







Bahasa
Halaman
Hukum


UNIVERSIDADE FEDERAL DE MINAS GERAIS INSTITUTO DE CIÊNCIAS BIOLÓGICAS
DEPARTAMENTO DE MORFOLOGIA
AVALIAÇÃO DA PERDA DE CÉLULAS ESPERMATOGÊNICAS
E DA CAPACIDADE DE COLONIZAÇÃO DAS ESPERMATOGÔNIAS-TRONCO, EM CAMUNDONGOS
EXPOSTOS A DIFERENTES REGIMES DE TRATAMENTO COM A CICLOFOSFAMIDA
Belo Horizonte
2011

UNIVERSIDADE FEDERAL DE MINAS GERAIS INSTITUTO DE CIÊNCIAS BIOLÓGICAS
DEPARTAMENTO DE MORFOLOGIA
AVALIAÇÃO DA PERDA DE CÉLULAS ESPERMATOGÊNICAS E DA CAPACIDADE DE COLONIZAÇÃO DAS
ESPERMATOGÔNIAS-TRONCO, EM CAMUNDONGOS EXPOSTOS A DIFERENTES REGIMES DE TRATAMENTO
COM A CICLOFOSFAMIDA
Programa de Pós-Graduação em Biologia Celular
Laboratório de Biologia Estrutural e Reprodução Departamento de Morfologia
ICB/UFMG
Doutoranda: Ana Luiza Drumond
Orientador: Dr. Hélio Chiarini-Garcia
Colaborador: Dr. Marvin L. Meistrich
Belo Horizonte 2011

ANA LUIZA DRUMOND
AVALIAÇÃO DA PERDA DE CÉLULAS ESPERMATOGÊNICAS E DA CAPACIDADE DE COLONIZAÇÃO DAS
ESPERMATOGÔNIAS-TRONCO, EM CAMUNDONGOS EXPOSTOS A DIFERENTES REGIMES DE TRATAMENTO
COM A CICLOFOSFAMIDA
Belo Horizonte
Instituto de Ciências Biológicas da UFMG
2011
Tese apresentada ao Programa de Pós-Graduação em Biologia Celular do Instituto de Ciências Biológicas da Universidade Federal de Minas Gerais, como requisito parcial para obtenção do título de Doutor. Área de concentração: Biologia Celular Orientador: Dr. Hélio Chiarini-Garcia

Drumond, Ana Luiza Alvarenga Avaliação da perda de células espermatogênicas e da capacidade de colonização das espermatogônias-tronco, em camundongos expostos a diferentes regimes de tratamento com a ciclofosfamida. [manuscrito] / Ana Luiza Alvarenga Drumond. – 2011. 115 f. : il. ; 29,5 cm. Orientador: Hélio Chiarini-Garcia. Tese (doutorado) – Universidade Federal de Minas Gerais, Instituto de Ciências Biológicas. 1. Testículos – Teses. 2. Biologia celular – Teses. 3. Ciclofosfamida. 4. Células tronco espermatogoniais. I. Chiarini-Garcia, Helio. II. Universidade Federal de Minas Gerais. Instituto de Ciências Biológicas. III. Título. CDU: 576.3: 615.7


Dedico esse trabalho aos meus tão bravos amigos,
que deixam o nosso país para se aventurar em terras estrangeiras,
e que, à procura de um crescimento profissional e/ou pessoal,
trabalham duro, enfrentam as diferenças e as saudades,
na busca insessante de se tornar uma pessoa melhor.

AGRADECIMENTOS:
Agradeço a todos que de alguma forma colaboraram para a realização deste trabalho, em
especial:
- Ao professor Hélio, pela orientação, pela amizade, pelo carinho, pelos puxões de orelha e por
tantos momentos compartilhados nesses longos sete anos;
- Thank you Dr. Marvin Meistrich, for having me at your lab, and for contributing in so many ways
to my professional and personal growth;
- Às professoras Fernanda e Gleydes;
- À Maria Luiza, técnica do Laber;
- Aos colegas do Laber, tantos alunos que por lá passaram nesses quatro anos, e àqueles que por
lá ainda estão: Aline, Amanda, Ana Luisa, Carolina, Daniel, Débora, Felipe, Francisco, Gisele,
Letícia, Letícia, Marilda, Michele, Nadine, Paula, Renata, Tatiane. Vocês também fazem parte
do meu crescimento;
- Thanks to all the crew from Dr. Meistrich’s lab, and to all the technicians and secretaries in the
Department of Experimental Radiation Oncology, who work hard, but always with a big smile,
and are “making cancer history”;
- Aos colegas do Programa de Biologia Celular, muitos dos quais já terminaram sua saga por aqui
e já se foram. Não há lugar para encontrar bons amigos e madrinhas de casamento como
nesse Programa;
- À Sibele, obrigada pela sua dedicação a todos nós, alunos e professores;
- Às funcionárias do Centro de Microscopia Eletrônica (CEMEL) do ICB/UFMG;
- Aos demais professores e funcionários do Programa de Pós-Graduação em Biologia Celular;
- Aos demais funcionários do Departamento de Morfologia;
- Aos meus pais, meus irmãos, e a toda a minha família, vocês suportaram e suportam a saudade,
mas ficam todos sempre na torcida, não importa a distância. Vocês são uma parte
indispensável de mim, não importa aonde eu vá;
- Aos meus queridos ex-houstonianos (outros nem tão ex assim), Tininha, Cris, Flavinha, Tiago,
Rodrigo, Vini, William, Miguel, Raquel e Leandro, vocês foram amigos e família. Em uma “festa
estranha, com gente esquisita”, me fizeram sentir como em casa, em Terra Brasillis. Os
momentos vividos por um ano vão ficar guardados pra sempre;
- Last, but definitely not least, thanks to my husband Mike, for his support, love, and care. For
showing so much interest in Cellular Biology, Biology of Reproduction, and so many things you
had no clue what they were about, most of the time. Thanks for being my partner in this crazy
deal, which is grad school, always by my side, no matter how far.
Or should I thank to my doctorate for bringing me to you?
I guess I should thank you both…

Esta tese foi desenvolvida no Laboratório de Biologia Estrutural e Reprodução
(Laber) do Departamento de Morfologia, do Instituto de Ciências Biológicas, da
Universidade Federal de Minas Gerais, sob a orientação do Prof. Dr. Hélio Chiarini-
Garcia;
Todos os experimentos aqui apresentados foram desenvolvidos no Departamento
de Radiologia Oncológica Experimental, do UT MD Anderson Cancer Center - EUA,
com a colaboração do Prof. Dr. Marvin L. Meistrich, e com o auxílio das seguintes
instituições:
- Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)
- Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)
- Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG)
- National Institute of Health (NIH) – EUA

“A mente que se abre a uma nova idéia,
jamais voltará ao seu tamanho original.”
(Albert Einstein)

SUMÁRIO
Resumo................................................................................................................................................ 1
1. INTRODUÇAO
1.1. A origem das células germinativas........................................................................................ 2
1.2. Processo de auto-renovação e diferenciação a partir das espermatogônias-tronco............ 2
1.3. Fatores que regulam as espermatogônias............................................................................
1.4. Estádios do ciclo do epitélio seminífero................................................................................
1.5. O uso da ciclofosfamida e seus efeitos colaterais na reprodução masculina.......................
1.6. Transplante de células germinativas.....................................................................................
4
6
7
9
2. JUSTIFICATIVA E OBJETIVOS.................................................................................................
13
3. MATERIAIS E MÉTODOS ......................................................................................................... 15
3.1. Animais...............................................................................................................................
Camundongos doadores.................................................................................................
Camundongos receptores...............................................................................................
3.2. Tratamentos com a ciclofosfamida.....................................................................................
Tratamento com dose única............................................................................................
Tratamento com múltiplas doses, aplicadas a cada 5 dias.............................................
Tratamento com múltiplas doses, aplicadas a cada 4 dias.............................................
3.3. Transplante de células germinativas........................................................................... ......
Obtenção da suspensão celular.....................................................................................
Injeção da suspensão celular........................................................................................
3.4. Coleta e fixação dos testículos..........................................................................................
3.5. Processamento histológico dos testículos.........................................................................
3.6. Contagem de espermatozóides testiculares......................................................................
3.7. TUNEL e imunoistoquímica................................................................................................
3.8. Análises morfológicas e morfométricas.............................................................................
Análise qualitativa............................................................................................................
Análise semi-quantitativa.................................................................................................
Número relativo de células germinativas por 100 nucléolos de células de Sertoli...........
Apoptoses espermatogoniais..........................................................................................
Número de colônias produzidas após o transplante........................................................
3.9. Análise estatística...............................................................................................................
15
15
17
18
18
19
20
21
21
22
22
24
25
26
27
27
27
27
28
28
29

4. RESULTADOS ...........................................................................................................................
4.1. Alterações causadas por uma única dose de ciclofosfamida.............................................
Avaliação dos pesos corporais e testiculares..................................................................
Indução de apoptose e depleção das células germinativas,............................................
Avaliações realizadas aos 28 dias após o tratamento......................................................
4.2. Alterações causadas múltiplas doses de ciclofosfamida.....................................................
Alterações no peso corporal durante e após o tratamento...............................................
Quantificação de morte celular.........................................................................................
Análises histológicas e de número celular........................................................................
4.3. Recuperação do epitélio seminífero ao fim do tratamento..................................................
Análises histológicas........................................................................................................
Análise da recuperação da função testicular....................................................................
4.4. Capacidade funcional das espermatogônias-tronco após transplante.................................
Número de células germinativas presente nos animais doadores....................................
Características das suspensões celulares........................................................................
Análises morfométricas dos camundongos receptores, após o transplante......................
30
30
30
31
33
35
35
36
37
40
40
40
41
41
43
43
5. DISCUSSÃO................................................................................................................................
47
6. CONCLUSÃO.............................................................................................................................. 52
7. REFERÊNCIAS BIBLIOGRÁFICAS.............................................................................................
ANEXO I - TRABALHO PUBLICADO..........................................................................................
ANEXO II - TRABALHO ENVIADO PARA PUBLICAÇÃO..............................................................
53
62
75

Resumo
1
Resumo
A espermatogênese é extremamente sensível a tratamentos quimioterápicos. A
ciclofosfamida é um exemplo de droga amplamente utilizada em pacientes, que pode causar
infertilidade nos homens. Baseando-nos no fato de que a ciclofosfamida diminui a contagem de
espermatozóides, e de que a recuperação da produção espermática é dependente da regeneração
das células-tronco, no presente estudo nós desenvolvemos um regime de tratamento com
ciclofosfamida capaz de eliminar células germinativas em diferenciação nos testículos de
camundongos, e avaliamos a abilidade das espermatogônias-tronco de regenerarem a
espermatogênese in situ e após transplante de células germinativas. Nós utilizamos um tratamento
com sete doses de ciclofosfamida, administradas com um intervalo de 4 dias entre doses, que
resultou em uma drástica redução do número de células germinativas em diferenciação, porém
aproximadamente 50% das espermatogônias do tipo A indiferenciadas ainda estavam presentes no
epitélio seminífero, com morfologia normal. Nós monitoramos a cinética da recuperação espermática
e encontramos que o número de espermatozóides testiculares foi restaurado para 64% do resultado
apresentado para animais controle, dentro do tempo necessário para que as células-tronco se tornem
espermatozóides. Quando as espermatogônias que sobreviveram ao tratamento com ciclofosfamida
foram transplantadas para camundongos com espermatogênese endógena deprimida por radiação
fracionada, a recuperação da espermatogênese foi reduzida para 12% do valor esperado, quando
comparados com animais criptorquídicos. Assim, nós concluímos que múltiplas doses de
ciclofosfamida não eliminaram completamente as espermatogônias-tronco, mas resultaram em perda
celular e dano funcional das células-tronco.

Introdução
2
1. Introdução
1.1. A origem das células germinativas
As células germinativas primordiais (CGPs) são células embrionárias extra-gonadais que dão
origem às células germinativas dos machos e fêmeas, respectivamente espermatogônias e oogônias.
As CGPs se originam a partir de uma pequena população de células do epiblasto (Lawson &
Pedersen, 1992) que, em camundongos, migram da base do alantóide, através do intestino primitivo,
até alcançarem as chamadas cristas urogenitais, entre os dias 9-11 pós-coito, onde terá início a
formação dos testículos.
Ao chegarem às cristas genitais, as CGPs são envolvidas pelas células de Sertoli, e pelas
células precursoras das células mióides, formando os cordões seminíferos. Após a formação dos
cordões, as células germinativas passam a ser denominadas gonócitos (Huckins & Clermont, 1968;
Revisão em Donovan & de Miguel, 2005). Estas células sofrem então divisões mitóticas por alguns
dias, até se estacionarem na fase G0/G1 do ciclo celular, estado em que permanecem até o momento
do nascimento, em camundongos (Huckins & Clermont, 1968). Logo após o nascimento, os gonócitos
voltam a proliferar e dão origem às espermatogônias-tronco (Vergouwen et al., 1991), e é a partir de
divisões destas células que se inicia a espermatogênes, processo que vai resultar na formação dos
espermatozóides.
1.2. Processo de auto-renovação e diferenciação a partir das espermatogônias-tronco
Em machos adultos, a célula mais primitiva no processo espermatogênico é a espermatogônia-
tronco. Nos camundongos, esta célula é também conhecida como espermatogônia Aisolada (Ais), uma
vez que, através do estudo de túbulos seminíferos em montagem in-toto, ela pode ser observada sem
quaisquer conexões citoplasmáticas com outras espermatogônias (de Rooij, 1998). As
espermatogônias Ais apresentam características de uma célula-tronco clássica, isto é, ao se dividirem

Introdução
3
podem formar células semelhantes a elas mesmas, através do processo de auto-renovação, ou
originar células mais diferenciadas, neste caso, comprometidas com a espermatogênese.
No processo de diferenciação, a mitose das espermatogônias Ais resulta na formação das
espermatogônias Apareadas (Apr). Um indicador morfológico de que as gerações espermatogoniais
subseqüentes estão comprometidas com a diferenciação é a ligação entre as células filhas da
espermatogônia Ais, através das chamadas pontes inter-citoplasmáticas (Weber & Russell, 1987). Por
sua vez, a divisão das espermatogônias Apr leva à formação de cadeias de 4, 8 e 16 células
denominadas de espermatogônias Aalinhadas (Aal). Estes tipos espermatogoniais, de Ais até Aal, são
também conhecidos como espermatogônias do tipo A indiferenciadas (Aind), ou imaturas, por
possuírem graus variáveis de indiferenciação, e ainda não podem ser distinguidos uns dos outros
através de marcadores morfológicos (Chiarini-Garcia & Russell, 2001; Chiarini-Garcia et al., 2003) ou
moleculares (Aponte et al., 2005).
Após a formação das espermatogônias Aal, as sucessivas divisões mitóticas geram os tipos
celulares denominados espermatogônias do tipo A em diferenciação (A1, A2, A3, e A4),
espermatogônias intermediárias (In) e espermatogônias do tipo B (B). Atualmente estas células mais
diferenciadas (de A1 até B) podem ser identificadas morfologicamente através das microscopias de
luz de alta resolução (Chiarini-Garcia & Russell, 2001) e eletrônica de transmissão (Chiarini-Garcia &
Russell, 2002). As espermatogônias do tipo B, as últimas células da linhagem espermatogonial, se
dividem gerando espermatócitos primários, que iniciam a fase meiótica da espermatogênese.
O esquema a seguir (Figura 1) ilustra a possível constituição de diferentes clones de
espermatogônias no camundongo desde a espermatogônia Ais até a última geração de
espermatogônia, do tipo B (de Rooij & Russell, 2000; de Rooij, 2001).
Ais ► Apr ► Aal (Aal-4, ► Aal-8 ► Aal-16) A1 ► A2 ► A3 ► A4 ► In ► B
Figura 1 – Divisões mitóticas (cabeças de seta) de cada um dos diferentes tipos espermatogoniais em
camundongos. São aproximadamente 9 divisões que vão formar, a partir de uma A is, em torno de 1024 espermatogônias do tipo B. As espermatogônias Ais são as células-tronco do processo espermatogênico, tendo a capacidade de se auto-renovarem (seta curva). Entre as espermatogônias Aal e A1 (seta clara) não ocorre divisão mas sim diferenciação celular.

Introdução
4
1.3. Fatores que regulam as espermatogônias
Quando se fala em fatores que regulam as espermatogônias, é importante relembrar que o
grupo de espermatogônias A indiferenciadas é constituído pelas espermatogônias A is, Apr e Aal, e que
dentre elas apenas a Ais é considerada como espermatogônia-tronco e, ainda, que os marcadores
descobertos até o momento não marcam especificamente a Ais, mas sim a todas as espermatogônias
indiferenciadas, indistintamente.
Embora não tenham sido identificados marcadores específicos para as espermatogônias-
tronco, vários fatores têm sido citados por exercerem um importante controle da regulação entre a
auto-renovação e a diferenciação das células-tronco em mamíferos. Exemplos de fatores que,
conhecidamente, regulam as espermatogônias indiferenciadas podem ser o GDNF (glial cell-line
derived neurotrophic factor; Creemers et al., 2002; Naughton et al., 2006; Yomogida et al., 2003), o
GFRα-1 (GDNF family receptor alpha 1; Buageaw et al., 2005;), o Plzf (promyelocytic leukemia zinc-
finger; Buaas et al., 2004; Costoya et al., 2004;), o Ngn3 (Neurogenina 3; Yoshida et al., 2004) e o
Sox3 (SRY related high mobility group-box; Raverot et al., 2005).
Dentre os vários fatores descritos por serem expressos nas células espermatogoniais está o
receptor c-kit, que se liga ao fator de célula-tronco (SCF, stem cell factor), produzido pelas células de
Sertoli. A importância desse marcador foi identificada em duas mutações, nos locus W/W White
spotting e Sl steel, que em camundongos resultam em esterilidade (Yoshinaga et al., 1991; Ohta et
al., 2000). Durante a fase inicial do desenvolvimento fetal, o receptor para SCF é expresso pelas
CGPs, sendo importante no sucesso da migração destas células para as gônadas. No entanto, após
a formação do testículo, o receptor c-kit passa a ser expresso nas espermatogônias do tipo A em fase
de diferenciação. Desta forma, as células de Sertoli regulam a proliferação das espermatogônias em
diferenciação através do receptor c-kit (Yoshinaga et al., 1991; Ohta et al., 2000).
Além do sistema c-kit/SCF, o GDNF é também importante para a proliferação e diferenciação
das células germinativas (Meng et al., 2000). Este fator é expresso pelas células de Sertoli, e se liga
nas espermatogônias-tronco através dos receptores c-ret e GFR-α1. Camundongos com a expressão
de GDNF elevada apresentam um aumento do número de espermatogônias indiferenciadas,
eventualmente, desenvolvendo tumores testiculares semelhantes ao seminoma observado em
homens (Meng et al., 2000). Estudos recentes, utilizando diversas abordagens, indicam que o GDNF

Introdução
5
estimula a proliferação das espermatogônias-tronco sem causar a diferenciação das mesmas (Kubota
et al., 2004; Buageaw et al., 2005; Hofmann et al., 2005; Kanatsu-Shinohara et al., 2005).
Outro marcador importante para as espermatogônias é o receptor ALK3 (alctivin receptor like
kinase 3), presente em espermatogônias do tipo A, que se liga à proteína morfogênica do osso do
tipo 4 (BMP4, bone morphogenetic protein) produzida pela célula de Sertoli. Experimentos in vitro
revelaram que a BMP4 proporciona aumento na proliferação e diferenciação das espermatogônias.
Tummers & Thesleff (2003) mostraram que a BMP4 também está envolvida no recrutamento das
células-tronco para regiões específicas do epitélio seminífero. A interação entre BMP4 e seu receptor
ALK3 regula uma via de transdução de sinal, que induz a expressão de c-kit, seguida pela
diferenciação da espermatogônia A indiferenciada (Pellegrini et al, 2003).
Ja o fator Plzf (promyelocytic leukemia zinc-finger) é um repressor de transcrição que regula o
estado epigenético das células indiferenciadas. Esse fator foi identificado como essencial para a
manutenção de auto-renovação das células-tronco (Buaas et al., 2004). Isso foi confirmado no epitélio
seminífero por Costoya e colaboradores (2004), que demonstraram que o Plzf é requerido para
regular a auto-renovação e manutenção do pool de espermatogônias-tronco. O Plzf parece ser
expresso em tipos celulares que não expressam ciclina D1 e/ou D2, conhecidas por serem expressas
em espermatogônias em diferenciação, sugerindo então serem as espermatogônias reguladas pelo
Plzf as mais indiferenciadas.
Finalmente, a Ngn 3 é um fator de transcrição expresso específicamente em espermatogônias
do tipo A indiferenciadas (Yoshida et al., 2004), sendo expresso em células que estão presentes em
todos os estádios do epitélio seminífero, observadas com pouca frequência, e presentes em clones
pequenos de células 2N. Além das características típicas de células indiferenciadas, as células
positivas para Ngn3 apresentaram-se negativas para o receptor c-kit. Após a transição para células
em diferenciação, a expressão de neurogenina 3 é suprimida. Recentemente, a Ngn3 foi utilizada, em
camundongos, na caracterização de espermatogônias-tronco em funcionamento normal, e após
drástica depleção de células germinativas (Yoshida, 2007). Os resultados sugerem que
espermatogônias que expressam uma quantidade de Ngn 3 maior são na verdade espermatogônias-
tronco potenciais, que sofreriam auto-renovação apenas durante a regeneração do epitélio
seminífero, quando este é danificado.

Introdução
6
1.4. Estádios do ciclo do epitélio seminífero
A classificação das secções transversais de túbulos seminíferos em estádios do ciclo do
epitélio seminífero (CES) é uma ferramente de grande valia para o estudo da cinética das células
germinativas, inclusive das espermatogônias (Russell et al, 1990; de Rooij, 1998). Ela permite
investigar a dinâmica, a progressão e a expansão das células em um determinado espaço de tempo.
A forma de classificação mais utilizada em camundongos é conhecida como método da vesícula
acrossômica, e permite o reconhecimento de doze estádios (Figura 2) a partir da observação das
diferentes evoluções morfológicas nucleares e acrossômicas das espermátides.
Como exemplo de eventos ocorrendo em estádios específicos do CES, temos a formação de
espermatogônias do tipo A1, que ocorre por diferenciação celular a partir das espermatogônias Aal nos
estádios VII e VIII (Figura 2). Uma vez que as espermatogônias do tipo A1 tenham sido formadas, as
divisões subseqüentes também são sincronizadas, novamente ocorrendo em estádios específicos do
ciclo espermatogênico. As espermatogônias A1 dividem-se para formar A2 no estádio IX, estas por
sua vez formam as espermatogônias A3 no estádio XI, sendo finalmente as espermatogônias A4
formadas no estádio I do próximo ciclo. Estas últimas se dividem em espermatogônias In nos estádios
II-III, dando origem às espermatogônias do tipo B no estádio IV (Figura 2).
No passado, os diferentes tipos de espermatogônias do tipo A não podiam ser reconhecidos
morfologicamente, tampouco existiam marcadores moleculares específicos para estas células. No
entanto, sabendo que os diferentes tipos celulares estavam presentes em estádios específicos, as
espermatogônias podiam ser identificadas de forma indireta. Por exemplo, numa secção transversal
de túbulo seminífero de camundongos no estádio XII podem ser observadas as espermatogônias do
tipo Ais, Apr Aal (presentes em todos os estádios, embora em número pequeno) e as espermatogônias
do tipo A3 (Figura 2). Hoje, com o uso de microscopia de luz de alta resolução (MLAR), é possível
identificar as células independentemente dos estádios. Estudos das gerações espermatogoniais já
podem ser realizados em animais que não possuem espermatogênese completa (Drumond et al,
2011), ou em animais nos quais os estádios do CES não podem ser identificados (Bolden-Tiller et al.,
2007; Zallio 2009).

Introdução
7
Figura 2 – Associações celulares durante os XII estádios do CES de camundongos. As espermatogônias A
indiferenciadas, que estão presentes em todos os estádios do ciclo, e se dividem entre os estádios X e III. As espermatogônias do tipo A em diferenciação (A1, A2, A3 e A4) estão presentes e se dividem (m, mitose) em estádios específicos do ciclo. Os espermatócitos formados vão passar pelas fases da prófase meiótica (preL, preleptoteno; L, leptóteno; Z, zigóteno; P, paquíteno; D, diplóteno), e as espermátides pelas 16 etapas de diferenciação (uma por estádio). Os tipos celulares presentes na mesma coluna são sempre encontrados juntos e constituem um estádio do ciclo. M!, divisões meióticas I e II.
1.5. O uso da ciclofosfamida e os seus efeitos colaterais na reprodução masculina
A ciclofosfamida é um agente alquilante, pertencente ao grupo de drogas conhecido como
mostardas nitrogenadas, assim denominadas por terem sido criadas a partir do gás mostarda, muito
utilizado na Segunda Guerra Mundial (Colvin, 2003). Essa droga reage fortemente com as bases
guanina presentes no DNA, adicionando um grupo alquil (CnH2n+1) às bases nitrogenadas. A reação
causa uma ligação cruzada entre as duas fitas do DNA, não permitindo a abertura da dupla hélice
para que haja replicação da molécula de DNA (Figura 3). O mecanismo de reparo da célula, ao
detectar dano no DNA, induz a morte das células afetadas (Colvin, 2003).
As células que apresentam maior sensibilidade ao dano causado pelos agentes alquilantes são
aquelas que estão em rápida atividade de proliferação, como as células cancerígenas (Colvin, 2003).
Por esse motivo, a ciclofosfamida vem sendo amplamente empregada para o tratamento de
neoplasias como linfomas, mielomas, leucemias, neuroblastomas, carcinoma de mama, dentre outros
(Ref.:Bula que acompanha o medicamento). Esta droga também pode ser utilizada no tratamento de
doenças auto-imunes, como o lupus eritematoso sistêmico, a anemia hemolítica e a artrite reumática.

Introdução
8
Figura 3 – Ação da ciclofosfamida na molécula de DNA. Promovendo a ligação cruzada entre as duas fitas,
através da ligação nas bases guanina, a droga impede que haja abertura da molécula para duplicação do DNA.
Uma séria reação ao tratamento com agentes alquilantes é o dano às células germinativas. A
bula que acompanha a droga ciclofosfamida afirma no tópico “advertências” que “A ciclofosfamida
pode causar infertilidade em ambos os sexos, dependendo da dose e do tempo de duração do
tratamento...”, e afirma ainda que “A infertilidade pode ser reversível”. Estudos demonstraram que os
efeitos da ciclofosfamida são dependentes da dose aplicada, sendo que doses maiores produzem
efeitos mais fortes (Ref.:Bula que acompanha o medicamento), e que o uso prolongado pode resultar
em infertilidade permanente.
Experimentos foram também realizados em ratos e camundongos, na tentativa de explicitar os
danos causados nas células germinativas pela ciclofosfamida. Lu e Meistrich (1979), por exemplo,
estudaram os efeitos causados por diferentes drogas quimioterápicas, dentre elas a ciclofosfamida,
na reprodução de camundongos C3H, após a aplicação de uma única dose. Esses pesquisadores
encontraram que com uma dose de 200mg de ciclofosfamida por Kg de peso corporal dos animais
(mg/Kg) as espermatogônias em diferenciação do tipo A2, A3, A4, In e B, e parte dos espermatócitos
do tipo pré-leptótenos, foram totalmente eliminados, enquanto as espermatogônias do tipo A1 e Aal
foram parcialmente danificadas, e as espermatogônias Ais e Apr não foram alteradas. Apesar de não
terem sido capazes de reconhecer específicamente os tipos espermatogoniais, os autores chegaram
a essa conclusão ao analisar os diferentes tipos de espermatócitos presentes 11 dias após a injeção,
e prevendo assim quais eram as células que estavam sendo eliminadas no momento da injeção. O
trabalho também mostrou que com uma única dose de 100mg/Kg de ciclofosfamida a contagem de
espermatozóides foi fortemente reduzida 29 dias após o tratamento, mas voltou à contagem normal,
prevista para os animais controle, 56 dias após o tratamento (Lu & Meistrich, 1979).

Introdução
9
Cai e colaboradores (1997) estudaram, em ratos machos, a indução de apoptose causada pela
ciclofosfamida em células germinativas. Eles encontraram que o pico de apoptoses aconteceu entre
12 e 24 horas após a aplicação de uma dose da droga. Também observaram que as
espermatogônias e os espermatócitos sofreram um número muito maior de apoptose induzida por
ciclofosfamida, do que as espermátides.
É importante destacar que, uma eventual recuperação da produção espermática em pacientes
que apresentam azoospermia ou oligospermia ao final do tratamento está relacionada com a
sobrevivência das espermatogônias-tronco, e com a sua habilidade de se diferenciar em células
germinativas mais avançadas, formando espermatozóides saudáveis. Por isso, se torna importante o
estudo dos efeitos da ciclofosfamida não só na reprodução como um todo, mas principalmente os
efeitos causados na linhagem espermatogonial.
1.6. Transplante de células germinativas
O transplante de espermatogônias é uma abordagem experimental que consiste na remoção
de células-tronco do testículo de um animal doador e a transferência das mesmas para o testículo de
um receptor, onde estas células irão se desenvolver e formar espermatozóides maduros com
características genéticas do doador (Figura 4). Essa nova e promissora técnica foi inicialmente
utilizada em camundongos por Brinster e colaboradores (Brinster & Avarbock, 1994; Brinster &
Zimmermann, 1994) e tem proporcionado enormes possibilidades para o estudo tanto da biologia das
células-tronco quanto do processo espermatogênico em si (Parreira et al., 1998; Hausler & Russell,
1999; Izadyar et al., 2000; Brinster, 2002; Dobrinski, 2005a,b; McLean, 2005). Antes de serem
transplantadas, as células germinativas são isoladas, utilizando-se para isso digestão enzimática
dupla (tripsina e colagenase) (Meistrich et al, 1973; Meistrich & Trostle, 1975). Apesar dos avanços na
área, as técnicas de isolamento realizadas a partir de animais adultos, não têm conseguido, até o
momento, obter suspensões celulares com uma concentração alta das espermatogônias-tronco.

Introdução
10
Figura 4 – Transplante de espermatogônias de um camundongo doador, exemplo GFP, para um camundongo receptor, exemplo C57Bl6. A) Coleta dos testículos do animal doador; B) Obtenção de suspensão celular, rica em espermatogônias, a partir dos testículos coletados; C) Injeção da suspensão celular nos testículos do animal receptor; D) Um determinado período após o transplante o animal receptor
passa a produzir espermatozóides com a carga genética do animal doador (verdes), além dos seus próprios espermatozóides (pretos). GFP (Green Fluorescent Protein), proteína fluorescente verde.
O sucesso do transplante depende dos tipos celulares contidos na suspensão obtida (Oatley &
Brinster, 2006). As suspensões celulares consideradas como ideais são aquelas que possuem
espermatogônias-tronco saudáveis, presentes em alta concentração, e um número baixo de
espermatócitos e espermátides. Nesse sentido, podemos citar diferentes modelos de animais
doadores que vêm sendo experimentados nos últimos anos. O emprego de animais jovens, por
exemplo, se revelou como um modelo eficiente, facil de ser obtido e de baixo custo financeiro
(Shinohara et al., 2001; McLean et al., 2003). Nestes animais a espermatogênese ainda não está
completa, o que facilita a obtenção de pools de células-tronco. Apesar de serem considerados bons
doadores, é importante estar atento à idade ideal do animal para obtenção das células a serem
transplantadas, uma vez que espermatogônias obtidas a partir de animais muito jovens podem não
estar presentes em quantidade ideal, além de não serem maduras o suficiente para produzir uma alta
eficiência de transplante (McLean et al., 2003).
Conforme já foi dito, o uso de animais adultos como doadores também tem sido empregado, no
entanto, por apresentarem o epitélio seminífero completo, com todas as células germinativas
presentes, os transplantes realizados a partir destes animais não resultam em altas eficiências de
colonização (Ogawa et al., 2003), sendo necessária a utilização da separação celular através de
Percoll (Koh et al., 2004). Uma alternativa ao uso do Percoll seriam tratamentos experimentais que
eliminem as células germinativas mais diferenciadas, mantendo as espermatogônias-tronco
saudáveis, aumentando assim a proporção de células-tronco que será injetada nos animais
receptores. O criptorquidismo experimental, é um exemplo de técnica que pode provocar a seleção de

Introdução
11
espermatogônias, pois o aumento da temperatura testicular leva à depleção das células germinativas
mais avançadas, sem causar dano às espermatogônias indiferenciadas (Nishimune et al., 1978; de
Rooij et al., 1999). Alguns experimentos demonstram o sucesso do emprego desses animais como
doadores de células-tronco para transplante (de Rooij et al., 1999; Shinohara et al., 2000; Orwig et al.,
2008). Além dos animais criptorquídicos, alguns animas mutantes também têm sido utilizados em
experimentos de transplante (de Rooij et al., 1999). Os camundongos mutantes para o gene steel
(Sl/Sl), por exemplo, possuem células germinativas saudáveis, que expressam o receptor para SCF,
c-kit, no entanto apresentam células de Sertoli defeituosas na produção do SCF, fazendo com que
esses animais sejam desprovidos de células germinativas mais avançadas (Brannan et al., 1992).
Como a alteração desses animais é no componente somático dos testículos, e não nas células
germinativas, estas células podem ser transplantadas com sucesso.
Outro fator importante para o sucesso do transplante é a escolha do receptor adequado. O
microambiente testicular dos animais receptores tem um papel crucial na dinâmica e na eficiência da
colonização, e no desenvolvimento das células germinativas transplantadas. Embora as células
germinativas dos animais doadores sejam capazes de colonizar o testículo de um animal receptor
normal, a eficiência do transplante depende também do sucesso na competição por nichos
disponíveis nos túbulos seminíferos, entre as células germinativas transplantadas e as células
germinativas endógenas (Shinohara et al., 2002). O receptor ideal para o transplante de
espermatogônias é aquele cujos túbulos seminíferos contêm apenas células de Sertoli, normais, pois
este aspecto garante a acessibilidade dos nichos às células germinativas dos doadores (Brinster et
al., 2003).
No que diz respeito aos animais receptores, podemos citar algumas técnicas aplicadas com o
fim de melhorar as condições do microambiente testicular receptor. A primeira delas, que vem sendo
utilizada desde os primeiros experimentos de transplante de células germinativas até hoje, é o uso do
tratamento com o quimioterápico bussulfan como forma de deprimir a produção endógena de células
germinativas, permitindo que apenas as células de Sertoli sobrevivam nos testículos (Brinster &
Avarbock, 1994; Brinster & Zimmermann, 1994; Nagano et al., 1999; Zhang et al., 2006). Apesar de
ser uma técnica bem sucedida no que diz respeito à eliminação das células germinativas, sem afetar
o componente somático, o bussulfan debilita muito os animais receptores. Vários trabalhos relatam a
necessidade de transplante de células da medula óssea para auxiliar na recuperação destes animais

Introdução
12
(Ogawa et al., 1999; Udagawa et al., 2001). Uma alternativa ao tratamento dos animais receptores
com bussulfan seria a utilização de animais mutantes. Os camundongos mutantes para o locus W/W,
por exemplo, apresentam alterações genéticas em suas células germinativas, não sendo capazes de
expressar o receptor c-kit para o fator SCF, produzido pelas células de Sertoli (Brinster & Avarbock,
1994; Brinster & Zimmermann, 1994; Parreira et al. 1998; Nagano et al., 1999). Estes animais, apesar
de desprovidos de células germinativas, possuem células de Sertoli em perfeito estado funcional,
sendo, por isso, receptores ideais para as células-tronco dos animais doadores. Apesar de ser uma
técnica biologicamente viável, a obtenção e criação dos animais mutantes requer um alto
investimento financeiro.
Recentemente, o emprego da radiação para depleção das células germinativas nos testículos
dos animais receptores se revelou uma técnica mais adequada, por ser menos prejudicial à saúde
dos animais do que o tratamento com o bussulfan, e mais acessível, em termos financeiros, do que a
obtenção dos animais mutantes. Por muitos anos o efeito da radiação na reprodução masculina vem
sendo estudado, mostrando como ela afeta as células germinativas em proliferação, sendo elas
espermatogônias-tronco ou não (Cattanach, 1974; Withers et al., 1974; Hasegawa, et al., 1997).
Esses estudos também mostram que doses de radiação fracionada são mais eficientes na depleção
das células germinativas do que doses únicas (Cattanach, 1974; Withers et al., 1974; de Ruiter-
Bootsma et al., 1977; Meistrich et al., 1984). Dessa forma, pesquisadores têm optado pelo uso da
radiação (Zhang et al., 2006; Barroca et al., 2009), representando uma excelente alternativa para a
produção de animais receptores.
A técnica de transplante de células germinativas tem sido utilizada para verificar a
funcionalidade espermatogênica das espermatogônias-tronco, e a possível morte celular causada
após tratamentos citotóxicos, eliminando-se a possibilidade de danos nas células Sertoli (Zhang et al.,
2007), uma vez que o receptor fornece um nicho saudável para as spermatogônias dos doadores se
desenvolverem. Este método foi utilizado para se determinar a queda no número de spermatogônias-
tronco após uma única injeção da droga busulfan, em camundongos, e subsequente recuperação
dessas células após transplante (Kanatsu-Shinohara et al., 2006). O transplante espermatogonial
também foi utilizado em macacos Rhesus (Hermman et al., 2007), tratados com busulfan, para se
acessar a funcionalidado das espermatogônias do tipo A indiferenciadas e como essas células se
comportavam após o tratamento com o quimioterápico.

