







Bahasa
Halaman
Hukum


Plasmid 50 (2003) 12–27
www.elsevier.com/locate/yplas
Display and release of the Plasmodium falciparumcircumsporozoite protein using the autotransporter MisL
of Salmonella enterica
Patricia Ruiz-Olvera,a,b Fernando Ruiz-P�eerez,a Nicol�aas Villegas Sepulveda,c
Araceli Santiago-Machuca,a Rogelio Maldonado-Rodr�ııguez,b
Guadalupe Garcia-Elorriaga,a and C�eesar Gonz�aalez-Bonillaa,*
a Unidad de Investigaci�oon M�eedica en Inmunolog�ııa e Infectolog�ııa, Hospital de Infectolog�ııa ‘‘Dr. Daniel M�eendez Hern�aandez,’’Centro M�eedico ‘‘La Raza,’’ IMSS, Mexico, D.F.
b Escuela Nacional de Ciencias Biol�oogicas del IPN, M�eexico, D.F.c Biomedicina Molecular, Centro de Investigaci�oon y de Estudios Avanzados del IPN, M�eexico, D.F.
Received 17 February 2003, revised 1 May 2003
Abstract
TheSalmonella entericaMisL (protein ofmembrane insertion and secretion) is an autotransporter with high homology
to AIDA-I (adhesin involved in diffuse adherence) of enteropathogenic Escherichia coli. Considering that it has been re-
ported that theMisL b translocator domain is able to display heterologous passenger peptides to the bacterial surface, wedeveloped a system to display proteins and release them to the external environment by means of proteolytic cleavage.
Plasmids were constructed encoding 8 or 53 repeats of the NANP (Asp–Ala–Asp–Pro) tetrapeptide, which is the main B
cell epitope of thePlasmodium falciparum circumsporozoitic protein (CSP), fused to the theMisL b-domain and includingthe recognition cleavage sequence from the E. coli OmpT surface protease. E. coli XL-10Gold and BL21(DE3) (OmpT
positive and negative, respectively) and Salmonella enterica serovar Typhimurium SL3261 (Aro A�) were transformed
with the plasmids and, both expression and localization of the fusion proteins were assessed by Western blot, indirect
immunofluorescence, and flow cytometry, using a monoclonal antibody against (NANP)3. Higher expression of the
(NANP)8 and (NANP)53 fusion proteins was demonstrated on the bacterial surface of the OmpT negative E. coli strains
and the (NANP)53 in the culture supernatant of E. coli XL-10Gold indicating a protease mediated cleavage. The flow
cytometry analysis suggested 71 and 98% cleavage efficiency for the (NANP)8 and (NANP)53, respectively, in E. coliXL-
10Gold. Similar results were obtained in S. enterica serovar Typhimurium SL3261, suggesting the involvement of other
proteases related to OmpT. These results demonstrate thatMisLmay be used for the autodisplay and release of passenger
proteins in attenuated Salmonella or E. coli strains, which may have several applications in vaccine design.
� 2003 Published by Elsevier Science (USA).
Keywords: Autodisplay; Salmonella enterica; MisL; Plasmodium falciparum; Live vector vaccines; Circumsporozoite protein
* Corresponding author. Fax: (55)5583-0626.
E-mail address: [email protected] (C. Gonz�aalez-Bonilla).
0147-619X/03/$ - see front matter � 2003 Published by Elsevier Science (USA).
doi:10.1016/S0147-619X(03)00047-7

P. Ruiz-Olvera et al. / Plasmid 50 (2003) 12–27 13
1. Introduction
Gram negative bacteria have evolved five differ-
ent mechanisms to secrete proteins to the external
environment. During the transportation process
from the cytosol to the bacterial surface, secreted
proteins have to travel to the internal membrane,
the periplasmic space, reach the outer membrane,
and frequently be processed and released to theexternal milieu. Proteins secreted by the type I, III,
and IV systems reach the bacterial surface through a
channel, which crosses both the inner and outer
membranes in a one step process without the inter-
vention of periplasmic chaperone proteins, whereas
proteins belonging to the type II and V systems are
first translocated to the periplasmic space following
the general secretion pathway (GSP) and thenthrough the outer membrane (Burns, 1999; Sal-
mond and Reeves, 1993; Sharff et al., 2001; Stath-
opoulos et al., 2000). The type IV secretion system is
similar the type III, although both may involve cell
to cell transfer, type IV systemmediate transfer of a
variety of biomolecules specially DNA–protein
complexes between bacterial cells or between bac-
teria and eukaryotic cells. The type V system com-prises the proteins also known as autotransporters
because they form their own pore to reach the outer
membrane by means of three different functional
domains. Classic autotransporters contain a N-
terminal leader peptide which allows the protein to
cross from the inner membrane to the periplasmic
space by a sec dependent mechanism. The leader
sequence is trimmed and the C-terminal transloca-tor b-domain forms a barrel which is inserted in theouter membrane, and finally, the N-terminal pas-
senger a-domain which travels through the pore tothe external environment and contains the biologi-
cal activity of the protein. In some autotransporters
the pore appears to be amonomer; like in the BrkA,
which is a 103 kDa outer membrane protein of
Bordetella pertussis that mediates resistance to an-tibody-dependent killing by complement (Shannon
and Fernandez, 1999). However, it has been dem-
onstrated that some autotransporters form more
complicated pores, the IgA protease fromNeisseria
gonorrhoeae assembles on the bacterial surface
forming a multimeric translocation channel (Veiga
et al., 2002). Once on the surface, the passenger
domain may remain anchored to the outer mem-brane or be cleaved by auto proteolytic mechanisms
or by the action of bacterial surface proteases (Fink
et al., 2001).
Autotransporters comprise a wide variety of
proteins related with bacterial virulence; which in-
clude IgA proteases of N. gonorrhoeae, Neisseria
menigitidis or Haemophilus influenzae (Henderson
and Nataro, 2001; Jose et al., 1995; Klauser et al.,1993; Maurer et al., 1997; Poulsen et al., 1992)
serine proteases from Enterobacteriaceae (Al Ha-
sani et al., 2000; Dozois et al., 2000; Guyer et al.,
2000; Henderson et al., 1999), adhesins from E. coli
(Benz and Schmidt, 1989; Leininger et al., 1991;
Lindenthal andElsinghorst, 1999; St III andCutter,
2000) and other virulence factors, such as VirG of
Shigellawhich is involved in the bacterial spreadingthrough epithelial cells (Suzuki et al., 1995).
Due to the relative simplicity of the their
transporting mechanism, the b-domains from
several autotransporters have been employed to
express recombinant ‘‘passenger’’ proteins on the
surface of enterobacteria with the idea to induce or
enhance humoral immune responses. These in-
clude the E. coli adhesin involved in diffuse ad-herence (AIDA-I), which has been demonstrated
to promote bacterial surface expression of the
peptide antigen tag PEYFK, the cholera toxin B
subunit (CtxB) (Maurer et al., 1997), and an en-
zymatically active b-lactamase (Lattemann et al.,2000); the N. gonorrhoeae IgA protease which was
also used to display the cholera toxin B subunit in
Salmonella typhimurium (Klauser et al., 1990).Although the factors determining efficient surface
expression of foreign antigens are still under in-
vestigation, the general notion is that protein size
and conformation are two main factors influencing
the translocation process. Large proteins, tightly
folded or those with internal disulfide bonds may
be difficult to translocate through the b-domain,thus the N. gonorrhoeae IgA protease b-domainwas able to translocate the CtxB only when the
disulphide induced folding in the periplasm was
prevented (Klauser et al., 1990). On the other
hand, it has been demonstrated that the N. gon-
orrhoeae IgA protease b-domain is able to pro-mote bacterial surface expression of single chain
variable antibody domains (scFv), although at a

14 P. Ruiz-Olvera et al. / Plasmid 50 (2003) 12–27
reduced level, in an active conformation with itsdisulphide bonds preformed in the periplasm
(Veiga et al., 1999). Once the passenger protein is
displayed on the bacterial surface, it can be pro-
cessed by surface proteases and released to the
external environment.
