







Bahasa
Halaman
Hukum


Nutrition, Metabolism & Cardiovascular Diseases (2009) 19, 20e26
ava i lab le a t www.sc iencedi rec t .com
j ourna l homepage : www.e lsev ier . com/ loca te /nmcd
Omega 3 has a beneficial effect on ischemia/reperfusion injury, but cannot reverse theeffect of stressful forced exercise
Daniele Mancardi*, Francesca Tullio, Antonio Crisafulli,Raffaella Rastaldo, Anna Folino, Claudia Penna, Pasquale Pagliaro
Dipartimento di Scienze Cliniche e Biologiche, Universita di Torino, Orbassano (TO), Italy
Received 16 August 2007; received in revised form 5 November 2007; accepted 17 January 2008
KEYWORDSu3;Forced exercise;Ischemia/reperfusion
* Corresponding author. Dipartimenlogiche, Universita di Torino, Ospeda10, 10043 Orbassano (TO), Italy. Tel.11 903 8639.
E-mail address: daniele.mancardi@
0939-4753/$ - see front matter ª 200doi:10.1016/j.numecd.2008.01.004
Abstract Background and aim: The beneficial effects of exercise in reducing the incidence ofcardiovascular diseases are well known. Several studies have demonstrated that forced exer-cise (FE) could activate a stress response similar to a restrain stress. Previous studies suggestthat heart protection to ischemic events would be improved by an omega 3 free fatty acid (u3-FFA)-enriched diet. Here, we investigate the impact of stressful FE and an omega 3-FFA-en-riched diet on cardiac tolerance to ischemic events over one month.Methods and results: Twenty-four Wistar rats were randomly assigned to one of the followingprotocols: 1) Sedentary (SED) animals who were regularly fed; 2) sedentary animals who weregiven 1 ml/day of fish oil for one month; 3) FE þ u3-FFA rats who were given 1 ml/day of fish oiland forced to run on a motorized wheel for 30 min every day, both for one month; and 4) FEanimals were forced to exercise as group 3 and fed with a regular diet. At the end of the treat-ments an isolated heart preparation was performed.
After a 30 min global ischemic event and 2 h reperfusion, hearts of sedentary-u3 animalsrecovered about 37% of left ventricular developed pressure, whereas FE, u3 þ FE and CTRL-SED animals recovered only about 15%, 5% and 8% respectively. Similarly, heart infarct sizewas significantly lower in sedentary-u3 animals compared to animals in the three other groups.Conclusions: Results indicate that one month of treatment with an u3-FFA-enriched diet im-proves cardioprotection upon ischemic events, whereas FE leads to a reduced heart toleranceto ischemic events, which cannot be reversed by an u3-FFA diet.ª 2008 Elsevier B.V. All rights reserved.
to di Scienze Cliniche e Bio-le S. Luigi, Regione Gonzole,: þ39 11 670 5430; fax: þ39
unito.it (D. Mancardi).
8 Elsevier B.V. All rights reserved
Introduction
Data supporting the inverse correlation of fish or long-chainomega-3 fatty acid (u3-FFA) supplement consumption and
.

Omega 3 has a beneficial effect on ischemia/reperfusion injury 21
coronary heart disease are inconclusive and may beconfounded by other lifestyle factors. Here we investigatedthe effects of stressful forced exercise (FE) and u3-FFA dietimplementation on cardiac tolerance to ischemia/reperfu-sion challenge.
Repeated bouts of exercise produce several physiologicadaptations that allow an organism to perform a greateramount of work with improved efficacy. These changes areoften referred to as training adaptations and are consid-ered beneficial for the health of an individual [1].
Beneficial effects of physical exercise on cardiovascularfunction are well established. However, several studieshave demonstrated that forced exercise could activatea stress response similar to a restrain stress [2e7] whichcan reduce antioxidant defense of the organs [5].
As a matter of fact, forced running is a common methodof training used by experimenters investigating physiologicadaptations produced by exercise [8,9]. Advantages of thisspecific mode of training include, for example, the repeat-ability of training volume since the researchers choose thetime, duration and intensity of the running. This type oftraining, however, forces the animals to run and it couldchronically activate a stress response.
Although it is well established that voluntary physicalexercise is beneficial both before and after myocardialinfarct how FE affects the outcomes of a subsequentepisode of ischemia/reperfusion is not clear.
The promise of a nutritional approach in considering thetreatment of disease is beginning to be recognized [10,11].For example, u3-FFA-rich diets are perceived to be gener-ally beneficial from a health standpoint, particularly in thecontext of the cardiovascular system [12,13]. However,retrospective studies associated benefits with a healthierdietary pattern rather than with a specific effect of anu3-FFA rich diet [14].
