







Bahasa
Halaman
Hukum


A Specific Interaction Between the NBD of the ABC-transporter HlyB and a C-Terminal Fragment of itsTransport Substrate Haemolysin A
Houssain Benabdelhak1†, Stephan Kiontke2†, Carsten Horn2
Robert Ernst2, Mark A. Blight1, I. Barry Holland1 and Lutz Schmitt2*
1Institut de Genetiqueet MicrobiologieBat. 409, Universitede Paris XI, 91405 OrsayFrance
2Institute of BiochemistryBiocenter N210University of FrankfurtMarie-Curie Str. 960439 Frankfurt, Germany
A member of the family of RTX toxins, Escherichia coli haemolysin A,is secreted from Gram-negative bacteria. It carries a C-terminal secretionsignal of approximately 50 residues, targeting the protein to the secretionor translocation complex, in which the ABC-transporter HlyB is a centralelement. We have purified the nucleotide-binding domain of HlyB(HlyB–NBD) and a C-terminal 23 kDa fragment of HlyA plus the His-tag(HlyA1), which contains the secretion sequence. Employing surfaceplasmon resonance, we were able to demonstrate that the HlyB–NBDand HlyA1 interact with a KD of approximately 4 mM. No interactionwas detected between the HlyA fragment and unrelated NBDs, OpuAA,involved in import of osmoprotectants, and human TAP1–NBD, involvedin the export of antigenic peptides. Moreover, a truncated version ofHlyA1, lacking the secretion signal, failed to interact with the HlyB–NBD. In addition, we showed that ATP accelerated the dissociation ofthe HlyB–NBD/HlyA1 complex. Taking these results together, we pro-pose a model for an early stage of initiation of secretion in vivo, in whichthe NBD of HlyB, specifically recognizes the C terminus of the transportsubstrate, HlyA, and where secretion is initiated by subsequent dis-placement of HlyA from HlyB by ATP.
q 2003 Elsevier Science Ltd. All rights reserved
Keywords: ABC-transporters; protein secretion; protein–proteininteraction; surface plasmon resonance*Corresponding author
Introduction
Haemolysin A (HlyA), a 110 kDa toxin, is amember of the RTX (repeats in toxins) family.1
These proteins share repetitive glycine-richsequences, which bind Ca2þ in the folded molecule.HlyA and all other members of this family aresecreted by type 1 secretion systems.2,3 Type 1
secretion is independent of the Sec pathway anddepends on the presence of an uncleaved secretionsignal at the C terminus of the toxin.4 Translocationthrough the cell envelope proceeds without aperiplasmic intermediate, resulting in a one-steptransfer of the HlyA toxin across both membranesof the cell envelope to the medium.5 The secretionsignal is localized in the last 50 to 60 C-terminalamino acid residues of HlyA.6,7 Mutational studieshave indicated a large functional redundancy inthe primary sequence of HlyA and other type 1transport substrates.8 – 13 Mutagenesis, complemen-tation and competition studies resulted in theidentification of an important functional regionextending at least from residues 215 to 246 withrespect to the C terminus. This region, which,although containing very few conserved residues,even compared with close relatives of HlyA,included at least three residues that were appar-ently involved in recognition of the translocator.9
Different genetic studies have provided evidencefor10,13 and against8,11 a specific secondary structure
0022-2836/03/$ - see front matter q 2003 Elsevier Science Ltd. All rights reserved
† H.B. and S.K. contributed equally to this work.Present address: S. Kiontke, Institute of Physiological
Chemistry, Philipps-University Marburg, Karl-von-Frisch Str. 1, 35033 Marburg, Germany.
E-mail address of the corresponding author:[email protected]
Abbreviations used: ABC, ATP-binding cassette; GST,glutathione-S-transferase; Hly, haemolysin; MFP,membrane fusion protein; NTA, N-nitrilo-triacetic acid;OpuAA, osmoprotectant uptake system A ATPase; pmf,proton motive force; SPR, surface plasmon resonance;RTX, repeat in toxins; RU, resonance unit; TMD,transmembrane domain; TMS, transmembranesegments.
doi:10.1016/S0022-2836(03)00204-3 J. Mol. Biol. (2003) 327, 1169–1179

in the signal region required for secretion. Struc-tural studies have so far failed to resolve thisparadox, leaving open the possibility that thesecretion signal is largely unstructured.14,15
The HlyA translocon is composed of three indis-pensable components: HlyB, an ABC-transporter inthe inner membrane;4,16 HlyD (a membrane fusionprotein (MFP)) anchored in the inner membranebut probably capable of spanning the periplasm;17
and TolC, an outer membrane protein. TolC formsa trimer that elongates into 12 antiparallel helices,remarkably constituting a tunnel across the peri-plasm.18 HlyD and TolC appear to interact, pre-sumably then forming a continuous chamberacross the periplasm to the external medium,19,20
thus forming the translocation pathway. HlyB isa member of the superfamily of ABC-trans-porters.16,21 HlyB contains an N-terminal trans-membrane domain (TMD) with six to eightpredicted transmembrane segments (TMSs)22,23
and a C-terminal ABC or nucleotide bindingdomain (NBD) in the cytoplasm. Although ATPhydrolysis by HlyB is essential for secretion ofHlyA,24 the step that is energized by HlyB isunknown and the proton motive force (pmf)25 isreported to be required for secretion of HlyA, thetransport substrate or allocrite.25
Both the ABC and MFP components of thetranslocon have been implicated in initial recog-nition of the corresponding allocrite.19,20,26
However, the regions involved in recognition,in either the allocrite or the ABC proteins, werenot examined in these previous studies. On theother hand, the short N-terminal, cytoplasmicregion of 50–60 residues of HlyD were shown tobe essential for secretion of HlyA.17 Balakrishnanet al. have shown that this region of HlyD isimplicated in binding to an unidentified regionof HlyA, apparently in order to trigger func-tional recruitment of TolC to the translocationcomplex.20
Koronakis et al. reported partial inhibition ofthe activity of the ATPase activity of a purifiedGST-HlyB–NBD fusion protein after addition ofHlyA.27 We have observed a similar inhibitionwith the His-tagged NBD, when incubated withthe C terminus of HlyA (our unpublished data).In addition, the ATPase activity of the ABC trans-porter, PrtD was reported to be partially inhibitedby the homologous C-terminal peptide.28 Theseresults indicated a possible interaction betweenthe ABC domain and the allocrite.
Here, we describe surface plasmon resonance(SPR) studies with the HlyB–NBD and a C-termi-nal 23 kDa fragment of HlyA. The results demon-strated a specific interaction between the HlyB–NBD and the HlyA C-terminal fragment, perhapsdirectly involving the region that contains thesecretion signal itself. Moreover, the complexformed by HlyB and the HlyA-fragment was dis-sociated rapidly in the presence of ATP. Thepossible implications of these observations arediscussed.
Results
Optimizing conditions for bindingprotein immobilization
A previous study indicated that the ATPaseactivity of a GST-fusion of the HlyB–NBD wasinhibited in the presence of the C-terminal secretionsignal of HlyA.27 Similarly, in this study, the ATPaseactivity of the HlyB-NBD was inhibited by the HlyAC-terminal fragment (data not shown), indicating apossible interaction between the two. In order tostudy such possible interactions, the His-taggedNBD was over-expressed in Escherichia coli andpurified as described.29 The C-terminal 23 kDa frag-ment of HlyA was also over-expressed as anN-terminal His-tagged fusion protein. This 23 kDapolypeptide was obtained as inclusion bodies, solu-bilized in guanidine hydrochloride, purified andrefolded in soluble form as described in Materialsand Methods. The purified fragment cross-reactedwith antibodies to HlyA and to the His-tag (data notshown). The identity of the fragments was verifiedby high-resolution mass spectrometry (MALDI-TOF). The measured molecular mass of 24,719.2 Daagreed within experimental error with the theoreticalmass of 24,719 Da and confirmed that this C-terminalHlyA fragment is composed of residues 807–1023 ofHlyA including the His-tag. In the following, werefer to this fragment as HlyA1.
