







Bahasa
Halaman
Hukum


REVIEW
Promoter diversity in multigene transformation
Ariadna Peremarti • Richard M. Twyman • Sonia Gomez-Galera •
Shaista Naqvi • Gemma Farre • Maite Sabalza • Bruna Miralpeix •
Svetlana Dashevskaya • Dawei Yuan • Koreen Ramessar • Paul Christou •
Changfu Zhu • Ludovic Bassie • Teresa Capell
Received: 2 November 2009 / Accepted: 11 March 2010 / Published online: 31 March 2010
� Springer Science+Business Media B.V. 2010
Abstract Multigene transformation (MGT) is becoming
routine in plant biotechnology as researchers seek to gen-
erate more complex and ambitious phenotypes in trans-
genic plants. Every nuclear transgene requires its own
promoter, so when coordinated expression is required, the
introduction of multiple genes leads inevitably to two
opposing strategies: different promoters may be used for
each transgene, or the same promoter may be used over and
over again. In the former case, there may be a shortage of
different promoters with matching activities, but repetitious
promoter use may in some cases have a negative impact on
transgene stability and expression. Using illustrative case
studies, we discuss promoter deployment strategies in
transgenic plants that increase the likelihood of successful
and stable multiple transgene expression.
Keywords Promoter � Transgene �Multigene transformation � Transcriptional silencing �Constitutive � Spatiotemporal � Inducible
Introduction
Genetic engineering has increased our fundamental under-
standing of how plants function. By introducing new genes,
we can study how plants grow, develop, and defend them-
selves against pests, diseases and harsh environments, how
photosynthesis is controlled, and the basis of primary and
secondary metabolism (Slater et al. 2003). Genetic engineer-
ing can also be applied to generate hardier crops, more nutri-
tious food, and plants that produce chemical precursors, novel
oils, industrial enzymes and pharmaceuticals (Farre et al.
2010; Zhu et al. 2007; Ma et al. 2003). The transferred genes
play the most significant role in determining the phenotypes of
transgenic plants but like stage hands working hard behind the
scenes, the promoters that control transgene expression are
essential components that often get overlooked. Promoters
allow transgene expression to be regulated, restricted and fine-
tuned, delivering more precise control over the manner in
which phenotypes are expressed (Twyman 2003).
The promoters used in plant biotechnology are tradition-
ally divided into three categories—constitutive (active con-
tinuously in most or all tissues), spatiotemporal (tissue-
specific or stage-specific activity) and inducible (regulated
by the application of an external chemical or physical signal;
Potenza et al. 2004). However, all are based on similar core
sequences usually including an initiator and TATA-box as
well specific cis-acting motifs that bind to transcription fac-
tors (Box 1). Promoter sequences are the same in all plant
tissues whether the gene is expressed or not, so the activity of
a promoter depends on the availability and activity of the
transcription factors. Those binding to constitutive promoters
are available and active all the time, whereas those binding
to spatiotemporal and inducible promoters are themselves
rationed and made available only in certain tissues or
developmental stages, or in response to external signals.
A. Peremarti � S. Gomez-Galera � S. Naqvi � G. Farre �M. Sabalza � B. Miralpeix � S. Dashevskaya � D. Yuan �K. Ramessar � P. Christou � C. Zhu � L. Bassie � T. Capell
Departament de Produccio Vegetal i Ciencia Forestal, ETSEA,
Universitat de Lleida, Av. Alcalde Rovira Roure 191, 25198
Lleida, Spain
R. M. Twyman
Department of Biological Sciences, University of Warwick,
Coventry CV4 7AL, UK
P. Christou (&)
Institucio Catalana de Recerca i Estudis Avancats, Bellaterra,
Spain
e-mail: [email protected]
123
Plant Mol Biol (2010) 73:363–378
DOI 10.1007/s11103-010-9628-1