Justificativa e Objetivos 13
2. Justificativa e Objetivos
O câncer vem desafiando os médicos e pesquisadores da área de saúde a muitos anos, devido
à sua ocorrência nos mais variados tipos de tecidos do corpo humano, e à agressividade com que
muitas vezes acomete os pacientes. Dentre os tratamentos empregados contra esta doença,
medicamentos como os agentes alquilantes são os mais utilizados, podendo ser combinados ou não
com outras formas de tratamento. Como visto na introdução, algumas drogas alquilantes agem nas
células que estão em atividade mitótica constante, não permitindo a replicação das moléculas de
DNA, consequentemente induzindo à morte celular.
Apesar de eficientes contra as neoplasias, os agentes alquilantes acarretam drásticas
alterações nas células germinativas dos pacientes. Logo após o início do tratamento é possível ainda
se observar espermatozóides sendo produzidos por até 2 meses, devido a certa resistência das
células germinativas em estágios mais avançados, como as espermátides (revisão em Meistrich,
2009). No entanto, mesmo em doses baixas, as drogas levam à redução da produção espermática
por até 3 meses após o fim do tratamento. Em um regime agressivo de quimioterapia pode ocorrer
forte redução do número de espermatozóides, ou até azoospermia do paciente. O tempo de duração
da azoospermia será então relacionado com a dose recebida pelo paciente, podendo durar de 1 a 3
anos, ou por um período mais prolongado, muitas vezes sendo permanente. Estudos em
camundongos revelaram que o intervalo necessário para recuperação da fertilidade está diretamente
relacionado ao grau de morte celular induzida nas espermatogônias-tronco (Meistrich, 1982). Assim
sendo, acredita-se que a eventual recuperação da produção espermática depende da sobrevivência
das espermatogônias-tronco, e da sua habilidade de se diferenciar em células mais avançadas,
produzindo espermatozóides saudáveis.
A ciclofosfamida é um exemplo de agente alquilante, que tem sido empregado no tratamento
de várias neoplasias, e também de doenças auto-imunes. Experimentos realizados em roedores

Justificativa e Objetivos 14
revelaram que a ciclofosfamida leva a morte principalmente de espermatogônias em diferenciação, e
espermatócitos do tipo pré-leptótenos (Lu & Meistrich, 1979), principalmente durante as primeiras 24
horas após a injeção da droga (Cai et al., 1997). Os estudos em camundongos revelam ainda que,
assim como em pacientes, a ciclofosfamida reduz drasticamente a quantidade de espermatozóides
produzida pelo epitélio seminífero, após o tratamento, porém essa situação pôde ser revertida em
animais tratados com doses menores (Lu & Meistrich, 1979).
Nesse sentindo, o estudo da ação dessa droga nas células germinativas, principalmente nas
espermatogônias, se torna extremamente importante, visando verificar tanto o impacto deletério nas
células germinativas, quanto a possibilidade de recuperação do processo espermatogênico,
dependendo do tipo de tratamento utilizado.
Objetivo Geral
Estudar as alterações causadas pelo tratamento com a ciclofosfamida nas células
espermatogênicas de camundongos, e avaliar o grau de recuperação das células germinativas após o
fim do tratamento.
Objetivos Específicos
Avaliar os efeitos de diferentes regimes de tratamento com uma única dose de ciclofosfamida;
Estudar o dano causado nas células germinativas após tratamentos envolvendo múltiplas
doses de ciclofosfamida;
Avaliar a recuperação de parâmetros como peso testicular e produção espermática, após o
fim do tratamento com múltiplas doses de ciclofosfamida;
Verificar a função espermatogênica das espermatogônias-tronco que sobreviveram ao
tratamento, através do transplante de células germinativas.

Materiais e Métodos
15
3. Materiais e Métodos
3.1. Animais
Para os tratamentos com ciclofosfamida, foram utilizados camundongos machos, adultos, da
linhagem C57BL6, provenientes da colônia do Departamento de Radiologia Oncológica Experimental,
do MD Anderson Cancer Center (ERO colony, Houston/TX, EUA). Os animais foram obtidos com
aproximadamente 6 semanas de idade, ambientados por uma semana, e mantidos no biotério do
departamento durante todo o tratamento, até o momento do sacrifício. Cada animal recebeu um
número de identificação individual, impresso em brinco de alumínio. O experimento foi aprovado pelo
comitê de ética do MD Anderson Cancer Center (IACUC, Institution Animal Care and Use Comitee).
Para os experimentos de transplante foram preparados grupos de animais doadores de células
germinativas, e grupos de animais receptores das suspensões celulares preparadas a partir dos
testículos dos camundongos doadores.
Camundongos doadores:
Como doadores de células germinativas, foram utilizados camundongos transgênicos para a
produção da proteína fluorescente verde (green fluorescent protein – GFP), para que as células dos
doadores pudessem ser reconhecidas no animal receptor, após o transplante.
Foram utilizados quatro grupos experimentais compostos de camundongos GFP: a) jovens,
b) adultos não tratados, c) adultos experimentalmente criptorquídicos e d) adultos tratados com
ciclofosfamida.
a) Animais jovens:
Foram realizados dois experimentos de transplante, nos quais foram utilizados 5
camundongos doadores, com idades entre 14 e 16 dias, para cada experimento, totalizando 10
animais. No momento da coleta os animais foram pesados, anestesiados (mistura de

Materiais e Métodos
16
ketamina e xilazina a 0,10 mL/10 g de peso corporal) e sacrificados. A suspensão celular
foi então obtida a partir dos testículos coletados.
b) Animais adultos não tratados:
Foram utilizados 5 animais, com aproximadamente 8 semanas de idade. No momento da
coleta os animais foram pesados e anestesiados, e após a coleta procedeu-se a eutanásia por
deslocamento cervical. A suspensão celular foi então obtida a partir dos testículos coletados.
c) Animais experimentalmente criptorquídicos:
Foram utilizados 10 animais, que tinham aproximadamente 8 semanas de idade no dia
da cirurgia. Para o procedimento de cirurgia de criptorquidismo, os animais foram pesados e
anestesiados com ketamina e xilazina (0,1mL/10g de peso corporal). Foi realizada tricotomia na
região abdominal, e em seguida a região foi higienizada com Iodo Povidine. Com bisturi
cirúrgico, foi feita uma incisão na região pelvica do animal, de aproximadamente 1,5 cm. Ambos
os testículos foram, separadamente, retirados do escroto e amarrados à parede abdominal com
linha de seda, sendo que o ponto de fixação foi localizado entre o testículo e o epidídimo.
Os animais foram mantidos criptorquídicos por 8 semanas, até o dia do transplante,
quando foi então obtida suspensão celular a partir dos testículos desses animais. No momento
da coleta os animais foram pesados, receberam a mesma dose de anéstesico do procedimento
cirurgico, e procedeu-se a eutanásia. Baseado em experimentos prévios (dados não
publicados), apenas testículos com valores de peso abaixo de 30 mg foram utilizados como
doadores para o transplante.
d) Animais adultos tratados com ciclofosfamida:
Foram realizados dois experimentos de transplante. Inicialmente previu-se o uso de 5
animais doadores para cada transplante, porém, devido à morte de um dos animais, utilizamos
5 doadores no primeiro experimento, e 4 doadores no segundo, totalizando 9 animais. Os
doadores tinham aproximadamente 8 semanas de idade ao início tratamento com
ciclofosfamida. Os animais foram submetidos a um tratamento com sete doses de
ciclofosfamida (150mg/Kg por dose), administradas por via oral a cada quatro dias. Os animais
foram pesados todos os dias antes da administração da droga. Quatro dias após a última dose
(28º dia de tratamento) os animais foram sacrificados e a suspensão celular foi obtida a partir

Materiais e Métodos
17
do testículo desses animais e injetada nos animais receptores, no mesmo dia. No momento da
coleta os animais foram pesados, anestesiados e sacrificados.
Além dos grupos de animais preparados para serem utilizados como doadores de células
germinativas para os transplantes, outros grupos de animais foram preparados, seguindo-se
exatamente os mesmo critérios estabelecidos nos grupos (a), (b), (c) e (d) anteriormente citados, para
serem processados para análise sob microscopia de luz de alta resolução - MLAR (n=4).
Camundongos receptores:
Como receptores para o transplante de espermatogônias foram utilizados ao todo 38
camundongos machos, adultos. Estes animais tiveram sua espermatogênese endógena deprimida
por tratamento com doses fracionadas de radiação γ, utilizando como fonte de radiação o Cs137
. Os
animais receberam a primeira dose de 1,5 Gy e uma segunda dose de 12 Gy, aplicada 24 horas após
a primeira. Este protocolo foi baseado nos experimentos de Zhang e colaboradores (2006).
Os animais receberam as doses de radiação nas regiões pélvica e do escroto, a fim de garantir
a exposição dos testículos. O restante do corpo do animal foi protegido com placa de cobre. Não foi
necessário anestesiar os animais no momento da irradiação, uma vez que foi possível a imobilização
dos mesmos dentro de uma caixa de acrílico.
Os animais irradiados foram separados nos seguintes grupos:
- Animais controle de 3 a 4 semanas após a radiação (n=3);
- Animais controle de 11 semanas após a radiação (n=3);
- Animais que receberam células de animais jovens (n=7);
- Animais que receberam células de animais adultos não tratados (n=6);
- Animais que receberam células de animais cirurgicamente criptorquídicos (n=7);
- Animais que receberam células de animais tratados com ciclofosfamida (n=12).
O transplante foi realizado 3 a 4 semanas após o tratamento com radiação, e a injeção da
suspenção celular foi feita manualmente, via dúctulo eferente. Os animais transplantados foram
sacrificados 8 semanas após o transplante (aproximadamente 11 semanas após a radiação). No
momento do sacrifício os animais foram pesados e receberam a mistura de ketamina e xilazina (0,10
Materiais e Métodos
18
mL/10 g de peso corporal). Os testículos foram coletados e pesados, e a eutanásia foi realizada após
a coleta.
3.2. Tratamentos com a ciclofosfamida
Tratamento com dose única:
Foram utilizados ao todo 33 camundongos. Os animais receberam injeções de ciclofosfamida
(Cytoxan®, Bristol-Myers Squibb Co), por via oral (p.o. - agulha de gavage 22G1”) e intraperitoneal
(i.p. - agulha de 26G½”). Foram injetadas doses de 150 e 200 mg/Kg de peso corporal do animal,
assim determinadas por base em resultados previamente obtidos no nosso laboratório. O peso
corporal dos animais foi obtido no dia da injeção da droga (dia 0) e duas vezes por semana (dias 3, 7,
10, 14, 17, 21, 24, 28 após o tratamento), até o dia do sacrifício.
Os animais foram divididos em onze (A-K) grupos (n=3), de acordo com a dose recebida, a via
de administração utilizada e o dia em que os animais foram sacrificados (Tabela 1). Animais dos
grupos controle (A-C) receberam injeções de salina a 0,9%.
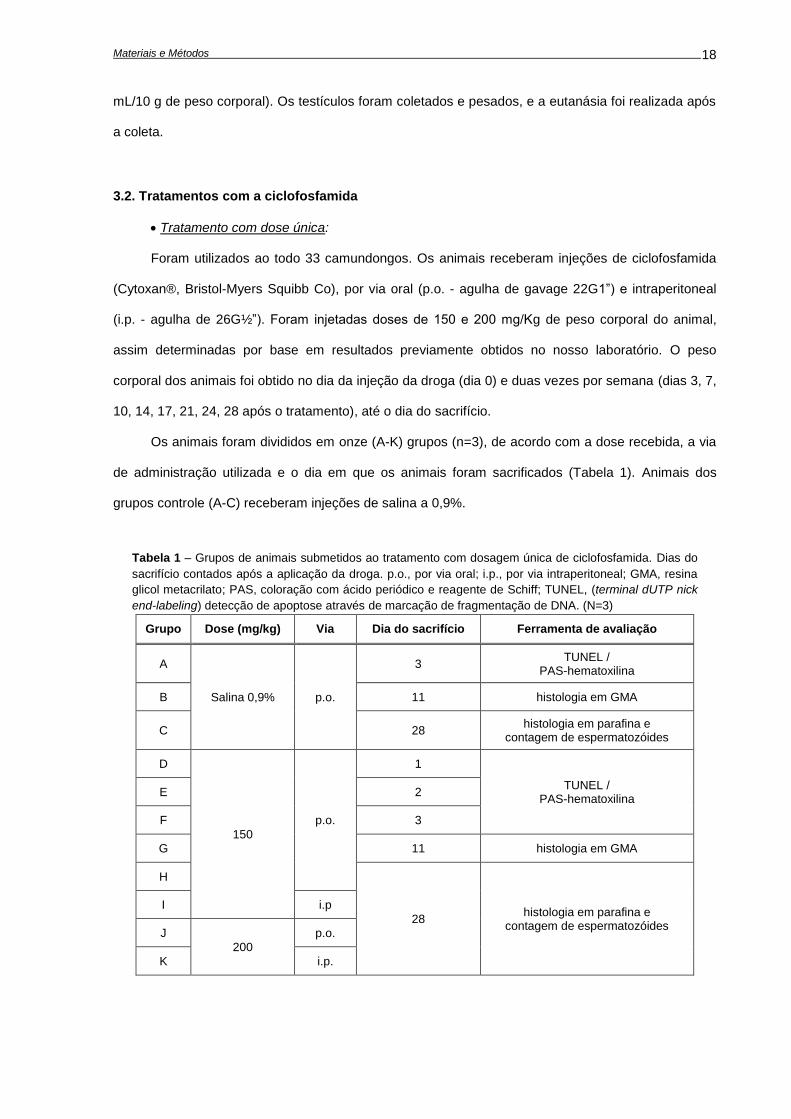
Tabela 1 – Grupos de animais submetidos ao tratamento com dosagem única de ciclofosfamida. Dias do
sacrifício contados após a aplicação da droga. p.o., por via oral; i.p., por via intraperitoneal; GMA, resina
glicol metacrilato; PAS, coloração com ácido periódico e reagente de Schiff; TUNEL, (terminal dUTP nick
end-labeling) detecção de apoptose através de marcação de fragmentação de DNA. (N=3)
Grupo Dose (mg/kg) Via Dia do sacrifício Ferramenta de avaliação
A
Salina 0,9% p.o.
3 TUNEL /
PAS-hematoxilina
B 11 histologia em GMA
C 28 histologia em parafina e
contagem de espermatozóides
D
150
p.o.
1
TUNEL / PAS-hematoxilina
E 2
F 3
G 11 histologia em GMA
H
28 histologia em parafina e
contagem de espermatozóides
I i.p
J
200
p.o.
K i.p.

Materiais e Métodos
19
Tratamento com múltiplas doses, aplicadas a cada 5 dias:
Foram utilizados ao todo 65 camundongos. Diferentes grupos de animais receberam 6 injeções
de ciclofosfamida por via oral, ou 6 injeções por via intraperitoneal, em doses de 150mg/Kg de peso
corporal, por injeção. Durante o tratamento, os animais foram separados em grupos de acordo com a
via de administração utilizada (Tabela 2). Os animais do grupo controle receberam injeções de salina
a 0,9% através de via oral, ou intraperitoneal.
Tabela 2 – Grupo de animais submetidos ao tratamento com múltiplas doses de ciclofosfamida, com
intervalos de 5 dias entre cada dose. p.o., por via oral; i.p., por via intraperitoneal.
Via (n) 1a injeção 2
a injeção 3
a injeção 4
a injeção 5
a injeção 6
a injeção
Controle (9)
Dia 0 Dia 5 Dia 10 Dia 15 Dia 20 Dia 25 p.o. (28)
i.p. (28)
Os animais foram pesados em todos os dias de injeção e, na primeira semana de tratamento,
em intervalos entre as injeções, (dias 0, 3, 5, 7, 10, 15, 20, 25, após o início do tratamento), a fim de
se avaliar a toxicidade do tratamento para os animais. Após o tratamento, os animais foram
agrupados (Tabela 3), de acordo com a data de sacrifício. Cada grupo recebeu três animais do grupo
controle, três animais do grupo p.o., e três animais do grupo i.p. (n=9), exceto para os animais que
foram sacrificados aos 28 dias (para avaliação sob MLAR), e aos 62 e 65 dias, que não receberam
animais do grupo controle (n=6).
Tabela 3 – Reagrupamento dos animais, após o tratamento com múltiplas doses de Ciclofosfamida, com
intervalos de 5 dias entre cada dose. GMA, resina glicol metacrilato; MLAR, microscopia de luz de alta resolução; PAS, coloração com ácido periódico e reagente de Schiff; TUNEL, (terminal dUTP nick end-labeling) detecção de apoptose através de marcação de fragmentação de DNA. (N=3)
Dia do sacrifício (dias após a primeira injeção)
28 Dias (n=9 / 6)
59 Dias (n=9)
62 Dias (n=6)
65 Dias (n=6)
70 Dias (n=9)
81 Dias (n=9)
histologia em GMA, TUNEL/PAS-hem,
contagem de espermatozóides,
MLAR (n=6)
histologia em parafina
e contagem de espermatozóides
Os dias do sacrifício foram determinados prevendo-se a recuperação da produção
espermática, a partir da sobrevivência dos diferentes tipos espermatogoniais potencialmente

Materiais e Métodos
20
afetados pela ciclofosfamida após o tratamento (Lu & Meistrich, 1979; de Rooij, 1998),
conforme a seguir:
Dia do sacrifício Dias após a última injeção Tipo celular produzindo espermatozóides
------- 59 -------- --------------- 34 --------------- Espermatogônia A2, estádio IX
------- 62 -------- --------------- 37 --------------- Espermatogônia Aalinhada, estádio VI
------- 65 -------- --------------- 40 --------------- Espermatogônia Apareada, estádio I
------- 70 -------- --------------- 45 --------------- Espermatogônia Aisolada, estádio VI
------- 81 -------- --------------- 56 --------------- Espermatogônia Aisolada, início do ciclo
Tratamento com múltiplas doses, aplicadas a cada 4 dias:
Foram utilizados ao todo 20 camundongos. Os animais do grupo tratado receberam 7 injeções
de ciclofosfamida, na dosagem de 150mg/Kg de peso corporal, por injeção. Animais do grupo controle
receberam 7 injeções de salina a 0,9%. Durante o tratamento os animais foram divididos em grupos,
de acordo com o tratamento recebido (Tabela 4).
Tabela 4 – Grupo de animais submetidos ao tratamento com múltiplas doses de ciclofosfamida, com
intervalos de 4 dias entre cada dose. p.o., por via oral.
Tratamento (n) 1a
injeção
2a
injeção
3a
injeção
4a
injeção
5a
injeção
6a
injeção
7a
injeção
Controle ( 2 ) Dia 0 Dia 4 Dia 8 Dia 12 Dia 16 Dia 20 Dia 24
p.o. ( 10 )
Os animais foram pesados em todos os dias de injeção (dias 0, 4, 8, 12, 16, 20, 24, após o
início do tratamento), a fim de se avaliar a toxicidade do tratamento para os animais. Após o
tratamento, os animais foram agrupados (Tabela 5), de acordo com a data de sacrifício.
Tabela 5 – Reagrupamento dos animais, após o tratamento com múltiplas doses de ciclofosfamida, com
intervalos de 4 dias entre cada dose. GMA, resina glicol metacrilato.
Data de sacrifício (dias após a primeira injeção)
28 Dias (n=5 + 1 controle) 81 Dias (n=5 + 1 controle)
contagem de espermatozóides, histologia com GMA,
contagem de espermatozóides, histologia com parafina,

Materiais e Métodos
21
Além disso, 10 camundongos foram submetidos ao mesmo regime de tratamento (7 injeções
de 150p.o., a cada 4 dias), e sacrificados 4 dias após a última dose. Esses animais tiveram seus
testículos coletados para obtenção de suspensão celular, que foi injetada em animais receptores,
para verificar a capacidade de colonização e diferenciação destas células.
3.3. Transplante de células germinativas
Obtenção da suspensão celular:
Após a coleta, em uma placa de cultura estéril, os testículos tiveram suas túnicas albugíneas
removidas e os parênquimas testiculares foram pesados todos juntos. Em seguida os testículos foram
colocados em uma garrafa estéril, com 15mL de solução de enzima colagenase a 0,5% (Worthington
– 233U/mg) em meio DFD (meio de cultura estéril DMEM/F12 (Gibco – Cat.# 11330-032) enriquecido
com aminoácidos, antibióticos e soro, contendo DNAse I (Sigma – Cat.# DN-25) na concentração de
100µg/mL). A primeira digestão, com colagenase, foi realizada por 20 minutos, em agitador
automático aquecido a 35oC por banho-maria, nas velocidades de 120rpm (jovens e adultos não
tratados) ou 300rpm (adultos tratados com ciclofosfamida e criptorquidismo). Foram utilizadas
velocidades de agitação diferentes para grupos de animais adultos tratados, por esses animais
possuírem uma estrutura testicular diferente. As velocidades de agitação foram determinadas,
baseando-se em experimentos prévios (dados não publicados).
Em seguida, a solução sobrenadante foi retirada, e sobre o tecido foram adicionados 15mL de
solução de enzima colagenase a 0,5% e enzima hialuronidase a 0,5% (Worthington – 550U/mg), em
meio DFD, em agitador automático aquecido a 35oC, por 20 minutos, na velocidade de 120rpm. O
tecido foi então lavado com solução estéril de tampão fosfato salina, para retirada das enzimas.
Finalmente, o tecido foi submetido a uma última digestão com 15mL de solução de enzima
tripsina a 0,1% (Worthington – Cat.# 3704), ácido etileno glicol tetra-acético (ethylene glycol
tetraacetic acid – EGTA) a 1mM (Sigma – Cat.# E4378) e 100µg/mL de DNase I, em tampão fosfato
salina. A digestão foi realizada por 10 minutos, em agitador automático aquecido a 35oC, na
velocidade de 120rpm. Após a digestão, a solução foi homogeneizada com pipeta de plástico e a
ação das enzimas foi neutralizada pela adição de 1,5mL de soro fetal bovino.
A solução final obtida foi filtrada com filtro de nylon de 40µm, para um tubo falcon de 50mL. Foi
retirada uma alíquota da suspensão celular para contagem em microscópio de contraste de fase com

Materiais e Métodos
22
hemocitômetro, e o restante da suspensão foi centrifugado, à temperatura ambiente, na velocidade de
1000rpm, por 10minutos. Depois de centrifugada a solução, o sobrenadante foi retirado e o pellet de
células restante foi re-suspendido com meio DFD, de acordo com a quantidade de células contadas
anteriormente, para uma concentração final de 2-6x107 células/mL de solução.
Após adição de solução de azul de tripan a 0,4% à suspensão celular final, foi possível verificar
a viabilidade da suspensão obtida, através da contagem celular em hemocitômetro, considerando-se
como mortas aquelas células coradas em azul. Os tubos contendo a suspensão celular foram
mantidos em gelo antes e durante o processo de injeção no animal receptor.
Injeção da suspensão celular:
No momento do transplante, os animais foram anestesiados com ketamina e xilazina
(0,1mL/10g de peso corporal) e foram colocados sobre um suporte que mantinha a parte abdominal
mais elevada que o resto do corpo, para facilitar a cirurgia. Foi realizada tricotomia da região
abdominal, seguida de higienização com Iodo Povidine. Com um bisturi foi feita uma incisão de
aproximadamente um centímetro na região abdominal, no sentido longitudinal ao corpo do animal. Os
testículos foram retirados pela cavidade abdominal, expostos e dissecados para melhor visualização
dos dúctulos eferentes.
A injeção da suspensão celular nos testículos foi feita através da inserção de agulhas,
produzidas a partir de capilares de vidro, nos dúctulos eferentes. A agulha estava conectada a um
longo tubo de plástico, que por sua vez estava conectado a uma seringa de diâmetro grande (50mL),
e a pressão de injeção se dava por pressionamento do embolo da seringa. Para melhor visualização
do campo cirúrgico, a cirurgia foi realizada com o auxílio de um microscópio estereoscópico.
Após a injeção da suspensão celular, os testículos foram recolocados na cavidade abdominal,
e a parede abdominal e a pele foram suturadas com linha de seda Vicryl 18”. Os animais foram
mantidos em observação até o término do efeito do anestésico, e em seguida foram alojados no
biotério.
3.4. Coleta e fixação dos testículos
Para todos os procedimentos de coleta, após serem pesados, os animais foram anestesiados
com uma mistura de ketamina (6,7mg/mL) e xilazina (1,3mg/mL) a uma proporção de 0,10 mL por 10

Materiais e Métodos
23
gramas de peso corporal do animal e a eutanásia se deu após o procedimento de coleta, através de
deslocamento cervical.
Nos estudos histológicos, para padronização do tratamento com ciclofosfamida, os testículos
foram fixados em solução de Bouin por 30 minutos e, após esse período, foram seccionados
transversalmente, sendo novamente colocados no mesmo fixador por mais 12 horas, à temperatura
ambiente. Em seguida os tecidos foram transferidos para solução saturada de carbonato de lítio em
álcool 70% e submetidos a várias trocas diárias, até completa descoloração do tecido. Finalmente os
tecidos foram armazenados em álcool 70%, até o momento da inclusão.
A fim de se estudar a morfologia espermatogonial com detalhes mais apurados, análises do
epitélio seminífero foram realizadas através da MLAR. Para isso, os animais foram submetidos ao
processo de perfusão intravascular. Aproximadamente 15 minutos antes do procedimento de
perfusão, injetou-se, pela via intraperitoneal, heparina (Liquemine Roche) na dosagem de 125 UI/kg
de peso corporal. Após a anestesia, por meio de injeção intraperitoneal de ketamina/xilazina (0,10
mL/10 g de peso corporal), procedeu-se à abertura da cavidade torácica com o objetivo de expor o
coração. Um cateter (20G - 1,1mm) foi introduzido no ventrículo esquerdo e o leito vascular lavado
com solução salina a 0,9%, a uma pressão de aproximadamente 80 mmHg. Após a retirada do
sangue com solução salina, que durou entre 5 a 10 minutos, iniciou-se o procedimento de fixação
através da perfusão com glutaraldeído a 5% em tampão cacodilato de sódio 0,05M (pH 7,3). Após um
período de perfusão de aproximadamente 25 minutos, os testículos foram retirados, pesados e
recortados em pequenos fragmentos de 1 a 2 mm de espessura. Estes fragmentos foram re-fixados
por imersão no mesmo fixador por mais 12 a 24 horas, a 4o C. Este procedimento baseou-se em
método já descrito por Sprando (1990) e Chiarini-Garcia & Meistrich (2008).
Para animais receptores do transplante, no momento da coleta, após serem pesados, os
testículos tiveram suas túnicas albugíneas removidas e foram fixados com paraformaldeído a 4%, em
tampão fosfato salina a 0,01M, por 24 horas. Após esse período os testículos foram seccionados
transversalmente, na região mediana do órgão, e colocados em tampão fosfato salina até o momento
da inclusão. Antes de serem processadas para inclusão, as metades dos testículos foram novamente
seccionadas transversalmente, aumentando-se assim a área testicular disponível para análises
morfométricas.

Materiais e Métodos
24
3.5. Processamento histológico dos testículos
O material fixado com solução de Bouin foi submetido à inclusão em parafina e/ou em glicol
metacrilato (GMA), conforme indicado nas tabelas 1, 3 e 5. Os blocos obtidos foram seccionados em
cortes na espessura de 5m, para confecção de lâminas.
As lâminas obtidas foram coradas com PAS (ácido periódico-reagente de Schiff), e contra-
coradas com hematoxilina. A reação de PAS permite a melhor visualização da vesícula acrossômica,
metodologia esta que vem sendo aplicada com sucesso para a classificação dos diferentes estádios
do ciclo do epitélio seminífero. Para tanto, as lâminas foram colocadas em ácido periódico por 10
minutos, sendo rapidamente lavadas em água destilada após o contato com o ácido. Em seguida as
lâminas foram colocadas em reagente de Schiff por 10 minutos, e em seqüência, colocadas em água
corrente por 10 minutos. A coloração com hematoxilina foi feita por 15 segundos, após os quais as
lâminas foram colocadas em água corrente por aproximadamente 5 minutos. Após a coloração, o
material foi submetido a uma série de desidratação em álcool (70%, 80%, 2x 90%, 2x 100), e a uma
substância solvente livre de água (Citrosolv ou xilol), sendo finalmente montada com Entellan (Merck
– Cat.# 107961).
Os testículos de animais receptores do transplante, fixados em paraformaldeído a 4%, foram
submetido à inclusão em parafina. Os blocos obtidos foram seccionados em cortes na espessura de
5m, para confecção de lâminas. As secções foram colocadas em lâminas cobertas
(Colorfrost®/Plus, Fisher Scientific – Cat.# 12-550-17).
As inclusões em parafina e glicol metacrilato, e a confecção das lâminas histológicas, foram
realizadas pelo centro de histologia do Departamento de Radiologia Oncológica Experimental, do MD
Anderson Cancer Center, (Houston/TX, EUA).
O material submetido a reações de TUNEL foi incluído em parafina, e os blocos seccionados
em cortes de 5 m de espessura. As secções foram colocadas em lâminas cobertas
(Colorfrost®/Plus, Fisher Scientific – Cat.# 12-550-17). As inclusões em parafina, e a confecção das
lâminas, também foram realizadas pelo centro de histologia do Departamento de Radiologia
Oncológica Experimental, do MD Anderson Cancer Center, (Houston/TX, EUA).
Os fragmentos de testículo preparados para MLAR foram submetidos à fixação secundária em
tetróxido de ósmio (EMS, Hatfield, PA – Cat.# 19110) reduzido (tetróxido de ósmio a 1% em
ferrocianeto de potássio a 1,5%) por 90 minutos, e em seguida desidratados em concentrações

Materiais e Métodos
25
crescentes de álcool (50º, 70º, 85º, 2 x 95º e 3 x 100º) e em acetona. Após a desidratação, os
fragmentos foram incluídos em resina Araldite 502 (EMS, Hatfield, PA – Cat.# 13900). Os tecidos
incluídos em blocos foram seccionados em cortes histológicos na espessura de 1m. As secções
histológicas obtidas foram coradas com azul de toluidina-borato de sódio a 1%, montadas com
Entellan (Merck – Cat.# 107961).
3.6. Contagem de espermatozóides testiculares
Para realização de contagem de espermatozóides testiculares, o testículo direito foi pesado,
com túnica albugínea, e colocado em tubos de plástico, contendo 1 mL de água destilada. Em
seguida o conteúdo dos tubos foi submetido a homogeneização, com homogeneizador Polytron
(Brinkmann Instruments, Westbury/NY, EUA), por 30 segundos, mantendo as amostras em balde com
gelo quando essas não estavam sendo homogeneizadas. Em seguida as amostras foram sonicadas
com Sonicador Brason 20-kHz (Modelo W185, Heat Systems, Ultrasonic, Plainview/NY, EUA),
durante 4 minutos, na calibragem número 7, sempre mantendo os tubos contendo as amostras em
água com gelo. É importante ressaltar que apenas as espermátides alongadas mais avançadas
(fases 13 a 16) e os espermatozóides são células resistentes ao processo de homogeneização,
seguido por sonicação, de forma que a utilização desses processos objetivou eliminar as células não
resistentes. A contagem de espermatozóides foi feita em microscópio de contraste de fase, utilizando
o aumento de 40x. As amostras foram contadas em duplicatas, colocando-se 7µL retirados das
amostras em cada lado de um hemocitômetro.
Para calcularmos o número de espermatozóides a seguinte fórmula foi aplicada:
Onde: = média dos valores contados
= número de quadrados disponíveis para contagem na área analisada do
hemocitômetro
= número de quadrados que foram contados na área analisada do
hemocitômetro
= Volume em que o tecido foi homogeneizado
= Peso Testicular

Materiais e Métodos
26
3.7. TUNEL e imunohistoquímica
Após o tratamento com ciclofosfamida, quantificamos a ocorrência de apoptoses
espermatogoniais, e de pré-leptótenos, através da técnica de TUNEL (Terminal dUTP nick end-labeling -
DeadEndTM
Colometric TUNEL System, Promega, USA – Cat.# G7130), seguindo o manual técnico
que acompanha o kit de reação. Após a reação com 3,3'-diaminobenzidina (DAB, Vector Labs – Cat.#
SK4100), as lâminas foram submetidas à lavagem em água destilada e foram coradas com PAS
(ácido periódico-reagente de Schiff) e contra-coradas com hematoxilina, seguindo-se o mesmo
protocolo utilizado no item 3.5 (Processamento histológico dos testículos). Como citado
anteriormente, a reação de PAS permite a melhor visualização da vesícula acrossômica, tornando
assim possível identificarmos os diferentes estádios do CES, e, conseqüentemente, determinar quais
os tipos espermatogoniais estavam sofrendo apoptose.
Após o transplante, utilizamos a técnica de imunohistoquímica para reconhecermos as colônias
produzidas pelas células dos animais doadores, nos testículos dos animais irradiados. Para isso,
foram utilizados anticorpos marcadores específicos para diferenciação celular (anti-GCNA-1 – germ
cell nuclear antigen, getilmente cedidos pelo Dr George Enders), e marcadores específicos para a
proteína fluorescente verde, presente nas células provenientes dos animais doadores (anticorpo
policlonal coelho-anti-GFP, Cat.# NB600-303; Novus Biologicals). Cortes seriados foram marcados
com os dois tipos de anticorpos separadamente, tornando assim possível identificar, dentre os túbulos
que apresentavam diferenciação celular (positivos para GCNA-1), quais túbulos eram formados por
células germinativas dos animais doadores (positivas para GFP).
As secções foram incubadas com os anticorpos primários (monoclonal rato anti-camundongo
para proteína GCNA-1, na diluição 1:100; e policlonal coelho anti-GFP na diluição de 1:300) em
câmara úmida, na temperatura de 4oC, durante 12 horas. Após incubação com os anticorpos
primários, as secções foram incubadas com anticorpo secundário biotinilado anti-camundongo (Vector
Labs – Cat.# PK6102, diluição 1:500), em temperatura ambiente, por um período de 30 minutos. Para
detecção da ligação dos anticorpos, após a incubação com anticorpo secudário, as lâminas foram
submetidas ao kit ABC, anti-coelho e anti-rato, (Vector Labs – Cat.# PK6102) por 30 minutos à
temperatura ambiente. Finalmente as secções foram reveladas com DAB (Vector Labs – Cat.#
SK4100), lavadas em água destilada, e coradas hematoxilina. Após a coloração, as secções foram
desidratadas e as lâminas foram montadas com meio de montagem permanente (Permount, Fisher

Materiais e Métodos
27
Scientific – Cat.# SP15-100). As lâminas produzidas foram analisadas sob microscópio de luz
(Olympus BX-41).
3.8. Análises morfológicas e morfométricas
Análise qualitativa:
As análises qualitativas, em parafina, foram realizadas para todos os animais que foram
submetidos ao tratamento com ciclofosfamida. Foram verificados os tipos de células germinativas
ausentes em cada um dos dias após tratamento avaliados, e as possíveis alterações morfológicas.
Além disso, análises morfológicas dos subtipos espermatogoniais foram realizadas através da
MLAR, a fim de se verificar possíveis alterações morfológicas das espermatogônias, quando
comparadas com os animais controle, em função do tratamento com ciclofosfamida.
Análise semi-quantitativa:
A análise semi-quantitativa, em GMA, foi realizada em animais dos grupos B (controle) e G
(150p.o.) (Tabela 1), submetidos ao experimento com dosagem única de ciclofosfamida. Com a
utilização de um quadro de pontuação, foi possível determinar quais os tipos celulares estavam sendo
afetados pela droga no momento da injeção (11 dias antes do sacrifício). Para isso, foi determinado,
nos túbulos seminíferos, se as células germinativas do tipo espermatogônias A4, Intermediária-In e B,
zigótenos, paquítenos e espermátides estavam presentes em (a) quantidades normais (mais que 20
células, para espermatócitos ou espermátides), (b) fortemente reduzidas (6 a 20 células),
(c) praticamente ausentes (1 a 5 células) ou (d) ausentes (0 células).
Número relativo de células germinativas por 100 nucléolos de células de Sertoli:
O número de cada subtipo espermatogonial (Aind, A1, A2, A3, A4, In e B), espermatócitos,
espermátides arredondadas e espermátides alongadas até a fase 12, presentes nos animais
avaliados foi calculado, e normalizado por 100 nucléolos de células de Sertoli, uma vez que esse tipo
celular apresenta-se em número invariável no epitélio seminífero de camundongos adultos. Foram
contados os números de núcleolos de células de Sertoli e de núcleos dos subtipos espermatogôniais
em aproximadamente 30 secções transversais de túbulos seminíferos, por animal.