In order to measure a possible interactionbetween the HlyB–NBD and the C-terminalfragment of HlyA, we employed SPR.30 We usedthe well-established strategy for the immobiliz-ation of either protein via the formation of a His-tag/NiNTA complex.31 To minimize non-specificinteraction of the proteins with the NTA-present-ing interface, a variety of such surfaces wasinvestigated.32 – 35 Non-detectable levels of non-specific interactions were obtained only with theNTA-dextran matrix.
The HlyB–NBD or the HlyA fragments wereimmobilized independently after a pre-loadingstep with the NTA-dextran surface via the N-termi-nal His-tag. We observed that the quantitativebehavior of the captured HlyB–NBD differedfrom that of HlyA1. The half-time of dissociationfor HlyA1 was more than ten hours, whilst thehalf-time of the immobilized HlyB–NBD wasaround 30 minutes (data not shown). Due to themore stable binding property of HlyA1, the proteinwas the preferred choice for immobilization via theHis-tag. However, all experiments were repeatedwith the HlyB–NBD being the first to be immobi-lized and identical results were obtained. In thefollowing results, only one example from thisreciprocal approach is usually shown.
The HlyB–NBD and HlyA form aspecific complex
After stable immobilization of HlyA1 via theN-terminal His-tag, various concentrations of the
1170 SPR Analysis of the HlyA/HlyB-NBD Interaction
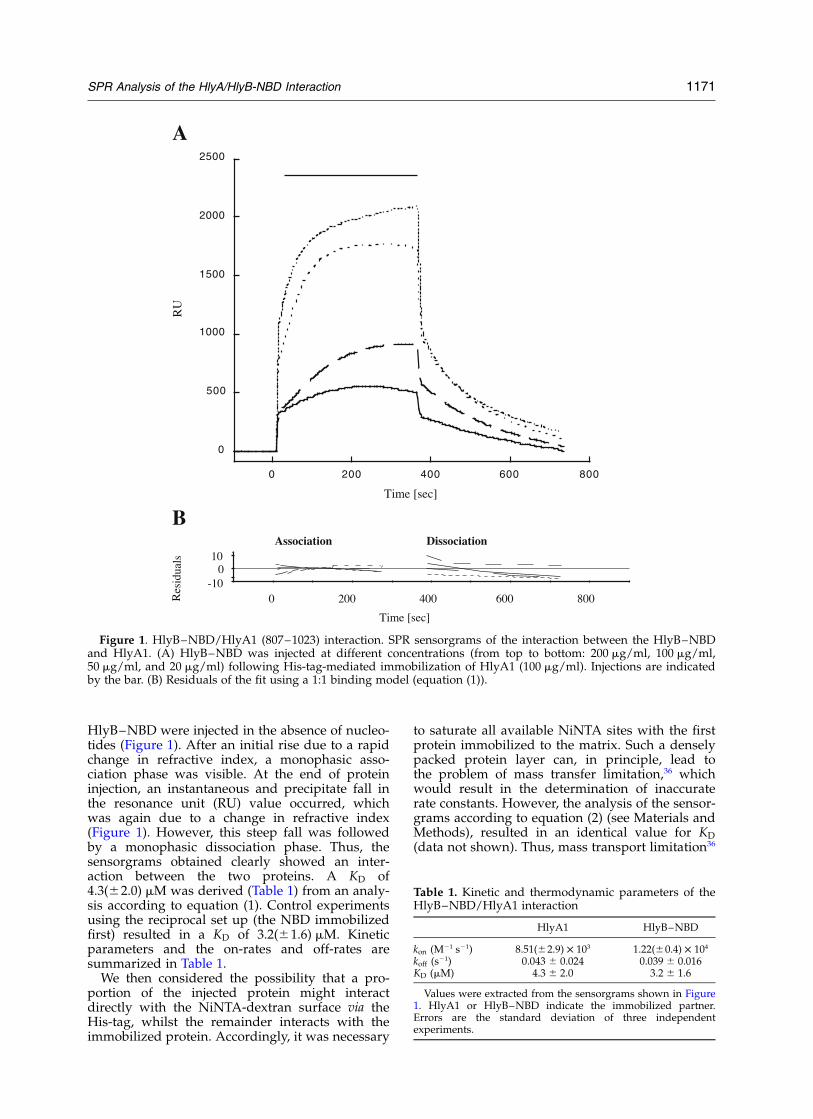
HlyB–NBD were injected in the absence of nucleo-tides (Figure 1). After an initial rise due to a rapidchange in refractive index, a monophasic asso-ciation phase was visible. At the end of proteininjection, an instantaneous and precipitate fall inthe resonance unit (RU) value occurred, whichwas again due to a change in refractive index(Figure 1). However, this steep fall was followedby a monophasic dissociation phase. Thus, thesensorgrams obtained clearly showed an inter-action between the two proteins. A KD of4.3(^2.0) mM was derived (Table 1) from an analy-sis according to equation (1). Control experimentsusing the reciprocal set up (the NBD immobilizedfirst) resulted in a KD of 3.2(^1.6) mM. Kineticparameters and the on-rates and off-rates aresummarized in Table 1.
We then considered the possibility that a pro-portion of the injected protein might interactdirectly with the NiNTA-dextran surface via theHis-tag, whilst the remainder interacts with theimmobilized protein. Accordingly, it was necessary
to saturate all available NiNTA sites with the firstprotein immobilized to the matrix. Such a denselypacked protein layer can, in principle, lead tothe problem of mass transfer limitation,36 whichwould result in the determination of inaccuraterate constants. However, the analysis of the sensor-grams according to equation (2) (see Materials andMethods), resulted in an identical value for KD
(data not shown). Thus, mass transport limitation36
Figure 1. HlyB–NBD/HlyA1 (807–1023) interaction. SPR sensorgrams of the interaction between the HlyB–NBDand HlyA1. (A) HlyB–NBD was injected at different concentrations (from top to bottom: 200 mg/ml, 100 mg/ml,50 mg/ml, and 20 mg/ml) following His-tag-mediated immobilization of HlyA1 (100 mg/ml). Injections are indicatedby the bar. (B) Residuals of the fit using a 1:1 binding model (equation (1)).
Table 1. Kinetic and thermodynamic parameters of theHlyB–NBD/HlyA1 interaction
HlyA1 HlyB–NBD
kon (M21 s21) 8.51(^2.9) £ 103 1.22(^0.4) £ 104
koff (s21) 0.043 ^ 0.024 0.039 ^ 0.016KD (mM) 4.3 ^ 2.0 3.2 ^ 1.6
Values were extracted from the sensorgrams shown in Figure1. HlyA1 or HlyB–NBD indicate the immobilized partner.Errors are the standard deviation of three independentexperiments.
SPR Analysis of the HlyA/HlyB-NBD Interaction 1171
does not influence the measurements. Additionally,the residuals of the fits (Figure 1(B)) fit to theapplied 1:1 binding model.
The results described above indicated thatthe C-terminal 23 kDa fragment of HlyA, whichcontains the secretion signal used in vivo,9 and theHlyB–NBD do interact. This interaction occurs inthe complete absence of the membrane domain ofHlyB. Thus, this is the first time that clear evidencefor a specific interaction between an NBD andits transport substrate has been obtained. Theseresults, however, do not preclude a further func-tional interaction of HlyA at some point in thesecretion process with the membrane domain ofHlyB or the HlyD protein.