In the early years of plant biotechnology, most trans-
genic plants contained two transgenes—one selectable
marker under the control of a constitutive promoter to
facilitate the selective propagation of transformed cells,
and a ‘primary transgene’ which could be under the control
of any sort of promoter and was intended to alter the plant’s
phenotype in a specific manner. MGT is now being
embraced as an approach to generate plants with more
ambitious phenotypes (Naqvi et al. 2009a). MGT allows
goals that were once impossible to be achieved—the
import of complex metabolic pathways, the expression of
entire protein complexes, the development of transgenic
crops simultaneously engineered to produce a spectrum of
added-value compounds (Zhu et al. 2007; Naqvi et al.
2009a). But each additional transgene requires its own
promoter, making it necessary to find different promoters
that achieve the same expression profile or use one pro-
moter multiple times in the same transgenic plant. In the
first case there may be a shortage of available promoters
with the desired expression profile, whereas in the second
case the deliberate introduction of repetitive sequences into
a transgenic locus has in some cases been linked with
undesirable negative effects on transgene expression and
stability (Mette et al. 1999, 2000; Mourrain et al. 2007).
Promoter diversity
The challenge of multiple coordinated transgene expres-
sion can be addressed using a promoter diversity approach,
where different promoters are used to drive different
transgenes with the same expression profile. This is
achievable with most expression strategies when the
number of transgene is small, but becomes increasingly
difficult as the number of transgenes increases due to the
lack of available promoters with suitable expression pro-
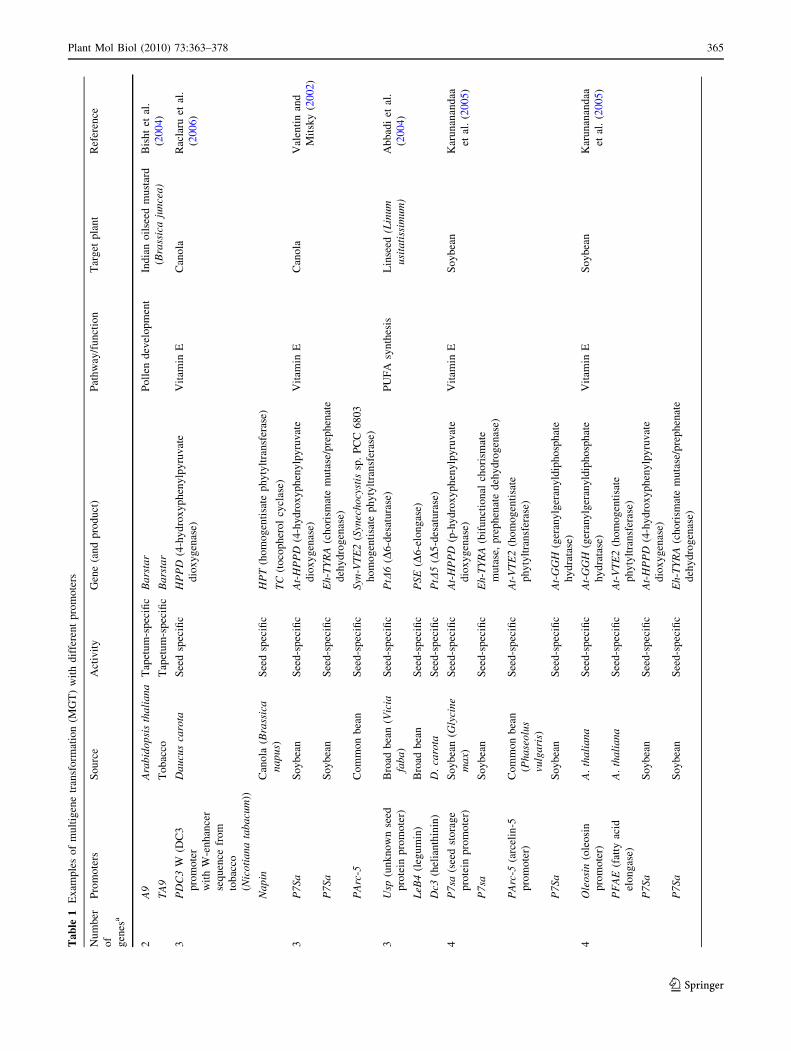
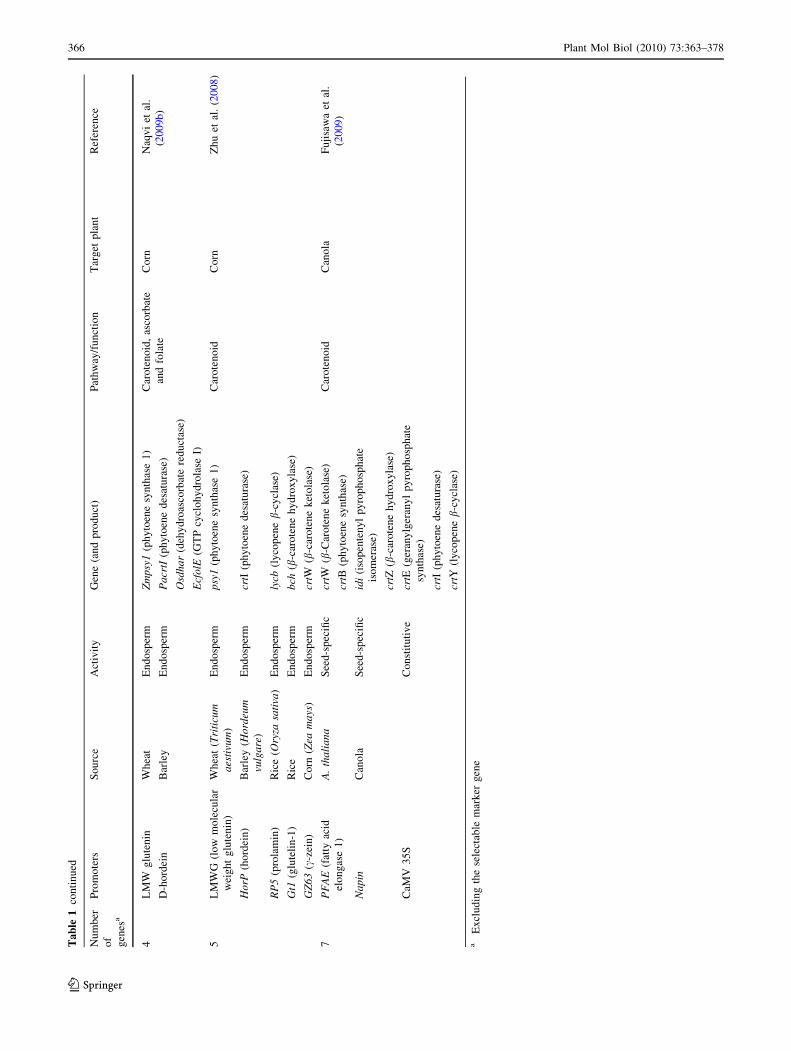
files. Table 1 provides examples where MGT has been
carried out with transgenes driven in a coordinated manner
by different promoters.
Constitutive promoters
Constitutive promoters show the most diversity because
there are two major sources—plant viruses and plant
housekeeping genes. Plant viruses have small genes that
are easy to define genetically, and small genomes that are
easy to manipulate in vitro, so many of the earliest con-
stitutive promoters were derived from plant viruses and are
still widely used today. The most prevalent of these is the
Cauliflower mosaic virus 35S promoter, which controls the
synthesis of the 35S major transcript (Odell et al. 1985;
Kay et al. 1987). The full-sized regulatory region for the
CaMV 35S major transcript is just under 3 kbp in length
(Odell et al. 1985). However, shorter cloned fragments are
active, and the typical CaMV 35S promoter used in plant
expression vectors is a 352-bp fragment spanning nucleo-
tides -343 to ?9 (Fang et al. 1989). The core promoter
containing the TATA box extends to position -46 and
confers basal activity. Full activity requires upstream pro-
moter elements, some of which are tissue-specific, which
Box 1 Plant promoters, the basics
The core promoters of protein-encoding genes in plants are similar to those found in animals. They often contain a TATA box positioned at
approximately position -25 relative to the transcriptional initiation site, with the consensus sequence TATAWAW. They may also possess
an initiator element, which is found immediately adjacent to the transcriptional start site and has the consensus sequence YYCARR. These
two elements help to position the general transcription factor TFIID, which contains the TATA-binding protein (TBP) and various TBP-
associated factors (TAFs). However, some promoters lack both a TATA box and an initiator, and the genes under their control tend to have
multiple transcriptional initiation sites reflecting an inability to position TFIID precisely. Other components of the transcriptional initiation
complex then assemble: TFIIA, TFIIB, RNA polymerase II (recruited to the complex by TFIIF), TFIIE and TFIIH. These proteins have been
isolated and functionally characterized in a number of plants, and appear closely related to their animal counterparts (Twyman 2003).
Adjacent to the core promoter, further motifs are clustered in what is known as the upstream promoter region, which extends 100–200 bp on
the 50 side of the transcriptional start site. These motifs bind transcription factors that interact with the transcriptional initiation complex and
facilitate its assembly, improve its stability or increase the efficiency of promoter escape (movement away from the promoter, into the gene
itself) once the transcriptional machinery sets off. Some of these transcriptional activators are constitutive while others may be cell-specific,
stage-specific or inducible by external signals. Therefore, although the DNA sequence of the promoter is the same in each plant cell, the
availability of the transcription factors that bind to individual motifs varies considerably, and this provides the main mechanism for
transcriptional regulation. As well as the upstream promoter, there are more distant cis-acting elements known as enhancers that fulfill a
similar function. In virus genomes, enhancers are often found quite near the gene and promoter, allowing the isolation of all regulatory
elements on a relatively short DNA fragment. In plant genomes, enhancers may be located many kbp away, and may be found upstream,
downstream or even within the introns of the genes they control. Because of their distant location relative to the core promoter, interactions
with the transcriptional initiation complex are achieved by looping out the intervening DNA. Other proteins that bind to the upstream
promoter disrupt the transcriptional initiation complex, either directly or by blocking the activity of transcriptional activators. These
transcriptional repressors also bind at more distant sites known as silencers. Transcriptional activators and repressors function in various
ways to influence the stability of the initiation complex. Some are DNA binding proteins while others interact at the protein level. Some may
form direct contacts with the initiation complex, or indirect contacts through a bridging complex called mediator, while others modify
chromatin structure or introduce bends or kinks into the promoter DNA to facilitate or disrupt other interactions. Some regulatory proteins
can act as either activators or repressors depending on promoter context and the other proteins that are present.
364 Plant Mol Biol (2010) 73:363–378
123
Ta
ble
1E
xam
ple
so
fm
ult
igen
etr
ansf
orm
atio
n(M
GT
)w
ith
dif
fere
nt
pro
mo
ters
Nu
mb
er
of
gen
esa
Pro
mo
ters
So
urc
eA
ctiv
ity
Gen
e(a
nd
pro
du
ct)
Pat
hw
ay/f
un
ctio
nT
arg
etp
lan
tR
efer
ence
2A
9A
rab
ido
psi
sth
ali
an
aT
apet
um
-sp
ecifi
cB
ars
tar
Po
llen
dev
elo
pm
ent
Ind
ian
oil
seed
mu
star
d
(Bra
ssic
aju
nce
a)
Bis
ht
etal
.
(20
04)
TA
9T
ob
acco
Tap
etu
m-s
pec
ific
Ba
rsta
r
3P
DC
3W
(DC
3
pro
mo
ter
wit
hW
-en
han
cer
seq
uen
cefr
om
tob
acco
(Nic
oti
an
ata
ba
cum
))
Da
ucu
sca
rota
See
dsp
ecifi
cH
PP
D(4
-hy
dro
xy
ph
eny
lpy
ruv
ate
dio
xy
gen
ase)
Vit
amin
EC
ano
laR
acla
ruet
al.
(20
06)
Na
pin
Can
ola
(Bra
ssic
an
ap
us)
See
dsp
ecifi
cH
PT
(ho
mo
gen
tisa
tep
hy
tylt
ran
sfer
ase)
TC
(to
cop
her
ol
cycl
ase)
3P
7S
aS
oy
bea
nS
eed
-sp
ecifi
cA
t-H
PP
D(4
-hy
dro
xy
ph
eny
lpy
ruv
ate
dio
xy
gen
ase)
Vit
amin
EC
ano
laV
alen
tin
and
Mit
sky
(20
02)
P7
Sa
So
yb
ean
See
d-s
pec
ific
Eh-T
YR
A(c
ho
rism
ate
mu
tase
/pre
ph
enat
e
deh
yd
rog
enas
e)
PA
rc-5
Co
mm
on
bea
nS
eed
-sp
ecifi
cS
yn-V
TE
2(S
ynec
ho
cyst
issp
.P
CC
68
03
ho
mo
gen
tisa
tep
hy
tylt
ran
sfer
ase)
3U
sp(u
nk
no
wn
seed
pro
tein
pro
mo
ter)
Bro
adb
ean
(Vic
iafa
ba)
See
d-s
pec
ific
PtD
6(D
6-d
esat
ura
se)
PU
FA
syn
thes
isL
inse
ed(L
inu
mu
sita
tiss
imu
m)
Ab
bad
iet
al.
(20
04)
LeB
4(l
egu
min
)B
road
bea
nS
eed
-sp
ecifi
cP
SE
(D6
-elo
ng
ase)
Dc3
(hel
ian
thin
in)
D.
caro
taS
eed
-sp
ecifi
cP
tD5
(D5
-des
atu
rase
)
4P
7sa
(see
dst
ora
ge
pro
tein
pro
mo
ter)
So
yb
ean
(Gly
cin
em
ax)
See
d-s
pec
ific
At-
HP
PD
(p-h
yd
rox
yp
hen
ylp
yru
vat
e
dio
xy
gen
ase)
Vit
amin
ES
oy
bea
nK
aru
nan
and
aa
etal
.(2
00
5)
P7
saS
oy
bea
nS
eed
-sp
ecifi
cE
h-T
YR
A(b
ifu
nct
ion
alch
ori
smat
e
mu
tase
,p
rep
hen
ate
deh
yd
rog
enas
e)
PA
rc-5
(arc
elin
-5
pro
mo
ter)
Co
mm
on
bea
n
(Ph
ase
olu
svu
lga
ris)
See
d-s
pec
ific
At-
VT
E2
(ho
mo
gen
tisa
te
ph
yty
ltra
nsf
eras
e)
P7
Sa
So
yb
ean
See
d-s
pec
ific
At-
GG
H(g
eran
ylg
eran
yld
iph
osp
hat
e
hy
dra
tase
)
4O
leo
sin
(ole
osi
n
pro
mo
ter)
A.
tha
lia
na
See
d-s
pec
ific
At-
GG
H(g
eran
ylg
eran
yld
iph
osp
hat
e
hy
dra
tase
)
Vit
amin
ES
oy
bea
nK
aru
nan
and
aa
etal
.(2
00
5)
PF
AE
(fat
tyac
id
elo
ng
ase)
A.
tha
lia
na
See
d-s
pec
ific
At-
VT
E2
(ho
mo
gen
tisa
te
ph
yty
ltra
nsf
eras
e)
P7
Sa
So
yb
ean
See
d-s
pec
ific
At-
HP
PD
(4-h
yd
rox
yp
hen
ylp
yru
vat
e
dio
xy
gen
ase)
P7
Sa
So
yb
ean
See
d-s
pec
ific
Eh-T
YR
A(c
ho
rism
ate
mu
tase
/pre
ph
enat
e
deh
yd
rog
enas
e)
Plant Mol Biol (2010) 73:363–378 365
123
Ta
ble
1co
nti
nu
ed
Nu
mb
er
of
gen
esa
Pro
mo
ters
So
urc
eA
ctiv
ity
Gen
e(a
nd
pro
du
ct)
Pat
hw
ay/f
un
ctio
nT
arg
etp
lan
tR
efer
ence
4L
MW
glu
ten
inW
hea
tE
nd
osp
erm
Zm
psy
1(p
hy
toen
esy
nth
ase
1)
Car
ote
no
id,
asco
rbat
e
and
fola
te
Co
rnN
aqv
iet
al.
(20
09
b)
D-h
ord
ein
Bar
ley
En
do
sper
mP
acr
tI(p
hy
toen
ed
esat
ura
se)
Osd
ha
r(d
ehy
dro
asco
rbat
ere
du
ctas
e)
Ecf
olE
(GT
Pcy
clo
hy
dro
lase
I)
5L
MW
G(l
ow
mo
lecu
lar
wei
gh
tg
lute
nin
)
Wh
eat
(Tri
ticu
ma
esti
vum
)
En
do
sper
mp
sy1
(ph
yto
ene
syn
thas
e1
)C
aro
ten
oid
Co
rnZ
hu
etal
.(2
00
8)
Ho
rP(h
ord
ein
)B
arle
y(H
ord
eum
vulg
are
)
En
do
sper
mcr
tI(p
hy
toen
ed
esat
ura
se)
RP
5(p
rola
min
)R
ice
(Ory
zasa
tiva
)E
nd
osp
erm
lycb
(ly
cop
ene
b-cy
clas
e)
Gt1
(glu
teli
n-1
)R
ice
En
do
sper
mb
ch(b
-car
ote
ne
hy
dro
xy
lase
)
GZ
63
(c-z
ein
)C
orn
(Zea
ma
ys)
En
do
sper
mcr
tW(b
-car
ote
ne
ket
ola
se)
7P
FA
E(f
atty
acid
elo
ng
ase
1)
A.
tha
lia
na
See
d-s
pec
ific
crtW
(b-C
aro
ten
ek
eto
lase
)
crtB
(ph
yto
ene
syn
thas
e)
Car
ote
no
idC
ano
laF
uji
saw
aet
al.
(20
09)
Na
pin
Can
ola
See
d-s
pec
ific
idi
(iso
pen
ten
yl
py
rop
ho
sph
ate
iso
mer
ase)
crtZ
(b-c
aro
ten
eh
yd
rox
yla
se)
CaM
V3
5S
Co
nst
itu
tiv
ecr
tE(g
eran
ylg
eran
yl
py
rop
ho
sph
ate
syn
thas
e)
crtI
(ph
yto
ene
des
atu
rase
)
crtY
(ly
cop
ene
b-c
ycl
ase)
aE
xcl
ud
ing
the
sele
ctab
lem
ark
erg
ene
366 Plant Mol Biol (2010) 73:363–378
123