Materiais e Métodos
28
Os números contados foram corrigidos pela fórmula de Abercrombie (Abercrombie, 1946)
utilizando-se os diâmetros nucleares de cada tipo de célula germinativa, calculados a partir da média
da medida de 10 núcleos, e o diâmetro nucleolar das células de Sertoli, calculado também a partir da
média da medida de 10 nucléolos.
Apoptoses espermatogoniais:
O número de apoptoses espermatogôniais foi calculado para os animais que receberam
injeções de ciclofosfamida, em diferentes momentos da padronização de tratamento.
Inicialmente, contamos o número de espermatogônias apoptóticas (TUNEL positivas) presentes
nos animais que receberam dosagem única de ciclofosfamida. Baseado nos dados de apoptose
obtidos por Cai e colaboradores (1997) para ratos tratados com ciclofosfamida, decidimos realizar as
análises nos dias 1, 2 e 3, após a administração da droga. Foram contadas espermatogônias TUNEL
positivas em secções transversais de túbulos seminíferos, determinando-se os estádios do CES
através da coloração de PAS. Foram contados aproximadamente 10 túbulos por estádio, calculando-
se a porcentagem de túbulos com células TUNEL positivas, por estádio.
A partir dos dados obtidos no tratamento com dosagem única de ciclofosfamida, optamos por
avaliar o índice apoptótico dos animais que receberam múltiplas dosagens, 3 dias após a última
administração da droga. Foram contadas aproximadamente 30 secções transversais de túbulos
seminíferos por animal, registrando-se o número de espermatogônias por túbulo, e de
espermatogônias TUNEL positivas por túbulo. O índice apoptótico foi calculado dividindo-se o número
total de espermatogônias TUNEL positivas pelo número total de espermatogônias contadas.
Número de colônias produzidas após o transplante:
Para as análises foram selecionadas lâminas obtidas a partir de testículos que sabidamente
receberam uma maior quantidade de células no momento do transplante. Foram selecionados 10
testículos para cada grupo, para os receptores que receberam células dos animais jovem, adultos
tratados com ciclofosfamida e criptorquídicos, e 7 testículos para o grupo que recebeu células de
animais adultos normais.
Durante a coleta os testículos foram seccionados em quatro partes, dessa forma foram
analisadas 4 secções transversais de testículos, por testículo selecionado. Nas análises, foram

Materiais e Métodos
29
contadas todas as secções transversais de túbulos seminíferos presentes em cada secção de
testículo.
Os túbulos seminíferos foram classificados quanto à positividade para GCNA-1, e os túbulos
positivos para GCNA-1 foram classificados quanto à positividade para GFP. Para se determinar a
eficiência do transplante, a partir do número de colônias formadas pelas células doadoras, e
compararmos os resultados obtidos nos diferentes grupos, a seguinte fórmula foi utilizada:
Onde: = número de túbulos GFP positivos
= número de túbulos contados
= número de células injetadas no momento do transplante
A partir das contagens e do cálculo foi possível obter o número total de colônias formadas nos
animais receptores, a porcentagem de colônias geradas por recuperação das espermatogônias tronco
endógenas, e a porcentagem de colônias formadas pelas células doadoras que colonizaram os
túbulos seminíferos dos animais receptores.
3.9. Análise estatística
Os dados morfométricos foram expressos em média erro padrão. Os grupos foram
comparados usando teste ANOVA, seguido dos testes Fisher’s LSD e Turkey’s. Diferenças foram
consideradas significativas quando p<0,05. Todas as análises foram realizadas no programa SPSS
18.0 (SPSS Inc. Chicago/USA).
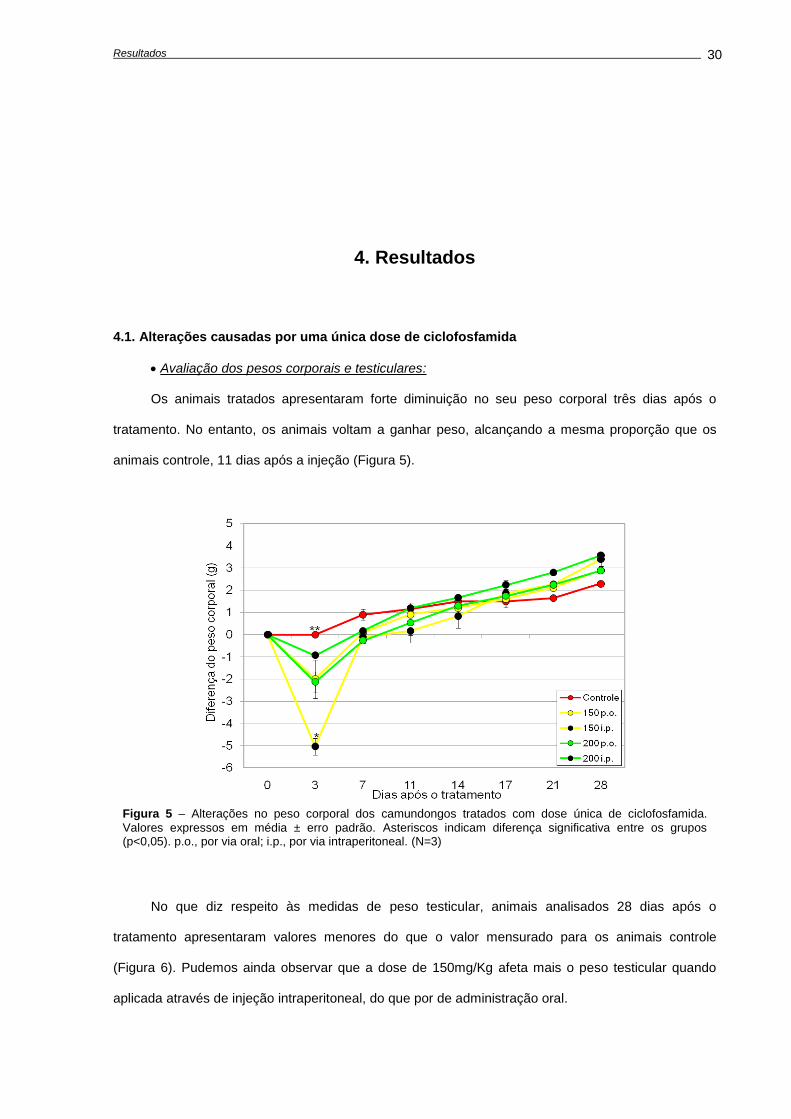
Resultados
30
4. Resultados
4.1. Alterações causadas por uma única dose de ciclofosfamida
Avaliação dos pesos corporais e testiculares:
Os animais tratados apresentaram forte diminuição no seu peso corporal três dias após o
tratamento. No entanto, os animais voltam a ganhar peso, alcançando a mesma proporção que os
animais controle, 11 dias após a injeção (Figura 5).
Figura 5 – Alterações no peso corporal dos camundongos tratados com dose única de ciclofosfamida.
Valores expressos em média ± erro padrão. Asteriscos indicam diferença significativa entre os grupos (p<0,05). p.o., por via oral; i.p., por via intraperitoneal. (N=3)
No que diz respeito às medidas de peso testicular, animais analisados 28 dias após o
tratamento apresentaram valores menores do que o valor mensurado para os animais controle
(Figura 6). Pudemos ainda observar que a dose de 150mg/Kg afeta mais o peso testicular quando
aplicada através de injeção intraperitoneal, do que por de administração oral.
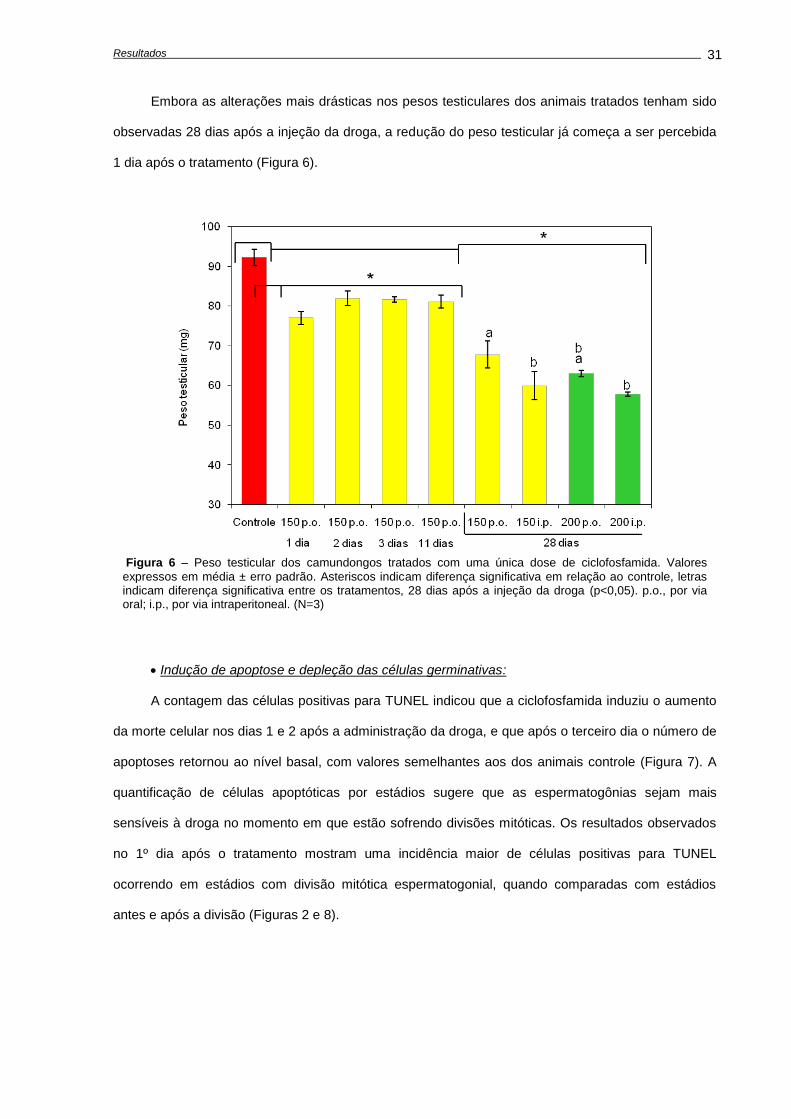
Resultados
31
Embora as alterações mais drásticas nos pesos testiculares dos animais tratados tenham sido
observadas 28 dias após a injeção da droga, a redução do peso testicular já começa a ser percebida
1 dia após o tratamento (Figura 6).
Figura 6 – Peso testicular dos camundongos tratados com uma única dose de ciclofosfamida. Valores
expressos em média ± erro padrão. Asteriscos indicam diferença significativa em relação ao controle, letras indicam diferença significativa entre os tratamentos, 28 dias após a injeção da droga (p<0,05). p.o., por via oral; i.p., por via intraperitoneal. (N=3)
Indução de apoptose e depleção das células germinativas:
A contagem das células positivas para TUNEL indicou que a ciclofosfamida induziu o aumento
da morte celular nos dias 1 e 2 após a administração da droga, e que após o terceiro dia o número de
apoptoses retornou ao nível basal, com valores semelhantes aos dos animais controle (Figura 7). A
quantificação de células apoptóticas por estádios sugere que as espermatogônias sejam mais
sensíveis à droga no momento em que estão sofrendo divisões mitóticas. Os resultados observados
no 1º dia após o tratamento mostram uma incidência maior de células positivas para TUNEL
ocorrendo em estádios com divisão mitótica espermatogonial, quando comparadas com estádios
antes e após a divisão (Figuras 2 e 8).
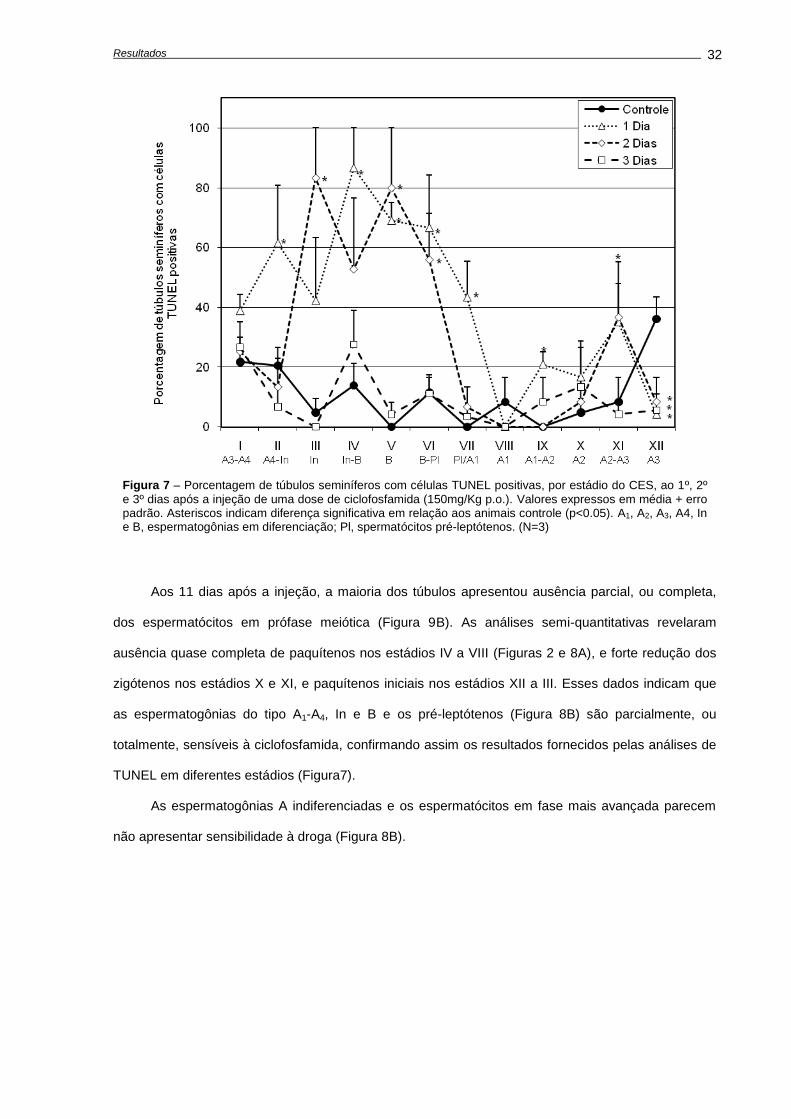
Resultados
32
Figura 7 – Porcentagem de túbulos seminíferos com células TUNEL positivas, por estádio do CES, ao 1º, 2º
e 3º dias após a injeção de uma dose de ciclofosfamida (150mg/Kg p.o.). Valores expressos em média + erro padrão. Asteriscos indicam diferença significativa em relação aos animais controle (p<0.05). A1, A2, A3, A4, In e B, espermatogônias em diferenciação; Pl, spermatócitos pré-leptótenos. (N=3)
Aos 11 dias após a injeção, a maioria dos túbulos apresentou ausência parcial, ou completa,
dos espermatócitos em prófase meiótica (Figura 9B). As análises semi-quantitativas revelaram
ausência quase completa de paquítenos nos estádios IV a VIII (Figuras 2 e 8A), e forte redução dos
zigótenos nos estádios X e XI, e paquítenos iniciais nos estádios XII a III. Esses dados indicam que
as espermatogônias do tipo A1-A4, In e B e os pré-leptótenos (Figura 8B) são parcialmente, ou
totalmente, sensíveis à ciclofosfamida, confirmando assim os resultados fornecidos pelas análises de
TUNEL em diferentes estádios (Figura7).
As espermatogônias A indiferenciadas e os espermatócitos em fase mais avançada parecem
não apresentar sensibilidade à droga (Figura 8B).

Resultados
33
Figura 8 – Células germinativas organizadas em doze estádios do CES. A) Células germinativas observadas 11 dias após uma injeçao de cicofosfamida. B) Células germinativas afetadas pela ciclofosfamida no
momento da injeçao. Quantidade de células foi cassificada em Ausentes; Praticamente ausentes; ou
Fortemente reduzidas. A1, A2, A3, A4, In e B, espermatogônias em diferenciação; preL, pre-leptótenos; L, leptótenos; Z, zigótenos; P, paquítenos; D, diplótenos; M!, divisões meióticas I e II.
Avaliações realizadas aos 28 dias após o tratamento:
As análises histológicas do epitélio seminífero 28 após a injeção de 150mg/Kg p.o. (Figura 9C)
mostraram que as espermatogônias, os espermatócitos e as espermátides se regeneraram
relativamente bem, a partir das espermatogônias-tronco sobreviventes. Apesar de a maior parte das
espermátides alongadas estar ausente (estádios II-VIII; Figuras 9B e 2), ainda foi possível observar
uma quantidade aparentemente normal de espermátides alongadas em estágios iniciais de
desenvolvimento nos estádios IX-XII. Em contraste, para os tratamentos utilizando 150mg/Kg i.p. e
A
B
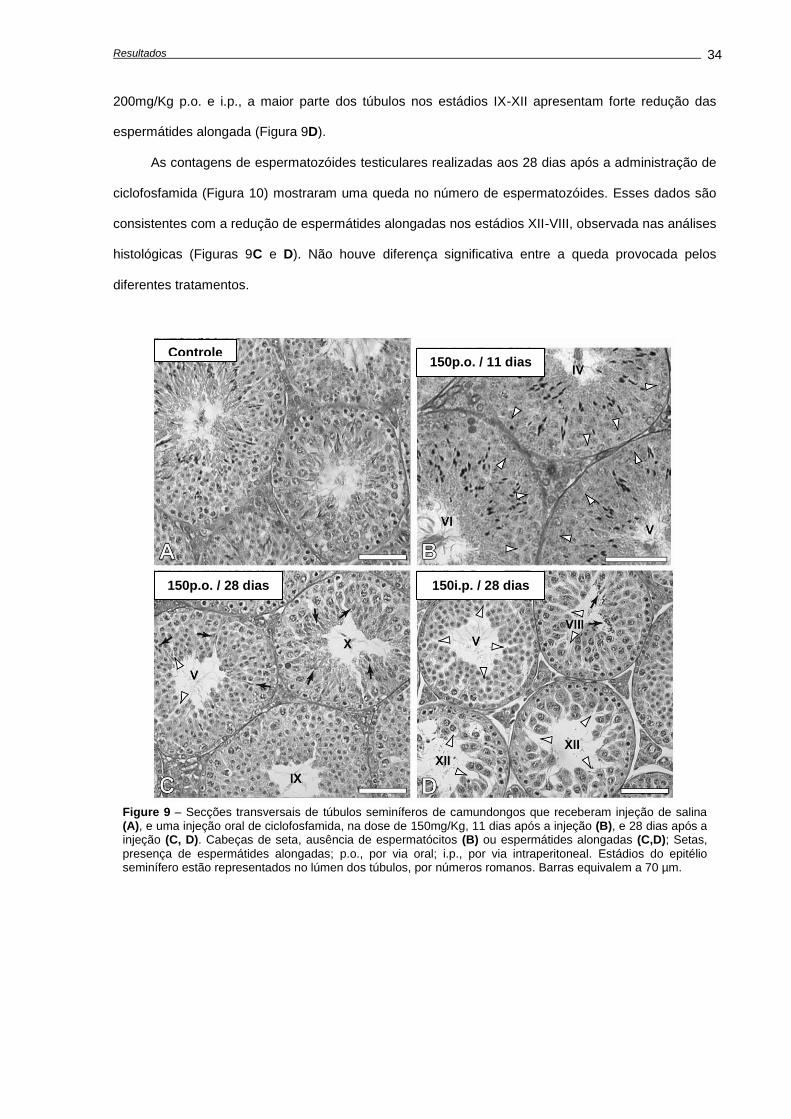
Resultados
34
200mg/Kg p.o. e i.p., a maior parte dos túbulos nos estádios IX-XII apresentam forte redução das
espermátides alongada (Figura 9D).
As contagens de espermatozóides testiculares realizadas aos 28 dias após a administração de
ciclofosfamida (Figura 10) mostraram uma queda no número de espermatozóides. Esses dados são
consistentes com a redução de espermátides alongadas nos estádios XII-VIII, observada nas análises
histológicas (Figuras 9C e D). Não houve diferença significativa entre a queda provocada pelos
diferentes tratamentos.
Figure 9 – Secções transversais de túbulos seminíferos de camundongos que receberam injeção de salina (A), e uma injeção oral de ciclofosfamida, na dose de 150mg/Kg, 11 dias após a injeção (B), e 28 dias após a injeção (C, D). Cabeças de seta, ausência de espermatócitos (B) ou espermátides alongadas (C,D); Setas,
presença de espermátides alongadas; p.o., por via oral; i.p., por via intraperitoneal. Estádios do epitélio seminífero estão representados no lúmen dos túbulos, por números romanos. Barras equivalem a 70 µm.
Controle 150p.o. / 11 dias
150p.o. / 28 dias 150i.p. / 28 dias
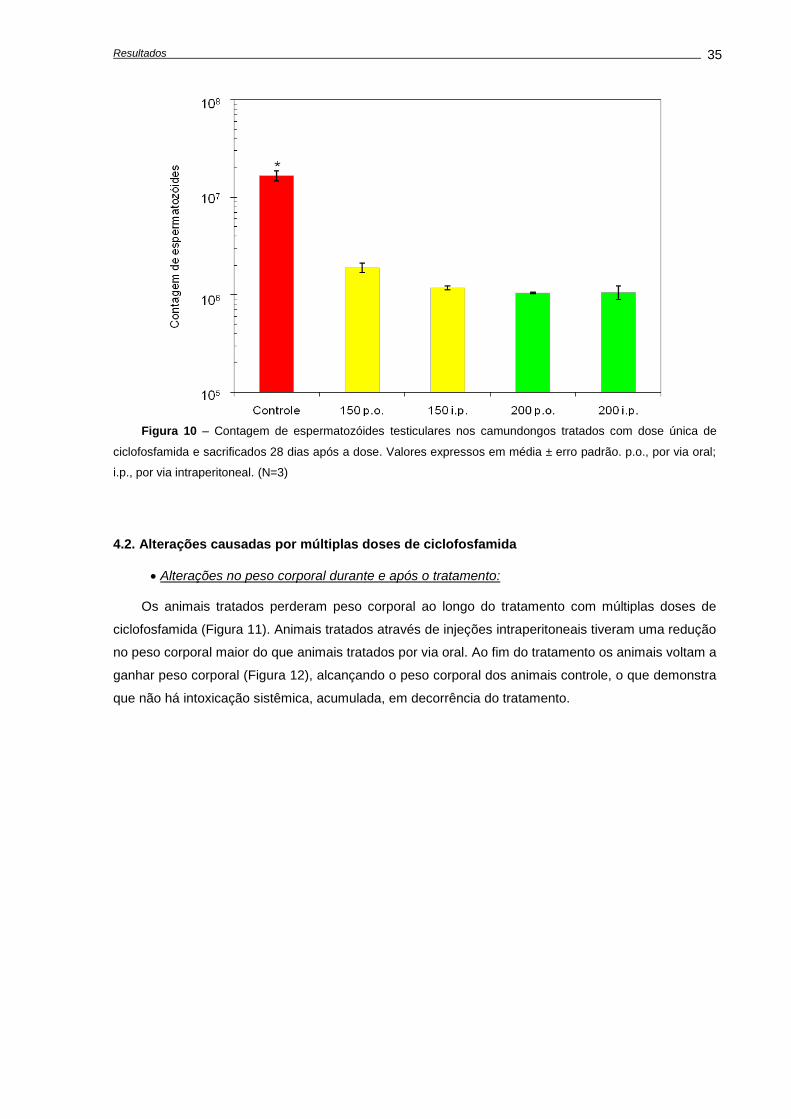
Resultados
35
Figura 10 – Contagem de espermatozóides testiculares nos camundongos tratados com dose única de
ciclofosfamida e sacrificados 28 dias após a dose. Valores expressos em média ± erro padrão. p.o., por via oral;
i.p., por via intraperitoneal. (N=3)
4.2. Alterações causadas por múltiplas doses de ciclofosfamida
Alterações no peso corporal durante e após o tratamento:
Os animais tratados perderam peso corporal ao longo do tratamento com múltiplas doses de
ciclofosfamida (Figura 11). Animais tratados através de injeções intraperitoneais tiveram uma redução
no peso corporal maior do que animais tratados por via oral. Ao fim do tratamento os animais voltam a
ganhar peso corporal (Figura 12), alcançando o peso corporal dos animais controle, o que demonstra
que não há intoxicação sistêmica, acumulada, em decorrência do tratamento.
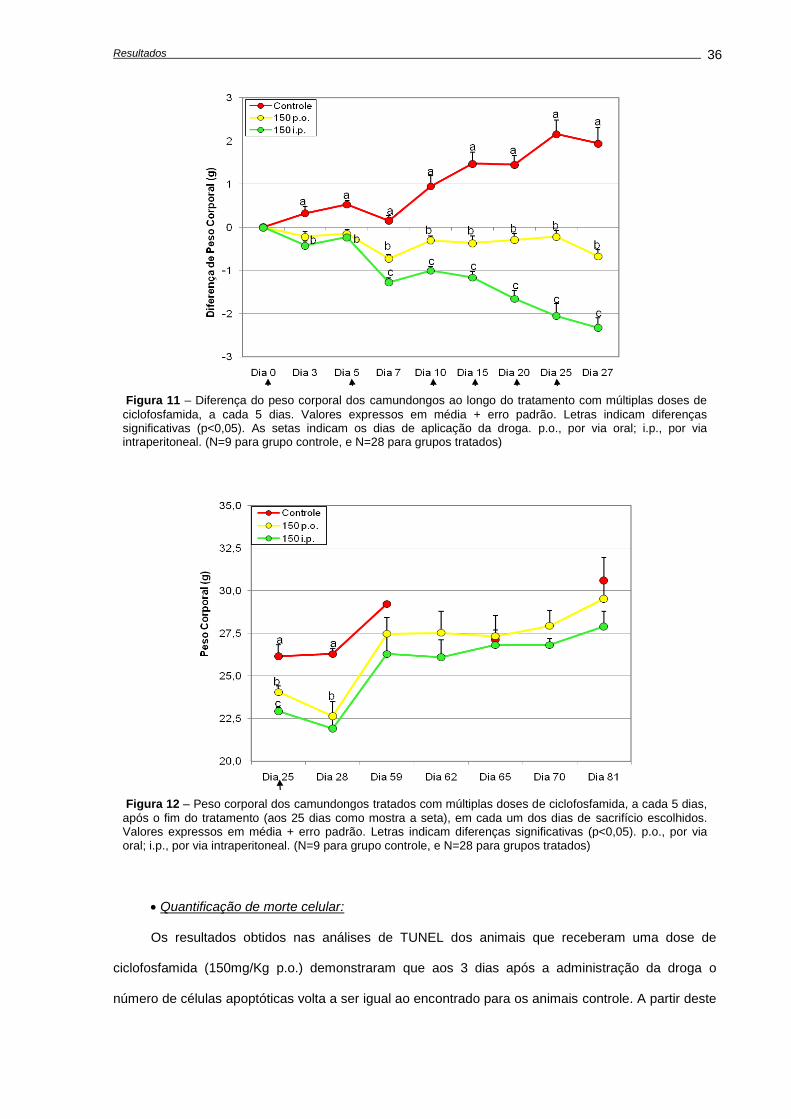
Resultados
36
Figura 11 – Diferença do peso corporal dos camundongos ao longo do tratamento com múltiplas doses de
ciclofosfamida, a cada 5 dias. Valores expressos em média + erro padrão. Letras indicam diferenças significativas (p<0,05). As setas indicam os dias de aplicação da droga. p.o., por via oral; i.p., por via intraperitoneal. (N=9 para grupo controle, e N=28 para grupos tratados)
Figura 12 – Peso corporal dos camundongos tratados com múltiplas doses de ciclofosfamida, a cada 5 dias,
após o fim do tratamento (aos 25 dias como mostra a seta), em cada um dos dias de sacrifício escolhidos. Valores expressos em média + erro padrão. Letras indicam diferenças significativas (p<0,05). p.o., por via oral; i.p., por via intraperitoneal. (N=9 para grupo controle, e N=28 para grupos tratados)
Quantificação de morte celular:
Os resultados obtidos nas análises de TUNEL dos animais que receberam uma dose de
ciclofosfamida (150mg/Kg p.o.) demonstraram que aos 3 dias após a administração da droga o
número de células apoptóticas volta a ser igual ao encontrado para os animais controle. A partir deste
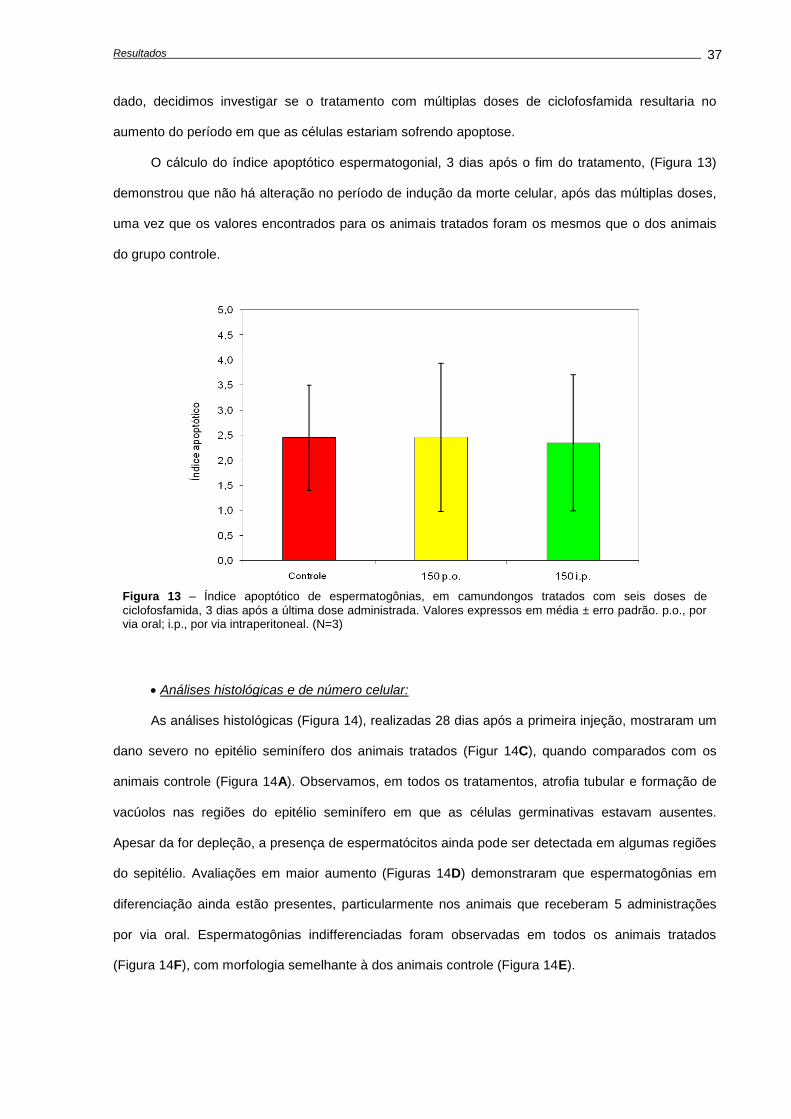
Resultados
37
dado, decidimos investigar se o tratamento com múltiplas doses de ciclofosfamida resultaria no
aumento do período em que as células estariam sofrendo apoptose.
O cálculo do índice apoptótico espermatogonial, 3 dias após o fim do tratamento, (Figura 13)
demonstrou que não há alteração no período de indução da morte celular, após das múltiplas doses,
uma vez que os valores encontrados para os animais tratados foram os mesmos que o dos animais
do grupo controle.
Figura 13 – Índice apoptótico de espermatogônias, em camundongos tratados com seis doses de
ciclofosfamida, 3 dias após a última dose administrada. Valores expressos em média ± erro padrão. p.o., por via oral; i.p., por via intraperitoneal. (N=3)
Análises histológicas e de número celular:
As análises histológicas (Figura 14), realizadas 28 dias após a primeira injeção, mostraram um
dano severo no epitélio seminífero dos animais tratados (Figur 14C), quando comparados com os
animais controle (Figura 14A). Observamos, em todos os tratamentos, atrofia tubular e formação de
vacúolos nas regiões do epitélio seminífero em que as células germinativas estavam ausentes.
Apesar da for depleção, a presença de espermatócitos ainda pode ser detectada em algumas regiões
do sepitélio. Avaliações em maior aumento (Figuras 14D) demonstraram que espermatogônias em
diferenciação ainda estão presentes, particularmente nos animais que receberam 5 administrações
por via oral. Espermatogônias indifferenciadas foram observadas em todos os animais tratados
(Figura 14F), com morfologia semelhante à dos animais controle (Figura 14E).

