
Mad
ella
etal
.(2
005)
,
Lu
etal
.(2
006)
No
taxa
tom
ical
sign
ifica
nce
Elo
nga
te
smo
oth
*
Elo
nga
teo
r
qu
adri
late
ral,
rect
angl
e,
trap
ezo
id,
lon
gce
llw
all
(aj)
Elo
nga
tew
ith
smo
oth
edge
<45
Ep
ider
mal
lon
gce
ll
Elo
nga
tep
hyt
oli
ths
are
mo
stly
abu
nd
ant
in
tem
per
ate
tosu
btr
op
ical
regi
on
s.
Tw
iss
etal
.(1
969)
,K
apla
net
al.
(199
2),
Mu
lho
llan
d&
Rap
p(1
992)
,P
ears
all
&
Eli
zab
eth
(199
2),
Tw
iss
(199
2),
Pip
ern
o&
Pea
rsal
l(1
998)
,
Mad
ella
etal
.(2
005)
,
Wan
g&
Lu
(199
3),
Lu
etal
.(2
006)
Elo
nga
te
ech
inat
e*
(al)
Elo
nga
tew
ith
ech
inat
e
edge
<10
0
Cu
nei
form
bu
llif
orm
cell
*
Fan
-sh
aped
,
bu
llif
orm
(y)–
(aa)
Cu
nei
form
<70
Lea
fb
ull
ifo
rm
cell
Bu
llif
orm
cell
sar
em
ost
ly
abu
nd
ant
insu
btr
op
ical
to
tro
pic
alre
gio
ns.
Tw
iss
etal
.(1
969)
,T
wis
s(1
992)
,
Mu
lho
llan
d&
Rap
p(1
992)
,
Pea
rsal
l&
Eli
zab
eth
(199
2),
Wan
g&
Lu
(199
3),
Bre
mo
nd
etal
.
(200
5),
Lu
&L
iu(2
005)
,
Mad
ella
etal
.(2
005)
,L
uet
al.
(200
6)
Par
alle
lep
iped
al
bu
llif
orm
cell
*
Squ
are,
rect
angu
lar
(x),
(ab
)P
aral
lele
pip
edal
<40
Un
cifo
rmh
air
cell
*
Po
int-
shap
ed,
hai
rce
ll,
lon
g
cell
app
end
age,
tric
ho
me
(ai)
,(a
k)Sc
uti
form
or
lan
ceo
late
<50
Hai
rce
ll
mes
op
hyl
lin
gras
ses
Un
cifo
rmh
air
cell
sar
e
mo
stly
abu
nd
ant
in
tem
per
ate
tosu
btr
op
ical
regi
on
s.
Tw
iss
etal
.(1
969)
,B
row
n
(198
4),
Pip
ern
o(1
988)
,K
apla
net
al.
(199
2),
Mu
lho
llan
d&
Rap
p(1
992)
,
Pea
rsal
l&
Eli
zab
eth
(199
2),
Tw
iss
(199
2),
Wan
g&
Lu
(199
3),
Lu
&L
iu
(200
3b),
Mad
ella
etal
.(2
005)
,
Lu
etal
.(2
006)
Cyp
erac
eae
Po
lyh
edro
nw
ith
con
ical
pro
ject
ion
Co
ne-
shap
ed,
hat
-sh
aped
,
con
es,
ron
del
con
cave
(am
)-(a
n)
To
pvi
ew,
po
lyh
edro
n
wit
hco
nic
alp
roje
ctio
ns,
usu
ally
wit
hp
itte
d
surf
ace
<40
Lea
fo
rse
ed
epid
erm
alce
ll
Mo
stly
dis
trib
ute
din
wet
hab
itat
.
Pip
ern
o(1
988,
1989
,20
06),
Bo
zart
h
(199
2),
Oll
end
orf
(199
2),
Wan
g&
Lu
(199
3),
Pea
rsal
let
al.
(200
6)
Y. Gu et al.
330 Journal of Biogeography 35, 325–341ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

Tab
le2
con
tin
ued
.
Maj
or
pla
nt
gro
up
Nam
eO
ther
Nam
esF
ig.
3M
orp
ho
typ
ed
escr
ipti
on
Size
ran
ge
(lm
)
An
ato
mic
al
Ori
gin
Pal
aeo
eco
logi
cal
sign
ifica
nce
Ref
eren
ces
Ast
erac
eae
Op
aqu
e
per
fora
ted
pla
tele
t
(ac)
Op
aqu
ep
erfo
rate
d
pla
tele
t
<80
Ep
ider
mal
cell
Mo
stly
dis
trib
ute
din
coo
l,
sem
iari
dre
gio
ns.
Pip
ern
o(1
988)
,B
oza
rth
(199
2),
Mer
cad
eret
al.
(200
0)
Fer
nE
lon
gate
un
du
lati
ng
(ao
)E
lon
gate
wit
htw
o
par
alle
led
un
du
lati
ng
rid
ges
Mo
stly
dis
trib
ute
din
war
m
and
wet
hab
itat
s.
Pip
ern
o(1
988)
Pri
smsi
nu
ate
(ap
)P
rism
-sh
aped
wit
h
sin
uat
eed
ges
<60
Wan
g&
Lu
(199
3)
Ob
lon
g
lacu
no
se*
Tab
ula
rw
ith
cava
teed
ges
and
surf
aces
<70
Gym
no
sper
mP
aral
lele
pip
edal
con
tort
ed*
Po
lyh
edra
l,
Po
lyh
edro
n-
shap
ed
blo
cky
or
elo
nga
te
(aq
)–(a
r)P
oly
hed
ron
,u
sual
ly
blo
cky
or
elo
nga
tew
ith
smo
oth
or
pit
ted
surf
ace
<80
Ep
ider
mal
or
sub
epid
erm
al,
end
od
erm
alan
d
trac
hei
d
Mo
stly
dis
trib
ute
din
coo
l
toco
ldan
dse
mia
rid
regi
on
sw
ith
hig
hla
titu
des
or
elev
atio
ns.
Bo
zart
h(1
993)
,W
ang
&L
u
(199
3),
Stro
mb
erg
(200
4),
Car
nel
liet
al.
(200
4)
Cla
vate
con
tort
ed*
Bo
ot-
shap
ed(b
c)B
ent
like
ab
oo
t,w
ith
gran
ula
tesu
rfac
e
<40
Wan
g&
Lu
(199
3)
Bro
ad-l
eave
dA
bb
revi
ated
stel
late
*
An
ticl
inca
l
epid
erm
is,
jigs
aw-s
hap
ed
(av)
–(a
w)
Has
sin
uo
us
shap
e,n
o
do
ub
leo
utl
ines
<50
Ep
ider
mal
cell
Ab
un
dan
tin
dec
idu
ou
s
tree
s.