The E. coli OmpT protease has been success-
fully used for this purpose as demonstrated by a
CtxB fused to the AIDA-1 which was cleavedwhen expressed in E. coli UT2300 (OmpT+), but
not in E. coli UT5600 or JK321 (OmpT-dsbA-)
(Maurer et al., 1997).
MisL (protein of membrane insertion and se-
cretion) is a protein of unknown function located in
the pathogenicity island SPI-3 of Salmonella ent-
erica, which has been proposed as an autotrans-
porter due to its high homology with AIDA-I. Wehave reported previously that the MisL b-domainis indeed an autotransporter, because it was able to
translocate four repeats of the tetrapetide NANP
(asp–Ala–Asp–Pro) to the bacterial surface, which
is the main B cell epitope of the Plasmodium falci-
parum circumsporozoitic protein (CSP) and more
importantly that antibodies raised against this
peptide were able to recognize the native protein onthe sporozoite surface (Ruiz-Perez et al., 2002).
Since it has been suggested that released antigens
may induce better antibody responses (Hess et al.,
1996) we designed a system to autodisplay larger
proteins and release them to the external milieu, by
adding cleavage sequences recognized by OmpT
between the translocator b-domain of MisL and
the passenger protein.
2. Material and methods
2.1. Bacterial strains, plasmids, and oligonucleotides
The bacterial strains, plasmids, and oligonucle-
otides used in this work are described in Table 1.
2.2. Culture and induction conditions
Escherichia coli strains transformed with plas-
mids pRO-NANP, pRO-T-NANP, pRO-CS,
pRO-T-CS, pnirBMisL or pUC19-CSP were cul-
tured in BHI agar plates at 37 �C with 100 lg/mlampicillin. S. enterica serovar Typhimurim was
supplemented with 0.01% 2,3-dihydroxybenzoicacid (DHB) (Sigma, St. Louis, MO). In order to
induce the nirB promoter, a single colony was in-
oculated in BHI broth supplemented with ampi-
cillin and DHB when necessary, and cultured with
agitation (200 rpm). Once the culture reached late
logarithmic growth phase (OD600 nm¼ 1.0), 150 ll(1.5� 108 cells) were transferred to 50ml of thio-glycolate medium (Difco Laboratories, Detroit,MI) and induced anaerobically at 37 �C in a BBLGasPak Anaerobic System (Becton–Dickinson
Microbiology Systems, Cockeysville, MD). Cul-
ture required 6 or 8 h for E. coli and Salmonella
strains, respectively, for IFA and FACS to ob-
serve maximum surface expression (optimal con-
ditions were determined in several experiments).
The bacteria were harvested after 5 or 6 h ofculture for E. coli and Salmonella strains, respec-
tively, for Western and dot blot. Bacterial strains
transformed with plasmid pUC19-CSP were in-
duced with 1mM IPTG for 3 h at 37 �C in aerobicconditions.
2.3. Genetic engineering
DNA handling was performed following the
recommendations of Sambrook (Sambrook and
Russell, 2001). Plasmids encoding the fusion pro-
teins MisLb-NANP and MisLb-CSP, including ornot the OmpT cleavage site, were constructed as
depicted in Figs. 1 and 2. First, pRO-NANP, con-
taining eight repeats of the tetrapeptideNANP,wasderived from pnirBMisL-NANP, which contains
four repeats of NANP following the strategy de-
scribed before (Ruiz-Perez et al., 2002). Briefly,
pnirBMisL-NANP was digested with NheI and
BamHI resulting in two fragments, a 1318 bp frag-
ment containing the MisL b-domain, which wassaved and a 2687 bp fragment, which was ligated to
a pair of complementary oligonucleotides (Nanp-1and Nanp-2) containing four repeats of the NANP
sequence flanked by open XbaI and BamHI re-
striction sites (from 50 to 30 ends) and internal NheI
and NsiI sites in the 30 end. The resulting plasmid
pRO-N was digested with NheI and BamHI and
then ligated to the 1318 bp fragment containing the
MisL b-domain (Figs. 1A and B). The resulting
plasmid pRO-NANP contains an extra NANP4
Table 1
Bacterial strains, plasmids and oligonucleotides used in this work
Name Characteristics Source
Bacterial strain
E. coli DH5a (sup E44, Dlac 169,
f80lacZDM15hsdR17, recA1endA1,
gyrA96thi-1relA1)
Laboratory collection
E. coli
BL21(DE3)
(B F-dcm ompT hsdS [rB-mB]) Laboratory collection
E. coli
XL-10Gold
(TetR D (mcrA)183D(mcrCB-
hsdSMR-mrr)173 endA1 supE44
thi-1 recA1 gyr A96 relA1 lac Hte
[F 0 pro AB lac Iq ZDM15 Tn10
(tetRÞ Amy CamR]a
Laboratory collection
S.enterica
serovar
Typhimurium
SL3261
(aroA::Tn10) Nature, 1981, 291, 238–239
Name Relevant characteristics Approximate molecular weight
(kDa)
Size (kb) Source or reference
Plasmids
pnirBMisL Cloning vector with nir B promoter,
signal sequence and translocator bMisL domain
55 3.9 Infect. Immun., 2002, 70, 3611–3620
pnirBMisL-
NANP
Derived from pnirLTBMisL bearing
four NANP repeats
57 4 Infect. Immun., 2002, 70, 3611–3620
pRO-NANP Derived from pnirLTBMisL bearing
eight NANP repeats
60 4 This work
pRO-CS Derived from pRO-NANP bearing a
truncated CSP protein
82 4.7 This work
pRO-T Cloning vector with the nir B pro-
moter, signal sequence and SCOTbUndetermined 2.7 This work
pRO-T-
NANP
Derived from pRO-T, bearing eight
NANP repeats, SCOT and the bMisL domain
62 4.1 This work
pRO-T-CS Derived from pRO-T-NANP, bear-
ing a truncated CSP protein, SCOT
and the b MisL domain
85 4.8 This work