Resultados
38
Figura 14 – Secções transversais de túbulos seminíferos de camundongos que receberam injeção de salina (A), e em maior aumento (B) mostrando células de Sertoli (S) e espermatogônias em diferenciação do tipo B (B). (C) túbulos seminíferos de camundongos que receberam múltiplas doses de ciclofosfamida (150mg/Kg
p.o.) com intervalos de 5 injeções, 28 dias após a primeira injeção, apresentando espermatócitos ainda presentes no epitélio seminífero (cabeça de seta). Em maior aumento (D) pode-se observar células de Sertoli
e espermatogônias do tipo B. Podem ser observadas ainda, espermatogônias do tipo A indifferenciadas em animais do grupo controle (E) e animais tratados (F). Em (G) túbulos seminíferos de animais tratados 81 dias
após a primeira injeção, mostrando recuperação na maior parte dos túbulos (**), porém alguns túbulos ainda se mostram danificados (*). Barras pretas equivalem a 70 µm, e barras brancas equivalem a 20 µm (B, D) e 10 µm (E,F).
As contagens de células germinativas mostraram que, aos 28 dias, todos os tratamentos
resultaram em um menor número de espermatogônias em diferenciação, espermatócitos e
espermátides (Figura 15). As análises de número de espermatogônias após 25 dias de tratamento
com ciclofosfamida por administração oral mostraram que algumas espermatogônias intermediárias e
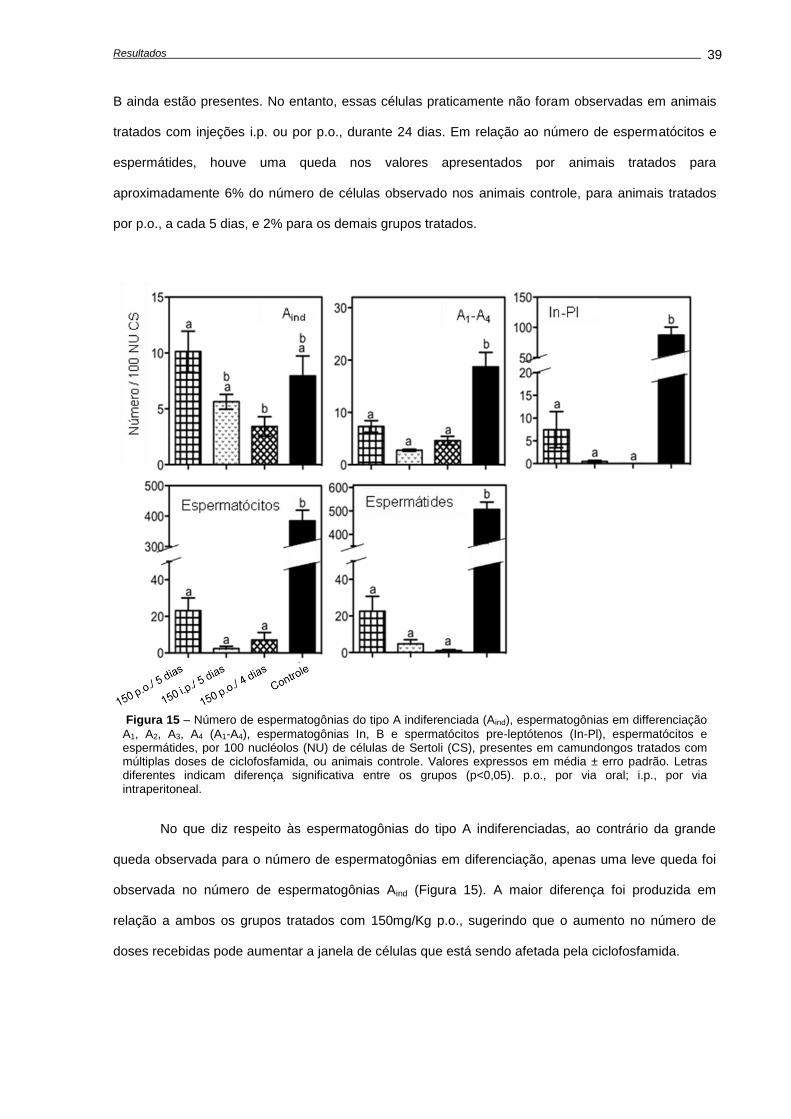
Resultados
39
B ainda estão presentes. No entanto, essas células praticamente não foram observadas em animais
tratados com injeções i.p. ou por p.o., durante 24 dias. Em relação ao número de espermatócitos e
espermátides, houve uma queda nos valores apresentados por animais tratados para
aproximadamente 6% do número de células observado nos animais controle, para animais tratados
por p.o., a cada 5 dias, e 2% para os demais grupos tratados.
Figura 15 – Número de espermatogônias do tipo A indiferenciada (Aind), espermatogônias em differenciação
A1, A2, A3, A4 (A1-A4), espermatogônias In, B e spermatócitos pre-leptótenos (In-Pl), espermatócitos e espermátides, por 100 nucléolos (NU) de células de Sertoli (CS), presentes em camundongos tratados com múltiplas doses de ciclofosfamida, ou animais controle. Valores expressos em média ± erro padrão. Letras diferentes indicam diferença significativa entre os grupos (p<0,05). p.o., por via oral; i.p., por via intraperitoneal.
No que diz respeito às espermatogônias do tipo A indiferenciadas, ao contrário da grande
queda observada para o número de espermatogônias em diferenciação, apenas uma leve queda foi
observada no número de espermatogônias Aind (Figura 15). A maior diferença foi produzida em
relação a ambos os grupos tratados com 150mg/Kg p.o., sugerindo que o aumento no número de
doses recebidas pode aumentar a janela de células que está sendo afetada pela ciclofosfamida.
*

Resultados
40
4.3. Recuperação do epitélio seminífero ao fim do tratamento
Análises histológicas:
As análises morfológicas, realizadas 56 dias após a última dose (80/81 dias após a primeira
dose), revelaram que o epitélio seminífero dos animais tratados se recupera relativamente bem
(Figura 14G). O diâmetro dos túbulos seminíferos voltou a ser semelhante ao dos animais controle
(Figura 14A), e não foram observados vacúolos, como os presentes aos 28 dias. Embora raros,
túbulos seminíferos danificados ainda podem ser observados nos animais tratados.
Análise da recuperação da função testicular:
A fim de verificar a funcionalidade das espermatogônias-tronco presentes ao fim do tratamento,
nós analisamos a recuperação da espermatogênese nos animais tratados, em diferentes períodos
após a primeira injeção. Ambos os parâmetros analisados, peso testicular (Figura 16A) e contagem
de espermatozóides testiculares (Figura 16B), demonstram que há uma recuperação da função
testicular. O tratamento utilizando injeções intraperitoneais apresentou a menor recuperação dentre
os grupos, e é possível que o tratamento via i.p. tenha causado danos não só ao funcionamento das
espermatogônias-trono, mas aos componentes somáticos do epitélio seminífero.
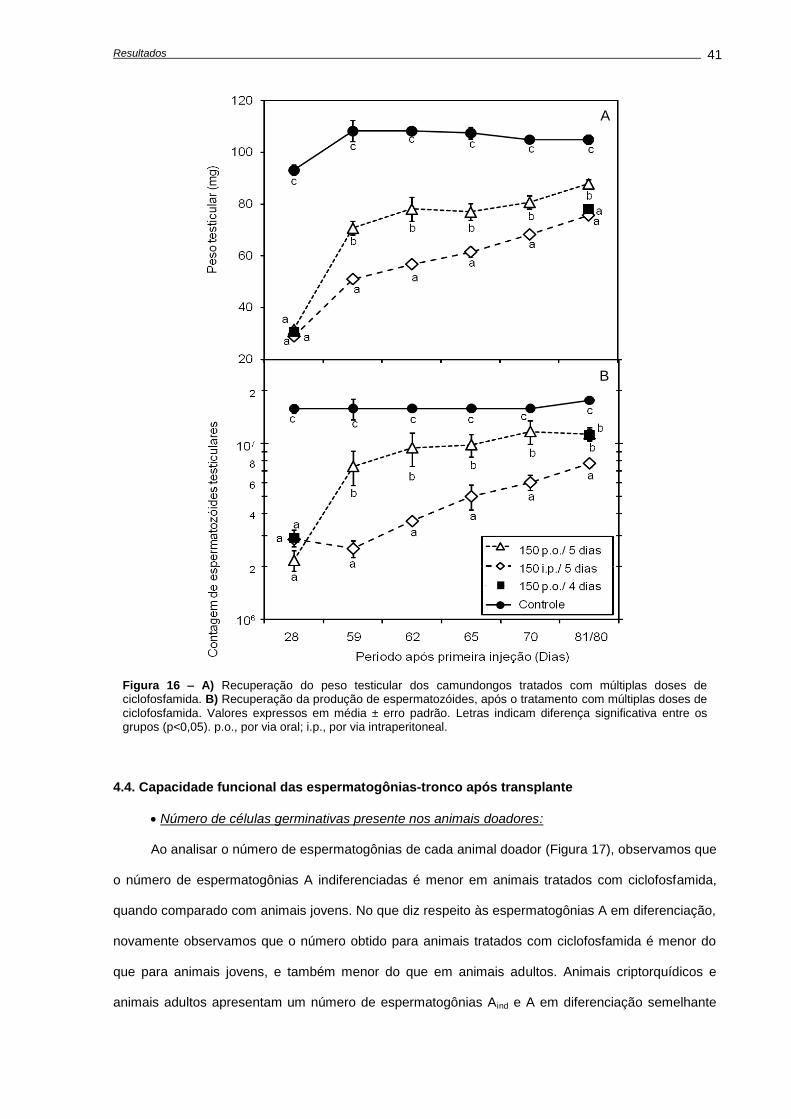
Resultados
41
Figura 16 – A) Recuperação do peso testicular dos camundongos tratados com múltiplas doses de ciclofosfamida. B) Recuperação da produção de espermatozóides, após o tratamento com múltiplas doses de
ciclofosfamida. Valores expressos em média ± erro padrão. Letras indicam diferença significativa entre os grupos (p<0,05). p.o., por via oral; i.p., por via intraperitoneal.
4.4. Capacidade funcional das espermatogônias-tronco após transplante
Número de células germinativas presente nos animais doadores:
Ao analisar o número de espermatogônias de cada animal doador (Figura 17), observamos que
o número de espermatogônias A indiferenciadas é menor em animais tratados com ciclofosfamida,
quando comparado com animais jovens. No que diz respeito às espermatogônias A em diferenciação,
novamente observamos que o número obtido para animais tratados com ciclofosfamida é menor do
que para animais jovens, e também menor do que em animais adultos. Animais criptorquídicos e
animais adultos apresentam um número de espermatogônias Aind e A em diferenciação semelhante
A
B
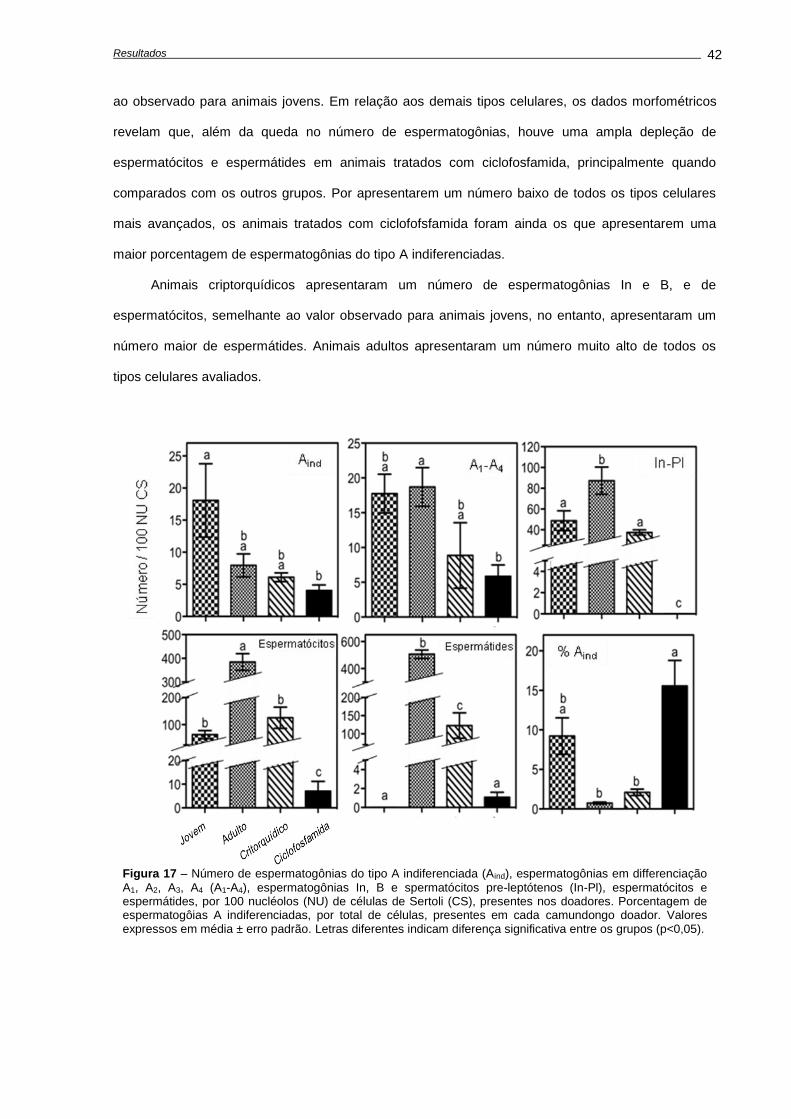
Resultados
42
ao observado para animais jovens. Em relação aos demais tipos celulares, os dados morfométricos
revelam que, além da queda no número de espermatogônias, houve uma ampla depleção de
espermatócitos e espermátides em animais tratados com ciclofosfamida, principalmente quando
comparados com os outros grupos. Por apresentarem um número baixo de todos os tipos celulares
mais avançados, os animais tratados com ciclofofsfamida foram ainda os que apresentarem uma
maior porcentagem de espermatogônias do tipo A indiferenciadas.
Animais criptorquídicos apresentaram um número de espermatogônias In e B, e de
espermatócitos, semelhante ao valor observado para animais jovens, no entanto, apresentaram um
número maior de espermátides. Animais adultos apresentaram um número muito alto de todos os
tipos celulares avaliados.
Figura 17 – Número de espermatogônias do tipo A indiferenciada (Aind), espermatogônias em differenciação
A1, A2, A3, A4 (A1-A4), espermatogônias In, B e spermatócitos pre-leptótenos (In-Pl), espermatócitos e espermátides, por 100 nucléolos (NU) de células de Sertoli (CS), presentes nos doadores. Porcentagem de espermatogôias A indiferenciadas, por total de células, presentes em cada camundongo doador. Valores expressos em média ± erro padrão. Letras diferentes indicam diferença significativa entre os grupos (p<0,05).

Resultados
43
Características das suspensões celulares:
As contagens celulares realizadas no momento da obtenção das suspensões, revelaram que
animais criptorquídicos e animais tratados com ciclofosfamida apresentaram peso testicular médio e o
número de células obtido na suspensão celular semelhantes entre si (Tabela 6). Camundongos
jovens, por sua vez, forneceram um número maior de células por testículo do que os grupos de
animais tratados. Como esperado, o peso testicular médio e o número de células obtidas na
suspensão celular dos animais adultos, não tratados, foram maiores que os dos outros animais,
devido ao fato de todas as células germinativas estarem presentes nesses animais. Vale ressaltar
que o protocolo de obtenção da suspensão celular foi o mesmo para todos os animais, de forma que
o procedimento não interferiu no resultado do número de células por testículo obtido.
Tabela 6 – Peso testicular dos camundongos doadores, número de testículos utilizados e número de
células obtido a partir dos animais doadores no momento da obtenção da suspensão celular. O peso
testicular foi obtido para todos os testículos juntos e dividido pelo número de testículos coletados. Para os
grupos jovens, criptorquídicos e tratados com ciclofosfamida, foram realizados 2 experimentos. Exp,
experimento.
Doador Peso Testicular (mg) Número de Testículos Células / Testículo (x106)
Adulto 104,0 10 21,2
Jovem 11,5 10 (1º Exp) - 6 (2º Exp) 1,8 (1º Exp) – 5,0 (2º Exp)
Adulto/Criptorquídico 27,1 6 (1º Exp) - 13 (2º Exp) 1,0 (1º Exp) - 1,4 (2º Exp)
Adulto/Ciclofosfamida 29,9 8 (1º Exp) - 10 (2º Exp) 1,0 (1º Exp) - 1,6 (2º Exp)
Análises morfométricas dos camundongos receptores, após o transplante:
Após a coloração com imunoistoquímica, foi possível reconhecer, dentre os túbulos seminíferos
que apresentavam diferenciação celular (positivos para GCNA1 - Figura 18A e C), quais eram
consequência de recuperação endógena da espermatogênese (negativos para GFP - Figura 18B) e
quais eram resultado da diferenciação das espermatogônias transplantadas (positivos para GFP -
Figura 18D).
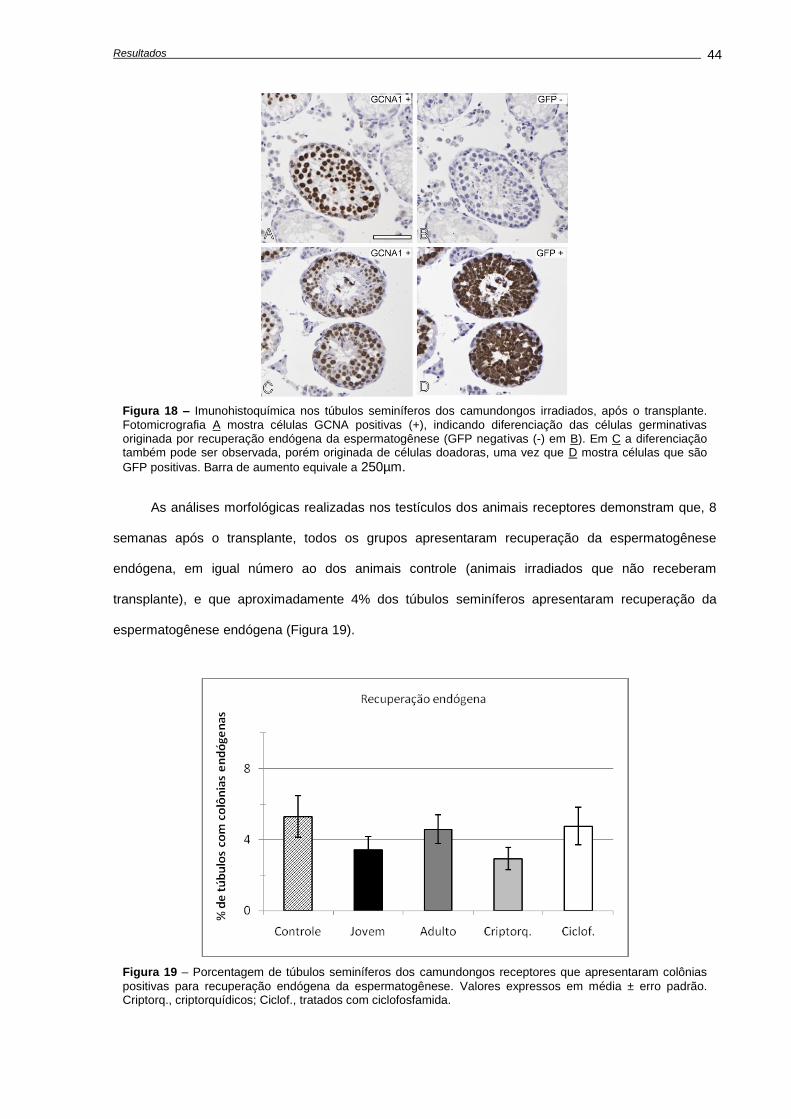
Resultados
44
Figura 18 – Imunohistoquímica nos túbulos seminíferos dos camundongos irradiados, após o transplante.
Fotomicrografia A mostra células GCNA positivas (+), indicando diferenciação das células germinativas originada por recuperação endógena da espermatogênese (GFP negativas (-) em B). Em C a diferenciação também pode ser observada, porém originada de células doadoras, uma vez que D mostra células que são
GFP positivas. Barra de aumento equivale a 250µm.
As análises morfológicas realizadas nos testículos dos animais receptores demonstram que, 8
semanas após o transplante, todos os grupos apresentaram recuperação da espermatogênese
endógena, em igual número ao dos animais controle (animais irradiados que não receberam
transplante), e que aproximadamente 4% dos túbulos seminíferos apresentaram recuperação da
espermatogênese endógena (Figura 19).
Figura 19 – Porcentagem de túbulos seminíferos dos camundongos receptores que apresentaram colônias
positivas para recuperação endógena da espermatogênese. Valores expressos em média ± erro padrão. Criptorq., criptorquídicos; Ciclof., tratados com ciclofosfamida.
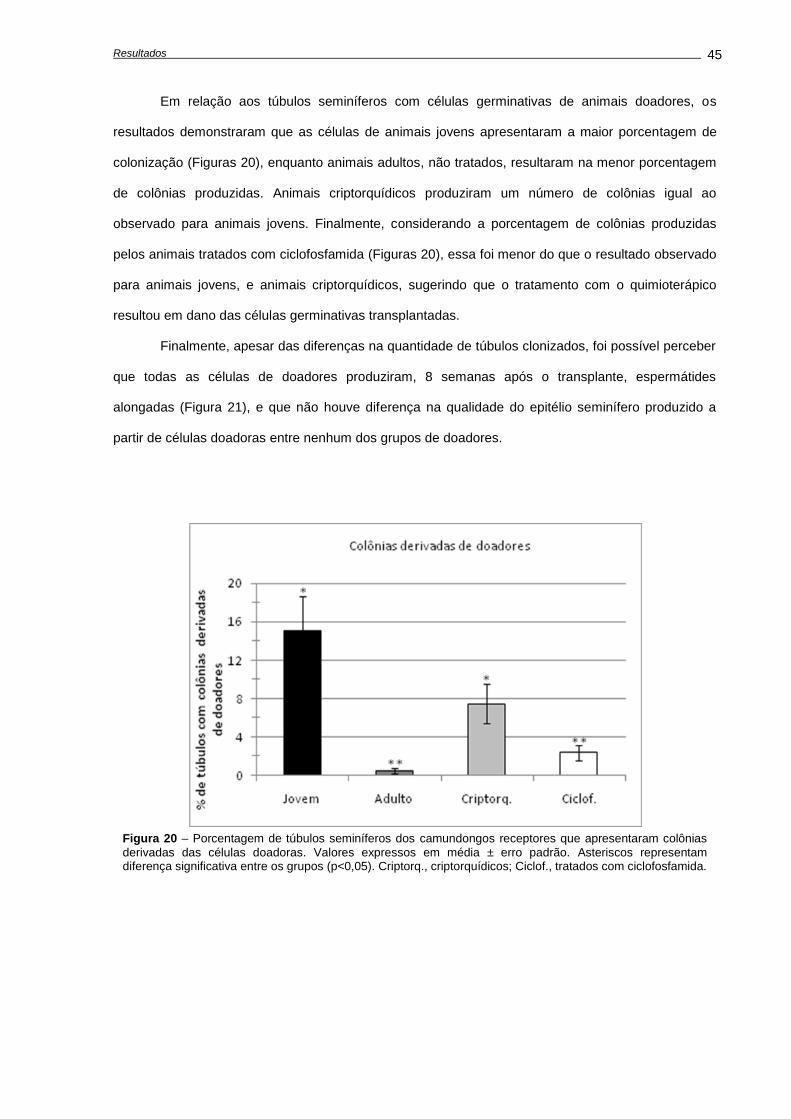
Resultados
45
Em relação aos túbulos seminíferos com células germinativas de animais doadores, os
resultados demonstraram que as células de animais jovens apresentaram a maior porcentagem de
colonização (Figuras 20), enquanto animais adultos, não tratados, resultaram na menor porcentagem
de colônias produzidas. Animais criptorquídicos produziram um número de colônias igual ao
observado para animais jovens. Finalmente, considerando a porcentagem de colônias produzidas
pelos animais tratados com ciclofosfamida (Figuras 20), essa foi menor do que o resultado observado
para animais jovens, e animais criptorquídicos, sugerindo que o tratamento com o quimioterápico
resultou em dano das células germinativas transplantadas.
Finalmente, apesar das diferenças na quantidade de túbulos clonizados, foi possível perceber
que todas as células de doadores produziram, 8 semanas após o transplante, espermátides
alongadas (Figura 21), e que não houve diferença na qualidade do epitélio seminífero produzido a
partir de células doadoras entre nenhum dos grupos de doadores.
Figura 20 – Porcentagem de túbulos seminíferos dos camundongos receptores que apresentaram colônias
derivadas das células doadoras. Valores expressos em média ± erro padrão. Asteriscos representam diferença significativa entre os grupos (p<0,05). Criptorq., criptorquídicos; Ciclof., tratados com ciclofosfamida.
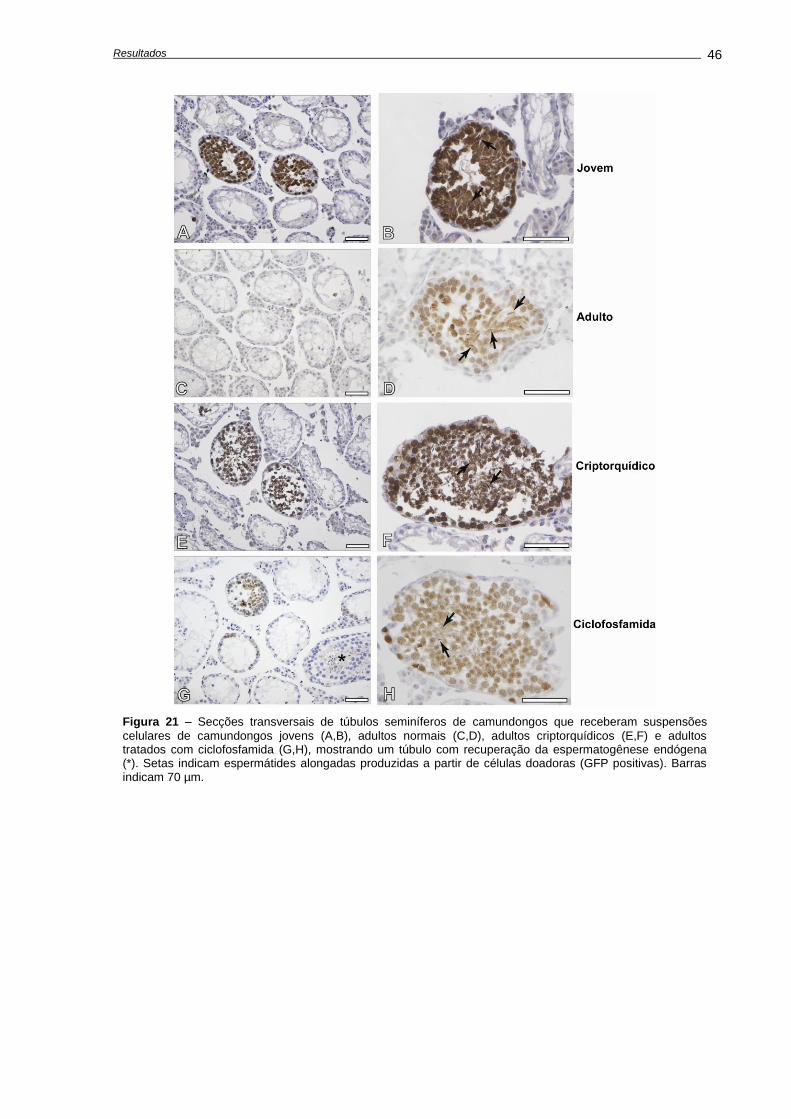
Resultados
46
Figura 21 – Secções transversais de túbulos seminíferos de camundongos que receberam suspensões
celulares de camundongos jovens (A,B), adultos normais (C,D), adultos criptorquídicos (E,F) e adultos tratados com ciclofosfamida (G,H), mostrando um túbulo com recuperação da espermatogênese endógena (*). Setas indicam espermátides alongadas produzidas a partir de células doadoras (GFP positivas). Barras indicam 70 µm.

Discussão 47
5. Discussão
No presente estudo nós desenvolvemos um regime de tratamento com múltiplas doses de
ciclofosfamida capaz de eliminar as células mais diferenciadas dos testículos, mantendo a população
de espermatogônias do tipo A indiferenciadas. Nós encontramos um balanço entre doses, vias de
administração e intervalos entre doses, ideais para eliminar desde espermatogônias em diferenciação
até espermátides mais avançadas, sendo menos tóxico para o animal, e ainda permitindo a
sobrevivência de um número considerável de células-tronco espermatogoniais, e a recuperação da
espermatogênese in situ, a partir das células sobreviventes. O regime de tratamento utilizado reduziu
o número de células diferenciadas para 1% dos valores observados para animais do grupo controle,
porém deixou aproximadamente 50% de espermatogônias do tipo Aind. O enriquecimento da
população de espermatogônias-tronco, através do tratamento com ciclofosfamida, poderia ser
utilizado como um passo inicial de purificação celular, para uso em estudos moleculares que
objetivam elucidar os efeitos de ciclofosfamida sobre essas células.
Esses resultados são consistentes com uma série de outros estudos presentes na literatura que
demonstram, em roedores, a sensibilidade das espermatogônias em diferenciação a tratamentos com
ciclofosfamida, porém uma relativa resistência das células-tronco a uma variedade de regimens de
tratamento (Lu & Meistrich, 1979; Karashima et al., 1988; Velez et al., 1989, Russel & Russel, 1991;
Cai et al., 1997). Isto é observado em contraste com outras drogas quimioterápicas, tais como
bussulfan, doxorrubicina e procarbazina, que efetivamente eliminam as espermatogônias-tronco,
assim como as espermatogônias em diferenciação, do epitélio seminífero (Meistrich et al., 1982;
Bucci & Meistrich, 1987; Brinster & Avarbock, 1994).
Ao final do tratamento, a presença de espermatogônias Aind aparentemente saudáveis permitiu a
observação de subsequente recuperação da espermatogênese nos animais tratados. Já aos 45 dias
após a última injeção, antes que as espermatogônias Ais tivessem tempo de sofrer auto-renovação e

Discussão 48
ainda produzir espermátides avançadas, a quantidade de espermatozóides testiculares observada
nos animais tratados foi de aproximadamente 63% dos resultados apresentados pelo grupo controle.
Apesar da sobrevivência das espermatogônias-tronco, e da recuperação da espermatogênese
in situ, essas células parecem apresentar uma perda de função espermatogênica quando
manipuladas para transplante. Apesar de animais tratados com ciclofosfamida terém apresentado as
maiores concentrações de células Aind como porcentagem de células germinativas totais, a habilidade
destas células de produzirem colônias após o transplante foi de apenas 16% dos valores
apresentados para animais jovens, e 32% dos valores apresentados para animais critorquídicos. Em
relação aos animais pre-púberes, é difícil comparar, quatitativamente, os resultados apresentados por
estes doadores com resultados observados em animais tratados ciclofosfamida, uma vez que o
número de células obtidas por mg de testículo foi muito maior para animais jovens. Isso pode ter
ocorrido porque as células de Sertoli nestes animais são ainda imaturas, e não estabeleceram
junções oclusivas, facilitando a recuperação destas células nas suspensões celulares. A presença de
células de Sertoli imaturas nas suspensões celulares transplantadas pode ter colaborado para o
sucesso de colonização das células de doadores, uma vez que as células de Sertoli podem ter
funcionado como nichos adicionais para a recuperação da espermatogênese nos animais receptores
(Kanatsu-Shinohara et al., 2005; Zhang et al, 2009), embora nós não tenhamos observado células de
Sertoli positivas para GFP nos camundongos receptores.
É possível estabelecer um paralelo entre animais criptorquídicos e animais tratados com
ciclofosfamida, uma vez que esses doadores parecem ser semelhantes em vários aspectos. Os
pesos testículares e o número de células obtidas por testículos foram os mesmos observados para
ambos os grupos, e o número de espermatogônias Aind por células de Sertoli também foram
semelhantes. Como ambos os modelos são provenientes de camundongos adultos, os tipos de
células somáticas, e a estrutura do epitélio seminífero deveriam ser basicamente os mesmos. Ainda
assim, embora a porcentagem de espermatogônias Aind entre as células germinativas tenha sido
aproximadamente 7 vezes maior nos animais tratados com ciclofosfamida, a colonização gerada por
células de animais tratados com ciclofosfamida foi muito menor do que a colonização gerada por
células de animais criptorquídicos. Embora a digestão com colagenase tenha removido a maioria das
células intersticiais, e tenha resultado na perda de muitas células de Sertoli durante a digestão dos
túbulos seminíferos (Mesitrich et al., 1973), é provável que algumas células ainda estejam presentes

Discussão 49
na suspensão celular. Mesmo que todas as células de Sertoli tenham sido recuperadas na
suspensão, a concentração de espermatogônias Aind ainda seria 2,6 vezes maior nas suspensões
celulares obtidas a partir de testículos de camundongos tratados com ciclofosfamida, e dessa forma
uma colonização 2,6 vezes maior era esperada para esses animais. Assim, a diferença na eficiência
de colonização sugere que as espermatogônias-tronco de animais tratados com ciclofosfamida
apresentem um dano.
É de comum acordo que as espermatogônias-tronco estão contidas na população celular
conhecida como espermatogônias Aind. Existem relatos na literatura de que espermatogônias
positivas para c-kit sejam capazes de colonizar testículos receptores (Barroca et al., 2008), sugerindo
que essas células possam ser espermatogônias em diferenciação do tipo A. Se isso fosse verdade,
então o número reduzido destas células em animais tratados com ciclofosfamida poderia ter
colaborado para uma eficiência de colonização baixa, resultante das células destes animais. Porém, a
espressão de c-kit começa a ser detectada em células Aal em estágio tardio de diferenciação
(Nakagawa et al., 2010). Dessa forma, a habilidade de colonização das células positivas para c-kit é
mais provalmente exercida por células do tipo Aal, que pertencem às espermatogônias
indiferenciadas, ou possivelmente por espermatogônia do tipo A1 em fase inicial de diferenciação. No
entanto o número de espermatogônias A1 não está reduzido nos animais tratados (dados não
mostrados), e a baixa habilidade de colonização não pode ser explicada pela ausência de
espermatogônias em diferenciação.
O método histológico empregado, microscopia de luz de alta resolução, permite a identificação
de espermatogônias Aind, porém não mostra o tramanho das correntes espermatogoniais, ou fornece
outros tipos de marcadores. A população de espermatogônias que contribui para o funcionamento
normal da espermatogênese está contida dentro da sub-população celular conhecida como Ais, no
entando espermatogônias do tipo Apr e Aal ocasionalmente contribuem para a população de células-
tronco, através da fragmentação das pontes intercitoplasmáticas (Nakagawa et al., 2010). Essas
células são consideradas células-tronco potenciais, e contribuem para a recuperação da
espermatogênese após uma grande depleção celular, como a que ocorre após o tratamento com o
agente quimioterápico bussulfan, ou após transplante de espermatogônias (Nakagawa et al., 2007).
Embora nós não tenhamos determinado a distribuiçao de Ais, Apr, e Aal dentro das espermatogônias
Aind, nós podemos concluir que a porcentagem de células-tronco propriamente ditas (Ais) deveria ser

Discussão 50
consideravelmente alta nos testículos de animais tratados com ciclofosfamida. A presença de
espermatogônias Apr e Aal, que constituem a maior parte das células Aind, aos 4 dias após a última
administração de ciclofosfamida, indica que as células progenitoras, células-tronco, tenham
sobrevivido às 6 injeções de ciclofosfamida, e dessa forma devem ter sobrevivido à sétima
administração.
A baixa eficiência de colonização, quando comparados com animais criptorquídicos, com
certeza reflete diferenças na qualidade das espermatogônias Aind desses dois grupos. Estudos que
permiticem a caracterização do tamanho dos clones celulares, e a observação dos marcadores de
diferenciação espermatogoniais na população de Aind sobrevivente, poderiam esclarecer as razões de
porque há diferenças nas espermatogônias-tronco de animais criptorquídicos e daqueles tratados
com ciclofosfamida. É importante destacar que os nichos que abrigam in situ as espermatogônias-
tronco de animais criptorquídicos e de animais tratados com ciclofosfamida podem ser muito
diferentes. Isso faz com que as espermatogônias, mesmo tendo sido transplantadas para ambientes
semelhantes (testículos de camundongos irradiados), se comportem de forma completamente
diferentes, explicando assim as diferenças na eficiência de colonização.
Também é possivel que tenha havido morte celular de pelo menos parte das espermatogônias-
tronco, mas que in situ a morte celular tenha sido compensada por fragmentação dos clones
celulares, e que tenha ocorrido aumento dos ciclos de auto-renovação das células-tronco (de Rooij,
1988). Apesar da morte de parte das espermatogônias-tronco, a recuperação in situ deve ter ocorrido
devido ao aumento nas divisões celulares de espermatogoônias Aal, e à redução da apoptose de
espermatogônias A em diferenciação (de Rooij, 2001). Assim, o transplante de células germinativas
se torna um ensaio direto da quantificação de espermatogônias-tronco, pois a recuperação endógena
da espermatogênese não é sensível a perdas celulares moderadas no número de células-tronco.
Dessa forma, a eficiência de colonização maior em animais criptorquídicos pode refletir a presença de
mais células-tronco saudáveis do que em animais tratados com ciclofosfamida.
Alternativamente, é possivel que um número maior de células-tronco saudáveis esteja presente
nos testículos de animals tratado com ciclofosfamida, no entato elas se recuperam melhor in situ, e
apresentam algum dano à habilidade de colonizarem testículos dos animais receptores. Nosso grupo
mostrou que a habilidade de colonização e diferenciação de células de doadores imaturos pode ser
melhorada após a supressão de testosterona nos animais receptores, antes e depois do transplante

Discussão 51
de células germinativas (Wang et al., 2010). Assim, talvez seja interessante avaliar se a supressão
hormonal poderia melhorar a atividade funcional das espermatogônias submetidas a ciclofosfamida,
nos testículos de animais receptores.
Em resumo, as espermatogônias-tronco de camundongos sobrevivem a múltiplas doses de
ciclofosfamida, e uma recuperação considerável da espermatogênese endógena é observada após o
tratamento. No entanto, ainda há evidências de perda de espermatogônias-tronco, ou de um dano
residual destas células quando elas são isoladas para transplante, porém o motivo pelo qual isso
ocorreria ainda tem que ser esclarecido. A observação da perda e dano celular em camundongos
tratados com ciclofosfamida pode fornecer métodos para melhor compreender a grande sensibilidade
dos testículos de pacientes à prologada azoospermia após o tratamento com ciclofsfamida (Hsu et al.,
1979; Fukutani et al, 1981).

Conclusão 52
6. Conclusão
Ao final do estudo, chegamos às seguintes conclusões:
Assim como em ratos, em camundongos uma dose de ciclofosfamida induz apoptose das
espermatogônias nos primeiros dois dias após tratamento;
O uso de múltiplas doses de ciclofosfamida não prolongou o período de apoptose induzida
por uma dose;
O tratamento com múltiplas doses de ciclofosfamida eliminou a maioria das células
germinativas em diferenciação;
Múltiplas doses de ciclofosfamida não alteraram drásticamente em número o grupo de
espermatogônias indiferenciadas;
Baseado nos dados de peso testicular e contagem de espermatozóides, seja com uma, ou
múltiplas doses, o tratamento com ciclofosfamida via intraperitoneal prejudica mais a
espermatogênese em camundongos, do que o tratamento administrado por via oral;
O tratamento com múltiplas doses de ciclofosfamida altera a capacidade de colonização das
espermatogônias-tronco.

Referências Bibliográficas 53
7. Referências Bibliográficas
Abercrombie M. (1946) Estimation of nuclear population from microtome sections.
Anatomical Records, v.94, p.239-247.
Aponte, P.M.; van Bragt. M.P.A.; de Rooij, D.G.; van Pelt, A.M.M. (2005) Spermatogonial
stem cells: characteristics and experimental possibilities. APMIS, v.113, p.727-42.
Barroca, V.; Lassalle, B.; Coureuil, M.; Louis, J.P.; Le Page, F.; Testart, J.; Allemand, I.;
Riou, L.; Fouchet, P. (2009) Mouse differentiating spermatogonia can generate germinal
stem cells in vivo. Nature Cell Biology, v.11, p.190-6.
Bolden-Tiller, O.U.; Chiarini-Garcia, H.; Poirier, C.; Alves-Freitas, D.; Weng, C.C.; Shetty, G.;
Meistrich, M.L. (2007). Genetic factors contributing to defective spermatogonial
differentiation in juvenile spermatogonial depletion (Utp14b/jsd) mice. Biology of
Reproduction, v.77, p.237-46.
Brannan, C.I.; Bedell, M.A.; Resnick, J.L.; Eppig, J.J.; Handel, M.A.; Williams, D.E. (1992)
Developmental abnormalities in Steel17H mice resul from a splicing defect in the stell
factor cytoplasmic tail. Genes & Development, v.6, p.1832-42.
Brinster, R.L. (2002) Germline stem cell transplantation and transgenesis. Science, v.296,
p.2174-6.
Brinster, R.L.; Avarbock, M.R. (1994) Germline transmission of donor haplotype following
spermatogonial transplantation. Developmental Biology, v.91, p.11303-7.
Brinster, C.J.; Ryn, B.Y.; Avarbock, M.R.; Karagenc, L.; Brinster, R.L. (2003) Restoration of
fertility by germ cell transplantation requires efficient recipient preparation. Biology of
Reproduction; v.69, p.412 –20.