Pip
ern
o(1
988)
,B
oza
rth
(199
2),
Wan
g&
Lu
(199
3),
Sase
&
Ho
son
o(2
001)
,L
u&
Liu
(200
5),
Pea
rsal
let
al.
(200
6)
Aci
cula
rh
air
cell
*
Hai
rce
ll(a
s)–(
at)
Has
ab
igb
ase
<60
Hai
rce
lls
mes
op
hyl
lin
sid
e
tree
s
Ab
un
dan
tin
tro
pic
al
regi
on
s.
Cyl
ind
ric
scle
reid
*
Scle
reid
(bk)
–(b
l)A
vari
ably
shap
ed
scle
ren
chym
ato
us
cell
wit
ha
cen
tral
spin
eo
r
rid
ge,
usu
ally
iso
dia
met
ric
<10
0P
aren
chym
ao
f
leaf
Ab
un
dan
tin
sub
tro
pic
al,
tro
pic
alre
gio
ns.
Pip
ern
o(1
988,
2006
),P
ears
all
&
Eli
zab
eth
(199
2),
Kea
lho
fer
&
Pip
ern
o(1
998)
,St
rom
ber
g
(200
4),
Pea
rsal
let
al.
(200
6)
Glo
bu
lar
face
tate
*
Hem
isp
her
e(b
d)–
(be)
Fac
eted
,to
pvi
ew,
usu
ally
sph
eric
alb
od
y,ed
ges
are
scal
lop
ed
<20
Ep
ider
mal
cell
Glo
bu
lar
face
tate
ph
yto
lith
s
are
abu
nd
ant
in
sub
tro
pic
al,
tro
pic
al
regi
on
s.
Kea
lho
fer
&P
iper
no
(199
8),
Ru
nge
(199
9),
Pea
rsal
let
al.
(200
6)
Glo
bu
lar
gran
ula
te*
Sph
eric
al
rugo
se,
sph
ero
id
(ax)
–(b
a)G
lob
ula
rsp
her
ical
wit
h
gran
ula
tesu
rfac
e
<30
Ab
un
dan
tin
sub
tro
pic
al
and
tro
pic
alre
gio
ns.
Pip
ern
o(1
988,
2006
),W
ang
&
Lu
(199
3),
Kea
lho
fer
&P
iper
no
(199
8),
Th
orn
(200
4)
Vegetation and fire history from phytolith and charcoal records
Journal of Biogeography 35, 325–341 331ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd
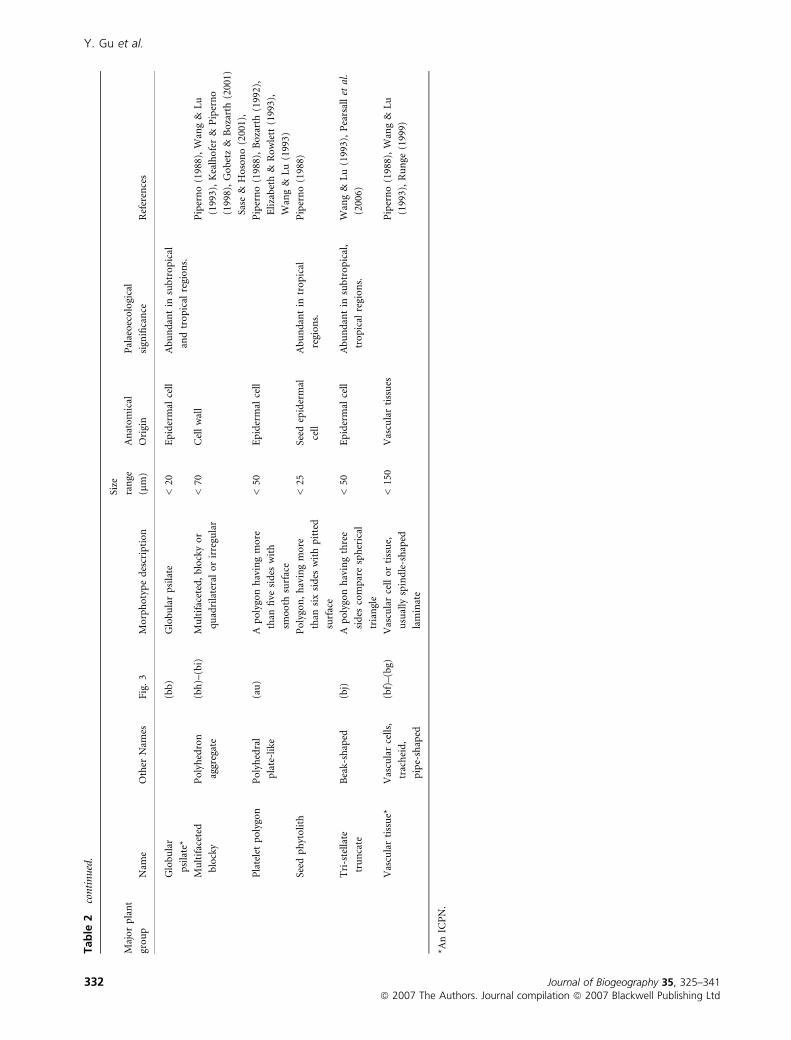
Tab
le2
con
tin
ued
.
Maj
or
pla
nt
gro
up
Nam
eO
ther
Nam
esF
ig.
3M
orp
ho
typ
ed
escr
ipti
on
Size
ran
ge
(lm
)
An
ato
mic
al
Ori
gin
Pal
aeo
eco
logi
cal
sign
ifica
nce
Ref
eren
ces
Glo
bu
lar
psi
late
*
(bb
)G
lob
ula
rp
sila
te<
20E
pid
erm
alce
llA
bu
nd
ant
insu
btr
op
ical
and
tro
pic
alre
gio
ns.
Mu
ltif
acet
ed
blo
cky
Po
lyh
edro
n
aggr
egat
e
(bh
)–(b
i)M
ult
ifac
eted
,b
lock
yo
r
qu
adri
late
ral
or
irre
gula
r
<70
Cel
lw
all
Pip
ern
o(1
988)
,W
ang
&L
u
(199
3),
Kea
lho
fer
&P
iper
no
(199
8),
Go
bet
z&
Bo
zart
h(2
001)
Sase
&H
oso
no
(200
1),
Pla
tele
tp
oly
gon
Po
lyh
edra
l
pla
te-l
ike
(au
)A
po
lygo
nh
avin
gm
ore
than
five
sid
esw
ith
smo
oth
surf
ace
<50
Ep
ider
mal
cell
Pip
ern
o(1
988)
,B
oza
rth
(199
2),
Eli
zab
eth
&R
ow
lett
(199
3),
Wan
g&
Lu
(199
3)
Seed
ph
yto
lith
Po
lygo
n,
hav
ing
mo
re
than
six
sid
esw
ith
pit
ted
surf
ace
<25
Seed
epid
erm
al
cell
Ab
un
dan
tin
tro
pic
al
regi
on
s.