pUC19-CSP Derived from pUC19, produces a
cytosolic CSP protein
60a 3.9 J. Infect. Dis. 1994, 169, 927–931
P.Ruiz-O
lveraetal./Plasmid50(2003)12–27
15
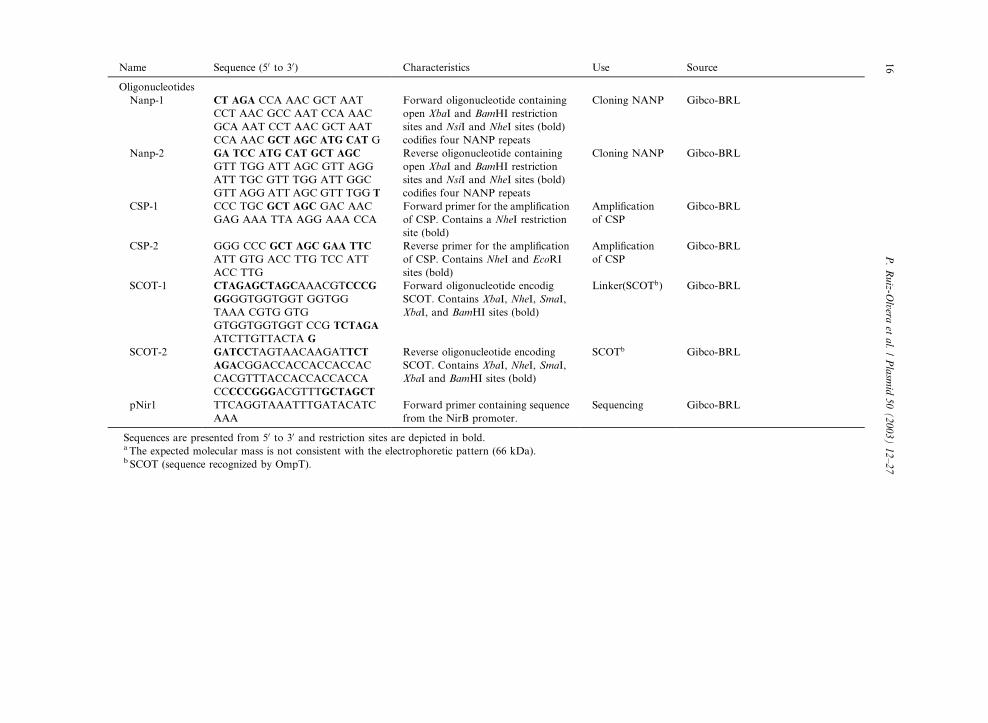
Name Sequence (50 to 30) Characteristics Use Source
Oligonucleotides
Nanp-1 CT AGA CCA AAC GCT AAT
CCT AAC GCC AAT CCA AAC
GCA AAT CCT AAC GCT AAT
CCA AAC GCT AGC ATG CAT G
Forward oligonucleotide containing
open XbaI and BamHI restriction
sites and NsiI and NheI sites (bold)
codifies four NANP repeats
Cloning NANP Gibco-BRL
Nanp-2 GA TCC ATG CAT GCT AGC
GTT TGG ATT AGC GTT AGG
ATT TGC GTT TGG ATT GGC
GTT AGG ATT AGC GTT TGG T
Reverse oligonucleotide containing
open XbaI and BamHI restriction
sites and NsiI and NheI sites (bold)
codifies four NANP repeats
Cloning NANP Gibco-BRL
CSP-1 CCC TGC GCT AGC GAC AAC
GAG AAA TTA AGG AAA CCA
Forward primer for the amplification
of CSP. Contains a NheI restriction
site (bold)
Amplification
of CSP
Gibco-BRL
CSP-2 GGG CCC GCT AGC GAA TTC
ATT GTG ACC TTG TCC ATT
ACC TTG
Reverse primer for the amplification
of CSP. Contains NheI and EcoRI
sites (bold)
Amplification
of CSP
Gibco-BRL
SCOT-1 CTAGAGCTAGCAAACGTCCCG
GGGGTGGTGGT GGTGG
TAAA CGTG GTG
GTGGTGGTGGT CCG TCTAGA
ATCTTGTTACTA G
Forward oligonucleotide encodig
SCOT. Contains XbaI, NheI, SmaI,
XbaI, and BamHI sites (bold)
Linker(SCOTb) Gibco-BRL
SCOT-2 GATCCTAGTAACAAGATTCT
AGACGGACCACCACCACCAC
CACGTTTACCACCACCACCA
CCCCCGGGACGTTTGCTAGCT
Reverse oligonucleotide encoding
SCOT. Contains XbaI, NheI, SmaI,
XbaI and BamHI sites (bold)
SCOTb Gibco-BRL
pNir1 TTCAGGTAAATTTGATACATC
AAA
Forward primer containing sequence
from the NirB promoter.
Sequencing Gibco-BRL
Sequences are presented from 50 to 30 and restriction sites are depicted in bold.a The expected molecular mass is not consistent with the electrophoretic pattern (66 kDa).b SCOT (sequence recognized by OmpT).
16
P.Ruiz-O
lveraetal./Plasmid50(2003)12–27

Fig. 1. Construction of plasmids pRO-NANP (A–B) and pRO-T-NANP (B–C). These plasmids encode for a fusion protein constituted
by theLTB signal sequence, eight repeats of the tetrapeptideNANPwhich is themainB cell epitope of theP. falciparumCSPand theMisL
b-domain. These constructs are under the control of the NirB promoter. In pRO-T-NANP a linker sequence containing two OmpTcleavage sites (K/R) was included between the passenger NANP8 peptide andMisL b-domain. The addition of new passenger sequenceswas achieved by ligation of double stranded oligonucleotides flanked with open restriction sites to compatible ends in the plasmid.
P. Ruiz-Olvera et al. / Plasmid 50 (2003) 12–27 17
sequence and downstream a NheI site. An adapter
oligonucleotide containing two OmpT recognitionK/R sites (designed here as SCOT) was inserted
downstream of the NANP encoding sequence fol-
lowing the same strategy. Plasmid pRO-NANPwas
digested with NheI and BamHI and the 1318 bp
fragment saved. The 2747 bp fragment was ligated
to a pair of complementary oligonucleotides
(SCOT-1 and SCOT-2) designedwith flankingXbaI
and BamHI restriction sites. The resulting plasmidpRO-T was ligated to the 1318 bp fragment previ-
ously obtained, generating plasmid pRO-T-NANP
(Figs. 1B and C). Due to the cloning strategy the
(NANP)8 peptide contains two flanking and one
internal alanine and arginine residues, which do not
correspond to the wild-type CSPB cell epitope from
P. falciparum (Fig. 2B). A truncated CSP encoding
amino acid residues 101–323, was amplified by PCRfrom pUC19-CSP using primers CSP-1 and CSP-2,
which are described in the Table 1. The amplifica-
tion was performed from 1pg of DNA, 1 pM of
primers, 200 lMdNTPs 1UVentDNApolymerase
(Biolabs), first denaturing at 94 �C for 5min, and
then following 30 cycles of annealing 66 �C for 1min, extension at 72 �C for 1min and denaturing
95 �C for 1min with a final extension at 72 �C(Thermocyclator Amplitron II, Thermoline). The
PCR product (669 bp) was inserted in the NheI
restriction site of plasmids pRO-NANP and pRO-
T-NANP resulting plasmids pRO-CS and pRO-T-
CS, respectively (Fig. 2A). Both constructions
contain 53 NANP repeats of the CSP protein fromP. falciparum. Fig. 2B shows the SCOT nucleo-
tide sequence. Figs. 2C and D show the predicted
amino acid sequence of the (NANP)8 peptide
and the CSP, respectively. Finally, all clones were
sequenced. Briefly, 1 lg of plasmid DNA was
sequencedwith theDNABigDyeTerminatorCycle
Sequencing Kit (Applied Biosystems Foster City,
CA, USA) using 8 pM of primer pNir1. The am-plification reaction was purified by CENTRISEP
Spin columns (Applied Biosystems) and examined
in a PE AbiPrism 310 genetic analyzer (Applied
Biosystems).

Fig. 2. Construction of plasmids pRO-CS and pRO-T-CS. These plasmids contain a fusion synthetic gene encoding the LTB signal
sequence, eight NANP repeats, a truncated CSP from P. falciparum, and the MisL b-domain. pRO-T-CS has an adapter with twoOmpT cleavage sites (K/R) between the CSP and MisL b-domain. (A) Ligation of the CSP to the plasmids containing eight repeats ofNANP. (B,C) Sequence containing OmpT cleavage sites (SCOT), nucleotides and amino acid translation, respectively. (D) Translated
amino acids sequence of the CSP.