Referências Bibliográficas 54
Brinster, R.L.; Zimmermann, J.W. (1994) Spermatogenesis following male germ-cell
transplantation. Developmental Biology, v.91, p.11298-302.
Buaas, F.W.; Kirsh, A.L., Sharma, M.; McLean, D.J.; Morris, J.L.; Griswold, M.D.; de Rooij,
D.G.; Braun R.E. (2004) Plzf is required in adult male germ cells for stem cell self-
renewal. Nature Genetics, v.36, p.551-3.
Buageaw, A.; Sukhwani, M.; Ben-Yehudah, A.; Ehmcke, J.; Rawe, V.Y.; Pholpramool, C.;
Orwig, K.E.; Schlatt, S. (2005) GDNF family receptor alpha1 phenotype of
spermatogonial stem cells in immature mouse testes. Biology of Reproduction, v.73,
p.1011-6.
Bucci, L.R.; Meistrich, M.L. (1987) Effects of busulfan on murine spermatogenesis:
cytotoxicity, sterility, sperm abnormalities and dominant lethal mutations. Mutation
Research, v.176, p.159-268.
Cai, L.; Hales, B.F.; Robaire, B. (1997) Induction of apoptosis in the germ cells of adult male
rats after exposure to cyclophosphamide. Biology of Reproduction, v.56, p.1490-7.
Cattanach (1974) Spermatogonial stem cell killing in the mouse following single and
fractioned X-ray doses, as assessed by length of sterile period. Mutation Research,
v.25, p.53-62.
Chiarini-Garcia, H.; Meistrich, M.L. (2008). High resolution light microscopic characterization
of spermatogonia. In: Germline Stem Cells. Ed. Hou S.X. & Singh S.R. Humana Press:
USA. 95-107p.
Chiarini-Garcia, H.; Raymer, A.M.; Russell, L.D. (2003) Non-random distribution of
spermatogonia in rats: evidence of niches in the seminiferous tubules. Reproduction,
v.126, p.669-80.
Chiarini-Garcia, H.; Russell, L.D. (2001) High resolution light microscopic characterization of
mouse spermatogonia. Biology of Reproduction, v.65, p.1170-78.
Chiarini-Garcia, H.; Russell, L.D. (2002) Characterization of mouse spermatogonial types by
electron microscopy. Reproduction, v.123, p.567-77.
Colvin, M. (2003) Chapter 51: Alkylating Agents and Platinum Antitumor Compounds. In:
Holland-Frei Cancer Medicine, Ed. Kufe, D.W.; Pollock, R.E.; Weichselbaum, R.R.;
Bast, R.C.; Gansler, T.S.; Holland, J.F.; Frei, E. BC Decker: Canada. 6th Edition.

Referências Bibliográficas 55
Costoya, J.A.; Hobbs, R.M.; Barna, M.; Cattoretti, G.; Manoya, K.; Sukhawani, M.; Orwig,
K.E.; Wolgemuth, D.J.; Pandolfi, P.P. (2004) Essential role of Plzf in maintenance of
spermatogonial stem cells. Nature Genetics, v.36, p.653-59.
Creemers, L.B.; Meng, X.; den Ouden, K.; van Pelt, A.M.; Izadyar, F.; Santoro, M.; Sariola,
H.; de Rooij, D. G. (2002) Transplantation of germ cells from glial cell line-derived
neurotrophic factor-overexpressing mice to host testes depleted of endogenous
spermatogenesis by fractioned irradiation. Biology of Reproduction; v.66, p.1579-84.
CYTOXAN: ciclofosfamida, Bristol Myers Squibb Co., Princeton/NJ, USA (2007). Bula de
remédio.
de Rooij, D.G. (1988) Regulation of the proliferation of spermatogonial stem cells. Journal of
Cell Science Supplement, v.10, p.181-194.
de Rooij, D.G. (1998) Stem cells in the testis. International Journal of Experimental
Pathology, v.79, p.67-80.
de Rooij, D.G. (2001). Proliferation and differentiation of spermatogonial stem cells.
Reproduction, v.121, p.347-54.
de Rooij, D.G.; Okabe, M.; Nishimune, Y. (1999) Arrest of spermatogonial differentiation in
jsd/jsd, Sl17H/Sl17H, and cryptorchid mice. Biology of Reproduction, v.61, p.842-47.
de Rooij, D.G.; Russell, L.D. (2000). All you wanted to know about spermatogonia but were
afraid to ask. Journal of Andrology, v.21, p.776-98.
de Ruiter-Bootsma, A.L.; Kramer, M.F.; de Rooij, D.G. (1977) Survival of spermatogonial
stem cells in the mouse after split-dose irradiation with fission neutrons of 1-MeV mean
energy or 300-kV X-rays. Radiation Research, v. 71, p. 579-92.
Dobrinski, I. (2005a) Transplantation. In: The Sertoli cell biology. Ed. Skinner M.K. &
Griswold M.D. Elsevier: New York, USA. 471-85p.
Donovan P.J.; de Migue M.P. (2005) Gonocyte development and differentiation. In: Sertoli
Cell Biology. Ed. Skinner M. K. & Griswold M. D. Elsevier: New York, USA. 417-435p.

Referências Bibliográficas 56
Dobrinski, I. (2005b) Germ cell transplantation and testis tissue xenografting in domestic
animals. Animal Reproduction Science, v.89, p.137-145.
Drumond, A.L., Meistrich, M.L., Chiarini-Garcia, H. (2011) Spermatogonial morphology and
kinetics during testis development in mice: a high resolution light microscopy approach.
Reproduction. In press.
Dym, M.; Kokkinaki, M.; He, Z. (2009) Spermatogonial stem cells: mouse and human
comparisons. Birth defects research. Part C, Embryo today, v.87, p.27-34.
Enders G.C, May J.J. (1994) Developmentally regulated expression of a mouse germ cell
nuclear antigen examined from embryonic day 11 to adult in male and female mice.
Developmental Biology, v.163, p.331-40.
Fukutani, K.; Ishida, H.; Shinohara, M.; Minowada, S.; Niijima, T.; Hijikata, K.; et al. (1981)
Supression of spermatogenesis in patients with behçet’s disease treated with
cyclophosphamide and colchicine. Fertility and Sterility, v.36, p.76-80.
Hasegawa, M.; Wilson, G.; Russell, L.D.; Meistrich, M.L. (1997) Radiation-induced cell death
in the mouse testis: relationship to apoptosis. Radiation Research, v.147, p.457-67.
Hausler C.L.; Russell, L.D. (1999) Prospects for spermatogonial transplantation in livestock
and endangered species. In: The male gamete: from basic science to clinical
applications. Ed. Gagnon C. Cache River Press: Vienna, USA, 7-45p.
Hermann, B.P.; Sukhwani, M.; Lin, C.C. ; Sheng, Y. ; Tomko, J. ; Rodriguez, M. ; et al.
(2007) Characterization, cryopreservation, and ablation of spermatogonial stem cells in
adult rhesus macaques. Stem Cells v.25, p.2330-2338.
Hofmann, M. C.; Braydich-Stolle, L.; Dym, M. (2005) Isolation of male germ-line stem cells;
influence of GDNF. Developmental Biology, v.279, p.114-24.
Huckins, C. & Clermont, Y. (1968) Evolution of gonocytes in the rat testis during late
embryonic and post-natal life. Archives of Anatomy, Histology and Embryology, v.51,
p.343-354.
Hsu, A.C.; Folami, A.O.; Bain, J.; Rance, C.P. (1979) Gonadal function in males treated with
cyclophosphamide for nephrotic syndrome. Fertility and Sterility, v.31, p.173-178.

Referências Bibliográficas 57
Izadyar, F.; Creemers, L.B.; van Dissel-Emiliani, F.M.F.; van Pelt, A.M.M.; de Rooij, D.G.
(2000) Spermatogonial stem cell transplantation. Molecular Cell Endocrinology; v.169,
p.21-6.
Kanatsu-Shinohara, M.; Toyokuni, S.; Shinohara, T. (2005) Genetic selection of mouse male
germline stem cell in vitro: offspring from single stem cells. Biology of Reproduction,
v.72, p.236-40.
Kanatsu-Shinohara, M.; Toyokuni, S.; Morimoto, T.; Matsui, S.; Honjo, T.; Shinohara, T.
(2006) Functional assestment of self-renewal actitivity of male germline stem cells
following cytotoxic damage and serial transplantation. Reproduction, v.68, p.1801-1807.
Karashima, T.; Zalatnai, A.; Schally, A.V. (1988) Protective effects of analogs of luteinizing
hormone-releasing hormone against chemotherapy-induced testicular damage in rats.
Proclaims National Academy of Sciences of USA, v.85, p.2329-2333.
Koh, K-B.; Komiyama, M.; Toyama, Y.; Adachi, T.; Mori, C. (2004) Percoll fractionation of
adult mouse spermatogonia improves germ cell transplantation. Asian Journal of
Andrology,v.6,p.63-98.
Kubota H.; Avarbock, M.R.; Brinster R.L. (2003) Spermatogonial stem cell share some, but
not all, phenotypic and functional characteristics with other stem cells. Proceedings of
the National Academy of Sciences of the USA, v.100, p.6487-92.
Lawson, K.A., Pedersen, R.A. (1992) Clonal analysis of cell fate during gastrulation and
early neurulation in the mouse. In: CIBA foundation symposium 165 Post implantation
development in the mouse. John Wileys & Sons: New York, USA, 3-26 p.
Lu, C.C.; Meistrich, M.L. (1979) Cytotoxic effects of chemotherapeutic drugs on mouse testis
cells. Cancer Research, v.39, p.3575-82.
McLean, D.J. (2005) Spermatogonial stem cell transplantation and testicular function. Cell
and Tissue Research, v.322, p.21-31.
McLean, D.J.; Friel, P.J.; Johnston, D.S.; Griswold, M.D. (2003) Characterization of
spermatogonial stem cell maturation and differentiation in neonatal mice. Biology of
Reproduction, v.69, p.2085-91.
Meistrich, M.L. (1982) Quantitative relation between testicular stem cell survival, sperm
production, and fertility in the mouse after treatment with different cytotoxic agents.
Journal of Andrology, v.3, p.58-68.

Referências Bibliográficas 58
Meistrich, M.L. (2009) Male gonadal toxicity. Pediatric Blood Cancer, v.53, p.261-6.
Meistrich, M.L.; Bruce, W.R.; Clermont, Y. (1973) Cellular composition of fractions of mouse
testis cells following velocity sedimentation separation. Experimental Cell Research, v
.79, p.213-27.
Meistrich, M.L.; Finch, M.; da Cunha, M.F.; Hacker, U.; Au, W.W. (1982) Damaging effects of
fourteen chemotherapeutic drugs on mouse testis cells. Cancer Research, v.42, p.122-
131.
Meistrich, M.L.; Finch, M.; Lu, C.C.; de Ruiter-Bootsma, A.L.; de Rooij, D.G. (1984) Strain
differences in the response of mouse testicular stem cells to fractionated radiation.
Radiation Research, v. 97, p. 478-87.
Meistrich, M.L.; Trostle, P.K. (1975) Separation of mouse testis cells by equilibrium density
centrifugation in renografin gradients. Experimental Cell Research, v.92, p.231-44.
Meng, X.; Lindahl, M.; Hyvonen, M.E.; Parvinen, M.; de Rooij, D.G; Hess, M.W. Raatikainen-
Ahokas, A.; Sainio, K.; Rauvala, H.; Lakso, M.; Pichel, J.G.; Westphal, H.; Saarma, M.;
Sariola, H. (2000) Regulation of cell fate decision of undifferentiated spermatogonia by
GDNF. Science, v.287, p.1489-93.
Nakagawa, T.; Nabeshima, Y.; Yoshida, S. (2007) Functional identification of the actual and
potential stem cell compartments in mouse spermatogenesis. Development Cell, v.12,
p.195-206.
Nakagawa, T.; Sharma, M.; Nabeshima, Y.; Braun, R.E.; Yoshida, S. (2010) Functional
hierarchy and reversibility within the murine spermatogenis stem cell compartment.
Science, v.328, p.62-67.
Nagano, M.; Avarbock, M.R.; Brinster, R.L. (1999) Pattern and kinetics of mouse donor
spermatogonial stem cell colonization in recipient testes. Biology of Reproduction, v. 60,
p. 1429-36.
Naughton, C.K.; Jain, S.; Strickland, A.M.; Gupta, A.; Milbrandt, J. (2006) Glial Cell-Line
Derived Neurotrophic Factor-Mediated RET Signaling Regulates Spermatogonial Stem
Cell Fate. Biology of Reproduction, v.74, p.314-21.
Nishimune, Y.; Aizawa, S.; and Komatsu, T. (1978). Testicular germ cell differentiation in
vivo. Fertility and Sterility,v.29, p.95–102.

Referências Bibliográficas 59
Oatley, J.M; Brinster, R.L. (2006) Spermatogonial stem cells. Methods in Enzimology, v.419,
p.259-82.
Ogawa, T.; Dobrinski, I.; Brinster, R.L. (1999) Recipient preparation is critical for
spermatogonial transplantation in rat. Tissue Cell, v. 31, p. 29-34.
Ogawa, T.; Ohmura, M.; Yumura, Y.; Sawada, H.; Kubota, Y. (2003) Expansion of murine
spermatogonial stem cells through serial transplantation. Biology of Reproduction, v.68,
p.316-22.
Orwig, K.E.; Ryu, B.Y.; Master, S.R.; Phillips, B.T.; Mack, M.; Avarbock, M.R.; Chodosh, L.;
Brinster, R.L. (2008) Genes involved in post-transcriptional regulation are
overrepresented in stem/progenitor spermatogonia of cryptorchid mouse testes. Stem
Cells, v.26, p.927-38.
Ohta, H.; Yomogida, K.; Dohmae, K.; Nishimune, Y. (2000) Regulation of proliferation and
differentiation in spermatogonial stem cells: the role of c-kit and its ligand SCF.
Development, v.127, p.2125-31.
Parreira, G. G.; Ogawa, T.; Avarbock, M. R.; Franca, L. R.; Brinster, R. L.; Russell, L. D.
(1998) Development of germ cell transplants in mice. Biology of Reproduction, v.59,
p.1360-70.
Pellegrini, M.; Grimaldi, P.; Rossi, P.; Geremia, R.; Dolci, S. (2003) Developmental
expression of BMP4/ ALK3/ SMAD5 signaling pathway in the mouse testis: A potential
role of BMP4 in spermatogonia differentiation. Journal of Cell Science, v.116, p.3363-
72.
Raverot G.; Weiss, J.; Park, S.Y.; Hurley, L.; Jameson, J.L. (2005) Sox3 expression in
undifferentiated spermatogonia is required for the progression of spermatogenesis.
Developmental Biology, v.283, p.215.
Russell, L.D.; Russell, J.A. (1991) Short-term morphological response of the rat testis to
administration of five chemotherapeutic agents. American Journal of Anatomy, v.192,
p.142-168.
Russell, L.D.; Ettlin, R.A.; Sinha Hikim, A. P.; Clegg, E.D. (1990). Histological and
histopathological evaluation of the testis. 1sted. Cache River Press: Vienna, USA. 280p.

Referências Bibliográficas 60
Shinohara T.; Avarbock, M.R.; Brinter, R.L. (1999) β1- and α6-integrin are surface markers
on mouse spermatogonial stem cells. Proceedings of the National Academy of Sciences
of the USA, v.96, p.5504-9.
Shinohara, T.; Avarbock, M.R.; Brinster, R.L. (2000) Functional analysis of spermatogonial
stem cells in steel and cryptorchid infertile mouse models. Developmental Biology,
v.220, p.401-11.
Shinohara, T.; Orwig, K.E.; Avarbock, M.R.; Brinster, R.L. (2001) Remodeling of the
postnatal mouse testis is accompanied by dramatic changes in stem cell number and
niche accessibility. PNAS, v.98, p.6186-91.
Shinohara, T.; Orwig, K.E.; Avarbock, M.R.; Brinster, R.L. (2002) Germ line stem cell
competition in postnatal mouse testes. Biology of Reproduction, v.66, p.1491-7.
Sprando, R.L. (1990) Perfusion of the rat testis through the heart using heparin. In:
Histological and Histopathological Evaluation of the Testis. Ed. Russell L.D., Ettlin R.A.,
Sinha Hikim A.P., Clegg E.D. Cache River Press: Vienna, USA. 277-80p.
Tummers, M.; Thesleff, I. (2003) Root or crown: A developmental choice orchestrated by the
differential regulation of the epithelial stem cell niche in the tooth of two rodent species.
Development, v.130, p.1049-57.
Udagawa, K.; Ogawa, T.; Watanabe, T.; Yumura, Y.; Takeda, M.; Hosaka, M. (2001) GnRH
analog, leuprorelin acetate, promotes regeneration of rat spermatogenesis after severe
chemical damage. International Journal of Urology, v.8, p.615-22.
Velez de Ia Calle, J.F.; de Queiroz, F.; Gamier, D.H.; Kercret, H.; Folliot, R.; Jegou, B.
(1989) Reproductive effects of the anticancer drug cyclophosphamide in male rats at
different ages. Archives of Androlology, v.22, p.251-263.
Vergouwen, R.P.F.A.; Jacobs, S.G.P.; Huiskamp, R.; Davids, J.A.G.; de Rooij, D.G. (1991)
Proliferative activity of gonocytes, Sertoli cells and interstitial cells during testicular
development in mice. Journal of Reproduction and Fertility, v.93, p.233-243.
Wang, G.; Shao, S.H.; Weng, C.C.; Wei, C.; Meistrich, M.L. (2010) Hormonal suppression
restores fertility in irradiated mice from both endogenous and donor-derived stem
spermatogonia. Toxicology Science, v.117, p.225-237.

Referências Bibliográficas 61
Weber, J. E.; Russell, L. D. (1987) A study of intercellular bridges during spermatogenesis in
the rat. American Journal Anatomy, v.180, p.1-24.
Withers, H.R.; Hunter, N.; Barkley Jr., H.T.; Reid, B.O. (1974) Radiation survival and
regeneration characteristics of spermatogenic stem cells of mouse testis. Radiation
Research, v.57, p.88-103.
Yoshida, S.; Takakura, A.; Ohbo, K.; Abe, K.; Wakabauashi, J.; Uamamoto, M., et al. (2004)
Neurogenin 3 delineates the earliest stages of spermatogenesis in the mouse testis.
Developmental Biology, v.269, p.447-58.
Yoshida, S.; Nabeshima, Y.; Nakagawa, T. (2007) Stem cell heterogeneity: actual and
potential stem cell compartments in mouse spermatogenesis. Annals of the New York
Academy of Sciences, v.1120, p.47-58.
Yoshinaga, K.; Nishikawa, S.; Ogawa, M.; Hayashi, S.; Kunisada, T.; & Fujimoto, T. (1991)
Role of c-kit in mouse spermatogenesis: identification of spermatogonia as a specific
site of c-kit expression and function. Development, v.113, p.689-99.
Zallio, R.C. (2009) Avaliação morfofuncional de espermatogônias de camundongos
mutantes Utp14bJSD
sob condições normais de temperatura e após criptorquidismo
experimental. Dissertação (Mestrado), Belo Horizonte, UFMG – Instituto de Ciências
Biológicas. 43p.
Zhang, Z.; Shao, S.; Meistrich, M.L. (2007) The radiation-induced block in spermatogonial
differentiation is due to damage to the somatic environment, not the germ cells. Journal
of Cell Physiology, v.211, p.149-158.
Zhang, Z.; Shao, S.; Meistrich, M.L. (2006) Irradiated mouse testes efficiently support
spermatogenesis derived from donor germ cells of mice and rats. Journal of Andrology,
v.27, p.365-375.
Zhang, Z.; Shao, S.; Shetty, G.; Meistrich, M. (2009) Donor Sertoli cells transplanted into
irradiated rat testes stimulate partial recovery of endogenous spermatogenesis.
Reproduction, v.137, p.497-508.
.

ANEXO I - TRABALHO PUBLICADO

R
EPRODUCTIONRESEARCHSpermatogonial morphology and kinetics duringtestis development in mice: a high-resolution lightmicroscopy approach
Ana Luiza Drumond, Marvin L Meistrich1 and Helio Chiarini-Garcia
Laboratory of Structural Biology and Reproduction, Department of Morphology, Institute of Biological Sciences,Federal University of Minas Gerais, Avenida Antonio Carlos, 6627 - Pampulha, 31.270-901 Belo Horizonte, MG,Brazil and 1Department of Experimental Radiation Oncology, M.D. Anderson Cancer Center, The University of Texas,Houston, Texas 77030-4095, USA
Correspondence should be addressed to H Chiarini-Garcia; Email: [email protected]
Abstract
Despite the knowledge of spermatogonial biology in adult mice, spermatogonial development in immature animals has not been fully
characterized. Thus, the aim of this study was to evaluate the ontogeny of the morphological development of the spermatogonial lineage
in C57BL/6 mouse testis, using high-resolution light microscopy. Spermatogonial morphology, chronology, and absolute number were
determined for different ages postpartum (pp). The morphology of spermatogonia in immature mice was similar to that of adult
spermatogonia, although their nuclear diameter was slightly smaller. The A1 spermatogonia were first observed on day 2 pp, and only
24 h later, differentiating type A3 and A4 spermatogonia were observed in the seminiferous cords. This result indicated a shortening of
the spermatogonial phase for immature mice of aboutw2.5 days when compared with adult mice and suggests that gonocytes and/or A1
spermatogonia could directly become A4 spermatogonia, skipping the developmental sequence of type A spermatogonia. These A4
spermatogonia are functional as they develop into type B spermatogonia by day 5 pp. At day 8 pp, while differentiation to spermatocytes
begins, the Aund spermatogonia reach their maximal numbers, which are maintained through adulthood. The various details of the
spermatogonial behavior in immature normal mice described in this study can be used as a baseline for further studies under
experimental or pathological conditions.
Reproduction (2011) 142 145–155
Introduction
At birth, the seminiferous cords of mammalian testescontain germ cells called gonocytes that are located inthe center of the seminiferous cords and are arrestedin the G0 phase of the cell cycle. In rodents, after birth,the gonocytes relocate toward the basal membrane(McGuinness & Orth 1992) and resume proliferation,giving rise, after several steps, to the adult spermatogo-nial stem cell (de Rooij & Grootegoed 1998, de Rooij& Russell 2000). The passage from gonocytes tospermatogonial stem cells is not completely understood.Some authors described that the gonocytes generatean intermediate cell type called pre-spermatogonia(Huckins & Clermont 1968), whereas others haveshown that the gonocytes can give rise to either sperma-togonial stem cells or differentiating spermatogonia(Kluin & de Rooij 1981, Yoshida et al. 2006).
In adult mammals, primarily based on the data fromrodents, the spermatogonial stem cell is known as anA single (As) spermatogonium and is capable of
q 2011 Society for Reproduction and Fertility
ISSN 1470–1626 (paper) 1741–7899 (online)
self-renewal or differentiation (de Rooij & Grootegoed1998, de Rooij & Russell 2000). Once it starts its pathtoward differentiation, the As spermatogonia give rise,through mitosis, to two daughter cells that remainconnected by intercellular bridges and are called Apaired (Apr) spermatogonia. These cells divide formingchains of spermatogonia, the A aligned (Aal) spermato-gonia, which can divide two or three times more inmice. The group formed by As, Apr, and Aal is known asundifferentiated type A spermatogonia (Aund). The Aal
spermatogonia will differentiate, with no mitoticdivision, to the differentiating type A1 spermatogonia.The type A1 spermatogonia undergo six successivemitotic divisions to yield A2, A3, A4, intermediate-type(In), and type B spermatogonia. The type B spermatogo-nia will divide giving rise to spermatocytes, starting themeiotic process.
Although previous studies could not distinguish thevarious A spermatogonial subtypes, high-resolution lightmicroscopy was able to differentiate the spermatogonial
DOI: 10.1530/REP-10-0431
Online version via www.reproduction-online.org

146 A L Drumond and others
types in adult mice (Chiarini-Garcia & Russell 2001).The ability to distinguish these spermatogonial subtypesunder light microscopy has made it possible tobetter understand details of spermatogonial biology inrodents under normal (Chiarini-Garcia et al. 2003),experimental (Nascimento et al. 2008), and geneticallypathological (Russell et al. 2002, Bolden-Tiller et al.2007) conditions. However, this new morphologicalapproach has not been used to evaluate the early eventsof spermatogenesis development.
Thus, the goal of this study was to provide a thoroughcharacterization of the morphology and developmentalkinetics of the spermatogonial subtypes during testisdevelopment, using a high-resolution method, in orderto characterize the ontogeny of this process. In additionto addressing specific questions in this process, such asthe transition from gonocytes to spermatogonia, it wasalso our goal to provide a standard that could be used asa baseline for future functional and pathological studies.
Results
The body weights, body lengths (0–14 days), andtesticular weights obtained from mice of different agesare shown in Table 1.
Germ cell emergence and spermatogonial synchrony
The timing of appearance of specific germ cells duringontogeny in the testes is shown in Table 2. Thedifferent types of spermatogonia are defined accordingto the criteria of Chiarini-Garcia & Russell (2001) andare shown in Fig. 1 that are to be also observed inimmature mice. The days of the first emergence of eachgerm cell type, from the gonocytes at birth up toelongated spermatids in adulthood, are presented in asemi-quantitative manner (Table 2). The morphology ofthe gonocytes is shown in Fig. 2 and the different
Table 1 Biometrical values from mice at different ages.
Age Body weight (g)Snout-to-rumpdistance (mm)
Testis weight(mg)
0 1.4G0.1 31G1 0.6G0.11 1.4G0.0 31G1 0.6G0.12 1.9G0.2 34G1 0.9G0.13 2.5G0.0 36G1 1.3G0.14 2.5G0.1 37G0 1.4G0.15 3.6G0.2 40G1 2.0G0.26 3.5G0.1 43G1 2.2G0.18 5.4G0.2 48G1 3.6G0.210 5.5G0.1 50G1 4.4G0.212 8.1G0.2 58G1 8.0G0.814 7.7G0.5 59G1 7.3G0.817 5.0G0.0 K 10G0.320 7.7G0.4 K 14G0.427 17.1G1.3 K 48G2.037 20.2G0.4 K 70G2.870 27.8G0.5 K 104G1.9
The values are expressed as meanGS.E.M.
Reproduction (2011) 142 145–155
subtypes are defined below. At birth and at day 1postpartum (pp), only gonocytes were observed inseminiferous cords (Fig. 2a and b).
On the second day, Aund and A1 spermatogonia werefirst seen, and on the third and fourth days, spermato-gonial types up to A3 and possibly A4 spermatogoniawere already observed. On days 5 and 6, all thespermatogonial types, as such Aund, A1, A2, A3, A4, In,and B spermatogonia were seen, although the mostdifferentiated ones were seen with low frequency (Fig. 2cand d). The meiotic cells were seen on day 8, whenpreleptotene spermatocytes were observed. At this time,it was possible to observe that the synchrony of theseminiferous epithelial stages between spermatogoniaand spermatocytes as defined in the adult was alreadyfound; for example, the type A2 spermatogonia wereassociated with leptotene spermatocytes (Fig. 3a). Basedon the appearance of cells, we calculated that the firstspermatogonial development in immature mice from A1
to A3 and possibly A4 spermatogonia occurred in w1 dayand from A1 to B spermatogonia in 3 days (Table 2).
The appearance of later stages of spermatocytesbeyond leptotene was observed starting on day 10 pp.After formation of the tubular lumen, at 12 days pp(Fig. 3b), pachytene spermatocytes were detected in lownumbers for the first time in the seminiferous tubules andwere frequently observed at 14 days (Fig. 3c) and wereassociated with type A3 and A4 spermatogonia. Meioticfigures were first seen at day 20 (Fig. 3d), when onceagain we could observe the synchrony as they wereassociated with type A3 spermatogonia and earlypachytene spermatocytes, as in adults.
The round spermatids were formed after day 20 andthe first elongated spermatids were observed at day 27(Fig. 3e), making it possible to identify all stages of theseminiferous epithelium by the acrosomal system. Onday 37 pp, the spermatogenic process was completelydeveloped (Fig. 3f).
Gonocyte and spermatogonial morphology
In the seminiferous cords, the gonocytes pass throughseveral morphological and positioning changes beforethey generate the type Aund spermatogonia. We dividedthese cells into three morphologically distinct subtypes,namely I, II, and III, to further study their numbers andkinetics in an attempt to separate them into functionallydifferent cells. All the gonocyte types had large nuclei,10–12 mm, and one to three nucleoli. Whereas thegonocyte subtype I had round nuclei, there was aprogressive change to a slightly oval shape by gonocytetype III. The morphological features that allowed usto separate these three subtypes of gonocytes are asfollows:
Gonocyte subtype I (Fig. 2a and e) containedcondensed round nucleoli and a regular cytoplasmicmembrane with no projections. They were usually
www.reproduction-online.org

Table
2O
ccurr
ence
of
germ
cell
s(G
C)
atsp
ecifi
cst
ages
of
dev
elopm
ent
duri
ng
post
nat
aldev
elopm
ent
inm
ice.
Days
01
23
45
68
10
12
a14
b17
c20
27
d37
70
Gonocy
tes
subty
pe
IC
CC
CC
CC
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
Gonocy
tes
subty
pe
IIC
CC
CC
CC
CK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KG
onocy
tes
subty
pe
III
CC
CC
CC
CC
CC
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
Aundiff
eren
tiat
edsp
erm
atogo
nia
KK
KK
KK
KK
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
C
Sper
mat
ogo
nia
A1
KK
KK
KK
KK
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
A2
KK
KK
KK
KK
KK
KK
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
A3
KK
KK
KK
KK
KK
KK
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
A4
KK
KK
KK
KK
KK
KK
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
InK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CB
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CPre
lepto
tene
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CLe
pto
tene
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
Zyg
ote
ne
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
Earl
ypac
hyt
ene
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
CIn
term
edia
tepac
hyt
ene
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
CC
CC
CC
CC
CC
CC
CC
CC
CC
CC
Late
pac
hyt
ene
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
CC
CC
CC
CC
CC
CC
CC
CC
CD
iplo
tene
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
CC
CC
CC
CC
CC
CSp
erm
atocy
tes
IIK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KC
CC
CC
CC
CC
CR
ound
sper
mat
ids
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
CC
CC
CC
CC
CEl
onga
ted
sper
mat
ids
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
KK
CC
CC
CC
C
KK
KK
,La
ckof
GC
;C
,fe
wG
C;C
C,in
term
edia
team
ount
of
GC
;C
CC
,hig
ham
ount
of
GC
.Th
enum
ber
of
germ
cell
sw
ascl
assi
fied
com
par
edw
ith
that
inth
esu
bja
cent
ages
;C
CC
,in
dic
ates
that
num
ber
of
germ
cell
sw
asnot
dis
tingu
ishab
lefr
om
that
ina
norm
alad
ult
mouse
.aThe
most
adva
nce
dpac
hyt
enes
her
ear
eas
soci
ated
wit
hpre
lepto
tenes
and
Bsp
erm
atogo
nia
.bTh
em
ost
adva
nce
dpac
hyt
enes
her
ear
eas
soci
ated
wit
hsp
erm
atogo
nia
A2.
cThe
most
adva
nce
dpac
hyt
enes
her
ear
eas
soci
ated
wit
hzy
gote
nes
.dEl
onga
ted
sper
mat
ids
inst
ep16.
Spermatogonial morphology during ontogeny 147
www.reproduction-online.org
located in the center of the seminiferous cords butalways separated from the basement membrane (Fig. 2a).
The nucleoli of gonocytes subtype II (Fig. 2f) were lesscondensed with a reticulated appearance more of anirregular shape. Although these cells were located inthe center of the cords, they had cytoplasmic projec-tions extending toward the basal membrane. Thesecells were frequently seen with a peculiar structureresembling acrosomal vesicles, located close to thenuclear envelope.
The nucleoli of gonocytes subtype III (Fig. 2b and g)were even less condensed and reticulated and hadan irregular shape and were usually close to thenuclear envelope; spots of heterochromatin wereobserved, associated with the nuclear envelope;cytoplasmic membrane was irregular, but with no pro-jections. These cells were seen lying on the basalmembrane (Fig. 2b).
In contrast to the gonocytes, the Aund spermatogoniahad a smaller nuclear diameter and a more oval shape,had more finely granular and darker euchromatin butlacked the spots of heterochromatin associated to thenuclear envelope of gonocytes, and the nucleoli weresomewhat more condensed but more irregular. Thedifferentiating type A spermatogonia were distinguishedfrom the Aund by their nuclear morphology (Fig. 1) asdescribed in detail previously (Chiarini-Garcia & Russell2001), becoming larger, more lightly staining, and withincreasing heterochromatin associated with the nuclearmembrane. The A2 to A4 spermatogonia were distin-guished from type III gonocytes by broader contact withthe basement membrane, a more oval nuclear shape,and more irregular nucleoli.
The only noticeable differences in a given spermato-gonial type, between postnatal (6 days) and adult(70 days) mice, were the nuclear diameters (seebelow). A morphologically peculiar type of A sperma-togonia was observed in young animals (Fig. 1c). Thesecells had many of the morphological characteristics ofdifferentiating type A2 and A3 spermatogonia, having anoval to round nucleus, with spots of heterochromatinaround the nuclear membrane and a large, reticulatednucleolus. However, they have nuclear vacuoles,described in the literature as being located only innuclei of type A undifferentiated cells (Chiarini-Garcia &Russell 2001). Such nuclear vacuoles in differentiatingspermatogonia were observed predominantly from 3 to6 days pp, being less common in the animals that were 8and 10 days old and were not observed in sexuallymature mice.
Although this study using an accurate histologicaltechnique allowed the precise identification of almost allgerm cell types during ontogenic development, the leastdifferentiated spermatogonial type, the Aund spermato-gonia, could not be more finely sub-classified to suggestwhich of these cells might be stem spermatogonia.
Reproduction (2011) 142 145–155

