







Bahasa
Halaman
Hukum


www.elsevier.com/locate/geoderma
Geoderma 132 (2
Morphological changes in Camponotus punctulatus
(Mayr) anthills of different ages
Norma B. Gorositoa,*, Pierre Curmib,1, Vincent Hallairec,2,
Patricia J. Folgarait a, Patrick M. Lavelled,3
aCentro de Estudios e Investigaciones (CEI), Universidad Nacional de Quilmes, Roque Saenz Pena 180,
B1876BXD Bernal, Buenos Aires, ArgentinabEtablissement National d’Enseignement Superieur Agronomique de Dijon (ENESAD), Equipe Milieu Physique et Environnement,
BP87999, 26 Bd du Dr Petitjean, 21079 Dijon Cedex, FrancecInstitut National de la Recherche Agronomique (INRA), Unite Sol et Agronomie, 65 rue St Brieuc, 35042 Rennes Cedex, France
dLaboratoire d’Ecologie de Sols Tropicaux (LEST)-Institut de Recherche pour le Developement (IRD),
32 rue Henri de Varagnat, F-93143 Bondy Cedex, France
Received 14 May 2004; received in revised form 16 May 2005; accepted 17 May 2005
Available online 23 June 2005
Abstract
In north-eastern Argentina, Camponotus punctulatus builds large numbers of big and coherent anthills after abandon-
ment of rice cultivation. These anthills easily reach 1 m in height and 2 m in diameter, and a density of 1800 nests ha�1.
We studied the internal morphology of C. punctulatus aged anthills of 4, 6 and 15 years, respectively, by describing and
quantifying, meso- and macroporosity of undisturbed soil samples using image analysis. Four different parts were
distinguished on these cone-shaped anthills: the loose granular cortex, the summit, the core of the anthill and the base
of the anthill at ground level. The percentage of macroporosity in anthills significantly differed between the parts of the
anthill, but changed little with age except for the 15 year old anthill that had increased percentages of mesopores and
macropores with rounded and irregular shapes. Walls of the chambers and galleries were highly compacted, highlighting
intense ant activity in the anthills, although it was localised mainly in the upper central part. After 6 years the anthills
became surrounded by a discontinuous peripheral depression, which likely affects water drainage and infiltration. In 15
0016-7061/$ - s
doi:10.1016/j.ge
* Correspondi
E-mail addr
[email protected] Fax: +33 3 82 Fax: +33 2 23 Fax: +33 1 4
006) 249–260
ee front matter D 2005 Elsevier B.V. All rights reserved.
oderma.2005.05.010
ng author. Fax: +54 11 4365 7182.
esses: [email protected] (N.B. Gorosito), [email protected] (P. Curmi), [email protected] (V. Hallaire),
edu.ar (P.J. Folgarait), [email protected] (P.M. Lavelle).
0 77 25 00.
3 48 54 30.
8 02 59 70.

N.B. Gorosito et al. / Geoderma 132 (2006) 249–260250
year old anthills the lateral depressions became a continuous ditch where water accumulates giving place to a constant
wetted zone inside the anthill. Our results support the previous consideration of C. punctulatus as an ecosystem engineer,
although in this case related to the changes produced on the soil surrounding the anthills which may affect the survival and
distribution of other soil organisms.
D 2005 Elsevier B.V. All rights reserved.
Keywords: Anthills; Anthills’ age; Camponotus punctulatus; Image analysis; Porosity
1. Introduction
Ants, throughout their activity, are acknowledged
to contribute to the structure and functioning of soils
(Baxter and Hole, 1967; Cox et al., 1992; Wang et al.,
1995; Folgarait, 1998). Ants influence soil properties
through the construction of their nests by changing
physical characteristics, such as porosity and infiltra-
tion (Wang et al., 1995). In addition, ant nests are
associated with high levels of nutrients and organic
matter (Lobry de Bruyn and Conacher, 1990; Folgar-
ait, 1998; Kristiansen and Amelung, 2001; Cammer-
aat et al., 2002).
From a pedological point of view ants can build
two types of nests (Paton et al., 1995). Type I, less
conspicuous in the landscape, are crater-shaped,
small in diameter and height; soil material is simply
deposited on the surface and the nests are highly
susceptible to erosion. Type II are larger, coherent
epigeic nests, often cemented, sometimes covered by
vegetation. These are very persistent through time
that may significantly affect the spatial heterogeneity
of the soil surface.
Camponotus punctulatus (Mayr) is a native ant
species widely distributed in Argentina from the
northern border (also present in Brazil and Uruguay)
to the latitude 448S (Bonetto et al., 1961; Kusnezov,
1951). This ant species may build small nests below
ground under the grass tussocks or large conspicuous
anthills (Kusnezov, 1951). In Northeast Argentina we
have found that the disturbances produced by agricul-
tural practices lead C. punctulatus to construct nests
of type II (Folgarait, 1998). This is particularly obvi-
ous in the fallow periods (4 to 15 years) following
paddy rice cultivation where densities of type II nests
may reach 1800 anthills ha�1 depending on the age of
abandonment of the rice fields (Folgarait et al., 2002).
In natural grasslands, we have also observed the
presence of anthills of type II, but the density was
much lower at 23 anthills ha�1. These cone-shaped
anthills, covered by vegetation, are extremely hard
when dry, and they can reach ~1 m in height and
~2 m in diameter (Folgarait et al., 2000). C. punctu-
latus does not present a direct hazard to the crop since
this ant is not herbivorous, feeding mostly on honey-
dew and other insects (Gorosito et al., 1997). Indeed,
C. punctulatus activity promotes specific plant com-
munities on the anthills, and anthill soil has a greater
fertility than the surrounding soil (Folgarait et al.,
2002). Activities within the anthill of C. punctulatus
also favour the development of bacteria, measured as
dehydrogenase activity (Gonzalez Polo et al., 2004),
and other mesofauna such as mites, and speed up the
mineralization of phosphorus inside the anthill (Paris
et al., 2001). Farmers, however, find these anthills
inconvenient and expensive to remove when returning
the area to cropping. Horses and livestock get injured
when stepping into the ditches that form around the
anthills and no tillage can be done if these structures
remain intact.