So far there has been no experimental evaluation on howstress-induced physio-pathologic changes can be altered bydifferent diets.
Therefore, the purpose of the present study was three-fold: 1) to ascertain whether an u3-FFA-enriched diet mayhave beneficial effects on I/R injuries; 2) to determinewhether the stress of forced exercise would result in alteredinjury to the myocardium after ischemia/reperfusion (I/R);and 3) to determine whether an u3-FFA diet may interferewith the effects of forced exercise upon I/R injury.
I/R injury was assessed using a Langendorff isolated ratheart preparation. This type of assay is reliably used todetermine acute I/R injury in the absence of interferencedue to neuro-hormonal and variable collateral flow, thatcan be confounding elements in an in vivo model. Yet theisolated heart model excludes the contribution to injurydue to leukocyte activation, which can be of importancein determining infarct size in the longer period.
Methods
Animals
Male Wistar rats (300e320 g, 6 week old) were purchasedfrom Harlan. Rats were housed in a controlled temperatureroom (22 �C) with a 12:12-h light-dark cycle (06.00e18.00
lights on) and ad libitum access to laboratory rat chowand water was provided. To reduce the stress associatedwith shipping, rats were housed in standard Plexiglas cagesfor 10 days before onset of any running activity. Care anduse of the animals were in accordance with protocolsapproved by the University of Torino and conformed tothe Italian law.
The animals were randomly assigned to one of thetreatment groups described below.
Rats were either forced to run in a motorized wheel(n Z 12) or remained sedentary (n Z 12).
Six rats of each group [6 forced exercise (FE) and 6sedentary (SED) rats] were fed with regular rat chow diet. Inthe remaining six rats of each group (6 FE þ u3-FFA rats and6 SED þ u3-FFA rats) the regular diet was supplementedwith 1 ml per day of fish oil (Fish oil from menhaden, Sigma)for one month. Fish oil from menhaden is reported to containapproximately 25% omega-3 (octadecatetraenoic, eicosa-pentaenoic and docosahexaenoic) fatty acids as triglycer-ides. Diet supplementation was carried out for five daysa week at 12.00 o’clock with an appropriate oesophagealcatheter, connected to a syringe; oil was therefore injectedinto the oesophagus of the animal at the desired quantity.
Exercise training
FE and FE þ u3-FFA rats were housed in clear plastic cagesand followed a moderate exercise training protocol shownto induce cardiovascular fitness in rats [15]. The 4-week ex-ercise training protocol consisted of running on a motorizedwheel (wheel radius 25 cm) five times per week with an in-cremental velocity of the wheel (from 5 to 12 laps per min)and running duration (from 10 to 30 min) to accommodateincreased fitness (mean velocity 15e30 cm/s circa). Thehighest level of activity [12 laps/min (i.e. 30 cm/s) for30 min] included a 5-min warm-up period at 5 laps/min,without a final period of warm down. Increase in velocityand/or duration were performed when the rats were ableto maintain the exercise intensity for 2e3 consecutive ses-sions. If the pace of the rat slowed below wheel rate theanimals were pushed by the operator to reassume the rightposition to run (physical prodding: the operator used a stickto force the rats to run).
No shocks were administered during the training session.In fact, Dunn and co-workers reported that treadmill-trained rats with no footshock showed changes in centralnervous system noradrenergic metabolism that are indica-tive of chronic stress [4]. The SED and SED þ u3-FFA ratswere housed in clear plastic cages and were handled iden-tically to the FE rats. The SED þ u3-FFA rats were fed eachday at the same time of day as FE þ u3-FFA rats.
Isolated heart perfusion
After 38 days of housing in the animal facility (i.e. two daysafter the last treatment-day for animals of GroupsSED þ u3-FFA, FE and FE þ u3-FFA rats), each animal wastreated with heparin (2500 U, i.m.). Then, 10 min later, an-imals were anaesthetized with urethane (1 g/kg, i.p.), thechest was opened, the heart was rapidly excised and placedin ice-cold buffer solution and weighed. Isolated rat hearts

22 D. Mancardi et al.
were attached to the perfusion apparatus and retrogradelyperfused with oxygenated KrebseHenseleit buffer (127 mMNaCl, 17.7 mM NaHCO3, 5.1 mM KCl, 1.5 mM CaCl2, 1.26 mMMgCl2, 11 mM D-glucose and gassed with 95% O2 and 5% CO2).The time required between excision and restoration of per-fusion was less than 30 s. The hearts were instrumented aspreviously described and pump-perfused at constant flow[16,17]. The constant flow was maintained to obtain a typi-cal coronary perfusion pressure of 85 mmHg. A small hole inthe left ventricular wall allowed drainage of the thebesianflow, and a polyvinyl-chloride balloon was placed into theleft ventricle and connected to an electromanometer forrecording of left ventricular pressure (LVP). The heartswere electrically paced at 280e300 bpm and kept in a tem-perature-controlled chamber (37 �C). Coronary perfusionpressure (CPP) and coronary flow (CF) were monitoredwith a second electromanometer and an electromagneticflow-probe, respectively, both placed along the perfusionline. Coronary flow, CPP and LVP were recorded and ana-lysed using a Lab-View software (National Instruments,USA), which also allowed quantification of the maximumrate of increase of LVP during systole (dP/dtmax).