S
(a) (b) (c)
(d) (e) (f)
(g) (h) (i)
(a) (b) (c)
(d) (e) (f)
(g) (h) (i)
S SS
S
S
A2A1
SS
S
S
S
SS
SS
InA4
S S
B
S
Aund Aund Adiff
A3
S SS
Figure 1 Spermatogonial lineage from mice at ages2 to 6 days postpartum. (a and b) Type Aundifferentiated spermatogonia, one of them witha nuclear vacuole (arrow); (c–i) differentiatingspermatogonia, (c) with a nuclear vacuole (arrow),(d–g) A1 to A4 spermatogonia, (h) Intermediate –In, and (i) B spermatogonia. S, immature Sertolicell and arrowhead, intercellular bridge. Barsrepresent 10 mm.
148 A L Drumond and others
Spermatogonial nuclear diameter
Figure 4 shows the average diameters of germ cell nucleifrom the Aund to type B spermatogonia from 2 up to70 days pp. It was observed that diameters varied duringthe germ cell and testes development. In all ages, theAund spermatogonia have a smaller nuclear diameterthan the differentiating type A spermatogonia. The typeA2 spermatogonia were found to be the cells with thelargest nuclear diameter, whereas the spermatogoniatype B were the smallest (Fig. 4). The nuclear diameter ofeach cell type from the Aund to In spermatogonia seemedto be larger in younger animals and decreased in sizeduring the development, reaching their smallest nucleardiameters by 37 days pp.
Gonocyte and spermatogonial numbers
The changes in the total numbers of each spermatogonialtype per testis during testicular development wereanalyzed by stereological methods. The number ofgonocytes decreased progressively from birth to 3 dayspp, whereas the numbers of differentiating type A, In,and B spermatogonia increased progressively up to37 days, at puberty (Fig. 5A and B). The number of Aund
spermatogonia at 8 days pp was 190 000 per testis,similar to the number of type Aund spermatogonia at allsubsequent ages (Fig. 5B), including the adult animals. Inaddition, the number of all differentiating spermatogoniareached the numbers found in the adult between 14 and37 days pp (Fig. 5B).
Gonocyte and spermatogonial kinetics
The total number of gonocytes remained constant duringthe first 24 h after birth (Fig. 6). When the number of
Reproduction (2011) 142 145–155
gonocytes of subtypes I and II went down, the number ofgonocytes of subtype III increased, indicating that theformer subtypes may differentiate into the latter subtype.However, it was observed that the number of gonocytesstarted to decline on day 2 pp coincident with theappearance of the first Aund spermatogonia that start tobe seen in the seminiferous cords. The decline in thetotal numbers of gonocytes on day 2 could possiblyaccount for the increase in Aund spermatogonia at thattime by direct transformation of gonocytes into Aund
spermatogonia. However, the further increase in Aund
spermatogonia on day 3 is too large to be accounted formerely by the transformation of gonocytes and mustinclude cell division.
Gonocyte and spermatogonial mitosis and apoptosis
The mitotic index of germ cells (gonocytes plusspermatogonia) showed an increase from birth to3 days pp (Fig. 7), remained high until the fifth day,and then started to decrease, reaching the adult level at10 days pp. The apoptotic index significantly increasedafter birth up to 2 days pp (Fig. 7); at this time only,gonocytes were found in the seminiferous cords,pinpointing this developmental age as the time of ahigh number of gonocyte deaths. The apoptotic indexthen decreased, and already at 3 days pp, it reached anumber as low as adult animals, remaining at this levelthroughout testis development.
Discussion
In this study, we have extended high-resolution lightmicroscopy techniques to quantify the numbers ofdifferent types of gonocytes and spermatogonia cells
www.reproduction-online.org
Day 0
Mi GonI
S
S
S
Mi
S
BV
Aund
S
S
A3
A2
S
S
2 Days
4 Days 6 Days
GonI GonII GonIII
S
GonIII
SMi
(a) (b)
(e) (g)(f)
(c) (d)
(a) (b)
(e) (g)(f)
(c) (d)
Figure 2 Seminiferous cords at day 0 (a), 2 days (b),4 days (c), and 6 days (d) postpartum. Note thedifferent morphological subtypes of gonocytes:gonocyte subtype I (e), gonocyte subtype II (f) witha structure close to the nuclear membrane (arrowhead), and subtype III (g). Structures identified inthe seminiferous cords and interstitium are labeledas follows: Sertoli cell (S), Sertoli cells in prophase(arrow), gonocyte subtype I (GonI), subtype II(GonII) and subtype III (GonIII), type A undiffer-entiated spermatogonia (Aund), type A2 and type A3
differentiating spermatogonia, myoid cells (Mi),and blood vessels (BV). Bars represent 10 mm.
Spermatogonial morphology during ontogeny 149
during postnatal development in the mouse. Theimplications of these observations for understandingthe development of spermatogonia from gonocytes,the proliferation, differentiation, and kinetics of sper-matogonial development and the formation of associ-ations of the cycle of the seminiferous epithelium isdiscussed below.
Although we have observed different subclasses ofgonocytes, which could be recognized by theirmorphology, number, and position in the seminiferouscords as gonocytes I, II, or III, the total number ofgonocytes did not increase after birth. The mitoticindex data showed that the gonocytes proliferateimmediately after birth but their number did not changedue to a balance between mitosis and apoptosis, whichwas also increased during this period. This is consistentwith previous observations showing that during thefirst days after birth, some of the gonocytes go throughmitosis (Kluin & de Rooij 1981), but a considerablenumber of gonocytes degenerate (Roosen-Runge &Leik 1968).
Kluin & de Rooij (1981) also reported that the daughtercells of the gonocyte divisions on day 1 consist of twocell types, the typical gonocytes, called type I, and cellswith larger nuclei, called type II cells. Huckins &Clermont (1968) showed that in rats, gonocytes divide
www.reproduction-online.org
to form cells larger and lighter than the Aund spermato-gonia normally found in adults and named these cellspre-A spermatogonia. Both descriptions of the gono-cytes’ daughter cells agree with our description for thesubtype III gonocytes. As the method applied in ourpaper allowed better cytological distinction of the germcells than the methods used previously, we coulddistinguish three gonocyte subtypes (I, II, and III) insteadof two types (I and II). Kluin & de Rooij (1981) considerthe type II gonocytes to be similar to newly formed A2
spermatogonia in adults. Our data suggest that the cellsfound to be type II by those authors were recognizedhere as subtype II and III, because the number of thesecells was 67% of the total gonocytes that are very closeto their percentage of type II cells (70%). Although weagree that these gonocytes are similar to type A2
spermatogonia, we could still morphologically recog-nize the spermatogonial subtypes in young mice, and itwas possible to show that the gonocytes II and III aremorphologically different from type A2 spermatogonia.
The emergence of type Aund and A1 spermatogonia, atday 2, morphologically similar to the those observed inadult mice, is consistent with the results of Yoshida et al.(2006), who demonstrated at 2 days pp the presence ofthe first germ cells that expressed KIT (c-Kit) receptor inthe seminiferous cords. Furthermore, they showed that
Reproduction (2011) 142 145–155
(a)
8 days
14 days
27 days 37 days
12 days
20 days
S
Lu
Pl
S
B
S
L
Lu
S
Lu
E
SP
S
PP P
R E
R
S
E
S
R
P
A4
P P
S II Sp
Me
S
S
DS
A3
A4
Mi
(a)
(c)(c)
(b)(b)
(d)(d)
(e)(e) (f)(f)
A2
Aund
Figure 3 Seminiferous epithelium at ages of 8 days(a), 12 days (b), 14 days (c), 20 days (d), 27 days (e),and 37 days (f) postpartum. Structures indicatedare Sertoli cells (S), type A undifferentiated (Aund),type A2 differentiating spermatogonia (A2), type A3
differentiating spermatogonia (A3), type A4 differ-entiating spermatogonia (A4), type B differentiatingspermatogonia (B), pre-leptotene (Pl), leptotene(L), pachytene (P), diplotene (D), secondaryspermatocyte (II Sp), round spermatid (R),elongated spermatid (E), and lumen (Lu). Barsrepresent 10 mm.
150 A L Drumond and others
these cells must correspond to a population of germ cellsthat skip at least one of the molecular steps ofspermatogonial differentiation that occurs in adults,because the KIT-positive cells that were rapidly formeddid not express neurogenin 3 (NGN3), which isexpressed in type A undifferentiated spermatogonia ofthe adult mice (Yoshida et al. 2006). This result indicatesthat the first type A differentiating spermatogonia (A1
and/or A2 spermatogonia) are part of a cell populationthat is directly derived from the gonocytes separatelyfrom the Aund spermatogonia. The later development ofdifferentiating spermatogonia, which have gone througha stage of NGN3 expression, must come from a subset ofthese Aund cells, which correspond both morpho-logically and functionally to the adult spermatogonialstem cells.
In this study, the first type A1 spermatogonia were seenin the seminiferous cords at 2 days pp, and surprisinglyonly 24 h later, spermatogonia morphologically similarto type A3 and possibly some A4 spermatogonia werealso observed. In adult mice, three mitotic divisions arerequired for a differentiating spermatogonia type A1 tobecome type A4 spermatogonia, and this processrequires a time interval of about 3.5 days (Clermont &
Reproduction (2011) 142 145–155
Trott 1969, de Rooij & Russell 2000). This short timesuggests that gonocytes and/or A1 spermatogonia aredirectly becoming A3 and/or A4 spermatogonia.Although it might be argued that the A3 and A4
spermatogonia produced on days 3 and 4 are notfunctionally the same as those in the adult, they do goon to produce B spermatogonia on day 5, preleptoteneson day 8, zygotene spermatocytes on day 10, andpachytene on day 12; thus, they are functionally actingas differentiated type A spermatogonia. The data inTable 2 can also be used to calculate that, in immaturemice, the formation of B spermatogonia (day 5 pp) fromA1 spermatogonia (day 2 pp) occurs in 3 days, whereasin adults this transition requires 6.1 days, showing thatthe process was shortened w3 days, which appears tohave largely been due to the acceleration of type Aspermatogonial development between days 2 and 3.Hence, our results indicate that there is accelerationof the spermatogonial phase, involving skippinggenerations of the developmental sequence of thetype A spermatogonia that are normally observed inadults. It should be noted that in this study, the stepsinvolved in this acceleration were only morphologicallydefined. The combination of immunohistochemistry
www.reproduction-online.org
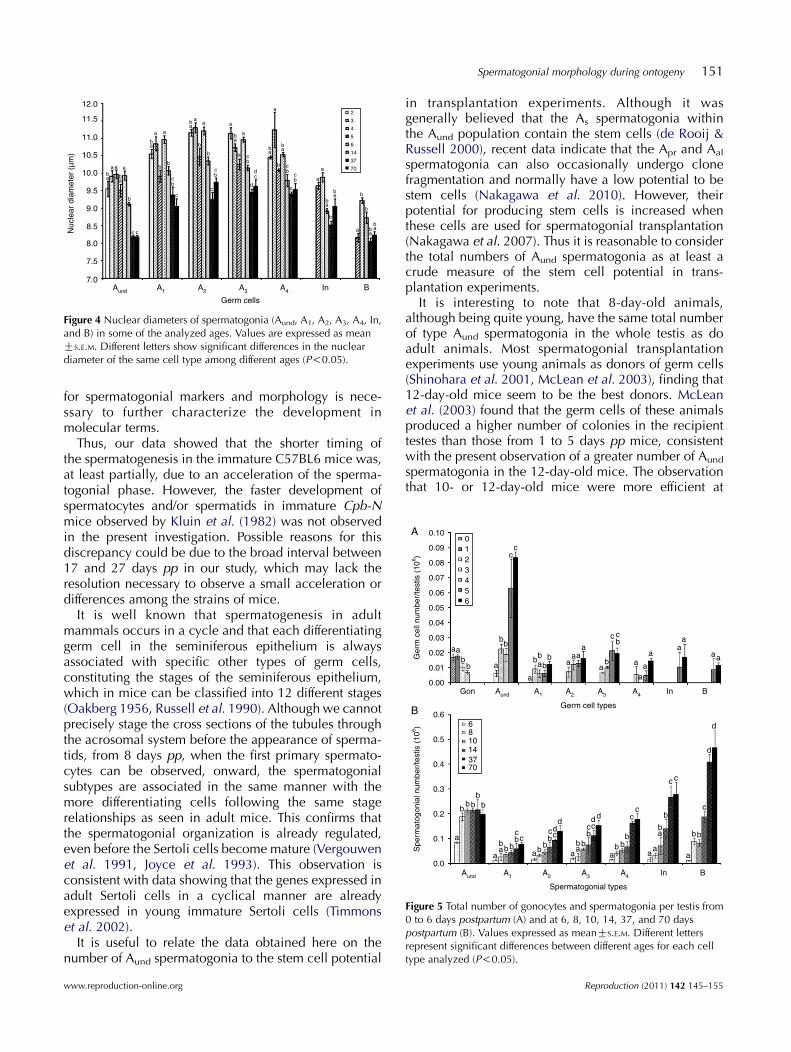
12.0
11.5
11.0
10.5
ab
a
a a
aa
a
a
b
b
d
b
c
b
ab
ab
ab
b
a
b
a
c
ab
ab
ab
ab
b
b
aab
a2
3
4
5
6
14
37
70bb
bc
bc
bc
bc
bcc
d
c
ab
b
c c
a a a10.0
Nuc
lear
dia
met
er (
µm)
9.5
9.0
8.5
8.0
7.5
7.0Aund A1 A2 A3
Germ cells
A4 In B
Figure 4 Nuclear diameters of spermatogonia (Aund, A1, A2, A3, A4, In,and B) in some of the analyzed ages. Values are expressed as meanGS.E.M. Different letters show significant differences in the nucleardiameter of the same cell type among different ages (P!0.05).
0.10
0.09
0.08
0.07
Ger
m c
ell n
umbe
r/te
stis
(10
6 )S
perm
atog
onia
l num
ber/
test
is (
106 )
0.06
0.05
0.04
0.03
0.02
0.01
0.00
0.66810143770
0.5
0.4
0.3
0.2
0.1
0.0Aund
a
bbb
bb
a ab bbccd
d
a ab b
b
c
aa
ab
b
c c
a
bb
c
d
d
c
abb
bccdd
ab
bbbc
c
A1 A2 A3
Spermatogonial types
Germ cell types
0A
B
123456
A4 In B
Gon Aund A1 A2 A3 A4 In B
aa
aa
a
aa
a
cc
ba
aaa
ab
babb
a
bb
bb
aa
a
b
cc
Figure 5 Total number of gonocytes and spermatogonia per testis from0 to 6 days postpartum (A) and at 6, 8, 10, 14, 37, and 70 dayspostpartum (B). Values expressed as meanGS.E.M. Different lettersrepresent significant differences between different ages for each celltype analyzed (P!0.05).
Spermatogonial morphology during ontogeny 151
for spermatogonial markers and morphology is nece-ssary to further characterize the development inmolecular terms.
Thus, our data showed that the shorter timing ofthe spermatogenesis in the immature C57BL6 mice was,at least partially, due to an acceleration of the sperma-togonial phase. However, the faster development ofspermatocytes and/or spermatids in immature Cpb-Nmice observed by Kluin et al. (1982) was not observedin the present investigation. Possible reasons for thisdiscrepancy could be due to the broad interval between17 and 27 days pp in our study, which may lack theresolution necessary to observe a small acceleration ordifferences among the strains of mice.
It is well known that spermatogenesis in adultmammals occurs in a cycle and that each differentiatinggerm cell in the seminiferous epithelium is alwaysassociated with specific other types of germ cells,constituting the stages of the seminiferous epithelium,which in mice can be classified into 12 different stages(Oakberg 1956, Russell et al. 1990). Although we cannotprecisely stage the cross sections of the tubules throughthe acrosomal system before the appearance of sperma-tids, from 8 days pp, when the first primary spermato-cytes can be observed, onward, the spermatogonialsubtypes are associated in the same manner with themore differentiating cells following the same stagerelationships as seen in adult mice. This confirms thatthe spermatogonial organization is already regulated,even before the Sertoli cells become mature (Vergouwenet al. 1991, Joyce et al. 1993). This observation isconsistent with data showing that the genes expressed inadult Sertoli cells in a cyclical manner are alreadyexpressed in young immature Sertoli cells (Timmonset al. 2002).
It is useful to relate the data obtained here on thenumber of Aund spermatogonia to the stem cell potential
www.reproduction-online.org
in transplantation experiments. Although it wasgenerally believed that the As spermatogonia withinthe Aund population contain the stem cells (de Rooij &Russell 2000), recent data indicate that the Apr and Aal
spermatogonia can also occasionally undergo clonefragmentation and normally have a low potential to bestem cells (Nakagawa et al. 2010). However, theirpotential for producing stem cells is increased whenthese cells are used for spermatogonial transplantation(Nakagawa et al. 2007). Thus it is reasonable to considerthe total numbers of Aund spermatogonia as at least acrude measure of the stem cell potential in trans-plantation experiments.
It is interesting to note that 8-day-old animals,although being quite young, have the same total numberof type Aund spermatogonia in the whole testis as doadult animals. Most spermatogonial transplantationexperiments use young animals as donors of germ cells(Shinohara et al. 2001, McLean et al. 2003), finding that12-day-old mice seem to be the best donors. McLeanet al. (2003) found that the germ cells of these animalsproduced a higher number of colonies in the recipienttestes than those from 1 to 5 days pp mice, consistentwith the present observation of a greater number of Aund
spermatogonia in the 12-day-old mice. The observationthat 10- or 12-day-old mice were more efficient at
Reproduction (2011) 142 145–155
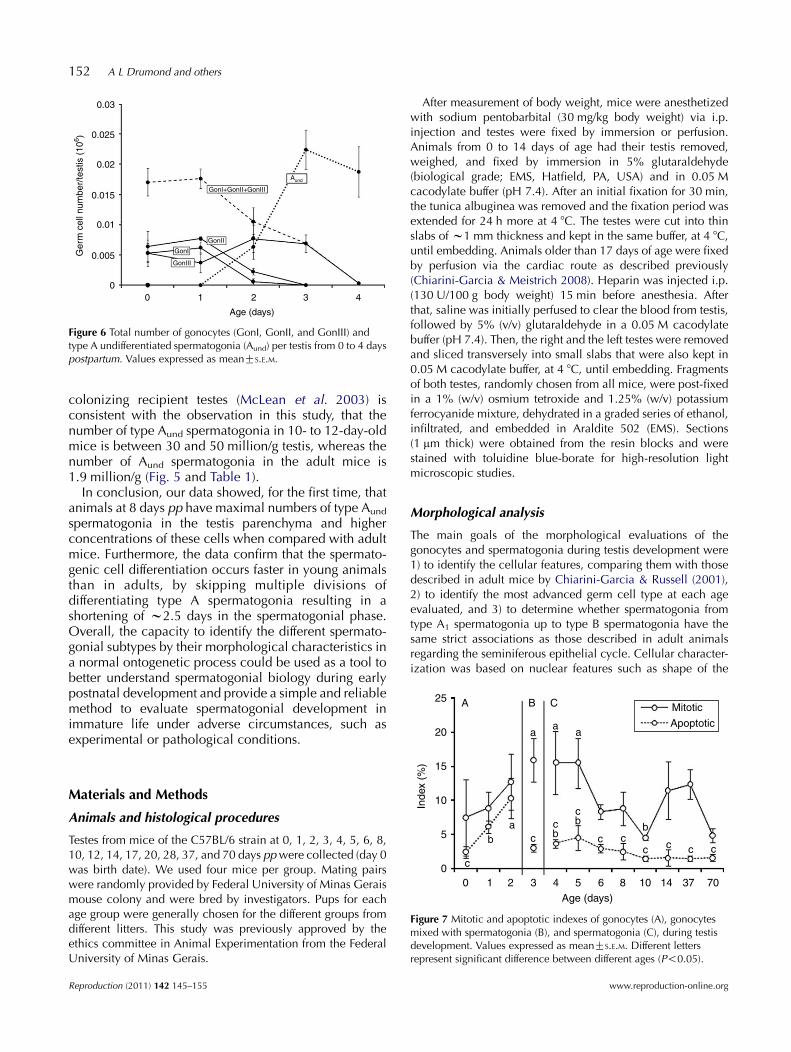
0.03
0.025
0.02
0.015
0.01
0.005Ger
m c
ell n
umbe
r/te
stis
(10
6 )
00
GonI
GonIII
GonII
GonI+GonII+GonIII
Aund
1 2
Age (days)
3 4
Figure 6 Total number of gonocytes (GonI, GonII, and GonIII) andtype A undifferentiated spermatogonia (Aund) per testis from 0 to 4 dayspostpartum. Values expressed as meanGS.E.M.
25 A B C
20 aa
Mitotic
Apoptotica
152 A L Drumond and others
colonizing recipient testes (McLean et al. 2003) isconsistent with the observation in this study, that thenumber of type Aund spermatogonia in 10- to 12-day-oldmice is between 30 and 50 million/g testis, whereas thenumber of Aund spermatogonia in the adult mice is1.9 million/g (Fig. 5 and Table 1).
In conclusion, our data showed, for the first time, thatanimals at 8 days pp have maximal numbers of type Aund
spermatogonia in the testis parenchyma and higherconcentrations of these cells when compared with adultmice. Furthermore, the data confirm that the spermato-genic cell differentiation occurs faster in young animalsthan in adults, by skipping multiple divisions ofdifferentiating type A spermatogonia resulting in ashortening of w2.5 days in the spermatogonial phase.Overall, the capacity to identify the different spermato-gonial subtypes by their morphological characteristics ina normal ontogenetic process could be used as a tool tobetter understand spermatogonial biology during earlypostnatal development and provide a simple and reliablemethod to evaluate spermatogonial development inimmature life under adverse circumstances, such asexperimental or pathological conditions.
15
Inde
x (%
)
10
5
00
c
ba
c bc b
c
c cc
b
c c c
1 2 3 4 5
Age (days)
6 8 10 14 37 70
Figure 7 Mitotic and apoptotic indexes of gonocytes (A), gonocytesmixed with spermatogonia (B), and spermatogonia (C), during testisdevelopment. Values expressed as meanGS.E.M. Different lettersrepresent significant difference between different ages (P!0.05).
Materials and Methods
Animals and histological procedures
Testes from mice of the C57BL/6 strain at 0, 1, 2, 3, 4, 5, 6, 8,10, 12, 14, 17, 20, 28, 37, and 70 days pp were collected (day 0was birth date). We used four mice per group. Mating pairswere randomly provided by Federal University of Minas Geraismouse colony and were bred by investigators. Pups for eachage group were generally chosen for the different groups fromdifferent litters. This study was previously approved by theethics committee in Animal Experimentation from the FederalUniversity of Minas Gerais.
Reproduction (2011) 142 145–155
After measurement of body weight, mice were anesthetizedwith sodium pentobarbital (30 mg/kg body weight) via i.p.injection and testes were fixed by immersion or perfusion.Animals from 0 to 14 days of age had their testis removed,weighed, and fixed by immersion in 5% glutaraldehyde(biological grade; EMS, Hatfield, PA, USA) and in 0.05 Mcacodylate buffer (pH 7.4). After an initial fixation for 30 min,the tunica albuginea was removed and the fixation period wasextended for 24 h more at 4 8C. The testes were cut into thinslabs of w1 mm thickness and kept in the same buffer, at 4 8C,until embedding. Animals older than 17 days of age were fixedby perfusion via the cardiac route as described previously(Chiarini-Garcia & Meistrich 2008). Heparin was injected i.p.(130 U/100 g body weight) 15 min before anesthesia. Afterthat, saline was initially perfused to clear the blood from testis,followed by 5% (v/v) glutaraldehyde in a 0.05 M cacodylatebuffer (pH 7.4). Then, the right and the left testes were removedand sliced transversely into small slabs that were also kept in0.05 M cacodylate buffer, at 4 8C, until embedding. Fragmentsof both testes, randomly chosen from all mice, were post-fixedin a 1% (w/v) osmium tetroxide and 1.25% (w/v) potassiumferrocyanide mixture, dehydrated in a graded series of ethanol,infiltrated, and embedded in Araldite 502 (EMS). Sections(1 mm thick) were obtained from the resin blocks and werestained with toluidine blue-borate for high-resolution lightmicroscopic studies.
Morphological analysis
The main goals of the morphological evaluations of thegonocytes and spermatogonia during testis development were1) to identify the cellular features, comparing them with thosedescribed in adult mice by Chiarini-Garcia & Russell (2001),2) to identify the most advanced germ cell type at each ageevaluated, and 3) to determine whether spermatogonia fromtype A1 spermatogonia up to type B spermatogonia have thesame strict associations as those described in adult animalsregarding the seminiferous epithelial cycle. Cellular character-ization was based on nuclear features such as shape of the
www.reproduction-online.org

Spermatogonial morphology during ontogeny 153
nucleus, presence, and arrangement of heterochromatin,granularity of euchromatin, and morphology and degree ofnucleolar compaction.
GC GC
SC
(a) (b)
(c) (d)
(e) (f)
(a) (b)
(c) (d)
(e) (f)
SC
(g)(g)
Figure 8 Morphological features of mitotic phases of germ and Sertolicells in neonatal testes: panels (a) and (c) represent prophase, andpanels (b) and (d) represent metaphase. GC, germ cells (a and b) andSC, Sertoli cells (c and d). Morphological features of apoptotic germcells: panel (e) represents a seminiferous cord at 3 days pp, withan apoptotic gonocyte in the center (arrow), panel (f) represents anapoptotic spermatogonium (arrow), located close to the basalmembrane, and panel (g) represents apoptotic spermatocyte (arrow).Bars represent 5 mm.
Morphometrical procedures
Morphometrical studies were performed to obtain the absolutenumber of different subtypes of gonocytes and differentspermatogonial types (Aund, A1, A2, A3, A4, In, B) per testisfor all ages studied. For this purpose, the following parameterswere obtained:
volume of testicular parenchyma – VT (mm3)volume density of seminiferous epithelium – Vve
absolute volume of seminiferous epithelium – Vte (mm3)nuclear volume density of each spermatogonial subtype – Vvn
absolute total nuclear volume of each spermatogonial subtype– Vtn (mm3)nuclear volume of each spermatogonial subtype – Vn (mm3)cellular number/testis – CN (106).
The weight of testicular parenchyma is the testis weightwithout the tunica albuginea weight. As the testicular densityis w1 g/ml (1.03–1.04; Sinha-Hikim et al. 1988), each gramof testis corresponds to a volume of 1012 mm3. Hence, VT
is obtained multiplying the testicular parenchyma weight (g)by 1012 mm3.
The volume density of the seminiferous epithelium wasestimated using the point counting method (Sinha-Hikim et al.1988, Russell et al. 2002). These data were obtained throughan Olympus BX-41 light microscope with a 40! objectiveand using a square lattice containing 441 intersections in a10! eyepiece. We counted w4410 intersections on theseminiferous epithelium and the rest of testicular parenchyma,distributed in ten randomly selected fields per animal (two tothree fields per histological section), for each age studied. Thevolume density of seminiferous epithelium (Vve) was obtaineddividing the sum of the points falling on seminiferousepithelium by the total number of points over the tissue. Thevolume of seminiferous epithelium (Vte) was estimatedas follows:
Vte ZVT!Vve:
The nuclear volume density of gonocytes and eachspermatogonial subtype was also estimated using pointcounting, applying the same 441 square lattice with a100! objective. In 40 randomly selected fields for eachanimal (four to five fields per histological section), theintersections over the nuclei of each cell type were recorded.The nuclear volume density of each spermatogonial subtype(Vvn) was obtained by dividing the sum of the points falling oneach cell subtype by the total points over the seminiferousepithelium. The total volume of each germ cell subtype (Vtn)was estimated as follows:
Vtn ZVte!Vvn:
To calculate the nuclear spermatogonial volumes (Vn), it isnecessary to first determine the nuclear diameter (D). Thediameter of ten nuclei of each germ cell subtype was measuredfor each animal, using a ruler fitted in a 10! eyepiece,
www.reproduction-online.org
calibrated with a micrometer ruler. For each spermatogonialcell type, the nuclei that appeared to have the largest profileswere selected to be measured, because that indicates thatthe sections were close to the center of the nucleus. The volumewas calculated as:
VnZpD3
6:
After the determination of the total volume of each germ cellsubtype (Vtn) and the spermatogonial nuclear volumes (Vn),it was possible to calculate the absolute cell number (CN),in millions (106), for each spermatogonial type per testisas follows:
CN ZVtnVn
:
Mitotic and apoptotic indexes of gonocytesand spermatogonia
To calculate the mitotic and apoptotic indexes, the followingcomponents of the seminiferous epithelium were counted:a) the number of gonocytes and spermatogonia, b) the numberof mitoses, and c) the number of apoptotic gonocytes andspermatogonia. Approximately 30 tubular cross sections were
Reproduction (2011) 142 145–155

154 A L Drumond and others
counted per animal. The mitotic and apoptotic indexes werecalculated by dividing the number of mitoses and/or apoptoticcells, respectively, by the sum of gonocytes, spermatogonia,mitoses, and apoptosis.
Specific criteria were needed to distinguish the mitoticspermatogonia from mitotic Sertoli cells. In prophase, the twocell types were clearly distinguished, as they still have thenuclear characteristics of the cells in interphase (Fig. 8a and c),whereas in other mitotic phases, the mitotic spermatogoniawere distinguished from mitotic Sertoli cells by their cyto-plasmic features. The mitotic figures of germ cells wereidentified by the outline of their cytoplasmic membrane,separating their cytoplasm from the Sertoli cells’ cytoplasm,and by the density of their cytoplasm, which was brighter thanthe Sertoli cells (Fig. 8b). The Sertoli cells in mitosis usuallydisplayed no cytoplasmic membrane delimitation, or anelongated outline, with processes toward the center of thecords/tubules, and their cytoplasmic density was morehomogenous with the cytoplasm around it (Fig. 8d).
The identities of the cells that produced the apoptotic bodieswere morphologically determined according to the borderblebbing and fragmentation of the chromatin (Fig. 8e, f, and g).When the cytoplasm is observed, the following featuresare considered: a) its shape (elongated in spermatogonia andround in spermatocytes), b) whether it is contacting basallamina (spermatogonia are in contact with the basal lamina)and c) how it is contacting (flattened in spermatogonia orpunctuated in spermatocytes). The nuclei have particularcharacteristics that enable their assignment to different celltypes such as a) shape, b) size, c) position in the cytoplasm,d) granularity of chromosome fragmentation, finely granular inspermatogonia (Fig. 8f) and coarsely granular in spermatocytes(Fig. 8g), e) number of joined apoptosis (normally spermatogoniaare alone), and f) the stage of the epithelium cycle where thespermatocytes are larger and usually far from the basal lamina.
Statistical analysis
The morphometrical data were expressed as meanGS.E.M.Groups were compared using ANOVA, followed by Fisher’sLSD and Turkey’s tests when necessary. The results wereconsidered to be statistically significant if P!0.05. All analyseswere performed using SPSS 15.0 software (SPSS, Inc., Chicago,IL, USA).
Declaration of interest
The authors declare that there is no conflict of interest thatcould be perceived as prejudicing the impartiality of theresearch reported.
Funding
This study was supported in part by the following BrazilianFoundations: Coordenacao de Aperfeicoamento de Pessoal deNıvel Superior (CAPES), Fundacao de Amparo a Pesquisa doEstado de Minas Gerais (FAPEMIG), and Conselho Nacional dePesquisa (CNPq).
Reproduction (2011) 142 145–155
Acknowledgements
We thank Maria Luiza da Silva and Adriano Moreira Ferreira forassistance in histological preparation.
References
Bolden-Tiller OU, Chiarini-Garcia H, Poirier C, Alves-Freitas D, Weng CC,Shetty G & Meistrich ML 2007 Genetic factors contributing to defectivespermatogonial differentiation in juvenile spermatogonial depletion(Utp14b/jsd) mice. Biology of Reproduction 77 237–246. (doi:10.1095/biolreprod.107.060087)
Chiarini-Garcia H & Meistrich ML 2008 High resolution light microscopiccharacterization of spermatogonia. Methods in Molecular Biology 45095–107. (doi:10.1095/biolreprod65.4.1170)
Chiarini-Garcia H & Russell LD 2001 High resolution light microscopiccharacterization of mouse spermatogonia. Biology of Reproduction 651170–1178. (doi:10.1095/biolreprod65.4.1170)
Chiarini-Garcia H, Raymer AM & Russell LD 2003 Non-randomdistribution of spermatogonia in rats: evidence of niches in the semi-niferous tubules. Reproduction 126 669–680. (doi:10.1530/rep.0.1260669)
Clermont Y & Trott M 1969 Duration of the cycle of the semini-ferous epithelium in the mouse and hamster determined by meansof 3H-thimidine and radioautography. Fertility and Sterility 20 805–817.
Huckins C & Clermont Y 1968 Evolution of gonocytes in the rat testis duringlate embryonic and post-natal life. Archives d’Anatomie, d’Histologie etd’Embryologie 51 343–354.
Joyce KL, Porcelli J & Cooke P 1993 Neonatal goitrogen treatment increasesadult testis size and sperm production in the mouse. Journal of Andrology14 448–455.
Kluin PM & de Rooij DG 1981 A comparison between the morphology andcell kinetics of gonocyte and adult type undifferentiated spermatogoniain the mouse. International Journal of Andrology 4 475–493. (doi:10.1111/j.1365-2605.1981.tb00732.x)
Kluin PM, Kramer MF & de Rooij DG 1982 Spermatogenesis in theimmature mouse proceeds faster than in the adult. InternationalJournal of Andrology 5 282–294. (doi:10.1111/j.1365-2605.1982.tb00257.x)
McGuinness MP & Orth JM 1992 Reinitiation of gonocytes mitosis andmovement of gonocytes to the basement membrane in testes of newbornrats in vivo and in vitro. Anatomical Records 233 527–537. (doi:10.1002/ar.1092330406)
McLean DJ, Friel PJ, Johnston DS & Griswold MD 2003 Characterization ofspermatogonial stem cell maturation and differentiation in neonatalmice. Biology of Reproduction 69 2085–2091. (doi:10.1095/biolreprod.103.017020)
Nakagawa T, Nabeshima Y & Yoshida S 2007 Functional identificationof the actual and potential stem cell compartments in mouse spermato-genesis. Developmental Cell 12 195–206. (doi:10.1016/j.devcel.2007.01.002)
Nakagawa T, Sharma M, Nabeshima Y, Braun RE & Yoshida S 2010Functional hierarchy reversibility within the murine spermatogenicstem cell compartment. Science 328 62–67. (doi:10.1126/science.1182868)
Nascimento HF, Drumond AL, Franca LR & Chiarini-Garcia H 2008Spermatogonial morphology, kinetics and niches in hamsters exposed toshort- and long-photoperiod. International Journal of Andrology 32486–497. (doi:10.1111/j.1365-2605.2008.00884.x)
Oakberg EF 1956 A description of spermiogenesis in the mouse and its usein analysis of the cycle of the seminiferous epithelium and germ cellrenewal. American Journal of Anatomy 99 391–414. (doi:10.1002/aja.1000990303)
de Rooij DG & Grootegoed JA 1998 Spermatogonial stem cell. CurrentOpinion in Cell Biology 10 694–701. (doi:10.1016/S0955-0674(98)80109-9)
de Rooij DG & Russell LD 2000 All you wanted to knowabout spermatogonia but were afraid to ask. Journal of Andrology 21776–798.
www.reproduction-online.org

Spermatogonial morphology during ontogeny 155
Roosen-Runge EC & Leik J 1968 Gonocyte degeneration in the postnatalmale rat. The American Journal of Anatomy 122 275–299. (doi:10.1002/aja.1001220208)
Russell LD, Ettlin RA, Sinha Hikim AP & Clegg ED 1990 Histologicaland Histopathological Evaluation of the Testis, edn 1. Vienna, IL: CacheRiver Press.
Russell LD, Chiarini-Garcia H, Korsmeyer SJ & Knudson CM 2002 Baxdependent spermatogonia apoptosis is required for testicular develop-ment and spermatogenesis. Biology of Reproduction 66 950–958.(doi:10.1095/biolreprod66.4.950)
Shinohara T, Orwing KE, Avarbock MR & Brinster RL 2001 Remodeling ofthe postnatal mouse testis is accompanied by dramatic changes in stemcell number and niche accessibility. PNAS 98 6186–6191. (doi:10.1073/pnas.111158198)
Sinha-Hikim AP, Bartke A & Russell LD 1988 Morphometric studies onhamster testes in gonadally active and inactive states: light microscopefindings. Biology of Reproduction 39 1225–1237. (doi:10.1095/biolre-prod39.5.1225)
www.reproduction-online.org
Timmons PM, Rigby PW & Poirirer F 2002 The murine seminiferousepithelial cycle is pre-figured in the Sertoli cells of the embryonic testis.Development 129 635–647.
Vergouwen RPFA, Jacobs SGP, Huiskamp R, Davids JAG & de Rooij DG1991 Proliferative activity of gonocytes, Sertoli cells and interstitial cellsduring testicular development in mice. Journal of Reproduction andFertility 93 233–243. (doi:10.1530/jrf.0.0930233)
Yoshida S, Sukeno M, Nakagawa T, Ohbo K, Nagamatsu G, Suda T &Nabeshima Y 2006 The first round of mouse spermatogenesis is adistinctive program that lacks the self-renewing spermatogonia stage.Development 133 1495–1505. (doi:10.1242/dev.02316)
Received 19 October 2010
First decision 9 December 2010
Revised manuscript received 25 February 2011
Accepted 26 April 2011
Reproduction (2011) 142 145–155

ANEXO II - TRABALHO ENVIADO PARA PUBLICAÇÃO

Effects of multiple doses of cyclophosphamide on mouse testes: Accessing the germ cells lost, and the functional damage of stem cells
Ana Luiza Drumond a, b, Connie C. Weng a, Gensheng Wang a, Helio Chiarini-Garcia b, Leticia Eras-Garcia b, Marvin L. Meistrich a
Ana Luiza Drumond: [email protected]; Connie C Weng: [email protected]; Gensheng Wang: [email protected]; Helio Chiarini-Garcia: [email protected]; Leticia Garcia: [email protected]; Marvin Meistrich: [email protected].
a Department of Experimental Radiation Oncology, M.D. Anderson Cancer Center, University of
Texas, 1515 Holcombe Blvd, 77030, Houston/TX, USA. b Laboratory of Structural Biology and Reproduction, Department of Morphology, Institute of
Biological Sciences, Federal University of Minas Gerais, Av. Antonio Carlos, 6627 - Pampulha, 31.270-901, Belo Horizonte/MG, Brazil.
Corresponding author: Ana Luiza Drumond, Department of Molecular and Human Genetics, Baylor College of Medicine, One Baylor Plaza, 77030, Houston/TX, USA; Phone: (713) 798-5964; FAX: (713) 798-7418; [email protected]. The authors declare that there are no conflicts of interest. Abbreviations: CY, cyclophosphamide; p.o., per oral; i.p., intraperitoneal; GCNA 1, germ cell nuclei antigen 1; Expt., experiment; SHC, sperm head count; HRLM, high resolution light microscopy; Pl and preL, preleptotene spermatocytes.