A research programme has been developed to
understand the reasons for anthill proliferation in
abandoned rice fields, their consequences, and to
combat the inconvenience they cause to the farmers
when returning the fields to cropping (Folgarait et
al., 2003). As part of this programme we have
studied the internal organization of the anthills of
C. punctulatus and its dynamics. We tested the
hypothesis that there were differences in the internal
morphology of anthills of different ages. A macro-
morphological and micromorphological characterisa-
tion of anthills allowed us to identify the main
fabrics and their genesis, using a qualitative ap-
proach and a quantitative estimation of porosity by
image analysis. This study was conducted on anthills
of different ages (4, 6 and 15 year old anthills)
representing a chronosequence of different fallow
periods used by farmers.

Table 1
Pore classification according to their size and shape for both scales
of resolution
Area (Am2) Shape classes
Size
classes
R E I
Ieb5 5b Ieb10 IeN10
Mesopores Areab1 d 103 1 R1 E1 I1
1 d 103b
areab1 d 1052 R2 E2 I2
AreaN1 d 105 3 R3 E3 I3
Macropores Areab1 d 106 4 R4 E4 I4
1 d 106b
areab1 d 1075 R5 E5 I5
AreaN1 d 107 6 R6 E6 I6
The shape was defined according to the Elongation Index (Ie). R:
rounded pores, E: elongated pores, I: irregular pores.
N.B. Gorosito et al. / Geoderma 132 (2006) 249–260 251
2. Materials and methods
2.1. Study site
Field work was carried out at the Aguaceritos farm,
~60 km north of Mercedes, Corrientes Province, in
Argentina (298S, 588W). The climate is subtropical,
without a definite dry season. Mean annual precipita-
tion is 1270 mm and the mean annual temperature is
20.1 8C (Fernandez et al., 1993). Soils are alfisols
(Soil Survey Staff, 1992), which have developed on a
several meters thick clayey saprolite from calcareous
sandstones. After rice cultivation, large compact clods
occurred in the ploughed layer and a distinct plough
pan was created by mechanical tillage at 15 cm depth
(see Lesturgez, 2000 for soil details). After 2 years of
fallow, the whole ploughed horizon is homogeneously
compacted due to cattle trampling and cannot be
distinguished from the plough pan.
2.2. Sampling
2.2.1. Anthills selection for general description
After the last rice harvest the land enters the fallow
period. During this period C. punctulatus, which were
not present in the rice field, re-colonised the aban-
doned rice field apparently from the natural grassland.
All the anthills of a given plot are therefore mostly
of the same age starting after the last harvest. In
January 2000, we chose one representative C. punc-
tulatus anthill in three rice fields abandoned (fallow
period) for 4, 6, and 15 years, respectively. The
dimensions of the chosen anthill were almost the
average size observed in rice plots (Folgarait et al.,
2000).
We used a mechanical shovel for cutting the anthills
vertically down to ca. 2 m depth in order to observe
their internal organization. Diameters of galleries were
measured at different depths on the anthill sections.
2.2.2. Micromorphological characterisation
Twelve undisturbed blocks of soil were taken in
order to have a representative sampling of the differ-
ent parts found inside anthills of different ages: in
each of three different anthills aged 4, 6 and 16 years,
respectively, 4 blocks (16 cm height�9 cm width�5
cm depth) were taken from the different parts regis-
tered within each anthill.
In order to avoid structural modifications, all the
blocks were impregnated with a polyester resin after
replacing water by acetone (Ringrose-Voase, 1996). A
fluorescent dye was also added to allow us to see the
macropore and mesopore space when illuminated with
UV light. After impregnation, three vertical sections
of 16�9 cm were prepared for each block, and they
were exposed to UV light to quantify the soil porosity
by computerised image analysis (Hallaire and Curmi,
1994).
Image acquisition was carried out at two levels of
resolutions (100 Am/pixel and 1.85 Am/pixel) on each
soil section. The first resolution, allowed us to see the
macropores whereas the second one was used to
quantify mesopores. Three images (768�576 pixels)
per section (i.e. nine per block) were taken for the
coarse resolution. Ten images (1798�1438 pixels)
were taken at random for the second scale. Addition-
ally, we took six images to evaluate the compaction
around the galleries (1798�1438 pixels). All images
were processed with the software Optimas 6.51 (Opti-
mas 6.51 Media Cybernetics).
Pores were classified into morphological classes
taking into account the area of their section and
their shape (Hallaire et al., 1997). Six size classes
were established according to their area. Pore shape
was determined using the Elongation Index, Ie (pe-
rimeter2/area) (Coster and Chermant, 1985), and three
classes were distinguished: rounded (R), elongated (E)
and irregular pores (I). Therefore, 18 morphological
classes resulted from the combination of different
shapes and sizes (Table 1).

N.B. Gorosito et al. / Geoderma 132 (2006) 249–260252
2.3. Statistical analysis
In the absence of normality and homoscedascity of
data, non-parametric analyses were performed (Stat-
view software, SAS Institute, 1992–1998). To com-
pare more than two variables we used the Kruskall–
Wallis test with a posteriori contrasts adjusted with
S
C
Old ploughed ho
Cortex
0.30 m
0 m
0.35 m
S
C
B
Old ploughed hor
Sub-vert
0.30 m
0 m
0.70 mCortex
S
C
B
Cortex
Old ploughed ho0.30 m
0 m
0.60 m
Wet
0 m
Fig. 1. Scheme of the internal organization of the 4, 6 and 15 year old anth
former ploughed horizon. Soil samples where taken where each letter is l
Bonferroni to keep an alpha level=0.05 and for
paired comparisons we chose Mann–Whitney test
(Daniel, 1990). For all the comparisons related with
the different parts (summit, core, base) within each
anthill, the number of images was 9 and 10 for the
first and second scale of observation, respectively.