The NBD–HlyA interaction occurs in a matrix-free phase
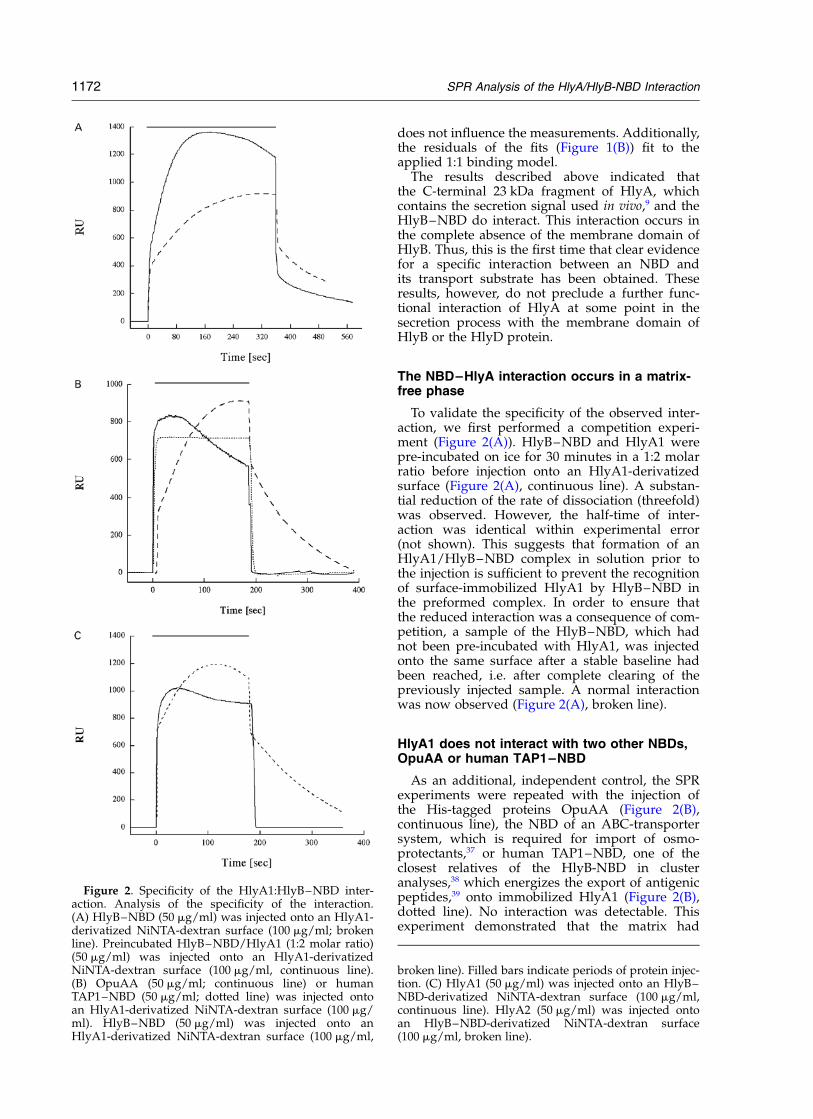
To validate the specificity of the observed inter-action, we first performed a competition experi-ment (Figure 2(A)). HlyB–NBD and HlyA1 werepre-incubated on ice for 30 minutes in a 1:2 molarratio before injection onto an HlyA1-derivatizedsurface (Figure 2(A), continuous line). A substan-tial reduction of the rate of dissociation (threefold)was observed. However, the half-time of inter-action was identical within experimental error(not shown). This suggests that formation of anHlyA1/HlyB–NBD complex in solution prior tothe injection is sufficient to prevent the recognitionof surface-immobilized HlyA1 by HlyB–NBD inthe preformed complex. In order to ensure thatthe reduced interaction was a consequence of com-petition, a sample of the HlyB–NBD, which hadnot been pre-incubated with HlyA1, was injectedonto the same surface after a stable baseline hadbeen reached, i.e. after complete clearing of thepreviously injected sample. A normal interactionwas now observed (Figure 2(A), broken line).
HlyA1 does not interact with two other NBDs,OpuAA or human TAP1–NBD
As an additional, independent control, the SPRexperiments were repeated with the injection ofthe His-tagged proteins OpuAA (Figure 2(B),continuous line), the NBD of an ABC-transportersystem, which is required for import of osmo-protectants,37 or human TAP1–NBD, one of theclosest relatives of the HlyB-NBD in clusteranalyses,38 which energizes the export of antigenicpeptides,39 onto immobilized HlyA1 (Figure 2(B),dotted line). No interaction was detectable. Thisexperiment demonstrated that the matrix had
Figure 2. Specificity of the HlyA1:HlyB–NBD inter-action. Analysis of the specificity of the interaction.(A) HlyB–NBD (50 mg/ml) was injected onto an HlyA1-derivatized NiNTA-dextran surface (100 mg/ml; brokenline). Preincubated HlyB–NBD/HlyA1 (1:2 molar ratio)(50 mg/ml) was injected onto an HlyA1-derivatizedNiNTA-dextran surface (100 mg/ml, continuous line).(B) OpuAA (50 mg/ml; continuous line) or humanTAP1–NBD (50 mg/ml; dotted line) was injected ontoan HlyA1-derivatized NiNTA-dextran surface (100 mg/ml). HlyB–NBD (50 mg/ml) was injected onto anHlyA1-derivatized NiNTA-dextran surface (100 mg/ml,
broken line). Filled bars indicate periods of protein injec-tion. (C) HlyA1 (50 mg/ml) was injected onto an HlyB–NBD-derivatized NiNTA-dextran surface (100 mg/ml,continuous line). HlyA2 (50 mg/ml) was injected ontoan HlyB–NBD-derivatized NiNTA-dextran surface(100 mg/ml, broken line).
1172 SPR Analysis of the HlyA/HlyB-NBD Interaction
been completely saturated with the first protein,preventing non-specific binding of the secondprotein to the NiNTA-surfaces via its own His-tag.Figure 2 shows that, following the passage ofOpuAA or human TAP1–NBD over the matrixcontaining the immobilized HlyA1, when theHlyB–NBD was subsequently injected onto thesame matrix, a specific interaction between HlyA1and the HlyB–NBD was again detected (Figure2(B), broken line).
HlyA1 lacking the C-terminal 57 residues doesnot interact with the NBD
As a further test of the authenticity of the HlyB–HlyA interaction, an independent experiment wasperformed with truncated forms of the C-terminalHlyA fragment, produced during the over-expression of the HlyA1 fragment, as spontaneousdegradation products of the A1 fragment (seeMaterials and Methods). This material, designatedHlyA2, was purified by immobilized metal ion-affinity chromatography (IMAC) as predominantlyan 18 kDa peptide. Mass spectrometry indicatedthat the major component constituted at least 80%of the mass, corresponding to residues 807–966 ofHlyA (17,738.1 Da) and was therefore lackingthe last 57 residues of the HlyA molecule. Such amolecule lacking the secretion signal would notbe secreted in vivo.25,40 When this material wasinjected onto an NiNTA surface containing theimmobilized HlyB–NBD, no protein–protein inter-action was observed (Figure 2(C), continuous line).In complete contrast, when a sample of the HlyA1protein, which contains the secretion signal (Figure2(C), broken line), was subsequently passed overthe same matrix, an interaction was observed.This result again demonstrates that under theseconditions, where the HlyB–NBD is present as adensely packed layer on the matrix, non-specificbinding via the His-tag to an HlyA fragment isprevented. More importantly, this experiment pro-vides the first indication that the site of interactionof the HlyB–NBD may specifically include theregion of HlyA containing the C-terminal secretionsignal used in vivo.
