







Bahasa
Halaman
Hukum


Observational Learning of New Movement Sequences IsReflected in Fronto-Parietal CoherenceJurjen van der Helden1,2*., Hein T. van Schie3., Christiaan Rombouts2
1 Donders Institute for Brain, Cognition and Behaviour, Radboud University, Nijmegen, The Netherlands, 2 Department of Cognitive Psychology and Ergonomics,
University of Twente, Enschede, The Netherlands, 3 Behavioural Science Institute, Radboud University Nijmegen, Nijmegen, The Netherlands
Abstract
Mankind is unique in her ability for observational learning, i.e. the transmission of acquired knowledge and behavioralrepertoire through observation of others’ actions. In the present study we used electrophysiological measures to investigatebrain mechanisms of observational learning. Analysis investigated the possible functional coupling between occipital(alpha) and motor (mu) rhythms operating in the 10Hz frequency range for translating ‘‘seeing’’ into ‘‘doing’’. Subjectsobserved movement sequences consisting of six consecutive left or right hand button presses directed at one of two target-buttons for subsequent imitation. Each movement sequence was presented four times, intervened by short pause intervalsfor sequence rehearsal. During a control task subjects observed the same movement sequences without a requirement forsubsequent reproduction. Although both alpha and mu rhythms desynchronized during the imitation task relative to thecontrol task, modulations in alpha and mu power were found to be largely independent from each other over time, arguingagainst a functional coupling of alpha and mu generators during observational learning. This independence wasfurthermore reflected in the absence of coherence between occipital and motor electrodes overlaying alpha and mugenerators. Instead, coherence analysis revealed a pair of symmetric fronto-parietal networks, one over the left and one overthe right hemisphere, reflecting stronger coherence during observation of movements than during pauses. Individualdifferences in fronto-parietal coherence were furthermore found to predict imitation accuracy. The properties of thesenetworks, i.e. their fronto-parietal distribution, their ipsilateral organization and their sensitivity to the observation ofmovements, match closely with the known properties of the mirror neuron system (MNS) as studied in the macaque brain.These results indicate a functional dissociation between higher order areas for observational learning (i.e. parts of the MNSas reflected in 10Hz coherence measures) and peripheral structures (i.e. lateral occipital gyrus for alpha; central sulcus formu) that provide low-level support for observation and motor imagery of action sequences.
Citation: van der Helden J, van Schie HT, Rombouts C (2010) Observational Learning of New Movement Sequences Is Reflected in Fronto-ParietalCoherence. PLoS ONE 5(12): e14482. doi:10.1371/journal.pone.0014482
Editor: Clayton T. Dickson, University of Alberta, Canada
Received July 21, 2010; Accepted December 10, 2010; Published December 31, 2010
Copyright: � 2010 van der Helden et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permitsunrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: HvS was supported by the European Union Integrated Project JAST (Joint Action Science and Technology), grant FP6-IST2-003747. Funders had no rolein study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Competing Interests: The authors have declared that no competing interests exist.
* E-mail: [email protected]
. These authors contributed equally to this work.
Introduction
Many behavioural skills that humans establish during their life are
acquired through observational learning. An important neurophys-
iological principle that has been hypothesized to underlie observa-
tional learning and imitation is motor resonance, i.e. the automatic
activation of motor representations during action observation.
Neuroimaging experiments in humans and single cell studies in
macaques have investigated the neurophysiological basis of motor
resonance during action observation [1]. In particular, motor
resonance has been described as a basic property of mirror neurons, a
selection of motor neurons in monkey higher-order premotor and
parietal areas that are activated in a comparable manner during
action execution and during the observation of a similar action by
another individual [2,3]. At a larger scale neuroimaging experiments
in humans confirm the existence of a mirror neuron system in man
comprised of the inferior frontal gyrus, and the inferior parietal lobe
supporting both action observation [4] and imitation [5,6].
The mirror neuron system (MNS) has been shown to be
activated to a stronger extent if observers view movements that
they already have in their repertoire. Calvo-Merino and others
[7,8] for instance found that the MNS of dancers that observed
dancing movements in the fMRI scanner, responded more
strongly if these movements were part of their dancing routine.
Orgs and others [9] showed that dancing experience was
correlated with motor resonance, reflected in mu- and beta-
suppression in the dancer’s electroencephalogram (EEG), when
they observed a familiar dance movement. Recent findings from
van Elk and others [10] further extend these findings to the
domain of natural motor development. In their study, EEG was
recorded from 14- to 16-month old infants during observation of
videos showing other infants crawling and walking. EEG mu- and
beta-suppression was found strongly related to the infant’s own
motor experience, suggesting that already early in life an
individuals’ action experience determines how the actions of
others are processed.
Whereas the above studies convincingly showed effects of motor
experience to influence the observation of actions by others,
involvement of the human MNS in the reverse direction, i.e.
during the acquisition of a new action repertoire, has until recently
PLoS ONE | www.plosone.org 1 December 2010 | Volume 5 | Issue 12 | e14482

remained untested [11]. Several fMRI studies in the last few years
suggest that activity in core areas of the MNS (inferior frontal
gyrus and inferior parietal lobe) and in cortical and subcortical
regions supporting motor learning and visual perception, increase
as a function of the complexity of the movements that are to be
acquired [6,12–14]. The results of these experiments are inspiring
as they reveal some of the underlying neural circuitry supporting
observational learning. However, still little is known about the
neural mechanisms and functional processes that operate within
the extent of spatial activation patterns as indicated by functional
imaging studies. For instance, not much is known about the
functional contribution of areas and the integration of information
between them in supporting functional aspects of observational
learning over time.
Two interesting proposals concerning the possible functional
integration of information between areas supporting observational
learning have recently been formulated. Iacoboni [5] proposes that
observational learning is supported by internal (inverse and
forward) models that dynamically link visual and motor processes
to support the observation and imitation of new behavior. Internal
models were first conceived to serve the execution of actions by
allowing sensory structures to anticipate the perceptual conse-
quences of initiated actions (via forward internal models), and use
the perceptual error – i.e. the difference between the perceived
and anticipated perceptual result – to generate corrective motor
commands (using inverse internal models). Iacoboni proposes that
similar to action execution, internal models may also be engaged
during the observation of others’ actions, providing an internal
simulation of the observed behavior. In his model, motor
representations in parietal and frontal parts of the MNS are
dynamically linked with perceptual representations in the posterior
superior temporal sulcus. Similarly, Pineda [15] proposes that
action observation (e.g. for observational learning) may cause
alpha and mu rhythms in respective visual and sensorimotor
structures to become transiently linked supporting the translation
of ‘‘seeing’’ and/or ‘‘hearing’’ into ‘‘doing’’. According to this
global entrainment hypothesis the coupling between perception and
action is realized via global alpha/mu entrainment, resulting in
widespread coherence between local alpha and mu generators in
visual, auditory and motor structures when tasks demand
integrated cognitive processing.
However, according to the direct matching hypothesis of Rizzolatti,
Fogassi, and Gallese [1] imitation learning is primarily associated
with resonance in the motor system and has to be distinguished
from activation in the visual system. According to this hypothesis
observational learning depends on the core areas of the MNS, the
inferior frontal gyrus and the inferior parietal lobe that are
supporting motor simulation of observed movements. Different
from the global entrainment hypothesis, the direct matching
hypothesis does not predict alpha and mu source activations to
become synchronized during observational learning. Instead, the
direct matching hypothesis would predict coupling between frontal
and parietal motor regions that make up the core MNS.