means that constitutive expression is achieved through the
additive effects of multiple tissue-specific motifs (Lam
et al. 1989). The region between nucleotides -90 and
-208 is an enhancer, which can increase promoter activity
in a linear fashion when up to four copies are present (Kay
et al. 1987). The enhancer can function in either orientation
and at different positions relative to the gene (Fang et al.
1989; Ohtsuki et al. 1998), and can also enhance the
activity of heterologous promoters to which it is attached
(Zheng et al. 2007).
Although widely used, the CaMV 35S promoter has
certain limitations such as its poor performance in mono-
cots, its suppression by feeding nematodes (Goddijn et al.
1993; Urwin et al. 1997) and the intellectual property
issues affecting its commercial deployment. For this rea-
son, alternative virus promoters with similar or improved
properties have been sought. Because the CaMV 35S
promoter has proven so successful, related caulimoviruses
have been the first port of call. Examples include promoters
from Figwort mosaic caulimovirus (FMV; Bhattacharyya
et al. 2002), Cassava vein mosaic virus (CsVMV; Verda-
guer et al. 1996), Cestrum yellow leaf curling virus
(CmYLCV; Stavolone et al. 2003), Mirabilis mosaic virus
(MiMV; Dey and Maiti 1999) and Peanut chlorotic streak
virus (PClSV; Maiti and Shepherd 1998; Bhattacharyya
et al. 2003). The CmYLCV promoter sequence is available
from Syngenta Biotechnology, Inc. for limited research
purposes, and has been used to express glyoxalase I (gly I)
to achieve salt tolerance in tobacco (Veena et al. 1999) and
mung bean (Bhomkar et al. 2008).
Badnaviruses are closely related to the caulimoviruses,
and they contain a strong constitutive promoter that tran-
scribes the entire genome. The Commelina yellow mottle
virus (CoYMV) promoter is active in tobacco (Medberry
et al. 1992), but more detailed studies have been carried out
in monocots where its activity is broad albeit not universal
(Torbert et al. 1998). The Sugarcane bacilliform virus
(ScBV) promoter is active in monocots (banana, corn,
millet and sorghum) and dicots (tobacco, sunflower, canola
and Nicotiana benthamiana) and has been shown to drive
high level gusA expression in transgenic banana and
tobacco plants (Schenk et al. 2001). A subcloned fragment
of the promoter was also shown to drive gusA expression in
sugarcane to at least the same level as achieved with the
corn Ubi-1 promoter (Braithwaite et al. 2004).
Subterranean clover stunt virus (SCSV) is a member of
the Circoviridae and has eight circular genome segments
each containing a promoter. Initial studies showed that the
promoters from segments four and seven were comparable
in strength to the CaMV 35S promoter in cotton, potato and
tobacco, and further analysis showed they were also suit-
able for expression in monocots, giving rise to the pPLEX
series of expression vectors (Schunmann et al. 2003a, b).
Plant housekeeping genes are another important source
of constitutive promoters, since housekeeping genes
encode proteins that are required by all cells for basic
functions such as core metabolism and maintenance of cell
structure and integrity. One major class of housekeeping
genes sees to the synthesis of cytoskeletal components, and
includes large families of genes encoding actins and tub-
ulins. The rice actin1 promoter drives strong transgene
expression in rice protoplasts transiently expressing gusA
(McElroy et al. 1990) and in most tissues of transgenic rice
plants (Zhang et al. 1991). Interestingly, McElroy and
colleagues showed that transient expression was abolished
in the absence of the first intron of the actin1 gene
(McElroy et al. 1990) and that the inclusion of this intron in
a chimeric CaMV 35S promoter enhanced its activity in
transgenic rice and corn plants by 40-fold (McElroy et al.
1991). In contrast, the first intron of the recently charac-
terized rice actin2 gene contains a negative regulatory
element whose removal is required for high-level promoter
activity; 2.6 kbp of upstream sequence contains all the 50
regulatory elements necessary for high-level constitutive
gusA expression in transgenic rice (He et al. 2009a). Other
actin promoters that have been used for constitutive or
near-constitutive expression include Arabidopsis ACT2
(An et al. 1996) and banana ACT1 (Hermann et al. 2001).
The ubiquitins are another highly conserved family of
housekeeping genes, some constitutively expressed (Ka-
walleck et al. 1993) others responsive to stress (Christensen
et al. 1992; Cornejo et al. 1993; Christensen and Quail
1996). For example, the Arabidopsis UBQ1 and UBQ6
promoters are active in all tobacco tissues, albeit at dif-
ferent levels (Callis et al. 1990), and the corn Ubi-1 pro-
moter is more than 10 times stronger than the CaMV35S
promoter in corn protoplasts when combined with the first
intron (Norris et al. 1993). Significant improvements in
activity have also been observed with three Arabidopsis
polyubiquitin promoters (UBQ3, UBQ10 and UBQ11) and
the potato ubi7 promoter (Garbarino et al. 1995).
Other housekeeping genes are involved in protein syn-
thesis and core metabolism. One example is NeIF4A-10, a
member of the tobacco eIF4A gene family—even a small
region of the promoter (to position -188) drives consti-
tutive expression in transgenic tobacco plants (Mandel
et al. 1995). Further characterization of the promoter
revealed a strong enhancer in the region -151 to -73 and
showed that an intron is not required to boost promoter
activity (Tiana et al. 2005). Other examples are the pro-
moters for genes encoding acetolactate synthase (ALS),
which catalyses the first step in the biosynthesis of the
branched chain amino acids leucine, isoleucine and valine,
and ACC synthase, which plays an important role in eth-
ylene synthesis. The Brassica ALS3 promoter is constitu-
tive and comparable in strength to the CaMV 35S promoter
Plant Mol Biol (2010) 73:363–378 367
123

(Baszczynski et al. 1997). VR-ACS1 is an auxin-inducible
ACC synthase gene from mung bean, whose promoter is
constitutive and 4–6 times more active than CaMV 35S in
Arabidopsis and tobacco (Cazzonelli et al. 2005). It is not
clear why the promoter is constitutive in a heterologous
background when it is tightly regulated in mung bean, but
this probably reflects the absence of a critical transcrip-
tional repressor in the transgenic plants.
With such a spectrum of diverse constitutive promoters
available it is perhaps surprising that there are so few
examples where different promoters have been incorpo-
rated into the same transgenic plant to drive multiple
transgenes (Table 1). In most of these examples, the CaMV
35S promoter has been used to regulate one transgene, with
the other(s) driven by a different viral or plant constitutive
promoter. However, there are cases where multiple pro-
moters other than the CaMV 35S promoter have been used
particularly in commercial varieties where the IP con-
straints surrounding the CaMV 35S promoter would make
commercial development more challenging.
Spatiotemporal promoters
Many different plant promoters have been described that
restrict expression to particular cells, tissues, organs or
developmental stages. Within this collection the number of
different promoters that can be used to drive coordinated
transgene expression (multiple transgenes with the same
expression profile) is somewhat more limited, and case
studies in which the promoter diversity strategy has been
applied are therefore fewer in number (Table 1).
Arguably the most abundant spatiotemporal promoters
are those that restrict transgene expression to seeds, and for
many reasons the seeds are often a favored target for
transgene expression, particularly if the goal of an experi-
ment is to force the accumulation of a heterologous product
that might interfere with vegetative growth at high con-
centrations or to improve the nutritional quality of seeds
used as staple foods. Many promoters have been identified
that target genes specifically to the seed, or to a particular
region of the seed. Storage proteins such as corn zein
(Schernthaner et al. 1988), rice glutelin (Leisy et al. 1989;
Takaiwa et al. 1991; Zheng et al. 1993), barley hordein
(Marris et al. 1988), rice prolamin (Qu and Takaiwa, 2004)
and wheat glutenin (Colot et al. 1987) have been rich
sources of seed-specific promoters, predominantly directing
expression to the endosperm (Wobus et al. 1995). Addi-
tional promoters have been shown to direct gene expression
to the embryo and aleurone (Opsahl-Sorteberg et al. 2004;
Qu and Takaiwa 2004; Furtado and Henry 2005).
The spatial and temporal activities of seed-specific
promoters reflect their peculiar combination of cis-acting
regulatory motifs (Onodera et al. 2001). The endosperm-
specific bZIP transcription factors O2 (Opaque-2; corn) and
SPA (storage protein activator; wheat) have been shown to
activate prolamin genes by binding to the GCN4-like motif
in the bifactorial endosperm box (Albani et al. 1997;
Schmidt et al. 1992). The GCN4 motif, a cis-acting ele-
ment that is highly conserved in the promoters of cereal
seed storage protein genes, plays a central role in con-
trolling endosperm-specific expression (Wu et al. 1998).
The barley endosperm-specific bZIP transcriptional acti-
vators BLZ1 and BLZ2 activate storage protein genes by
interacting with the GCN4 motif in barley B1-hordein
promoters (Vicente-Carbajosa et al. 1998; Onate et al.
1999), and the rice bZIP transcription factor, RISBZ1
works in a similar manner (Onodera et al. 2001). It has
been demonstrated that GCN4 and AACA motifs are
conserved in all glutelin gene promoters isolated thus far
(Takaiwa et al. 1996), and the GCN4 motif is also observed
in the PPK, SBE1 and 10, 13 (PG5a) and 16 kDa prolamin
promoters. This motif acts as a key element conferring
aleurone and subaleurone-specific expression (Wu et al.
1998; Qu and Takaiwa 2004).
Anther-specific promoters are also very useful because
they can be used to control male fertility, an important trait
in crop breeding. Numerous anther-specific and pollen-
specific promoters have been identified in a variety of
plants, including the TA29 promoter from tobacco (Kol-
tunow et al. 1990), the A9 promoter from Arabidopsis (Paul
et al. 1992) and the RA8 promoter from rice (Jeon et al.
1999). A cis-acting element found between 285 and
2,207 bp upstream of the TA29 gene confers strong, tape-
tum-specific expression and appears to work in heterolo-
gous settings, as it has been used to create male-sterile
tobacco and rapeseed plants through the expression of
Bacillus amyloliquefaciens barnase, a potently cytotoxic
ribonuclease (Mariani et al. 1990). Fertility can be restored
in F1 plants if female barnase transgenic parents are cros-
sed with males expressing the barnase inhibitor barstar
(Mariani et al. 1992), but the frequency of full restoration
can be low due to poor control over transgene expression.
Bisht et al. (2004, 2007) addressed this by enhancing and
augmenting the expression of barstar using a combination
of gene optimization, promoter engineering and MGT. The
wild-type and synthetic barstar genes were expressed using
either the TA29 and A9 promoters separately, or a chimeric
promoter combining elements from each. Although the
chimeric promoter was more active than either of the
individual promoters, the expression of different versions
of barstar using the two different tapetum-specific pro-
moters achieved the greatest frequency of restoration to
fertility.
Many case studies have been published in which
multiple transgenes are expressed in specific tissues by
combining the use of spatiotemporally-regulated and
368 Plant Mol Biol (2010) 73:363–378
123

constitutive promoters but there have been few examples of
studies in which different spatially-restricted promoters
have been used in the same plant. The barnase/barstar case
study described above is one example (Bisht et al. 2004,
2007). Another involved the use of five different promoters
to drive five transgenes in corn endosperm, i.e., corn
phytoene synthase 1, Pantoea ananatis phytoene desatur-
ase, Gentiana lutea lycopene b-cyclase, G. lutea b-caro-
tene hydroxylase, and Paracoccus sp. b-carotene ketolase,
controlled by the low molecular weight wheat glutenin,
barley hordein, rice prolamin, rice glutelin-1 and corn
c-zein promoters, plus the selectable marker gene under
constitutive control (Zhu et al. 2008). This study showed
how combinatorial transformation with multiple genes
could be used to generate a library of plants with different
phenotypes representing carotenoid biosynthesis. The dif-
ferent promoters ensured that, in any combination, it would
be possible to isolate plants without multiple copies of the
same promoter thus reducing the likelihood of transcrip-
tional silencing.
Inducible promoters
Arguably the most useful promoters in plants are those
responsive to the changing environment. Many different
inducible promoters have been identified in plants and
these generally fall into three categories—(1) those
responsive to endogenous signals (plant hormones); (2)
those responsive to external physical stimuli (abiotic and
biotic stresses); and (3) those responsive to external
chemical stimuli. Such promoters provide immense scope
for the precise regulation of transgene expression through
external control, ranging from the precise control of
transgene activation/inactivation in experimental settings
to the ability to activate transgenes on an agricultural scale
by the application of chemical sprays. Like constitutive
promoters, there are two major sources for inducible pro-
moters—endogenous genes and heterologous systems.
Inducible promoters have been combined with both con-
stitutive and spatiotemporal promoters in transgenic plants
but are generally not combined with each other, i.e., where
multiple genes need to be induced together it is standard
practice to place them under the control of the same pro-
moter. The inducible promoters most commonly used for
MGT strategies include hormone responsive promoters,
heat shock promoters and those responding to pathogens.
The most widely-used hormone-responsive promoters
are those induced by auxins, gibberellins and abscisic acid,
although promoters responsive to heterologous hormones
(from insects and mammals) are also useful because they
do not activate endogenous pathways. For example, Mar-
tinez et al. (1999) developed a hybrid system consisting of
the tobacco budworm ecdysone receptor ligand-binding
domain fused to the mammalian glucocorticoid receptor
DNA-binding domain and the VP16 transactivation
domain. The receptor responds to tebufenozide (an insec-
ticide better known by its trade name CONFIRM). Simi-
larly, Padidam et al. (2003) have developed a system that is
based on the spruce budworm ecdysone receptor ligand-
binding domain, and responds to another common insec-
ticide, methoxyfenozide (INTREPID). Another system
based on the European corn borer ecdysone receptor also
responds to this insecticide (Unger et al. 2002).
The application of exogenous auxins induces certain
genes within minutes so auxin-inducible promoters are
suitable for rapid responses. The cis-acting auxin response
elements in these genes comprise the motif TGTCTC adja-
cent to or overlapping a constitutive motif, such that the
auxin-response element is occupied and the constitutive
element repressed when the auxin concentration is low, and
the opposite occurs when the concentration is higher. Syn-
thetic auxin response elements can be engineered to contain
multiple TGTCTC repeats, and two domains from the pea
PS-IAA4/5 promoter can induce transcription from the
CaMV 35S basal promoter following auxin application
(Ballas et al. 1995). Additional auxin response elements
include the as-1/ocs-like element (TGACG(T/C)AAG(C/
G)(G/A)(C/A) T(G/T)ACG(T/C)(A/C)(A/C); Ellis et al.
1993). Auxin-response elements are often found combined
with response elements to other hormones, allowing com-
plex control. For example, the promoter of the brassinolide-
inducible BLE3 gene in rice also contains auxin response
elements which allow dual regulation by brassinolides and
IAA (Yang et al. 2006). Gibberellins also induce gene
expression rapidly, but in this case signaling is achieved
when the presence of gibberellic acid (GA) induces the
degradation of DELLA proteins, which bind to silencer
elements in GA-inducible genes and interfere with the
binding of transcriptional activators to GA response ele-
ments (GARE). In the absence of DELLA proteins, the GA-
regulated MYB transcription factor (GAMYB) binds to
GAREs and activates transcription (Sun and Gubler 2004;
Woodger et al. 2003). GA-regulated gene expression has
been studied exhaustively in the barley a-amylase genes,
revealing the core of the GARE to be a TAACAAA-like box
which can be repressed by abscisic acid (ABA). Skriver et al.
(1991) reported that when the GARE is fused as a tandem
repeat of six units it can confer GA responsiveness to a
CaMV 35S minimal promoter. Other studies have identified
further elements that lie a conserved distance from the
GARE and, in different combinations, help to modify the
expression profile. These elements combined with the
GARE constitute what is known as a GAR complex, which
can include the pyrimidine box (C/TCTTTT), the TATC-
CAC box, the CAACTC box and the Box1/O2S-like ele-
ment. Promoters that are inducible by ABA usually contain
Plant Mol Biol (2010) 73:363–378 369
123

