







Bahasa
Halaman
Hukum


ORIGINAL PAPER
S-genotyping of old apple cultivars from the Carpathianbasin: methodological, breeding and evolutionary aspects
Julia Halász & Attila Hegedűs & Zsuzsanna György &
Éva Pállinger & Magdolna Tóth
Received: 1 February 2011 /Revised: 27 March 2011 /Accepted: 11 May 2011 /Published online: 14 June 2011# Springer-Verlag 2011
Abstract Apple exhibits self-incompatibility controlledby the multiallelic S-locus. Twenty-three old applecultivars were S-genotyped using three differentapproaches (allele-specific polymerase chain reaction(PCR)+ cleaved amplified polymorphic sequences(CAPS), consensus PCR+sequencing and consensusPCR+CAPS) to compare the robustness and reliability ofthese techniques and characterise genotypes from theCarpathian basin that might be useful in resistancebreeding. Best results were obtained using the ASPF3and ASPR3S consensus primer pair that detected 96% ofall alleles carried by the 23 cultivars tested. Flowcytometry analysis was also needed to control thecompleteness of the genotypes as was seen in case of atetraploid cultivar with only three assigned S-alleles. Thegenetic disparity between the old Carpathian basin andmodern apple cultivars was indicated by differences inallele frequency data (S9, S24 and S26) as well as single
nucleotide polymorphisms in S1, S2, S7 S24 and S26 andindels in S20 and S26 alleles. An alignment of partialgenomic sequences indicated trans-specific and trans-generic evolution of S-ribonuclease alleles in the Maloi-deae subfamily (S26 and S28) and a possibly recentintrogression event (S1) between Malus×domestica andMalus sylvestris. These data suggest that the genome ofold cultivars from the Carpathian basin was enriched byseveral Malus taxa and are free from the consequences ofmodern breeding. These cultivars may contribute to thewidening of the genetic basis of cultivated apple andprevent genetic erosion in future commercial cultivars.
Keywords Apple . Ploidy . S-RNase . S-allele diversity .
Self-incompatibility . S-genotyping . SNP
Introduction
Similarly to most fruit trees in the Rosaceae family, applerequires cross-pollination for adequate fruit set since self-fertilisation is inhibited by gametophytic self-incompatibility.This is a genetically controlled mechanism enabling styles toreject self-pollen (de Nettancourt 2001). When the pollen S-haplotype matches one of those carried by the pistil tissue,pollen tube growth is arrested and fertilisation fails. This traitis governed by the highly polymorphic, multiallelic S-locus,in which the pistil component is an S-ribonuclease protein(S-RNase) (McClure et al. 1989). S-genotyping is a crucialtask for fruit trees especially for species like apple, whereself-compatible cultivars are unavailable to commercialgrowing (Bošković and Tobutt 1999; Broothaerts 2003).Knowledge on the S-genotypes of cultivars is required forchoosing appropriate pollen donors in commercial orchardsand breeding programs.
Communicated by E. Dirlewanger
Electronic supplementary material The online version of this article(doi:10.1007/s11295-011-0401-7) contains supplementary material,which is available to authorized users.
J. Halász (*) :A. Hegedűs : Z. GyörgyDepartment of Genetics and Plant Breeding,Corvinus University of Budapest,PO Box 53, Budapest 1518, Hungarye-mail: [email protected]
É. PállingerResearch Group for Inflammation Biology and Immunogenomicsof Hungarian Academy of Sciences and Semmelweis University,PO Box 370, Budapest 1445, Hungary
M. TóthDepartment of Pomology, Corvinus University of Budapest,PO Box 53, Budapest 1518, Hungary
Tree Genetics & Genomes (2011) 7:1135–1145DOI 10.1007/s11295-011-0401-7

First report on the self-incompatibility of applecultivars was written by Kobel et al. (1939), whodescribed 11 S-alleles (S1–S11) by monitoring pollen tubegrowth after controlled pollinations. Molecular S-genotyp-ing of apple cultivars was initiated by Sassa et al. (1994)using two-dimensional polyacrylamide gel electrophoresisanalyses for the identification of 6 S-alleles labelled byalphabetical codes (Sa–Sf). This was later extended bythree additional alleles (Sg–Si) (Komori et al. 2000). Then,S-genotypes of the 14 diploid cultivars proposed in theclassic work of Kobel et al. (1939) were confirmed byBošković and Tobutt (1999) using non-equilibrium pH gelelectrofocusing and isoelectric focusing. They proposed14 novel S-RNase alleles (S12–S25). The first DNAsequence data for apple S-RNases were reported by aBelgian research group who sequenced fragments from 15S-RNase alleles (Broothaerts 2003; Broothaerts et al.1995; Janssens et al. 1995; Van Nerum et al. 2001).Similarly, eight alleles were sequenced in Japan (Kitaharaand Matsumoto 2002a, b; Kitahara et al. 2000; Matsumotoand Kitahara 2000; Sassa et al. 1996). Additional alleles,S29 (Matityahu et al. 2005), S31 and S32 (Kim et al. 2008),S33–S41 (Li et al., unpublished data; Zhang et al.,unpublished data), S42–S43 (Dreesen et al. 2010) andS44–S46 (Long et al. 2010) were also identified.
Until now, several genotyping methods based on theuse of S-allele specific primers were published (Broothaerts2003; Broothaerts et al. 2004; Janssens et al. 1995;Kitahara et al. 2000; Matsumoto et al. 2007). However,specific primer-based methods could sometimes generatefalse-negative/positive results (Ershadi and Talaii 2007;Van Nerum et al. 2001). Matsumoto and Kitahara (2000)designed a consensus primer pair for the conservedregions of the S-RNase gene, which could simultaneouslyamplify nine alleles. Later, Kim et al. (2006, 2009)developed a new pair of consensus primers and a cleavedamplified polymorphic sequences (CAPS) system for theidentification of 22 apple S-alleles. It could overcome thefalse results; however, genomic sequences of a few appleS-RNase alleles (S8, S15 and S18) are not available inpublic databases.