Nucleotides can disrupt the HlyA1–HlyB complex
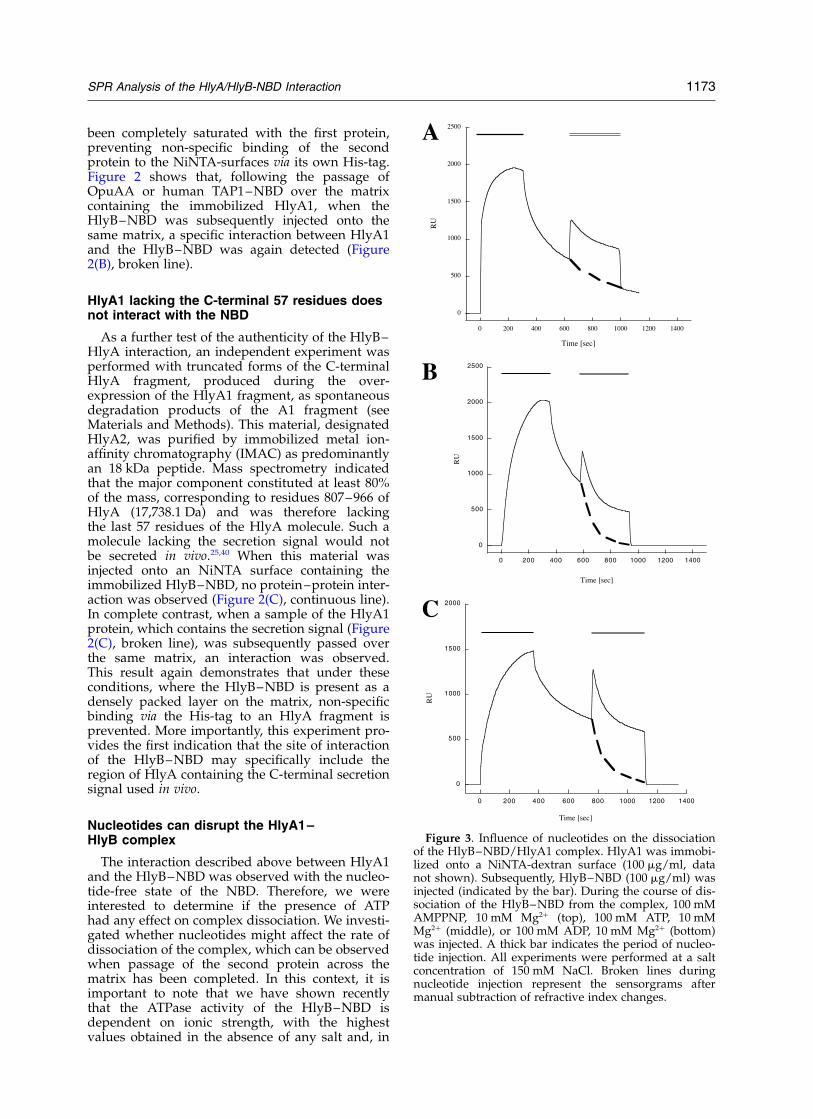
The interaction described above between HlyA1and the HlyB–NBD was observed with the nucleo-tide-free state of the NBD. Therefore, we wereinterested to determine if the presence of ATPhad any effect on complex dissociation. We investi-gated whether nucleotides might affect the rate ofdissociation of the complex, which can be observedwhen passage of the second protein across thematrix has been completed. In this context, it isimportant to note that we have shown recentlythat the ATPase activity of the HlyB–NBD isdependent on ionic strength, with the highestvalues obtained in the absence of any salt and, in
Figure 3. Influence of nucleotides on the dissociationof the HlyB–NBD/HlyA1 complex. HlyA1 was immobi-lized onto a NiNTA-dextran surface (100 mg/ml, datanot shown). Subsequently, HlyB–NBD (100 mg/ml) wasinjected (indicated by the bar). During the course of dis-sociation of the HlyB–NBD from the complex, 100 mMAMPPNP, 10 mM Mg2þ (top), 100 mM ATP, 10 mMMg2þ (middle), or 100 mM ADP, 10 mM Mg2þ (bottom)was injected. A thick bar indicates the period of nucleo-tide injection. All experiments were performed at a saltconcentration of 150 mM NaCl. Broken lines duringnucleotide injection represent the sensorgrams aftermanual subtraction of refractive index changes.
SPR Analysis of the HlyA/HlyB-NBD Interaction 1173

contrast, no detectable steady-state ATPase activityabove 300 mM NaCl. This reversible loss of activityat high concentrations of salt was accompanied byreduced binding of 8-azido-ATP.41
Therefore, we analyzed the influence of nucleo-tides (AMPPNP, ATP, and ADP) on the dissociationbehavior of the HlyB–NBD/HlyA1 complex atthree different ionic strengths, 50 mM, 150 mM,and 300 mM NaCl. Lower concentrations of salt insuch experiments were precluded, since no stablebaseline could be obtained. As a first set of experi-ments, HlyB–NBD was injected on to an HlyA1-derivatized surface in the presence of 5 mM ATP,2 mM Mg2þ at salt concentrations of 150 mM NaCl(data not shown). These experiments demonstratedthat the presence of ATP reduced the interactionbetween the proteins. However, we were not ableto inhibit the interaction completely. This is likelydue to the presence of a mixed population ofNBD molecules (nucleotide-free and ATP-bound)present in the initial injected sample. Thus, thenucleotide-free form would still be able to interactwith the immobilized HlyA1. Therefore, in orderto test the effect of nucleotides on the binding toHlyA more specifically, we changed our experi-mental set-up and injected nucleotides at highconcentration (100 mM), but only after nucleotide-free HlyB–NBD was injected onto an HlyA1-derivatized surface. The experiments in Figure 3were carried out with 150 mM NaCl. Injection ofAMPPNP (Figure 3, top panel), even at the veryhigh concentration of 100 mM, had no influenceon the dissociation rate of the HlyB–NBD/HlyA1complex, since the RU value reached afterAMPPNP injection was identical with thatobtained in the absence of AMPPNP (as indicatedby the broken line during AMPPNP injection).This result also suggested that ATP hydrolysiswas essential for disruption of the complex, atleast under these conditions. Indeed, the situationwas quite different for ATP (Figure 3, middlepanel) and ADP (Figure 3, bottom panel), sinceaddition of either nucleotide resulted in animmediate, faster dissociation phase. At the endof nucleotide injection, no HlyB–NBD/HlyA1complex remained. From these sensorgrams,an acceleration factor of 12(^2) and 11(^2),respectively, was determined for ATP and ADP(Table 2). At 300 mM NaCl (sensorgrams notshown), injections of AMPPNP or ATP, even atconcentrations of 100 mM, during the HlyB–NBDdissociation phase had no detectable influence onthe off-rate, consistent with greatly reduced bindingof 8-azido-ATP under these conditions.41 In contrastto this, all three nucleotides promoted a roughly 15-fold faster dissociation rate at 50 mM NaCl, wherethe ATPase is most active. Thus, under these con-ditions, the fixation of ATP, presumably inducing aconformational change in the HlyB–NBD,27 was suf-ficient to promote rapid displacement of HlyA1 fromthe complex (see Discussion).