Experimental protocols
Each heart was allowed to stabilize for 20 min at whichtime baseline parameters were recorded. Typically, CF(9 � 2 ml/min per g wet weight) was adjusted to obtainthe desired CPP (85 mmHg) within the first 5 min and keptconstant thereon. During the stabilization period the ven-tricular volume (VV) was adjusted, and kept constantthroughout the experiment, to obtain an end diastolic leftventricular pressure of 5 mmHg [16].
All hearts were then subjected to 30 min of global,normothermic, no-flow ischemia followed by 120 min ofreperfusion (I/R). Pacing was discontinued at the beginningof the ischemic period and restarted after the third minuteof reperfusion [16,18].
Assessment of ventricular function
Changes in left ventricular end-diastolic pressure (LVEDP),developed LVP (dLVP), and dP/dtmax values induced by theI/R protocol were continuously monitored. The differencebetween LVEDP before the end-reperfusion (mean valueof the last minute of reperfusion) and during pre-ischemicconditions (mean value of the last minute of washout/buffer only) was used as an index of the extent of contrac-ture development. Maximal recovery of developed LVP anddP/dtmax during reperfusion was also compared with re-spective pre-ischemic values.
Assessment of myocardial injury
Infarct areas were assessed by an independent observer atthe end of the experiment. Directly after reperfusion,hearts were rapidly removed from the perfusion apparatusand sliced across the long axis of the left ventricle, fromapex to base, into 2e3 mm thick transverse sections andthen incubated in 0.1% solution of nitro blue tetrazolium(Sigma Chemical, St. Louis, MO) in phosphate buffer (pH
7.4) at 37 �C for 15 min [16,19]. In the heart subjected toa global ischemia, the entire ventricle is at risk of infarctionand therefore measurement of collateral flow is notrequired. Global ischemia resulted in multiple blue-tetrazolium positive areas. We were careful to separatethe areas of viable and necrotic tissue using a surgical blade.The tissues were separated and then weighed by an indepen-dent observer who did not know the origin of the hearts. Thevolumes of the infarcted zone and the area at risk were thenobtained and infarct size expressed as a percentage of leftventricular volume for each heart [16,19]. In fact, thoughin this model the whole heart underwent ischemia, onlythe LV had a fixed volume and pre-load; therefore only theLV mass was considered as the risk area.
LDH release during reperfusion was determined as pre-viously described [16].
Statistical analysis
All values are presented as means � S.E.M. To compare thecardiac responses of different groups we also report data aspercent changes from baseline values. Two sets of compar-isons were made for each of the considered hemodynamicvariables: one among baseline data and the other compar-ing maximal effects in response to I/R. Moreover, areasunder the curves were calculated and comparisons wereperformed using two-way analysis of variance (ANOVA) forrepeated measures (factors: group and condition). One-way ANOVA was used to compare, between subsets, thecardiac weight, LV weight and the cardiac to body weightratio. For these parameters post hoc comparison betweengroups at an individual time point was performed usingthe Student’s t-test for unpaired data. Contrasts betweenconditions in the same subset were analysed using Stu-dent’s t-test for paired data. Student’s t-test was per-formed with Bonferroni correction. Significance wasaccepted at a P level of <0.05.
Results
Final cardiac weight (1.33 � 0.01 g; n Z 24), LV weight(0.89 � 0.006 g; n Z 24) and the cardiac to body weight ra-tio (0.003 � 0.00003; n Z 24) were equivalent among thefour Groups (n Z 6 for each group). Neither initial and finalbody weight nor final heart weight to body weight ratiowere statistically different between groups.
Pre-ischemic function
Baseline cardiac function after stabilization and prior toischemia observed in the four groups of hearts are reportedin Table 1. No significant differences were observed amonggroups.
Perfusion pressure during ischemia-reperfusion
The area under the curve of PP for sedentary-63 hearts wassignificantly smaller than that of FE, 63 þ FE and SEDanimals (Fig. 1). These data suggest that vasoconstriction de-velopment during I/R is less severe in sedentary-63 hearts.