Electroencephalograpic (EEG) measurements provide an excel-
lent opportunity to investigate the contribution of visual and motor
processes and their hypothesized integration during observational
learning. Both EEG power and EEG coherence measures may
provide relevant information to uncover the possible relationship
between perceptual and action processes in observational learning.
Variations in alpha and mu power over time, i.e. during different
stages of observational learning, may help to determine a possible
relationship between these two components. Also, EEG coherence
[16], which provides a measure of the functional connectivity as
expressed by the relative phase stability between oscillations
recorded from different sites, may be used to uncover relations
between different cortical areas supporting observational learning.
Previous EEG studies in the domain of action observation have
mostly focused on EEG power and found that observation of a
movement is accompanied by a transient drop in the mu and beta
power. Mutukumaraswamy and others, for instance [17,18] have
shown the amplitude of the mu rhythm to attenuate when subjects
observe a goal-directed grasping movement in much the same way
as it is attenuated when subjects performed this same movement
themselves. Surprisingly, whereas previous studies have consis-
tently focused on desynchronization of the mu rhythm, modula-
tions in alpha power during action observation have been largely
neglected. Similar to the mu rhythm, alpha, which is strongest
over visual cortices, is found to desynchronize to the presentation
of stimuli (e.g. actions) and during tasks that require visual
attention, e.g. for selecting, anticipating or remembering visual
stimuli [15,19–22]. Consequently, in previous studies alpha and
mu may have been confounded and the possible individual and
joint effects of either component to observational learning remain
to be determined. Note that although beta power (,16–30Hz),
similar to mu, is found to desynchronize during action observation
[9,23,33], its sources are mainly centrally distributed and thus not
likely to be confounded with a posterior source.
The present study focuses on the learning of new action
sequences through observation, analogue to the observations of
etiologists and psychologists [1,24–26] that new actions may be
learned by recognizing the elementary motor acts and the specific
order or sequence between them. For this type of observational
learning to work, it is necessary that all movement elements are
already within the motor repertoire of the observer. In the present
study we used button presses that are well established in the motor
repertoire of adult subjects and known for their ability to generate
motor activation in an observer as shown by neuroimaging [27–
29] and electro- and magnetoencephalographic studies [30–33].
Sequences of movements were repeated four times to allow
subjects to effectively memorize the correct order of movements
for subsequent reproduction (see Figure 1). Furthermore, each
action sequence repetition was intervened by a short pause to
study the neural processes supporting retention and rehearsal of
these movements. That is, whereas much attention has been
directed at action observation, an important requirement for
successful imitation appears to lie in the ability of the observer to
retain or rehearse the order of successive motor acts for subsequent
imitation. Most likely, efficient rehearsal involves a process of
mental imagery in which subjects mentally simulate the motor and
perceptual effects that would accompany the actual execution of
such actions [11,34]. Consistent with this view, Buccino and
colleagues [6] found that subjects activated large parts of the
MNS, dorsal motor regions and the dorsolateral prefrontal cortex
during a pause interval that intervened between the observation
and execution phase in an experiment.
To capture the specific neural systems supporting imitation
learning, a control task was included in which subjects viewed the
same action sequences, without a requirement for subsequent
reproduction, but had to subsequently report deviant movements.
Results
The data of one of the participants contained too many artifacts
to allow a reliable estimate of coherence. Data of this participant
was excluded from both power and coherence analyses. Analysis of
performance showed that the imitation task was challenging for
most subjects. No participant succeeded in flawless performance.
The amount of errors varied between 2 and 33 of the 40 trials
Observational Learning
PLoS ONE | www.plosone.org 2 December 2010 | Volume 5 | Issue 12 | e14482

between participants (mean percentage correct = 61.5%,
SD = 23.3%).
Power analysisStatistical analysis of power in the alpha/mu frequency range
was tested using a 2626264 within-subject ANOVA, with the
factors Location (posterior electrodes pairs, central electrode
pairs), Task (imitation, detection), Period (observation, pause),
and Repetition (S1, S2, S3, S4). The percentage of correct
sequence repetitions of each subject was added as a covariate to
investigate if individual variations in alpha and mu power were
systematically related to individual differences in imitation ability.
Figure 2 presents the grand average activity in the mu/alpha
frequency range over central and posterior sites. Alpha power over
extrastriate visual areas at electrodes PO3 and PO4 was found to
be stronger than mu power over motor regions at electrodes C3
and C4, as reflected by an effect of Location, F(1, 13) = 21.6,
p,.001. Furthermore, a main effect of Task was found, indicating
that both mu and alpha were more strongly desynchronized in the
imitation task than in the detection task, F(1, 13) = 18.8, p,.001.
Mu suppression over central areas was stronger during pauses than
during actual movement observation, while over posterior
electrodes the reverse was found, i.e. alpha was suppressed more
strongly during observation than pause intervals, reflected by an
interaction between Location and Period, F(1, 13) = 29.6, p,.001.
Importantly, the power increase in alpha over visual areas during
the pause was stronger in the detection task than in the imitation
task. On the contrary, over motor regions a stronger suppression
of mu power was found during the pause of the imitation task as
compared to the pause of the detection task, reflecting the
interaction between Location, Task, and Period, F(1, 13) = 4.4,
p = .056. This latter effect points to a functional difference between
mu and alpha in their contribution to observation and retention of
movements in the two tasks. In order to further specify this three-
way interaction two post-hoc analyses were conducted focusing on
posterior electrodes PO3/PO4 and central electrodes C3/C4,
separately. The results of these analyses are presented hereafter.
