







Bahasa
Halaman
Hukum


Dominant-negative interference with defence signalling bytruncation mutations of the tomato Cf-9 disease resistancegene
Claire L. Barker1,†, Brett K. Baillie1, Kim E. Hammond-Kosack2,‡, Jonathan D. G. Jones2 and David A. Jones1,*
1Plant Cell Biology Group, Research School of Biological Sciences, The Australian National University, Canberra ACT 0200,
Australia, and2Sainsbury Laboratory, John Innes Centre, Colney Lane, Norwich, Norfolk NR4 7UH, UK
Received 4 October 2005; accepted 15 December 2005.*For correspondence (fax þ61 2 6125 4331; e-mail [email protected]).†Present address: CSIRO Plant Industry, GPO Box 350, Glen Osmond, SA 5064, Australia.‡Present address: Wheat Pathogenesis Programme, Plant-Pathogen Interactions Division, Rothamsted Research, Harpenden, Hertfordshire AL5 2JQ, UK.
Summary
The tomato Cf-9 gene confers resistance to races of the leaf mould fungus Cladosporium fulvum that carry the
Avr9 avirulence gene. Cf-9 was isolated by transposon tagging using a modified maize Dissociation (Ds)
element. This generated an allelic series of Ds-induced mutations of Cf-9, of which two were found to confer
novel phenotypes in a screen for mutants affecting wild-type Cf-9 function in trans. Genetic and molecular
analysis of these mutants suggested semidominant, Avr9-dependent, negative-interfering mutations invol-
ving Ds insertions in a defined subregion of Cf-9. Interference was associated with expression of the 5¢-end of
Cf-9 upstream of the Ds insertions in these mutants, suggesting that truncated Cf-9 proteins were the likely
cause of interference. Transgenic tomato lines harbouring Cf-9 constructs with premature stop codons in
positions similar to the Ds insertions also showed interference, indicating that the presence of Ds was not
required for interference to occur. Interestingly, interference in these transgenic lines was completely
dominant and was associated with a pronounced developmental phenotype that was dependent on co-
expression of Cf-9, Avr9 and a truncated Cf-9 transgene. However, interference with a weakly autoactive Hcr9
gene was Avr9-independent and did not cause a developmental phenotype, suggesting that localized
restoration of Cf-9/Avr9-dependent cell death was responsible for the developmental phenotype. The
restricted region in which truncation of Cf-9 results in dominant-negative interference suggests that leucine-
rich repeats (LRR) 16–19 of Cf-9 may mediate dimerization of Cf-9 and LRRs 20–23 may mediate interactions
with downstream partner proteins required for Cf-9 signalling, or vice versa.
Keywords: negative interference, plant disease resistance, Cladosporium fulvum, leucine-rich repeat, trans-
poson.
Introduction
Plant disease resistance genes encode proteins capable of
recognizing specific, corresponding avirulence proteins
produced by pathogens during infection, allowing early and
rapid activation of localized defence responses that prevent
progression of the disease (Dangl and Jones, 2003; Ham-
mond-Kosack and Jones, 1996). A large number of disease
resistance genes have been cloned and sequenced in recent
years, and the proteins that they encode are structurally
related. Most resistance genes encode modular, cytosolic
proteins that possess a central nucleotide-binding site (NBS)
linked to a C-terminal leucine-rich repeat (LRR) domain
(Dangl and Jones, 2003). However, a small number of
resistance genes encode transmembrane proteins that
possess large, extracytosolic LRR domains. Of these, the rice
Xa21 and Xa26 proteins are unique in possessing cytosolic
kinase domains (Song et al., 1995; Sun et al., 2004). How-
ever, the tomato Cf proteins, which confer resistance to
Cladosporium fulvum (Dixon et al., 1996, 1998; Jones et al.,
1994; Thomas et al., 1997), the apple HcrVf2 protein, which
confers resistance to Venturia inaequalis (Belfanti et al.,
ª 2006 The Authors 385Journal compilation ª 2006 Blackwell Publishing Ltd
The Plant Journal (2006) 46, 385–399 doi: 10.1111/j.1365-313X.2006.02699.x

2004) and the Arabidopsis RPP27 protein, which confers
resistance to Peronospora parasitica (Tor et al., 2004), all
possess short cytosolic tails. As these proteins do not pos-
sess obvious catalytic domains, it is a challenge to under-
stand how these receptor-like proteins function.
The tomato Cf-9 protein is one of the best-studied
examples of a receptor-like resistance protein. Cf-9 has an
extracellular LRR domain composed of 27 LRRs that is split
into two parts by a region of non-LRR sequence (Jones et al.,
1994). The Cf-9 protein mediates recognition of Avr9, an
avirulence protein that is secreted into the intercellular space
by C. fulvum during infection (De Wit and Spikman, 1982;
van Kan et al., 1991). Although Cf-9 has the hallmarks of a
receptor, binding studies have been unable to detect a direct
interaction between Cf-9 and Avr9 (Luderer et al., 2001).
Instead, Avr9 binds to a high-affinity binding site present on
the cell surface of all solanaceous plants regardless of the
presence or absence of Cf-9 (Kooman-Gersmann et al.,
1996). Therefore, it is likely that Cf-9 must interact with
partner proteins both during perception of the Avr9 stimulus
and during subsequent activation of defence signalling
pathways. These presumptive protein–protein interactions
are likely to involve different regions of Cf-9, as domain
swapping experiments between Cf-9 and Cf-4 have identi-
fied the N-terminal region of the LRR domain as critical for
Avr9 recognition (van der Hoorn et al., 2001; Wulff et al.,
2001), whereas sequence conservation between Cf proteins
in the C-terminal half of the LRR domain may indicate a site
of interaction with a common signalling partner (Jones and
Jones, 1997; Thomas et al., 1997).
Proteins that contain LRR domains are important compo-
nents of many signalling pathways in plants and animals
(Dievart and Clark, 2004; Jones and Jones, 1997). In order to
understand how these proteins function, mutation studies
have been performed that often eliminate protein function.
However, in some cases, production of apparently non-
functional mutant proteins can affect wild-type protein
function when expressed in the same cell, a phenomenon
termed interference. For example, the Toll gene of Dro-
sophila encodes a membrane-anchored LRR receptor
involved in embryo development and adult innate
immunity. Truncation mutations of Toll that cause loss of
the C-terminal transmembrane and cytosolic signalling
domains were themselves inactive, but resulted in activation
of wild-type Toll function in regions of the Drosophila
embryo in which it would not normally be active, indicative
of positive interference with wild-type Toll function by the
truncated Toll protein (Schneider et al., 1991). In contrast,
mutants of the Arabidopsis ERECTA gene have been gener-
ated that display negative interference. ERECTA is a mem-
brane-bound receptor kinase with an extracellular LRR
domain that controls organ shape and inflorescence archi-
tecture (Torii et al., 1996). Deletion of the ERECTA kinase
domain resulted in an erecta loss-of-function phenotype,
even though a wild-type ERECTA gene was present in the
genetic background of transgenic plants expressing the
deletion mutant (Shpak et al., 2003). Examples of interfer-
ence by mutated signalling proteins have also been
observed for other structural classes of receptor and,
commonly, interference is explained in terms of interactions
between mutated receptor proteins and their signalling
partners that inhibit the formation of functional signalling
complexes by the wild-type protein (Avery Ince et al., 1993;
Nagaya and Jameson, 1993).
The tomato Cf-9 genewas isolated by transposon tagging,
using an Avr9 transgene to select for inactivation of Cf-9 by
insertion of a modified maize Dissociation (Ds) element,
following mobilization by a stabilized maize Activator (sAc)
element (Jones et al., 1994). A large number of mutants
were recovered from the tagging experiment that carried Ds
elements within the Cf-9 coding sequence. Ds insertion in
any orientation or reading frame would have introduced an
in-frame termination codon within 132 bp of the junction
between Cf-9 and Ds sequences, so it was likely that
expression of these genes would result in the production
of truncated Cf-9 proteins. We used this allelic series of non-
functional Ds-insertion mutants to search for mutations that
might cause interference with wild-type Cf-9 function. Two
Ds-insertion mutants were identified that reduced wild-type
Cf-9 function in trans, consistent with negative interference.
Moreover, transgenic plants harbouring Cf-9 constructs with
premature stop codons at positions similar to these two Ds
insertions caused negative interference with wild-type Cf-9
function, associatedwith pronounced developmental abnor-
malities. This study examines some of the genetic and
molecular genetic properties of these negative interference
mutations and the insight they provide into Cf-9 function.
Results
The M75 and M102 interference phenotype
Cf-9 coding sequences adjacent to the Ds-insertion sites of
19 independent Ds-insertion mutants were amplified by
polymerase chain reaction (PCR) and sequenced to deter-
mine the location of the Ds element (Table 1). These mutant
plants (heterozygous for Cf-9::Ds and Avr9) were crossed to
Cf9 tomato plants (homozygous for Cf-9) and segregation of
the seedling lethal phenotype caused by interaction be-
tween Cf-9 and Avr9 was examined among the progeny to
look for evidence of alteration in wild-type Cf-9 function. A
1:1 segregation of normal plants:dead seedlings was ob-
served among the progeny of 17 of the 19 mutants, sug-
gesting that the Cf-9::Ds genes in thesemutants do not affect
Cf-9 function in trans. In contrast, a 2:1:1 segregation of
normal plants:dead seedlings:surviving seedlings showing
progressive chlorosis and necrosis later in development was
observed among the progeny of two mutants, M75 and
386 Claire L. Barker et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 46, 385–399