It is important to note that we chose high con-centrations of nucleotides (100 mM) in the above
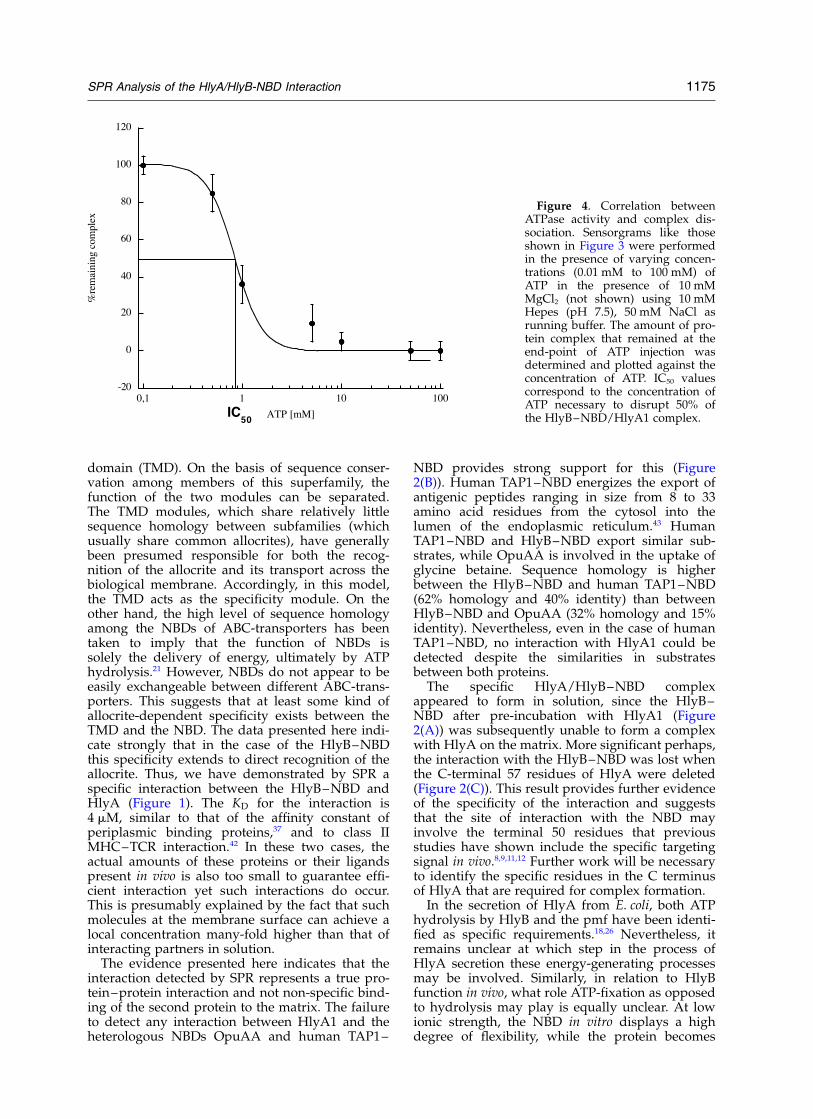
experiments in order to instantaneously saturateall nucleotide-binding sites of the NBD and there-fore to preclude the presence of a mixed popu-lation of the NBD, e.g. nucleotide-loaded andnucleotide-free protein. However, at 300 mMNaCl, neither 100 mM AMPPNP nor 100 mM ATPhad any effect on the dissociation behavior. Thesame observation was made for 100 mM AMPPNPat 150 mM NaCl. This demonstrates that the highconcentration of nucleotides employed did notimpose any non-specific, electrostatic effects onthe stability of the complex. Nevertheless, weinvestigated the dependence of complex dis-sociation on the concentration of the applied ATPat 50 mM and 150 mM NaCl (Table 2 and Figure4). The amount of HlyA1/HlyB–NBD complexremaining after injection of varying amounts ofATP/Mg2þ, 10 mM to 100 mM ATP, with a constantconcentration of 10 mM Mg2þ (this alone had noeffect on dissociation of the complex), was deter-mined (Table 2 and Figure 4). The amount ofcomplex remaining versus concentration of ATPresulted in a sigmoidal dependence (Figure 4),with disruption of the complex by ATP concen-trations observed in the physiological range. TheIC50 value was 2.3(^1.2) mM at 150 mM NaCl and0.8(^0.4) mM at 50 mM NaCl (Table 2). Here, theIC50 value is defined as the concentration of nucleo-tide necessary to disrupt 50% of the HlyB–NBD/HlyA1 complex. The situation at 300 mM NaClwas not analyzed, since neither AMPPNP norATP had a detectable influence on complex dis-sociation. It may be noted that the KM values ofHlyB–NBD and the determined IC50 values areclosely related under these conditions (Table 2).Importantly, the results do show clearly that the highexcess of nucleotide used in some experiments doesnot create an artificial disruption of the complex.
Discussion
ABC-transporters consist of two duplicated mod-ules, the cytoplasmic NBD and the transmembrane
Table 2. Summary of the influence of ionic strength andnucleotides (in the presence of 10 mM Mg2þ) on the dis-sociation of the HlyB–NBD–HlyA1 complex and thecorrelation between ATPase activity of the HlyB–NBDand IC50 values
Acceleration factor[NaCl](mM)
KM
(mM)a
IC50
(mM) AMPPNP ATP ADP
50 0.9 ^ 0.1 0.8 ^ 0.2 14 ^ 2 13 ^ 2 17 ^ 2150 3.2 ^ 0.2 2.3 ^ 0.7 1 12 ^ 2 11 ^ 2300 ND NM 1 1 13 ^ 2
Acceleration factors are defined as the ratio of the off-rate inthe presence of the corresponding nucleotides ðkoff;nucleotideÞ andthe off-rate in the absence of nucleotides ðkoffÞ: koff;nucleotide wasdetermined from the parts of the sensorgrams shown as brokenlines in Figure 3. ND, not detectable; NM, not measured.
a Taken from Benabdelhak et al.41
1174 SPR Analysis of the HlyA/HlyB-NBD Interaction
domain (TMD). On the basis of sequence conser-vation among members of this superfamily, thefunction of the two modules can be separated.The TMD modules, which share relatively littlesequence homology between subfamilies (whichusually share common allocrites), have generallybeen presumed responsible for both the recog-nition of the allocrite and its transport across thebiological membrane. Accordingly, in this model,the TMD acts as the specificity module. On theother hand, the high level of sequence homologyamong the NBDs of ABC-transporters has beentaken to imply that the function of NBDs issolely the delivery of energy, ultimately by ATPhydrolysis.21 However, NBDs do not appear to beeasily exchangeable between different ABC-trans-porters. This suggests that at least some kind ofallocrite-dependent specificity exists between theTMD and the NBD. The data presented here indi-cate strongly that in the case of the HlyB–NBDthis specificity extends to direct recognition of theallocrite. Thus, we have demonstrated by SPR aspecific interaction between the HlyB–NBD andHlyA (Figure 1). The KD for the interaction is4 mM, similar to that of the affinity constant ofperiplasmic binding proteins,37 and to class IIMHC–TCR interaction.42 In these two cases, theactual amounts of these proteins or their ligandspresent in vivo is also too small to guarantee effi-cient interaction yet such interactions do occur.This is presumably explained by the fact that suchmolecules at the membrane surface can achieve alocal concentration many-fold higher than that ofinteracting partners in solution.
The evidence presented here indicates that theinteraction detected by SPR represents a true pro-tein–protein interaction and not non-specific bind-ing of the second protein to the matrix. The failureto detect any interaction between HlyA1 and theheterologous NBDs OpuAA and human TAP1–
NBD provides strong support for this (Figure2(B)). Human TAP1–NBD energizes the export ofantigenic peptides ranging in size from 8 to 33amino acid residues from the cytosol into thelumen of the endoplasmic reticulum.43 HumanTAP1–NBD and HlyB–NBD export similar sub-strates, while OpuAA is involved in the uptake ofglycine betaine. Sequence homology is higherbetween the HlyB–NBD and human TAP1–NBD(62% homology and 40% identity) than betweenHlyB–NBD and OpuAA (32% homology and 15%identity). Nevertheless, even in the case of humanTAP1–NBD, no interaction with HlyA1 could bedetected despite the similarities in substratesbetween both proteins.