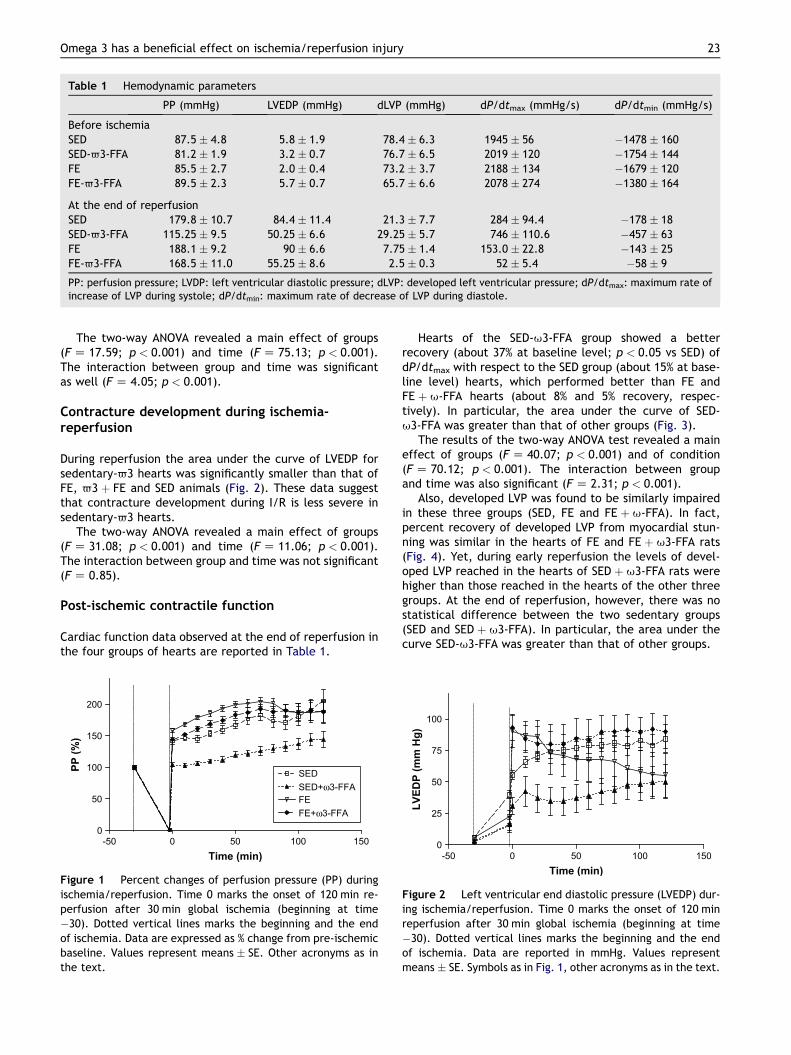
Table 1 Hemodynamic parameters
PP (mmHg) LVEDP (mmHg) dLVP (mmHg) dP/dtmax (mmHg/s) dP/dtmin (mmHg/s)
Before ischemiaSED 87.5 � 4.8 5.8 � 1.9 78.4 � 6.3 1945 � 56 �1478 � 160SED-63-FFA 81.2 � 1.9 3.2 � 0.7 76.7 � 6.5 2019 � 120 �1754 � 144FE 85.5 � 2.7 2.0 � 0.4 73.2 � 3.7 2188 � 134 �1679 � 120FE-63-FFA 89.5 � 2.3 5.7 � 0.7 65.7 � 6.6 2078 � 274 �1380 � 164
At the end of reperfusionSED 179.8 � 10.7 84.4 � 11.4 21.3 � 7.7 284 � 94.4 �178 � 18SED-63-FFA 115.25 � 9.5 50.25 � 6.6 29.25 � 5.7 746 � 110.6 �457 � 63FE 188.1 � 9.2 90 � 6.6 7.75 � 1.4 153.0 � 22.8 �143 � 25FE-63-FFA 168.5 � 11.0 55.25 � 8.6 2.5 � 0.3 52 � 5.4 �58 � 9
PP: perfusion pressure; LVDP: left ventricular diastolic pressure; dLVP: developed left ventricular pressure; dP/dtmax: maximum rate ofincrease of LVP during systole; dP/dtmin: maximum rate of decrease of LVP during diastole.
Omega 3 has a beneficial effect on ischemia/reperfusion injury 23
The two-way ANOVA revealed a main effect of groups(F Z 17.59; p < 0.001) and time (F Z 75.13; p < 0.001).The interaction between group and time was significantas well (F Z 4.05; p < 0.001).
Contracture development during ischemia-reperfusion
During reperfusion the area under the curve of LVEDP forsedentary-63 hearts was significantly smaller than that ofFE, 63 þ FE and SED animals (Fig. 2). These data suggestthat contracture development during I/R is less severe insedentary-63 hearts.