M102 (Figure 1). This indicates incomplete (i.e. semidomi-
nant) negative interference with Cf-9 function by Cf-9::DsM75
and Cf-9::DsM102. PCR analysis confirmed the presence of
both Ds and Avr9 in these surviving plants, consistent with
this interpretation (data not shown).
Both of the original M75 and M102 plants were variega-
ted for necrosis, which indicated the presence of sAc
(Table 1) and suggested simple insertions of single Ds
elements into Cf-9. However, the 2:1:1 segregation ratio
observed in progeny of crosses between Cf9 tomato plants
and M75 or M102 suggested that the interference pheno-
type was not dependent on sAc, which also segregated
among the progeny of each cross. This was confirmed by
PCR analysis, which enabled plants displaying the inter-
ference phenotype, but lacking sAc, to be identified (data
not shown).
The M75 and M102 interference plants showed leaf
chlorosis followed by leaf necrosis progressing from the
base of the plant towards the apex (Figure 1). The phenotype
of the M102 interference plants was less severe than that for
M75 suggesting that Cf-9::DsM102 causes greater interfer-
ence with Cf-9 function than Cf-9::DsM75 (Figure 1). Normal
progeny carrying Cf-9::Ds, but not Avr9, from crosses
between Cf9 tomato plants and either M75, M102 or the
non-interfering Ds-insertion mutants, M29 or M88, were
screened by inoculation with C. fulvum race 5 to determine
whether interference with Cf-9 function affected disease
resistance (Table 2). More pathogen growth was allowed on
Cf9 · M75 and Cf9 · M102 plants than on Cf9 · M29 or
Cf9 · M88 plants (Table 2), consistent with interference in
Cf-9-mediated pathogen resistance. However, sporulation
was not observed on Cf9 · M75 or Cf9 · M102 plants,
consistent with incomplete interference.
Analysis of Cf-9::Ds expression
Reverse transcriptase (RT)-PCR analysis was conducted to
determine whether interference was correlated with
expression of the 5¢- or 3¢-end of Cf-9::Ds using RNA
extracted from mature leaves of plants carrying the Cf-
9::Ds genes from M75, M102 or M248, but lacking Cf-9 and
Avr9. M248 is a non-interfering mutant that contains a Ds
insertion in Cf-9 at a position close to those found in M75
and M102 (Table 1). Amplification of transcripts using Cf-9-
specific primers in conjunction with Ds-specific primers
confirmed that Cf-9::Ds transcripts corresponding to the 5¢-end of Cf-9 could be detected for each of the three mutants
tested (Figure 2). However, transcripts corresponding to
the 3¢-end of Cf-9 could only be detected for M75 and
M248. Interestingly, the Ds elements in these two mutants
are inserted in the opposite orientation to the Ds element
in M102 (Table 1). Despite repeated attempts, no product
was ever obtained from M102 that corresponded to a 3¢ Cf-9::Ds transcript. These data indicate that interference is
likely to be associated with expression of the 5¢-end of
specific Cf-9::Ds genes.
Table 1 Positions and interference proper-ties of Ds-insertion mutants of Cf-9
MutantOrientationof Dsa
8 bpduplication
Domainaffectedb
Avr9-dependentinterference sAc present
23 ‹ CTTGTATT A No No142 fi AATGGAAT LRR8 No No20 ‹ CTCATGAC LRR8 No No96 fi GTGTGAAT LRR8 No No29 fi ATCTTGAA LRR10 No Yes52 fi CACAATAA LRR16 No No
219 fi GTTGAGAG LRR18 No Yes247 ‹ ATTTTAAG LRR19 No No102 fi GTCCCATC LRR21 Yes Yes75 ‹ TTTATGGG LRR22 Yes Yes
248 ‹ ATTCCAAG LRR24 No Yes258 ‹ CATTCCTT LRR27 No Yes88 fi GATGCATC LRR27 No No30 fi GTGAAGAT D/E No No41 fi CCAATGAT E No No
234 ‹ GGGGTTCT F No Yes255 fi ACGGTTGT F No No51 fi GTTGTGGA F No Yes
141 ‹ GGTCAACT F No No
aArrows indicate Ds orientation relative to the Cf-9 coding sequence, with fi indicating senseand ‹ indicating antisense orientation of the transposase gene of the Ac element from whichthe Ds element was derived, but antisense and sense orientation, respectively, of the 35S:gusAgene located within the Ds element.bDomains are named according to those described by Jones et al. (1994).
Interfering mutations of Cf-9 387
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 46, 385–399

Generation of transgenic plants containing artificially
truncated Cf-9 genes
To eliminate the possibility that the Ds element played a role
in the negative interference observed in Cf9 · M75 or
Cf9 · M102 plants, two C-terminal deletion (CTD) constructs,
CTD75 and CTD102, were produced in which stop codons
were introduced into the Cf-9 coding region at positions
identical to the sites of Ds insertion in M75 and M102
respectively. The non-interfering mutants M247 and M248
carried the closest flanking Ds insertions, located in the re-
gion of Cf-9 encoding LRRs 19 and 24 respectively (Table 1).
Therefore, four additional CTD constructs (CTD20–23) were
made with stop codons causing truncations in LRRs 20–23,
to further define the window of interference activity (Fig-
ure 3a). For all six constructs, the native Cf-9 promoter and
terminator sequences were used and the 3¢-end of the Cf-9
coding region, downstream of the introduced stop codon,
was deleted to ensure that the truncated Cf-9 (Cf-9tr) tran-
scripts possessed 3¢-untranslated region (UTR) sequences
identical to the wild-type Cf-9 transcript (Figure 3b).
The CTD constructs were used to transform Cf9 tomato
plants and six to 10 independent lines were recovered for
each construct. These were crossed as female parents to
SLJ6201B, a Cf0 line (i.e. lacking Cf-9) homozygous for a
35S:Avr9 transgene (Hammond-Kosack et al., 1994), and the
seedling progeny were screened for interference. The nature
of this cross ensured that all progeny carried Cf-9 and Avr9,
but segregated for Cf-9tr. If the introduced transgene had no
effect on wild-type Cf-9 function, or had been lost due to
segregation, the expression of Cf-9 and Avr9 in seedlings
should result in their death. However, if Cf-9tr were capable
of causing negative interference, then seedlings should
survive, despite expression of Cf-9 and Avr9 in the plant.
At least 10 seedlings were screened for each line and
seven lines were identified for which seedling survival
indicated dominant-negative interference (Table 3). Two
CTD102 lines showed interference, demonstrating that the
phenomenon was not dependent on the presence of the Ds
sequence but could be caused by a construct in which Cf-9
was truncated at the same position as in the Cf-9::Ds gene
fromM102. One CTD20 line, two CTD21 lines and two CTD22
lines also exhibited interference. However, no surviving
seedlings were present in the progeny from seven inde-
pendent CTD75 lines or nine independent CTD23 lines
crossed to SLJ6201B (Avr9).
The ratio of healthy to necrotic seedlings in the progeny of
transformed lines that displayed interference in crosses to
SLJ6201B (Avr9) allowed the number of loci causing inter-
ference to be determined. For the independent transgenic
lines, CTD102G, CTD21E and CTD22D, only one locus was
predicted to cause interference and this was supported by an
analysis of the number of T-DNA loci and the number of T-
DNAs integrated into the genome (Table 3). As only three
Figure 1. Interference of M75 and M102 with the seedling death phenotype
resulting from interaction between Cf-9 and Avr9.
(a) Two-week-old seedlings resulting from a cross between Cf9 tomato
(homozygous for Cf-9) and SLJ6201B (homozygous for an Avr9 transgene).
(b) Six-week-old siblings produced from a cross between an M102 plant
carrying Avr9 (Cf-9::DsM102/þ Avr9/)) and Cf9 tomato (Cf-9/Cf-9 )/)). The plant
on the left is wild type in appearance and lacks Avr9, whereas the plant on the
right contains Avr9 and has survived due to dominant-negative interference
with wild-type Cf-9 function. The chlorosis and necrosis evident on the lower
leaves of this plant indicates that interference is incomplete.
(c) Eight-week-old siblings produced from a cross between an M75 plant
carrying Avr9 (Cf-9::DsM75/þ Avr9/)) and Cf9 tomato (Cf-9/Cf-9 )/)). As in (b),
the plant on the right survived due to dominant-negative interference
with wild-type Cf-9 function. However, the phenotype of survivors that carried
Cf-9::DsM75 was more severe than that observed for interference produced by
Cf-9::DsM102.
388 Claire L. Barker et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 46, 385–399