Pip
ern
o(1
988)
Tri
-ste
llat
e
tru
nca
te
Bea
k-sh
aped
(bj)
Ap
oly
gon
hav
ing
thre
e
sid
esco
mp
are
sph
eric
al
tria
ngl
e
<50
Ep
ider
mal
cell
Ab
un
dan
tin
sub
tro
pic
al,
tro
pic
alre
gio
ns.
Wan
g&
Lu
(199
3),
Pea
rsal
let
al.
(200
6)
Vas
cula
rti
ssu
e*V
ascu
lar
cell
s,
trac
hei
d,
pip
e-sh
aped
(bf)
–(b
g)V
ascu
lar
cell
or
tiss
ue,
usu
ally
spin
dle
-sh
aped
lam
inat
e
<15
0V
ascu
lar
tiss
ues
Pip
ern
o(1
988)
,W
ang
&L
u
(199
3),
Ru
nge
(199
9)
*An
ICP
N.
Y. Gu et al.
332 Journal of Biogeography 35, 325–341ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

clavate contorted phytoliths (Bozarth, 1993; Wang & Lu, 1993;
Sase & Hosono, 2001; Carnelli et al., 2004; Stromberg, 2004).
Broad-leaved trees produce many kinds of phytoliths, inclu-
ding the following types: multifaceted blocky, platelet polygon,
abbreviated stellate, globular psilate, globular granulate, vas-
cular tissue (spindle laminate), globular facetate, tri-stellate
truncate, cylindrical sclereid, acicular hair cell (tree-type), and
seed phytoliths. The first three are abundant in deciduous trees
(Pearsall et al., 2006; Piperno, 1988; Bozarth, 1992; Wang &
Lu, 1993; Kealhofer & Piperno, 1998; Sase & Hosono, 2001;
Gobetz & Bozarth, 2001; Lu & Liu, 2005); the rest are
abundant in subtropical and tropical regions, and originate
mostly from ligneous dicots (Piperno, 1988; Barboni et al.,
1999).
Analysis of fire evidence
Direct evidence for fire was established by counting burnt
phytoliths and charcoal particles (see Appendix S1 in the
supplementary material). Burning is associated with surface
vegetation clearance and increased slope erosion on hills.
Increased erosion leads to increased and immediate transport
of phytoliths, which reduces the time for which phytoliths are
exposed in the source areas. The degree of weathering on large
phytoliths, such as bulliform cells, is therefore an indirect
indicator of fire events. Compared with fresh bulliform cells,
highly weathered bulliform cells have rebuilt edges and
surfaces marked with a number of cavities (see Appendix
S2). The frequency of occurrence of highly weathered bulli-
form cells should decrease in sediments formed during fire
episodes. As an indirect indicator, highly weathered bulliform
cells have a positive relationship with wet and warm condi-
tions, which suggests that they indicate the recovery of surface
vegetation and stable weathering processes.
Micro-charcoal particles in slides can be classified into two
size groups, namely 100–25 and < 25 lm. Micro-charcoals of
size 100–25 lm are abundant in fire episodes, whereas micro-
charcoals < 25 lm are abundant in pre-fire periods (see
Appendix S3). Micro-charcoal particles found in phytolith
slides, together with burnt phytoliths and highly weathered
bulliform cells, were counted for 20 fields at 250 · magni-
fication. Tallies of all phytoliths were made for the same
fields. Abundances of burnt phytoliths, highly weathered
bulliform cells, and micro-charcoal particles relative to the
total number of phytoliths counted for each sample were
calculated, providing a relative measure of the severity of the
fire episode.
RESULTS
Phytolith assemblages and indices
On the basis of phytolith identifications and phytolith indices,
six zones of phytolith assemblages can be discriminated and
interpreted in terms of climate and major vegetation constit-
uents (Figs 4 & 5).
Zone 1 (100–87 cm):
The Ic (climatic index) and Iph (aridity index) ratios vary
from 29% to 64%, and from 64% to 83%, respectively.
Poaceae phytoliths make up the greatest part of the
assemblage, varying from 67% to 76%, with bulliform cells
and long cells making the greatest contributions. The
proportion of fern and Asteraceae phytoliths increase within
Zone 1 together with Poaceae. Broad-leaved tree types occur
in reverse proportion to Poaceae phytoliths, decreasing from
16% to 10%. Globular granulate, acicular hair cells, vascular
tissues and sclereids are common broad-leaved trees types.
Gymnosperm phytoliths remain stable. It should be noted
that the short cells in this zone are present in relatively low
quantities, which might influence the high Iph value of 83%
(see Appendix S4).
Zone 2 (87–72 cm):
Poaceae phytoliths still make up the greatest part of the
assemblage. Bulliform cells, long cells, and Bambusoideae,
Panicoideae, and Pooideae short cells make the greatest
contributions. Broad-leaved tree phytoliths are present in
inverse proportion to the Poaceae phytoliths; globular granu-
late and sclereid forms are dominant, and multi-faceted
blocky, platelet polygon, and vascular tissues are common.
Ferns and Asteraceae decline slightly, together with Poaceae.
Gymnosperm phytoliths increase slightly.
Zone 3 (72–58 cm):
As in Zone 1, Poaceae phytoliths are dominant in assemblage
zone 3. Bulliform cells, long cells, and Bambusoideae, Arun-
dinoideae, and Pooideae short cells are abundant. Arundino-
ideae phytoliths increase, while Bambusoideae phytoliths
decrease. Broad-leaved tree phytoliths still occur in inverse
proportion to Poaceae phytoliths. Globular granulate, globular
psilate, and sclereids are dominant; multifaceted blocky,
globular facetate, acicular hair cell, and vascular tissues are
common. Fern and gymnosperm phytoliths decrease gradually.
Zone 4 (58–44 cm):
This zone is similar to Zone 2. Poaceae phytoliths are still
dominant. The assemblage of broad-leaved tree phytoliths is
dominated by globular granulate, globular psilate, multifaceted
blocky, and sclereids. Globular facetate, acicular hair cell, and
vascular tissues also occur.