The two-way ANOVA revealed a main effect of groups(F Z 31.08; p < 0.001) and time (F Z 11.06; p < 0.001).The interaction between group and time was not significant(F Z 0.85).
Post-ischemic contractile function
Cardiac function data observed at the end of reperfusion inthe four groups of hearts are reported in Table 1.
-50 0 50 100 1500
50
100
150
200
SED+ω3-FFASED
FEFE+ω3-FFA
Time (min)
PP
(%
)
Figure 1 Percent changes of perfusion pressure (PP) duringischemia/reperfusion. Time 0 marks the onset of 120 min re-perfusion after 30 min global ischemia (beginning at time�30). Dotted vertical lines marks the beginning and the endof ischemia. Data are expressed as % change from pre-ischemicbaseline. Values represent means � SE. Other acronyms as inthe text.
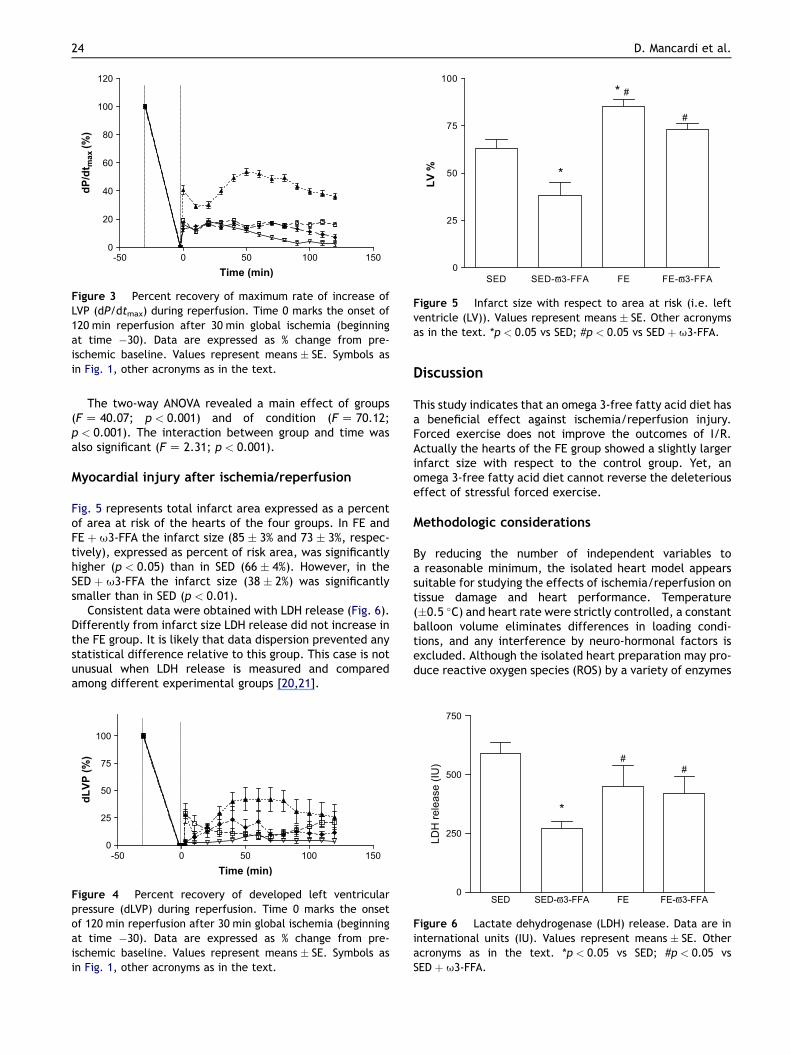
Hearts of the SED-u3-FFA group showed a betterrecovery (about 37% at baseline level; p < 0.05 vs SED) ofdP/dtmax with respect to the SED group (about 15% at base-line level) hearts, which performed better than FE andFE þ u-FFA hearts (about 8% and 5% recovery, respec-tively). In particular, the area under the curve of SED-u3-FFA was greater than that of other groups (Fig. 3).
The results of the two-way ANOVA test revealed a maineffect of groups (F Z 40.07; p < 0.001) and of condition(F Z 70.12; p < 0.001). The interaction between groupand time was also significant (F Z 2.31; p < 0.001).
Also, developed LVP was found to be similarly impairedin these three groups (SED, FE and FE þ u-FFA). In fact,percent recovery of developed LVP from myocardial stun-ning was similar in the hearts of FE and FE þ u3-FFA rats(Fig. 4). Yet, during early reperfusion the levels of devel-oped LVP reached in the hearts of SED þ u3-FFA rats werehigher than those reached in the hearts of the other threegroups. At the end of reperfusion, however, there was nostatistical difference between the two sedentary groups(SED and SED þ u3-FFA). In particular, the area under thecurve SED-u3-FFA was greater than that of other groups.