progeny were recovered from the cross involving CTD102B,
it was not possible to obtain a statistically meaningful
estimate of the number of interference loci for this line.
However, the fact that only one T-DNA was detected in the
genome of CTD102B suggests that this line also possessed a
single interference locus. In contrast, although only one
interference locus was predicted for CTD21C, analysis of the
number of integrated T-DNA sequences and the number of
T-DNA loci revealed that multiple copies were present at
more than one location in the genome. Consequently, it
appears that not all T-DNA insertions were capable of
causing interference in this line. CTD22B also possessed
multiple T-DNA copies at different locations in the genome,
but at least four loci appeared to be active in this line as no
necrotic seedlingswere recovered among the progeny of the
cross to SLJ6201B (Avr9).
Cf-9tr expression in CTD plants
The low frequency of recovery of interference lines from
plants transformed with CTD constructs prompted an
investigation of Cf-9tr expression in transgenic lines, to
determine if construct expression was related to the ability
to interfere with wild-type Cf-9 function. Reverse transcrip-
tase-PCR analysis was conducted using primers that span
both the region deleted from CTD constructs and the 115 bp
intron in the 3¢-UTR of the Cf-9 gene, allowing products
amplified from Cf-9tr transcripts to be readily distinguished
from those amplified from wild-type Cf-9 transcripts. First,
we examined transgene expression in the 10 independent
CTD102 lines, of which only two displayed interference. As
shown in Figure 4(a), all 10 CTD102 lines expressed the
transgene, including the two interference lines, CTD102B
and CTD102G. Secondly, we examined transgene
expression in six of the seven lines that exhibited interfer-
ence. Although significant contamination of cDNA samples
by genomic DNA was evident, we were clearly able to detect
expression of spliced Cf-9tr transcripts in all six of the inter-
ference lines examined (Figure 4b). Finally, we examined Cf-
9tr expression in the surviving progeny of crosses between
CTD102B · SLJ6201B (Avr9) and CTD102G · SLJ6201B
(Avr9). As shown in Figure 4(c), Cf-9tr expression could be
detected in transgenic lines that did not contain Avr9 and in
surviving plants that contained Cf-9, Avr9 and the CTD102
construct. Together these data indicate that truncated Cf-9
genes are expressed in transgenic plants that carry CTD
constructs and that expression of Cf-9tr is evident in progeny
displaying dominant-negative interference. However, as Cf-
9tr transcripts can also be detected in transgenic lines that do
Table 2 Phenotypes of semidominantAvr9-dependent interfering mutations ofCf-9
Genotype Location of Ds
Infection phenotype
Avr9 transgene phenotypeHost Pathogena
Cf-9/þ Normal L: 2–4 Seedling deathResistance D: 1–2
Cf-9/Cf-9::DsM29 LRR10 Normal L: 2–4 Seedling deathResistance D: 1–2
Cf-9/Cf-9::DsM102 LRR21 Reduced L: 18–25 Survival with delayed necrosisResistance D: 4–6
Cf-9/Cf-9::DsM75 LRR22 Reduced L: 8–14 Survival with delayed necrosisResistance D: 2–4
Cf-9/Cf-9::DsM88 LRR27 Normal L: 2–4 Seedling deathResistance D: 1–2
aL, lateral hyphal growth of Cladosporium fulvum race 5 permitted in plant cell diameters;D, depth of hyphal growth permitted in plant cell diameters.
Figure 2. Expression of Cf-9::Ds genes in Ds-tagged mutants.
Mature leaf RNA was used to generate first-strand cDNA in reactions
containing (RTþ) or lacking (RT)) reverse transcriptase. The PCRs were
performed using Cf-9-specific primers (F6 and F90) in conjunction with Ds-
specific primers (5¢-Ds and 3¢-Ds) to amplify the 5¢- and 3¢-ends of Cf-9::Ds
fusion transcripts (see Experimental procedures). The choice of primer
combination was dictated by the orientation of the Ds element within the
Cf-9 gene (Table 1). To detect expression of the 3¢-end of Cf-9::Ds fusion
transcripts, amplification was performed across the site of a 115 bp intron
found within the 3¢-UTR of the Cf-9 gene. The PCRwas performed using cDNA
reactions or genomic DNA from the appropriatemutant (DNA) as the template
source. Approximate product sizes are shown in base pairs.
Interfering mutations of Cf-9 389
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 46, 385–399