Compaction around the galleries was first compared
4 yrs
6 yrs
rizon (P)
izon (P)
15 yrs
ical gallery
Continuous Peripheral depression
Discontinuous
Peripheral depression
Sub-vertical galleryrizon (P)
ted zone
1.50 m
ills. S: summit; C: core; B: base of the epigeic part of the anthill; P:
ocated.

a
6.4 mm
b
6.4 mm
N.B. Gorosito et al. / Geoderma 132 (2006) 249–260 253
within the different parts found inside each anthill,
then among the anthills, and finally we compared the
porosity around the galleries with the mesoporosity
found in the whole anthill (the parts altogether). We
considered a level of significance of pb0.05 for all
comparisons.
e
6.4 mm
d6.4 mm
6.4 mm
f
c
6.4 mm
Fig. 2. Different types of pores and structures found inside the
anthills. All images were 768�576 pixels in size. (a) Rounded
pore, (b) elongated pore, (c) interconnected pores, (d) irregular pore
filled with angular microaggregates, (e) traces of ant mandibles, (f
gallery wall compaction.
3. Results
3.1. Morphological description
A vertical cut of the anthills allowed us to identify
four different parts based on the abundance and mor-
phology of the macropores (Fig. 1): the coherent
summit of the anthill (S), the core (C) and the base
at the ground level of the anthill (B), and finally below
ground the remnants of the former ploughed horizon
(P). A loose cortex of fine granular and irregular
aggregates covered the outside upper part of the ant-
hill. We did not make a micromorphological analysis
of this area due to its lack of cohesion.
Observation of each area inside the anthills led us
to distinguish three types of macropores: rounded,
elongated and irregular (Fig. 2). The rounded pores
(Fig. 2a) corresponded to chambers usually used for
brood caring, while the elongated ones represented
connections among chambers and galleries (Fig.
2b,c) (used for carrying soil particles, brood or
food supplies). Finally, the irregular pores resulted
from the filling of chambers and galleries with
angular microaggregates displaced by ants (Fig.
2d). In some sections we have also observed cham-
bers or galleries filled with rounded earthworms
faecal pellets and oval termites ones. Using a field
magnifier, we observed prints of ant mandibles on
the walls of the galleries and chambers (Fig. 2e),
and also soil compaction around chambers and gal-
leries (Fig. 2f).
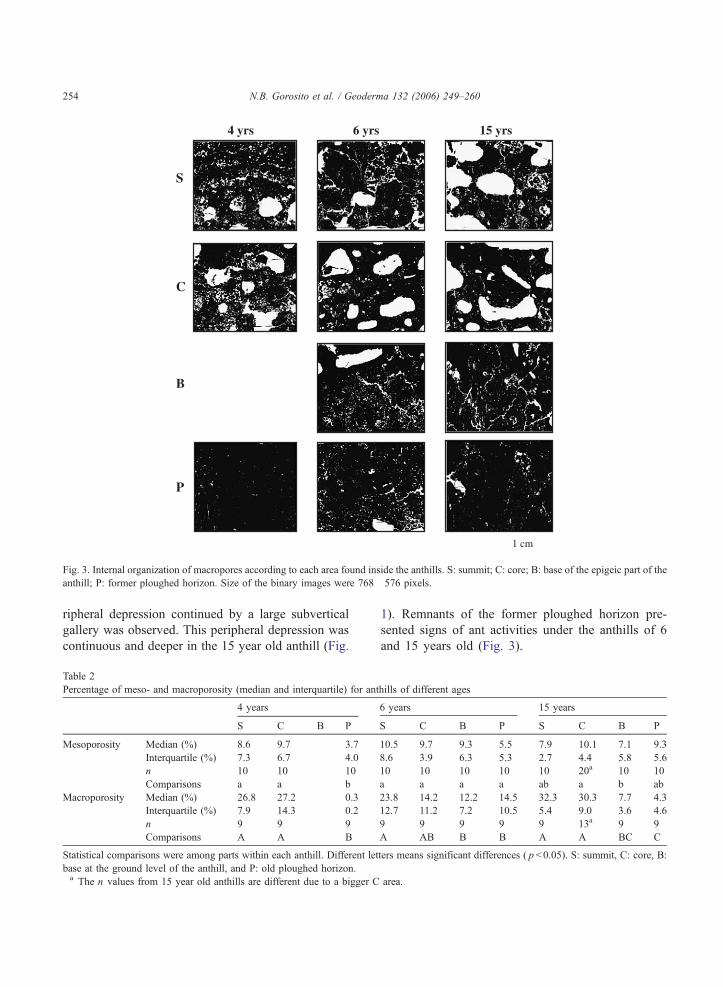
The 4 year old anthill, 0.35 m in height and 0.90 m
in diameter (Vol.: 0.09 m3) (Fig. 1), had the thickest
cortex (~10 cm) of fine aggregates (Fig. 1). The
summit was the most friable as shown by the loose
assemblage of small aggregates (Fig. 3). The mean
diameter of the galleries was 1.03 cm (n =16). The 4
year old anthill did not yet have a base as observed in
the older anthills and the core was very hard when dry.
Galleries and chambers were the most abundant in the
)
core (Fig. 3). Remnants of the former ploughed hori-
zon were found under the anthill at the same level as
the ploughed horizon outside the anthill. The
ploughed horizon was the most compacted under the
anthill.
The 6 year old anthill was 0.60 m in height and
1.50 m in diameter (Vol.: 0.17 m3) (Fig. 1) and the
surface cortex was thinner than in the younger ant-
hill. At this stage, the mean diameter of galleries was
0.87 cm (n =34). The 15 year old anthill was 0.70 m
in height and 1.50 m in diameter (Vol.: 0.54 m3) and
had the thinnest cortex (2–3 cm). The hardness of the
anthill was higher in comparison to the younger
ones. At this stage, mean diameter of galleries was
0.76 cm (n =21). The size of the core changed with
age, with a clear enlargement in the 15 year old
anthill (Fig. 1). The base part was present in the 6
year old anthill, and it was more compacted in
comparison to the summit and the core. Finally, the
compaction at the base of the anthill was most
important in the 15 year old anthill. Around the
base of the 6 year old anthill, a discontinuous pe-
6 yrs4 yrs
S
15 yrs
P
C
B
1 cm
Fig. 3. Internal organization of macropores according to each area found inside the anthills. S: summit; C: core; B: base of the epigeic part of the
anthill; P: former ploughed horizon. Size of the binary images were 768�576 pixels.