18 P. Ruiz-Olvera et al. / Plasmid 50 (2003) 12–27
2.4. Bacterial fractions
Different bacterial fractions were obtained from
E. coli XL-10Gold and BL21(DE3) transformedwith plasmids pRO-NANP pRO-CS, pRO-T-
NANP, pRO-T-CS, pnirBMisL or pUC19-CSP.
Briefly, 50ml of bacterial cultures obtained under
inducing conditions were harvested by centrifu-
gation at 5000g/5min, washed twice with PBS, pH
7.4 and adjusted to 1� 109 cells/ml (OD600 nm¼1.0). Crude protein preparations consisting of
solubilized bacteria, were obtained by mixing
1� 108 cells with 100 ll of sample buffer (0.5MTris, pH 6.8, 2% SDS, 5% b-mercaptoethanol,10% glycerol, and 0.1% bromophenol blue), boil-
ing for 10min and collecting the supernatant aftercentrifugation at 13,000g for 2min. Periplasmic
proteins were obtained from 1� 1010 cells resus-pended in 10ml of 30mM Tris–HCl with 20%
sucrose, pH 8.0, in 1mM EDTA. The cells were
incubated in agitation for 10min at room tem-
perature, the suspension was centrifuged at 8000g/
10min, the pellet was resuspended in 300 ll cold5mM MgSO4 and shaken on ice for 10min. The

P. Ruiz-Olvera et al. / Plasmid 50 (2003) 12–27 19
suspension was centrifuged at 8000g/10min at 4 �Cand the supernatant containing the periplasmic
fraction was collected. In order to obtain outer
membrane proteins (OMP), 1� 1010 cells were re-suspended in 10ml of 10mM Tris–HCl, pH 8.0,
and 10mM EDTA and then disrupted in a French
press (20,000 psi, twice). Undisrupted cells were
removed by centrifugation and the supernatant
was incubated with 1% Triton X-100 for 30minand centrifuged to 100,000g for 60min at 4 �C. Thepellet containing OMP was dissolved in 10mM
Tris–EDTA. Protein concentration was quantified
in all bacterial fractions and 40 lg samples werefurther analyzed by electrophoresis under reducing
conditions. Culture supernatants were prepared as
described (Hess et al., 1990) with some modifica-
tions. Briefly, the different bacterial strains werecultured overnight at 37 �C on agar plates sup-
plemented with ampicillin. Then, single colonies
were inoculated in 40ml of thioglycolate and cul-
tured in anaerobic conditions for 6 h. Culture su-
pernatants were prepared by first removing the
cells by centrifugation at 1500g for 15min at 4 �Cafter adding trichloroacetic acid (TCA) to a final
concentration of 10% (w/v). Precipitated proteinswere harvested by centrifugation at 13,000g for 1 h
at 4 �C. The samples were neutralized using a Tris-saturated solution and finally the proteins were
solubilized in SDS sample buffer (0.5M Tris, pH
6.8, 2% SDS, 5% b-mercaptoethanol, 10% glyc-
erol, and 0.1% bromophenol blue) and 50 lgsamples were further analyzed by electrophoresis
under reducing conditions.
2.5. MisLb-NANP and MisLb-CSP expression
depending on the OmpT cleavage sequence
Expression of the four fusion proteins, MisLb-CSP, MisLb-NANP, MisLb-T-CSP, and MisLb-T-NANP (with or without the OmpT cleavagesite) was evaluated by Western blot in the different
bacterial fractions obtained as described above.
Protein samples were electrophoresed in denatu-
rating polyacrylamide gels (SDS–PAGE) (Lae-
mmli, 1970) under reducing and denaturing
conditions and then stained with Coomassie blue
dye. Western and dot blot was performed by
transferring the proteins to nylon membranes
(Millipore, Bedford, MA) (Towbin et al., 1979),after blocking the membranes with PBS 5% skim
milk for 1 h at 37 �C, they were incubated for 12 hat 37 �C with 2 lg/ml of monoclonal antibody2A10 diluted in PBS 5% skim milk (2A10 recog-
nizes the NANP epitope from P. falciparum and
was kindly donated by Dr. Elizabeth Nardin from
the Department of Medical and Molecular Para-
sitology New York University School of Medicine,New York, NY). The membranes were then in-
cubated with a goat-anti mouse IgG-peroxidase
conjugate (1 lg/ml in PBS 5% skim milk) for 1 h at
37 �C. The immunological reaction was revealedwith 4-chloro-1-naphthol 30% H2O2 in PBS, pH
7.4, and between incubations the membranes
were washed with PBS 5% skim milk 0.01% Tween
20.
2.6. Evaluation of MisLb-CSP and MisLb-NANPfusion proteins surface expression determined by the
OmpT cleavage site
The surface expression of the four fusion pro-
teins, MisLb-CSP and MisLb-NANP bearing ornot the SCOT, was assessed by indirect immuno-
fluorescence (IFA) and flow cytometry in E. coli
XL-10Gold and BL21(DE3), and S. enterica ser-
ovar Typhimurium SL3261. The bacterial behav-ior was compared in the strains transformed with
these plasmids. As controls, strains transformed
with pnirBMisL and pUC19-CSP were included.
After inducing them as described in Section 2, the
bacterial strains were harvested, washed twice with
PBS, and then incubated with monoclonal anti-
body 2A10 for 30min at room temperature with
agitation. Then they were washed twice with PBSand incubated with a goat anti-mouse IgG-FITC
conjugate (10 lg/ml in PBS plus 1% BSA) for
30min at room temperature in the dark. The
washings were repeated and the samples were re-
suspended in propidium iodide–PBS (4mg/L) and
spread on slides to view them under fluorescence
microscopy (Olympus BX40). Flow cytometry was
performed with 400 lL of the cell suspension(1� 104 cells) using a Flow cytometer (Beckton–
Dickinson, Bedford, MA) and analyzed with the
Lysis II (Ver 1.1). Statistical analysis included
Kolmogorov–Smirnov test (Watson, 2001).