not cause interference, additional factors may influence the
ability of CTD constructs to interfere with wild-type Cf-9
function.
Cf-9 and Avr9 expression in CTD102 plants
The survival of plants carrying Cf-9,Avr9 and CTD constructs
could be due to inactivation of the Cf-9 or Avr9 genes.
Therefore, we investigated wild-type Cf-9 and Avr9 expres-
sion in transgenic plants. Cf-9 is a member of a large
multigene family in tomato (Parniske et al., 1999), so we
designed primers that spanned the intron in the 3¢-UTR of
the Cf-9 gene and capitalized on the presence of a poly-
morphic nucleotide present in Cf-9 but absent from all other
sequenced Hcr9 (homologue of C. fulvum resistance gene
Cf-9) genes, to generate a novel BstBI restriction site only in
products derived from Cf-9 transcripts. To confirm the spe-
cificity of our primers, we amplified products from Cf0 and
Cf9 genomic DNAs or cDNAs and digested them with BstBI.
Only the products amplified fromCf9 genomic DNA or cDNA
were cleaved by BstBI (Figure 5a). The RT-PCR product
amplified from Cf9 cDNA was cloned and five clones were
sequenced. These were all found to correspond to tran-
scripts of the Cf-9 gene (data not shown).
To determine if the wild-type Cf-9 gene was expressed in
plants displaying interference, RNA was extracted from two
surviving siblings obtained from CTD102B · SLJ6201B
(Avr9) and CTD102G · SLJ6201B (Avr9) crosses. Two sib-
lings carrying CTD102 obtained by self-fertilization of
CTD102B and CTD102G were also examined. Reverse tran-
scriptase-PCR analysis indicated that Cf-9 was expressed in
all CTD102B- and CTD102G-derived plants (Figure 5b), inclu-
ding surviving plants obtained from crosses with SLJ6201B
(Avr9). The RT-PCR products amplified from transcripts of
surviving plants carrying Cf-9, Avr9 and CTD102 were
cloned. Five clones obtained from each individual were
sequenced and all were found to correspond to transcripts of
the Cf-9 gene (data not shown).
Avr9 expression was also examined in plants derived
from the CTD102B and CTD102G (Figure 5b). As expected,
no PCR products were amplified from self-progeny of
CTD102B and CTD102G transformants, which lacked the
Avr9 gene, but products were amplified from surviving
progeny obtained by crossing CTD102B and CTD102G to
SLJ6201B (Avr9). Therefore, we conclude that the survival of
plants carrying Cf-9, Avr9 and CTD constructs is not caused
by inactivation of the wild-type Cf-9 or Avr9 genes.
CTD75 and CTD23 show interference in an agroinfiltration
assay
The absence of interference in the progeny of CTD75 lines
crossed to SLJ6201B (Avr9), appeared to contradict the
evidence for interference in the M75 mutant upon which
(a)
(b)
Figure 3. C-terminal deletion constructs generated to determine the region
of Cf-9 in which truncation results in dominant-negative interference.
(a) The positions of Ds insertions and engineered truncations are shown
relative to the amino acid sequence for LRRs 18–24 of Cf-9. M75, M102, M219,
M247 and M248 are Ds-insertion mutants. CTD75, CTD102, CTD219 and
CTD20–23 are Cf-9 constructs that contain premature translation-termination
signals, resulting in C-terminal deletions of the Cf-9 protein. Residues shown
in bold type represent the last amino acid encoded by Cf-9::Ds or Cf-9tr
transcripts prior to termination. Residues underlined represent amino acids
mutagenized as a result of construct design.
(b) C-terminal deletion constructs were generated that contained premature
stop codons introduced into the Cf-9 coding region at seven different
locations. Constructs retained the wild-type Cf-9 promoter and 3¢-UTR
complete with 115 bp intron. The location of the start (ATG) and stop (TAG)
codons of wild-type Cf-9 are indicated and the coding region is shown as a
rectangle. Regions encoding each LRR are shown in grey. The dashed line
represents Cf-9 coding sequence deleted from each CTD construct and each
artificially introduced stop codon is shown as an asterisk.
390 Claire L. Barker et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 46, 385–399
CTD75 was based. This prompted experiments to determine
whether interference could be detected using an agroinfil-
tration assay and, if so, whether CTD75 and CTD23 were
capable of causing interference. Initially, leaves of mature
SLJ6201A (Avr9) tobacco plants were co-agroinfiltrated with
pCLB3, a construct containing the wild-type Cf-9 and native
promoter, and either pCTD102 or pCTD219 (Figure 3), cor-
responding to Ds-insertion mutants M102, which shows
interference, and M219, which does not. Necrosis was ob-
served in leaf panels treated with pCLB3 and pCTD219,
indicating a lack of interference. However, no necrosis was
observed in leaf panels treated with pCLB3 and pCTD102,
indicating negative interference (Figure 6). Agroinfiltration
of pCLB3 and either pCTD75 or pCTD23 also failed to trigger
necrosis, indicating that both pCTD75 and pCTD23, like
pCTD102, are capable of causing interference (Figure 6).
Interference mutants display a distinct, developmental
phenotype dependent on Cf-9, Cf-9tr and Avr9
When Ds-tagged mutants M75 and M102 were crossed to
Cf9 tomato, progeny carrying Cf-9, Avr9 and Cf-9::Ds dis-
played a delayed necrotic phenotype. In contrast, when
transgenic plants harbouring CTD constructs were crossed
to SLJ6201B (Avr9), none of the surviving progeny carrying
Cf-9, Cf-9tr and Avr9 exhibited delayed necrosis. Instead, all
surviving plants displayed a distinct developmental pheno-
type, the severity of which differed between survivors de-
rived from different CTD transgenic lines (Figure 7). The
least severely affected surviving plants were those from the
CTD22B and CTD22D crosses, which appeared almost wild
type, with only slight stunting evident (Figure 7a). The
stunting of surviving plants from the CTD102G and CTD20C
crosses was more severe than that observed for the two
CTD22 lines, yet these plants still grew to maturity, flowered
and set fruit. Surviving CTD20C · SLJ6201B (Avr9) plants
exhibited slight twisting of the petioles and reduced leaf size
in comparison with wild-type controls. The petiole-twisting
trait was more severe in surviving CTD102G · SLJ6201B
(Avr9) plants and leaves of these plants appearedmisshapen
andwaxy, with stiff mid-veins (Figure 7b,c). Surviving plants
derived from the CTD21E · SLJ6201B (Avr9) cross were se-
verely stunted in comparison with wild-type controls and
rarely flowered (Figure 7d). Any flowers that did form often
became necrotic soon after bud formation. Leaves were very
small, twisted and misshapen (Figure 7e). Petiole twisting
was severe and mature, fully expanded leaflets generally
grew only 1–2 cm in length, whereas the leaflets of wild-type
plants were approximately 6–8 cm in length (data not
shown).
The most severely affected surviving plants were those
from the CTD102B and CTD21C crosses, which were char-
acterized by dramatic stunting and reduced mature leaf
formation (Figure 7f,g). In each case, seedlings germinated
normally and the cotyledons expanded, but subsequent
growth was strongly impaired. Mature leaves were highly
irregular in shape, thickened, waxy and translucent in
comparison with wild-type controls. The ability to form
mature leaves appeared to decline with plant age, and
abortive organ formation was evident as small regions of
necrotic tissue (Figure 6h,i). In the most severe cases, the
ability to specify organs deteriorated as the apical meristem
became an amorphous mass of cells. Flowers never formed
on these plants, and at 9 weeks of agemature plants reached
approximately 10 cm in height whereas wild-type controls
were approximately 50–100 cm tall (data not shown).
All primary CTD transformants were wild type in appear-
ance and abnormal developmental phenotypes were only
observed in the surviving progeny of crosses between the
primary CTD transformants and SLJ6201B (Avr9) tomato.
This indicated that the developmental phenotype observed
was Cf-9tr and Avr9 co-dependent. To determine if the
developmental phenotype also required the wild-type Cf-9
gene, surviving plants from the cross between CTD102G and
SLJ6201B (Avr9) were allowed to self-pollinate. The
CTD102G line was chosen for study as it only contains one
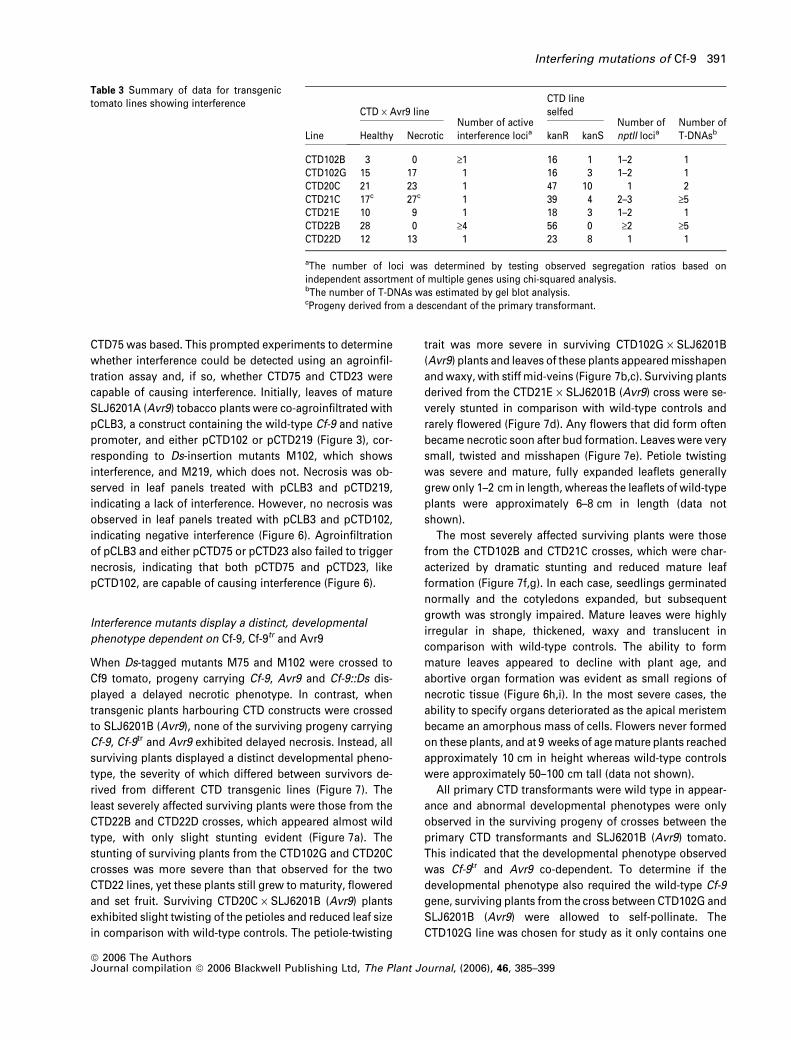
Table 3 Summary of data for transgenictomato lines showing interference
Line
CTD · Avr9 lineNumber of activeinterference locia
CTD lineselfed
Number ofnptII locia
Number ofT-DNAsbHealthy Necrotic kanR kanS
CTD102B 3 0 ‡1 16 1 1–2 1CTD102G 15 17 1 16 3 1–2 1CTD20C 21 23 1 47 10 1 2CTD21C 17c 27c 1 39 4 2–3 ‡5CTD21E 10 9 1 18 3 1–2 1CTD22B 28 0 ‡4 56 0 ‡2 ‡5CTD22D 12 13 1 23 8 1 1
aThe number of loci was determined by testing observed segregation ratios based onindependent assortment of multiple genes using chi-squared analysis.bThe number of T-DNAs was estimated by gel blot analysis.cProgeny derived from a descendant of the primary transformant.
Interfering mutations of Cf-9 391
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 46, 385–399

T-DNA and interference locus (Table 3), so surviving plants
used for self-pollination were all expected to be heterozy-
gous for Cf-9, Cf-9tr and Avr9. The progeny were scored for
segregation of the developmental phenotype and from 190
plants tested, 95 werewild type, 75 displayed developmental
abnormalities and 20 died as seedlings. This conforms to the
28:27:9 ratio for wild type:developmentally abnormal:nec-
rotic seedlings expected for independent assortment of
three heterozygous loci if the developmental phenotype is
dependent on the presence of Cf-9tr, Avr9 and the wild-type
Cf-9 gene (see Table S1 for a detailed analysis).
Interference with an autoactive Hcr9 protein is not
dependent on Avr9
Although interference in Cf-9 function is dependent on Avr9,
the role of Avr9 in interference is not clear. Recognition of
Avr9 by full-length Cf-9 protein may simply provide a back-
ground necrosis phenotype upon which interference by
truncated Cf-9 is detected. However, the M75, M102 and CTD
truncations extendwell beyond the LRR domains involved in
Avr9 recognition (van der Hoorn et al., 2001; Thomas et al.,
1997; Wulff et al., 2001), raising the possibility that recog-
nition of Avr9 by truncated Cf-9 proteins might also be
required for interference. To determine whether Avr9 is
required for interference to occur or simply required for its
detection, CTD102G · SLJ6201B plants showing the devel-
opmental phenotype, and therefore heterozygous for Cf-9,
Cf-9tr and Avr9, were crossed to tomato plants heterozygous
for Hcr9-M205, which encodes an autoactive protein
(a)
(b)
Figure 5. Cf-9 and Avr9 expression in plants displaying interference.
(a) Primers Hcr9F2 and Hcr9R1 were used to amplify products from Cf0 and
Cf9 genomic DNA and cDNA. The PCR products are shown that were either
uncut ()) or cut (þ) with BstBI. The size difference between products amplified
from genomic DNA and those amplified from cDNA is consistent with splicing
of the intron found in the 3¢-UTR of Hcr9 transcripts. Approximate product
sizes are shown in base pairs.
(b) Polymerase chain reaction amplification using primers Hcr9F2 and Hcr9R1
followed by digestionwithBstBI was used to detectCf-9 expression in CTD102
plants displaying dominant-negative interference. Plants 1 and 2 are progeny
obtained by selfing of CTD102B or CTD102G, whereas plants 3 and 4 are
surviving progeny of the cross between the respective primary transformant
and SLJ6201B (Avr9). Polymerase chain reaction was also performed using
AVR9F/AVR9R2 to detect Avr9 expression and GAPDHF/GAPDHR to detect
expression of the housekeeping gene glyceraldehyde-3-phosphate dehy-
drogenase (GAPDH). For size comparison, DNA controls were amplified from
genomic DNA extracted from the transgenic line, CTD102G (Cf-9 and GAPDH
panels), or pSLJ6201 plasmid DNA (Avr9 panel). Approximate product sizes
are shown in base pairs.
(a)
(b)
(c)
Figure 4. Cf-9tr expression in transgenic plants.
Mature leaf RNA was extracted and used to generate first-strand cDNA for
amplification with the primers Hcr9F1 and Hcr9R1. As these primers amplify
across a 115 bp intron in the Cf-9 3¢-UTR, products amplified from cDNA are
smaller than those amplified from genomic DNA due to intron splicing.
Reactions were also performed using primers that amplify the housekeeping
gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH) to confirm the
use of approximately equal concentrations of cDNA. Approximate product
sizes are shown in base pairs.
(a) Hcr9F1/Hcr9R1 products amplified from 10 independent transgenic lines
containing CTD102 are shown. The product amplified from genomic DNA
extracted from the transgenic line, CTD102G, is also shown (DNA) for size
comparison.
(b) Hcr9F1/Hcr9R1 products amplified from six independent transgenic lines
that cause dominant-negative interference are shown. As Hcr9F1 and Hcr9R1
span the region deleted from Cf-9 in CTD constructs, the size of the product
amplified differs between constructs. The product amplified from plasmid
DNA containing each CTD construct is also shown (DNA) for size comparison.
(c) Hcr9F1/Hcr9R1 products amplified from transgenic CTD102 lines that
exhibit dominant-negative interference are shown. CTD102B and CTD102G
are primary transformants that lack Avr9, whereas 102B · A9 and 102G · A9
are surviving progeny from crosses between the respective primary trans-
formants and SLJ6201B (Avr9). No products of similar size were amplified
from wild-type Cf0 or Cf9 DNA and the product amplified from genomic DNA
extracted from the transgenic line, CTD102G, is shown (DNA) for size
comparison.
392 Claire L. Barker et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 46, 385–399