N.B. Gorosito et al. / Geoderma 132 (2006) 249–260254
ripheral depression continued by a large subvertical
gallery was observed. This peripheral depression was
continuous and deeper in the 15 year old anthill (Fig.
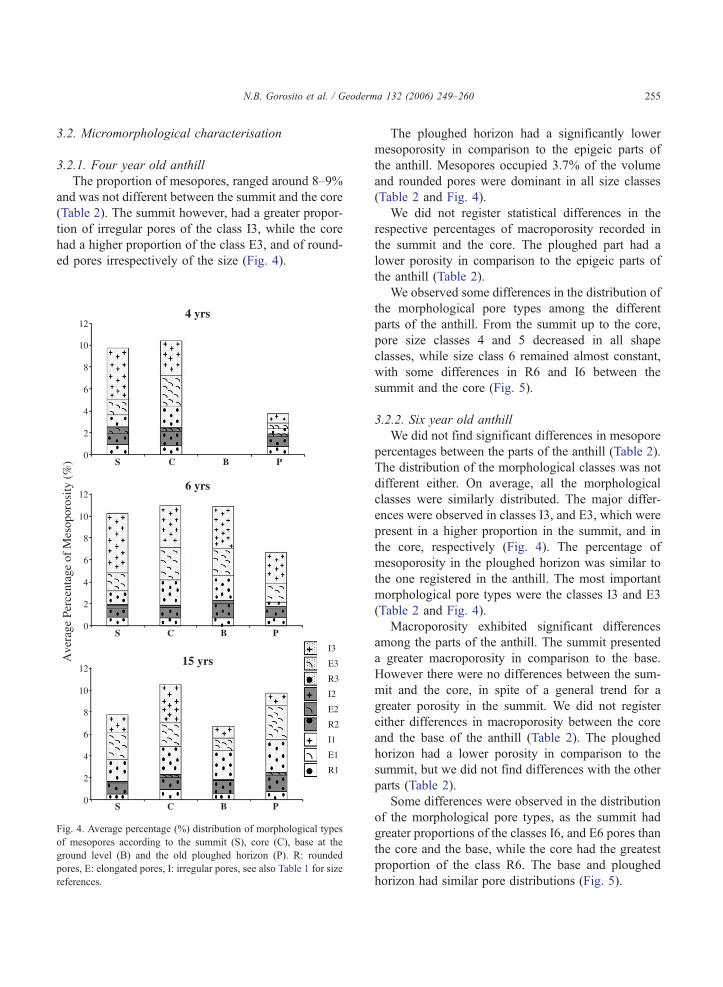
Table 2
Percentage of meso- and macroporosity (median and interquartile) for ant
4 years
S C B P
Mesoporosity Median (%) 8.6 9.7 3.7
Interquartile (%) 7.3 6.7 4.0
n 10 10 10
Comparisons a a b
Macroporosity Median (%) 26.8 27.2 0.3
Interquartile (%) 7.9 14.3 0.2
n 9 9 9
Comparisons A A B
Statistical comparisons were among parts within each anthill. Different let
base at the ground level of the anthill, and P: old ploughed horizon.a The n values from 15 year old anthills are different due to a bigger C
1). Remnants of the former ploughed horizon pre-
sented signs of ant activities under the anthills of 6
and 15 years old (Fig. 3).
hills of different ages
6 years 15 years
S C B P S C B P
10.5 9.7 9.3 5.5 7.9 10.1 7.1 9.3
8.6 3.9 6.3 5.3 2.7 4.4 5.8 5.6
10 10 10 10 10 20a 10 10
a a a a ab a b ab
23.8 14.2 12.2 14.5 32.3 30.3 7.7 4.3
12.7 11.2 7.2 10.5 5.4 9.0 3.6 4.6
9 9 9 9 9 13a 9 9
A AB B B A A BC C
ters means significant differences ( p b0.05). S: summit, C: core, B:
area.
N.B. Gorosito et al. / Geoderma 132 (2006) 249–260 255
3.2. Micromorphological characterisation
3.2.1. Four year old anthill
The proportion of mesopores, ranged around 8–9%
and was not different between the summit and the core
(Table 2). The summit however, had a greater propor-
tion of irregular pores of the class I3, while the core
had a higher proportion of the class E3, and of round-
ed pores irrespectively of the size (Fig. 4).
4 yrs
0
2
4
6
8
10
12
S B P
6 yrs
0
2
4
6
8
10
12
15 yrs
0
2
4
6
8
10
12
Ave
rage
Per
cent
age
of M
esop
oros
ity (
%)
I3
E3
R3
I2
E2
R2
I1
E1
R1
C
S B PC
S B PC
Fig. 4. Average percentage (%) distribution of morphological types
of mesopores according to the summit (S), core (C), base at the
ground level (B) and the old ploughed horizon (P). R: rounded
pores, E: elongated pores, I: irregular pores, see also Table 1 for size
references.
The ploughed horizon had a significantly lower
mesoporosity in comparison to the epigeic parts of
the anthill. Mesopores occupied 3.7% of the volume
and rounded pores were dominant in all size classes
(Table 2 and Fig. 4).
We did not register statistical differences in the
respective percentages of macroporosity recorded in
the summit and the core. The ploughed part had a
lower porosity in comparison to the epigeic parts of
the anthill (Table 2).
We observed some differences in the distribution of
the morphological pore types among the different
parts of the anthill. From the summit up to the core,
pore size classes 4 and 5 decreased in all shape
classes, while size class 6 remained almost constant,
with some differences in R6 and I6 between the
summit and the core (Fig. 5).