The first ANOVA repeated measures contrast analysis directed
at posterior alpha power revealed a main effect of Task reflecting
stronger alpha desynchronization during the imitation task as
compared to the detection task, F(1, 13) = 14.9, p,.01. In both
tasks alpha desynchronized during observations of movements as
compared to the subsequent pauses, as indicated by a main effect
of Period, F(1, 13) = 11.2, p,.005. The topographical distribution
of this effect is shown in the bottom left of Figure 2. Furthermore,
a significant interaction between Task and Period was found, F(1,
Figure 1. Schematic Overview of the Task Structure. (A) Each trial consisted of 4 repetitions (S1 – S4) of an observation interval (6 s) andsubsequent pause interval (5 s). Following these 4 repetitions, subjects either tried to imitate the movement sequence from memory (imitation task)or responded if they had observed a catch movement in the action sequence (detection task). (B) Each observation epoch consisted of six buttonpress movements, followed by a 5000 ms pause interval. Each button press movement consisted of a 500 ms display showing the left and right handat the respective left and right start buttons (S) and a subsequent 500 ms apparent motion display (M) showing either the left or the right handpressing an ipsi- or contralateral target button. (C) Examples of stimuli: Start display (S) showing the two hands pressing the start buttons; Example ofan ipsilateral movement (M1); Example of a contralateral movement (M2); During the Pause display the central red button on the response box wasilluminated.doi:10.1371/journal.pone.0014482.g001
Observational Learning
PLoS ONE | www.plosone.org 3 December 2010 | Volume 5 | Issue 12 | e14482
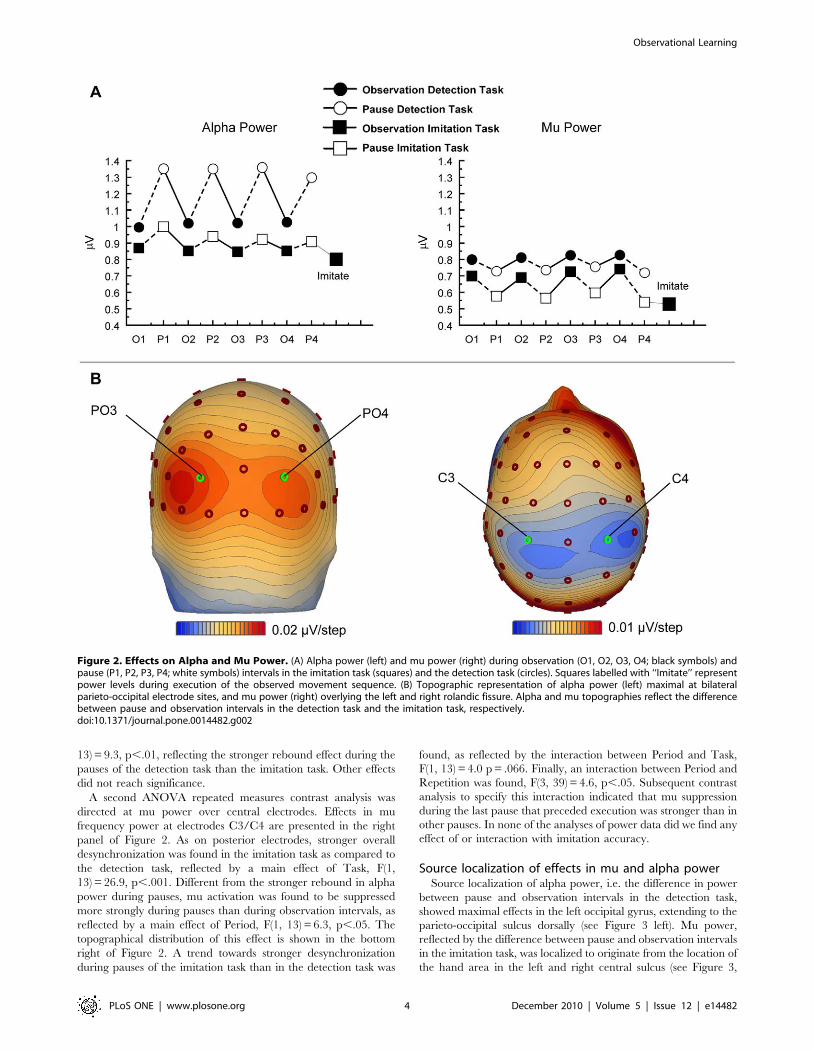
13) = 9.3, p,.01, reflecting the stronger rebound effect during the
pauses of the detection task than the imitation task. Other effects
did not reach significance.
A second ANOVA repeated measures contrast analysis was
directed at mu power over central electrodes. Effects in mu
frequency power at electrodes C3/C4 are presented in the right
panel of Figure 2. As on posterior electrodes, stronger overall
desynchronization was found in the imitation task as compared to
the detection task, reflected by a main effect of Task, F(1,
13) = 26.9, p,.001. Different from the stronger rebound in alpha
power during pauses, mu activation was found to be suppressed
more strongly during pauses than during observation intervals, as
reflected by a main effect of Period, F(1, 13) = 6.3, p,.05. The
topographical distribution of this effect is shown in the bottom
right of Figure 2. A trend towards stronger desynchronization
during pauses of the imitation task than in the detection task was
found, as reflected by the interaction between Period and Task,
F(1, 13) = 4.0 p = .066. Finally, an interaction between Period and
Repetition was found, F(3, 39) = 4.6, p,.05. Subsequent contrast
analysis to specify this interaction indicated that mu suppression
during the last pause that preceded execution was stronger than in
other pauses. In none of the analyses of power data did we find any
effect of or interaction with imitation accuracy.
Source localization of effects in mu and alpha powerSource localization of alpha power, i.e. the difference in power
between pause and observation intervals in the detection task,
showed maximal effects in the left occipital gyrus, extending to the
parieto-occipital sulcus dorsally (see Figure 3 left). Mu power,
reflected by the difference between pause and observation intervals
in the imitation task, was localized to originate from the location of
the hand area in the left and right central sulcus (see Figure 3,
Figure 2. Effects on Alpha and Mu Power. (A) Alpha power (left) and mu power (right) during observation (O1, O2, O3, O4; black symbols) andpause (P1, P2, P3, P4; white symbols) intervals in the imitation task (squares) and the detection task (circles). Squares labelled with ‘‘Imitate’’ representpower levels during execution of the observed movement sequence. (B) Topographic representation of alpha power (left) maximal at bilateralparieto-occipital electrode sites, and mu power (right) overlying the left and right rolandic fissure. Alpha and mu topographies reflect the differencebetween pause and observation intervals in the detection task and the imitation task, respectively.doi:10.1371/journal.pone.0014482.g002
Observational Learning
PLoS ONE | www.plosone.org 4 December 2010 | Volume 5 | Issue 12 | e14482

right). In addition, the minimum norm displays an effect in the
right occipital gyrus reflecting the difference in alpha power
between observation and pause intervals of the imitation task that
was found concurrently over posterior areas.
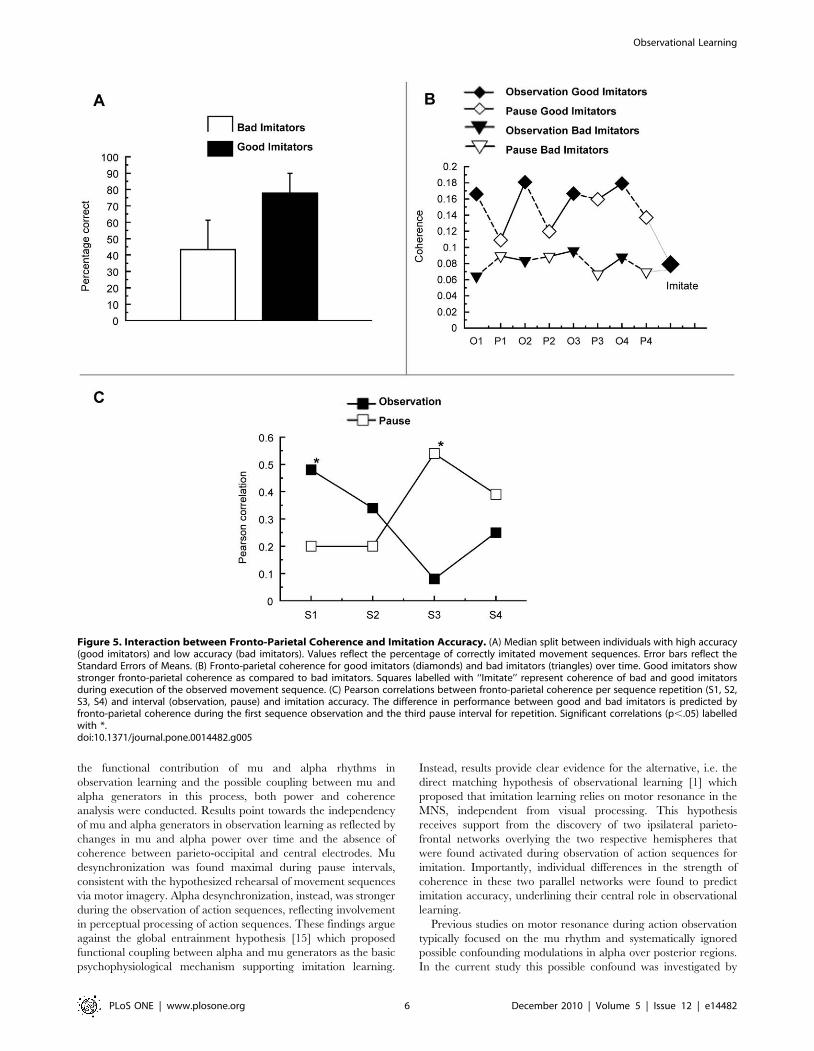
Coherence analysisFigure 4 presents the network found to be activated more
strongly during the observation of movements than during pause
intervals (see Method section below for details on the procedure).