-50 0 50 100 1500
25
50
75
100
Time (min)
LV
ED
P (m
m H
g)
Figure 2 Left ventricular end diastolic pressure (LVEDP) dur-ing ischemia/reperfusion. Time 0 marks the onset of 120 minreperfusion after 30 min global ischemia (beginning at time�30). Dotted vertical lines marks the beginning and the endof ischemia. Data are reported in mmHg. Values representmeans � SE. Symbols as in Fig. 1, other acronyms as in the text.
-50 0 50 100 1500
20
40
60
80
100
120
Time (min)
dP
/d
tm
ax (%
)
Figure 3 Percent recovery of maximum rate of increase ofLVP (dP/dtmax) during reperfusion. Time 0 marks the onset of120 min reperfusion after 30 min global ischemia (beginningat time �30). Data are expressed as % change from pre-ischemic baseline. Values represent means � SE. Symbols asin Fig. 1, other acronyms as in the text.
0
25
SED FESED-ϖ3-FFA FE-ϖ3-FFA
50
75
100
LV
%
*
* #
#
Figure 5 Infarct size with respect to area at risk (i.e. leftventricle (LV)). Values represent means � SE. Other acronymsas in the text. *p < 0.05 vs SED; #p < 0.05 vs SED þ u3-FFA.
24 D. Mancardi et al.
The two-way ANOVA revealed a main effect of groups(F Z 40.07; p < 0.001) and of condition (F Z 70.12;p < 0.001). The interaction between group and time wasalso significant (F Z 2.31; p < 0.001).
Myocardial injury after ischemia/reperfusion
Fig. 5 represents total infarct area expressed as a percentof area at risk of the hearts of the four groups. In FE andFE þ u3-FFA the infarct size (85 � 3% and 73 � 3%, respec-tively), expressed as percent of risk area, was significantlyhigher (p < 0.05) than in SED (66 � 4%). However, in theSED þ u3-FFA the infarct size (38 � 2%) was significantlysmaller than in SED (p < 0.01).
Consistent data were obtained with LDH release (Fig. 6).Differently from infarct size LDH release did not increase inthe FE group. It is likely that data dispersion prevented anystatistical difference relative to this group. This case is notunusual when LDH release is measured and comparedamong different experimental groups [20,21].
-50 0 50 100 1500
25
50
75
100
Time (min)
dL
VP
(%
)
Figure 4 Percent recovery of developed left ventricularpressure (dLVP) during reperfusion. Time 0 marks the onsetof 120 min reperfusion after 30 min global ischemia (beginningat time �30). Data are expressed as % change from pre-ischemic baseline. Values represent means � SE. Symbols asin Fig. 1, other acronyms as in the text.
Discussion
This study indicates that an omega 3-free fatty acid diet hasa beneficial effect against ischemia/reperfusion injury.Forced exercise does not improve the outcomes of I/R.Actually the hearts of the FE group showed a slightly largerinfarct size with respect to the control group. Yet, anomega 3-free fatty acid diet cannot reverse the deleteriouseffect of stressful forced exercise.
Methodologic considerations
By reducing the number of independent variables toa reasonable minimum, the isolated heart model appearssuitable for studying the effects of ischemia/reperfusion ontissue damage and heart performance. Temperature(�0.5 �C) and heart rate were strictly controlled, a constantballoon volume eliminates differences in loading condi-tions, and any interference by neuro-hormonal factors isexcluded. Although the isolated heart preparation may pro-duce reactive oxygen species (ROS) by a variety of enzymes
0
250
500
750
LDH
rele
ase
(IU)
*
##
SED FESED-ϖ3-FFA FE-ϖ3-FFA
Figure 6 Lactate dehydrogenase (LDH) release. Data are ininternational units (IU). Values represent means � SE. Otheracronyms as in the text. *p < 0.05 vs SED; #p < 0.05 vsSED þ u3-FFA.

Omega 3 has a beneficial effect on ischemia/reperfusion injury 25
in endothelial cells and cardiomyocytes, in this model thereare no leukocytes which produce a great amount of ROS inthe I/R scenario. This can be an advantage because it al-lows a separation of damage due to an intrinsic mechanismfrom those due to leukocytes, but it can also be a disadvan-tage because the extension of damage may be differentfrom that inducible in in vivo conditions.
Therefore, the isolated heart is an experimental‘‘clean’’ condition, in which all types of cells and matrixelements with their interactions are represented, whileexternal ‘‘disturbing’’ events (i.e. leukocytes) are excludedor, if necessary, included under a well-defined experimen-tal control.
Implication of the findings
This study confirms many others that have found beneficialcardiovascular effects of an u3-FFA diet [13]. The benefi-cial effects of an u3-FFA enriched diet are evidenced bya reduced contracture development, an improved heartcontractile recovery and a reduced extension of infarctsize after I/R, with respect to a control standard diet.