one or more ABA response elements (ABREs), which may
include a G-box/C-box as well as MYC and MYB binding
sites. There have been many reports of promoters whose
activity has been modified by the incorporation of such
elements. Sequence analysis of the cotton late embryogen-
esis-abundant (LEA) D113 gene promoter (Luo et al. 2008)
revealed the presence of ABREs, a drought-response ele-
ment, and MYB and MYC sites; a 158 bp fragment was
shown to be sufficient for ABA induced reporter gene
expression. Some promoters can also be negatively regu-
lated by ABA. The corn HyPRP gene is specifically
expressed in immature embryos and its expression is
inhibited when ABA levels increase during maturation. A
construct containing 2 kbp of the HyPRP promoter joined to
the gusA gene allowed the repression of GUS activity in corn
and tobacco following the application of ABA. Two ACGT
elements at positions -260 and -93 bp in the promoter are
likely to mediate this effect (Jose-Estanyol et al. 2005).
Many metabolic genes need to respond sensitively to the
levels of specific metabolites to facilitate feedback regu-
lation loops that tie gene expression to metabolic balance.
Therefore, many promoters respond to the presence of
certain chemicals, and these can be extremely useful to
regulate transgene expression in plants because unlike
hormones they do not have pleiotropic effects on growth
and development. Sugar responsive promoters contain a
variety of response elements and these can confer sugar-
sensitivity on heterologous genes. For example, elements
from sporamin and amylase promoters have been studied in
detail and the minimal a-amylase 3 promoter makes the
normally constitutive rice actin1 promoter sensitive to the
presence of sugar (Lu et al. 1998). SURE (SUcrose
REsponsive) elements are found in sugar-inducible patatin
promoters (Grierson et al. 1994; Zourelidou et al. 2002)
and in the promoter of the VvTH1 gene, which encodes a
hexose transporter (Atanassova et al. 2003).
Plant defense genes often have promoters that are
inducible by chemicals (elicitors) produced by pests and
pathogens. Different combinations of trans-acting factors
are responsible for the elicitor-mediated activation of
defense related genes and numerous cis-acting elements
have been identified in the corresponding promoters. They
can be found as single or multiple copies and in different
combinations, allowing some genes to respond to more
than one elicitor, and particular pathogens to elicit distinct,
functionally-related sets of genes. The cis-acting elements
are highly conserved among different plant species so they
are very useful for making transgenes pathogen-inducible
in a range of heterologous settings. Individual elicitor
response elements appear to work well in a different pro-
moter context allowing a certain degree of ‘plug-and-play’
between promoters. The number of copies of each element
also appears to have an effect on the strength of induction,
thus promoters with one or two copies are best suited to
mediate pathogen-specific expression (Rushton et al.
2002). Many plants also respond to the physical aspects of
pest activity (wounding) by expressing a set of protective
genes, many of which are also inducible by jasmonate or
ethylene. Wound-inducible genes include those encoding
proteinase inhibitor II (pinII; Godard et al. 2007; Keinon-
en-Mettala et al. 1998; Keil et al. 1989; Xu et al. 1993), the
Agrobacterium tumefaciens enzymes nopaline synthase
(An et al. 1990) and mannopine synthase (Guevara-Garcıa
et al. 1993; Ni et al. 1996), and peroxidases such as rice
R2329 (Sasaki et al. 2007). The use of wound-inducible
promoters in transgenic plants shows that wound signaling
pathways are functionally conserved across taxonomic
groups (Wu et al. 1999; Yevtushenko et al. 2004).
Elevated temperatures result in the expression of so-
called heat shock proteins (HSPs), which protect plants
from the effects of heat by helping to stabilize proteins and
other cellular components thus conferring thermotolerance.
HSP genes have heat-inducible promoters that are also
strongly conserved across species. For example, the soy-
bean hsp17.3B promoter was used to drive gusA expression
in the moss Physcomitrella patens resulting in no GUS
activity at 25�C but increasing GUS activity at higher
temperatures (Saidi et al. 2005). Heat shock elements
(HSEs) were identified in the tomato ftsH6 promoter, and
were recognized by another HSP acting as a transcription
factor (heat shock factor, HSF; Sun et al. 2006). The heat
shock response may be universal or tissue specific. The
Arabidopsis GolS1 promoter confers universal GUS
activity in transgenic Arabidopsis plants following a tem-
perature shift (Panikulangara et al. 2004), whereas the
barley hspl7 promoter confers heat-induced expression
only in the xylem bundles of the stem and petioles—this
expression profile is conserved in transgenic monocot and
dicot plants (Raho et al. 1996).
Repetitious promoters and transcriptional
gene silencing
The promoter diversity approach discussed above is
employed to avoid repetitious use of the same promoter,
which can in some cases promote transcriptional gene
silencing through promoter methylation and inactivation.
This effect is usually constitutive but may be tissue-specific
(Kloti et al. 2002). However, there have been plenty of
reports describing transgenic plants carrying multiple
transgenes under the control of the same promoter yet
showing strong and stable expression. For example,
although Zhu et al. (2008) used five different endosperm-
specific promoters in corn to achieve the high-level
expression of carotenogenic genes, Naqvi et al. (2009b)
370 Plant Mol Biol (2010) 73:363–378
123

achieved strong expression of four genes in the same sys-
tem using the barley D-hordein promoter to control each
transgene, with no adverse effects. The literature suggests
that repetitious use of the same promoter is often suc-
cessful, so what are the exact limitations of using the same
promoter to control more than one transgene?
To answer this question, it is necessary to look more
closely at the reasons for gene silencing caused by repetitive
promoter use. In plants, transcriptional gene silencing
resulting from sequence homology in promoter regions is
correlated with increased promoter methylation (Kooter
et al. 1999) and appears to be driven by the production of
double-stranded RNA (dsRNA) matching the promoter
sequence (Mette et al. 2000). This has been demonstrated
by deliberately expressing dsRNA corresponding to the nos
promoter in transgenic plants carrying a second transgene
driven by the nos promoter (Mette et al. 1999) and by
constructing transgenic plants in which the transgene locus
triggers both transcriptional and post-transcriptional
silencing simultaneously, by producing dsRNA corre-
sponding to the promoter and transcribed sequences of
different target genes (Mourrain et al. 2007). The compo-
nents of this RNA-driven DNA methylation system have
been systematically sought and investigated, and include
RNA-dependent RNA polymerase 2 (RDR2) and Dicer-like
3 (DCL3; Xie et al. 2004), Pol IV (Herr et al. 2005; Kanno
et al. 2005; Onodera et al. 2005), Pol IVb/Pol V (Pontier
et al. 2005; Wierzbicki et al. 2008; He et al. 2009b), Arg-
onaute 4 (AGO4; Zilberman et al. 2003), chromatin
remodeling protein DRD1 (Kanno et al. 2004), and de novo
DNA methyltransferase DRM2 (Cao and Jacobsen 2002).
In the absence of deliberately-created promoter dsRNA,
the transcriptional silencing seen in some transgenic plants
carrying multiple copies of the same promoter appears to
arise from dsRNA produced serependitiously. The organi-
zation of a transgenic locus is difficult to control, and it is
therefore a common occurrence in both Agrobacterium-
mediated transformation and direct DNA transfer that the
juxtaposition of transgenes or fragments thereof can result
in the creation of hairpin promoter structures at the DNA
level that are transcribed into aberrant dsRNA species.
Such arrangements can be obvious and easy to detect, but
even where gross rearrangements are absent it is possible
that undetected ‘micro-rearrangements’ are present in the
transgenic locus, as observed by Mehlo et al. (2000) when
investigating the structure of a transgenic locus in corn
generated by direct DNA transfer. The siRNAs that trigger
RNA-dependent DNA methylation are just 24 bp in length,
so it is conceivable that inverted repeats of \50 bp could
be sufficient for transgene silencing, and such structures
would be undetectable using the coarse analysis methods
typically employed to study transgenic plants, such as
Southern blot hybridization.
There are at least four mechanisms by which promoter-
specific hairpin RNAs could be generated in transgenic
plants carrying different transgenes under the control of the
same promoter. In many cases, the creation of hairpin RNA
structures may be a consequence of the relative position of
intact transgenes, which is itself a reflection of the mecha-
nism of transgene integration. The organization of integrated
T-DNA sequences differs among Agrobacterium strains, but
a common feature of nopaline-type derivatives such as C58
is the preferential integration of T-DNA as dimers with an
inverted repeat configuration, linked either at the left or right
borders (Jones et al. 1987; Jorgensen et al. 1987). Where
cotransformation is carried out with two T-DNAs containing
different genes, the different T-DNAs often integrate as
heterodimeric inverted repeats, preferentially around the
right border (De Block and Debrouwer 1991). If the same
promoter is used for both genes, this would favor the for-
mation of hairpin structures that could be transcribed from
the opposite strand. The structure of loci generated by direct
DNA transfer is more variable, but inverted repeat structures
involving promoter sequences are not uncommon, allowing
the same silencing mechanism to operate (Kohli et al. 2003).
The second mechanism reflects the frequent occurrence
of spontaneous transgene rearrangements (truncation,
fragmentation, recombination) which can be thought of as
‘collateral damage’ occurring to DNA during the transfor-
mation process due to the activities of nucleases and repair
complexes that facilitate the integration of exogenous DNA.
There have been few systematic studies of this phenomenon
in Agrobacterium-mediated transformation, but Afolabi
et al. (2004) and Zhu et al. (2006) found that nonintact
T-DNAs were present in [70% of transgenic rice lines, in
most cases reflecting loss of the mid to right border portion
of the T-DNA. Similarly, Rai et al. (2007) found that about
50% of rice plants transformed with a T-DNA containing
the phytoene synthase (psy) and phytoene desaturase (crtI)
genes showed evidence of rearrangements, and in the
majority of cases the rearrangements occurred in the crtI
expression cassette, which was adjacent to the right T-DNA
border. Rearrangements involving the left border are often
characterized by the insertion of variable-size regions of
filler DNA, which may be derived from the T-DNA
sequence or from plant genomic DNA, providing further
scope for the creation of hairpin structures (De Buck et al.
1999; Kumar and Fladung 2000, 2002).
A third mechanism for silencing triggered by repetitive
promoter usage reflects the structure of the promoter itself.
Rearrangements affecting the promoter of a transgene may
be more common than expected due to the presence of
specific recombinogenic sequences, such as the imperfect
palindromic structure that is present in the CaMV 35S
promoter (Kohli et al. 1999). This sequence has the ability
to adopt a cruciform secondary structure, which may
Plant Mol Biol (2010) 73:363–378 371
123