comprising the C-terminus of Cf-9 and the N-terminus of the
Cf-9 paralogue Hcr9-9A (Barker et al., 2006). Hcr9-M205
confers a semidominant Avr9-independent gain-of-function
phenotype comprising stunting, wilting, acropetal chlorosis
and necrosis, and constitutive activation of plant defence
genes (Barker et al., 2006). In this cross, half the plants re-
ceived Hcr9-M205 and should have displayed the M205
phenotype. The progeny of the cross were scored for seg-
regation of the developmental phenotype and the M205
phenotype, and from 56 plants tested, 32 were wild type, 10
displayed the M205 phenotype, 10 displayed developmental
abnormalities and four died as seedlings. The 52 surviving
plants were assayed for the presence of the Hcr9-M205 gene
as described by Barker et al. (2006) and 22 were found to
carry Hcr9-M205. Of these, nine plants were wild type, three
were developmentally abnormal and only 10 displayed the
M205 phenotype, indicating that suppression of the M205
phenotype had occurred (Figure 8). The Cf-9 haplotype,
comprising Hcr9-9A, Cf-9B, Cf-9, Hcr9-9D and Hcr9-9E, has
been shown to delay development of the M205 phenotype
under ideal environmental conditions of low light and high
humidity (Barker et al., 2006), but the 12 plants showing
suppression of the M205 phenotype showed no sign of the
M205 phenotype at any stage in their development, even
under non-ideal conditions of high light and low humidity.
The segregation of the visible phenotypes observed among
the progeny of the CTD102G · SLJ6201B cross conforms to
a 9:3:2:2 ratio for wild type:M205:developmentally abnor-
mal:necrotic seedlings expected if suppression is dependent
on the presence of Cf-9tr but not on Cf-9 or Avr9 (see
Table S2 for a detailed analysis).
Discussion
Specifically truncated Cf-9 genes interfere negatively with
wild-type Cf-9 function
The allelic series of Ds-insertion mutations generated by
transposon tagging of the Cf-9 gene are non-functional as
resistance genes, as indicated by their inability to confer a
seedling lethal phenotype when expressed in tomato plants
carrying an Avr9 transgene (Jones et al., 1994). However, we
have shown that two Ds-insertion mutations, Cf-9::DsM75
and Cf-9::DsM102, cause semi-dominant, Avr9-dependent,
negative interference with wild-type Cf-9 function. Interfer-
ence was unrelated to the presence of sAc, the orientation of
the Ds element in the Cf-9 coding region or the expression of
the 3¢-end of Cf-9::Ds transcripts. However, interference was
correlated with the expression of the 5¢-end of Cf-9::Ds
genes, suggesting that specifically truncated Cf-9 proteins
might be responsible for the observed interference.
The Ds insertions of M75 and M102 were located in
regions encoding LRRs 22 and 21 of Cf-9 respectively.
Therefore, to further examine the phenomenon of negative
interference by truncated Cf-9 proteins, we generated a
series of Cf-9 constructs carrying artificially introduced stop
codons at six different positions within the region encoding
LRRs 20–23 and have shown, by production of transgenic
tomato lines and transient expression via agroinfiltration
that all six constructs have the ability to negatively interfere
with wild-type Cf-9 function. In contrast, a construct gener-
ated to mimic the truncated protein predicted to result from
Cf-9::DsM219 expression did not cause negative interference,
consistent with the hypothesis that truncation of Cf-9 only at
positions within LRRs 20–23 allows negative interference to
occur.
Potentially, the survival of transgenic plants containing Cf-
9, Avr9 and Cf-9tr could have been due to inactivation of the
Avr9 or wild-type Cf-9 genes. However, several lines of
evidence make this possibility unlikely. For five of the seven
Figure 6. Detection of interference with wild-type Cf-9 function by agroinfil-
tration.
Leaf panels of SLJ6201A (Avr9) tobacco plants were agroinfiltrated with a 6:1
ratio of Agrobacterium tumefaciens strain GV3101 containing either no binary
vector (GV3101) or a binary vector with a Cf-9 CTD: A. tumefaciens strain
GV3101 containing a binary vector with wild-type Cf-9 promoter and coding
region (pCLB3). Panels agroinfiltrated with GV3101 and pCLB3 or pCTD219
and pCLB3 (negative controls) showed necrosis, indicating lack of interfer-
ence in Cf-9 function, whereas panels agroinfiltratedwith pCLB3 and pCTD102
(positive control), pCTD75 or pCTD23 all showed no necrosis, indicating
interference in Cf-9 function. The leaf shown is representative of results
obtained for 21 leaves on 15 plants in four separate experiments. Similar
results were also obtained using 10:1, 8:1 and 4:1 ratios of Agrobacterium
containing pCTD construct: Agrobacterium containing pCLB3.
Interfering mutations of Cf-9 393
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 46, 385–399

CTD lines that displayed interference, both necrotic and
healthy seedlings were observed in the progeny of crosses
to SLJ6201B (Avr9). This indicates that a functional Cf-9 gene
was inherited from the primary transformants. Furthermore,
necrotic seedlings were observed among the progeny
obtained by self-pollination of surviving CTD102G
· SLJ6201B (Avr9) plants, indicating the inheritance of a
Cf-9 gene with normal function from plants displaying an
interference phenotype. Finally, RT-PCR analysis indicated
that both Avr9 and Cf-9were expressed in interference lines,
eliminating the possibility that the survival of seedlings was
due to gene silencing. Therefore, our data are consistent
with the proposal that seedling survival is due to dominant-
negative interference of wild-type Cf-9 function caused by
the expression of Cf-9tr genes.
Not all independent lines harbouring the same CTD
construct exhibited interference, despite the fact that RT-
PCR analysis detected expression of Cf-9tr genes in
all CTD102 lines tested. This suggests that the location of
T-DNA insertion in CTD transformants may have played a
role in interference by placing the CTD constructs in different
sequence contexts that may have affected their expression.
In support of this proposal, genetic analysis of the CTD21C
interference line identified at least five T-DNA copies
Figure 7. Abnormal developmental phenotype of surviving plants that expressed Cf-9, Avr9 and Cf-9tr.
Eight-week-old CTD transgenic lines are shown that were grown under identical glasshouse conditions. All plants contain Cf-9 and Cf-9tr. Plants were chosen that
displayed a representative phenotype from a pool of five or six individuals of like genotype.
(a) Mature CTD22B plants that lack (left) or contain (right) Avr9.
(b) Mature leaf taken from a CTD102G plant that lacked Avr9 and was wild type in appearance.
(c) Mature leaf taken from a surviving CTD102G plant that contained Avr9 and displayed the petiole-twisting characteristic of interference plants.
(d) Mature CTD21E plants that lack (left) or contain (right) Avr9.
(e) Surviving plant derived from a cross between CTD21E and SLJ6201B (Avr9). Plants were moderately stunted in comparison with wild type and leaf shape was
highly irregular. Leaves appeared feathery, with narrow, thickened leaflets.
(f) Surviving plant derived from a cross between CTD21C and SLJ6201B (Avr9). Plants were strongly stunted in comparison with wild type and leaf shape was highly
irregular.
(g) Mature CTD21C plants that lack (left) or contain (right) Avr9.
(h) Shoot apex of a surviving plant derived from a cross between CTD21C and SLJ6201B (Avr9). This plant displayed a severe phenotype, with the virtual absence of
a shoot apex structure and browning of the tissue in this region. Amature leaf can be seen on the left, with two necrotic patches at positions that might have formed
leaflets.
(i) Shoot apex of a surviving plant derived from a cross between CTD21C and SLJ6201B (Avr9) that was sibling of the plants shown in (g) and (h). The phenotype of
this plant was less severe than the plant shown in (h) and a shoot apex was evident. Browning of patches of tissue on a developing leaf can be seen.
394 Claire L. Barker et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 46, 385–399