Since breeding of new disease-resistant cultivarsremains an important objective, the identification oflocally adapted resistant genotypes are of great value inbreeding programs. For breeding and cultivation, cross-(in)compatibility relationships are very important both forsuccessful mating and reliable fruit set. Until now, a largenumber of apple cultivars have been S-genotyped.Besides the main commercial cultivars, S-genotypes werealso assigned for several Czech (Melounová et al. 2005),Iranian (Ershadi and Talaii 2007), Swedish (Garkava-Gustavsson et al. 2008), German, English, Norwegianand Danish apple cultivars (Nybom et al. 2008). Our study
connects to these efforts by determining the S-genotype ofold apple cultivars originated in the Carpathian basin. Inaddition, DNA sequence polymorphisms detected inseveral S-alleles may have further relevancies in terms ofcrop evolution.
Materials and methods
Plant material
A total of 23 old apple cultivars were analysed (Table 1).All are maintained at the germplasm collection of CorvinusUniversity of Budapest, Department of Pomology inSoroksár. The most important apple cultivars of theCarpathian basin were sent to the National Fruit Collections(NFC), Brogdale, UK in 1948 (Tóth 2005). Since many ofthem had disappeared from Hungarian germplasm collec-tions, propagation materials were obtained from NFC, andcultivars (1–17) were replanted in the orchard of CUB.Others were collected from Visk, Ukraine (18–22) or theAggtelek National Park, Hungary (23). Total genomic DNAwas extracted from young leaves using Qiagen DNeasyPlant Mini Kit (Hilden, Germany).
PCR and CAPS analysis
Allele-specific primers were used to detect single alleles S1,S2, S3, S5, S7, S9, S10, S23, S24, S26 and S28 or a group ofalleles, S4/16/22 and S6/14/17/20/21 (Broothaerts 2003). Todetect S25-allele, a primer pair published by Kitahara andMatsumoto (2002a) was used. For the simultaneousamplification of more S-alleles, the consensus FTQQYQprimer (Ishimizu et al. 1999) was used in combination withthe anti-I/MIWPNV primer (Matsumoto and Kitahara 2000).In addition, ASPF3 primer (Kim et al. 2006) was also usedwith ASPR3S primer (Kim et al. 2009).
PCR was carried out in a PTC 200 thermocycler (MJResearch, Budapest, Hungary) using the program de-scribed for the primers. Approximately 20–80 ng ofgenomic DNA was used for PCR amplification in a 25-μlreaction volume, containing 1× PCR buffer (Sigma, St.Louis, USA) with final concentrations of 10 mM Tris–HCl (pH 8.3), 50 mM KCl, 1.5 mM MgCl2, 0.2 mM ofdNTPs, 0.4 μM of each primer of the pair and 0.625 U ofTaq DNA polymerase (Sigma). The PCR products wereseparated on 2% TAE agarose gels for 3 h at 100 V, andDNA bands were stained with ethidium bromide. Fragmentsizes were estimated by comparison with the 1-kb+DNAladder (Promega, Madison, USA).
Restriction enzymes were used after allele-specific ampli-fication for the allele S2 (OWB122 and OWB123 primers;EcoRV) and S3 (OWB134 and OWB145; PstI) (Broothaerts
1136 Tree Genetics & Genomes (2011) 7:1135–1145
et al. 1995), S4/16/22 (FTC5 and OWB249, TaqI) (Broothaerts2003), S5 (ASPF3 and ASPR3S; HindIII) and S7 (ASPF3and ASPR3S; HindIII) (Kim et al. 2009).
DNA sequencing and sequence analysis
Genomic DNA fragments were extracted and purified fromthe agarose gels using the QIAquick Gel Extraction Kit(Qiagen) and cloned into a pGEM-T Easy plasmid vector(Promega). PlasmidDNAwas isolatedwith the Rapid PlasmidDNA Daily Miniprep Kit (V-gene, Szeged, Hungary) andsequenced in an automated sequencer ABI PRISM 3100Genetic Analyser (Applied Biosystems, Budapest, Hungary).For each fragment, the nucleotide sequences of three to sixclones obtained from separate PCRs were determined in bothdirections. DNA and deduced amino acid sequences werecompared using BLASTN at NCBI and ClustalW program(Thompson et al. 1994).
Flow cytometry analysis