stimulate recombination events. Notably, many other pro-
moters contain palindromic sequences of variable length
within 100 bp of the transcription start site, often because
they bind dimeric transcription factors, so it is possible that
promoters may be disproportionately involved in transgene
rearrangement events, a hypothesis that will need to be
examined through the detailed analysis of transgenic pop-
ulations carrying multiple copies of other promoters.
Evidence from many transformation experiments indi-
cates that there is no simple correlation between transgene
copy number and expression level, with the exception of
certain carefully controlled experiments using boundary
elements. In some cases, higher copy numbers have sup-
pressed overall expression levels (e.g., Cannell et al. 1999;
Spencer et al. 1992) whereas in others higher copy numbers
have enhanced expression (Stoger et al. 1998; Gahakwa
et al. 2000). Where suppression effects have occurred, it has
been suggested that ‘‘runaway expression’’ resulting in the
generation of aberrant RNAs lacking polyadenylate tails
has triggered potent silencing through the post-transcrip-
tional silencing pathway (Schubert et al. 2004). From the
prospect of promoter usage, this means in a practical sense
that multiple copies of the same promoter can appear in a
transgenic locus because of the presence of multiple copies
of the same transgene, so why should it matter if the pro-
moter is also represented with different transgenes? A
potential answer to this question comes from experiments
involving the use of the CaMV 35S promoter for large-scale
screening, and reveals a fourth mechanism that may trigger
transcriptional silencing. The CaMV 35S promoter and
enhancer have been used as a random insertional mutagen
to hyperactivate adjacent genes and generate gain-of-func-
tion phenotypes. It has been noted that such screens using
T-DNA cassettes containing the enhancer elements from
the CaMV 35S promoter return a lower than expected fre-
quency of morphological mutants (Chalfun et al. 2003).
Detailed analysis revealed a correlation between the num-
ber of T-DNA insertion sites, the methylation status of the
enhancer sequence and enhancer activity. All plants con-
taining more than a single T-DNA insert were methylated
on the enhancer and its activity was reduced, with the
amount of methylation and the reduction of enhancer
activity correlating with the number of T-DNA copies,
particularly those with right border inverted repeats (Chal-
fun et al. 2003). Even so, methylation was still detected at a
lower frequency in plants without right border inverted
repeats suggesting other triggers were active in these lines.
Alternative solutions
Research into the structure and activity of plant promoters
has allowed the identification of specific cis-acting elements
with defined functions, so the next generation of promoters
is likely to comprise a mix and match of cis-acting elements
that confer upon transgenes exactly the properties required
by the experimenter. To a certain extent this has already
been achieved through the use of heterologous recombinant
promoters such as the inducible promoters based on insect
hormones (see above), but the key disadvantage of these
systems is that they require the corresponding trans-acting
factors to be imported, a problem that would not be
encountered when using recombined endogenous elements
to generate bespoke plant promoters. In some cases, the aim
is simply to increase the activity of an endogenous pro-
moter, which can be achieved in the case of the CaMV 35S
promoter by duplicating its enhancer elements, a strategy
that also works when these elements are imported into other
promoters. Chimeric promoters have also been engineered
using sequences from the CoYMV and CsVMV major
transcript promoters as activating sequences to augment the
CaMV 35S promoter, resulting in higher activity compared
to CaMV 35S in stably transformed tobacco plants and
higher activity compared to corn c-zein in corn endosperm
(Rance et al. 2002). In other cases, it is desirable to con-
strain promoter activity in some way, which has been
achieved with synthetic auxin response elements that make
heterologous promoters more auxin-sensitive. In such cases,
the TGTCTC element is placed either as tandem or inverted
repeats with appropriate spacing, making the promoters up
to 10 times more sensitive to exogenous auxins (Guilfoyle
1999). The ‘super c-zein promoter’ contains a duplication
(region -444 to -174) from the corn endosperm-specific
27 kDa c-zein promoter, making it more active in endo-
sperm tissue (Marzabal et al. 1998). This has been used to
increase the expression of the bacterial genes crtB and crtI
resulting in greater accumulation of b-carotene (Aluru et al.
2008). Many plant promoters can also be enhanced by
including an intron (e.g., Vain et al. 1996; Mitsuhara et al.
1996; Fiume et al. 2004; Chiera et al. 2007).
Can this strategy of modulating promoter structure be
used to reduce promoter duplication in MGT? Several
reports have described the creation of artificial promoters
specifically to avoid repetition. For example, Sawant et al.
(2001) have described a strategy in which the sequences of
the strongest plant promoters were compared, to facilitate
the design of ideal artificial consensus promoters. By
combining the ideal minimal cassette (TATA box, initiator
and Kozak sites) with the ideal upstream activator, they
were able to develop a 450-bp synthetic promoter that was
active in cotton leaves, potato tubers and cabbage stems,
and was stronger than the CaMV 35S promoter in trans-
genic tobacco plants. In theory, several such artificial pro-
moters could be developed and used in combination, or with
native promoters. Bhullar et al. (2003) have developed two
alternative approaches to avoid promoter homology, one in
372 Plant Mol Biol (2010) 73:363–378
123

which functional cis-acting elements are embedded in dif-
ferent synthetic DNA sequences, and another in which the
functional cis-acting elements of different promoters with
the same activity are swapped to create non-homologous
chimeras. A set of modified CaMV 35S promoters was
developed and tested in transgenic tobacco plants, showing
that the chimeric promoters were generally weaker than the
native CaMV 35S promoter but that promoters constructed
from cis-elements embedded in a synthetic DNA sequence
were generally comparable in strength.
Most promoters used in plant biotechnology are unidi-
rectional, but bidirectional promoters are becoming
increasingly useful for MGT experiments as they allow the
simultaneous expression of two gene products. In many
promoters, only the core promoter sequence needs to be
oriented with respect to the coding sequence of the gene
to ensure transcription in the correct direction. Often,
upstream promoter elements can be present in either ori-
entation, and enhancer elements enjoy even more freedom
in terms of position and orientation. If a minimal promoter
consisting of a TATA box and initiator is duplicated and
placed facing outwards, cis-acting elements between the
minimal promoters can be used to drive transcription in
two directions simultaneously. The duplicated minimal
CaMV 35S promoter has been used for bidirectional tran-
scription in several different species (Xie et al. 2001;
Zhang et al. 2008) and can also be combined with other
promoters or synthetic response elements to make bidi-
rectional chimeric regulatory complexes (Chaturvedi et al.
2006). Xie et al. (2001) fused the minimal CaMV 35S
promoter to the constitutive PClSV promoter on one side
and the JA-sensitive OPR1 promoter at the other, and fused
a minimal promoter derived from the senescence-specific
SAG12 gene to the 50 end of the CaMV 35S promoter.
Naturally bidirectional promoters have been reported in
plants, including those directing the oleosin and methionine
sulphoxide reductase genes in canola (Keddie et al. 1994;
Sadanandom et al. 1996), the Arabidopsis cab1 and cab2
genes (chlorophyll a/b-binding protein; Mitra et al. 2009)
and the rice Ocip1 and Ocpi2 genes (chymotrypsin prote-
ase inhibitor; Singh et al. 2009). Bioinformatic analysis of
Arabidopsis, rice and poplar genome sequences has iden-
tified several putatively paired genes whose promoters may
be useful in the future for bidirectional gene expression
constructs (Dhadi et al. 2009). The different promoter
strategies discussed above are summarized in Fig. 1.
Conclusions and outlook
MGT is here to stay and in the future it will be desirable,
even essential, to increase the number of simultaneously
introduced transgenes still further (Naqvi et al. 2009a).
Currently, the creation of transgenic plants with stable
transgene expression relies on a healthy dose of luck, with
the best performing plants being selected from large pop-
ulations, the remaining plants being discarded if they fail to
live up to expectations. As MGT experiments become more
ambitious, the recovery rates will start to suffer from the
law of diminishing returns. How many plants will need to
be screened to find a line that stably expresses 10–20 or
more different transgenes? This is a realistic outlook for the
forthcoming decade, and in order to achieve such ambitious
objectives it will be necessary to minimize the likelihood
of transgene silencing triggered by the repetitive use of the
same promoter sequence. This will be addressed by a
number of strategies, including promoter diversity (facili-
tated by the discovery and characterization of new pro-
moters with relevant expression profiles), the use of
synthetic and modified promoters to reduce the extent of
homology between transgenes, and the use of bidirectional
promoters to reduce the promoter to transgene ratio
(a)
(b)
(c)
(d)
(e)
(f)
Fig. 1 Strategies for promoter deployment in nuclear multigene
transformation. a Repetitious use of the same promoter is a
convenient approach that often works effectively, but some research-
ers have found that it increases the risk of homology-dependent
transcriptional gene silencing. The following alternative approaches
may therefore be chosen instead. b Promoter diversity involves the
use of different promoters for each transgene, although this depends
on availability and becomes more difficult with larger numbers of
transgenes. c Synthetic consensus promoters can be developed from
the conserved features of native promoter sequences, and can add to
the repertoire of available promoters (Sawant et al. 2001). d Chimeric
promoters comprising the functional segments of native promoters
can be prepared by domain swapping (e.g., Bisht et al. 2004) or by
importing specific functional motifs from one promoter into another
(e.g., introducing the CaMV 35S enhancer or actin1 intron into
heterologous promoters to increase activity, or introducing hormone
response elements into constitutive promoters to make them induc-
ible). e The functional cis-elements of natural promoters can be
embedded in a synthetic DNA sequence (e.g., Bhullar et al. 2003).
f Bidirectional promoters can halve the number of promoters required
for MGT. Strategies d, e and f can be combined in different ways to
generate functionalized unidirectional and bidirectional chimeric
promoters with potentially limitless diversity
Plant Mol Biol (2010) 73:363–378 373
123