integrated into the genome at two or more loci, but only one
locus was responsible for interference with Cf-9 function
(Table 3). Although RT-PCR confirmed that Cf-9tr genes were
expressed in CTD lines displaying interference, the methods
used were qualitative, not quantitative. Therefore, the pos-
sibility remains that the difference between CTD lines that
do, or do not, show interference when transformed with the
same construct may be related to the intensity or timing of
construct expression. Alternatively, this difference may be
related to the level of protein production rather than
transcript production. This might explain why, although
CTD75 and CTD23 could interfere with Cf-9 function in an
agroinfiltration assay, no transgenic lines were recovered
that gave rise to surviving seedlings when crossed to
SLJ6201B (Avr9) tomato.
The molecular basis for negative interference
All Hcr9 proteins, and even the more distantly related Hcr2
proteins, show strong sequence similarity in the C-terminal
half of the LRR domain, which in Cf-9 corresponds to LRRs
16–27 (see Figure S1). This led to the suggestion that the
conserved C-terminal LRRs might be involved in binding
partner proteins required for the activation of defence sig-
nalling pathways (Dixon et al., 1996; Jones and Jones, 1997).
Our data support this hypothesis, as the restricted region in
which truncation of Cf-9 results in dominant-negative inter-
ference is located within the conserved region and suggests
a specific effect on protein interactions in the vicinity of LRRs
20–23. The nature of the interference caused by truncated Cf-
9 proteins is currently unclear but, as proposed for
dominant-negative mutants of LRR receptor-like kinase
proteins and NBS-LRR resistance proteins, it is possible that
interfering proteins sequester signalling partner proteins
that are present in limiting quantities in the cell (Dievart
et al., 2003; Dinesh-Kumar et al., 2000; Shpak et al., 2003;
Tao et al., 2000).
Any model to explain our observations of interference
must account for the fact that truncation within a specific
region of Cf-9 results in negative interference, whereas
longer and shorter truncations do not. To date, the only
protein known to directly interact with Cf-9 is CITRX, a
negative-regulatory thioredoxin protein that binds the
cytosolic C-terminus of Cf-9 (Rivas et al., 2004). However,
it is unlikely that disruption to CITRX binding is responsible
for interference, as all of the truncation mutants of Cf-9
examined in this study lacked the cytosolic domain with
which CITRX interacts. Alternatively, interference could be
associated with interaction between truncated Cf-9 proteins
and an upstream partner protein required for Avr9 percep-
tion, a candidate for which is the plasma membrane-bound
Avr9 high-affinity binding site described by Kooman-Gers-
mann et al. (1996). Given that interference with Hcr9-M205
can occur in the absence of Avr9, the interaction between
Cf-9tr and the upstream partner protein would also have to
occur in the absence of Avr9. All of the residues critical for
Avr9 recognition are located in LRRs 10–15 (van der Hoorn
et al., 2001; Wulff et al., 2001) and, as these residues are
present in all truncated Cf-9 proteins that show dominant-
negative interference, it is possible that truncated Cf-9
proteins can bind the upstream partner protein, thereby
making it inaccessible to wild-type Cf-9 protein. However,
this proposal also seems unlikely, as it does not explain
interference with Hcr9-M205, which, given its autoactivity
and inability to recognize Avr9, either does not require an
upstream partner protein or requires a different upstream
partner protein. Instead, the simplest interpretation of our
observations is that LRRs 20–23 has one or other of two
distinct functions, either binding with a downstream part-
ner protein or Cf-9 dimerization, and two related models
can be proposed to explain how negative interference
occurs. The first scenario assumes that sequences in the
conserved LRRs preceding LRRs 20–23 (i.e. LRRs 16–19)
allow recruitment of a downstream partner protein required
for signal output and LRRs 20–23 contain sequences
required for Cf-9 dimerization. In this case, dominant-
negative truncated Cf-9 proteins would possess the partner
recruitment domain but lack the adjacent downstream
dimerization domain, allowing the truncated proteins to
sequester a downstream partner protein that might be
present in limiting quantities in the cell into a non-
functional protein complex. Alternatively, the dimerization
domain might precede the partner protein-binding site,
allowing dominant-negative truncated Cf-9 proteins to
dimerize with wild-type Cf-9 proteins, but not perform a
critical function mediated by the downstream domain that
Figure 8. Suppression of the M205 phenotype in plants carrying Hcr9-M205
and Cf-9tr.
An 8-week-old plant carrying Hcr9-M205 and Cf-9tr (left) from a cross between
a CTD102G · SLJ6201B plant carrying Cf-9tr, Avr9 and wild-type Cf-9 and a
M205 plant carrying Hcr9-M205 shows no M205 phenotype whereas a sibling
carrying Hcr9-M205 but not Cf-9tr (right) shows the M205 phenotype.
Interfering mutations of Cf-9 395
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 46, 385–399

is required in both molecules of the dimer for function to
occur, e.g. dimerization of a signalling partner might be
required for signalling to occur. In this case, wild-type Cf-9
is sequestered by the interfering protein. Common to both
scenarios, shorter truncations do not interfere as they lack
both the recruitment and dimerization domains and so do
not form complexes with proteins that are present in
limiting quantities in the cell, whereas longer truncations
that possess both domains can form functional signalling
complexes, but only upon dimerization with an activated
wild-type Cf protein.
Dominant-negative interference with Cf-9 function affects
plant development
Transgenic CTD plants able to interfere with Cf-9 function
displayed unexpected developmental abnormalities
dependent on the presence of Cf-9tr,Avr9 and wild-type Cf-9.
These abnormalities could have been caused by an inter-
action between defence signalling and plant developmental
signalling. Constitutive association of Cf-9, Avr9 and an
Avr9-binding protein in a non-functional complex might
have had an epistatic effect on plant developmental signal-
ling by sequestering a protein required for both Cf-9-medi-
ated and developmental signalling pathways. Alternatively,
the abnormal developmental phenotypes could have been
caused by localized breakdown in interference leading to
somatic restoration of Cf-9 function, due either to develop-
mental downregulation of Cf-9tr or to developmental accu-
mulation of a component sequestered by dominant-
negative forms of Cf-9 to a level that is no longer limiting.
Consistent with this idea, plants from the original Cf-9 tag-
ging experiment (Jones et al., 1994) carrying Cf-9::Ds, sAc
and Avr9, which were variegated for Cf-9-dependent necro-
sis as a consequence of somatic Ds excision, often showed
developmental abnormalities associated with the restor-
ation of Cf-9 function similar to those observed in interfer-
ence plants (see Figure S2).
In past studies, dominant-negative interference in plants
co-expressing mutated and wild-type resistance genes has
been demonstrated locally at sites of inoculation with the
corresponding avirulent pathogens (Dinesh-Kumar et al.,
2000; Tao et al., 2000). In contrast, the plants produced in
this study allowed expression of an avirulence gene, its
cognate resistance gene and a dominant-negative form of
the resistance gene throughout the plant over its entire life
cycle. Generation of interference plants for other combina-
tions of resistance and avirulence genes might provide a
useful tool, not only for dissection of defence signalling
pathways but also for studying the interplay between
defence signalling pathways and developmental cues that
cannot normally be done due to the localized and lethal
nature of resistance gene activation.
Experimental procedures
Plant material and cultivation
The production of Ds-insertion mutants of Cf-9 has been describedpreviously (Jones et al., 1994). Lycopersicon esculentum cv.Moneymaker (Cf0) lacks functional Cf resistance genes, whereas Cf9tomato is a near-isogenic line harbouring an introgression fromLycopersicon pimpinellifolium that contained the Cf-9 haplotype.Production of a Cf0 line homozygous for an Avr9 transgene(SLJ6201B) was described by Hammond-Kosack et al. (1994). Allplants were grown under glasshouse conditions (25�C by day, 18�Cby night, ambient light intensity and relative humidity).
Phenotypic characterization of Ds-induced mutants of Cf-9
Cultures and inocula of C. fulvum race 5 were prepared and used toinoculate mutant tomato plants or their progeny as described byHammond-Kosack and Jones (1993). Inoculated leaves were stained14 days after inoculation with lactophenol trypan blue and exam-ined microscopically as described by Hammond-Kosack and Jones(1993).
CTD construct generation
Plasmid SLJ10221B consists of a pDBS KS(þ) vector with a 5.3 kbBamHI fragment that contains the Cf-9 promoter, coding regionand 3¢-UTR. The sequence of the vector and insert were modifiedto introduce an XbaI site 9 bp upstream of the Cf-9 stop codonand to delete an XbaI site from the plasmid polylinker. CTD con-structs were generated by introducing an in-frame stop codon,followed by an XbaI restriction site, into the coding sequence ofCf-9 in pSLJ10221B by site-directed mutagenesis (Kunkel et al.,1987). Mutagenic primers are detailed in Table 4. Mutagenizedplasmids were cut with XbaI to release an approximately 1 kbinternal fragment of the 3¢-end of the Cf-9 coding region and re-circularized to form the deletion constructs CTD75, CTD102,CTD219 and CTD20–23. At this stage constructs were sequencedacross the junction points of the deletions to determine theirfidelity. The CTD constructs were then inserted as BamHI frag-ments into the BamHI site of the pSLJ7292 binary vector (des-cribed at http://www.jic.bbsrc.ac.uk/sainsbury-lab/jj/plasmid.html)and clones were selected for those in which the direction oftranscription of the Cf-9 construct and the nptII marker gene wereconvergent.
Generation of transgenic plants for analysis of negative
interference
Cf9 tomato was used for Agrobacterium tumefaciens-mediatedplant transformation as previously described (Jones et al., 1994).Screening of seedlings for kanamycin resistance was performed onMurashige and Skoog medium containing 150 lg ml)1 kanamycinsulphate to provide selection for plants expressing the nptII genecontained within binary vectors. To generate seed for analysis ofinterference, primary CTD transformants were crossed as femaleparents to the Avr9-containing line SLJ6201B (Hammond-Kosacket al., 1994). Individual seeds were sown into separate cells ofmulticompartmented trays and seedling phenotypes were monit-ored for a period of 4 weeks after sowing, at which time the num-bers of healthy and necrotic seedlings were recorded. Surviving
396 Claire L. Barker et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 46, 385–399