Abstract
Spermatogenesis is sensitive to the chemotherapeutic drug cyclophosphamide,
which decreases the patients’ sperm count. Since the recovery of fertility is dependent on
regeneration from stem cells, in the present study we designed a cyclophosphamide-5
regimen to eliminate differentiating germ cells in mouse, and evaluated the ability of
cyclophosphamide-exposed stem spermatogonia to regenerate spermatogenesis in situ
and after transplantation. Seven doses of cyclophosphamide given at 4-day intervals
largely eliminated the differentiating germ cells but left ~50% of the undifferentiated type A
spermatogonia. We monitored the recovery and found that sperm production recovered to 10
64% of control within the time expected. When the cyclophosphamide-surviving
spermatogonia were transplanted into recipient mice, the recovery of spermatogenesis
from the cyclophosphamide-exposed donor cells was reduced to 12% of that expected,
when compared to cryptorchid donors. Thus, multidose regimens of cyclophosphamide
did not eliminate the stem spermatogonia, but resulted in cell loss and residual damage. 15
Key Words: cyclophosphamide, mice, spermatogonial stem cells, testis
20

1. Introduction
25
The seminiferous epithelium of the testis is very susceptible to damage caused by
toxicants, particularly chemotherapy and radiotherapy used in the treatment of cancer. As
a result, young patients cured of cancer often suffer prolonged and sometimes permanent
reductions in sperm counts or azoospermia and are infertile.
The most sensitive testicular cells to chemotherapeutic drugs and radiation are the 30
ones that are undergoing constant mitotic activity, which are the differentiating
spermatogonia [1-3]. The death of these cells results in maturation depletion of later
stages of germ cells and reductions in sperm counts.
The eventual recovery of sperm production depends on the survival of the
spermatogonial stem cells and their ability to differentiate to spermatozoa. In rodents, the 35
spermatogonial stem cells have been identified morphologically as individual isolated cells
called A single spermatogonia (As) [4]. When these cells divide without complete
separation of their cytoplasm, they form A paired spermatogonia (Apr), which undergo
further mitotic divisions, forming a chain of cells known as A aligned spermatogonia (Aal).
Finally, the Aal spermatogonia can form A1 differentiating spermatogonia, which begin the 40
precisely timed sequence of mitotic divisions and the differentiation process towards the
formation of the spermatozoa.
If stem cells survive and their microenvironment is not affected, recovery of
spermatogenesis will occur rapidly [2]. However, if some spermatogonial stem cells are
killed, as occurs with radiation and some chemotherapeutic agents, recovery is more 45
gradual [5,6]. In mice, it was shown that the interval needed for the recovery of fertility is
directly proportional to the level of killing of the spermatogonial stem cells. There is also
evidence that these agents can produce damage to the somatic tissue, and in rat testes,
even though numerous stem spermatogonia survived the cytotoxic treatment, there was no
recovery of the seminiferous epithelium [7,8]. 50
The first methods developed to assess functional damage to the stem cells in the
testis relied on quantifying their ability to regenerate spermatogenesis after the toxic insult.
These included counts of regenerating tubules or sperm production at the time when the
sperm would have been derived from surviving stem spermatogonia [2,9,10]. Although
efficient for evaluating damage caused by a cytotoxic agent on the differentiation of stem 55
cells, measurement of these endpoints did not account for possible damage to the somatic
microenvironment of the seminiferous epithelium which could reduce the level of germ cell

differentiation [8]. More recently, it has been demonstrated that spermatogonial
transplantation [11] can be used to assess stem cell numbers and function after cytotoxic
treatment, eliminating the possible effects of damage to the endogenous somatic cells. 60
This method was used to determine the decrease in the stem cell number after a single
injection of the chemotherapeutic drug busulfan, and the subsequent recovery of stem cell
numbers in these mice. Spermatogonial transplantation has also been combined with
assessment of undifferentiated A spermatogonia to demonstrate stem cell ablation by
busulfan in rhesus monkeys [12]. 65
One chemotherapeutic drug that is widely used for cancer treatments and
immunosuppression for nephrotic disorders is cyclophosphamide (CY). This drug causes
reduction in patients’ sperm counts during treatment. Recovery from moderate doses of
CY is possible, sometimes after a prolonged period of azoospermia; however high doses
of CY cause permanent azoospermia in humans [13,14]. To better elucidate the action of 70
CY on spermatogonial stem cell survival and the subsequent recovery of
spermatogenesis, studies using experimental animal models systems are needed.
In mice, Lu and Meistrich [2] have studied the comparative effects of single
injections of several different chemotherapeutic drugs, given as single doses, on
spermatogenesis. One 200-mg/Kg dose of CY totally eliminated differentiating 75
spermatogonia (A2 stage and later) and early preleptotene spermatocytes and this resulted
in a decline in testicular sperm count 29 days after treatment. In contrast the
undifferentiated A spermatogonia, which include the spermatogonial stem cells, did not
appear to be affected by this treatment and testicular sperm count was completely
recovered 56 days after treatment. 80
Similar results were observed in rats. A single injection of CY induced a peak of
apoptosis among basally located germ cells within 1 day, primarily in stages where A4 to
intermediate spermatogonia were present [15]. However even high doses CY resulted in
very little spermatogonial stem cell killing as evidenced by recovery of sperm counts 9
weeks after injection [16]. 85
Based on the above observations of the high sensitivity of rodent differentiating
spermatogonia to CY but the relative resistance of the stem cells, one of the initial goals of
the present study was to use multiple doses of CY to eliminate most of the later germ cells
and produce an enriched population of mouse stem spermatogonia. We determined if the
stem cells left in the epithelium were functional in situ, by showing endogenous recovery of 90
spermatogenesis in the treated host. Finally, after the isolation of a pool of cells, we used

germ cell transplantation assay to assess the functional ability of treated stem cells, and
determine whether the recovery in the recipient mice is compatible with the results in situ.
2. Materials and Methods 95
2.1. Animals and treatments
Male mice of the C57BL/6Law strain were used unless otherwise noted. Animals
were treated at approximately 8 weeks of age. All procedures were approved by the M. D.
Anderson Cancer Center Institutional Animal Care and Use Committee. 100
Animals received injections of CY (Cytoxan, Bristol-Myers Squibb Co, Princeton,
NJ) through oral (p.o., 22G gavage needle) or intraperitoneal (i.p) administration. Control
animals received saline injections. Several experiments were performed to examine
effects of different treatment protocols
In Experiment 1, mice were treated with a single injection of CY (Fig. 1). Animals 105
received single doses of CY at150 mg/Kg or 200 mg/Kg, either p.o. or i.p.. Animals were
euthanized at 1, 2, 3, 11 and 28 days after administration of the drug, and various
parameters were evaluated for each endpoint.
In Experiment 2, mice were treated with six doses of CY, at a dose of 150 mg/Kg
each. The drug was given at 5-day intervals (Fig. 1), through both the p.o. and i.p. routes, 110
over a period of 25 days. Animals were euthanized on days 28, 59, 62, 65, 70 and 81 (3,
34, 37, 40, 45, and 56 days after the last injection). The specific time points were chosen
based on the time intervals for the different cell types to produce testicular sperm [2,4]
(Table 1).
In Experiment 3, mice were treated with seven doses of CY, given p.o. at 150 115
mg/Kg each. The drug was given with 4-day intervals between doses (Fig. 1), over a
period of 24 days. Animals were euthanized on days 28 and 80 (4 and 56 days after last
injection, respectively).
For the transplantation assay, male C57BL/6-Tg (CAG-EGFP) mice expressing
green fluorescent protein (GFP; Jackson Laboratory, Bar Harbor, ME) were used as 120
donors. Four groups of donors were used: 1) young, 14- to 16-day old mice; 2) untreated
adult 8-week old mice; c) surgically cryptorchid adult mice, and 4) adults treated with CY.
The adult cryptorchid mice were 8 weeks old at the time of the surgery, and had
their testicles raised by suturing the epididymal fat pad to the upper portion of the lateral
abdominal wall [17] for 8 weeks prior being used as donors for the transplantation 125

experiment. Only testicles that weighed less than 30 mg at the time of harvest and
appeared healthy (no sign of necrosis) were used for the cell suspension.
Adult CY-treated mice were 8 weeks old at the beginning of the treatment. Mice
received CY at a dose of 150 mg/Kg p.o., administered every 4 days over a 24-day period
(as in Experiment 3). These animals were used as donors for transplantation 4 days after 130
the last injection of CY.
Recipient animals were male C57BL/6Law mice, at approximately 8 weeks of age,
treated with 2 doses (1.5 +12 Gy) of ionizing radiation separated by 24 hours [21]. They
were used for transplantations 3 to 4 weeks after irradiation.
135
2.2. Histology and morphological analysis
Testes were weighed, fixed with Bouin’s solution and embedded in paraffin or
glycol methacrylate resin. Blocks were sectioned at 5 µm thickness, and sections were
stained with periodic acid-Schiff’s (PAS) and hematoxylin.
For animals evaluated 11 days after injection, tubular cross-sections were scored to 140
determine the stage of the seminiferous epithelial cycle, and whether the layers of cell
types (A4 spermatogonia to zygotene, pachytene to meiosis, and round to step 12
spermatids) were present in normal amounts (>20 in the case of spermatocytes or
spermatids), appreciably reduced (6 to 20), nearly completely absent (1 to 5) or absent (0).
After scoring, we were able to calculate the cell types affected by the drug, at the moment 145
of injection 11 days before the analysis, by back-calculation based on the kinetics of germ
cell development [2].
In order to distinguish the spermatogonial types in the testes, samples were
prepared for high resolution light microscopy [18,19]. Thus, animals were perfusion-fixed
with 5% glutaraldehyde in 0.05 M cacodylate buffer (pH 7.4). Testes were removed, 150
weighed and sliced in small slabs. Fragments were post-fixed in osmium
tetroxide/potassium ferrocyanide mixture and embedded in Araldite 502 (EMS, Hatfield,
PA). One-µm thick sections were obtained from the resin blocks and were stained with
toluidine blue-borate for light microscopy studies.
Morphometric studies were performed to obtain the numbers of each different 155
spermatogonial type (Aund, A1, A2, A3, A4, In, B), preleptotene spermatocytes, meiotic
spermatocytes, and spermatids per 100 Sertoli cells present in seminiferous tubule cross
sections; the Sertoli cells were only counted when the nucleolus was observed. Aund are
undifferentiated type A spermatogonia and include the single, paired, and aligned cells.

Approximately 30 tubular cross sections were counted per animal, using multiple 160
fragments from both testes. The numbers of each germ cell type, and Sertoli cells was
corrected using the Abercrombie formula [20], using the average nuclear diameter of each
cell type, calculated after measuring 10 cells, and the average Sertoli cell nucleoli
diameter, also calculated after measuring 10 nucleoli.
165
2.3. TUNEL staining for apoptosis
The testes were fixed with Bouin’s solution and embedded in paraffin. Five-µm
sections were obtained and stained with the TUNEL reaction (DeadEndTM Colorimetric
TUNEL system, Promega, Madison, WI) to identify apoptotic cells, followed by
PAS/hematoxylin staining. 170
For those animals analyzed 1 to 3 days after one dose of CY (Expt. 1), the stage of
the cycle of the seminiferous epithelium was determined. Only the TUNEL-positive cells
found adjacent to the basal membrane were counted, and these cells were consistent with
the morphology of spermatogonia or preleptotene spermatocytes. Approximately 10
tubular cross-sections were counted per stage, and the percentage of tubules with TUNEL-175
positive cells was calculated for each stage.
For animals assayed 3 days after receiving the last of six doses of CY (Expt. 2),
staging was not possible and the spermatogonial apoptotic index was calculated. The
percentage of spermatogonia undergoing apoptosis was calculated by dividing the number
of TUNEL-positive spermatogonia by the total number of spermatogonia counted. 180
Approximately 30 tubular cross-sections were counted per animal.
2.4. Sperm head count
For sperm head count [10], the right testis was weighed with the tunica albuginea
intact, and placed in 1 ml distilled water. The contents were homogenized, for 30 seconds 185
with a Polytron homogenizer (Brinkmann Instruments, Westbury, NY), keeping the
samples on ice. After homogenization, the samples were sonicated for 4 minutes, in a cold
water bath using a Branson Sonicator (Heat Systems, Ultrasonic, Plainview, NY). Sperm
heads were counted in a hemacytometer with a 40x objective and phase contrast optics.
190

2.5. Transplantation
To obtain the cell suspensions from the testis, the animals were anesthetized and 195
the testicles were collected and weighed. Donor cell suspensions were obtained by
successive enzymatic digestions of GFP mice testes in a 35oC water bath [21], and
filtration of the solution obtained through a 40 µm nylon filter. The final germ cell
suspensions were adjusted to concentrations of between 2 and 6 x 107 per ml prior to
injection into the recipient testes. Approximately 10 µL of the donor cell solution was 200
injected through the efferent ducts.
The recipient animals were euthanized 8 weeks after transplantation and their
testes were fixed with 4% paraformaldehyde, and embedded in paraffin. Serial sections 5
µm thick, were obtained and immunohistochemistry for germ cell nuclei antigen 1 (GCNA1,
rat monoclonal IgM anti-mouse GCNA1, a gift from Dr George Enders) and green 205
fluorescent protein (GFP, rabbit polyclonal anti-GFP; Cat #2956; Cell Signaling, Danvers,
MA) was performed using the ABC Elite kit, appropriate secondary antibodies, and
diaminobenzidine (Vector Laboratories, Burlingame, CA) according to the manufacturer’s
protocol. Sections were counterstained with hematoxylin. The percentages of tubules with
colonies derived from endogenous stem cells (GFP-negative) and from donor stem cells 210
(GFP-positive) were counted in about 30 tubular cross-sections per testis. For comparison
between different donor types, the percentages of tubule cross-sections with donor-derived
colonies found in each testis were normalized to the average numbers of cells injected.
2.6. Statistical analysis 215
Data are expressed as mean standard error. Groups were compared using
ANOVA, followed by Fisher’s LSD or Turkey’s tests when appropriate. Differences were
considered to be significant when p<0.05. All analyses were performed with SPSS 15.0
software (SPSS Inc. Chicago, IL).
220
3. Results
3.1. Effects of a single dose of cyclophosphamide
Initial experiments were performed to characterize the effects of singe injections of
cyclophosphamide (Expt. 1, Fig. 1). With both gavage and i.p administration of either 150 225
or 200 mg/Kg there was a transient loss of body weights 3 days after CY treatment, but

body weights returned to pre-treatment levels by day 7, and were similar to controls
thereafter (data not shown).
Detailed analysis of cell loss was performed with the 150 mg/Kg dose administered
p.o. The TUNEL analysis revealed that CY increased the incidence of apoptotic cells 1 230
and 2 days after administration (Fig. 2). These increases were observed, especially in
tubules in stages II-VII of the cycle of the seminiferous epithelium which contain
intermediate or B spermatogonia and preleptotene spermatocytes, which means that the
cells that were A4 to B spermatogonia at the time of injection were sensitive to induction of
apoptosis. These are the stages during which the largest numbers of mitotically active 235
spermatogonia are present in the testis. The apoptosis returned to the basal level on the
third day after injection, indicating that the cells that underwent apoptosis were cleared
from the tissue during the first two days after exposure to CY.
At 11 days after injection, most tubules showed partial or complete absence of
spermatocytes in meiotic prophase (Fig. 3B). Pachytenes were completely absent from 240
stages IV to IX, and nearly completely absent in stages I to III and stage X (Fig. 4A).
Zygotenes and early pachytenes in stage XII were markedly reduced. Back-calculation
based on the kinetics of spermatogenesis (Fig. 4B) revealed that this dose of CY
specifically killed the differentiating spermatogonia A1-A4, In and B spermatogonia and the
newly formed preleptotene cells, with the A4 to B spermatogonia being the most sensitive. 245
Both earlier (Aund spermatogonia) and later stage cells (spermatocytes in meiotic
prophase), did not appear to be appreciably affected by this dose. These results agree
with the analysis of apoptotic cells showing that the spermatogonia that are undergoing
active proliferation and mitotic divisions are the main target of CY cell killing.
In order to evaluate the relative effectiveness of different doses and routes of 250
administration in depletion of differentiating spermatogonia (types A1-B) mice were treated
with single doses of CY, of either 150 mg/Kg or 200 mg/Kg, both given p.o. and i.p. and
spermatogenesis was evaluated 28 days later. This is the time point at which late
spermatids would be derived from the surviving spermatogonia.
Histological analysis of the seminiferous epithelium (Fig. 3C), 28 days after 255
injection with 150 mg/Kg p.o., showed that the spermatogonia, spermatocytes, and early
spermatids regenerate quite well from surviving undifferentiated spermatogonia. Although
it was common to see appreciable numbers of elongating and condensing spermatids in
tubules at stages IX-XII, most of late elongated spermatids are absent (stages II-VIII). In
contrast, with treatments using 200 mg/Kg p.o. and 150 or 200 mg/Kg i.p, most of the 260

tubules at stages IX-XII had reduced numbers of elongating and condensing spermatids
(Fig. 3D). These results are consistent with the moderate 30% losses in testis weights
observed (Table 2). In addition, the testis weight data showed that intraperitoneal
injections of CY seemed to be slightly more effective at reducing the numbers of
recovering cells than did gavage administrations. 265
The testicular sperm head counts (Table 2) revealed a large decrease in the
numbers of sperm heads present in treated animals. Treatment with 150 mg/Kg p.o.
produced a 10-fold decline in sperm counts. Treatments with 200 mg/Kg by both routes
and 150 mg/Kg i.p produced about 15-fold declines.
270
3.1.2 Acute effects of multiple injections of cyclophosphamide
In order to develop a protocol using the minimal dose to eliminate as many of the
differentiating germ cells as possible, mice were treated with multiple injections of CY to kill
the differentiating spermatogonia as they developed. Animals received 6 doses with a 5-
day interval between injections over a span of 25 days, through either the p.o. or i.p. routes 275
(Expt. 2, Fig. 1), or with 7 doses p.o. with a 4-day interval (Expt. 3). There were minor
toxic effects of the p.o. administration as the mice lost 0.7 ± 0.2 g body weight with the 5-
day intervals and 0.9 ± 0.4 g with the 4-day interval, compared to a 1.9 ± 0.4 g weight gain
in sham-treated controls. However there was more severe toxicity in those treated i.p. as
there was a 2.3 ± 0.2 g loss of body weight. 280
All of the multiple injection regimens caused more severe depletion in the
seminiferous epithelium than did the single dose treatments. The testis weights in animals
treated with multiple doses, 28 days after the first injection, were about 30 mg compared to
60-68 mg with single doses (Table 2).
Histological analyses showed major depletion of the seminiferous epithelium 28 285
days after first injection (Fig. 5C). At this time point there was severe seminiferous tubular
atrophy with all three treatment regimens. There was a very dramatic reduction of most
spermatocytes and spermatids, although a few could still be observed. In addition, there
were some differentiated spermatogonia (Fig. 5D) that survived or recovered from earlier
stage spermatogonia in the 3 days from the last injection, as well as undifferentiated 290
spermatogonia (Fig. 5F) that had the same morphology as those from control animals (Fig.
5E). Nearly all of the spermatogonia appeared normal and viable, only 2.4% were
undergoing apoptosis, which was similar to the percentage in control animals. Thus any

apoptosis that was induced by the last CY injection 3 days previously had been completed,
consistent with what was observed after a single injection of CY. 295
Quantitative counts of cells from the histological material showed that, at 28 days,
all multidose regimens produced large decreases in the numbers of differentiating
spermatogonia, spermatocytes and spermatids (Fig. 6). Analysis of the numbers of
differentiating cells following the 6-dose/ 5-day interval regimens showed that, when given
p.o., there were still some intermediate spermatogonia to preleptotene spermatocytes (7% 300
of control), meiotic spermatocytes (5% of control), and spermatids (4% of control) present.
However, numbers of these cells were effectively reduced to about 1% of control by giving
the same dose i.p., or giving 7 doses p.o. with a shorter interval.
In contrast to the large decreases of the numbers of differentiating germ cells, there
was at most a small effect of multiple doses of CY on the numbers of undifferentiated type 305
A spermatogonia (Fig. 5F and Fig. 6). There was no effect of giving 6 doses of CY p.o. at
150 mg/Kg with a 5-day interval on these cells. There were trends towards reductions in
undifferentiated type A spermatogonial numbers with i.p. injection or the 7-dose/ 4-day
interval p.o. treatment, and they were at most about 50%. Although the treatments with
150 mg/Kg i.p. (5-day intervals) and with 150 mg/Kg p.o. (4-day intervals) produced very 310
similar germ cell counts, the i.p. regimen produced a greater weight loss during treatment,
suggesting this treatment to be more toxic.
3.3. Endogenous recovery of spermatogenesis from surviving stem cells
Since there were remaining stem cells at the end of the treatment, despite the 315
nearly complete tubular atrophy, it gave us an opportunity to assess the time course of the
recovery from this depleted state. The recovery of the seminiferous epithelium after the
end of treatment was evaluated at different times after the multidose treatment regimens.
Qualitative histological analysis performed 56 days after the last injection (80/81
days after the first injection), when the surviving stem spermatogonia should be producing 320
spermatozoa, showed recovery of most of the seminiferous epithelium (Fig. 5G), although
there were still some damaged seminiferous tubules, deficient in germ cells, for all
treatment regimens.
The recovery of spermatogenesis was quantified by measurements of testis weight
and sperm head count (Fig. 7). Both parameters steadily increased after the end of the 325
treatment. At 56 days after the last injection among the animals receiving 6 treatments at
5-day intervals, those receiving i.p. injections only reached testis weights and sperm head

counts that were 70% and 40% of control, respectively, whereas those receiving p.o.
administration had significantly greater recoveries reaching 80% and 64% of control,
respectively. It was noted that none of the treated animals had reached the values 330
measured for control animals. Thus we conclude that recovery of spermatogenesis
following p.o. treatment, although incomplete, is better than after i.p. treatment. The lower
recovery after i.p. injections, despite the equivalent numbers of Aund spermatogonia, might
be due to additional damage to the spermatogonia, to the testicular tissue, and to
extratesticular targets affecting spermatogenesis. 335
Comparing the p.o. treatments, 7 administrations given 4 days apart (Fig. 1, Expt 3)
showed recovery of testis weight to 74% of control, observed 56 days after the last
injection (80 days after first injection), lower than those receiving 6 treatments at 5-day
intervals. However, the numbers of sperm produced, which should be more directly a
measure of recovery of spermatogenesis from the surviving stem cells, at this time point 340
were approximately the same (64% of control) (Fig. 7B); therefore the surviving stem cells
seem to be effectively differentiating even after the more intense treatment regimen.
3.4. Functional evaluation of cyclophosphamide-treated stem cells
Next we tested whether the stem cells that survived CY treatment, and were able to 345
produce recovery of spermatogenesis in situ in the host testis, were fully functional after
germ cell transplantation. For this purpose, the animals were treated with 7 doses of 150
mg/Kg p.o. with 4-day intervals between injections, which we found to be the most efficient
treatment for eliminating differentiating germ cells, with only limited toxicity, and a
seemingly modest effect on the numbers of undifferentiated type A spermatogonia. We 350
compared the efficiency of transplantation with known donor models of spermatogonial
transplantation, including prepubertal mice and cryptorchid mice, which have a high
reported concentrations of stem cells, and untreated adult mice, which have a low reported
concentration of stem cells.
To obtain a prediction of the relative numbers of stem cells in the cell suspension, 355
we counted the numbers of different types of germ cells in the prepubertal, adult,
cryptorchid, and CY-treated testes (Fig. 8). The total numbers of type A undifferentiated
spermatogonia per Sertoli cell were not significantly different among all the adult groups,
but were lower for adult mice treated with CY than for young animals The adult mice had
the highest numbers of later spermatogonia (In and B) and preleptotene spermatocytes and 360
very high numbers of spermatocytes and spermatids. The cryptorchid B6 mice showed

significantly reduced numbers of In spermatogonia to preleptotene spermatocytes and 3- to
4-fold reductions in the numbers of later germ cells, such as spermatocytes and
spermatids. However, we have rarely been able to achieve the complete loss of these
differentiated cells as reported by Nishimune and Aizawa [17]. Immature mice had nearly 365
as many spermatogonia per Sertoli cells as the adult but lower numbers of spermatocytes
and no spermatids. The CY-treated mice had the lowest number of differentiating germ
cells at all stages and very markedly reduced numbers of cells from the intermediate
spermatogonia through spermatids. These results taken together demonstrate that the
percentage of germ cells that were of type Aund spermatogonia was highest for CY animals 370
(Fig. 8), because these animals had fewer differentiated germ cells than all the other three
groups. However, if we include Sertoli cells, which would also be in the cell suspension in
the counts, the percentage of cells that are Aund is highest for the young animals but still
would be next highest for the CY-treated ones.
The number of cells obtained per testis from mice treated with CY, after the cell 375
suspension preparation, was similar to what was observed for cryptorchid animals, but was
lower than the numbers observed for young and adult animals (Table 3). We injected
similar numbers of cells from the cell suspensions (Table 4), obtained from animals of each
of the four groups, through the efferent ducts, in order to produce colonization of recipient
testis after transplantation. 380
Histological analysis of the testes of recipient animals (Fig. 9) harvested 8 weeks
after transplantation were performed by immunostaining for GCNA-1, in order to detect
differentiation of the germ cells, and for GFP, to distinguish endogenous and donor
colonies, among the GCNA-1-positive tubular cross-sections. There was recovery of
spermatogenesis from endogenous stem cells in about 3 to 4% of the tubules of recipient 385
animals with no significant differences between animals transplanted with cells from the
different donor groups. Donor cells from all sources used were capable of colonizing the
recipient testis and to differentiate, forming elongated spermatids (Fig. 9B, D, F and H);
there were no apparent differences in the stages of differentiation or the quality of colonies
produced between the different donor animals. The percentages of tubules that contained 390
GFP positive colonies (donor derived) were assessed and normalized based on the
numbers of cells injected in each group during the transplantation (Table 4). The results
showed that the prepubertal mice and adult cryptorchid mice had the highest
concentrations of functional stem cells in their testes. Despite the higher concentration of
Aund spermatogonia and presumably stem cells present in their cell suspensions (Fig. 8), 395

cells from CY-treated donors resulted in significantly lower levels of colonization than did
those from cryptorchid mice. These results suggest that the loss of functional stem cells
might have been greater than what the in situ recovery suggested or that there was
functional damage to the stem spermatogonia caused by CY so that the numbers of Aund
are not a good measure of functional stem cells. Nevertheless, the cells from CY-treated 400
mice still appeared to show greater colonization than did those from untreated adult mice.
4. Discussion
In the present study we developed a multidose regimen of treatment with CY capable 405
of depleting the differentiated cells in the testis, while maintaining the population of
undifferentiated spermatogonia, which includes the stem cell population. We were able to
find a balance of dose, route of administration, and interval between doses, that eliminated
a wide window of cells, from differentiating spermatogonia to late spermatids, with minimal
animal toxicity, and still allowing survival of an appreciable number of undifferentiated 410
spermatogonia and apparent good recovery of spermatogenesis from surviving
spermatogonial stem cells. This regimen reduced the numbers of differentiated cells to 1%
of control but left ~50% of the undifferentiated type A spermatogonia remaining. The
enrichment of undifferentiated spermatogonia by CY-treatment could potentially be used
as an initial step in the purification of these cells for further molecular characterization of 415
the effects of CY on them.
These results are consistent with and extend other studies in the literature on rodents
showing the sensitivity of differentiating spermatogonia but relative resistance of stem cells
to a wide variety of CY regimens [2,15,22-24]. This is in contrast to some other
chemotherapeutic drugs, such as busulfan, doxorubicin, thiotepa, and procarbazine that 420
effectively kill the stem spermatogonia as well as differentiating spermatogonia [2,25-27].
The presence of healthy looking Aund spermatogonia at the end of the treatment
regimen is the source of the subsequent recovery of spermatogenesis in the treated
testes. Already at 45 days after last injection, before the surviving As had time to go
through further self–renewal divisions and still produce late spermatids, the sperm 425
produced from these stem cells were at approximately 63% of the control levels.
Despite the survival of the stem cells and their active function for recovery of
spermatogenesis in situ in the testis, these cells seemed to have some loss of function
when assayed by transplantation. Notwithstanding the fact that the CY-treated testis had

the highest concentration of Aund cells as a percentage of germ cells, the ability of the stem 430
cells to produce colonies upon transplantation of a given number of cells was only 16% of
that of the suspensions from immature testes and 32% of that from cryptorchid testes.
Concerning the results observed for young mice as donors, it may be difficult to
quantitatively compare with the CY-treated as the yield of cells was much higher per mg-
testis for the young animals. That could occur because the Sertoli cells are still immature 435
and have not yet established the tight junctions facilitating the recovery of these cells and
the basally located spermatogonia in suspension. Inclusion of immature Sertoli cells in the
suspension that was transplanted could possibly provide additional niche function for
development of spermatogenesis [28,29], although we were not able to detect GFP-
stained Sertoli cells in the recipient mice. 440
However comparisons can be made between the cryptorchid and CY-treated animals
as they appear to be very similar in many respects. The testis weights and yields of cells
per testis were the same for both groups and the numbers of Aund per Sertoli cell were
similar. Since both models were derived from treating already adult mice, the somatic cell
types and structure should be largely the same. Yet although the percentage of germ cells 445
that were Aund spermatogonia was ~7 times higher in the CY-treated animals than in the
cryptorchid ones, the number of tubules with colonies generated by CY treated donors was
only 32% of the colonization promoted by cryptorchid cells. Although the initial collagenase
digestion resulted in the removal of most of the interstitial cells, and loss of many Sertoli
cells during the digestion of tubules [30], some of these cells might be present in the 450
suspension. Even if all of the Sertoli cells were recovered in the suspension, the
concentration of Aund spermatogonia would still be 2.6-fold higher in the suspensions from
the CY-treated mice than from the cryptorchid ones, and we expected at least 2.6 times
the number of stem-cell derived colonies upon transplantation. Thus the 32% yield of
colonies from the CY-treated mice compared to the cryptorchid donors is only 12% of 455
expectation, further supporting the conclusion that the stem cells from the former were
functionally impaired.
There is general agreement that the stem cells are contained within the population of
Aund spermatogonia. There is one report in the literature that c-kit-positive spermatogonia
are capable of colonizing recipient testes [31], implying that these must be differentiated 460
spermatogonia. If this were true, then the reduced number of differentiated spermatogonia
in the CY-treated mice could possibly account for the lower transplantation efficiency.
However, c-kit expression begins in late stage Aal [32] so the colonization ability of the c-kit

positive cells could be due to Aal, which are undifferentiated spermatogonia, or possibly
early stage differentiated type A cells. However the numbers of A1 spermatogonia are not 465
reduced by the CY-treatment (data not shown), so this cannot explain the lower colonizing
ability of the cells from CY-treated mice.
The histology method we used, high-resolution light microscopy of tissue sections,
permits identification of the Aund spermatogonia but does not show their chain sizes or
provide other markers. The stem cell pool that contributes to steady state spermatogenesis 470
is largely contained within a subpopulation of As spermatogonia, however the Apr and Aal
can occasionally contribute to the stem cell pool by fragmentation [32]. These chains are
considered to be potential stem cells and contribute more markedly to the self-renewing
stem cells during recovery from cell depletion, as occurs after treatment with a
chemotherapy agent such as busulfan or after transplantation [33]. Although we have not 475
determined the distribution of As, Apr, and Aal within these Aund spermatogonia, we can
conclude that the percentage of these that are stem cells should be at least as high as in
the untreated testis. Since this treatment involves 7 CY doses, the presence of appreciable
numbers of Apr and Aal cells, which constitute the bulk of the Aund population, at 4 days after
the last treatment indicates that their progenitors, the stem cells, must have survived the 480
first 6 injections and hence should have survived the seventh injection. The lower
colonizing efficiency of cells from the CY-treated mice than from the cryptorchid mice may
reflect differences within the Aund compartment. Further studies characterizing the clone
size and stem and differentiation markers in the surviving Aund population from cryptorchid
and CY-treated mice might further elucidate reasons for the differences in stem cell 485
functional abilities.
It is also possible that each CY-treatment is killing a fraction of the stem cells, but the
loss is compensated by increased clone fragmentation and increased self-renewal that
occurs after cell loss [34]. Sperm production could be maintained after recovery from CY-
treatment, despite killing of stem cells, by increased division of the Aal and reduction of 490
differentiated type A spermatogonial apoptosis [35]. Thus the transplantation assay may
be more directly measuring the loss of surviving functional stem cells, whereas
endogenous recovery is not sensitive to small to moderate losses in stem cell numbers.
Thus, the more efficient transplantation from the cryptorchid mice actually reflects higher
numbers of functional stem cells than in the CY-treated mice. 495
Alternatively it is possible that a high number of functional stem cells are indeed
present in the CY-treated testes, and although they are functional in situ, they have some

damage that affects their ability to colonize an irradiated recipient testis. We have
previously shown that the colonization and differentiation ability of donor cells from
immature mice were improved in the recipient testis about 6-fold by suppression of 500
testosterone in the recipient mice before and after the transplantation [36]. Therefore, it
may be worthwhile testing whether hormone suppression could even more dramatically
enhance the functionality of the CY treated stem cells in the recipient testis.
In summary, the stem spermatogonia in mice seem to survive multiple doses of CY,
and very pronounced in situ recovery is observed. Still there is evidence for some loss of 505
stem cells or residual damage to them when they are isolated from the testis and
transplanted, but the specific reason why it occurs still remains to be elucidated. The
observation of stem cell loss or damage in CY-treated mice may provide methods for
further understanding of the apparently greater sensitivity of the human testis to prolonged
azoospermia after CY treatment [14,37]. 510
Acknowledgements
The present study was funded by the NIH, the Florence M. Thomas Professorship in
Cancer Research, and by the program CAPES-UT, established between the University of Texas
and the Brazilian agency CAPES. We thank Shan Shao, Nalini Patel and Kuriakose Abraham, from
the MDACC, and Maria Luiza Silva, from UFMG, for their technical assistance with histological
preparations and immunostaining. We thank Dr G. Enders for kindly supplying anti GCNA1 antibody
and Dr R. Behringer for opening his laboratory for making glass pipettes.
References
[1] Dym M, Clermont Y. Role of spermatogonia in the repair of the seminiferous epithelium
following x-irradiation of the rat testis. Am J Anat 1970;128:265-82.
[2] Lu CC, Meistrich ML. Cytotoxic effects of chemotherapeutic drugs on mouse testis cell.
Cancer Res 1979;39:3575-82.
[3] Kangasniemi M, Veromaa TI, Kulmala J, Kaipia A, Parvinen M, Toppari 1. DNA-flow
cytometry of defined stages of rat seminiferous epithelium: effects of 3 Gv of high-energy X-
irradiation. J Androl 1990;11:312-7.
[4] de Rooij DG. Stem cells in the testis. Int J Exp Pathol 1998;79:67-80.
[5] Meistrich ML, Hunter NR, Suzuki N, Trostle PK, Withers HR. Gradual regeneration of mouse
testicular stem cells after exposure to ionizing radiation. Radiat Res 1978;74:349-62.

[6] Meistrich ML. Quantitative relation between testicular stem cell survival, sperm production,
and fertility in the mouse after treatment with different cytotoxic agents. J Androl 1982;3:58-
68.
[7] Kangasniemi M, Huhtaniemi I, Meistrich ML. Failure of spermatogenesis to recover despite
the presence of A spermatogonia in the irradiated LBNF1 rat. Biol Reprod 1996;54:1200-8.
[8] Zhang Z, Shao S, Meistrich ML. The radiation-induced block in spermatogonial differentiation
is due to damage to the somatic environment, not the germ cells. J Cell Physiol
2007;211:149-58.
[9] de Ruiter-Bootsma AL, Kramer MF, de Rooij DG. Response of stem cells in the mouse testis
to fission neutrons of 1 MeV mean energy and 300 kV x rays. Methodology, doseresponse
studies, relative biological effectiveness. Radiat Res 1976;67:56–68.
[10] Meistrich ML, Van Beek M. Spermatogonial stem cells: Assessing their survival and ability
to produce differentiated cells. In: Chapin RE, Heindel JJ, editors. Methods in Toxicology
vol.3A, San Diego: Academic Press Inc; 1993, p. 106-23.
[11] Brinster RL, Zimmermann JW. Spermatogenesis following male germ-cell transplantation.
Dev Biol 1994;91:11298-302.
[12] Hermann BP, Sukhwani M, Lin CC, Sheng Y, Tomko J, Rodriguez M, et al.
Characterization, cryopreservation, and ablation of spermatogonial stem cells in adult
rhesus macaques. Stem Cells 2007;25:2330-8.
[13] Pryzant RM, Meistrich ML, Wislon G, Brown B, McLaughlin P. Long-term reduction in sperm
count after chemotherapy with and without radiation therapy for non-hodgkin’s lymphomas.
J Clin Oncol 1993;11:239-47.
[14] Fukutani K, Ishida H, Shinohara M, Minowada S, Niijima T, Hijikata K, et al. Supression of
spermatogenesis in patients with behçet’s disease treated with cyclophosphamide and
colchicine. Fertil Steril 1981;36:76-80.
[15] Cai L, Hales BF, Robaire B. Induction of apoptosis in the germ cells of adult male rats after
exposure to cyclophosphamide. Biol Reprod 1997;56:1490-7.
[16] Meistrich ML, Parchri N, Wilson G, Kurdoglu B, Kangasniemi M. Hormonal protection from
cyclophosphamide-induced inactivation of rat stem spermatogonia. J Androl 1995;16:334-
41.
[17] Nishimune Y, Aizawa S. Temperature sensitivity of DNA synthesis in mouse testicular germ
cells in vitro. Exp Cell Res 1978;113:403-8.
[18] Chiarini-Garcia H, Russell LD. High resolution light microscopic characterization of mouse
spermatogonia. Biol Reprod 2001;65:1170-8.
[19] Chiarini-Garcia H, Meistrich ML. High resolution light microscopic characterization of
spermatogonia. In: Hou SX, Singh SR, editors. Germline Stem Cells, New York: Humana
Press Inc; 2008, p. 95-107p.