(Fig. 1). Novel approaches, such as the use of episomal
constructs like plant artificial chromosomes will also play a
role in the future, although for the time being such systems
are limited to a few model species and are technically
demanding.
Acknowledgments Work in our laboratory is supported by the
Ministry of Science and Innovation, Spain (BFU2007-61413 and
BIO2007-30738-E), a European Research Council Advanced Grant
(BIOFORCE) to PC, and the CONSOLIDER Agrigenomics program
funded by MICINN, Spain.
References
Abbadi A, Domergue F, Bauer J, Napier JA, Welti R, Zahringer U,
Cirpus P, Heinz E (2004) Biosynthesis of very-long-chain
polyunsaturated fatty acids in transgenic oilseeds: constraints on
their accumulation. Plant Cell 16:2734–2748
Afolabi AS, Worland B, Snape JW, Vain P (2004) A large-scale study
of rice plants transformed with different T-DNAs provides new
insights into locus composition and T-DNA linkage configura-
tions. Theor Appl Genet 109:815–826
Albani D, Hammond-Kosack MCU, Smith C, Conlan S, Colot V,
Holdsworth M, Bevan MW (1997) The wheat transcriptional
activator SPA: a seed-specific bZIP protein that recognizes the
GCN4-like motif in the bifactorial endosperm box of prolamin
genes. Plant Cell 9:171–184
Aluru M, Xu Y, Guo R, Wang Z, Li S, White W, Wang K, Rodermel
S (2008) Generation of transgenic maize with enhanced provi-
tamin A content. J Exp Bot 59:3551–3562
An G, Costa MA, Ha SB (1990) Nopaline synthase promoter is
wound inducible and auxin inducible. Plant Cell 2:225–233
An YQ, McDowell JM, Huang S, McKinney EC, Chambliss S,
Meagher RB (1996) Strong, constitutive expression of the
Arabidopsis ACT2/ACT8 actin subclass in vegetative tissues.
Plant J 10:107–121
Atanassova R, Leterrier M, Gaillard C, Agasse A, Sagot E, Coutos-
Thevenot P, Delrot S (2003) Sugar-regulated expression of a
putative hexose transport gene in grape. Plant Physiol 131:326–
334
Ballas N, Wong LM, Ke M, Theologis A (1995) Two auxin-
responsive domains interact positively to induce expression of
the early indoleacetic acid-inducible gene PS-IAA4/5. Proc Natl
Acad Sci USA 92:3483–3487
Baszczynski CL, Barbour E, Miki B (1997) ALS3 promoter. US
Patent 5659026
Bhattacharyya S, Dey N, Maiti IB (2002) Analysis of cis-sequence of
subgenomic transcript promoter from the Figwort mosaic virusand comparison of promoter activity with the Cauliflower mosaicvirus promoters in monocot and dicot cells. Virus Res 90:47–62
Bhattacharyya S, Pattanaik S, Maiti IB (2003) Intron-mediated
enhancement of gene expression in transgenic plants using
chimeric constructs composed of the Peanut chlorotic streakvirus (PClSV) promoter-leader and the antisense orientation of
PClSV ORF VII (p7R). Planta 218:115–124
Bhomkar P, Upadhyay C, Saxena M, Muthusamy A, Prakash NS,
Sarin NB (2008) Salt stress alleviation in transgenic Vignamungo L. Hepper (blackgram) by overexpression of the glyox-
alase I gene using a novel Cestrum yellow leaf curling virus(CmYLCV) promoter. Mol Breed 22:169–181
Bhullar S, Chakravarthy S, Advani S, Datta S, Pental D, Burma PK
(2003) Strategies for development of functionally equivalent
promoters with minimum sequence homology for transgene
expression in plants: cis-elements in a novel DNA context versus
domain swapping. Plant Physiol 132:988–998
Bisht NC, Jagannath A, Gupta V, Kumar Burma P, Pental D (2004) A
two gene–two promoter system for enhanced expression of a
restorer gene (barstar) and development of improved fertility
restorer lines for hybrid seed production in crop plants. Mol
Breed 14:129–144
Bisht NC, Jagannath A, Burma PK, Pradhan AK, Pental D (2007)
Retransformation of a male sterile barnase line with the barstar
gene as an efficient alternative method to identify male sterile-
restorer combinations for heterosis breeding. Plant Cell Rep
26:727–733
Braithwaite KS, Geijskes RJ, Smith GR (2004) A variable region of
the sugarcane bacilliform virus (SCBV) genome can be used to
generate promoters for transgene expression in sugarcane. Plant
Cell Rep 23:319–326
Callis J, Raasch JA, Vierstras RD (1990) Ubiquitin extension proteins
of Arabidopsis thaliana. Structure, localization, and expression
of their promoters in transgenic tobacco. J Biol Chem 265:
12466–12493
Cannell ME, Doherty A, Lazzeri PA, Barcelo P (1999) A population
of wheat and tritordeum transformants showing a high degree of
marker gene stability and heritability. Theor Appl Genet 99:
772–784
Cao X, Jacobsen SE (2002) Locus-specific control of asymmetric and
CpNpG methylation by the DRM and CMT3 methyltransferase
genes. Proc Natl Acad Sci USA 99:16491–16498
Cazzonelli CI, McCallum EJ, Lee R, Botella JR (2005) Character-
ization of a strong, constitutive mung bean (Vigna radiata L.)
promoter with a complex mode of regulation in planta.
Transgenic Res 14:941–967
Chalfun A, Mes JJ, Mlynarova L, Aarts MGM, Angenent GC (2003)
Low frequency of T-DNA based activation tagging in Arabid-
opsis is correlated with methylation of CaMV 35S enhancer
sequences. FEBS Lett 555:459–463
Chaturvedi CP, Sawant SV, Kiran K, Mehrotra R, Lodhi N, Ansari
SA, Tuli R (2006) Analysis of polarity in the expression from a
multifactorial bidirectional promoter designed for high-level
expression of transgenes in plants. J Biotechnol 123:1–12
Chiera JM, Bouchard RA, Dorsey SL, Park E, Buenrostro-Nava MT,
Ling PP, Finer JJ (2007) Isolation of two highly active soybean
(Glycine max (L.) Merr.) promoters and their characterization
using a new automated image collection and analysis system.
Plant Cell Rep 26:1501–1509
Christensen AH, Quail PH (1996) Ubiquitin promoter-based vectors
for high-level expression of selectable and/or screenable marker
genes in monocotyledonous plants. Transgenic Res 5:213–218
Christensen AH, Sharrock RA, Quail PH (1992) Maize polyubiquitin
genes: structure, thermal perturbation of expression and tran-
script splicing, and promoter activity following transfer to
protoplasts by electroporation. Plant Mol Biol 18:675–689
Colot V, Robert LS, Kavanagh TA, Bevan MW, Thompson RD
(1987) Localization of sequences in wheat endosperm protein
genes which confer tissue-specific expression in tobacco. EMBO
J 6:3559–3564
Cornejo MJ, Luth D, Blankenship KM, Anderson OD, Blechl AE
(1993) Activity of a maize ubiquitin promoter in transgenic rice.
Plant Mol Biol 23:567–581
De Block M, Debrouwer D (1991) Two T-DNAs co-transformed into
Brassica napus by a double Agrobacterium tumefaciens infec-
tion are mainly integrated at the same locus. Theor Appl Genet
82:257–263
De Buck S, Jacobs A, Van Montagu M, Depicker A (1999) The DNA
sequences of T-DNA junctions suggest that complex T-DNA
loci are formed by a recombination process resembling T-DNA
integration. Plant J 20:295–304
374 Plant Mol Biol (2010) 73:363–378
123

Dey N, Maiti IB (1999) Structure and promoter/leader deletion
analysis of mirabilis mosaic virus (MMV) full-length tran-
script promoter in transgenic plants. Plant Mol Biol 40:
771–782
Dhadi SR, Krom N, Ramakrishna W (2009) Genome-wide compar-
ative analysis of putative bidirectional promoters from rice,
Arabidopsis and Populus. Gene 429:65–73
Ellis JG, Tokuhisa JG, Llewellyn DJ, Bouchez D, Singh K, Dennis
ES, Peacock WJ (1993) Does the ocs-element occur as a
functional component of the promoters of plant genes? Plant J
4:433–443
Fang RX, Nagy F, Sivasubramaniam S, Chua NH (1989) Multiple cis-
regulatory elements for maximal expression of the Cauliflowermosaic virus 35S promoter in transgenic plants. Plant Cell
1:141–150
Farre G, Ramessar K, Twyman RM, Capell T, Christou P (2010) The
humanitarian impact of plant biotechnology: recent break-
throughs vs bottlenecks for adoption. Curr Opin Plant Biol (in
press; doi:10.1016/j.pbi.2009.11.002)
Fiume E, Christou P, Giani S, Breviario D (2004) Introns are key
regulatory elements of rice tubulin expression. Planta 218:
693–703
Fujisawa M, Takita E, Harada H, Sakurai N, Suzuki H, Ohyama K,
Shibata D, Misawa N (2009) Pathway engineering of Brassicanapus seeds using multiple key enzyme genes involved in
ketocarotenoid formation. J Exp Bot 60:1319–1332
Furtado A, Henry RJ (2005) The wheat Em promoter drives reporter
gene expression in embryo and aleurone tissue of transgenic
barley and rice. Plant Biotechnol J 3:421–434
Gahakwa D, Maqbool SB, Fu X, Sudhakar D, Christou P, Kohli A
(2000) Transgenic rice as a system to study the stability of
transgene expression: multiple heterologous transgenes show
similar behaviour in diverse genetic backgrounds. Theor Appl
Genet 101:388–399
Garbarino JE, Oosumi T, Belknap WR (1995) Isolation of a
polyubiquitin promoter and its expression in transgenic potato
plants. Plant Physiol 109:1371–1378
Godard KA, Byun-McKay A, Levasseur C, Plant A, Seguin A,
Bohlmann J (2007) Testing of a heterologous, wound- and
insect-inducible promoter for functional genomics studies in
conifer defense. Plant Cell Rep 26:2083–2090
Goddijn OJM, Lindsey K, van der Lee FM, Klap JC, Sijmons PC
(1993) Differential gene expression in nematode induced feeding
structures of transgenic plant harbouring promoter-gusA fusion
constructs. Plant J 4:863–873
Grierson C, Du JS, de Torres Zabala M, Beggs K, Smith C,
Holdsworth M, Bevan M (1994) Separate cis sequences and
trans factors direct metabolic and developmental regulation of a
potato tuber storage protein gene. Plant J 5:815–826
Guevara-Garcıa A, Mosqueda-Cano G, Arguello-Astorga G, Simpson
J, Herrera-Estrella L (1993) Tissue-specific and wound-inducible
pattern of expression of the mannopine synthase promoter is
determined by the interaction between positive and negative cis-
regulatory elements. Plant J 4:495–505
Guilfoyle TJ (1999) Auxin-regulated genes and promoters. In:
Hooykaas PJJ, Hall M, Libbenga KL (eds) Biochemistry
and molecular biology of plant hormones. Elsevier, London,
pp 423–459
He C, Lin Z, McElroy D, Wu R (2009a) Identification of a rice actin2gene regulatory region for high-level expression of transgenes in
monocots. Plant Biotechnol J 7:227–239
He XJ, Hsu YF, Pontes O, Zhu J, Lu J, Bressan RA, Pikaard C, Wang
CS, Zhu JK (2009b) NRPD4, a protein related to the RPB4
subunit of RNA polymerase II, is a component of RNA
polymerases IV and V and is required for RNA-directed DNA
methylation. Genes Dev 23:318–330
Hermann SR, Harding RM, Dale JL (2001) The banana actin 1
promoter drives near-constitutive transgene expression in veg-
etative tissues of banana (Musa spp.). Plant Cell Rep 20:525–530
Herr AJ, Jensen MB, Dalmay T, Baulcombe DC (2005) RNA
polymerase IV directs silencing of endogenous DNA. Science
308:118–120
Jeon J-S, Chung Y-Y, Lee S, Yi G-H, Oh B-G, An G (1999) Isolation
and characterization of an anther-specific gene, RA8, from rice
(Oryza sativa, L). Plant Mol Biol 39:35–44
Jones JDG, Gilbert DE, Grady KL, Jorgensen RA (1987) T-DNA
structure and gene expression in petunia plants transformed by
Agrobacterium tumefaciens C58 derivates. Mol Gen Genet
207:478–485
Jorgensen RA, Snyder C, Jones JDG (1987) T-DNA is organized
predominantly in inverted repeat structures in plants transformed
with Agrobacterium tumefaciens C58 derivatives. Mol Gen
Genet 207:471–477
Jose-Estanyol M, Perez P, Puigdomenech P (2005) Expression of the
promoter of HyPRP, an embryo-specific gene from Zea mays in
maize and tobacco transgenic plants. Gene 356:146–152
Kanno T, Mette MF, Kreil DP, Aufsatz W, Matzke M, Matzke AJ
(2004) Involvement of putative SNF2 chromatin remodeling
protein DRD1 in RNA-directed DNA methylation. Curr Biol
14:801–805
Kanno T, Huettel B, Mette MF, Aufsatz W, Jaligot E, Daxinger L,
Kreil DP, Matzke M, Matzke AJ (2005) Atypical RNA
polymerase subunits required for RNA-directed DNA methyla-
tion. Nature Genet 37:761–765
Karunanandaa B, Qi Q, Hao M, Baszis SR, Jensen PK, Wong YH,
Jiang J, Venkatramesh M, Gruys KJ, Moshiri F, Post-Beittenm-
iller D, Weiss JD, Valentin HE (2005) Metabolically engineered
oilseed crops with enhanced seed tocopherol. Metab Eng 7:
384–400
Kawalleck P, Somssich IE, Feldbrugge M, Hahlbrock K, Weisshaar B
(1993) Polyubiquitin gene expression and structural properties of
the ubi 2–4 gene in Petroselinum crispum. Plant Mol Biol
21:673–684
Kay R, Chan A, Daly M, McPherson J (1987) Duplication of CaMV
35S promoter sequences creates a strong enhancer for plant
genes. Science 236:1299–1302
Keddie JS, Tsiantis M, Piffanelli P, Cella R, Hatzopoulos P, Murphy
DJ (1994) A seed-specific Brassica napus oleosin promoter
interacts with a G-box-specific protein and may be bi-directional.
Plant Mol Biol 24:327–340
Keil M, Sanchez-Serrano JJ, Willmitzer L (1989) Both wound-
inducible and tuber-specific expression are mediated by the
promoter of a single member of the potato proteinase inhibitor II
gene family. EMBO J 5:1323–1330
Keinonen-Mettala K, Pappinen A, Von Weissenberg K (1998)
Comparisons of the efficiency of some promoters in silver birch
(Betula pendula). Plant Cell Rep 17:356–361
Kloti A, He X, Potrykus I, Hohn T, Futterer J (2002) Tissue-specific
silencing of a transgene in rice. Proc Natl Acad Sci USA
99:10881–10886
Kohli A, Griffiths S, Palacios N, Twyman RM, Vain P, Laurie DA,
Christou P (1999) Molecular characterization of transforming
plasmid rearrangements in transgenic rice reveals a recombina-
tion hotspot in the CaMV 35S promoter and confirms the
predominance of microhomology-mediated recombination. Plant
J 17:591–601
Kohli A, Twyman RM, Abranches A, Wegel E, Shaw P, Christou P,
Stoger E (2003) Transgene integration, organization and inter-
action in plants. Plant Mol Biol 52:247–258
Koltunow AM, Truettner J, Cox KH, Walroth M, Goldberg RB
(1990) Different temporal and spatial gene expression patterns
occur during anther development. Plant Cell 2:1201–1224
Plant Mol Biol (2010) 73:363–378 375
123