3.2.2. Six year old anthill
We did not find significant differences in mesopore
percentages between the parts of the anthill (Table 2).
The distribution of the morphological classes was not
different either. On average, all the morphological
classes were similarly distributed. The major differ-
ences were observed in classes I3, and E3, which were
present in a higher proportion in the summit, and in
the core, respectively (Fig. 4). The percentage of
mesoporosity in the ploughed horizon was similar to
the one registered in the anthill. The most important
morphological pore types were the classes I3 and E3
(Table 2 and Fig. 4).
Macroporosity exhibited significant differences
among the parts of the anthill. The summit presented
a greater macroporosity in comparison to the base.
However there were no differences between the sum-
mit and the core, in spite of a general trend for a
greater porosity in the summit. We did not register
either differences in macroporosity between the core
and the base of the anthill (Table 2). The ploughed
horizon had a lower porosity in comparison to the
summit, but we did not find differences with the other
parts (Table 2).
Some differences were observed in the distribution
of the morphological pore types, as the summit had
greater proportions of the classes I6, and E6 pores than
the core and the base, while the core had the greatest
proportion of the class R6. The base and ploughed
horizon had similar pore distributions (Fig. 5).
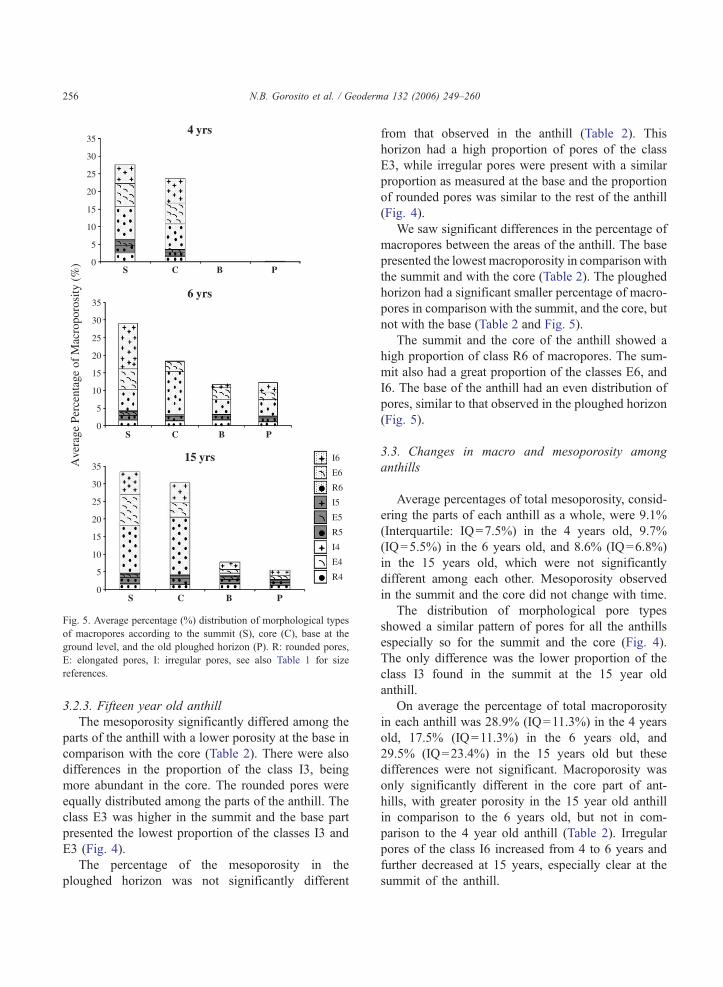
4 yrs
0
5
10
15
20
25
30
35
S
6 yrs
0
5
10
15
20
25
30
35
15 yrs
0
5
10
15
20
25
30
35I6
E6
R6
I5
E5
R5
I4
E4
R4
Ave
rage
Per
cent
age
of M
acro
poro
sity
(%
)
C B P
S C B P
S C B P
Fig. 5. Average percentage (%) distribution of morphological types
of macropores according to the summit (S), core (C), base at the
ground level, and the old ploughed horizon (P). R: rounded pores,
E: elongated pores, I: irregular pores, see also Table 1 for size
references.
N.B. Gorosito et al. / Geoderma 132 (2006) 249–260256
3.2.3. Fifteen year old anthill
The mesoporosity significantly differed among the
parts of the anthill with a lower porosity at the base in
comparison with the core (Table 2). There were also
differences in the proportion of the class I3, being
more abundant in the core. The rounded pores were
equally distributed among the parts of the anthill. The
class E3 was higher in the summit and the base part
presented the lowest proportion of the classes I3 and
E3 (Fig. 4).
The percentage of the mesoporosity in the
ploughed horizon was not significantly different
from that observed in the anthill (Table 2). This
horizon had a high proportion of pores of the class
E3, while irregular pores were present with a similar
proportion as measured at the base and the proportion
of rounded pores was similar to the rest of the anthill
(Fig. 4).
We saw significant differences in the percentage of
macropores between the areas of the anthill. The base
presented the lowest macroporosity in comparison with
the summit and with the core (Table 2). The ploughed
horizon had a significant smaller percentage of macro-
pores in comparison with the summit, and the core, but
not with the base (Table 2 and Fig. 5).
The summit and the core of the anthill showed a
high proportion of class R6 of macropores. The sum-
mit also had a great proportion of the classes E6, and
I6. The base of the anthill had an even distribution of
pores, similar to that observed in the ploughed horizon
(Fig. 5).
3.3. Changes in macro and mesoporosity among
anthills
Average percentages of total mesoporosity, consid-
ering the parts of each anthill as a whole, were 9.1%
(Interquartile: IQ=7.5%) in the 4 years old, 9.7%
(IQ=5.5%) in the 6 years old, and 8.6% (IQ=6.8%)
in the 15 years old, which were not significantly
different among each other. Mesoporosity observed
in the summit and the core did not change with time.
The distribution of morphological pore types
showed a similar pattern of pores for all the anthills
especially so for the summit and the core (Fig. 4).