Zone 5 (44–9 cm):
The Ic and Iph ratios are variable within this zone. The
majority of the Ic values are lower than 50%, and fluctuate
around 30%. The pattern is similar for the Iph ratio. Poaceae
phytoliths are dominant in this zone. Bulliform cells, long cells,
and Bambusoideae, Pooideae and Panicoideae short cells are
Vegetation and fire history from phytolith and charcoal records
Journal of Biogeography 35, 325–341 333ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

abundant; Chloridoideae and Arundinoideae short cells and
hair cell (grass-type) phytoliths are common. The percentage
of Pooideae and Asteraceae phytoliths changes in inverse
proportion to Panicoideae phytoliths. Chloridoideae and
Arundinoideae phytoliths occur in inverse proportion to
Bambusoideae phytoliths. Broad-leaved tree phytoliths fluctu-
ate in inverse proportion to Poaceae phytoliths. The broad-
leaved tree assemblage is dominated by globular granulate,
(a)
(o)
(v) (w) (x) (y) (z) (aa) (ab) (ac)
(ad) (ae) (af) (ag) (ah) (ai)
(aj) (ak) (al) (am) (an)
(ao) (ap) (aq) (ar) (as) (at)
(au) (av) (aw) (ax) (ay) (az) (ba) (bb)
(bc) (bd) (be) (bf) (bg) (bh)
(bl)(bk)(bj)(bi)
(p) (q) (r) (s) (t) (u)
(h)(g) (i) (j) (k) (l) (m) (n)
(b) (c) (d) (e) (f)
Y. Gu et al.
334 Journal of Biogeography 35, 325–341ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

globular psilate, acicular hair cell, and sclereids; and platelet
polygon, globular facetate, and vascular tissues are also present.
The percentage of sponge spicules is low, but diatoms occur
frequently in this zone, and approach the highest percentage
(5%) at 16 cm.
Zone 6 (9–0 cm):
The Ic and Iph ratios decrease gradually. Poaceae phytoliths
make up the greatest part of this assemblage zone; hair cells
and Chloridoideae and Arundinoideae short cells are com-
mon. Broad-leaved tree phytoliths change in inverse pro-
portion to Poaceae phytoliths, and globular granulate;
acicular hair cells, vascular tissues, and sclereids are com-
mon.
Fire episodes based on phytolith and charcoal
evidence
Macro-charcoal and micro-charcoal particles in the sediments
vary in shape from angular to irregular (see Appendix S1).
Most burnt phytoliths are from Poaceae, ferns, and broad-
leaved trees. These phytoliths contain black occluded carbon
(Piperno, 1988; Kealhofer & Penny, 1998), and reveal burning
directly. They are also occasionally slightly distorted in shape
(see Appendix S2). On the basis of charcoal and burnt
Figure 4 Percentage of main phytolith morphotypes and assemblages in sediments (count based on 10 fields at 400 ·).
Figure 3 Major phytolith morphotypes from geological sediments from the tropical rain forest in Xishuangbanna. Poaceae phytoliths (a)–
(al) and non-Poaceae phytoliths (am)–(bl) (the bar is 10 lm): (a)–(d) bilobates (Panicoideae type); (e), (g)–(h) crosses (Panicoideae type);
(f), (i)–(k) oblong concave saddles (Bambusoideae type); (l) Bambusoideae variant 3 bilobate; (m)–(n) trapeziform saddles (Arundinoideae
type); (o)–(s) square saddles (Chloridoideae type); (t)–(u) rondels (Pooideae type); (v)–(w) complex rondels (unknown origin); (x), (ab)
parallelepipedal bulliform cells; (y)–(aa) cuneiform bulliform cells; (ac) opaque perforated platelets (Asteraceae); (ad)–(ae) cylindrical
polylobates (Panicoideae type); (af)–(ag) trapeziform polylobates (Pooideae type); (ah) trapeziform sinuates (Pooideae type); (ai), (ak)
unciform hair cell (grass type); (aj) elongate smooth; (al) elongate echinate; (am)–(an) polyhedrons with conical projection (Cyperaceae,
Cyperus sp.) (Piperno, 1989); (ao) elongate undulating (fern type); (ap) prism sinuate (fern type); (aq)–(ar) parallelepipedal contorted
(gymnosperm type); (as)–(at) acicular hair cell (tree type); (au) platelets polygon (broad-leaved tree type); (av)–(aw) abbreviated stellate
(broad-leaved tree type); (ax)–(ba) globular granulate (broad-leaved tree type); (bb) globular psilate (broad-leaved tree type); (bc) clavate
contorted phytoliths (gymnosperm type); (bd)–(be) globular facetates (broad-leaved tree type); (bf)–(bg) vascular tissues (broad-leaved tree
type); (bh)–(bi) multifaceted blocky (broad-leaved tree type); (bj) tri-stellate truncate (broad-leaved tree type); (bk)–(bl) cylindrical sclereid
(broad-leaved tree type).
Vegetation and fire history from phytolith and charcoal records
Journal of Biogeography 35, 325–341 335ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

phytolith occurrences, 12 episodes and six zones of fire events
can be identified in the profile (Fig. 6).
Episode 1:
The percentage of burnt phytoliths decreases gradually from 90%
to 70%, from the start to the end of the episode, corresponding to
Zone 1. The number of micro-charcoal particles decreases
gradually from 130 to 65 particles, corresponding to the decrease
in burnt phytoliths. The percentage of highly weathered bulli-
form cells changes in inverse proportion to the percentage of
burnt phytoliths, fluctuating from 5% to 10%.
Episode 2:
Corresponding to Zone 2, the percentage of burnt phytoliths
decreases rapidly within this zone; there is a peak at 80 cm,
corresponding to fire Episode 2. Micro-charcoal particles show
a similar pattern. The percentage of highly weathered bulliform
cells is again inversely proportional to the percentage of burnt
phytoliths.
Episodes 3–4:
Corresponding to Zone 3, the percentage of burnt phytoliths
and the sum of micro-charcoal particles increase rapidly in this
zone. The percentage of highly weathered bulliform cells
decreases in inverse proportion to the percentage of burnt
phytoliths.
Episode 5:
As in Episode 2, the percentages of burnt phytoliths and micro-
charcoal particles decrease rapidly in Zone 4. There is a peak at
Figure 6 Evidence of fire occurrence and
positive feedbacks between drought, fire and
vegetation composition.
Figure 5 The abundance of grass sub-
families and results for phytolith indices.
Y. Gu et al.
336 Journal of Biogeography 35, 325–341ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

50 cm, which corresponds to fire episode 5. The percentage of
highly weathered bulliform cells increases in inverse propor-
tion to the percentage of burnt phytoliths.
Episodes 6–11:
The percentages of burnt phytoliths, charcoal, and highly
weathered bulliform cells show frequent and rapid variability.
Burnt phytoliths have six evident peaks varying from 50% to
96%, corresponding to fire episodes 6–11. The sum of
micro-charcoal particles changes in concert with the burnt
phytoliths. The percentage of highly weathered bulliform
cells is inversely proportional to the percentage of burnt
phytoliths.
Episode 12:
The percentage of burnt phytoliths and charcoal declines
rapidly; there is a single peak at 6 cm, corresponding to fire
episode 12. The percentage of highly weathered bulliform cells
again demonstrates an inverse relationship with the percentage
of burnt phytoliths (Fig. 7a).
Nearly every fire episode involved in the burning of grasses
and trees, and burnt phytoliths occur mostly in plant taxa from
the Poaceae, ferns, and broad-leaved trees. This is critical in
determining the intensity and range of fire episodes in the
study area.