This network basically consists of two individual networks each
connecting parietal, central and frontal areas in each respective
hemisphere.
An ANCOVA with Period (observation, pause) and Repetition
(S1, S2, S3, S4) as within-subject factors and imitation accuracy as
a covariate tested if coherence within the pair of fronto-parietal
networks during observation for imitation was related to individual
differences in imitation performance. Within the networks,
coherence varied as a function of imitation accuracy, F(1,
12) = 4.8, p = .049, showing larger coherence for subjects with
higher imitation accuracy. Furthermore, imitation accuracy
significantly interacted with Period and Repetition, F(3,
36) = 4.0, p = .026. This interaction effect is reflected in
Figure 5B where the coherence data is split between subjects with
high and low imitation accuracy (Figure 5A). To further specify
how coherence in the fronto-parietal networks is supporting
imitation accuracy over time, a series of correlation analyses were
performed between imitation accuracy and each consecutive
observation and pause. One subject with exceptionally high
coherence values (.0.6) was removed from the correlation analysis
to avoid spurious correlations. This did not affect the overall
pattern of correlation results. The results of this correlation
analysis are represented in Figure 5C. Interestingly, a positive
correlation was found between imitation accuracy and fronto-
parietal coherence during the first observation of the sequence,
r = 0.48, p = .05. No significant correlation was found in the
subsequent pause, r = 0.20, p = .25. Interestingly, a reversed
pattern of results was found during the third repetition, where
imitation accuracy correlated positively with fronto-parietal
coherence during the pause, r = 0.54, p = .027, but not during
observation of movement sequences. These findings suggest that
the critical difference between individuals who do well in imitation
learning and those who do not, is that the former will i) coherently
activate fronto-parietal systems during the initial observations of
the movement sequence, and ii) are able to reproduce fronto-
parietal activation during pause intervals after having observed
several repetitions of the same movement sequence.
Discussion
In the present study we investigated the neural mechanisms
supporting the acquisition of new action sequences focusing on
brain oscillations in the mu/alpha frequency range. To determine
Figure 3. Localization of the Effects in Alpha and Mu Power. The alpha and mu effects (pause – observation) were calculated using minimum-norm analysis in BESA 5.1.6. (Brain Electrical Source Analysis, MEGIS Software, Germany). The minimal norm was computed with spatiotemporalweighting according to [49] fitted to an individual Talairach transformed brain surface. Effects are scaled with respect to the maximum differencebetween pause and observation intervals (100%, in blue). The left sources represent the alpha power, reflecting the difference between pause andobservation intervals in the detection task, and the right sources represent the mu power, reflecting the difference between pause and observation inthe imitation task.doi:10.1371/journal.pone.0014482.g003
Figure 4. Fronto-parietal Coherence Network. Pairs of electrodesshowing higher mu-frequency coherence during the observation ofmovement sequences than during the pause intervals. Only electrodepairs with the largest normalized coherence effects (1.z.2, z.2) areshown.doi:10.1371/journal.pone.0014482.g004
Observational Learning
PLoS ONE | www.plosone.org 5 December 2010 | Volume 5 | Issue 12 | e14482
the functional contribution of mu and alpha rhythms in
observation learning and the possible coupling between mu and
alpha generators in this process, both power and coherence
analysis were conducted. Results point towards the independency
of mu and alpha generators in observation learning as reflected by
changes in mu and alpha power over time and the absence of
coherence between parieto-occipital and central electrodes. Mu
desynchronization was found maximal during pause intervals,
consistent with the hypothesized rehearsal of movement sequences
via motor imagery. Alpha desynchronization, instead, was stronger
during the observation of action sequences, reflecting involvement
in perceptual processing of action sequences. These findings argue
against the global entrainment hypothesis [15] which proposed
functional coupling between alpha and mu generators as the basic
psychophysiological mechanism supporting imitation learning.
Instead, results provide clear evidence for the alternative, i.e. the
direct matching hypothesis of observational learning [1] which
proposed that imitation learning relies on motor resonance in the
MNS, independent from visual processing. This hypothesis
receives support from the discovery of two ipsilateral parieto-
frontal networks overlying the two respective hemispheres that
were found activated during observation of action sequences for
imitation. Importantly, individual differences in the strength of
coherence in these two parallel networks were found to predict
imitation accuracy, underlining their central role in observational
learning.
Previous studies on motor resonance during action observation
typically focused on the mu rhythm and systematically ignored
possible confounding modulations in alpha over posterior regions.
In the current study this possible confound was investigated by
Figure 5. Interaction between Fronto-Parietal Coherence and Imitation Accuracy. (A) Median split between individuals with high accuracy(good imitators) and low accuracy (bad imitators). Values reflect the percentage of correctly imitated movement sequences. Error bars reflect theStandard Errors of Means. (B) Fronto-parietal coherence for good imitators (diamonds) and bad imitators (triangles) over time. Good imitators showstronger fronto-parietal coherence as compared to bad imitators. Squares labelled with ‘‘Imitate’’ represent coherence of bad and good imitatorsduring execution of the observed movement sequence. (C) Pearson correlations between fronto-parietal coherence per sequence repetition (S1, S2,S3, S4) and interval (observation, pause) and imitation accuracy. The difference in performance between good and bad imitators is predicted byfronto-parietal coherence during the first sequence observation and the third pause interval for repetition. Significant correlations (p,.05) labelledwith *.doi:10.1371/journal.pone.0014482.g005
Observational Learning
PLoS ONE | www.plosone.org 6 December 2010 | Volume 5 | Issue 12 | e14482

studying discrepancies between alpha and mu power during the
different phases of imitation learning. Consistent with the
prediction from the global entrainment hypothesis that imitation
learning would be supported by both visual and motor processes
[15] alpha and mu rhythms were both found to desynchronize
when participants observed and remembered sequences of
movements for subsequent imitation, as compared to a control
task in which subjects detected deviating (thumb) movements. This
suggests that both visual and motor processes contributed to the
current task of observational learning. Importantly however, no
functional coherence was detected between visual and motor
processes within the alpha/mu band as was predicted by the global
entrainment hypothesis [15] and modulations in alpha and mu
power were found to be largely independent from each other,
arguing against the functional integration of alpha and mu
oscillations in support of action observation and observational
learning.