All cultivars were analysed for ploidy level. Young leaveswere prepared for analysis according to Galbraith (1990).Approximately 1 cm2 of apple leaves were chopped andplaced in a 5-cm Petri dish containing 2 ml extractionbuffer and 2 μl Triton X-100. The extraction bufferconsisted of 20 mmol/l 3-(N-morpholino)propanesulfonicacid (pH 7.0), 34 mmol/l Na3 citrate and 45 mmol MgCl2/l.The tissue was homogenised, and samples were filteredthrough 30-μm nylon meshes into a labelled test tube.Samples were kept on ice throughout. The solution wastransferred to a 1.5-ml conical tube and incubated for20 min at 37°C. Following incubation, propidium iodide(Sigma) was added to each sample to the final concen-tration of 4 μg/ml. The measurement was carried outusing a FACSCalibur flow cytometer (Becton Dickinson,San Jose, USA) on the day of the staining, collecting
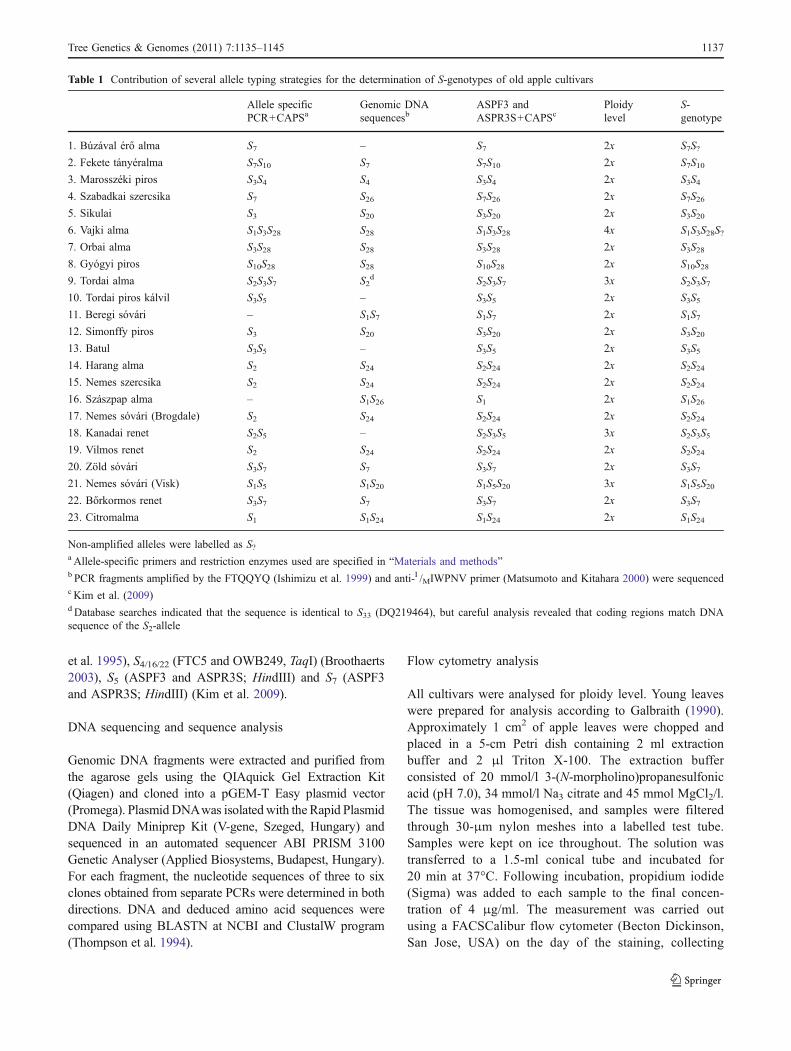
Table 1 Contribution of several allele typing strategies for the determination of S-genotypes of old apple cultivars
Allele specificPCR+CAPSa
Genomic DNAsequencesb
ASPF3 andASPR3S+CAPSc
Ploidylevel
S-genotype
1. Búzával érő alma S7 – S7 2x S7S?2. Fekete tányéralma S7S10 S7 S7S10 2x S7S103. Marosszéki piros S3S4 S4 S3S4 2x S3S44. Szabadkai szercsika S7 S26 S7S26 2x S7S265. Sikulai S3 S20 S3S20 2x S3S206. Vajki alma S1S3S28 S28 S1S3S28 4x S1S3S28S?7. Orbai alma S3S28 S28 S3S28 2x S3S288. Gyógyi piros S10S28 S28 S10S28 2x S10S289. Tordai alma S2S3S7 S2
d S2S3S7 3x S2S3S710. Tordai piros kálvil S3S5 – S3S5 2x S3S511. Beregi sóvári – S1S7 S1S7 2x S1S712. Simonffy piros S3 S20 S3S20 2x S3S2013. Batul S3S5 – S3S5 2x S3S514. Harang alma S2 S24 S2S24 2x S2S2415. Nemes szercsika S2 S24 S2S24 2x S2S2416. Szászpap alma – S1S26 S1 2x S1S2617. Nemes sóvári (Brogdale) S2 S24 S2S24 2x S2S2418. Kanadai renet S2S5 – S2S3S5 3x S2S3S519. Vilmos renet S2 S24 S2S24 2x S2S2420. Zöld sóvári S3S7 S7 S3S7 2x S3S721. Nemes sóvári (Visk) S1S5 S1S20 S1S5S20 3x S1S5S2022. Bőrkormos renet S3S7 S7 S3S7 2x S3S723. Citromalma S1 S1S24 S1S24 2x S1S24
Non-amplified alleles were labelled as S?a Allele-specific primers and restriction enzymes used are specified in “Materials and methods”b PCR fragments amplified by the FTQQYQ (Ishimizu et al. 1999) and anti-I /MIWPNV primer (Matsumoto and Kitahara 2000) were sequencedc Kim et al. (2009)d Database searches indicated that the sequence is identical to S33 (DQ219464), but careful analysis revealed that coding regions match DNAsequence of the S2-allele
Tree Genetics & Genomes (2011) 7:1135–1145 1137

25,000–100,000 cells/tube. For the analysis of results,CellQuest Pro Software 3.1 acquisition and analysingprogram was used.
Results
For the first approach to determine S-genotype of localapple cultivars from the Carpathian basin, allele-specificprimers were applied. Eight of the 14 allele-specific primersused gave amplification from 21 cultivars. Primers selec-tively amplifying the S1-, S2-, S3-, S4/16/22-, S5-, S7-, S10-and S28-alleles worked satisfactorily (Fig. 1), while thosedesigned for the detection of S6/14/17/20/21-, S9-, S23-, S24-,S25- and S26-alleles gave no amplification. For two cultivars(‘Beregi sóvári’ and ‘Szászpap alma’), none of the allele-specific primers amplified any fragment (Table 1).
To detect the rest of alleles, another strategy has beenapplied. The consensus primers, FTQQYQ primer (Ishimizuet al. 1999) combined with the anti-I/MIWPNV primer(Matsumoto and Kitahara 2000) designed from the con-served regions of Maloideae S-RNases were tested. Usingthis primer pair, one or two fragments were detected in theassayed samples with sizes ranging from 320 to 540 bp.
However, fragment size polymorphism was low, andhence, the resolving power of the agarose gel was noteligible to identify S-alleles of very similar fragment sizes.After cloning and sequencing of the PCR products, thefollowing alleles were identified, and their sequence wasdeposited in the GenBank/EMBL/DDBJ database: S1(from four cultivars, HQ689386-9), S2 (HQ693077), S4(HQ689390), S7 (from four cultivars, HQ689391-4), S20(from three cultivars, HQ689395-7), S24 (from fivecultivars, HQ693064-7, HQ6930649), S26 (HQ693070and HQ693072) and S28 (from three cultivars,HQ693074-6) (Table 2 of the Electronic supplementarymaterial). The amplification of the approximately 1,300-bpfragment characteristic for the S3- and S5-RNases failed. Inour experiment, this primer set could detect three additionalalleles (S20, S24 and S26) for which allele-specific primerswere not available or did not give amplification.