20 P. Ruiz-Olvera et al. / Plasmid 50 (2003) 12–27
3. Results
3.1. Construction of MisLb-NANP and MisLb-CSP fusion proteins
With the purpose to evaluate the ability of the
MisL translocator b-domain to promote bacterialsurface display of proteins with different size and
the possibility to release them to external milieuafter proteolytic cleavage, two fusion proteins
were constructed, each with two different variants
of the same repetitive tetrapeptide NANP as de-
scribed in Section 2. The MisLb-NANP containingeight repeats of the tetrapeptide NANP and the
MisLb-CSP which contains a truncated form of
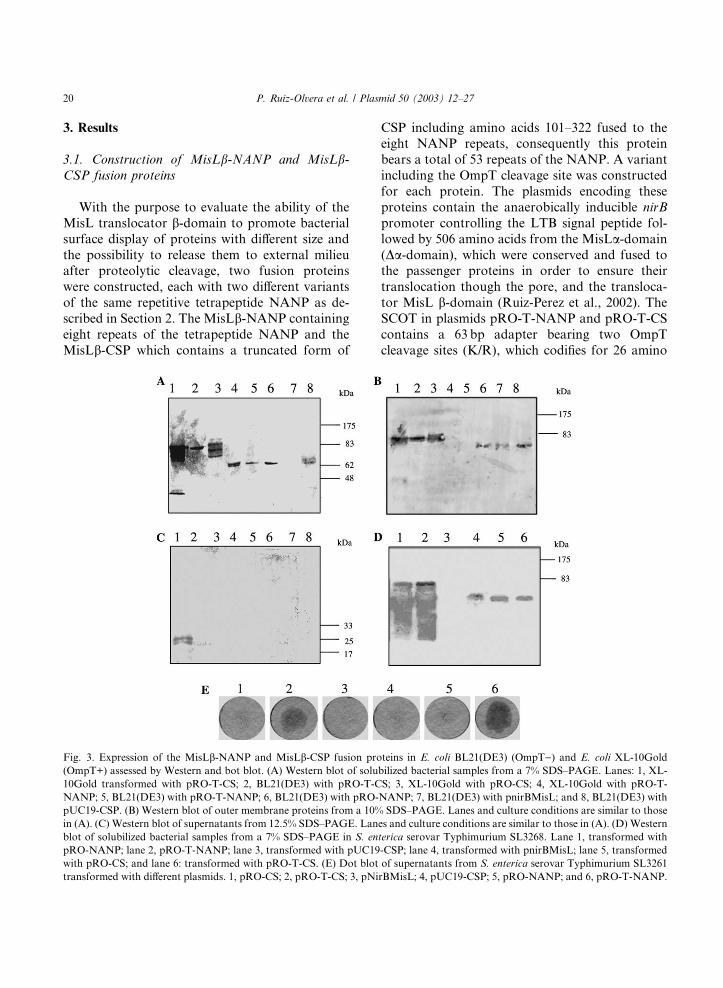
Fig. 3. Expression of the MisLb-NANP and MisLb-CSP fusion pro(OmpT+) assessed by Western and bot blot. (A) Western blot of solu
10Gold transformed with pRO-T-CS; 2, BL21(DE3) with pRO-T-C
NANP; 5, BL21(DE3) with pRO-T-NANP; 6, BL21(DE3) with pRO-
pUC19-CSP. (B) Western blot of outer membrane proteins from a 10%
in (A). (C) Western blot of supernatants from 12.5% SDS–PAGE. Lan
blot of solubilized bacterial samples from a 7% SDS–PAGE in S. en
pRO-NANP; lane 2, pRO-T-NANP; lane 3, transformed with pUC19
with pRO-CS; and lane 6: transformed with pRO-T-CS. (E) Dot blot
transformed with different plasmids. 1, pRO-CS; 2, pRO-T-CS; 3, pNi
CSP including amino acids 101–322 fused to theeight NANP repeats, consequently this protein
bears a total of 53 repeats of the NANP. A variant
including the OmpT cleavage site was constructed
for each protein. The plasmids encoding these
proteins contain the anaerobically inducible nirB
promoter controlling the LTB signal peptide fol-
lowed by 506 amino acids from the MisLa-domain(Da-domain), which were conserved and fused tothe passenger proteins in order to ensure their
translocation though the pore, and the transloca-
tor MisL b-domain (Ruiz-Perez et al., 2002). TheSCOT in plasmids pRO-T-NANP and pRO-T-CS
contains a 63 bp adapter bearing two OmpT
cleavage sites (K/R), which codifies for 26 amino
teins in E. coli BL21(DE3) (OmpT)) and E. coli XL-10Goldbilized bacterial samples from a 7% SDS–PAGE. Lanes: 1, XL-
S; 3, XL-10Gold with pRO-CS; 4, XL-10Gold with pRO-T-
NANP; 7, BL21(DE3) with pnirBMisL; and 8, BL21(DE3) with
SDS–PAGE. Lanes and culture conditions are similar to those
es and culture conditions are similar to those in (A). (D) Western
terica serovar Typhimurium SL3268. Lane 1, transformed with
-CSP; lane 4, transformed with pnirBMisL; lane 5, transformed
of supernatants from S. enterica serovar Typhimurium SL3261
rBMisL; 4, pUC19-CSP; 5, pRO-NANP; and 6, pRO-T-NANP.

P. Ruiz-Olvera et al. / Plasmid 50 (2003) 12–27 21
acid residues as described by Hanke et al. (1992)with some modifications and was inserted between
the passenger protein and the Da-domain.
3.2. The MisLb-NANP and MisLb-CSP fusion
proteins are produced in E. coli
Expression of the fusion proteins MisLb-NANPand MisLb-CSP was examined by Western blot inprotein preparations obtained from different bac-
terial compartments. E. coli XL-10 Gold (OmpT+)
and BL21(DE3) (OmpT)) were transformed withthe four different plasmids, using as controls
pUC19-CSP (cytoplasmic CSP expression) and
pnirBLTBMisL (lacks the passenger domain). The
fusion proteins consisted of the following constit-
uents, the MisL b-domain (280 amino acids,
�32 kDa), an Da-domain (226 amino acids,
�26 kDa) and the passenger domains NANP (32amino acids, �4 kDa) or CSP (223 amino acids,�25 kDa). All bacterial strains were first inducedby anaerobiosis, except for pUC19-CSP which was
induced with IPTG. The Western blot clearly
demonstrated the expression of the (NANP)8 and
CSP passenger proteins in solubilized bacteria and
OMP preparations (Fig. 3). TheMisLb-CSP fusionprotein exhibited the expected �90 kDa molecularweight in both E. coli XL-10Gold and BL21(DE3),but when this fusion protein included the OmpT
cleavage (K/R) site and was expressed in E. coli
(OmpT+) an extra band of �25 kDa was observedin the crude protein samples (Fig. 3A, lane1). This
band was not observed in OmpT negative strain
transformed with pRO-T-CS (Fig. 3A, lane 2).
These results suggest that OmpT protease mediated
the protein processing, although other proteasesmissing in E. coli BL21(DE3) such as Lon or other
expressed by E. coli XL-10Gold could be involved
in the protein cleavage.
The Western blot shows several bands suggest-
ing protein degradation by other proteases (Fig.
3A, lanes 1 and 3), However, in E. coli BL21(DE3)
(OmpT)) presented a single band (Fig. 3A, lane2).It has to be considered that the OmpT proteasemay cut also between two basic amino acids and
the CSP has a R/K sequence six amino acids from
the beginning of the cloned sequence. However,
this result is consistent with other recombinant
CSP constructs expressed by other bacterial strainswhich show several bands in Western blot (Ruiz-
Perez et al., 2002). Moreover, the fusion proteins
were not detected in periplasmic extracts, therefore
it may be suggested that they are not retained in
this compartment and may be promptly translo-
cated to the external membrane (data not shown).
The expected �60 kDa band corresponding to theMisLb-NANP fusion protein was revealed in totalprotein and OMP samples (Fig. 3A, lanes 4–6 and
3B, lanes 6–8), although the fusion protein could
not be demonstrated in the culture supernatant of
E. coli XL-10Gold transformed with the plasmid
bearing the SCOT, this may be explained by the
small size of the passenger peptide (NANP)8. As
expected, control bacterial strains transformed
with pnirBLTBMisL did not show any band be-cause this plasmid did not contain a passenger
protein (Fig. 3A, lane 7 and 3B, lane 7), while E.
coli BL21(DE3) transformed with pUC19-CSP
exhibited a band in solubilized bacterial prepara-
tions but not in OMP extracts because this plasmid
produces a cytosolic CSP (Fig. 3A lane 8 and 3B,
lane 8). When comparing all bacterial strains
transformed with pRO-T-CS (encodes for the CSPand the linker with the OmpT cleavage site), the
most important finding was that a �25 kDa bandwas revealed by the 2A10 antibody only in super-
natant precipitates from E. coli XL-10Gold
(OmpT+), thus suggesting specific OmpT mediated
proteolytic cleavage between the Da-MisL domainwith release of the (NANP)53 (Fig. 3C, lane 1).