The specific HlyA/HlyB–NBD complexappeared to form in solution, since the HlyB–NBD after pre-incubation with HlyA1 (Figure2(A)) was subsequently unable to form a complexwith HlyA on the matrix. More significant perhaps,the interaction with the HlyB–NBD was lost whenthe C-terminal 57 residues of HlyA were deleted(Figure 2(C)). This result provides further evidenceof the specificity of the interaction and suggeststhat the site of interaction with the NBD mayinvolve the terminal 50 residues that previousstudies have shown include the specific targetingsignal in vivo.8,9,11,12 Further work will be necessaryto identify the specific residues in the C terminusof HlyA that are required for complex formation.
In the secretion of HlyA from E. coli, both ATPhydrolysis by HlyB and the pmf have been identi-fied as specific requirements.18,26 Nevertheless, itremains unclear at which step in the process ofHlyA secretion these energy-generating processesmay be involved. Similarly, in relation to HlyBfunction in vivo, what role ATP-fixation as opposedto hydrolysis may play is equally unclear. At lowionic strength, the NBD in vitro displays a highdegree of flexibility, while the protein becomes
Figure 4. Correlation betweenATPase activity and complex dis-sociation. Sensorgrams like thoseshown in Figure 3 were performedin the presence of varying concen-trations (0.01 mM to 100 mM) ofATP in the presence of 10 mMMgCl2 (not shown) using 10 mMHepes (pH 7.5), 50 mM NaCl asrunning buffer. The amount of pro-tein complex that remained at theend-point of ATP injection wasdetermined and plotted against theconcentration of ATP. IC50 valuescorrespond to the concentration ofATP necessary to disrupt 50% ofthe HlyB–NBD/HlyA1 complex.
SPR Analysis of the HlyA/HlyB-NBD Interaction 1175

more rigid at higher ionic strength, parallelingreduced binding of ATP and activity.41 The resultsshown in Table 2 demonstrate that ATP fixation atlow ionic strength (50 mM NaCl) is sufficient toaccelerate complex dissociation, without hydroly-sis. However, we have observed that at 150 mMNaCl, AMPPNP, unlike ATP, is not able to promotedissociation of the complex (Figure 3 and Table 2).At 300 mM NaCl, only ADP is sufficient to acceler-ate complex dissociation. Thus, caution is requiredin concluding that ATP binding alone may be suf-ficient, particularly in vivo, to disrupt the complex.Nevertheless, the observed acceleration of theoff-rates of the HlyB–NBD/HlyA1 complex(Table 2) implies a relation between hydrolysisand dis-sociation.
The results in this study clearly differ from theevidence, well established, of specific binding sitesin the TMD of multidrug transporters such asPgp (Mdr1), where drug molecules stimulate ATPhydrolysis by binding to specific sites in theTMD.44 In this study, although without stimulatingATPase activity, rather the converse (data notshown),27,28 we find a specific interaction betweenthe NBD and HlyA. Consequently, what are theimplications in relation to the function of HlyB?First, ABC-transporters such as Mdr1, whichexport hydrophobic drugs directly from the innerleaflet of the membrane bilayer, perhaps not sur-prisingly have a specific drug-binding site withinthe membrane domain, which, when loaded,appears to communicate with the NBD.45 For thetotally different allocrites of the type 1 secretionsystem, large polypeptides, often with extensive
hydrophilic regions, there is no evidence of initialbinding to the bilayer and we must presume thatthey approach the transport channel from the cyto-plasm. It would be reasonable to anticipate that thetransport of such molecules may involve quitedifferent mechanisms of allocrite recognition andcorrespondingly some different features in inter-and intramolecular signalling processes. In Figure5, we have summarized our data and propose amodel, in which the HlyB–NBD docks specificallywith HlyA, involving a binding site located veryclose to, if not within, the region covered by theC-terminal secretion signal. The stoichiometry ofthe HlyA–HlyB–NBD complex detected in vitrohas not been established clearly, but preliminarydata indicate a 1:1 stoichiometry in the nucleotide-free state (data not shown). In vivo, a random bind-ing mode for HlyA or ATP to HlyB is possible.However, we propose that ATP binding and/orhydrolysis through the induction of conforma-tional changes24,27 and/or dimerization41 to acomplex already containing HlyA, will result incomplex dissociation and initiation of secretion bydisplacement of HlyA towards the interior of thetranslocator. In order to achieve this directionalbias, in the movement of the displaced HlyAmolecule, both the membrane domain of HlyBand certainly the N-terminal of HlyD,17,19,20 whichis implicated in the early stages of HlyA secretion,are likely to be involved.
Materials and Methods
Construction of clones and protein expression
The HlyB–NBD corresponding to residues 467–707with an N-terminal His tag, was prepared and purifiedby Ni2þ-affinity chromatography as described.29 Thepurification of OpuAA will be described elsewhere. TheNBD of human TAP1 was purified as reported.39
The hlyA DNA fragment encoding the residues N807–A1023 was generated as an Nhe I–Pae I fragment usingPCR and cloned under the control of an arabinose-inducible pBAD promoter.46 A His6 tag was engineeredinto the amino terminus of the HlyA-fragment by incor-poration of the corresponding nucleotide sequencedownstream of the Nhe I site. Cultures were grown at37 8C until an absorbance at 600 nm of 1.0 and inducedby the addition of arabinose (final concentration 4 mM)and harvested after 90 minutes.
Purification of HlyA1 and the truncated C-terminalfragment, HlyA2
Harvested cells were washed in buffer A (10 mMTris–HCl (pH 8.0), 100 mM KCl, 1 mM PMSF, 0.2 mMEDTA, 10% (v/v) glycerol) and resuspended in thesame buffer. Cells were broken by four passages througha French pressure cell at 1000 psi (1 psi < 6.9 kPa).Unbroken cells, cell debris, and inclusion bodies(HlyA1), constituting 95% of the induced protein, werecollected. The remaining protein was present as a solublebut truncated form (HlyA2). The pellet of inclusionbodies, containing what we designate as HlyA1, was
Figure 5. Proposed model for the initial recognitionof the HlyB–NBD by HlyA via its secretion signal andsubsequent initiation of the transport process. HlyA isshown as open diamond, HlyB–NBD as a box. Theoligomeric state of the HlyB–NBD during the differentfunctional states has been assumed to be identical withthe oligomeric states of NBDs and full-length ABC-transporters, for which an X-ray structure has beenreported.39,47 – 51 Since conformational changes and theoligomeric state of the TMDs of HlyB are not known,the TMDs have been excluded from our model. Forsimplicity, possible interactions of HlyA with the cyto-solic domain of HlyD are not included. For furtherdetails, see the text.