We can argue that this protective effect may be due toa modulation of global gene expression profile in cardio-myocytes, as recently reported by Bordoni et al. [22]. Infact, these authors reported that many genes related tolipid transport and metabolism are up-regulated, whilemany genes related to inflammation, including thoserelated to ROS generation in cardiomyocytes, are down-regulated in rat cardiomyocytes exposed to fatty acids.
ROS are involved in both cell membrane damage andcontractile dysfunction. A reduction in cell death and animprovement of contractile recovery of viable cells may berelated to a reduced ROS generation. Moreover, contrac-ture during I/R may be related to damage induced in thecontractile apparatus due to ROS and Ca2þ overload.Reduced contracture may be a better index of cardiopro-tection than improved contractility after I/R [23].
Despite the beneficial effect in unstressed animals,u3-FFA are unable to prevent the deleterious effect ofstressors. It can be argued that among the reasons fornegative/contradictory results with diet intervention maybe included a stressful life. This is an important issue and itis in line with the fact that there is wide variation in theability of nutritional factors to modulate the expression ofdetrimental or protective proteins at an individual level. Itis thus mandatory to identify ‘good responders’ and ‘poorresponders’ to diet therapy. The present study representsa small step-forward in this direction.
It is well known that chronic exposure to psychologicand/or physical stress can also result in maladaptiveconsequences of the stress response, such as suppressionof many aspects of the immune response and an increasedincidence of disease [24]. Psychologic challenges that arecapable of activation of the stress response often shareseveral common features, including ‘‘loss of control’’ andexposure to adverse events. In addition to the physicalstress of exercise, forced running would seem likely toproduce chronic mental stress because the animals areboth forced to run (loss of control) and often are aversivelymotivated (physical prodding, footshock and loud sounds).
The demands of running at the intensity levels commonlyused [i.e., 70e90% maximal O2 consumption (VO2 max)]are known to produce short-term neural and endocrineresponses indicative of physical stress or challenge [25].In addition, Yancey and Overton [2] and Dunn et al. [4]found significant increases in mean arterial pressure, heartrate and mesenteric blood flow at the outset of forcedtreadmill, but not voluntary running. Therefore, psycho-logic factors associated with physical activity may also con-tribute to the stress of activity that is measured in exercisestudies [26].
Forced exercise has been seen to induce maladaptivechanges in both brain and heart tissues [2e7]. In particular,the levels of 8-hydroxydeoxyguanosine, indices of oxidativedamages, are about doubled in the hearts of rats forced torun with respect to voluntary running rats. These observa-tions may explain why we observed a worsening and notan improvement of I/R outcomes in the hearts of ratsforced to run.
In conclusion, we show that an u3-FFA enriched diet canreduce the I/R injury in unstressed animals. Yet theprotocol of forced exercise we used results in a worseningof I/R injury, which cannot be reversed by an u3-FFAenriched diet.
Acknowledgements
We thank ‘‘Compagnia di San Paolo’’, Torino, the RegionePiemonte, the Istituto Nazionale per la Ricerca Cardiovas-colare (INRC) and the Italian Ministry of University andResearch (MIUR) for their financial contributions.
References
[1] Brooks GA, Fahey TD, White TP. Exercise physiology: humanbioenergetics and its applications. Mountain View, CA:Mayfield Publishing; 1995.
[2] Yancey SL, Overton JM. Cardiovascular responses to voluntaryand treadmill exercise in rats. J Appl Physiol 1993;75:1334e40.
[3] Pedersen BK, Bruunsgaard H, Klokker M, Kappel M,MacLean DA, Nielsen HB, et al. Exercise-induced immunomo-dulationdpossible roles of neuroendocrine and metabolicfactors. Int J Sports Med 1997;18(Suppl. 1):S2e7.
[4] Dunn AL, Reigle TG, Youngstedt SD, Armstrong RB,Dishman RK. Brain norepinephrine and metabolites aftertreadmill training and wheel running in rats. Med Sci SportsExerc 1996;28:204e9.
[5] Asami S, Hirano T, Yamaguchi R, Tsurudome Y, Itoh H, Kasai H.Effects of forced and spontaneous exercise on 8-hydroxydeox-yguanosine levels in rat organs. Biochem Biophys Res Commun1998;243:678e82.
[6] Koller A, Mertelseder S, Moser H. Is exercise-induced myocar-dial injury self-abating? [see comment]. Med Sci Sports Exerc2001;33:850e1.
[7] Scopel D, Fochesatto C, Cimarosti H, Rabbo M, Bello-Klein A,Salbego C, et al. Exercise intensity influences cell injury inrat hippocampal slices exposed to oxygen and glucose depri-vation. Brain Res Bull 2006;71:155e9.