The independence between visual and motor oscillations in the
mu/alpha range differs from other studies that found posterior –
central coherence in the 8–13 Hz frequency range [35,36]. In an
experiment by Classen and others [36], for instance, subjects had
to exert an oscillating force with their right index finger on a strain
gauge. It was shown that central-posterior alpha coherence was
highest when the force exerted had to be synchronized with a
visually presented oscillating target dot. Another example is
provided by Hummel and Gerloff [35] who simultaneously
presented a tactile Braille letter to the right index finger tip and
a visual Braille letter on a computer screen, resulting in a phasic
increase in alpha coherence between central and posterior
electrodes that was higher if subjects were successful in verifying
if the visual and tactile letter matched or not. The main difference
between these studies and the current study is probably that task
properties of these other studies demand the immediate integra-
tion between visual and sensorimotor information, whereas for
observational learning the requirement for temporal integration
between visual and motor information is less demanding. One
other possibility may be that for observational learning to be
effective, motor representations are to be protected from visual
input that may interfere with the retention or rehearsal of action
sequences, as was indicated by behavioral studies that investigated
interference effects during learning of whole body movement
patterns [37]. Irrespective, however, of the precise reason for the
absence of visuomotor integration during imitation learning, the
coherence data of the current experiment suggest that such an
integration of visual and sensorimotor information is not a
necessary requirement for observational learning to take place.
In line with the conclusion that visual and motor processes
operated largely independently from each other, several dissoci-
ations between visual and motor processes as reflected in alpha
and mu power were found. One prominent difference in mu and
alpha power was that alpha desynchronization was strongest
during the observation interval, suggesting maximal sensitivity to
visual input, whereas mu desynchronization was strongest during
the pause intervals, suggesting active involvement in the rehearsal
of action sequences for subsequent reproduction. The latter
finding of enhanced mu desynchronization during pause intervals
is particularly interesting because it suggests that subjects engaged
in active motor imagery supporting the maintenance and/or
encoding of action sequences in memory for subsequent
reproduction. Consistent with this interpretation, desynchroniza-
tion effects in mu power were found to be stronger in the imitation
task as compared to the detection task. Source localization of mu
desynchronization using a minimum norm approach showed
activation to originate from pre- and post-central gyri around the
location of the hand area in the left and right central sulcus.
Consistent with the interpretation that mu desynchronization
during pauses may be a measure of motor imagery, EEG studies
have reliably found motor imagery accompanied by desynchro-
nization in mu oscillations originating from primary sensorimotor
areas [38,39].
Previous studies investigating action observation of finger
movements, facial expressions and goal directed grasping
[17,18,40] have firmly implicated mu desynchronization as a
measure of motor resonance. This view has furthermore been
strengthened by studies that found stronger mu desynchronization
to the observation of actions that were already established in the
observer’s motor repertoire (e.g. dancing moves by professional
dancers [9], or infants’ ability to crawl or walk [10]) as compared
to actions that were not mastered yet. The present findings further
extend the functional role of mu desynchronization beyond action
observation by showing that mu is not only involved in the
observation of already learned actions but also directly supports
the acquisition of new actions. Importantly, although finger
movements are probably well established in the observers’ motor
repertoire, covert motor activation was stronger during the pause
intervals than during actual action observation. This result points
towards the involvement of mu in both action observation and
motor imagery, and suggests that, when studying imitation and
learning of new motor repertoire, one should be careful to
distinguish between motor activation resulting from action
observation and motor activation reflecting motor imagery.
Consistent with the direct matching hypothesis, coherence
analysis rendered two ipsilateral fronto-parietal networks, one over
the left and one over the right hemisphere, revealing stronger
fronto-parietal coherence during the observation of movement
sequences than during subsequent pauses. The properties of these
two networks, i.e. their fronto-parietal distribution, their ipsilateral
organization and their sensitivity to the observation of movements,
matches closely with the known properties of the MNS as
uncovered by anatomical and electrophysiological studies in the
macaque brain [41]. These studies have found strong parallel and
reciprocal connections between ipsilateral regions of the frontal
and parietal lobes supporting specific sensorimotor transforma-
tions for action execution and action observation [42]. The present
study is the first to find a possible correlate of the MNS reflected in
fronto-parietal coherence. These findings call for further confir-
mation, e.g. by studies that combine EEG and fMRI.
Importantly, the strength of fronto-parietal coherence during
imitation learning was found to be positively related to the
accuracy of subsequent imitation. This suggests that the strength of
the functional coupling between parietal and frontal parts of the
network for imitation learning is decisive for the accuracy of newly
acquired action sequences. Neuroimaging studies have suggested
different functional roles for frontal and parietal parts of the MNS
[43,44], for processing the goals of actions (i.e. the end-location or
the object being targeted) and the means of observed actions (i.e.
the selected grasp or movement trajectory). In the present
sequence learning task participants needed to integrate informa-
tion about the hand that was used (action means) and the specific
target button visited (action goal) for each consecutive pointing
action. A reasonable inference is that enhanced coherence
between frontal and parietal MNS regions may support the
integration of information about action goals and means during
action observation and rehearsal for subsequent imitation.
Consistent with this interpretation, Fogassi et al [45] found that
the discharge rates of cells supporting the execution and
observation of grasping acts in the macaque inferior parietal lobe
were sensitive to the final goals of grasping actions (i.e. grasping to
Observational Learning
PLoS ONE | www.plosone.org 7 December 2010 | Volume 5 | Issue 12 | e14482

place vs. to eat), i.e. reflecting the capacity of the MNS to
anticipate and integrate successive motor acts (grasping and
placing) in an action chain. Interestingly, the anticipatory effects of
action chaining were found both during execution and observation
of similar grasping acts by an experimenter, confirming the
functional contribution of the MNS to both action production and
action observation.
Regarding the present study, correlation analyses between
imitation accuracy and fronto-parietal coherence per repetition
and period, indicated that performance differences were most
clearly expressed during the first observation of the movement
sequence and the third pause interval. This pattern suggests that,
in addition to the general difference in fronto-parietal coherence
between good and bad imitators, good imitators pay better
attention to, or are better able to integrate information, about
action goals (left/right button) and action means (left/right hand)
during the first sequence presentation. Furthermore, the correla-
tion during the third pause interval suggests that the functional
contribution of fronto-parietal coherence is not limited to action
observation only, but may also support the active rehearsal of a
movement sequence in memory [45]. The positive correlation
between imitation accuracy and fronto-parietal coherence during
the third pause interval suggests that good imitators were better
able to activate a correct representation of consecutive goal-means
relations in memory for subsequent reproduction.
An interesting dissociation is that imitation accuracy correlated
with fronto-parietal coherence, but not with mu and alpha power.
The absence of a correlation between imitation accuracy and
power measures confirms the interpretation that both alpha and
mu power represent activation in relative peripheral systems, i.e
visual extrastriate regions and sensorimotor cortex, that may not
be directly involved in representing the order of an action
sequence or the integration between goals and means, but seem to
provide low level support during the observation and motor
imagery of action sequences in memory. Instead, both the
topography of fronto-parietal coherence overlying higher-order
motor systems in frontal and parietal cortices and the relation
between coherence strength and imitation accuracy point towards
a central role in imitation learning.