progeny were maintained under glasshouse conditions for theremainder of their life cycle.
Analysis of negative interference by agroinfiltration
Agroinfiltration was carried out essentially as described by Barkeret al. (2006). Agrobacterium tumefaciens cultures were pelleted andresuspended to an optical density of 0.5 at 600 nm before beingused to make mixed suspensions containing 10:1, 8:1, 6:1 or 4:1ratios of A. tumefaciens strain GV3101 containing the CTD constructunder study to A. tumefaciens strain GV3101-containing pCLB3, abinary vector carrying the wild-type Cf-9 gene (Barker et al., 2006).The mixed suspensions were infiltrated into leaves of budding andearly flowering SLJ6201A tobacco plants homozygous for Avr9(Hammond-Kosack et al., 1998). Development of Cf-9-dependentnecrosis occurred 4–14 days later, depending on environmentalconditions.
RNA extraction and RT-PCR analysis
Five mature leaflets per plant were ground in liquid nitrogen and100 mg of tissue was used for total RNA extraction using theRNeasy Plant Mini Kit (Qiagen, Valencia, CA, USA). For RT-PCRanalysis, 1.2 lg total RNA was incubated in a volume of 12 ll con-taining 1X DNase buffer, 1.2 U RQ1 RNase-free DNase (Promega,Madison, WI, USA), 26.175 U RNAguard Porcine RNase Inhibitor(Amersham Pharmacia Biotech, Piscataway, NJ, USA) for >4 h at37�C, then for 15 min at 65�C. First-strand cDNAwas then generatedfrom 5 ll of DNase-treated RNA in a reaction volume of 18 ll con-taining 50 mM Tris-HCl pH 8.3, 75 mM KCl, 10 mM dithiothreitol,
5 mM of each nucleotide (dNTP), 3 mM MgCl2 and 200 ng oligodeoxy-thymidine [oligo dT(12)18)]. The reaction was incubated at65�C for 5 min, then at 37�C for 10 min before adding 200 USuperScript II reverse transcriptase (Invitrogen, Carlsbad, CA, USA)and 8.725 U RNAguard Porcine RNase Inhibitor (Amersham Phar-macia Biotech). The reaction volume was adjusted to 20 ll beforeincubation at 37�C for 60 min, then at 95�C for 5 min.
To detect Cf-9::Ds transcripts, PCRs were conducted in 25 llvolumes using appropriate combinations of primers F6, F90, 5¢-Dsand 3¢-Ds and 40 thermal cycles were performed. To detect Avr9 andglyceraldehyde-3-phosphate dehydrogenase (GAPDH) transcripts,PCR was performed using AVR9F and AVR9R2, or GAPDHF andGAPDHR, respectively, in a 15 ll reaction and 30 thermal cycleswere performed. All amplifications were performed using eitherBIOTAQ Red DNA polymerase (Bioline, Sydney, Australia) orREDTaq DNA polymerase (Sigma-Alrich, St Louis, MO, USA),according to the manufacturer’s instructions. To detect Cf-9tr andCf-9 expression, PCR was performed using Hcr9F1 and Hcr9R1, orHcr9F2 and Hcr9R1 respectively. Polymerase chain reactions werecarried out in 15 ll volumes using Platinum Taq DNA polymeraseHigh Fidelity (Invitrogen) and 40 thermal cycles were performed.Hcr9F2/Hcr9R1 products were digested in a 35 ll volume containing10 U BstBI and incubated at 65�C for 2 h. The primers used in RT-PCR are shown in Table 5.
Acknowledgements
We thank Sara Perkins for plant care in Norwich, and Steve Demp-sey for plant care in Canberra. We thank Peter Scott and AndrewDavis for photography in Norwich and Jeff Wilson for photography
Table 4 Primers used for site-directedmutagenesis of Cf-9 Primera Sequence (5¢ fi 3¢)b Mutation
CTD219 GCGAAAGGTATTCGTTCTAGACAAC-CACGCATTGTGGG E499V, R500stop in LRR18CTD20 GTGTCATTCTAGATATTGTTACC M561I, L562stop in LRR20CTD21 TTGATCTCTAGATTAAAATCTTC S580I, L581stop in LRR21CTD102 CCCTGAAGTCTAGATGGGACC K591stop in LRR21CTD75 CAAGAATCTAGAGTCACATAAACAAG G601stop in LRR22CTD22 ATTCCCTCTAGATCAATTAGATGAT G611stop in LRR22CTD23 CATCAATTTTCTAGATGGTTTGCAA M627I, K628stop in LRR23
aPrimers are in the antisense orientation with respect to the Cf-9 coding sequence.bAltered bases are in bold, the introduced stop codons are underlined and introduced XbaI sitesare in italics.
Table 5 Primers used for RT-PCR analysisPrimer Sequence (5¢ fi 3¢) Region amplified Primer orientationa
3¢-Ds CCCATCCTACTTTCATCCCTG Ds 3¢ fi5¢-Ds CGTTACCGACCGTTTTCATCC Ds 5¢ ‹F6 GAGTTCAAGTCCAAAACATTAAGTG Cf-9 LRR14/15 fiHcr9F1 GCACGGGAATAAGCTAACGG Cf-9 LRR19 fiHcr9F2 AAAGTTGGAACACATAATTACTTC Cf-9 domain G ‹F90 CAAGCAACTGGGAACTCGGAAATC Cf-9 3¢-UTR ‹Hcr9R1 GAACATATCTACGTGCCATAGAC Cf-9 3¢-UTR ‹AVR9F GTTCTCTTTTCACAATTGCCTTC Avr9 transgene fiAVR9R2 CTCAATGTACACATTGAAGC Avr9 transgene ‹GAPDHF TGTTGTGGGTGTCAACGAG GAPDH fiGAPDHR GATGTTGAATGAAGCAGCTC GAPDH ‹
aArrows indicate primer orientation with fi indicating sense and ‹ indicating antisenseorientation relative to the coding sequence.
Interfering mutations of Cf-9 397
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 46, 385–399