[8] Apple FS, Billadello JJ. Expression of creatine kinase M and BmRNAs in treadmill trained rat skeletal muscle. Life Sci 1994;55:585e92.
[9] Ong JM, Simsolo RB, Saghizadeh M, Goers JW, Kern PA. Effectsof exercise training and feeding on lipoprotein lipase gene

26 D. Mancardi et al.
expression in adipose tissue, heart, and skeletal muscle of therat. Metabolism 1995;44:1596e605.
[10] Archer MC, Clarkson TW, Strain JJ. Genetic aspects ofnutrition and toxicology: report of a workshop. J Am CollNutr 2001;20:119e28.
[11] Paoloni-Giacobino A, Grimble R, Pichard C. Genetics andnutrition. Clin Nutr 2003;22:429e35.
[12] Nair SS, Leitch JW, Falconer J, Garg ML. Prevention of cardiacarrhythmia by dietary (n-3) polyunsaturated fatty acids andtheir mechanism of action. J Nutr 1997;127:383e93.
[13] Billman GE, Kang JX, Leaf A. Prevention of sudden cardiacdeath by dietary pure omega-3 polyunsaturated fatty acidsin dogs. Circulation 1999;99:2452e7.
[14] Cundiff DK, Lanou AJ, Nigg CR. Relation of omega-3 Fatty Acidintake to other dietary factors known to reduce coronaryheart disease risk. Am J Cardiol 2007;99:1230e3.
[15] Mazzeo RS, Brooks GA, Horvath SM. Effects of age on meta-bolic responses to endurance training in rats. J Appl Physiol1984;57:1369e74.
[16] Pagliaro P, Mancardi D, Rastaldo R, Penna C, Gattullo D,Miranda KM, et al. Nitroxyl affords thiol-sensitive myocardialprotective effects akin to early preconditioning. Free RadicBiol Med 2003;34:33e43.
[17] Rastaldo R, Paolocci N, Chiribiri A, Penna C, Gattullo D,Pagliaro P. Cytochrome P-450 metabolite of arachidonic acidmediates bradykinin-induced negative inotropic effect. Am JPhysiol Heart Circ Physiol 2001;280:H2823e32.
[18] Penna C, Alloatti G, Cappello S, Gattullo D, Berta G,Mognetti B, et al. Platelet-activating factor induces cardio-protection in isolated rat heart akin to ischemic precondi-tioning: role of phosphoinositide 3-kinase and proteinkinase C activation. Am J Physiol Heart Circ Physiol 2005;288:H2512e20.
[19] Ma XL, Gao F, Liu GL, Lopez BL, Christopher TA, Fukuto JM,et al. Opposite effects of nitric oxide and nitroxyl on postis-chemic myocardial injury. Proc Natl Acad Sci U S A 1999;96:14617e22.
[20] Roe CR, Cobb FR, Starmer CF. The relationship betweenenzymatic and histologic estimates of the extent of myocar-dial infarction in conscious dogs with permanent coronaryocclusion. Circulation 1977;55:438e49.
[21] Cappello S, Angelone T, Tota B, Pagliaro P, Penna C,Rastaldo R, et al. Human recombinant chromograninA-derived vasostatin-1 mimics preconditioning via an adenosi-ne/nitric oxide signaling mechanism. Am J Physiol e HeartCirc Physiol 2007;293:H719e27.
[22] Bordoni A, Astolfi A, Morandi L, Pession A, Danesi F, DiNunzio M, et al. N-3 PUFAs modulate global gene expressionprofile in cultured rat cardiomyocytes. Implications in cardiachypertrophy and heart failure. FEBS Lett 2007;581:923e9.
[23] Gelpi RJ, Morales C, Cohen MV, Downey JM. Xanthine oxidasecontributes to preconditioning’s preservation of left ventricu-lar developed pressure in isolated rat heart: developedpressure may not be an appropriate end-point for studies ofpreconditioning. Basic Res Cardiol 2002;97:40e6.
[24] Kiecolt-Glaser JK, Dura JR, Speicher CE, Trask OJ,Glaser R. Spousal caregivers of dementia victims: longitudi-nal changes in immunity and health. Psychosom Med 1991;53:345e62.
[25] Pederson BK, Bruunsgaard H, Klokker M, MacLean HB,Rohde T, Ullum H, et al. Exercise-induced immunomodula-tion-possible roles of neuroendocrine and metabolic factors.Int J Sports Med 1997;18:52e7.
[26] Hoffman-Goetz L, Pedersen BK. Exercise and the immunesystem: a model of the stress response? Immunol Today1994;15:382e7.
Top Related
Copyright © 2022 FDOKUMEN