The only difference was the lower proportion of the
class I3 found in the summit at the 15 year old
anthill.
On average the percentage of total macroporosity
in each anthill was 28.9% (IQ=11.3%) in the 4 years
old, 17.5% (IQ=11.3%) in the 6 years old, and
29.5% (IQ=23.4%) in the 15 years old but these
differences were not significant. Macroporosity was
only significantly different in the core part of ant-
hills, with greater porosity in the 15 year old anthill
in comparison to the 6 years old, but not in com-
parison to the 4 year old anthill (Table 2). Irregular
pores of the class I6 increased from 4 to 6 years and
further decreased at 15 years, especially clear at the
summit of the anthill.

Table 3
Statistical comparisons between the percentage of mesoporosity
around the galleries (G) and the percentage of mesoporosity in
the matrix of each anthill (epigeic part of the anthill)
Age Area Median (%) Interquartile (%) N p
4 years G 5.9 3.6 12 p =0.067
M 9.1 7.5 20
6 years G 4.8 3.4 18 p =0.001
M 9.7 5.4 30
15 years G 5.8 2.9 12 p =0.013
M 8.6 4.2 20
Level of significance p b0.05.
N.B. Gorosito et al. / Geoderma 132 (2006) 249–260 257
Significant changes were observed, however, for
macropores in the ploughed horizon. Below the 4 year
old anthill the ploughed horizon did have almost no
macropores and had a low percentage of mesopores.
After 15 years macroporosity was still low, and the
mesoporosity achieved values similar to that ones
found at the base of the anthill (Figs. 4 and 5).
3.4. Comparison between the porosity around the
galleries and the mesoporosity of the anthills
Mesoporosity around the galleries showed similar
patterns among the parts within the anthills (4 years
old, p =0.83, n =6; 6 years old, p =0.45, n =6; 15 years
old, p =0.87, n =6) and among the anthills of different
ages ( p =0.31, n =18). However, when comparing
mesoporosity around the galleries with the mesopor-
osity present in the epigeic part of the anthill (matrix),
the mesoporosity around the galleries was significantly
smaller in all cases, although this trend was not sig-
nificantly different in the 4 year old anthill (Table 3).
4. Discussion
4.1. External changes to the anthills
The thickness of the loose cortex, that covers the
anthills, is directly associated to the ant building activ-
ity, decreasing with the age of the anthill and disappear-
ing when dead. Once compacted and incorporated into
the anthill, elements from the loose cortex become
highly coherent giving to the anthill a persistence that
is a characteristic of anthills of type II described by
Paton et al. (1995). Additionally, the cortex covering
the anthills likely plays an important role in absorbing
the impact of the rain drops and in improving the water
infiltration inside the anthills (Lobry de Bruyn and
Conacher, 1994).
After 6 years, we have observed the development of
peripheral depressions of several centimetres depth
around some parts of the anthills, and after 15 years,
the depressions became a continuous ditch around the
anthill. The presence of these ditches in older anthills
had been formerly interpreted as a consequence of their
own weight since an anthill of 7 years old exerts a
pressure of 2.22 kPa (Folgarait, unpublished data).
However, after analysing the structure of the anthills
we alternatively propose that as the remnants of the
former ploughed horizon are observed below the anthill
at the same level than outside, no sinking effect of the
anthill due to its own weight seems to have occurred.
On the other hand, the peripheral depression around the
anthills could be the result of a combination of the
specific physico-hydric behaviour of the soil at the
periphery of the anthill and the effects of livestock.
Water runoff along the anthill induces a wetter zone
around them which had been softened by ant activities.
Therefore ditches may be due to cattle trampling in this
peripheral wetter and softer zone.
4.2. Internal organization of the anthills
In anthills aged 4 years, the summit and the core
parts of the anthill were completely developed, but the
compacted base was still absent. We assume that this
is a consequence of the relative youth of the anthill,
which did not have enough time for differentiating the
areas described. However, in the other two anthills the
base was clearly developed. The porosity in this area
was smaller than the one registered in the summit for
the 6 year old anthill and also smaller than that found
in the core of the 15 year old anthill.
At the base of the 6 and 15 year old anthills, we
observed few active galleries, which were normally
found in the upper part of the anthill (summit and
core) at the three ages. This pattern of gallery distribu-
tion was also found in Lasius neoniger (Wang et al.,
1995). We state that the formation of the base through
time is the result of filling galleries and chambers by C.
punctulatus ants. In this way, galleries and chambers
belonging to the core, once filled, become part of the
base. The observed increased in mesoporosity further
supported this hypothesis. We observed an increase in

N.B. Gorosito et al. / Geoderma 132 (2006) 249–260258
the proportion of irregular pores (especially the larger
ones) from 4 years to 6 years old, and then a decrease at
15 years old, especially so, for the summit of the ant-
hills across ages. This change in the proportion of
irregular pores is a clear indication that ant would fill
with particles of soil part of the macropores. We hy-
pothesize that ants filled the galleries and chambers in
order to stabilize the anthill.
4.3. Stabilization of the mound by wall gallery’s
compaction
The micromorphological analysis points out that
the soil particles around the galleries were more
compacted than at other parts of the mound (matrix).
The soil particles were closer among each other
producing a reduction in the porosity. Under a mag-
nifier, we observed on the walls of the chambers, the
imprinting of ant mandibles, which ensure to us the
origin of the arrangement of the soil particles that
produce the compaction (see Fig. 2e,f). This com-
paction suggests that the ants were trying to stabilise
the walls of the galleries, necessary to resist the
increasing weight of the soil that is being accumu-
lated while the anthill is growing. In fact, many
myrmecologists have found behaviours related to
wall galleries stabilisation. They suggest that the
walls may have more resistance due to the re-orga-
nization of soil particles mixed with ceraceous sub-
stances, coming from the body of the ants
(Holldobler and Wilson, 1990; Wang et al., 1995).