An important question to ask is to what extent the fronto-
parietal coherence that we found to support observational learning
is representative for the learning of new motor repertoire in more
naturalistic settings. One property of the present task is that was
quite difficult for subjects to accomplish faultless imitation of the
entire sequence. The complexity and difficulty of the present
sequence learning task may not, however, be that different from
natural conditions in which individuals e.g. have to learn a new
sequence of dance steps, or learn the correct order of actions
(switching gears, use indicator, looking, etc.) when approaching a
crossroad during driving lessons. It is still unclear however to what
extent fronto-parietal coherence may support other types of
observational learning where the focus is not on combining
established movement elements in a new arrangement, but on the
learning of an entirely new movement primitive, for instance when
children in elementary school have to imitate a teacher’s posture of
holding a pen to allow writing.
In conclusion, the present study was successful in uncovering
different neural mechanisms and their potential roles in observa-
tional learning. Observational learning was found to be supported
by a pair of fronto-parietal coherence networks, one overlaying the
left and one overlaying the right hemisphere. Coherence in these
fronto-parietal networks likely provides a direct measure of
activation and coherence within the human MNS. Fronto-parietal
coherence predicted the accuracy of subsequent imitation,
indicating a central role for these structures in imitation learning.
In addition, observational learning was found to be supported by
activation in visual extrastriate and central motor regions, reflected
by power modulations in alpha and mu components. Although
visual and motor processes reflected in alpha and mu power may
be jointly activated in support of action observation and motor
imagery, both the variance in mu and alpha power over time and
the absence of coherence between occipito-parietal and central
motor regions argue against the global entrainment hypothesis and
the hypothesized coupling between alpha and mu oscillations in
support of observational learning. Although the present study
indicates a role for visual processes in imitation learning, the
current findings generally confirm the direct-matching hypothesis
of imitation learning which states that observational learning
depends on motor resonance in high-level motor structures
making up the core MNS. In addition the results obtained during
the pause intervals make clear that imitation learning, for a good
part, relies on motor imagery and that studies investigating
observational learning will need to distinguish between motor
activation resulting from action observation and motor imagery for
subsequent imitation. We hope that the current study will inspire
other researchers to further investigate the neural and functional
basis of one of most central of human abilities, i.e. the capacity to
learn new behavioral repertoire, through action observation.
Materials and Methods
ParticipantsThe study was approved by the ethics committee of the Faculty
of Behavioural Sciences from the University of Twente. Fifteen
university students (9 females) participated in the experiment and
were given course credits as reward. All participants were between
18 and 26 years of age, with a mean age of 20.5 years (SD = 2y).
Subjects were right handed as assessed by the Annett Handedness
Inventory [46] and provided written informed consent before the
study.
ApparatusA custom made response-box was used, consisting of four
buttons that were arranged in a square shape and a fifth button
located in the middle (see Figure 1). Each button had a built-in red
LED which could be illuminated. The response-box was
connected to a PC and positioned in front of a 17 inch computer
monitor (Philips 107T5, refresh-rate 85 Hz) that was used for
presenting visual stimuli to subjects. The experiment was
programmed and controlled in E-prime (Version 1.2., Psychology
Software Tools, Inc. Pittsburgh, USA).
ProcedureIn each trial, participants were presented with a sequence of 6
button presses on the computer screen. For each button press
movement two subsequent pictures were presented, the first one
showing the index fingers of both hands at the left and right
(proximal) starting buttons of the response box, and a second
picture showing one of the hands pressing one of two (distal) target
button (see Figure 1). Six individual button press movements were
presented in succession with each movement lasting 1000 ms (12
pictures of 500 ms each). The end-locations of each movement
were selected from a total of four pictures that reflected the
different combinations of hand (left, right) and button (left, right).
Each of these four movements was presented at least once in every
sequence of movements, but never thrice, and the same movement
was never repeated more than once in succession. Directly
following the sequence of six movements, a 5000 ms pause was
Observational Learning
PLoS ONE | www.plosone.org 8 December 2010 | Volume 5 | Issue 12 | e14482

administered during which a picture was presented showing the
hands in the starting posture and the central red button
illuminated to demarcate the pause interval from the sequence
repetitions.
In the imitation task subjects were asked to replicate the
observed sequence of movements after having seen 4 sequence
repetitions, using the same response box that was depicted on the
computer screen. Subjects first pushed the two buttons that
represented the starting positions. As a result, the central button on
the response pad was illuminated for 5000 ms, during which a text
was presented on the screen informing the subject that, as soon as
the centre button on the response pad was turned off, they had to
execute the observed movement sequence as accurately as
possible. Accuracy of each movement was evaluated with respect
to whether subjects released the correct starting button (i.e.
whether they used the correct hand) and whether they pushed the
correct target button. If either the wrong hand was used or the
wrong target button was pushed in one of the movements, subjects
received feedback that they made one or more errors. If the
sequence of movements was executed correctly, the time to
execute the entire sequence was presented on the screen. Subjects
pushed a button on the button pad when they were ready to start a
new sequence.
In the detection task, the movements and sequences were the
same as in the imitation task with the exception that in 20% of the
presented sequences one of the movements was replaced by a
catch movement in which a target button was pushed with the
thumb instead of the index finger. After each presentation of the
sequence, subjects had to report if a catch movement had
occurred. If so, they had to push any button on the response-box
within the 5000 ms during which the visual pause was adminis-
tered. If the subject missed a catch movement or responded falsely,
i.e. when no deviant was shown, ‘‘WRONG!’’ was projected over
the warning display for 1000 ms, after which a new, neutral
warning signal was displayed for 5 s, and the experiment
continued. Sequences containing catch movements or false alarms
were omitted from the EEG analyses to rule out the possibility of
response preparation contaminating the detection task.
Electrophysiological recordingSixty-one-channel EEG was recorded according to the
International 10–20 System of Electrode Placement [47], using a
BrainVision QuickAmp amplifier (Brain Products GmbH, Mu-
nich, Germany) in combination with shielded Ag/AgCl electrodes.
Electrode impedances were kept below 10 kV. EEG was sampled
at 500 Hz with a 140 Hz low pass filter and a 50 Hz notch filter.
An average reference was used during acquisition of the EEG
signal.
Because arm and hand movements are known to influence the
mu rhythm, participants were instructed to position their hands
along each side of the response box on the table and to move as
little as possible during sequence observation in both tasks.
Electromyograms (EMG) were measured using bipolar electrodes
over flexor carpi radialis and extensor pollicis longus on both forearms, so
that covert movements made by participants could be detected
[17]. Vertical and horizontal electro-oculograms (EOG) were
recorded using bipolar electrodes placed at the supraorbital and
infraorbital ridge of the right eye and the outer canthi of the left
and right eye respectively. Participants were instructed to fixate on
the middle button of the response box presented on the computer
monitor during sequence observation in both tasks. EEG, EMG
and EOG data were analyzed offline using BrainVision Analyzer
v1.05 (Brain Products GmbH, Munich, Germany) and BESA
5.1.6. (Megis Software GmbH, Grafelfing, Germany).