in Canberra. The Sainsbury Laboratory is supported by the GatsbyCharitable Foundation and the UK Biotechnology and BiologicalSciences Research Council (BBSRC).
Supplementary Material
The following supplementary material is available for this articleonline:Table S1 This table contains a detailed analysis of the segregationdata showing that the developmental phenotype associated withinterference requires Cf-9 as well as Cf-9tr and Avr9Table S2 This table contains a detailed analysis of the segregationdata showing that interference with the M205 phenotype conferredby the weakly autoactiveHcr9-M205 gene requires Cf-9tr but not Cf-9or Avr9Figure S1. This figure contains an alignment of the conserved C-terminal regions of the Cf-2 and Cf-9 proteins and highlights theconservation of non-consensus residues in the LRR domains.Figure S2. This figure contains images of plants showing develop-mental abnormalities caused by somatic restoration of Cf-9/Avr9-dependent necrosis due to somatic excision of a Ds elementinserted at various positions in Cf-9.This material is available as part of the online article from http://www.blackwell-synergy.com
References
Avery Ince, B., Zhuang, Y., Wrenn, C.K., Shapiro, D.J. and Katze-
nellenbogen, B.S. (1993) Powerful dominant negative mutants ofthe human estrogen receptor. J. Biol. Chem. 268, 14026–14032.
Barker, C.L., Talbot, S.J., Jones, J.D.G. and Jones, D.A. (2006) Atomato mutant that shows stunting, wilting, progressive necrosisand constitutive expression of defence genes contains a recom-binant Hcr9 gene encoding an auto-active protein. Plant J.DOI: 10.1111/j.1365-313X.2006.02698.x.
Belfanti, E., Silfverberg-Dilworth, E., Tartarini, S., Patocchi, A.,
Barbieri, M., Zhu, J., Vinatzer, B.A., Gianfranceschi, L., Gessler, C.
and Sansavini, S. (2004) The HcrVf2 gene from a wild appleconfers scab resistance to a transgenic cultivated variety. Proc.Natl Acad. Sci. USA, 101, 886–890.
Dangl, J.L. and Jones, J.D.G. (2003) Plant pathogens and integrateddefence responses to infection. Nature, 411, 826–833.
De Wit, P.J.G.M. and Spikman, G. (1982) Evidence for theoccurrence of race- and cultivar specific elicitors of necrosis inintercellular fluids of compatible interactions of Cladosporiumfulvum (syn. Fulvia fulva) and tomato. Plant Physiol. 77, 642–647.
Dievart, A. and Clark, S.E. (2004) LRR-containing receptors regula-ting plant development and defense. Development, 131, 251–261.
Dievart, A., Dalal, M., Tax, F.E., Lacey, A.D., Huttly, A., Li, J. and
Clark, S.E. (2003) CLAVATA1 dominant-negative alleles revealfunctional overlap between multiple receptor kinases thatregulate meristem and organ development. Plant Cell, 15,1198–1211.
Dinesh-Kumar, S.P., Tham, W.H. and Baker, B.J. (2000) Structure-function analysis of the tobacco mosaic virus resistance gene N.Proc. Natl Acad. Sci. USA, 97, 14789–14794.
Dixon, M.S., Jones, D.A., Keddie, J.S., Thomas, C.M., Harrison, K.
and Jones, J.D.G. (1996) The tomato Cf-2 disease resistance locuscomprises two functional genes encoding leucine-rich repeatproteins. Cell, 84, 451–459.
Dixon, M.S., Hatzixanthis, K., Jones, D.A., Harrison, K. and Jones,
J.D.G. (1998) The tomato Cf-5 disease resistance gene and six
homologs show pronounced allelic variation in leucine-rich re-peat copy number. Plant Cell, 10, 1915–1925.
Hammond-Kosack, K.E. and Jones, J.D.G. (1993) Incomplete dom-inance of tomato Cf genes for resistance to Cladosporium fulvum.Mol. Plant Microbe Interact. 7, 58–70.
Hammond-Kosack, K.E. and Jones, J.D.G. (1996) Resistance gene-dependent plant defense responses. Plant Cell, 8, 1773–1791.
Hammond-Kosack, K.E., Harrison, K. and Jones, J.D.G. (1994)Developmentally regulated cell death on expression of thefungal avirulence gene Avr9 in tomato seedlings carrying thedisease-resistance gene Cf-9. Proc. Natl Acad. Sci. USA, 91,10445–10449.
Hammond-Kosack, K.E., Tang, S., Harrison, K. and Jones, J.D.G.
(1998) The tomato Cf-9 disease resistance gene functions in to-bacco and potato to confer responsiveness to the fungal aviru-lence gene product Avr9. Plant Cell, 10, 1251–1266.
van der Hoorn, R.A.L., Roth, R. and de Wit, P.J.G.M. (2001) Identi-fication of distinct specificity determinants in resistance proteinCf-4 allows construction of a Cf-9 mutant that confers recognitionof avirulence protein AVR4. Plant Cell, 13, 273–285.
Jones, D.A. and Jones, J.D.G. (1997) The role of leucine-rich repeatproteins in plant defences. Adv. Bot. Res. 24, 89–167.
Jones, D.A., Thomas, C.M., Hammond-Kosack, K.E., Balint-Kurti,
P.J. and Jones, J.D.G. (1994) Isolation of the tomato Cf-9 gene forresistance to Cladosporium fulvum by transposon tagging. Sci-ence, 266, 789–793.
van Kan, J.A.L., van den Ackerveken, G.F.J.M. and de Wit, P.J.G.M.
(1991) Cloning and characterization of cDNA of avirulence geneavr9 of the fungal pathogen Cladosporium fulvum, causal agentof tomato leaf mold. Mol. Plant Microbe Interact. 4, 52–59.
Kooman-Gersmann, M., Honee, G., Bonnema, G. and de Wit,
P.G.J.M. (1996) A high-affinity binding site for the AVR9 peptideelicitor of Cladosporium fulvum is present on plasmamembranesof tomato and other Solanaceous plants. Plant Cell, 8, 929–938.
Kunkel, T.A., Roberts, J.D. and Zakour, R.A. (1987) Rapid and effi-cient site-specific mutagenesis without phenotypic selection.Methods Enzymol. 154, 367–382.
Luderer, R., Rivas, S., Nurnberger, T. et al. (2001) No evidence forbinding between resistance gene product Cf-9 of tomato andavirulence gene product AVR9 of Cladosporium fulvum. Mol.Plant Microbe Interact. 14, 867–876.
Nagaya, T. and Jameson, J.L. (1993) Thyroid hormone receptordimerization is required for dominant negative inhibition bymutations that cause thyroid hormone resistance. J. Biol. Chem.268, 15766–15771.
Parniske, M., Wulff, B.B.H., Bonnema, G., Thomas, C.M., Jones, D.A.
and Jones, J.D.G. (1999) Homologues of the Cf-9 disease resist-ance gene (Hcr9s) are present at multiple loci on the short arm oftomato chromosome 1. Mol. Plant Microbe Interact. 12, 93–102.
Rivas, S., Rougon-Cardoso, A., Smoker, M., Schauser, L., Yoshioka,
H. and Jones, J.D.G. (2004) CITRX thioredoxin interacts with thetomato Cf-9 resistance protein and negatively regulates defence.EMBO J. 23, 2156–2165.
Schneider, D.S., Hudson, K.L., Lin, T.Y. and Anderson, K.V. (1991)Dominant and recessive mutations define functional domains ofToll, a transmembrane protein required for dorsal-ventral polarityin the Drosophila embryo. Genes Dev. 5, 797–807.
Shpak, E.D., Lakeman, M.B. and Torii, K.U. (2003) Dominant-neg-ative receptor uncovers redundancy in the Arabidopsis ERECTAleucine-rich repeat receptor-like kinase signalling pathway thatregulates organ shape. Plant Cell, 15, 1095–1110.
Song, W.Y., Wang, G.L., Chen, L.L. et al. (1995) A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21.Science, 270, 1804–1806.
398 Claire L. Barker et al.
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 46, 385–399

Sun, X., Cao, Y., Yang, Z., Xu, C., Li, X., Wang, S. and Zhang, Q.
(2004) Xa26, a gene conferring resistance to Xanthomonas oryzaepv. oryzae in rice, encodes an LRR receptor kinase-like protein.Plant J. 37, 517–527.
Tao, Y., Yuan, F., Leister, R.T., Ausubel, F.M. and Katagiri, F. (2000)Mutational analysis of the Arabidopsis nucleotide binding site-leucine-rich repeat resistance gene RPS2. Plant Cell, 12, 2541–2554.
Thomas, C.M., Jones, D.A., Parniske, M., Harrison, K., Balint-Kurti,
P.J., Hatzixanthis, K. and Jones, J.D.G. (1997) Characterisation ofthe tomato Cf-4 gene for resistance to Cladosporium fulvumidentifies sequences that determine recognitional specificity inCf-4 and Cf-9. Plant Cell, 9, 2209–2224.
Tor, M., Brown, D., Cooper, A., Woods-Tor, A., Sjolander, K., Jones,
J.D.G. and Holub, E.B. (2004) Arabidopsis downy mildew resist-ance gene RPP27 encodes a receptor-like protein similar to CLA-VATA2 and tomato Cf-9. Plant Physiol. 135, 1100–1112.
Torii, K.U., Mitsukawa, N., Oosumi, T., Matsuura, Y., Yokoyama, R.,
Whittier, R.F. and Komeda, Y. (1996) The Arabidopsis ERECTAgene encodes a putative receptor protein kinase with extracellularleucine-rich repeats. Plant Cell, 8, 735–746.
Wulff, B.B.H., Thomas, C.M., Smoker, M., Grant, M. and Jones,
J.D.G. (2001) Domain swapping and gene shuffling identify se-quences required for induction of an Avr-dependent hypersensi-tive response by the tomato Cf-4 and Cf-9 proteins. Plant Cell, 13,255–272.
Interfering mutations of Cf-9 399
ª 2006 The AuthorsJournal compilation ª 2006 Blackwell Publishing Ltd, The Plant Journal, (2006), 46, 385–399
Top Related
Copyright © 2022 FDOKUMEN