Expression of MisLb-CSP and MisLb-NANPfusion proteins was also confirmed in Salmonella
by Western blot from solubilized bacterial samples
(Fig. 3D), when S. enterica serovar Typhimurium
SL3261 was transformed with pRO-CS or pRO-T-
CS a �83 kDa protein was demonstrated (Fig. 3D,lanes 5 and 6), whereas strains transformed with
pRO-NANP and pRO-T-NANP showed a a
�60 kDa protein (Fig. 3D, lanes 5 and 6). Inter-estingly important protein degradation was ob-
served in those Salmonella strains producing
(NANP)53. The small NANP and CSP released to
the external milieu from MisLb-NANP and Mis-Lb-CSP fusion proteins containing the SCOT weredetected by dot blot in Salmonella strains (Fig. 3E,
lanes 2 and 6).

Fig. 4. Surface expression of the MisLb-NANP and MisLb-CSP fusion proteins, and their variants bearing the OmpT cleavage se-quence, in E. coli BL21(DE3) (OmpT)) and XL-10Gold (OmpT+) strains. The (NANP)3 epitope was detected by IFA using the 2A10monoclonal antibody and a FITC-conjugated goat anti-mouse IgG as secondary antibody. (A) E. coli BL21(DE3) transformed with
pRO-T-NANP. (B) E. coli XL-10Gold transformed with pRO-T-NANP; (C) E. coli BL21(DE3) transformed with pRO-T-CS; (D) E.
coli XL-10Gold transformed with pRO-T-CS; (E) E. coli XL-10Gold transformed with pnirBMisL; (F) E. coli XL-10Gold transformed
with pUC19-CSP; (G) S. enterica serovar Typhimurium SL3261 transformed with pRO-NANP; and (H) S. enterica serovar
Typhimurium SL3261 transformed with pRO-CS.
22 P. Ruiz-Olvera et al. / Plasmid 50 (2003) 12–27
3.3. The MisLb-NANP and MisLb-CSP fusion
proteins are expressed on the bacterial surface of E.
coli and Salmonella strains
Fig. 4 compares the surface expression of the
MisLb-NANP and MisLb-CSP fusion proteins inOmpT positive or negative bacterial strains andSalmonella strains, which was assessed by IFA
performed with the 2A10 monoclonal antibody.
Controls included E. coli strains transformed with
pUC19-CSP (cytosolic CSP expression), which
should not show fluorescence, thus demonstrating
that under the experimental conditions the
monoclonal antibody recognized the (NANP)3epitope only on the bacterial surface (not shown).Clear surface expression of the fusion proteins
MisLb-NANP and MisLb-CSP containing the
SCOT was demonstrated in E. coli BL21(DE3)
(OmpT)) (Figs. 4A and C), in contrast only slightfluorescence was observed in E. coli XL-10Gold
(OmpT+) (Figs. 4B and D), this result was ex-
pected and suggests that the passenger antigen was
released from the bacterial surface. The proteinexpression in Salmonella strains were positive
the same manner that expressed in E. coli strains
(Fig. 4G and H).
The passenger protein expression on the bacte-
rial surface in OmpT positive and negative E. coli
strains was measured by flow cytometry (Fig. 5).
Whereas fluorescence intensity was similar in all
E. coli BL21(DE3) (OmpT)) transformed plas-
mids bearing the passenger protein (NANP)8(Figs. 5B and E) or (NANP)53 (Figs. 5H and K), itwas clearly decreased in those E. coli XL-10Gold
(OmpT+) strains producing the fusion proteins
with the SCOT (NANP)8 (Figs. 5A and D) and
(NANP)53 (Figs. 5G and J), thus suggesting an
OmpT dependent proteolytic surface processing.
The statistical analysis of the surface expression
comparing OmpT positive or negative bacterial
strains demonstrated 71 and 98% cleavage for theMisLb-NANP and MisLb-CSP fusion proteins,
respectively.
3.4. Expression and cleavage of MisLb-NANP andMisLb-CSP fusion proteins in S. enterica serovar
Typhimurium SL3261
Salmonella enterica serovar Typhimurium
SL3261 (a DaroA� auxotrophic mutant) was
transformed with pRO-NANP, pRO-T-NANP,
pRO-CS, and pRO-T-CS. The Western blot and

Fig. 5. Flow cytometry analysis. MisLb-NANP and MisLb-CSP fusion protein expression on the surface of different bacterial strains.(A) E. coli XL-10Gold transformed with pRO-NANP; (B) E. coli BL21(DE3) transformed with the same plasmid; (C) histogram
comparing A and B; (D) E. coli XL-10Gold transformed with pRO-T-NANP; (E) E. coli BL21(DE3) transformed with the same
plasmid; (F) histogram comparing D and E; (G) E. coli XL-10Gold transformed with pRO-CS; (H) BL21(DE3) transformed with the
same plasmid; (I) histogram comparing G and H; (J) E. coli XL-10Gold transformed with pRO-T-CS; (K) E. coli BL21(DE3)
transformed with the same plasmid; and (L) histogram comparing (J) and (K).
P. Ruiz-Olvera et al. / Plasmid 50 (2003) 12–27 23
IFA of these strains were consistent with the be-
havior of the fusion proteins in the E. coli (datanot shown). The flow cytometry analysis demon-
strated that both the (NANP)8 and (NANP)53passenger proteins were expressed on the bacterial
surface of Salmonella and that they were processed
when the SCOT was included (Fig. 6). The cleav-
age efficiency similar to the E. coli strains, ap-
proximately 50% when comparing pRO-NANP
with pRO-T-NANP (Figs. 6A–C) and almost100% when comparing pRO-CS with pRO-T-CS
(Fig. 6D–F). Nevertheless, whereas the maximum
cleavage activity in E. coli was observed after 6 h
of induction by culturing in anaerobic conditions,in Salmonella it was observed after 8 h of culture.
4. Discussion
Gram negative attenuated bacterial strains are
attractive live vectors of passenger proteins because
they have been demonstrated to elicit humoral andcellular immune response against viral, bacterial,
and parasite antigens. Although the main factors

Fig. 6. Flow cytometry analysis in S. enterica serovar Typhimurium SL3261. Surface expression of MisLb-NANP and MisLb-CSP,and their variant containing the SCOT. (A) Salmonella transformed with pRO-NANP; (B) Salmonella transformed with pRO-T-
NANP; (C) histogram comparing A and B; (D) Salmonella transformed with pRO-CS; (E) Salmonella transformed with pRO-T-CS;
and (F) histogram comparing D and E.
24 P. Ruiz-Olvera et al. / Plasmid 50 (2003) 12–27
determining the immune response to these passen-
ger antigens are still under discussion, the general
notion is that antigen amount and accessibility
during the induction phase of the adaptative im-
mune response are two critical factors in its out-
come. Antigens profusely displayed on the bacterial
surface may induce better antibody response than
those produced in the bacterial cytosol. For thisreason and because of their simplicity, autotrans-
porters are attractive tools for the development of
live bacterial vector vaccines. Several autotrans-
porters have been employed with success to export
heterologous passenger proteins to the bacterial
surface (autodisplay) (Maurer et al., 1997; Ruiz-
Perez et al., 2002; Thanassi and Hultgren, 2000;
Veiga et al., 1999). Nevertheless, translocation ofpassenger antigens may be related to their size and
folding, and may be different for each autotrans-
porter b-domain. Therefore, it is difficult to predictwhether a protein may be secreted by these systems,
and this has to be determined in individual cases.