This should be particularly the case as the anthills of
C. punctulatus are among the hardest observed
among ants (Folgarait, personal communication). A
similar behaviour has been observed in termites,
which made the wall chambers more resistant and
persistent through time by changing the mineralogy
of clay that is deposited for building the wall cham-
bers (Jouquet et al., 2002). It would be very inter-
esting to check for this kind of change in clay
mineralogy also for the anthills of C. punctulatus.
4.4. Changes in the compacted topsoil under ant
activity
The old ploughed horizon remained almost un-
modified below the 4 year old anthill. The macro-
pores were almost absent and there was a small
proportion of mesopores. In beginning a colony C.
punctulatus queens develop their nests at the base of
tussocks or rosettes, and most of the time at the
btaipasQ (name given locally for the ridges that sep-
arate parcels in the rice field) (personal observation).
We do not know how long it may take to the queen
to have a sufficient worker force to build the above-
ground nest. We think that C. punctulatus ants do
not excavate the old ploughed horizon until they are
a huge worker force. Meanwhile the ants may take
the soil from the upper few centimetres of the soil
surface (0–10 cm) and build aerated structures that
they consolidate by mixing with ceraceous sub-
stances coming from their body, and compacting
them with their mandibles and legs. Buhl et al.
(2004) have shown experimentally that the volume
of galleries excavated by the ant Mesor sancta Forel
was positively correlated with the number of work-
ers. On the other hand, at the beginning of the
anthill building, the ant C. punctulatus might not
need to dig a lot of new galleries, all the ants may
re-use the already excavated galleries. The tendency
for an individual to dig where other individuals have
previously dug has been shown in several ant spe-
cies (Hangarten, 1969; Rasse, 1999; Buhl et al.,
2004). After the statement of these hypotheses, we
conclude that C. punctulatus did not modify the old
ploughed horizon below the 4 year old anthill, which
was massive and compact after rice plantation and
cattle trampling during the fallow period. However
an increase in the macropores proportion through
time was clear from 4 to 6 years and then there
was a decrease at 15 years (Table 2), due to the
activity of filling the galleries and chambers as seen
by the increase in the proportion of mesopores,
especially the irregular ones from 4 to 15 years.
Despite some increment in soil porosity in the old
ploughed horizon, it was surprising to find this layer
almost unmodified 15 years after rice field abandon-
ment. The presence of the old ploughed horizon
gave us an idea of the degree of persistence of the
effects of some agricultural practices, which cannot
be rapidly modified by the biological activity. This
results contrasts with observations made in central
Amazonia by, e.g., Barros et al. (2001) who showed
restoration of soil physical properties of a degraded
pasture soil 1 year after exposure to the diverse soil
macrofauna of the original rainforest.

N.B. Gorosito et al. / Geoderma 132 (2006) 249–260 259
5. Conclusions
Anthills of C. punctulatus ants are very hard when
dry, and they persist through time even when aban-
doned. On average the percentage of soil porosity
(macroporosity) observed in the anthills was ~24.6%
in comparison to 8.3% found in the surrounding soil
(Gorosito et al., unpublished). The effect of C. punctu-
latus on soil physical properties was also observed
through the increase of porosity shown in the old
ploughed horizon as the anthills aged. Moreover, the
ditches surrounding old anthills also demonstrate an
enormous indirect effect on the surrounding soil. The
great compaction surrounding galleries and chambers
as well as the filling of those observed at the base of the
anthill additionally demonstrates the capacity of this
ant to consolidate the soil structure. Considering the
high density of anthills registered in abandoned rice
fields (1800 nests ha�1), their enormous size, high
resistance to erosion (we have seen 25 year old active
anthills), the ditches they produce, the enormousmove-
ment of soil particles from the soil to the surface, C.
punctulatus surely affects soil properties such as water
infiltration, soil aeration, and soil chemical properties,
and further affects life conditions for a large number of
organisms. Therefore, this study gives more evidence
to support the idea ofC. punctulatus as a soil ecosystem
engineer (Folgarait, 1998; Folgarait et al., 2002). These
effects are so important that they do not seem to be
naturally reversible in 10–30 years, the time period
covered by our observations. More detailed informa-
tion on the physical and chemical mechanisms in-
volved at the microaggregate scale should explain
how ants and termites manage to build long lasting
structures from apparently fragile soil materials.
Acknowledgements
We are indebted to the staff of the Agricultural
Experimental Station of INTA-Mercedes especially
to Rafael Pizzio for his continual support and logistic
help. We also thank the owners of the establishment
Aguaceritos, especially their managers J.C. Calfuan,
and P. Preliasco for their continuous help. We would
like also to thank Yannick Benard (INRA-Rennes)
for preparing the soil sections. This work was funded
by Institut de Recherche pour le Developement, IRD
(fellowship to NBG), by ECOS-SEPCyT (grant
A99B01 to PJF and PML), by Universidad Nacional
de Quilmes (grant 827-0201/99 and 0235 to PJF)
and IFS (grant C/2437-2 to PJF).
References
Barros, E., Curmi, P., Hallaire, V., Chauvel, A., Lavelle, P., 2001.
The role of macrofauna in the transformation and reversibility of
soil structure of an oxisol in the process of forest to pasture
conversion. Geoderma 100, 193–213.
Baxter, F.P., Hole, F.D., 1967. Ant (Formica cinerea) pedoturba-
tion in a prairie soil. Soil Science American Proceedings 31,
425–428.
Bonetto, A.A., Manzi, R., Pignalberi, C., 1961. Los btacuruesQ deCamponotus punctulatus. Physis 22, 217–224.
Buhl, J., Gautrais, J., Deneubourg, J.-L., Theraulaz, G., 2004. Nest
excavation in ants: group size effects on the size and structure of
tunneling networks. Naturwissenschaften 91, 602–606.
Cammeraat, L.H., Willot, S.J., Compton, S.G., Incoll, L.D., 2002.
The effects of ants’ nests on the physical, chemical and hydro-
logical properties of rangeland soil in semi-arid Spain. Geoderma
105, 1–20.
Coster, M., Chermant, J.C., 1985. Precis d’Analyse d’image.
Presses du C N.R.S., Paris, p. 321.