DISCUSSION
Climate change and palaeoenvironmental analysis
Previous studies have demonstrated that the aridity (Iph) and
climate (Ic) indices have significant potential as climatic
indicators for sedimentary palaeoenvironmental interpretation
(Twiss, 1987, 1992; Barboni et al., 1999; Parker et al., 2004).
Mean annual precipitation, together with mean annual tem-
perature, has a decisive influence on the spatial distributions of
phytolith types in the study area (Lu et al., 2006). Our results
are consistent with modern soil phytolith assemblages in
southern tropical China (Lu et al., 2006): bulliform cells
(except for fan-reed), long saddles (oblong concave saddle),
Palmae (Arecaceae, palm family), and broad-leaf types are very
abundant; dumbbell (bilobate), elongate, point-shaped (hair
cell), pteridophyte (fern) and gymnosperm types are moder-
ately abundant; and short saddles (square saddle) and
Pooideae are rare (Fig. 4). An Iph value of 30% was selected
here as the boundary for humidity–aridity, consistent with the
approach used in intertropical Africa (Alexandre et al., 1997).
On this basis, the humidity history of the study area is
characterized by alternating wet and dry intervals over the past
2000 years. In particular, 11 drought events have been
identified (a1 to a11 in Fig. 5), which might be related to
decreased precipitation from the Indian summer monsoon
(Denniston et al., 2000). Comparable with the Iph index, the
Ic index also indicates a trend of fluctuating warmth-coolness,
with 11 cold events (temperature below average) observed in
the study region over the past 2000 years (Fig. 5). The majority
of the Ic values are < 50%, and the relatively high Ic value
(> 50%) could be explained by the presence of Sporobolus,
which produces Pooideae-type phytoliths even though it
belongs to the Chloridoideae group (Barboni et al., 1999).
Comparable fluctuations in regional climate have been docu-
mented during the late Pleistocene–early Holocene in
Xishuangbanna (Liu et al., 1986; Walker, 1986). Modern
meteorological data from Mengla County also show that,
during a 40-year interval, 25% of years experienced low
temperatures (Tan et al., 2002). Moreover, historical docu-
ments and lake sediments in the neighbouring areas (e.g.
Kunming and Erhai) produce markedly fluctuating tempera-
ture records for the last 2000 years (Sun et al., 1986; Zhang
et al., 2001), which is consistent with the phytolith record
reported herein. Our results demonstrate the significant
potential of phytoliths for identifying subtle climate changes
on a short time-scale.
To summarize, our phytolith records document six climate
periods marked by occurrence of drought events and cold
events. Zone 1 is marked by cool and dry conditions, with the
presence of Pooideae, Chloridoideae, Bambusoideae, and
Asteraceae. Zone 2 is characterized by warm and wet condi-
tions, with the presence of Panicoideae, Pooideae, and
Bambusoideae. In addition, a brief cold and dry interval
occurs at 80 cm. Zone 3 is marked by cool–dry conditions,
with the presence of Pooideae, Chloridoideae, Bambusoideae,
(a) (b)
Figure 7 Correlation analysis among burnt
phytoliths, highly weathered bulliform cells
and Iph (aridity index): (a) burnt phytoliths
are negatively correlated with highly weath-
ered bulliform cells; (b) burnt phytoliths are
positively correlated with Iph.
Vegetation and fire history from phytolith and charcoal records
Journal of Biogeography 35, 325–341 337ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

Panidoideae, Arundinoideae, ferns, and Asteraceae. There is a
relatively wet interval at 65 cm, which is the lithological
boundary between silt and clay in the profile. Zone 4 is
dominated by warm and wet conditions, with the presence of
Panicoideae, Pooideae, Bambusoideae and Arundinoideae,
corresponding to the Medieval Warm Period in the Erhai
area (Zhang et al., 2001). Within this zone, there are brief cold
and dry intervals at about 50 cm. Zone 5 is dominated by
alternating cold–dry and warm–wet intervals, corresponding to
the Little Ice Age in the neighbouring areas (Sun et al., 1986;
Zhang et al., 2001). Zone 6 is characterized by rapid warming
and wet conditions, with the presence of Panicoideae, Pooideae,
Bambusoideae, and Arundinoideae. There is a cold event at
6 cm. The rapid warming and wet conditions of the most recent
zone suggest that rain forest is sensitive to global warming. It is
evident that Panicoideae, Bambusoideae, Cyperaceae, sponge
spicules, and diatoms occur in warm and wet conditions.
However, Pooideae and Asteraceae exist in cool–dry or
cool–wet circumstances (Figs 4 & 5).
Drought, fire and vegetation succession
Modern species composition, physiognomy and plant diversity
surveys show that trees dominate rain forest and have a higher
abundance and diversity than shrubs and herbs (Zhu et al.,
1999; Zhu & Cai, 2005; Li et al., 2004). Rain-forest compo-
sition and structure might change and grasses increase rapidly
in an open habitat after a fire occurrence and disturbance.
Owing to recurring fire episodes, grasses (Poaceae) and ferns
increase as a result of recurring episodes of fire caused by
drought and other factors at the expense of broad-leaved and
gymnosperm trees. In contrast, broad-leaved and gymnosperm
trees increase during pre-fire and wet conditions at the expense
of grasses and ferns. The vegetation structure of the forest is
thus disturbed again and again by the fire regime. It is
therefore likely that both drought and fire, the latter in
particular, have a significant influence on vegetation compo-
sition and structure in the study area. A similar kind of
relationship is observed today in the Amazonian rain forest,
where modern drought and fire have damaged the forest,
causing fragmentation, forest loss, and climate change (Will-
iamson et al., 2000; Laurance & Williamson, 2001).
It is noteworthy that nearly all fire episodes identified in the
profile are associated with evidence for droughts (Fig. 6). The
significant positive correlation between the Iph index and
burnt phytoliths (Fig. 7b) sheds further light on the relation-
ship between fire and drought. More significantly, the relative
abundances of vegetation elements such as grasses, gymno-
sperms, and broad-leaved trees show positive feedbacks to pre-
fire and fire episodes over the past 2000 years.
It should be noted that a fire episode occurs at 30 cm in
association with a relatively low Iph value of 25%. This implies
that fire can occur when the Iph index approaches 25%, and that
this value may be a cut-off value for this region. An exceptional
fire episode, Episode 12, occurs at 6 cm, in association with
decreasing abundances of both grasses and trees. This might
record human interference in the rain forest, corresponding to
the rapid growth of local populations during the 19th and 20th
centuries (Lee, 1982; Hansen, 1999; Fukao, 2004).
CONCLUSIONS
This first phytolith and charcoal record from tropical rain-
forest sediments in south-western China provides an excellent
framework for analysing the interaction between climate, fire,
and vegetation history over the last 2000 years. Our main
findings are as follows.