We reported previously that the MisL b-do-main was able to translocate four repeats of the
tetrapeptide NANP to the bacterial surface, dem-onstrating that it is indeed an autotransporter
(Ruiz-Perez et al., 2002). Herein we report the
ability of the MisL b-domain to display bigger
passenger proteins, and amethod to release them to
the external milieu by protease cleavage. The con-
structs were assessed in an attenuated Salmonella
strain, which offers several theoretical advantages
over other bacterial live vectors, because it is able to
reach swiftly macrophages and dendritic cells, de-livering antigens directly to these main antigen
presenting cells, therefore inducing local and
systemic, both humoral and cellular immune re-
sponses. Moreover, a surprising property of Sal-
monella vaccine vectors is their ability to induce
MHC class I restricted CD8þ cytotoxic responses
against the passenger antigens (Shata et al., 2000).
Two variants of the main surface protein fromP. falciparum sporozoites were fused to the MisL
b-domain. One consists of eight repeats of the tet-rapeptide NANP, while the other consists of 53 re-
peats. Both proteins were successfully displayed on
the bacterial surface of E. coli and Salmonella as
demonstrated by IFA and flow cytometry. The
Western blot from solubilized E. coli strains
showed a �90 kDa band corresponding to thecomplete MisLb-CSP fusion protein (b-domain

P. Ruiz-Olvera et al. / Plasmid 50 (2003) 12–27 25
�32 kDa,Da-domain�26 kDa, andCSP�25 kDa),and �60 kDa corresponding to the MisLb-NANP(b-domain,Da-domain and eight repeats of NANP,�4 kDa). More importantly, the CSP passengerprotein was processed and released by proteolytic
cleavage in E. coli XL-10Gold as demonstrated by
the�25 kDa band obtained from supernatants. The(NANP)8 was not detectable probably due to its
small size. Western blot analysis also suggested thatthe fusion proteins were not retained in the peri-
plasmic space. However we detected (NANP)8peptide in supernatants from Salmonella by dot
blot. These results may indicate that cleavage and
release is feasible in both E. coli and Salmonella
strains.
Release of the CSP and (NANP)8 passenger
proteins was achieved by inserting the OmpTcleavage site between the passenger proteins and
the MisL b-domain. OmpT from E. coli is member
of the family of outer membrane proteases known
as omptins, which are virulence factors of Gram
negative bacteria related to the plasminogen acti-
vator of Yersinia (Sodeinde and Goguen, 1989).
Their proteolytic activity depends on the interac-
tion with lipopolysaccharide (LPS) and they cleavebetween two consecutive basic amino acid residues
(Kramer et al., 2000), mainly K/R, although other
amino acid recognition sequences have been re-
ported (Maurer et al., 1997). Omptins, such as
OmpT and OmpP, may also degrade recombinant
proteins, thus interfering with their production
(Matsuo et al., 1999). In the case of passenger
autodisplayed proteins with putative OmpT rec-ognition sites in their amino acid sequence, the
evaluation of proteolytic processing have led to
contradictory results, whereas some authors have
reported protein cleavage when the heterologous
protein is produced in OmpT positive bacterial
strains (Lattemann et al., 2000), others find no
evidence of proteolytic cleavage (Shi and Wen,
2001). Hanke et al. (1992) reported two hybridproteins consisting of the cholesterol esterase/li-
pase from Pseudomonas fused to the E. coli hem-
olysin (HlyA). These authors also achieved protein
release to the external milieu including an adapter
sequence that contained cleavage sites for OmpT.
Similar results to E. coli strains were observed
in S. enterica serovar Typhimurium SL3261, the
flow cytometry demonstrated surface display ofMisLb-NANP and MisLb-CSP and fluorescence
displacement to the left with MisLb-T-NANP andMisLb-T-CSP, suggesting release of the passengerproteins. Cleavage in S. enterica serovar Ty-
phimurium SL3261 may be explained by the ac-
tivity of other proteases related to OmpT.
Grodberg et al. have demonstrated that the Sal-
monella E protein encoded by pgtE is highly ho-mologous to OmpT (Grodberg and Dunn, 1989).
We estimated the cleavage efficiency for CS and
(NANP)8 passenger proteins in E. coli after 6 h of
culture to be approximately 98 and 71%, respec-
tively, whereas in S. enterica serovar Typhimurium
SL3261 it was 100 and 50%, respectively, after 8 h
of culture in anaerobic conditions.
Taken together the data presented here revealthat observations made with other autotrans-
porters apply to MisL as well, and demonstrate
this protein is able to autodisplay passenger
proteins which can be released to the external
environment if a protease cleavage recognition
site is included after the pore forming translo-
cator domain. To our knowledge this is the first
report of successful utilization of the OmpTcleavage sequence in S. enterica serovar Ty-
phimurium for the release of passenger proteins
to the external milieu.
Acknowledgments
The authors thank Dr. Leopoldo Aguilar Faisalfor his technical assistance. This work was in part
supported by CONACYT Grant M28958 (P.I:
C.G.B) and IMSS Grant FP-2001/056, Reg.2000-
693-0012 (P.I: C.G:B.). P.R.O., A.S:M., and
F.R.P. were recipients of fellowship from CO-
NACYT and IMSS.
References
Al Hasani, K., Henderson, I.R., Sakellaris, H., Rajakumar, K.,
Grant, T., Nataro, J.P., Robins-Browne, R., Adler, B.,
2000. The sig A gene which is borne on the she pathoge-
nicity island of Shigella flexneri 2a encodes an exported
cytopathic protease involved in intestinal fluid accumula-
tion. Infect. Immun. 68, 2457–2463.

26 P. Ruiz-Olvera et al. / Plasmid 50 (2003) 12–27
Benz, I., Schmidt, M.A., 1989. Cloning and expression of an
adhesin (AIDA-I) involved in diffuse adherence of entero-
pathogenic Escherichia coli. Infect. Immun. 57, 1506–1511.
Burns, D.L., 1999. Biochemistry of type IV secretion. Curr.
Opin. Microbiol. 2, 25–29.
Dozois, C.M., Dho-Moulin, M., Bree, A., Fairbrother, J.M.,
Desautels, C., Curtiss III, R., 2000. Relationship between
the Tsh autotransporter and pathogenicity of avian Escher-
ichia coli and localization and analysis of the Tsh genetic
region. Infect. Immun. 68, 4145–4154.
Fink, D.L., Cope, L.D., Hansen, E.J., Geme III, J.W., 2001.
The Haemophilus influenzae Hap autotransporter is a
chymotrypsin clan serine protease and undergoes autopro-
teolysis via an intermolecular mechanism. J. Biol. Chem.
276, 39492–39500.
Grodberg, J., Dunn, J.J., 1989. Comparison of Escherichia coli
K-12 outer membrane protease OmpT and Salmonella
typhimurium E protein. J. Bacteriol. 171, 2903–2905.
Guyer, D.M., Henderson, I.R., Nataro, J.P., Mobley, H.L.,
2000. Identification of sat, an autotransporter toxin pro-
duced by uropathogenic Escherichia coli. Mol. Microbiol.
38, 53–66.
Hanke, C., Hess, J., Schumacher, G., Goebel, W., 1992.