Kooter J, Matzke MA, Meyer P (1999) Listening to the silent genes:
transgene silencing, gene regulation and pathogen control.
Trends Plant Sci 4:340–346
Kumar S, Fladung M (2000) Transgene repeats in aspen: molecular
characterisation suggests simultaneous integration of indepen-
dent T-DNAs into receptive hotspots in the host genome. Mol
Gen Genet 264:20–28
Kumar S, Fladung M (2002) Transgene integration in aspen:
structures of integration sites and mechanism of T-DNA
integration. Plant J 31:543–551
Lam E, Benfey PN, Gilmartin PM, Fang RX, Chua NH (1989) Site-
specific mutations alter in vitro factor and change promoter
expression pattern in transgenic plants. Proc Natl Acad Sci USA
86:7890–7894
Leisy DJ, Hnilo J, Zhao Y, Okita TW (1989) Expression of a rice
glutelin promoter in transgenic tobacco. Plant Mol Biol 14:
41–50
Lu CA, Lim EK, Yu SM (1998) Sugar response sequence in the
promoter of a rice alpha-amylase gene serves as a transcriptional
enhancer. J Biol Chem 273:10120–10131
Luo K, Zhang G, Deng W, Luo F, Qiu K, Pei Y (2008) Functional
characterization of a cotton late embryogenesis-abundant D113gene promoter in transgenic tobacco. Plant Cell Rep 27:707–717
Ma JK, Drake PM, Christou P (2003) The production of recombinant
pharmaceutical proteins in plants. Nat Rev Genet 4:794–805
Maiti IB, Shepherd RJ (1998) Isolation and expression analysis of
Peanut chlorotic streak caulimovirus (PClSV) full-length tran-
script (FLt) promoter in transgenic plants. Biochem Biophys Res
Commun 244:440–444
Mandel T, Fleming AJ, Krahenbuhl R, Kuhlemeier C (1995)
Definition of constitutive gene expression in plants: the trans-
lation initiation factor 4A gene as a model. Plant Mol Biol
29:995–1004
Mariani C, De Beuckeleer M, Truettner J, Leemans J, Goldberg RB
(1990) Induction of male sterility in plants by a chimeric
ribonuclease gene. Nature 347:737–741
Mariani C, Gossele V, De Beuckeleer M, De Block M, Goldberg RB,
De Greef W, Leemans J (1992) A chimaeric ribonuclease-
inhibitor gene restores fertility to male sterile plants. Nature
357:384–387
Marris C, Gallois P, Copley J, Kreis M (1988) The 50 flanking region
of a barley B hordein gene controls tissue and developmental
specific CAT expression in tobacco plants. Plant Mol Biol
10:359–366
Martinez A, Sparks C, Hart CA, Thompson J, Jepson I (1999)
Ecdysone agonist inducible transcription in transgenic tobacco
plants. Plant J 19:97–106
Marzabal P, Busk PK, Ludevid MD, Torrent M (1998) The bifactorial
endosperm box of gamma-zein gene: characterisation and
function of the Pb3 and GZM cis-acting elements. Plant J
16:41–52
McElroy D, Zhang W, Cao J, Wu R (1990) Isolation of an efficient
actin promoter for use in rice transformation. Plant Cell 2:
163–171
McElroy D, Blowers AD, Jenes B, Wu R (1991) Construction of
expression vectors based on the rice actin 1 (Act1) 50 region for
use in monocot transformation. Mol Gen Genet 231:150–160
Medberry SL, Lockhart BEL, Olszewski NE (1992) The Commelinayellow mottle virus promoter is a strong promoter in vascular and
reproductive tissues. Plant Cell 4:185–192
Mehlo L, Mazithulela G, Twyman RM, Boulton MI, Davies JW,
Christou P (2000) Structural analysis of transgene rearrange-
ments and effects on expression in transgenic maize plants
generated by particle bombardment. Maydica 45:277–287
Mette MF, van der Winden J, Matzke MA, Matzke AJM (1999)
Production of aberrant promoter transcripts contributes to
methylation and silencing of unlinked homologous promoters
in trans. EMBO J 18:241–248
Mette MF, Aufsatz W, van der Winden J, Matzke MA, Matzke
AJM (2000) Transcriptional silencing and promoter methyla-
tion triggered by double-stranded RNA. EMBO J 19:5194–
5201
Mitra A, Han J, Zhang ZJ, Mitra A (2009) The intergenic region of
Arabidopsis thaliana cab1 and cab2 divergent genes functions as
a bidirectional promoter. Planta 229:1015–1022
Mitsuhara I, Ugaki M, Hirochika H, Ohshima M, Murakami T, Gotoh
Y, Katayose Y, Nakamura S, Honkura R, Nishimiya S, Ueno K,
Mochizuki A, Tanimoto H, Tsugawa H, Otsuki Y, Ohashi Y
(1996) Efficient promoter cassettes for enhanced expression of
foreign genes in dicotyledonous and monocotyledonous plants.
Plant Cell Physiol 37:49–59
Mourrain P, van Blokland R, Kooter JM, Vaucheret H (2007) A
single transgene locus triggers both transcriptional and post-
transcriptional silencing through double-stranded RNA produc-
tion. Planta 225:365–379
Naqvi S, Farre G, Sanahuja G, Capell T, Zhu C, Christou P (2009a)
When more is better: multigene engineering in plants. Trends
Plant Sci 15:48–56
Naqvi S, Zhu C, Farre G, Ramessar K, Bassie L, Breitenbach J, Perez
Conesa D, Ros G, Sandmann G, Capell T, Christou P (2009b)
Transgenic multivitamin corn through biofortification of endo-
sperm with three vitamins representing three distinct metabolic
pathways. Proc Natl Acad Sci USA 106:7762–7767
Ni M, Cui D, Gelvin SB (1996) Sequence-specific interactions of
wound-inducible nuclear factors with mannopine synthase 20
promoter wound-responsive elements. Plant Mol Biol 30:
77–96
Norris SR, Meyer SE, Callis J (1993) The intron of Arabidopsisthaliana polyubiquitin genes is conserved in location and is a
quantitative determinant of chimeric gene expression. Plant Mol
Biol 21:895–906
Odell JT, Nagy F, Chua NH (1985) Identification of DNA sequences
required for activity of the Cauliflower mosaic virus 35S
promoter. Nature 313:810–812
Ohtsuki S, Levine M, Cai HN (1998) Different core promoters
possess distinct regulatory activities in the Drosophila embryo.
Genes Dev 12:547–556
Onate L, Vicente-Carbajosa J, Lara P, Dıaz I, Carbonero P (1999)
Barley BLZ2, a seed-specific bZIP protein that interacts with
BLZ1 in vivo and activates transcription from the GCN4-like
motif of B-hordein promoters in barley endosperm. J Biol Chem
274:9175–9182
Onodera Y, Suzuki A, Wu CY, Washida H, Takaiwa F (2001) A rice
functional transcriptional activator, RISBZ1, responsible for
endosperm-specific expression of storage protein genes through
GCN4 motif. J Biol Chem 276:14139–14152
Onodera Y, Haag JR, Ream T, Nunes PC, Pontes O, Pikaard CS
(2005) Plant nuclear RNA polymerase IV mediates siRNA and
DNA methylation-dependent heterochromatin formation. Cell
120:613–622
Opsahl-Sorteberg HG, Divon HH, Nielsen PS, Kalla R, Hammon-
Kosach M, Shimamoto K, Kohli A (2004) Identification of a 49-
bp fragment of the HvLTP2 promoter directing aleurone cell
specific expression. Gene 341:49–58
Padidam M, Gore M, Lu DL, Smirnova O (2003) Chemical-inducible,
ecdysone receptor-based gene expression system for plants.
Transgenic Res 12:101–109
Panikulangara TJ, Eggers-Schumacher G, Wunderlich M, Stransky H,
Schoffl F (2004) Galactinol synthase1. A novel heat shock factor
target gene responsible for heat-induced synthesis of raffinose
family oligosaccharides in Arabidopsis. Plant Physiol 136:3148–
3158
376 Plant Mol Biol (2010) 73:363–378
123