Cox, G.W., Mills, J.N., Ellis, B.A., 1992. Fire ants (Hymenoptera:
Formicidae) as major agents of landscape development. Austra-
lian Journal of Soil Research 22, 95–107.
Daniel, W.W., 1990. Applied Nonparametric Statistics, 2nd rd.
PWS-Kent, p. 507.
Fernandez, J.G., Benıtez, C.A., Royo Pallares, O., Pizzio, R., 1993.
Principales forrajeras nativas del medio este de la provincia de
Corrientes. Serie Tecnica, vol. 23. Estacion Experimental Agro-
pecuaria, Mercedes, p. 91.
Folgarait, P.J., 1998. Ant biodiversity and its relationship to eco-
system functioning: a review. Biodiversity and Conservation 6,
1293–1307.
Folgarait, P., Gorosito, N., Pizzio, R., Fernandez, J., 2000. Demog-
raphy of Camponotus punctulatus ants along a gradient of
agricultural disturbance and through time. Abstracts of XXI
International Congress of Entomology. Foz do Iguacu, Brazil.
20–26 August 2000.
Folgarait, P.J., Perelman, S., Gorosito, N., Pizzio, R., Fernandez, J.,
2002. Effects of Camponotus punctulatus ants on plant com-
munity composition and soil properties across land use histories.
Plant Ecology (ex Vegetatio) 163, 1–13.
Folgarait, P.J., Thomas, F., Desjardins, T., Grimaldi, M., Tayasu,
I., Curmi, P., Lavelle, P.M., 2003. Soil properties and the
macrofauna community in abandoned irrigated rice fields of
north-eastern Argentina. Biology and Fertility of Soils 38,
349–357.
Gonzalez Polo, M., Folgarait, P.J., Martınez, A., 2004. Evaluacion
estacional del efecto de los nidos de Camponotus punctulatus
sobre la biomasa y actividad microbiana en una pastura sub-
tropical de Argentina. Ecologıa Austral 14, 9–16.

N.B. Gorosito et al. / Geoderma 132 (2006) 249–260260
Gorosito, N., Zipeto, G., Folgarait, P.J., 1997. Las preferencias
alimenticias de Camponotus punctulatus en pasturas naturales
e implantadas. XVIII Reunion de la Asociacion Argentina de
Ecologıa. ASAE, Buenos Aires.
Hallaire, V., Curmi, P., 1994. Image analysis of pore space mor-
phology in soil sections, in relation to water movement. In:
Ringroase-Voase, A.J., Humphreys, G.S. (Eds.), Soil Micromor-
phology: Studies in Management and Genesis, Proceedings of
the Developments in Soil Science, vol. 22. Elsevier, Amster-
dam, p. 559–678.
Hallaire, V., Curmi, P., Widiatmaka, W., 1997. Morphologie de la
porosite et circulations preferentielles en sature: cas des horizons
d’un systeme pedologique armoricain. Etudes et Gestion des
Sols 4, 115–126.
Hangarten, W., 1969. Carbon dioxide, a releaser for digging behav-
ior in Solenopsis geminata (Hymenoptera: Formicidae). Psyche
76, 58–67.
Holldobler, B., Wilson, E.O., 1990. The Ants. Belknap Press,
Harvard, p. 732.
Jouquet, P., Mamou, L., Lepage, M., Velde, B., 2002. Effects of
termites on clay minerals in tropical soils: fungus-growing
termites as weathering agents. European Journal of Soil Science
53, 521–527.
Kristiansen, S.M., Amelung, W., 2001. Abandoned anthills of For-
mica polytecna and soil heterogeneity in a temperate deciduous
forest: morphology and organic matter composition. European
Journal of Soil Science 52, 355–363.
Kusnezov, N., 1951. El genero Camponotus en la Argentina. Acta
Zoologica Lilloana 12, 183–252.
Lesturgez, G., 2000. Proprietes des sols sous savane du Nord-Est de
l’Argentine. Impact de la riziculture. Memoire DEA National de
Science du Sol. Institut National Agronomique Paris-Grignon,
France.
Lobry de Bruyn, L.A., Conacher, A.J., 1990. The role of termites
and ants in soil modification: a review. Australian Journal of
Soil Research 28, 55–93.
Lobry de Bruyn, L.A., Conacher, A.J., 1994. The effect of ant
biopores on water infiltration in soils in undisturbed bushland
and in farmland in a semi-arid environment. Pedobiologia 38,
193–207.
Optimas Image Analysis Software, version 6.51. Media Cybernet-
ics, Silver Springs MD 20910, USA.
Paris, C.I., Folgarait, P.J., Garbagnoli, C., Somma, G., 2001. Biodi-
versidad de mesofauna y abundancia de nematodes en suelo de
tacurues: Como afectan la descomposicion de la pastura Setaria
sphacelata. XX Reunion Argentina de Ecologıa/X Reunion de
la Sociedad de Ecologıa de Chile, Bariloche, Argentina.
Paton, T.R., Humphreys, G.S., Mitchell, P.B., 1995. Soils—A
Global View. UCL Press, London, p. 213.
Rasse, P., 1999. Etude sur la regulation de la taille et sur la
structuration du nid souterrain de la fourmi Lasius niger. PhD
thesis, Universite Libre de Bruxelles, Brussels.
Ringrose-Voase, A.J., 1996. Measurement of soil macropore geom-
etry by image analysis of sections through impregnated soil.
Plant and Soil 183, 27–47.
Statview Software, SAS Fulfillment Services Departament, SAS
Campus Drive, Cary, NC 27513–2414.
Soil Survey Staff, 1992. Keys to soil taxonomy. (fifth edition).
SMSS Technical Monograph, vol. 19. Pocahontas Press, Inc.,
Blackburg, VA, USA, p. 541.
Wang, D., McSweeney, K., Lowery, B., Norman, J.M., 1995. Nest
structure of ant Lasius neoniger Emery and its implications to
soil modification. Geoderma 66, 259–272.
Top Related
Copyright © 2022 FDOKUMEN