This strategy also offered the possibility to comparethe DNA sequence of alleles with the respective allelesavailable from the public database. Single nucleotidepolymorphisms (SNPs) were identified in several allelesincluding S1, S2, S7, S20 and S24 from all or only some ofthe assayed cultivars, while others (S4, S26 and S28) wereidentical with the sequences previously obtained from
Fig. 1 PCR products in 23 oldapple cultivars from theCarpathian basin using a theconsensus ASPF3 (Kim et al.2006) with ASPR3S (Kim et al.2009) and b S1–S5, S7, S10 andS28 allele-specific primers toamplify a range of alleles ofMalus S-RNase gene. M DNAladder, 1–23 numerical order ofsamples corresponds to thenumbers in Table 1
1138 Tree Genetics & Genomes (2011) 7:1135–1145

other cultivars (Table 2 of the Electronic supplementarymaterial). From the four partial S1-RNase sequences, theone isolated from ‘Nemes sóvári (Visk)’ differed in fourSNPs (three of which located within the intron region andone corresponded to a synonymous substitution) from the‘Fuji’ S1 sequence. Very interestingly, this allele was moresimilar to Malus sylvestris S1’ than Malus×domestica S1(Fig. 2a). Similarly to S1, the S7-allele was identical with‘Indo’ and ‘Akane’ S7 in case of three cultivars, while S7from ‘Fekete tányéralma’ differed in two intronic SNPs(Table 2 of the Electronic supplementary material). AllS24-alleles sequenced from Carpathian basin cultivarsdiffered from ‘Akane’ S24 in four to five intronic SNPs(Fig. 2b).
In the intron region of S20-alleles one nucleotide (T)insertion was detected in each assayed cultivar. Two indelswere evident in the intron of ‘Szászpap alma’ S26compared with the S26 identified in ‘Szabadkai szercsika’that completely matched ‘David’ S26. The coding se-quence of the allele S26 was clarified to be identical toMalus sieversii S35, while S28 was rather similar to Pyruscommunis Sd- and Pyrus ussuriensis S35-alleles (Table 2 ofthe Electronic supplementary material). Non-synonymousSNPs were detected in ‘Citromalma’ S1 and ‘Tordai alma’ S2(Fig. 2, Table 2 of the Electronic supplementary material).
In the third set of experiments, a recently developedprimer pair (Kim et al. 2009) was used, which has beenreported to detect simultaneously 22 different S-alleles.This primer set indeed provided a considerably poly-morphic electrophoresis pattern (Fig. 1), amplifying all 11alleles detected. The amplification was successful betweenthe fragment lengths of approximately 600 and 2,250 bp.The biggest fragments representing alleles S3, S5 and S10and the shortest fragment representing allele S7 wereunambiguously distinguished in the agarose gel becausetheir expected size was quite different from all others. Inaddition, allele assignment was confirmed by CAPSanalysis in some cases (S2, S5 and S7). The rest of thefragments were separated into two size groups: group I(618–674 bp) and group II (777–844 bp) according to Kimet al. (2009). To discriminate between these fragments, Kimet al. (2009) developed a CAPS system using 17 restrictionenzymes. In our experiment, it proved unnecessary to useall restriction enzymes because a longer running periodachieved much better resolution and allele sequencesdetermined from several cultivars allowed reliable identifi-cation of the corresponding fragment sizes described byKim et al. (2009).
Using several rounds of PCR and sequencing, a total of11 different S-alleles were identified in the tested 23cultivars. For one cultivar (‘Búzával érő alma’), only oneS-allele was identified, while two and three alleles wereassigned for 18 and four cultivars, respectively. The
presence of more than two alleles may be the consequenceof polyploidy. Since former data were not available on theploidy level of these old cultivars, flow cytometry analysiswas inevitable to decide whether the presence of two orthree alleles represents full or partial S-genotypes. Alto-gether, 19 cultivars proved to be diploid, three cultivars,‘Tordai alma’, ‘Kanadai renet’ and ‘Nemes sóvári (Visk)’,were triploid, while ‘Vajki alma’ was the only tetraploidcultivar out of the 23 genotypes assayed (Table 1, Fig. 3).
The genotypes and ploidy level determined for testedcultivars are shown in Table 1. By combining the resultsprovided by three different S-genotyping techniques andflow cytometry analysis, the complete S-allele constitutionsof 21 cultivars were determined. In general, two S-alleleswere found in diploids with one exception showing onlyone allele, and three alleles were detected in triploids. Theonly tetraploid cv. ‘Vajki alma’ also showed three alleles.Some cultivars possessed the same S-genotype: ‘Batul’ and‘Tordai piros kálvil’ (S3S5), ‘Bőrkormos renet’ and ‘Zöldsóvári’ (S3S7), ‘Sikulai’ and ‘Simonffy piros’ (S3S20), or‘Harang alma’, ‘Nemes szercsika’, ‘Nemes sóvári (Brogdale)’and ‘Vilmos renet’ (S2S24).
The most commonly occurring S-alleles of thoseidentified in the present study were S3 (22%), followed byS7 (14%), S2 and S24 (12%), S1 (10%), S5 (8%), S20 and S28(6%) and S10 and S26 (4%), while S4-allele was only foundin one cultivar (2%).
Discussion
Robustness and reliability of the S-genotyping approaches
Knowledge of self-incompatibility genotype of apple iscrucial for breeding and orchard planning since self-compatible cultivars are unknown and parthenocarpy cannotgive economically reasonable fruit set. Therefore, different S-genotyping strategies have been elaborated. In contrast toPrunus where two introns are inserted in the S-RNase gene(Halász et al. 2007), the maloideous S-RNases contain onlyone intron, which is characterised by relatively moderatelength variations among alleles (Broothaerts et al. 1995).However, the introns being free from the effects of purifyingselection and the hypervariable exonic region exposed topositive selection are variable in sequence so that primerscould be designed to selectively amplify specific alleles.We tested the performance of 14 allele-specific primers(Broothaerts 2003; Kitahara and Matsumoto 2002a), eightof which gave amplification. Two of the primers were latershown to give false negative results for the S24- and S26-alleles. False negative problems were also evident for S1 in‘Beregi sóvári’ and ‘Szászpap alma’, S3 in ‘Kanadai renet’and S7 in ‘Beregi sóvári’. This might be attributed to low
Tree Genetics & Genomes (2011) 7:1135–1145 1139

a
b
1140 Tree Genetics & Genomes (2011) 7:1135–1145

quality and/or quantity DNA samples (Kim et al. 2009).False-positive results did not occur possibly due to the use ofimproved and more specific primers (Broothaerts 2003).Altogether, 70% of the S-alleles present in the samplepopulation were detected using allele-specific primers.