Paul W, Hodge R, Smartt S, Draper J, Scott R (1992) The isolation
and characterization of the tapetum specific Arabidopsis thalianaA9 gene. Plant Mol Biol 19:611–622
Pontier D, Yahubyan G, Vega D, Bulski A, Saez-Vasquez J, Hakimi
MA, Lerbs-Mache S, Colot V, Lagrange T (2005) Reinforce-
ment of silencing at transposons and highly repeated sequences
requires the concerted action of two distinct RNA polymerases
IV in Arabidopsis. Genes Dev 19:2030–2040
Potenza C, Aleman L, Sengupta-Gopalan C (2004) Targeting
transgene expression in research, agricultural, and environmental
applications: promoters used in plant transformation. In Vitro
Cell Dev Biol Plant 40:1–22
Qu LQ, Takaiwa F (2004) Evaluation of tissue specificity and
expression strength of rice seed component gene promoters in
transgenic rice. Plant Biotechnol J 2:113–125
Raclaru M, Gruber J, Kumar R, Sadre R, Luhs W, Zarhloul MK,
Friedt W, Frentzen M, Weier D (2006) Increase of the
tocochromanol content in transgenic Brassica napus seeds by
overexpression of key enzymes involved in prenylquinone
biosynthesis. Mol Breed 18:93–107
Raho G, Lupotto E, Hartings H, Della Torre AP, Perrotta C,
Marmiroli N (1996) Tissue-specific expression and environmen-
tal regulation of the barley Hvhspi7 gene promoter in transgenic
tobacco plants. J Exp Bot 47:1587–1594
Rai M, Datta K, Parkhi V, Tan J, Oliva N, Chawla HS, Datta SK
(2007) Variable T-DNA linkage configuration affects inheritance
of carotenogenic transgenes and carotenoid accumulation in
transgenic indica rice. Plant Cell Rep 26:1221–1231
Rance I, Norre F, Gruber V, Theisen M (2002) Combination of viral
promoter sequences to generate highly active promoters for
heterologous therapeutic protein over-expression in plants. Plant
Sci 162:833–842
Rushton PJ, Reinstadler A, Lipka V, Lippok B, Somssich IE (2002)
Synthetic plant promoters containing defined regulatory ele-
ments provide novel insights into pathogen- and wound-induced
signaling. Plant Cell 14:749–762
Sadanandom A, Piffanelli P, Knott T, Robinson C, Sharpe A, Lydiate
D, Murphy D, Fairbairn DJ (1996) Identification of a peptide
methionine sulphoxide reductase gene in an oleosin promoter
from Brassica napus. Plant J 10:235–242
Saidi Y, Finka A, Chakhporanian M, Zryd JP, Schaefer DG,
Goloubinoff P (2005) Controlled expression of recombinant
proteins in Physcomitrella patens by a conditional heat-shock
promoter: a tool for plant research and biotechnology. Plant Mol
Biol 59:697–711
Sasaki K, Yuichi O, Hiraga S, Gotoh Y, Seo S, Mitsuhara I, Ito H,
Matsui H, Ohashi Y (2007) Characterization of two rice
peroxidase promoters that respond to blast fungus-infection.
Mol Genet Genomics 278:709–722
Sawant S, Singh PK, Madanala R, Tuli R (2001) Designing of an
artificial expression cassette for the high-level expression of
transgenes in plants. Theor Appl Genet 102:635–644
Schenk PM, Remans T, Sagi L, Elliott AR, Dietzgen RG, Swennen R,
Ebert PR, Grof CPL, Manners JM (2001) Promoters for
pregenomic RNA of banana streak badnavirus are active for
transgene expression in monocot and dicot plants. Plant Mol Biol
47:399–412
Schernthaner JP, Matzke MA, Matzke AJM (1988) Endosperm-
specific activity of a zein gene promoter in transgenic tobacco
plants. EMBO J 7:1249–1255
Schmidt RJ, Ketudat M, Aukerman MJ, Hoschek G (1992) Opaque-2
is a transcriptional activator that recognizes a specific target site
in 22-kD zein genes. Plant Cell 4:689–700
Schubert D, Lechtenberg B, Forsbach A, Gils M, Bahadur S, Schmidt
R (2004) Silencing in Arabidopsis T-DNA transformants: the
predominant role of a gene-specific RNA sensing mechanism
versus position effects. Plant Cell 16:2561–2572
Schunmann PHD, Llewellyn DJ, Surin B, Boevink P, Feyter RCD,
Waterhouse PM (2003a) A suite of novel promoters and
terminators for plant biotechnology. Functional Plant Biol
30:443–452
Schunmann PHD, Surin B, Waterhouse PM (2003b) A suite of novel
promoters and terminators for plant biotechnology. II. The
pPLEX series for use in monocots. Functional Plant Biol
30:453–460
Singh A, Sahi C, Grover A (2009) Chymotrypsin protease inhibitor
gene family in rice: genomic organization and evidence for the
presence of a bidirectional promoter shared between two
chymotrypsin protease inhibitor genes. Gene 428:9–19
Skriver K, Olsen FL, Rogers JC, Mundy J (1991) Cis-acting DNA
elements responsive to gibberellin and its antagonist abscisic
acid. Proc Natl Acad Sci USA 88:7266–7270
Slater A, Scott N, Fowler M (2003) Plant Molecular Biology. Oxford
University Press, Oxford
Spencer TM, O’Brien JV, Start WG, Adams TR, Gordon-Kamm WJ,
Lemaux PG (1992) Segregation of transgenes in maize. Plant
Mol Biol 18:201–210
Stavolone L, Ragozzino A, Hohn T (2003) Characterization of
Cestrum yellow leaf curling virus: a new member of the
caulimoviridae family. J Gen Virol 84:3459–3464
Stoger E, Williams S, Keen D, Christou P (1998) Molecular
characteristics of transgenic wheat and the effect on transgene
expression. Transgenic Res 7:463–471
Sun TP, Gubler F (2004) Molecular mechanism of gibberellin
signaling in plants. Annu Rev Plant Biol 55:197–223
Sun AQ, Yi SY, Yang JY, Zhao CM, Liu J (2006) Identification and
characterization of a heat-inducible ftsH gene from tomato
(Lycopersicon esculentum Mill.). Plant Sci 170:551–562
Takaiwa F, Oono K, Kato A (1991) Analysis of the 50 flanking region
responsible for the endosperm-specific expression of a rice
glutelin chimeric gene in transgenic tobacco. Plant Mol Biol
16:49–58
Takaiwa F, Yamanouchi U, Yoshihara T, Washida H, Tanabe F, Kato
A, Yamada K (1996) Characterization of common cis-regulatory
elements responsible for the endosperm-specific expression of
members of the rice glutelin multigene family. Plant Mol Biol
30:1207–1221
Tiana L, Wub K, Hannama C, Latoszek-Greena M, Sibbalda S, Huc
M, Browna DCW, Mikic B (2005) Analysis and use of the
tobacco eIF4A–10 promoter elements for transgene expression. J
Plant Physiol 162:1355–1366
Torbert KA, Gopalraj M, Medberry SL, Olszewski NE, Somers DA
(1998) Expression of the Commelina yellow mottle viruspromoter in transgenic oat. Plant Cell Rep 17:284–287
Twyman RM (2003) Growth and development: control of gene
expression, regulation of transcription. In: Thomas B, Murphy
DJ, Murray B (eds) Encyclopedia of Applied Plant Sciences.
Elsevier Science, London, pp 558–567
Unger E, Cigan AM, Trimnell M, Xu RJ, Kendall T, Roth B,
Albertsen M (2002) A chimeric ecdysone receptor facilitates
methoxyfenozide-dependent restoration of male fertility in ms45
maize. Transgenic Res 11:455–465
Urwin PE, Moller SG, Lilley CJ, McPherson MJ, Atkinson HJ (1997)
Continual green fluorescent protein monitoring of Cauliflowermosaic virus 35S promoter activity in nematode-induced feeding
cells in Arabidopsis thaliana. Mol Plant Microbe Interact
10:394–400
Vain P, Finer KR, Engler DE, Pratt RC, Finer JJ (1996) Intron-
mediated enhancement of gene expression in maize (Zea maysL.) and bluegrass (Poa pratensis L.). Plant Cell Rep 15:489–494
Plant Mol Biol (2010) 73:363–378 377
123

Valentin HE, Mitsky TA (2002) TYRA genes and uses thereof. Patent
Application WO 02/089561 A1
Veena, Reddy VS, Sopory SK (1999) Glyoxalase I from Brassicajuncea: molecular cloning, regulation and its over-expression
confer tolerance in transgenic tobacco under stress. Plant J
17:385–395
Verdaguer B, de Kochko A, Beachy RN, Fauquet C (1996) Isolation
and expression in transgenic tobacco and rice plants, of the
Cassava vein mosaic virus (CsVMV) promoter. Plant Mol Biol
31:1129–1139
Vicente-Carbajosa J, Onate L, Lara P, Diaz I, Carbonero P (1998)
Barley BLZ1: a bZIP transcriptional activator that interacts with
endosperm-specific gene promoters. Plant J 13:629–640
Wierzbicki AT, Haag JR, Pikaard CS (2008) Noncoding transcription
by RNA polymerase Pol IVb/Pol V mediates transcriptional
silencing of overlapping and adjacent genes. Cell 135:635–648
Wobus U, Borisjuk L, Panitz R, Manteuffel R, Baumlein H, Wohlfart
T, Heim U, Weber H, Misera S, Weschke W (1995) Control of
seed storage protein gene expression: new aspects on an old
story. J Plant Physiol 145:592–599
Woodger F, Millar A, Murray F, Jacobsen J, Gubler F (2003) The role
of GAMYB transcription factors in GA-regulated gene expres-
sion. J Plant Growth Regul 22:176–184
Wu CY, Suzuki A, Washida H, Takaiwa F (1998) The GCN4 motif in
a rice glutelin gene is essential for endosperm-specific gene
expression and is activated by Opaque-2 in transgenic rice
plants. Plant J 14:673–683
Wu H, Michler CH, LaRussa L, Davis JM (1999) The pine Pschi4promoter directs wound-induced transcription. Plant Sci
142:199–207
Xie M, He Y, Gan S (2001) Bidirectionalization of polar promoters in
plants. Nature Biotechnol 19:677–679
Xie Z, Johansen L, Gustafson A, Kasschau K, Lellis A, Zilberman D,
Jacobsen S, Carrington J (2004) Genetic and functional diver-
sification of small RNA pathways in plants. PLoS Biol 2:E104
Xu D, McElroy D, Thornburg RW, Wu R (1993) Systemic induction
of a potato pin2 promoter by wounding, methyl jasmonate, and
abscisic acid in transgenic rice plants. Plant Mol Biol 22:
573–588
Yang G, Nakamura H, Ichikawa H, Kitano H, Komatsu S (2006)
OsBLE3, a brassinolide-enhanced gene, is involved in the growth
of rice. Phytochemistry 67:1442–1454
Yevtushenko DP, Sidorov VA, Romero R, Kay WW, Misra S (2004)
Wound-inducible promoter from poplar is responsive to fungal
infection in transgenic potato. Plant Sci 167:715–724
Zhang W, McElroy D, Wu R (1991) Analysis of rice Act1 50 region
activity in transgenic rice plants. Plant Cell 3:1155–1165
Zhang C, Gai Y, Wang W, Zhu Y, Chen X, Jiang X (2008)
Construction and analysis of a plant transformation binary vector
pBDGG harboring a bi-directional promoter fusing dual visible
reporter genes. J Genet Genomics 35:245–249
Zheng Z, Kawagoe Y, Xiao S, Li Z, Okita TW, Hau TL, Lin A, Murai
N (1993) 50 Distal and proximal cis-acting regulation elements
are required for developmental control of a rice seed storage
protein glutelin gene. Plant J 4:357–366
Zheng X, Deng W, Luo K, Duan H, Chen Y, McAvoy R, Song S, Pei
Y, Li Y (2007) The Cauliflower mosaic virus (CaMV) 35S
promoter sequence alters the level and patterns of activity of
adjacent tissue- and organ-specific gene promoters. Plant Cell
Rep 26:1195–1203
Zhu QH, Ramm K, Eamens AL, Dennis ES, Upadhyaya NM (2006)
Transgene structures suggest that multiple mechanisms are
involved in T-DNA integration in plants. Plant Sci 171:308–322
Zhu C, Naqvi S, Gomez-Galera S, Pelacho AM, Capell T, Christou P
(2007) Transgenic strategies for the nutritional enhancement of
plants. Trends Plant Sci 12:548–555
Zhu C, Naqvi S, Breitenbach J, Sandmann G, Christou P, Capell T
(2008) Combinatorial genetic transformation generates a library
of metabolic phenotypes for the carotenoid pathway in maize.
Proc Natl Acad Sci USA 105:18232–18237
Zilberman D, Cao X, Jacobsen SE (2003) ARGONAUTE4 control of
locus-specific siRNA accumulation and DNA and histone
methylation. Science 299:716–719
Zourelidou M, de Torres-Zabala M, Smith C, Bevan MW (2002)
Storekeeper defines a new class of plant-specific DNA-binding
proteins and is a putative regulator of patatin expression. Plant J
30:489–497
378 Plant Mol Biol (2010) 73:363–378
123
Copyright © 2022 FDOKUMEN