AnalysisPower analysis. EEG recordings in the imitation and
detection tasks were analyzed separately for observation intervals
and pause intervals to distinguish between activation generated by
the observation and retention of movements. An additional
analysis was directed at the execution interval of the imitation
task during which participants performed the movement sequence
from memory. Observation intervals (6000 ms interval consisting
of 6 consecutive button presses of 1 s each), pause segments
(5000 ms interval following each sequence repetition), and the
execution interval (movement onset until movement end), were
segmented into equally sized epochs (of 512 time points). Epochs
containing EEG artefacts or EMG activity (execution interval
excluded) were discarded from analysis using an automated
procedure. This resulted in a rejection of 15.5% of all epochs in
the detection task and 8.8% of all epochs in the imitation task.
Furthermore, segments in which eye blinks were detected were
discarded, resulting in an additional rejection of 10.4% of all trials
of the detection task and 14.9% of the imitation task. For each
individual subject, Fast Fourier Transforms (FFT) were performed
on the artefact free EEG segments (512 points, Hanning window,
10% envelope), and averaged to create separate power frequency
spectra for the observation and pause intervals per repetition in the
imitation and detection tasks.
Each participant’s individual mu rhythm band was determined
by subtracting the power frequency spectrum of the sequence
execution phase during the imitation task (which contained the
lowest mu activation due to hand movements) from the power
frequency spectrum during the observation phase of the detection
task (which contained the highest mu activation). For each
participant this difference was topographically mapped and the
location of maximal mu modulation identified to determine each
participant’s individual mu frequency [48]. Mu power was
calculated using a +/20.5 Hz frequency bandwidth around each
individual’s mu frequency. Similarly, for each participant the
individual alpha rhythm was determined by subtracting the
frequency spectrum in the condition with the lowest alpha power
(observation of movements in the imitation task) from the
condition with the highest power of alpha (pause in the detection
task). For each individual, the posterior location and the frequency
at which alpha power was maximal were identified. Alpha power
was calculated using a +/20.5 Hz bandwidth around the each
individual’s alpha frequency. For statistical analysis bilateral
electrodes were identified at which modulations in mu power
(C3 and C4) and alpha power (PO3 and PO4) were found
maximal over subjects. Data from left and right hemispheres were
pooled in the statistical analyses.
Source localization. Localization of the effects in alpha and
mu power (pause-observation) was established using minimum-
norm analysis in BESA 5.1.6. (Brain Electrical Source Analysis,
MEGIS Software, Germany). The minimal norm was computed
with spatiotemporal weighting according to [49] fitted to an
individual Talairach transformed brain surface.
Coherence analysis. Raw data were filtered off-line (1–
50 Hz, slope 12 dB/octave) in advance of coherence analysis.
Epochs containing artefacts were discarded from analysis using an
automatic procedure. Segments with eye blinks were rejected.
Before coherence analysis a current source density analysis was
conducted (order of splines = 4, 10 polynomials) to remove
spreading of nearby source activation to neighbouring electrodes
[50,51]. An FFT was performed on each segment before
calculating the coherences within each segment. Coherence
values between electrode pairs were calculated for the
frequencies within the 8–13 Hz frequency range. Similar to the
Observational Learning
PLoS ONE | www.plosone.org 9 December 2010 | Volume 5 | Issue 12 | e14482

analysis of power, each subject’s individual coherence frequency
was identified by averaging coherence across all electrode pairs
and tasks and determining the peak frequency with maximum
coherence. Individual coherence values were calculated by taking
the average coherence activation in the +/20.5 Hz band
encompassing each individual’s coherence peak frequency.
The aim of the coherence analysis was to identify functional
networks differentiating between the observation and retention of
movements for imitation. To identify the networks that show
differential coherence between movement observation and
subsequent pauses, coherence averages of the observation interval
were subtracted from coherence averages during the pause. These
coherence difference scores were transformed to z-values to
identify electrode pairs with strongest coherence during the
observation of movements (z-values smaller than -1) and electrode
pairs with strong coherence during pause intervals (with z-values
larger than 1). Inspection of the resulting networks indicated a
specific pair of fronto-parietal networks that were maximally
coherent during the observation of movements. Statistical analysis
of coherence in the observation network was analyzed using a
within-subjects ANOVA repeated measures analysis to determine
effects of Period (pause, observation) and Repetition (S1, S2, S3,
S4), with imitation accuracy as a covariate to investigate
associations with individual differences in imitation ability.
Author Contributions
Conceived and designed the experiments: JvdH HTvS CR. Performed the
experiments: JvdH HTvS CR. Analyzed the data: JvdH HTvS CR. Wrote
the paper: JvdH HTvS.
References
1. Rizzolatti G, Fogassi L, Gallese V (2001) Neurophysiological mechanisms
underlying the understanding and imitation of action. Nat Rev Neurosci 2:
661–670.
2. Gallese V, Fadiga L, Fogassi L, Rizzolatti G (1996) Action recognition in the
premotor cortex. Brain 119(Pt 2): 593–609.
3. Gallese V (2005) Embodied simulation: From neurons to phenomenal
experience. Phenomenol Cogn Sci 4: 23–48.
4. Buccino G, Binkofski F, Fink GR, Fadiga L, Fogassi L, et al. (2001) Action
observation activates premotor and parietal areas in a somatotopic manner: an
fMRI study. Eur J Neurosci 13: 400–404.
5. Iacoboni M (2005) Neural mechanisms of imitation. Curr Opin Neurobiol 15:
632–637.
6. Buccino G, Vogt S, Ritzl A, Fink GR, Zilles K, et al. (2004) Neural circuits
underlying imitation learning of hand actions: an event-related fMRI study.
Neuron 42: 323–334.
7. Calvo-Merino B, Glaser DE, Grezes J, Passingham RE, Haggard P (2005)
Action observation and acquired motor skills: an FMRI study with expert
dancers. Cereb Cortex 15: 1243–1249.
8. Calvo-Merino B, Grezes J, Glaser DE, Passingham RE, Haggard P (2006)
Seeing or doing? Influence of visual and motor familiarity in action observation.
Curr Biol 16: 1905–1910.
9. Orgs G, Dombrowski JH, Heil M, Jansen-Osmann P (2008) Expertise in dance
modulates alpha/beta event-related desynchronization during action observa-
tion. Eur J Neurosci 27: 3380–3384.
10. van Elk M, van Schie HT, Hunnius S, Vesper C, Bekkering H (2008) You’ll
never crawl alone: Neurophysiological evidence for experience-dependent motor
resonance in infancy. Neuroimage. pp 808–814.
11. Jeannerod M (1994) The representing brain: neural correlates of motor intention
and imagery. Behav Brain Sci 17: 187–245.
12. Frey SH, Gerry VE (2006) Modulation of neural activity during observational
learning of actions and their sequential orders. J Neurosci 26: 13194–13201.
13. Molnar-Szakacs I, Kaplan J, Greenfield PM, Iacoboni M (2006) Observing
complex action sequences: The role of the fronto-parietal mirror neuron system.
Neuroimage 33: 923–935.
14. Vogt S, Buccino G, Wohlschlager AM, Canessa N, Shah NJ, et al. (2007)
Prefrontal involvement in imitation learning of hand actions: effects of practice
and expertise. Neuroimage 37: 1371–1383.
15. Pineda JA (2005) The functional significance of mu rhythms: translating
‘‘seeing’’ and ‘‘hearing’’ into ‘‘doing’’. Brain Res Brain Res Rev 50: 57–68.