The false-negative results as well as time-consumingnature of the analysis, including many subsequent runs ofPCRs, have prompted experts to find an alternativetechnique. The consensus primers FTQQYQ andanti-I/MIWPNV (Ishimizu et al. 1999; Matsumoto andKitahara 2000) amplified more alleles simultaneously,although fragments showed very limited size variations,which made allele-specific digestion or sequencing neces-
sary for genotyping. In total, only 46% of all alleles couldbe detected based on this strategy, which might be due tothe matching fragment sizes of different alleles andpreferential amplification (Broothaerts et al. 1995). Whencomplex bands were isolated from the agarose gels andcloned into vector, some alleles might be overrepresentedamong clones. However, it is worth mentioning that onlythis approach would allow the identification of novel allelesfor which detection assays are not yet available. In addition,SNPs identified in sequences may help to clarify evolutionaryconnections between genotypes (see later).
In the third set of experiments, ASPF3 primer (Kim et al.2006) was used with ASPR3S (Kim et al. 2009), a primercombination that was reported to amplify 22 S-RNase alleles.This primer set also gave reliable amplification for all 11alleles present in the sample population in the present study,with the exception of S26 in ‘Szászpap alma’. Altogether,96% of all alleles carried by the assayed cultivars weredetected, and hence, the primers proved to be especiallyrobust in the S-genotyping of old apple cultivars.
In conclusion, consensus primers proved preferable toallele-specific primers, since false-negative allele-specificassays may prevent correct genotyping. DNA sequencingor using restriction enzymes are relatively expensive, while
Fig. 3 Ploidy levels of old apple cultivars from the Carpathian basin.a ‘Idared’ and ‘Jonica’ were used as diploid (red) and triploid (green)control cultivars; blue curve indicates the relative fluorescence values
of the samples b ‘Marosszéki piros’ (2n=2x), c ‘Tordai alma’ (2n=3x)and d ‘Vajki alma’ (2n=4x)
Fig. 2 Alignment of partial (C2-RHV-intron-RHV region) nucleotidesequence of Malus S-RNases. Comparison of partial S1-RNasesequences from the commercial cultivar ‘Fuji’, Carpathian basincultivars, ‘Citromalma’, ‘Szászpap alma’, ‘Beregi sóvári’ and ‘Nemessóvári (Visk)’ and M. sylvestris (a) as well as partial S24 sequencesfrom the commercial cultivar ‘Akane’ and Carpathian basin cultivars,‘Vilmos renet’, ‘Harang alma’, ‘Nemes sóvári (Brogdale)’ and ‘Nemesszercsika’ (b). Black boxes indicate different residues between thealigned sequences. The hyper variable region (RHV) is boxed asindicated by Sassa et al. (1996); broken arrows points into the deducedintron regions
R
Tree Genetics & Genomes (2011) 7:1135–1145 1141

consensus primer approach may also be burdened withfalse-negative effects, and hence the S-genotyping of anunknown material will require a combination of some ofthese techniques. At first, we propose using the ASPF3 andASPR3S primer pair, and in case of undetected or doubtfulalleles, it should be completed with other techniques. Ifploidy level is unknown, flow cytometry should also beused to verify the completeness of genotypes.
For the tetraploid cultivar ‘Vajki alma’ (S1S3S28S?),only three S-alleles were assigned. It is supposed that thedetection of the fourth allele failed. Another explanationmight be that ‘Vajki alma’ indeed carries only threedifferent alleles, and one of them is found in two copiesin the tetraploid genome. The rising of tetraploid plantsrequires the fusion of diploid male and female gametes. Aheteroallelic diploid pollen grain carrying two different S-alleles will be able to achieve fertilisation on any style dueto competitive interaction (de Nettancourt 2001). Lewisand Modlibowska (1942) presented evidence that, in pear,a heteroallelic pollen (e.g. S1S2) of a tetraploid plant wouldsucceed on a style even if they have both alleles incommon (resulting in the genotype of S1S1S2S2 in ourexample), whereas a homoallelic pollen (e.g. S1S1 or S2S2)will fail. A possible scenario is that the seed parent wasfertilised by a heteroallelic diploid pollen (carrying one ofthe two alleles present in the diploid gamete of the seedparent along with another allele different from those). Inthis case, the offspring will have three different S-alleles,and its tetraploid nature can only be detected using flowcytometry.
Breeding and crop evolutionary perspectives
All four cross-incompatibility groups established in thepresent study were previously identified among the 60groups described by Broothaerts et al. (2004), and hence,our work extended these groups by incorporating oldCarpathian basin cultivars. This information might be usedin apple breeding programs.
The S-genotype of the cultivar ‘Kanadai renet’ wasdetermined as S2S3S5. Previously, a cultivar with the samename (‘Kanada Reinette’) obtained from the NFC atBrogdale, UK was genotyped as S1S2S3 (Bošković andTobutt 1999; Broothaerts 2003), indicating that our materialcollected and known in Ukraine under this name is notidentical with that sent to Brogdale from Hungary in 1948(Tóth 2005), although their triploid nature and the twomatching S-alleles indicate their relatedness, presumably aparent–offspring relationship. More interesting is thedifference between the ‘Nemes sóvári’ trees obtained fromUkraine and Brogdale. It indicates that cultivars kept underthe same name in different germplasm collections mightrepresent different genotypes. S-allele assignment and other
molecular markers are appropriate tools to check theidentity of such cultivars.
One of the main purposes in apple breeding programs isto obtain cultivars with increased tolerance against wide-spread diseases like scab, powdery mildew and fire blight.Several locally adapted genotypes (‘Sikulai’, ‘Tordai piroskálvil’, ‘Batul’, ‘Vilmos renet’, ‘Zöld sóvári’, etc.) havebeen found at the Carpathian basin with valuable pomo-logical characteristics and moderate resistance to Erwiniaamylovora and scab (Tóth 2005; Tóth et al. 2005), whichmakes them attractive for using as donor genotypes inbreeding programs for resistance, fruit size, favourabletaste, etc.