1. Phytolith assemblages and indices indicate that the region
has experienced six periods of climate change marked by
alternating warm–wet and cool–dry conditions. Eleven
drought events and cold events occurred. Recent rapid
warming and wet conditions indicate that rain forest is
sensitive to global warming.
2. Burnt phytoliths, highly weathered bulliform cells, and
charcoal particles reveal the occurrence of 12 fire episodes.
3. Fire apparently influences drought occurrence and exerts a
strong influence on vegetation structure. Nearly every fire
episode, coupled with a climatic shift, triggered rapid changes
in the composition of the flora. Grasses expanded in response
to an increased occurrence of fire, which confirms the
expectation that drought interacts with fire to affect rain-
forest ecosystems. In the last fire episode, grasses and trees
both declined. This suggests that human activity in recent
centuries has affected fire occurrence and vegetation.
ACKNOWLEDGEMENTS
This project was funded by the National Natural Science
Foundation of China (grants 40502015, 40232025 and
40525008). The authors are grateful for the assistance of the
Xushuangbanna Tropical Botanical Garden of the Chinese
Academy of Sciences, and for helpful information and sugges-
tions provided by Professor Zhu Hua and Mr Wang Hong. We
thank to Professor Yang Fengqing and Dr Wang Hongmei for
help with the fieldwork. We are grateful to Professor Zhou
Xiugao and master candidate Qin Yangmin, who helped with
sediment laboratory processing. Many thanks should be given
to the referees for their valuable comments and constructive
suggestions on an earlier version of the manuscript.
REFERENCES
Alexandre, A., Meunier, J.D., Lezine, A.M., Vincens, A. &
Schwartz, D. (1997) Phytoliths: indicators of grassland
dynamics during the late Holocene in intertropical Africa.
Palaeogeography, Palaeoclimatology, Palaeoecology, 136, 213–
229.
Barboni, D., Bonnefille, R., Alexandre, A. & Meunier, J.D.
(1999) Phytoliths as palaeoenvironmental indicators, West
Side Middle Awash Valley, Ethiopia. Palaeogeography,
Palaeoclimatology, Palaeoecology, 152, 87–100.
Y. Gu et al.
338 Journal of Biogeography 35, 325–341ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

Bozarth, S.R. (1992) Classification of opal phytoliths formed in
selected dicotyledons native to the Great Plains. Phytolith
systematics: emerging issues (ed. by G. Rapp Jr and S.C. Mul-
holland), pp. 193–214. Plenum Press, New York.
Bozarth, S.R. (1993) Biosilicate assemblages of boreal forests
and aspen. Current research in phytolith analysis: applications
in archaeology and palaeoecology, Volume 10 (ed. by D.M.
Pearsall and D.R. Piperno), pp. 95–105. MASCA, The
University Museum of Archaeology and Anthropology,
University of Pennsylvania, Philadelphia.
Bremond, L., Alexandre, A., Hely, C. & Guiot, J. (2005) A
phytolith index as a proxy of tree cover density in tropical
areas: calibration with Leaf Area Index along a forest-
savanna transect in southeastern Cameroon. Global and
Planetary Change, 45, 277–293.
Brown, D.A. (1984) Prospects and limits of a phytolith key for
grasses in the central United States. Journal of Archaeological
Science, 11, 345–368.
Cao, M. & Zhang, J.H. (1997) Tree species diversity of tropical
forest vegetation in Xishangbanna, SW China. Biodiversity
and Conservation, 6, 995–1006.
Carnelli, A.L., Theurillat, J.P. & Madella, M. (2004) Phytolith
types and type-frequencies in subalpine-alpine plant species
of the European Alps. Review of Palaeobotany and Palynol-
ogy, 129, 39–65.
Carter, J.A. (2002) Phytolith analysis and palaeoenvironmental
reconstruction from Lake Poukawa Core, Hawkes Bay, New
Zealand. Global and Planetary Change, 33, 257–267.
Denniston, R.F., Gonzalez, L.A., Asmerom, Y., Sharma, R.H. &
Reagan, M.K. (2000) Speleothem evidence for changes in
Indian Summer Monsoon precipitation over the last
2300 years. Quaternary Research, 53, 196–202.
Diester-Haass, L., Schrader, H.J. & Thiede, J. (1973) Sedi-
mentological and palaeoclimatological investigations of two
pelagic-ooze cores off Cape Barbas, North-West Africa.
Meteor, Forschung-Ergebnisse, 16, 19–66.
Elizabeth, H.D. & Rowlett, R.M. (1993) Vegetation changes at
the Shriver Palaeo-Indian site, NW Missouri: phytolith
analysis as an aid in environmental reconstruction. Current
research in phytolith analysis: applications in archaeology and
palaeoecology, Volume 10 (ed. by D.M. Pearsall and D.R.
Piperno), pp. 73–82. MASCA, The University Museum of
Archaeology and Anthropology, University of Pennsylvania,
Philadelphia.
Fredlund, G.G. & Tieszen, L.T. (1994) Modern phytolith
assemblages from the North American Great Plains. Journal
of Biogeography, 21, 321–335.
Fredlund, G.G. & Tieszen, L.T. (1997) Calibrating grass
phytolith assemblages in climatic terms: application to late
Pleistocene assemblages from Kansas and Nebraska.
Palaeogeography, Palaeoclimatology, Palaeoecology, 136, 199–
211.
Fukao, Y. (2004) Socioecological transformation triggered by
national rubber plantations in Yunnan, China: the impact of
Han-Chinese immigration into Xishuangbanna since the
1950s. Southeast Asian Studies, 42, 294–327.
Gobetz, K.E. & Bozarth, S.R. (2001) Implications for late
Pleistocene mastodon diet from opal phytoliths in tooth
calculus. Quaternary Research, 55, 115–122.
Hansen, M.H. (1999) The call of Mao or money? Han Chinese
setters on China’s Southwestern borders. The China Quar-
terly, 158, 394–413.
Harrison, R.D. (2000) Repercussions of El Nino: drought
causes extinction and the breakdown of mutualism in Bor-
neo. Proceedings of the Royal Society B: Biological Sciences,
267, 911–915.
Harrison, R.D. (2001) Drought and the consequences of El
Nino in Borneo: a case study of figs. Population Ecology, 43,
63–75.
Kaplan, L., Smith, M.B. & Sneddon, L.A. (1992) Cereal grain
phytoliths of Southwest Asian and Europe. Phytolith sys-
tematics: emerging issues (ed. by G. Rapp Jr and S.C. Mul-
holland), pp. 149–174. Plenum Press, New York.
Kealhofer, L. & Penny, D. (1998) A combined pollen and
phytolith record for fourteen thousand years of vegetation
change in northeastern Thailand. Review of Palaeobotany
and Palynology, 103, 83–93.
Kealhofer, L. & Piperno, D.R. (1998) Opal phytoliths in
Southeastern Asian Flora. Smithsonian Contributions to
Botany, 88, 1–39.