1176 SPR Analysis of the HlyA/HlyB-NBD Interaction

resuspended in 25 ml of buffer A supplemented with6 M guanidine–HCl and dialyzed exhaustively againstbuffer A plus PMSF (1 mM) and EDTA (0.2 mM). Thisallowed quantitative recovery of a soluble, refoldedform. Refolded HlyA1 and the cytosolic soluble truncate,HlyA2, were loaded onto separate HiTrap chelatingcolumns in buffer A plus 10 mM imidazole (Pharmacia;5 ml bed volume). Protein was eluted by a lineargradient of 10 mM–350 mM imidazole. The fractionscontaining the HlyA fragments were pooled, concen-trated and stored at 280 8C. Analysis by SDS-PAGErevealed bands of 25 kDa and 18 kDa (with smalleramounts of two additional bands) for HlyA1 andHlyA2, respectively. Western blotting confirmed thatHlyA1 and the three bands of HlyA2 reacted with anti-bodies to both HlyA and the His-tag (data not shown).This indicates that the HlyA2 fragments were truncateddegradation products of the HlyA1 fragment. Mass spec-trometry indicated that the 18 kDa fragment contributedat least 80% of the mass of the HlyA2 preparation andcorresponds to residues 807–966 of HlyA. This estimatewas in line with an SDS-PAGE of HlyA2 (gel notshown). The measured molecular mass of 17,738.1 Daagreed with the expected, calculated mass of17,736.4 Da. This therefore corresponds to a fragment ofHlyA, which lacks the last C-terminal 57 residues. Sucha deletion would in vivo be sufficient to abolish secretionof HlyA.25,40 The second major fragment contributedapproximately 15% of the mass, and apparently lacked14 residues from the C terminus, sufficient to reducesecretion levels in vivo at least by fivefold.12 The greatmajority of the molecules in the HlyA2 preparationtherefore lacked a functional secretion signal. Mass spec-trometry confirmed the expected molecular mass of24,719.2 Da for HlyA1, corresponding to residues 807–1023 of HlyA and therefore with the intact signalsequence.
Mass spectrometry
Measurements were performed with a PerSeptivebiosystems Voyager mass spectrometer (PerSeptive Bio-systems). All proteins (100 mg/ml) were dialyzed against10 mM Tris–HCl (pH 8.0) prior to the measurements.
Surface plasmon resonance
All experiments were performed with a BIAcore X(BIAcore, Freiburg, Germany) at 22(^1) 8C, with a con-stant flow-rate of 10 ml/minute. Buffer B (10 mM Hepes(pH 7.5), 150 mM NaCl) was used as running buffer,except where indicated. Proteins were diluted from astock solution (1–2 mg/ml) in 10 mM phosphate (pH8.0), 100 mM KCl (HlyB–NBD), 10 mM phosphate (pH7.6), 150 mM NaCl (OpuAA), 10 mM Tris–HCl (pH 8.0),100 mM KCl, 10% glycerol (HlyA1 and A2) or 10 mMphosphate (pH 8.0), 150 mM NaCl, 10% glycerol(TAP1–NBD) into buffer B. All sensorgrams presentedwere not corrected for background, i.e. sensorgrams inthe absence of immobilized protein were not subtracted.
To determine the influence of ionic strength on com-plex dissociation with respect to the functional state ofthe NBD, different ionic strengths were used. Therefore,the salt concentration of buffer B was changed to 50 mMor 300 mM NaCl. All other parameters were keptconstant. 100 mM nucleotides and 10 mM MgCl2 weredissolved in 10 mM Tris–HCl at the appropriate salt
concentration and pH adjusted with concentratedNaOH to pH 7.5.
Data analysis
All sensorgrams were analyzed using the BiaEvalua-tion software version 3.1. A 1:1 binding model wasassumed for data analysis by a global fitting routine.Here, kon and koff were fitted simultaneously accordingto equation (1):
d½B�
dt¼ 2ðkon½A�½B�2 koff½AB�Þ;B½0� ¼ 0
d½AB�
dt¼ ðkon½A�½B�2 koff½AB�Þ;AB½0� ¼ 0
ð1Þ
Here, [A] denotes the injected analyte, [B] denotes thecaptured protein, [0] indicates time zero, and kon andkoff are the intrinsic rate constants of the association anddissociation reaction, respectively. In the experiments,densely packed protein surfaces were prepared to avoidinteraction of the injected analyte with the surface, sinceall proteins contained a His-tag. Under these conditions,the analysis can become dependent on mass transferlimitations.36 Under such circumstances, the determinedassociation rate constant reflects the diffusion of theanalyte to the captured receptor, which becomes therate-limiting step and not the investigated recognitionprocess. However, if mass transfer limitations do apply,actual affinity constants can still be derived from theaffinity model:36
Req ¼Rmax
KD
½A�þ 1
ð2Þ
Here, Req denotes the equilibrium RU value, Rmax themaximal RU value at a given concentration of analyteA, [A] the concentration of injected analyte, and KD thedissociation constant.
Acknowledgements
We thank Jorg Kahnt and Silke Hutschenreiterfor assistance in mass spectrometry, Suman Latafor stimulating discussions and critical reading ofthe manuscript, and Robert Tampe for constantencouragement. We are indebted to Chris van derDoes for many fruitful discussions and criticalreading of the manuscript. H. B. acknowledges thesupport of FRM (Fondation pour la RechercheMedicale) and the Societe de Secours des Amisdes Sciences. The support of CNRS and theUniversite Paris-Sud is gratefully acknowledged.The Deutsche Forschungsgemeinschaft supportedthis work (Emmy Noether program, grantSchm1279/2-1 to L.S.).
References
1. Welch, R. A. (1991). Pore-forming cytolysins ofGram-negative bacteria. Mol. Microbiol. 5, 521–528.
SPR Analysis of the HlyA/HlyB-NBD Interaction 1177

2. Thanassi, D. G. & Hultgren, S. J. (2000). Multiplepathways allow protein secretion across the bacterialouter membrane. Curr. Opin. Cell Biol. 12, 420–430.
3. Buchanan, S. K. (2001). Type I secretion and multi-drug efflux: transport through the TolC channel-tunnel. Trends Biochem. Sci. 26, 3–6.
4. Holland, I. B. & Blight, M. A. (1996). Structure andfunction of HlyB, the ABC-transporter essential forhaemolysin secretion from Escherichia coli. In Hand-book of Biological Physics (Konings, W. N., Kaback,H. R. & Lolkema, J. S., eds), 2, pp. 111–135, NorthHolland, Amsterdam.
5. Blight, M. A. & Holland, I. B. (1994). Heterologousprotein secretion and the versatile Eschericichia colihaemolysin translocator. Trends Biotechnol. 12,450–455.
6. Mackman, N., Baker, K., Gray, L., Haigh, R., Nicaud,J. M. & Holland, I. B. (1987). Release of a chimericprotein into the medium from Escherichia coli usingthe C-terminal secretion signal of haemolysin.EMBO J. 6, 2835–2841.
7. Jarchau, T., Chakraborty, T., Garcia, F. & Goebel, W.(1994). Selection for transport competence ofC-terminal polypeptides derived from Escherichiacoli haemolysin: the shortest peptide capable ofautonomous HlyB/HlyD-dependent secretion com-prises the C-terminal 62 amino acids of HlyA. Mol.Gen. Genet. 245, 53–60.
8. Kenny, B., Taylor, S. & Holland, I. B. (1992). Identifi-cation of individual amino acids required forsecretion within the haemolysin (HlyA) C-terminaltargeting region. Mol. Microbiol. 6, 1477–1489.
9. Kenny, B., Chervaux, C. & Holland, I. B. (1994).Evidence that residues 215 to 246 of the haemo-lysin secretion signal are involved in early steps insecretion, leading to recognition of the translocator.Mol. Microbiol. 11, 99–109.
10. Hui, D., Morden, C., Zhang, F. & Ling, V. (2000).Combinatorial analysis of the structural require-ments of the Escherichia coli haemolysin signalsequence. J. Biol. Chem. 275, 2713–2720.
11. Chervaux, C. & Holland, I. B. (1996). Random anddirected mutagenesis to elucidate the functionalimportance of helix II and F-989 in the C-terminalsecretion signal of Escherichia coli hemolysin.J. Bacteriol. 178, 1232–1236.
12. Stanley, P., Koronakis, V. & Hughes, C. (1991). Muta-tional analysis supports a role for multiple structuralfeatures in the C-terminal secretion signal ofEscherichia coli haemolysin. Mol. Microbiol. 5,2391–2403.