16. Nunez PL, Srinivasan R, Westdorp AF, Wijesinghe RS, Tucker DM, et al.
(1997) EEG coherency. I: Statistics, reference electrode, volume conduction,
Laplacians, cortical imaging, and interpretation at multiple scales. Electroen
Clin Neuro 103: 499–515.
17. Muthukumaraswamy SD, Johnson BW (2004) Changes in rolandic mu rhythm
during observation of a precision grip. Psychophysiology 41: 152–156.
18. Muthukumaraswamy SD, Johnson BW, McNair NA (2004) Mu rhythm
modulation during observation of an object-directed grasp. Brain Res Cogn
Brain Res 19: 195–201.
19. Rihs TA, Michel CM, Thut G (2007) Mechanisms of selective inhibition in
visual spatial attention are indexed by alpha-band EEG synchronization.
Eur J Neurosci 25: 603–610.
20. Gomarus HK, Althaus M, Wijers AA, Minderaa RB (2006) The effects of
memory load and stimulus relevance on the EEG during a visual selective
memory search task: An ERP and ERD/ERS study. Clin Neurophysiol 117:
871–884.
21. Klimesch W (1996) Memory processes, brain oscillations and EEG synchroni-
zation. Int J Psychophysiol 24: 61–100.
22. Thut G, Nietzel A, Brandt SA, Pascual-Leone A (2006) Alpha-band
electroencephalographic activity over occipital cortex indexes visuospatial
attention bias and predicts visual target detection. J Neurosci 26: 9494–9502.
23. Kilner JM, Marchant JL, Frith CD (2009) Relationship between Activity in
Human Primary Motor Cortex during Action Observation and the MirrorNeuron System. PLoS ONE 4(3): e4925. doi:10.1371/journal.pone.0004925.
24. Rizzolatti G (2005) The mirror neuron system and imitation. In: Hurley S,
Chater N, eds. Perspectives on Imitation: From Neuroscience to Social Science.Cambridge: MIT Press. pp 55–76.
25. Byrne RW, Russon AE (1998) Learning by imitation: a hierarchical approach.Behav Brain Sci 21: 667–684.
26. Byrne RW (2005) Detecting, Understanding, and Explaining Imitation by
Animals. In: Hurley S, Chater N, eds. Perspectives on Imitation: FromNeuroscience to Social Science. Cambridge: MIT Press. pp 225–242.
27. Koski LM, Wohlschlager A, Bekkering H, Woods RP, Dubeau MC, et al. (2002)
Modulation of motor and premotor activity during imitation of target-directedactions. Cereb Cortex 12: 847–855.
28. Iacoboni M, Woods RP, Brass M, Bekkering H, Mazziotta JC, et al. (1999)
Cortical mechanisms of human imitation. Science 286: 2526–2528.
29. Aziz-Zadeh L, Koski LM, Zaidel E, Mazziotta JC, Iacoboni M (2006)Lateralization of the human mirror neuron system. J Neurosci 26: 2964–2970.
30. Babiloni C, Babiloni F, Carducci F, Cincotti F, Cocozza G, et al. (2002) Human
cortical electroencephalography (EEG) rhythms during the observation of simpleaimless movements: a high-resolution EEG study. Neuroimage 17: 559–572.
31. van Schie HT, Mars RB, Coles MGH, Bekkering H (2004) Modulation of
activity in medial frontal and motor cortices during error observation. NatNeurosci 7: 549–554.
32. van Schie HT, KoelewijnThomas, Jensen O, Oostenveld R, Maris E, et al.
(2008) Evidence for fast, low-level motor resonance to action observation: AnMEG study. Soc Neurosci 3: 213–228.
33. KoelewijnThomas, van Schie HT, Bekkering H, Oostenveld R, Jensen O (2008)
Motor-cortical beta oscillations are modulated by correctness of observed action.Neuroimage 40: 767–775.
34. Johnson SH, Rotte M, Grafton ST, Hinrichs H, Gazzaniga MS, et al. (2002)
Selective activation of a parietofrontal circuit during implicitly imaginedprehension. Neuroimage 17: 1693–1704.
35. Hummel F, Gerloff C (2005) Larger interregional synchrony is associated with
greater behavioral success in a complex sensory integration task in humans.Cereb Cortex 15: 670–678.
36. Classen J, Gerloff C, Honda M, Hallett M (1998) Integrative visuomotor
behavior is associated with interregionally coherent oscillations in the humanbrain. J Neurophysiol 79: 1567–1573.
37. Smyth MM, Pendleton LR (1990) Space and movement in working memory.
Q J Exp Psychol 42: 291–304.
38. Pfurtscheller G, Neuper C (1997) Motor imagery activates primary sensorimotorarea in humans. Neurosci Lett 239: 65–68.
39. Neuper C, Pfurtscheller G (2001) Event-related dynamics of cortical rhythms:
frequency-specific features and functional correlates. Int J Psychophysiol 43:41–58.
40. Muthukumaraswamy SD, Johnson BW, Gaetz WC, Cheyne DO (2006) Neuralprocessing of observed oro-facial movements reflects multiple action encoding
strategies in the human brain. Brain Res 1071: 105–112.
41. Rizzolatti G, Luppino G, Matelli M (1998) The organization of the corticalmotor system: new concepts. Electroen Clin Neuro 106: 283–296.
42. Rizzolatti G, Luppino G (2001) The cortical motor system. Neuron 31: 889–901.
43. Hamilton AF, Grafton ST (2006) Goal representation in human anterior
intraparietal sulcus. J Neurosci 26: 1133–1137.
44. Newman-Norlund R, van Schie HT, van Hoek ME, Cuijpers RH, Bekkering H(2010) The role of inferior frontal and parietal areas in differentiating meaningful
and meaningless object-directed actions. Brain Research 1315: 63–74.
45. Fogassi L, Ferrari PF, Gesierich B, Rozzi S, Chersi F, et al. (2005) Parietal lobe:from action organization to intention understanding. Science 308: 662–667.
46. Annett M (1970) A classification of hand preference by association analysis.
Brit J Psycho 61: 303–321.
Observational Learning
PLoS ONE | www.plosone.org 10 December 2010 | Volume 5 | Issue 12 | e14482

47. Pivek RT, Broughton RJ, Coppola R, Davidson RJ, Fox N, et al. (1993)
Guidelines for the recording and quantative analysis of electroencephalographic
activity in research contexts. Psychophysiology 30: 547–558.
48. Pfurtscheller G, Lopes da Silva FH (1999) Event-related EEG/MEG
synchronization and desynchronization: basic principles. Clin Neurophysiol
110: 1842–1857.
49. Dale AM, Sereno MI (1993) Improved localization of cortical activity by
combining EEG and MEG with MRI cortical surface reconstruction: A linearapproach. J Cogn Neurosci 5: 162–176.
50. Andrew C, Pfurtscheller G (1996) Dependence of coherence measurements on
EEG derivation type. Med Biol Eng Comput 34: 232–238.51. Kayser J, Tenke CE (2006) Principal components analysis of Laplacian
waveforms as a generic method for identifying ERP generator patterns: I.Evaluation with auditory oddball tasks. Clin Neurophysiol 117: 348–368.
Observational Learning
PLoS ONE | www.plosone.org 11 December 2010 | Volume 5 | Issue 12 | e14482
Top Related
Copyright © 2022 FDOKUMEN