Kellman, M. & Meave, J. (1997) Fire in the tropical gallery
forests of Belize. Journal of Biogeography, 24, 23–34.
Laurance, W.F. & Williamson, G.B. (2001) Positive feedbacks
among forest fragmentation, drought, and climate change in
the Amazon. Conservation Biology, 15, 1529–1535.
Laurance, W.F., Delamonica, P., Laurance, S.G., Vasconcelos,
H.L. & Lovejoy, T.E. (2000) Rainforest fragmentation kills
big trees. Nature, 404, 836.
Lee, J. (1982) Food supply and population growth in south-
west China. The Journal of Asian Studies, 41, 711–746.
Li, Z.S., Tang, J.W., Zheng, Z., Li, Q.J., Luo, C.K., Liu, Z.A., Li,
Z.N., Duan, W.Y. & Guo, X.M. (2004) A study on plant
diversity of tropical montane rain forests in Xishuangbanna,
Yunnan. Acta Phytoecologica Sinica, 28, 833–843 (in Chinese
with English abstract).
Li, Q., Xu, D.K. & Lu, H.Y. (2005) Morphology of phytolith in
Bambusoideae (Gramineae) and its ecological significance.
Quaternary Sciences, 25, 777–784 (in Chinese with English
abstract).
Liu, W.J. & Li, H.M. (1996) Climatic feature, tendency and its
influence on tropical plants in Menglun region. Tropical
Plant Research, 38, 16–22 (in Chinese).
Liu, J.L., Tan, L.Y., Qiao, Y.L., Head, M.J. & Walker, D. (1986)
Late Quaternary vegetation history at Menghai, Yunnan
province, southwest China. Journal of Biogeography, 13, 399–
418.
Lu, H.Y. & Liu, K.B. (2003a) Morphological variations of
lobate phytoliths from grasses in China and the southeastern
USA. Diversity and Distributions, 9, 73–87.
Lu, H.Y. & Liu, K.B. (2003b) Phytoliths of common grasses in
the coastal environments of southeastern USA. Estuarine,
Coastal and Shelf Science, 58, 587–600.
Vegetation and fire history from phytolith and charcoal records
Journal of Biogeography 35, 325–341 339ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

Lu, H.Y. & Liu, K.B. (2005) Phytolith assemblages as indicators
of coastal environmental changes and hurricane overwash
deposition. The Holocene, 15, 965–972.
Lu, H.Y., Wu, N.Q., Yang, X.D., Jiang, H., Liu, K.B. & Liu,
T.S. (2006) Phytoliths as quantitative indicators for the
reconstruction of past environmental conditions in China I:
phytolith-based transfer functions. Quaternary Science
Reviews, 25, 945–959.
Madella, M., Alexandre, A. & Ball, T. (ICPN Working Group)
(2005) International code for phytolith nomenclature 1.0.
Annals of Botany, 96, 253–260.
Marchant, R. (2005) Special issue: late Quaternary tropical
ecosystem dynamics. Quaternary Research, 64, 283.
Mercader, J., Runge, F., Vrydaghs, L., Doutrelepont, H.,
Ewango, C.E.N. & Juan-Tresseras, J. (2000) Phytoliths
from Archaeological Sites in the Tropical Forest of Ituri,
Democratic Republic of Congo. Quaternary Research, 54,
102–112.
Mulholland, S.C. (1989) Phytoliths shape frequencies in North
Dakota grasses: a comparison to general patterns. Journal of
Archaeological Science, 16, 489–511.
Mulholland, S.C. & Rapp, G., Jr (1992) A morphological
classification of grass silica-bodies. Phytolith systematics:
emerging issues (ed. by G. Rapp, Jr and S.C. Mulholland), pp.
65–90. Plenum Press, New York.
van Nieuwstadt, M.G.L. & Sheil, D. (2005) Drought, fire and
tree survival in a Borneo rain forest, East Kalimantan,
Indonesia. Jounal of Ecology, 93, 191–201.
Ollendorf, A.L. (1992) Towards a classification scheme of
Sedge (Cyperaceae) phytoliths. Phytolith systematics: emer-
ging issues (ed. by G. Rapp Jr and S.C. Mulholland), pp. 91–
106. Plenum Press, New York.
Parker, A.G., Eckersly, L., Smith, M.M., Goudie, A.S.,
Stokes, S., Ward, S., White, K. & Hodson, M.J. (2004)
Holocene vegetation dynamics in the northeastern Rub’al-
Khali desert, Arabian peninsula: a phytolith, pollen and
carbon isotope study. Journal of Quaternary Science, 19,
665–676.
Pearsall, D.M. (2000) Palaeoethnobotany: a handbook of pro-
cedures, 2nd edn. Academic Press, San Diego.
Pearsall, D.M. & Elizabeth, H.D. (1992) Developing a phytolith
classification system. Phytolith systematics: emerging issues
(ed. by G. Rapp, Jr and S.C. Mulholland), pp. 37–64. Ple-
num Press, New York.
Pearsall, D.M., Biddle, A., Chandler-Ezell, K., Collins, S.,
Duncan, N., Logan, A., O’Brien, M., Stewart, S., Veintimilla,
C., Zhao, Z.J. & Grimm, B. (2006) Phytoliths in the Flora of
Ecuador: the University of Missouri Online Phytolith
Database (Website: http://www.missouri.edu/~phyto).
Piperno, D.R. (1988) Phytolith analysis: an archaeological and
geological perspective. Academic Press, San Diego.
Piperno, D.R. (1989) The occurrence of phytoliths in the
reproductive structures of selected tropical angiosperms and
their significance in tropical paleoecology, paleoethnobota-
ny, and systematics. Review of Palaeobotany and Palynology,
61, 147–173.
Piperno, D.R. (2006) Phytoliths: a comprehensive guide to
archaeologists and palaeoecologists. Altamira Press, Lanham,
MD.
Piperno, D.R. & Pearsall, D.M. (1998) The silica bodies of
tropical American grasses: morphology, taxonomy, and
implications for grass systematics and fossil phytolith
identification. Smithsonian Contributions to Botany, 85, 1–
40.
Potts, M.D. (2003) Drought in a Bornean everwet rain forest.
Journal of Ecology, 91, 467–474.
Pu, Y.S., Zhang, J.H. & Cao, M. (2001) A strategic study on
biodiversity conservation in Xishuangban. Journal of Forestry
Research, 12, 25–30.
Runge, F. (1999) The opal phytolith inventory of soils in
central Africa - quantities, shapes, classification, and spectra.
Review of Palaeobotany and Palynology, 107, 23–53.