[20] Abercrombie M. Estimation of nuclear population from microtome sections. Anat Rec 1946;
94:239-47.
[21] Zhang Z, Shao S, Meistrich ML. Irradiated mouse testes efficiently support spermatogenesis
derived from donor germ cells of mice and rats. J Androl 2006;27:365-75.
[22] Karashima T, Zalatnai A, Schally AV. Protective effects of analogs of luteinizing hormone-
releasing hormone against chemotherapy-induced testicular damage in rats. Proc Natl Acad
Sci USA 1988; 85:2329-33.
[23] Velez de Ia Calle JF, de Queiroz F, Gamier DH, Kercret H, Folliot R, Jegou, B. Reproductive
effects of the anticancer drug cyclophosphamide in male rats at different ages. Arch Androl
1989;22:251-63.
[24] Russell LD, Russell JA. Short-term morphological response of the rat testis to administration
of five chemotherapeutic agents. Am J Anat. 1991;192:142-68.
[25] Meistrich ML, Finch M, da Cunha MF, Hacker U, Au WW. Damaging effects of fourteen
chemotherapeutic drugs on mouse testis cells. Cancer Res 1982;42:122-31.
[26] Brinster RL, Avarbock MR. Germline transmission of donor haplotype following
spermatogonial transplantation. Proc Natl Acad Sci USA 1994;91:11303-7.
[27] Bucci LR, Meistrich ML. Effects of busulfan on murine spermatogenesis: cytotoxicity,
sterility, sperm abnormalities and dominant lethal mutations. Mutat Res 1987;176:159-268.
[28] Kanatsu-Shinohara M, Miki H, Inoue K, Ogonuki N, Toyokuni S, Ogura A, Shinohara T.
Germline niche transplantation restores fertility in infertile mice. Hum Reprod 2005;20:
2376-82.
[29] Zhang Z, Shao S, Shetty G, Meistrich M. Donor Sertoli cells transplanted into irradiated rat
testes stimulate partial recovery of endogenous spermatogenesis. Reprod 2009;137:497-
508.
[30] Meistrich ML, Bruce WR, Clermont Y. Cellular composition of fractions of mouse testis cells
following velocity sedimentation separation. Exp Cell Res 1973;79:213-27.
[31] Barroca V, Lassalle B, Coureuil M, Louis JP, Le Page F, Testart J, et al. Mouse
differentiating spermatogonia can generate germinal stem cells in vivo. Nat Cell Biol
2008;11:190-201.
[32] Nakagawa T, Sharma M, Nabeshima Y, Braun RE, Yoshida S. Functional hierarchy and
reversibility within the murine spermatogenis stem cell compartment. Science 2010;328:62-
7.
[33] Nakagawa T, Nabeshima Y, Yoshida S. Functional identification of the actual and potential
stem cell compartments in mouse spermatogenesis. Dev Cell 2007;12:195-206.
[34] de Rooij DG. Regulation of the proliferation of spermatogonial stem cells. J Cell Sci Suppl
1988;10:181-94.
[35] de Rooij DG. Proliferation and differentiation of spermatogonial stem cells. Reproduction
2001;121:347-54.

[36] Wang G, Shao SH, Weng CC, Wei C, Meistrich ML. Hormonal suppression restores fertility
in irradiated mice from both endogenous and donor-derived stem spermatogonia. Toxicol
Sci 2010;117:225-37.
[37] Hsu AC, Folami AO, Bain J, Rance CP. Gonadal function in males treated with
cyclophosphamide for nephrotic syndrome. Fertil Steril 1979;31:173-8.
Tables
Table 1 – Assessment intervals for sperm counts chosen based on predicted
recovery of sperm production from different cell types potentially affected by
cyclophosphamide.
Days after last injection Latest stage of cell capable of producing testicular
spermatozoa at indicated time a
34 37 40 45 56
A2 spermatogonia, stage IX Aaligned spermatogonia, stage VI Apaired spermatogonia, stage I Asingle spermatogonia, stage VI
Asingle spermatogonia, earlier cycle a Based on different stage of the seminiferous epithelial cycle [4].
Table 2 - Testis weight and sperm head count per testis 28 days
the start of treatment. Treatment was given as a single dose
(Expt. 1) or with 6 doses given at 5-day intervals (Expt. 2) or 7
doses at 4-day intervals (Expt. 3) (N=3). Different letters indicate
statistically significant differences among all groups (p<0.05).
Testis Weight Sperm Head Count
Control 83.8 ± 2.6 a 16.7 ± 1.9 x10
6 a
Expt. 1 150 p.o. 67.8 ± 3.3
b 1.9 ± 0.2 x10
6 b,c
150 i.p. 59.9 ± 3.6 c 1.2 ±0.05 x10
6 c
200 p.o. 63.0 ± 0.8 b,c
1.0 ±0.02 x106 c
200 i.p. 57.8 ± 0.5 c 1.1 ± 0.2 x10
6 c
Expt. 2 150 p.o. / 5 days 31.3 ± 2.8
d 2.2 ± 0.3 x10
6 b
150 i.p. / 5 days 30.3 ± 3.3 d 2.9 ± 0.1 x10
6 b
Expt. 3 150 p.o. / 4 days 30.5 ± 1.1
d 2.9 ± 0.3 x10
6 b
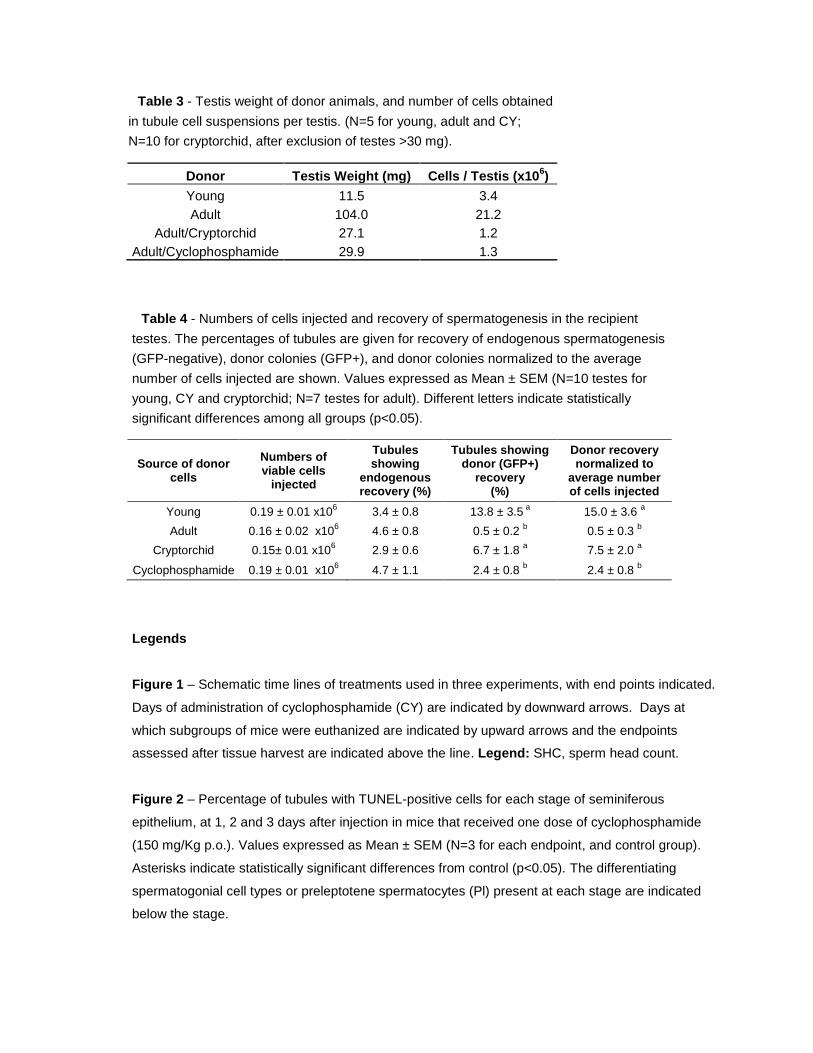
Table 3 - Testis weight of donor animals, and number of cells obtained
in tubule cell suspensions per testis. (N=5 for young, adult and CY;
N=10 for cryptorchid, after exclusion of testes >30 mg).
Donor Testis Weight (mg) Cells / Testis (x106)
Young 11.5 3.4
Adult 104.0 21.2
Adult/Cryptorchid 27.1 1.2
Adult/Cyclophosphamide 29.9 1.3
Table 4 - Numbers of cells injected and recovery of spermatogenesis in the recipient
testes. The percentages of tubules are given for recovery of endogenous spermatogenesis
(GFP-negative), donor colonies (GFP+), and donor colonies normalized to the average
number of cells injected are shown. Values expressed as Mean ± SEM (N=10 testes for
young, CY and cryptorchid; N=7 testes for adult). Different letters indicate statistically
significant differences among all groups (p<0.05).
Source of donor cells
Numbers of viable cells
injected
Tubules showing
endogenous recovery (%)
Tubules showing donor (GFP+)
recovery (%)
Donor recovery normalized to
average number of cells injected
Young 0.19 ± 0.01 x106 3.4 ± 0.8 13.8 ± 3.5
a 15.0 ± 3.6
a
Adult 0.16 ± 0.02 x106 4.6 ± 0.8 0.5 ± 0.2
b 0.5 ± 0.3
b
Cryptorchid 0.15± 0.01 x106 2.9 ± 0.6 6.7 ± 1.8
a 7.5 ± 2.0
a
Cyclophosphamide 0.19 ± 0.01 x106 4.7 ± 1.1 2.4 ± 0.8
b 2.4 ± 0.8
b
Legends
Figure 1 – Schematic time lines of treatments used in three experiments, with end points indicated.
Days of administration of cyclophosphamide (CY) are indicated by downward arrows. Days at
which subgroups of mice were euthanized are indicated by upward arrows and the endpoints
assessed after tissue harvest are indicated above the line. Legend: SHC, sperm head count.
Figure 2 – Percentage of tubules with TUNEL-positive cells for each stage of seminiferous
epithelium, at 1, 2 and 3 days after injection in mice that received one dose of cyclophosphamide
(150 mg/Kg p.o.). Values expressed as Mean ± SEM (N=3 for each endpoint, and control group).
Asterisks indicate statistically significant differences from control (p<0.05). The differentiating
spermatogonial cell types or preleptotene spermatocytes (Pl) present at each stage are indicated
below the stage.

Figure 3 – Seminiferous tubule cross-sections of testes of mice that received saline injection (A),
and those that received one injection of cyclophosphamide, p.o. at 150 mg/Kg, 11 days previously
(B), or 28 days previously (C, D). Arrowheads: areas where spermatocytes (B) or elongated
spermatids (C, D) are missing; Arrows: areas where there are remaining elongating and condensed
spermatids; Stages of seminiferous epithelium are indicated in the lumen of the tubules. (A,C,D)
paraffin embedded; (B) methacrylate embedded. Bars indicate 70 micrometers.
Figure 4 – Germ cells organized in the twelve stages of seminiferous epithelium cycle. Stages/germ
cells that are affected by the cyclophosphamide are shaded.
(A) Cells observed in testes 11 days after p.o. CY injection at 150 mg/Kg. (B) Calculated cells
affected at time of injection. Arrows indicate mitotic divisions and spermatogonial differentiation. A1,
A2, A3, A4, In and B, differentiating spermatogonia; preL, preleptotene; L, leptotene; Z, zygotene; P,
pachytene; D, diplotene; M, meiotic divisions. Appreciably reduced; Nearly completely
absent; Absent.
Figure 5 – Seminiferous tubule cross-sections of mice that received saline injections (A,B) showing
Sertoli cells (S) and type B differentiating spermatogonia . (C,D), Seminiferous tubules of mice that
received multiples doses of cyclophosphamide (Expt. 2 -150 mg/Kg p.o., 6 doses, at 5-day
intervals), 28 days after first injection showing severe damage to the germinal epithelium at the end
of the treatment. The tubules contain predominantly Sertoli cells (S). Some remaining
spermatocytes (arrowheads) and spermatids are observed as well as some type B (“B”) and type A
spermatogonia. (E,F) Undifferentiated type A spermatogonia in the control and the treated animals,
respectively. (G) Seminiferous tubules of the treated animals, 81 days after first injection, showing
recovery in most of the tubules (**), but some tubules are still damaged (*). Tissues in A, C, and G
were Bouin’s fixed, paraffin embedded and stained with PAS-hematoxylin. Those in B, D, E, and F
were prepared for HRLM by glutaraldehyde fixation, araldite embedding, and toluidine blue staining.
Black bars represent 70 micrometers, and white bars represent 20 micrometers (B,D) or 10
micrometers (E,F).
Figure 6 – Relative numbers of different types of germ cells per 100 Sertoli cells in tubules of
animals treated with 150 mg/Kg dose of cyclophosphamide p.o. and i.p., every 5 days, and 150
mg/Kg p.o., every 4 days. Legend: In-Pl, differentiating In and type B spermatogonia plus
preleptotene spermatocytes. Aund, undifferentiated spermatogonia type A; A1-A4, differentiating type
A spermatogonia. Values expressed as Mean ± SEM (N=3). Different letters indicate statistically
significant differences among all groups (p<0.05).

Figure 7 – A) Testis weight and B) sperm head count per testis for mice treated with multiple doses
of cyclophosphamide (150 mg/Kg p.o. or i.p.), starting 28 days after initiation of treatment, showing
the recovery of spermatogenesis. Note only 28-day and 80-day points were obtained for Expt. 3
(p.o doses at 4 day intervals) A sham-treated control group is shown for comparison. Times are
given on the X-axis from the time of the first injection. Values expressed as Mean ± SEM (N=3).
Different letters indicate statistically significant differences among all groups (p<0.05).
Figure 8 – Relative numbers of different types of germ cells per 100 Sertoli cells in tubules of young
mice, and adults with no treatment, cryptorchidization, or treatment with cyclophosphamide (CY),
and percentage of germ cells that are A undifferentiated spermatogonia. Legend: In-Pl,
differentiating In and type B spermatogonia plus preleptotene spermatocytes. Aund, undifferentiated
type A spermatogonia; A1-A4, differentiating type A spermatogonia. Values expressed as Mean ±
SEM (N=3). Different letters indicate statistically significant differences among all groups (p<0.05).
Figure 9 – Histological and immunohistochemical analysis of seminiferous tubules of the recipient
animals transplanted with cells from prepubertal donors (A,B), adult donors (C,D), adult cryptorchid
donors (E,F) and adults treated with cyclophosphamide (CY) (G,H), at 8 weeks after
transplantation. Slides were immunostained with antibodies to GFP and visualized with the
diaminobenzidine chromophore (brown). An endogenous colony is indicated by "*" and donor
colonies are stained brown. At higher magnification (B,D,F,G) the production of elongated
spermatids (arrows) at this time point is observed in donor colonies. White bars indicate 70
micrometers.
NOTE: This figure is intended for color reproduction on the web and in print.
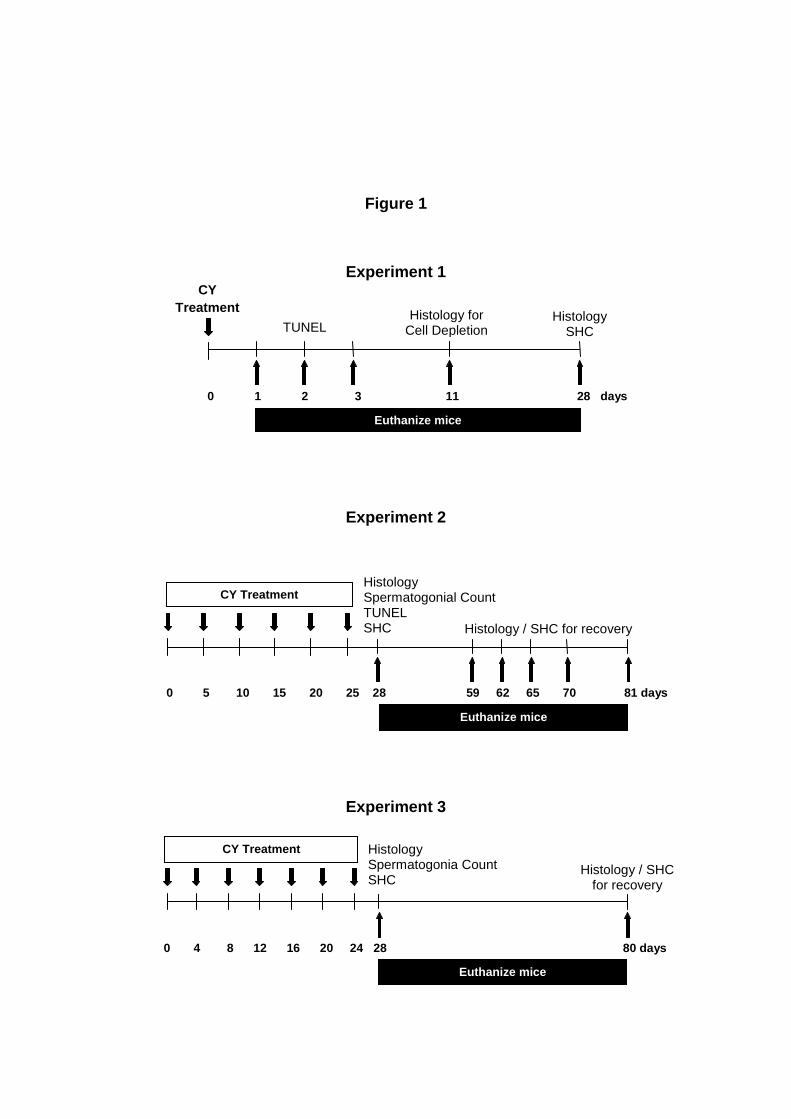
Figure 1
Experiment 1
0 1 2 3 11 28 days
Experiment 2
0 5 10 15 20 25 28 59 62 65 70 81 days
Experiment 3
0 4 8 12 16 20 24 28 80 days
Euthanize mice
Euthanize mice
CY
Treatment
Euthanize mice
CY Treatment
CY Treatment
Histology / SHC for recovery
Histology Spermatogonial Count TUNEL SHC
Histology / SHC for recovery
Histology Spermatogonia Count SHC
TUNEL Histology
SHC
Histology for Cell Depletion
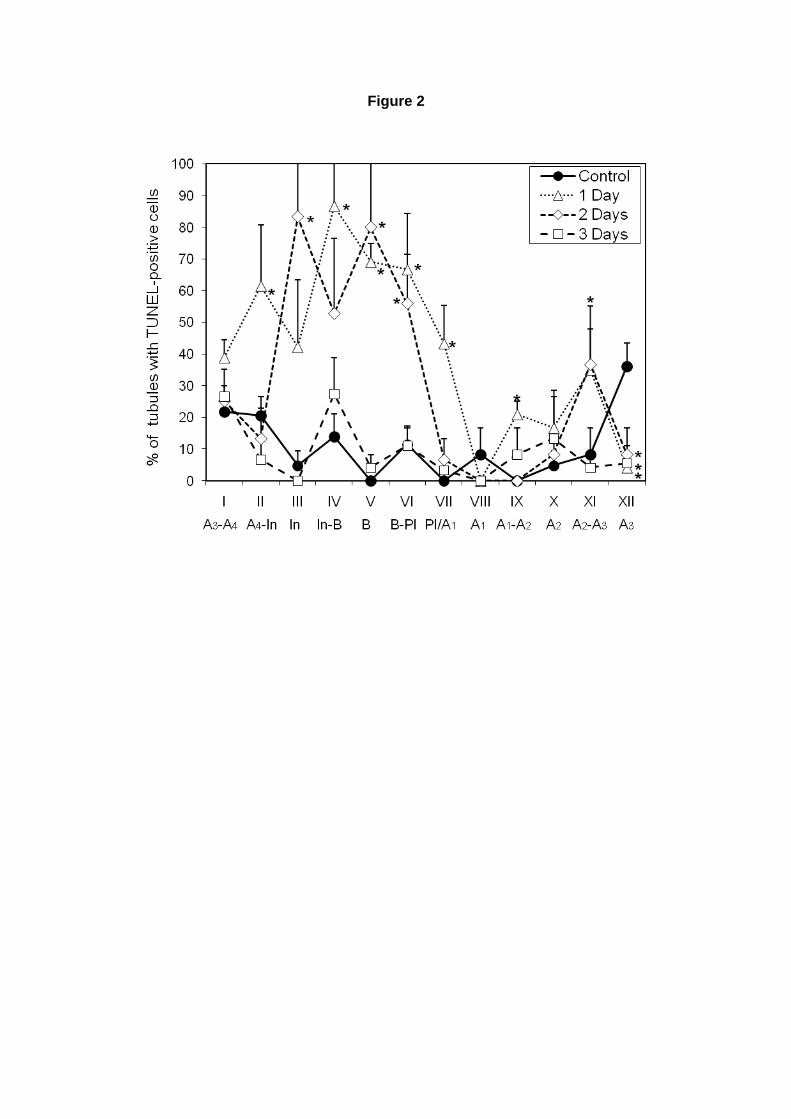
Figure 2
Figure 3
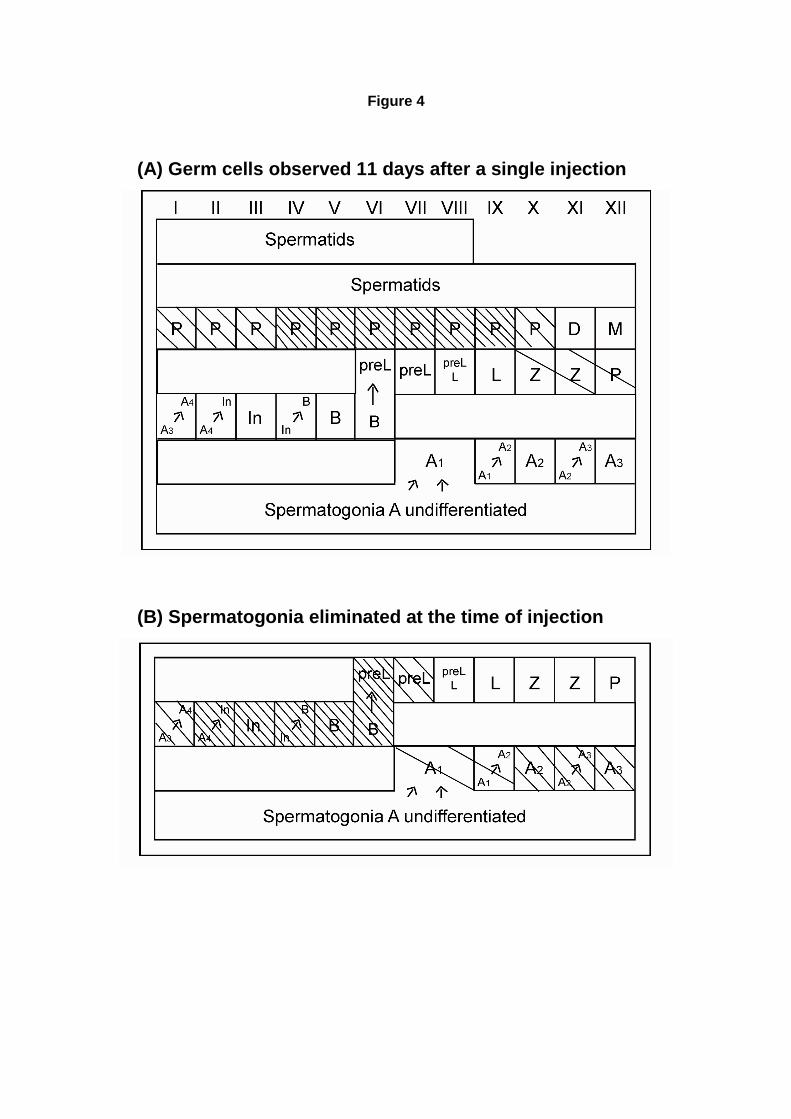
Figure 4
(A) Germ cells observed 11 days after a single injection
(B) Spermatogonia eliminated at the time of injection
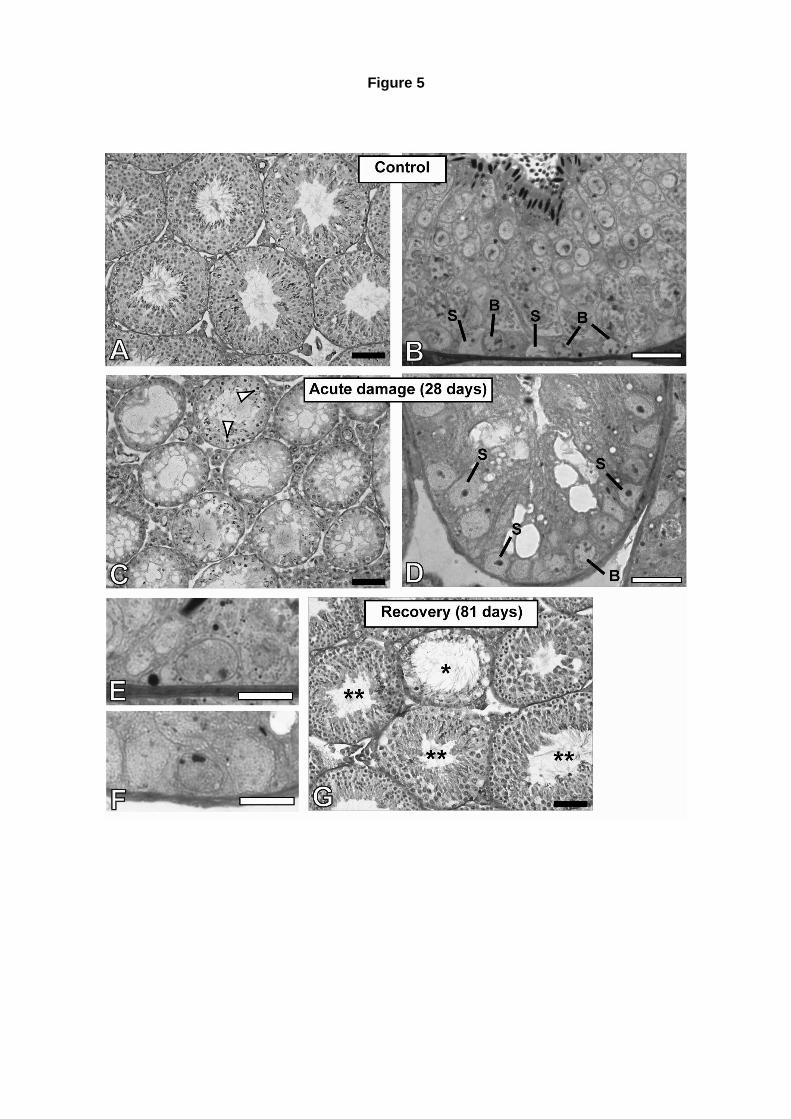
Figure 5
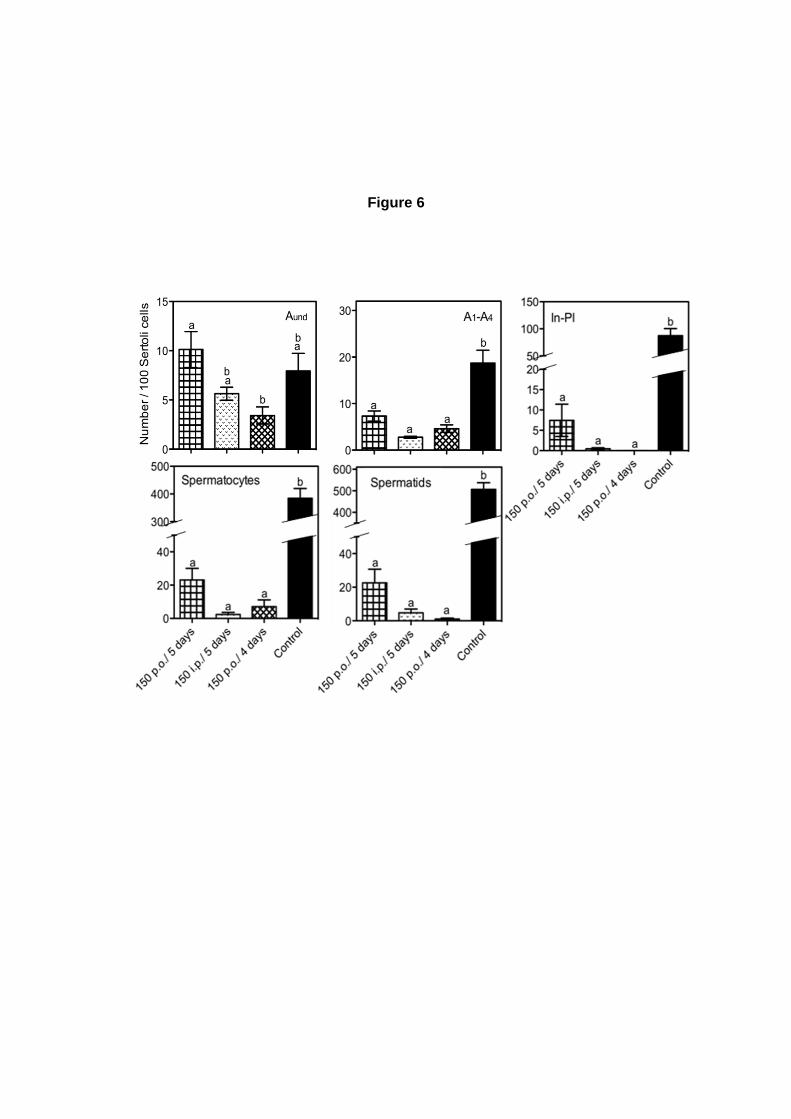
Figure 6
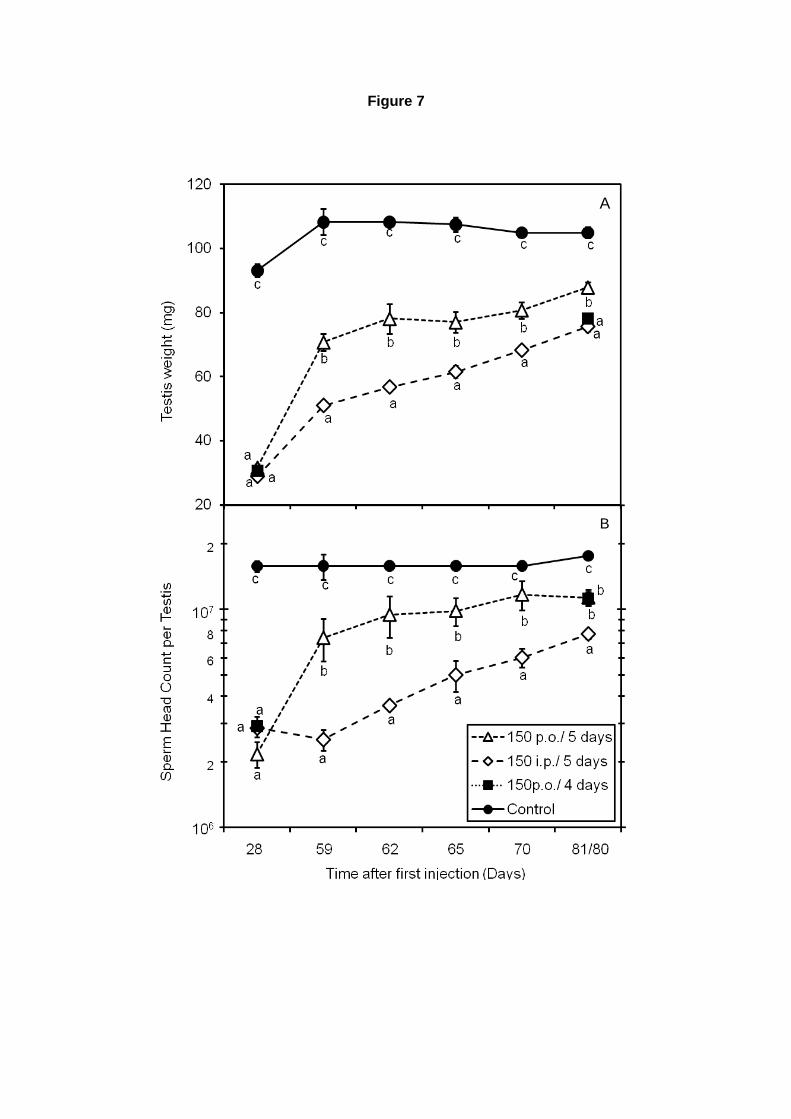
Figure 7
B
A
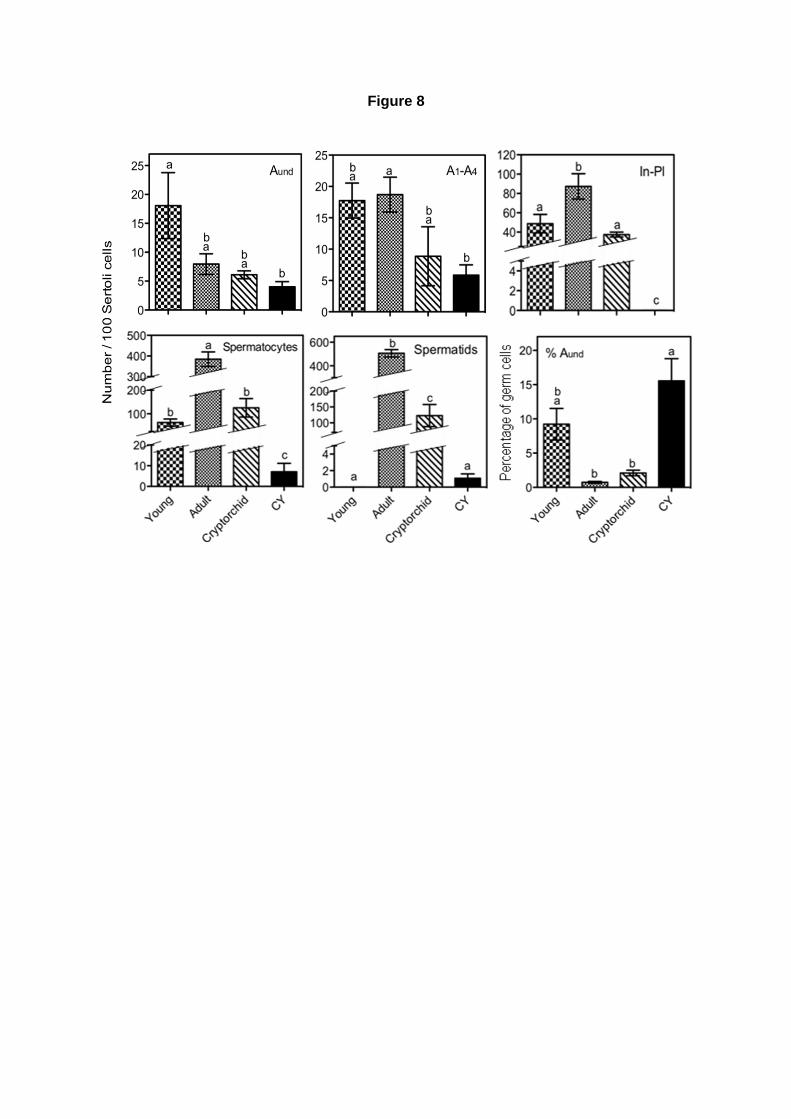
Figure 8
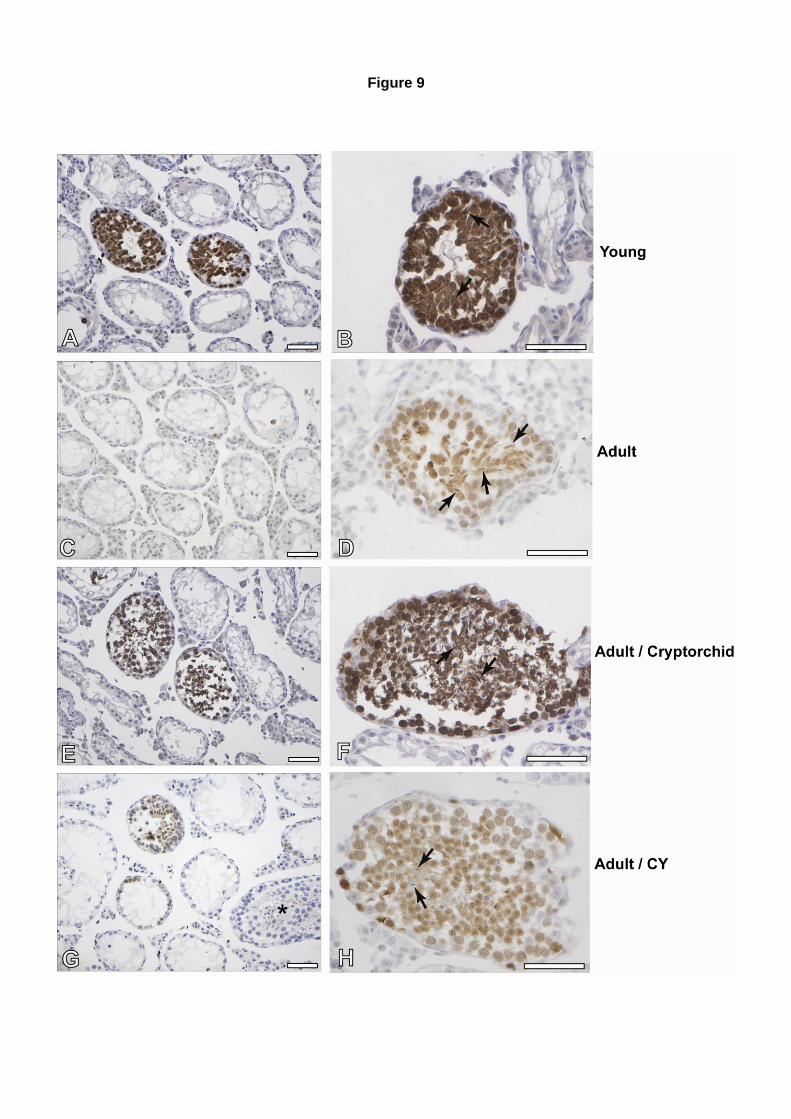
Figure 9
Top Related
Copyright © 2022 FDOKUMEN