Scab-resistant apple cultivars were shown to carry theS2- or S3-allele (Broothaerts and Van Nerum 2003), whichwas attributed to the use of common breeding parents. It isinteresting to consider that all cultivars characterised by atleast low susceptibility to Venturia inaequalis (‘Sikulai’,‘Tordai piros kálvil’, ‘Batul’ and ‘Vilmos renet’) are alsoharbouring the S2- or S3-alleles. Some of these cultivarshave been known in the Carpathian basin for more than200 years and are even dated back to the Turkish occupationof Transylvania (Bereczki 1899), which may rule outprevious explanations like sharing common ancestry. Itshould be checked if the presence of these alleles issomehow coupled with the increased scab tolerance. S5 wasalso frequently observed in some resistant cultivars (Melou-nová et al. 2005) and was detected in two of the four scabtolerant cultivars analysed in the present study. For futurebreeding to develop scab-resistant cultivars that do notcontain this allele, ‘Sikulai’ or ‘Vilmos renet’ would be agood choice, allowing also to check for co-segregation ofthese alleles and scab resistance in their progenies.
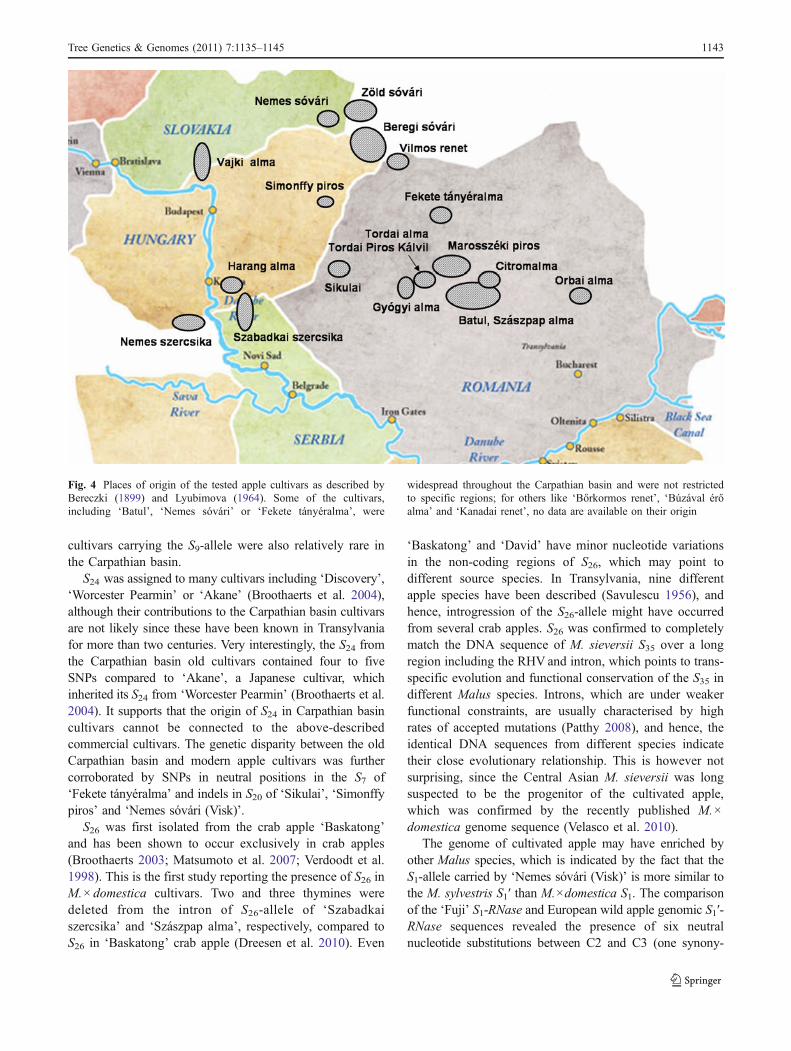
The places of origin of the S-genotyped 23 cultivarsrepresent different regions in the Carpathian basin (Fig. 4)and so might have interesting implications for their geneticpotential and relatedness to apples from other territories.Although the number of assayed cultivars would beexpected to influence the allele frequency data, thecomparison of studies carried out on an uneven number ofcultivars revealed very similar allelic frequencies (Dreesenet al. 2010; Nybom et al. 2008). Our data show that theallele frequencies of S1–S5, S7, S10, S20 and S28 were verysimilar to most studies. However, the frequency of S24 was12%, in contrast with the ranges 3–4% determined inprevious studies. The frequency of S26 (4%) also exceededthe values published in other studies (Broothaerts et al.2004). The absence of the allele S9 is also striking, as it had14–16% frequency in many other studies. However, innorthern European cultivars, it was also a very rare allele(1%) possibly due to the exclusion of some not sufficientlywinter hardy cultivars (many of which carried S9) frombreeding (Nybom et al. 2008). Our data indicate that
1142 Tree Genetics & Genomes (2011) 7:1135–1145
cultivars carrying the S9-allele were also relatively rare inthe Carpathian basin.
S24 was assigned to many cultivars including ‘Discovery’,‘Worcester Pearmin’ or ‘Akane’ (Broothaerts et al. 2004),although their contributions to the Carpathian basin cultivarsare not likely since these have been known in Transylvaniafor more than two centuries. Very interestingly, the S24 fromthe Carpathian basin old cultivars contained four to fiveSNPs compared to ‘Akane’, a Japanese cultivar, whichinherited its S24 from ‘Worcester Pearmin’ (Broothaerts et al.2004). It supports that the origin of S24 in Carpathian basincultivars cannot be connected to the above-describedcommercial cultivars. The genetic disparity between the oldCarpathian basin and modern apple cultivars was furthercorroborated by SNPs in neutral positions in the S7 of‘Fekete tányéralma’ and indels in S20 of ‘Sikulai’, ‘Simonffypiros’ and ‘Nemes sóvári (Visk)’.
S26 was first isolated from the crab apple ‘Baskatong’and has been shown to occur exclusively in crab apples(Broothaerts 2003; Matsumoto et al. 2007; Verdoodt et al.1998). This is the first study reporting the presence of S26 inM.×domestica cultivars. Two and three thymines weredeleted from the intron of S26-allele of ‘Szabadkaiszercsika’ and ‘Szászpap alma’, respectively, compared toS26 in ‘Baskatong’ crab apple (Dreesen et al. 2010). Even
‘Baskatong’ and ‘David’ have minor nucleotide variationsin the non-coding regions of S26, which may point todifferent source species. In Transylvania, nine differentapple species have been described (Savulescu 1956), andhence, introgression of the S26-allele might have occurredfrom several crab apples. S26 was confirmed to completelymatch the DNA sequence of M. sieversii S35 over a longregion including the RHV and intron, which points to trans-specific evolution and functional conservation of the S35 indifferent Malus species. Introns, which are under weakerfunctional constraints, are usually characterised by highrates of accepted mutations (Patthy 2008), and hence, theidentical DNA sequences from different species indicatetheir close evolutionary relationship. This is however notsurprising, since the Central Asian M. sieversii was longsuspected to be the progenitor of the cultivated apple,which was confirmed by the recently published M.×domestica genome sequence (Velasco et al. 2010).