Processing by OmpT of fusion proteins carrying the HlyA
transport signal during secretion by the Escherichia coli
hemolysin transport system. Mol. Gen. Genet. 233, 42–48.
Henderson, I.R., Czeczulin, J., Eslava, C., Noriega, F., Nataro,
J.P., 1999. Characterization of pic, a secreted protease of
Shigella flexneri and enteroaggregative Escherichia coli.
Infect. Immun. 67, 5587–5596.
Henderson, I.R., Nataro, J.P., 2001. Virulence functions of
autotransporter proteins. Infect. Immun. 69, 1231–1243.
Hess, J., Gentschev, I., Goebel, W., Jarchau, T., 1990. Analysis
of the haemolysin secretion system by PhoA–HlyA fusion
proteins. Mol. Gen. Genet. 224, 201–208.
Hess, J., Gentschev, I., Miko, D., Welzel, M., Ladel, C.,
Goebel, W., Kaufmann, S.H., 1996. Superior efficacy of
secreted over somatic antigen display in recombinant
Salmonella vaccine induced protection against listeriosis.
Proc. Natl. Acad. Sci. USA 93, 1458–1463.
Jose, J., Jahnig, F., Meyer, T.F., 1995. Common structural
features of IgA1 protease-like outer membrane protein
autotransporters. Mol. Microbiol. 18, 378–380.
Klauser, T., Pohlner, J., Meyer, T.F., 1990. Extracellu-
lar transport of cholera toxin B subunit using Neisse-
ria IgA protease beta-domain: conformation-dependent
outer membrane translocation. EMBO J. 9, 1991–1999.
Klauser, T., Pohlner, J., Meyer, T.F., 1993. The secretion
pathway of IgA protease-type proteins in Gram-negative
bacteria. Bioessays 15, 799–805.
Kramer, R.A., Zandwijken, D., Egmond, M.R., Dekker, N.,
2000. In vitro folding, purification and characterization of
Escherichia coli outer membrane protease ompT. Eur. J.
Biochem. 267, 885–893.
Laemmli, U.K., 1970. Cleavage of structural proteins during
the assembly of the head of bacteriophage T4. Nature 227,
680–685.
Lattemann, C.T., Maurer, J., Gerland, E., Meyer, T.F., 2000.
Autodisplay: functional display of active beta-lactamase on
the surface of Escherichia coli by the AIDA-I autotrans-
porter. J. Bacteriol. 182, 3726–3733.
Leininger, E., Roberts, M., Kenimer, J.G., Charles, I.G.,
Fairweather, N., Novotny, P., Brennan, M.J., 1991. Pert-
actin, an Arg–Gly–Asp-containing Bordetella pertussis sur-
face protein that promotes adherence of mammalian cells.
Proc. Natl. Acad. Sci. USA 88, 345–349.
Lindenthal, C., Elsinghorst, E.A., 1999. Identification of a
glycoprotein produced by enterotoxigenic Escherichia coli.
Infect. Immun. 67, 4084–4091.
Matsuo, E., Sampei, G., Mizobuchi, K., Ito, K., 1999. The
plasmid F OmpP protease, a homologue of OmpT, as a
potential obstacle to E. coli-based protein production.
FEBS Lett. 461, 6–8.
Maurer, J., Jose, J., Meyer, T.F., 1997. Autodisplay: one-
component system for efficient surface display and release of
soluble recombinant proteins from Escherichia coli. J.
Bacteriol. 179, 794–804.
Poulsen, K., Reinholdt, J., Kilian, M., 1992. A comparative
genetic study of serologically distinct Haemophilus influen-
zae type 1 immunoglobulin A1 proteases. J. Bacteriol. 174,
2913–2921.
Ruiz-Perez, F., Leon-Kempis, R., Santiago-Machuca, A.,
Ortega-Pierres, G., Barry, E., Levine, M., Gonzalez-Bonilla,
C., 2002. Expression of the Plasmodium falciparum immu-
nodominant epitope (NANP)(4) on the surface of Salmo-
nella enterica using the autotransporter MisL. Infect.
Immun. 70, 3611–3620.
Salmond, G.P., Reeves, P.J., 1993. Membrane traffic wardens
and protein secretion in Gram-negative bacteria. Trends
Biochem. Sci. 18, 7–12.
Sambrook, J., Russell, D.W., 2001. Molecular Cloning: A
Laboratory Manual, third ed Cold Spring Harbor
Laboratory Press, Cold Spring Harbor, New York,
USA.
Shannon, J.L., Fernandez,R.C., 1999.TheC-terminal domainof
theBordetella pertussis autotransporter BrkA forms a pore in
lipid bilayer membranes. J. Bacteriol. 181, 5838–5842.
Sharff, A., Fanutti, C., Shi, J., Calladine, C., Luisi, B., 2001.
The role of the TolC family in protein transport and
multidrug efflux. From stereochemical certainty to mecha-
nistic hypothesis. Eur. J. Biochem. 268, 5011–5026.
Shata, T.M., Stevceva, L., Agwale, S., Lewis, K.G., Hone,
D.M., 2000. Recent advances with recombinant bacterial
vaccine vectors. Mol. Med. Today 6, 66–71.
Shi, H., Wen, S.W., 2001. Display of green fluorescent protein
on Escherichia coli cell surface. Enzyme Microb. Technol.
28, 25–34.
Sodeinde, O.A., Goguen, J.D., 1989. Nucleotide sequence of
the plasminogen activator gene of Yersinia pestis: rela-
tionship to omp T of Escherichia coli and gene E of
Salmonella typhimurium. Infect. Immun. 57, 1517–
1523.
St III, G.J., Cutter, D., 2000. The Haemophilus influenzae Hia
adhesin is an autotransporter protein that remains unc-

P. Ruiz-Olvera et al. / Plasmid 50 (2003) 12–27 27
leaved at the C terminus and fully cell associated. J.
Bacteriol. 182, 6005–6013.
Stathopoulos, C., Hendrixson, D.R., Thanassi, D.G., Hultgren,
S.J., St III, G.J., Curtiss III, R., 2000. Secretion of virulence
determinants by the general secretory pathway in Gram-
negative pathogens: an evolving story. Microbes Infect. 2,
1061–1072.
Suzuki, T., Lett, M.C., Sasakawa, C., 1995. Extracellular
transport of VirG protein in Shigella. J. Biol. Chem. 270,
30874–30880.
Thanassi, D.G., Hultgren, S.J., 2000. Multiple pathways allow
protein secretion across the bacterial outer membrane. Curr.
Opin. Cell Biol. 12, 420–430.
Towbin, H., Staehelin, T., Gordon, J., 1979. Electro-
phoretic transfer of proteins from polyacrylamide
gels to nitrocellulose sheets: procedure and some
applications. Proc. Natl. Acad. Sci. USA 76, 4350–
4354.
Veiga, E., de, L.V, Fernandez, L.A., 1999. Probing secretion
and translocation of a beta-autotransporter using a reporter
single-chain Fv as a cognate passenger domain. Mol.
Microbiol. 33, 1232–1243.
Veiga, E., Sugawara, E., Nikaido, H., de, L.V., Fernandez,
L.A., 2002. Export of autotransported proteins proceeds
through an oligomeric ring shaped by C-terminal domains.
EMBO J. 21, 2122–2131.
Watson, J.V., 2001. Proof without prejudice revisited: immu-
nofluorescence histogram analysis using cumulative fre-
quency subtraction plus ratio analysis of means. Cytometry
43, 55–68.
Communicated by S. Levy
Top Related
Copyright © 2022 FDOKUMEN