Sase, T. & Hosono, M. (2001) Phytolith record in soils inter-
stratified with late Quaternary tephras overlying the eastern
region of Towada volcano, Japan. Phytoliths: application in
earth sciences and human history (ed. by J.D. Meunier and
F. Colin), pp. 58–71. A.A. Balkema Publishers, Rotterdam.
Stromberg, C.A.E. (2004) Using phytolith assemblages to
reconstruct the origin and spread of grass-dominated hab-
itats in the great plains of North America during the late
Eocene to early Miocene. Palaeogeography, Palaeoclimatolo-
gy, Palaeoecology, 207, 239–275.
Stuiver, M. & Reimer, P.J. (1993) Extended 14C data base and
revised CALIB 3.0 14C age calibration program. Radiocarbon,
35, 215–230.
Sun, X.J., Wu, Y.S., Qiao, Y.L. & Walker, D. (1986) Late
Pleistocene and Holocene vegetation history at Kunming,
Yunnan Province, southwest China. Journal of Biogeography,
13, 441–476.
Tan, Y.Z., Gao, X.S., Huang, W. L., Deng, Y. & Pu, G.M.
(2002) Analysis of chilling injury weather and the disaster-
reducing countermeasures in Xishuangbanna. Chinese
Agriculture Meteorology, 23, 44–48 (in Chinese with English
abstract).
Tang, L.Y. (1992) Vegetation and climate history at Menghai,
Yunnan during the past 42,000 years. Acta Micropalaeon-
tology Sinica, 9, 433–455 (in Chinese with English sum-
mary).
Thorn, V.C. (2004) Phytoliths from subantarctic Campbell
Island: plant production and soil surface spectra. Review of
Palaeobotany and Palynology, 132, 37–59.
Twiss, P.C. (1987) Grass opal phytoliths as climatic indicators
of the Great Plains Pleistocene. Quaternary environments of
Kansas (ed. by W.C. Johnson), pp. 79–188. Kansas Geolo-
gical Survey, guidebook series, Kansas.
Twiss, P.C. (1992) Predicted world distribution of C3 and C4
grass phytoliths. Phytolith systematics: emerging issues (ed. by
G. Rapp, Jr and S.C. Mulholland), pp. 13–128. Plenum
Press, New York.
Twiss, P.C., Suess, E. & Smith, R.M. (1969) Morphological
classification of grass phytoliths. Soil Science Society of
America Proceedings, 33, 109–115.
Y. Gu et al.
340 Journal of Biogeography 35, 325–341ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd

Walker, D. (1986) Late Pleistocene-early Holocene vegetation
and climatic changes in Yunnan Provinces, southwest Chi-
na. Journal of Biogeography, 13, 477–486.
Walsh, R.P.D. (1996) Climate. The tropical rain forest, 2nd edn.
by P.W. Richards), pp. 159–205. Cambridge University
Press, Cambridge.
Walsh, R.P.D. & Newbery, D.M. (1999) The ecoclimatology of
Danum, Sabah, in the context of the world’s rainforest re-
gions, with particular reference to dry periods and their
impact. Philosophical Transactions of the Royal Society B:
Biological Sciences, 354, 1869–1883.
Wang, Y.J. & Lu, H.Y. (1993) Phytolith Study and its Appli-
cation. China Ocean Press, Beijing (in Chinese).
Williamson, G.B., Laurance, W.F., Oliveira, A.A., Delamonica,
P., Gascon, C., Lovejoy, T.E. & Pohl, L. (2000) Amazonian
tree morality during the 1997 El Nino drought. Conservation
Biology, 14, 1538–1542.
Woods, P. (1989) Effects of logging, drought and fire on
structure and composition of tropical forests in Sabah,
Malaysia. Biotropica, 21, 290–298.
Zhang, K.Y. (1988) The climatic dividing line between SW and
SE monsoon and their differences in climatology and ecol-
ogy in Yunnan Province of China. Climatological Notes, 38,
197–207.
Zhang, Z.K., Wu, R.J., Shen, J., Pan, H.X., Xia, W.L. & Wu,
Y.H. (2001) Lacustrine records showing climatic changes in
Erhai lake, Yunnan Province since the past 2000 years.
Marine Geology and Quaternary Geology, 21, 31–35 (in
Chinese with English abstract).
Zhu, H. (1997) Ecological and biogeographical studies on the
tropical rain forest of south Yunnan, SW China with a
special reference to its relation with rain forests of tropical
Asia. Journal of Biogeography, 24, 647–662.
Zhu, H. & Cai, L. (2005) Biogeography of the tropical rain
forest of Yunnan and some implications to geological his-
tory. Advances in Earth Science, 20, 1–80 (in Chinese with
English abstract).
Zhu, H., Wang, H., Li, B.G. & Xu, Z.F. (1998) Research on the
tropical seasonal rainforest of Xishuangbanna, South Yun-
nan. Guihaia, 18, 371–384 (in Chinese with English
abstract).
Zhu, H., Wang, H., Li, B.G. & Xu, Z.H. (1999) Tropical
vegetation types and urgent conservation for plant diversity
in Xishuangbanna, Yunnan. Tropical Plant Research, 44,
1–10 (in Chinese).
SUPPLEMENTARY MATERIAL
The following supplementary material is available for this
article online:
Appendix S1 Macro-charcoal, micro-charcoal and burnt
phytoliths in sediments.
Appendix S2 Burnt phytoliths, highly weathered bulliform
cells and other silicon remains.
Appendix S3 Statistics of burnt phytoliths, highly weathered
bulliform cells and micro-charcoals in 20 fields at 250 · mag-
nification expressed as percentages.
Appendix S4 Statistics of phytolith types expressed as
percentages.
This material is available as part of the online article from:
http://www.blackwell-synergy.com/doi/abs/10.1111/j.1365-2699.
2007.01763.x
Please note: Blackwell Publishing are not responsible for the
content or functionality of any supplementary materials sup-
plied by the authors. Any queries (other than missing material)
should be directed to the corresponding author for the article.
BIOSKETCHES
Yansheng Gu has a PhD from Wuhan University. Currently
he is an associate professor at the Institute of Ecology and
Environment, China University of Geosciences. He is interes-
ted in Quaternary geology and environmental changes in
southern China.
Deborah Pearsall has a PhD from the University of Illinois.
Currently she is a professor in the Department of Anthropol-
ogy, director of the Palaeoethnobotany Laboratory, University
of Missouri-Columbia, and a palaeoethnobotanist specializing
in macro-remain analysis, phytolith analysis, and starch-grain
analysis.
Shucheng Xie has a PhD from the China University of
Geosciences. Currently he is a professor at the Key Laboratory
of Biogeology and Environmental Geology, China University
of Geosciences. He is interested in Quaternary vegetation and
climate change.
Editor: Peter Linder
Vegetation and fire history from phytolith and charcoal records
Journal of Biogeography 35, 325–341 341ª 2007 The Authors. Journal compilation ª 2007 Blackwell Publishing Ltd
Top Related
Copyright © 2022 FDOKUMEN