The genome of cultivated apple may have enriched byother Malus species, which is indicated by the fact that theS1-allele carried by ‘Nemes sóvári (Visk)’ is more similar tothe M. sylvestris S1′ than M.×domestica S1. The comparisonof the ‘Fuji’ S1-RNase and European wild apple genomic S1′-RNase sequences revealed the presence of six neutralnucleotide substitutions between C2 and C3 (one synony-
Fig. 4 Places of origin of the tested apple cultivars as described byBereczki (1899) and Lyubimova (1964). Some of the cultivars,including ‘Batul’, ‘Nemes sóvári’ or ‘Fekete tányéralma’, were
widespread throughout the Carpathian basin and were not restrictedto specific regions; for others like ‘Bőrkormos renet’, ‘Búzával érőalma’ and ‘Kanadai renet’, no data are available on their origin
Tree Genetics & Genomes (2011) 7:1135–1145 1143

mous SNP in exon and five SNPs in intron). Since S1 from‘Nemes sóvári (Visk)’ shared four out of the six nucleotideswith M. sylvestris, it points to a putative introgression event.Such introgression was also suggested for some oldCarpathian basin apple cultivars, including ‘Nemes sóvári’based on morphological characters (Borza 1921).
The S28-allele isolated from three old cultivars wasidentical to that reported from the ‘Delicious’ cultivar. Thisallele is rather similar to P. communis Sd, which is identicalto P. ussuriensis S35. It indicates that this allele might havebeen inherited from the common ancestor of these taxa, as97–98% identity of trans-specific and trans-generic S-alleles within theMaloideae family was shown (Bokszczaninet al. 2009). Non-synonymous SNPs were detected in theRHV region of ‘Citromalma’ S1 and ‘Tordai alma’ S2; theirfunction requires verification.
New alleles were not detected in the Carpathian basincultivars, which might be due to the many apple allelesalready described and also to the restricted genetic variabilityof cultivated apples. Domestication of apple has been shownto increase genetic uniformity (Dreesen et al. 2010), whichwas further impoverished through modern breeding usingonly a limited set of genotypes for hybridisation (Broothaertset al. 2004; Nybom et al. 2008). However, alterations inallele frequencies, the presence of S26 and SNPs withinspecific S-alleles of the Carpathian basin cultivars indicatetheir genetic disparity. The occurrence of such alterationsmay also be associated with historical events, e.g. Turkishoccupation over the sixteenth and seventeenth centuries, aswas also confirmed in case of apricots (Halász et al. 2010).There are historical records supporting the Turkish origin ofsome of the tested apple cultivars like ‘Sikulai’ and others(Bereczki 1899). Introducing such old cultivars into breedingprograms may contribute to the widening of the genetic basisof cultivated apple and help to prevent genetic erosion infuture commercial cultivars.
Our data suggest that the genome of old cultivars fromthe Carpathian basin was enriched by several Malus taxaand are free from the consequences of modern breeding.The information presented in this paper can be equallyuseful for breeders and pomologists and in following thekey stages of crop evolution. The historical link betweenHungary and the supposed Central Asian territory of appleorigin is also commemorated by the Hungarian name of thisfruit ‘alma’, which derives from the Kazakh word for apple.Apples have been present in the Carpathian basin for a longtime, and hence, this region forms an important connectionzone between the main growing countries of Europe andthe Central Asian gene centre.
Acknowledgements This work was funded by the NKTH-OTKAK68921 project and also supported by the János Bolyai ResearchScholarship of the Hungarian Academy of Sciences (Júlia Halász andAttila Hegedűs). The authors thank Ildikó Király for collecting plant
material and performing DNA extraction and István Forgács for hiskind help in sample preparation for flow cytometry.
References
Bereczki M (1899) Gyümölcsészeti vázlatok, vol I–V, 2nd edn. PestiLoyd Társulat, Budapest
Bokszczanin K, Palucha A, Przybyla A (2009) Description of anew trans-generic Skb-RNase allele in apple. Euphytica 166(1):83–94
Borza A (1921) Flora gradinilor taranesti romane. I. Marul, vol 1. BulInform Grad Muz Bot Univ Cluj. Grad. Muz. Bot. Univ. Cluj,Cluj-Napoca
Bošković R, Tobutt KR (1999) Correlation of stylar ribonucleaseisoenzymes with incompatibility alleles in apple. Euphytica 107(1):29–43
Broothaerts W (2003) New findings in apple S-genotype analysisresolve previous confusion and request the re-numbering of someS-alleles. Theor Appl Genet 106(4):703–714
Broothaerts W, Van Nerum I (2003) Apple self-incompatibilitygenotypes: an overview. Acta Hort 622:379–387
Broothaerts W, Janssens GA, Proost P, Broekaert WF (1995) cDNAcloning and molecular analysis of two self-incompatibility allelesfrom apple. Plant Mol Biol 27(3):499–511
Broothaerts W, Van Nerum I, Keulemans J (2004) Update on andreview of incompatibility (S-) genotypes of apple cultivars.Hortscience 39:943–947
de Nettancourt D (2001) Incompatibility and incongruity in wild andcultivated plants, 2nd edn. Springer, New York
Dreesen R, Vanholme B, Luyten K, Van Wynsberghe L, Fazio G,Roldán-Ruiz I, Keulemans J (2010) Analysis of Malus S-RNasegene diversity based on a comparative study of old and modernapple cultivars and European wild apple. Mol Breed 26(4):693–709
Ershadi A, Talaii A (2007) Identification of S-alleles in 40 apple(Malus x domestica BORKH) cultivars by allele-specific PCRamplification. Acta Hort 760:111–116
Galbraith DW (1990) Flow cytometric analysis of plant genomes.Meth Cell Biol 33:549–562
Garkava-Gustavsson L, Brantestam AK, Sehic J, Nybom H (2008)Molecular characterisation of indigenous Swedish apple cultivarsbased on SSR and S-allele analysis. Hereditas 145(3):99–112
Halász J, Pedryc A, Hegedus A (2007) Origin and dissemination of thepollen-part mutated SC-haplotype which confers self-compatibilityin apricot (Prunus armeniaca). New Phytol 176(4):792–803
Halász J, Pedryc A, Ercisli S, Yilmaz KU, Hegedus A (2010) S-genotyping supports the genetic relationships between Turkish andHungarian apricot germplasm. J Am Soc Hort Sci 135(5):410–417
Ishimizu T, Inoue K, Shimonaka M, Saito T, Terai O, Norioka S (1999)PCR-based method for identifying the S-genotypes of Japanesepear cultivars. Theor Appl Genet 98(6):961–967
Janssens GA, Goderis IJ, Broekaert WF, Broothaerts W (1995) Amolecular method for S-allele identification in apple based onallele-specific PCR. Theor Appl Genet 91(4):691–698
Kim H, Hattori G, Hirata Y, Kim D, Hwang J, Shin Y, Nou I (2006)Determination of self-locompatibility genotypes of Korean applecultivars based on S-RNase PCR. J Plant Biol 49(6):448–454
Kim H, Park J, Hirata Y, Nou I (2008) Molecular characterization ofnew S-RNases (‘S31’ and ‘S32’) in apple (Malus × domesticaBorkh.). J Plant Biol 51(3):202–208
Kim H, Kakui H, Kotoda N, Hirata Y, Koba T, Sassa H (2009)Determination of partial genomic sequences and development ofa CAPS system of the S-RNase gene for the identification of 22 Shaplotypes of apple (Malus × domestica Borkh.). Mol Breed 23(3):463–472
1144 Tree Genetics & Genomes (2011) 7:1135–1145

Kitahara K, Matsumoto S (2002a) Cloning of the S25 cDNA from‘McIntosh’ apple and an S25-allele identification method. J HorticSci Biotechnol 77:724–728
Kitahara K, Matsumoto S (2002b) Sequence of the S10 cDNA from'McIntosh' apple and a PCR-digestion identification method.Hortscience 37(1):187–190
Kitahara K, Soejima J, Komatsu H, Fukui H, Matsumoto S (2000)Complete sequences of the S-genes, Sd- and Sh-RNase cDNA inapple. Hortscience 35(4):712–715
Kobel F, Steinegger P, Anliker J (1939) Weitere Untersuchungen überdie Befruchtungsverhältnisse der Apfel- und Birnsorten. LandwJb Schweiz 53:160–191
Komori S, Soejima J, Abe K, Kotoda N, Kato H (2000) Analysis of S-allele genotypes and genetic diversity in the apple. Acta Hort538:83–86
Lewis D, Modlibowska I (1942) Genetical studies in pears. J Genet 43(1):211–222
Long S, Li M, Han Z, Wang K, Li T (2010) Characterization of threenew S-alleles and development of an S-allele-specific PCRsystem for rapidly identifying the S-genotype in apple cultivars.Tree Genet Genomes 6(2):161–168
Lyubimova L (1964) Jabloni zá Kárpáttiá. UzhgorodMatityahu A, Stern RA, Schneider D, Goldway M (2005) Molecular
identification of a new apple S-RNase—S29—cloned from‘Anna’, a low-chilling-requirement cultivar. Hortscience 40(3):850–851
Matsumoto S, Kitahara K (2000) Discovery of a new self-incompatibility allele in apple. Hortscience 35(7):1329–1332
Matsumoto S, Eguchi T, Bessho H, Abe K (2007) Determination andconfirmation of S-RNase genotypes of apple pollinators andcultivars. J Hort Sci Biotech 82(2):323–329
McClure BA, Haring V, Ebert PR, Anderson MA, Simpson RJ,Sakiyama F, Clarke AE (1989) Style self-incompatibility geneproducts of Nicotlana alata are ribonucleases. Nature 342(6252):955–957
MelounováM, Vejl P, Sedlák P, Blazek J, Zoufalá J, Milec Z, Blazková H(2005) Alleles controlling apple skin colour and incompatibility in
new Czech apple varieties with different degrees of resistanceagainst Venturia inaequalis CKE. Plant Soil Environ 51:65–73
Nybom H, Sehic J, Garkava-Gustavsson L (2008) Self-incompatibilityalleles of 104 apple cultivars grown in northern Europe. J HortSci Biotech 83:339–344
Patthy L (2008) Protein evolution, 2nd edn. Blackwell, MaldenSassa H, Mase N, Hirano H, Ikehashi H (1994) Identification of self-
incompatibility-related glycoproteins in styles of apple (Malus ×domestica). Theor Appl Genet 89(2):201–205
Sassa H, Nishio T, Kowyama Y, Hirano H, Koba T, Ikehashi H (1996)Self-incompatibility (S) alleles of the Rosaceae encode membersof a distinct class of the T2/S ribonuclease superfamily. Mol GenGenet 250(5):547–557
Savulescu T (1956) Flora republicii populare romane, 4th edn. AcadR.P.R, Bucharest
Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W:improving the sensitivity of progressive multiple sequencealignment through sequence weighting, position-specific gappenalties and weight matrix choice. Nucl Acids Res 22:4673–4680. doi:10.1093/nar/22.22.4673
Tóth M (2005) Exploring and preserving old apple cultivars of theCarpathian basin. Int J Hort Sci 11:9–13
Tóth M, Baliko E, Szani Z (2005) Evaluation of fruit quality of oldapple cultivars originating from the foot of the CarpathianMountains, for utilization in breeding and in organic farming.Int J Hort Sci 11(3):15–21
Van Nerum I, Geerts M, Van Haute A, Keulemans J, Broothaerts W(2001) Re-examination of the self-incompatibility genotype ofapple cultivars containing putative ’new’ S-alleles. Theor ApplGenet 103(4):584–591
Velasco R, Zharkikh A, Affourtit J et al (2010) The genome of thedomesticated apple (Malus x domestica Borkh.). Nat Genet 42(10):833–839
Verdoodt L, Van Haute A, Goderis IJ, De Witte K, Keulemans J,Broothaerts W (1998) Use of the multi-allelic self-incompatibilitygene in apple to assess homozygocity in shoots obtained throughhaploid induction. Theor Appl Genet 96(2):294–300
Tree Genetics & Genomes (2011) 7:1135–1145 1145
Top Related
Copyright © 2022 FDOKUMEN