13. Hui, D. & Ling, V. (2002). A combinatorial approachtoward analyzing functional elements of theEscherichia coli hemolysin signal sequence. Bio-chemistry, 41, 5333–5339.
14. Wolff, N., Delepelaire, P., Ghigo, J. M. & Delepierre,M. (1997). Spectroscopic studies of the C-terminalsecretion signal of the Serratia marcescens haem acqui-sition protein (HasA) in various membrane-mimeticenvironments. Eur. J. Biochem. 243, 400–407.
15. Yin, Y., Zhang, F., Ling, V. & Arrowsmith, C. H.(1995). Structural analysis and comparison of theC-terminal transport signal domains of hemolysin Aand leukotoxin A. FEBS Letters, 366, 1–5.
16. Holland, I. B. & Blight, M. A. (1999). ABC-ATPases,adaptable energy generators fuelling transmembranemovement of a variety of molecules in organismsfrom bacteria to humans. J. Mol. Biol. 293, 381–399.
17. Pimenta, A. L., Young, J., Holland, I. B. & Blight,M. A. (1999). Antibody analysis of the localisation,expression and stability of HlyD, the MFP com-ponent of the E. coli haemolysin translocator. Mol.Gen. Genet. 261, 122–132.
18. Koronakis, V., Hughes, C. & Koronakis, E. (1991).Energetically distinct early and late stages of HlyB/HlyD-dependent secretion across both Escherichiacoli membranes. EMBO J. 10, 3263–3272.
19. Thanabalu, T., Koronakis, E., Hughes, C. & Korona-kis, V. (1998). Substrate-induced assembly of a con-tiguous channel for protein export from E. coli:reversible bridging of an inner-membrane translo-case to an outer membrane exit pore. EMBO J. 17,6487–6496.
20. Balakrishnan, L., Hughes, C. & Koronakis, V. (2001).Substrate-triggered recruitment of the TolC channel-tunnel during type I export of hemolysin byEscherichia coli. J. Mol. Biol. 313, 501–510.
21. Higgins, C. F. (1992). ABC transporters: from micro-organisms to man. Annu. Rev. Cell Biol. 8, 67–113.
22. Gentschev, I. & Goebel, W. (1992). Topological andfunctional studies on HlyB of Escherichia coli. Mol.Gen. Genet. 232, 40–48.
23. Wang, R. C., Seror, S. J., Blight, M., Pratt, J. M.,Broome-Smith, J. K. & Holland, I. B. (1991). Analysisof the membrane organization of an Escherichia coliprotein translocator, HlyB, a member of a largefamily of prokaryote and eukaryote surface transportproteins. J. Mol. Biol. 217, 441–454.
24. Koronakis, E., Hughes, C., Milisav, I. & Koronakis, V.(1995). Protein exporter function and in vitro ATPaseactivity are correlated in ABC-domain mutants ofHlyB. Mol. Microbiol. 16, 87–96.
25. Koronakis, V., Koronakis, E. & Hughes, C. (1989).Isolation and analysis of the C-terminal signaldirecting export of Escherichia coli hemolysin proteinacross both bacterial membranes. EMBO J. 8,595–605.
26. Binet, R. & Wandersman, C. (1995). Protein secretionby hybrid bacterial ABC-transporters: specific func-tions of the membrane ATPase and the membranefusion protein. EMBO J. 14, 2298–2306.
27. Koronakis, V., Hughes, C. & Koronakis, E. (1993).ATPase activity and ATP/ADP-induced confor-mational change in the soluble domain of thebacterial protein translocator hlyb. Mol. Microbiol. 8,1163–1175.
28. Delepelaire, P. (1994). PrtD, the integral membraneATP-binding cassette component of the Erwiniachrysanthemi metalloprotease secretion system,exhibits a secretion signal-regulated ATPase activity.J. Biol. Chem. 269, 27952–27957.
29. Benabdelhak, H., Blight, M. A., Stoven, V., Vallois, D.,Boucher, J. & Holland, I. B. (2003). Over expression,purification and initial structural and biochemicalcharacterization of the ABC-ATPase domain of HlyBrequired for secretion of haemolysin from E. coli.Eur. J. Biochem., in press.
30. Lofas, S. (1995). Dextran modified self-assembledmonolayer surfaces for use in biointeraction analysiswith surface plasmon resonance. Pure Appl. Chem.67, 829–834.
31. Hochuli, E. (1990). Purifaction of recombinantproteins with metal chelate adsorbent. Genet. Eng.12, 87–98.
32. Schmitt, L., Dietrich, C. & Tampe, R. (1994). Synthesisand characterization of chelator-lipids for reversibleimmobilization of engineered proteins at
1178 SPR Analysis of the HlyA/HlyB-NBD Interaction

self-assembled lipid interfaces. J. Am. Chem. Soc. 116,8485–8491.
33. Sigal, G. B., Bandad, C., Barberis, A., Strominger, J. &Whitesides, G. M. (1996). A self-assembled mono-layer for the binding and study of histidine-taggedproteins by surface plasmon resonance. Anal. Chem.68, 490–497.
34. Schmitt, L., Ludwig, M., Gaub, H. E. & Tampe, R.(2000). A metal-chelating microscopy tip as a newtoolbox for single molecule experiments by atomicforce microscopy. Biophys. J. 78, 3275–3285.
35. Nieba, L., Nieba-Axmann, S. E., Persson, A.,Hamalainen, M., Edebratt, F., Hansson, A. et al.(1997). BIACORE analysis of histidine-tagged pro-teins using a chelating NTA sensor chip. Anal. Bio-chem. 252, 217–228.
36. Karlsson, R. (1999). Affinity analysis of non-steady-state data obtained under mass transport limitedconditions using BIAcore technology. J. Mol. Recognit.12, 285–292.
37. Kempf, B. & Bremer, E. (1995). OpuA, an osmoticallyregulated binding protein-dependent transport sys-tem for the osmoprotectant glycine betaine in Bacillussubtilis. J. Biol. Chem. 270, 16701–16713.
38. Saurin, W., Hofnung, M. & Dassa, E. (1999). Gettingin or out: early segregation between importers andexporters in the evolution of ATP-binding cassette(ABC) transporters. J. Mol. Evol. 48, 22–41.
39. Gaudet, R. & Wiley, D. C. (2001). Structure of theABC ATPase domain of human TAP1, the trans-porter associated with antigen processing. EMBO J.20, 4964–4972.
40. Gray, L., Mackman, N., Nicaud, J. M. & Holland, I. B.(1986). The carboxy-terminal region of haemolysin2001 is required for secretion of the toxin fromEscherichia coli. Mol. Gen. Genet. 205, 127–133.
41. Benabdelhak, H., Schmitt, L., Horn, C., Jumel, K.,Blight, M. A. & Holland, I. B. (2003). The ABC-ATPase domain of HlyB in vitro displays positiveco-operativity, but dimers appear highly unstable.J. Mol. Biol., submitted.
42. Matsui, K., Boniface, J. J., Reay, P. A., Schild, H.,Fazekas, B., de St. Groth, F. & Davis, M. M. (1991).
Low affinity interaction of peptide–MHC complexeswith T cell receptors. Science, 254, 1788–1791.
43. Schmitt, L. & Tampe, R. (2000). Affinity, specificity,diversity: a challenge for the ABC transporter TAPin cellular immunity. ChemBioChem, 1, 16–35.
44. Loo, T. W. & Clarke, D. M. (1999). Identification ofresidues in the drug-binding domain of humanP-glycoprotein. Analysis of transmembrane segment11 by cysteine-scanning mutagenesis and inhibitionby dibromobimane. J. Biol. Chem. 274, 35388–35392.
45. Loo, T. W. & Clarke, D. M. (2000). Identification ofresidues within the drug-binding domain of thehuman multidrug resistance P-glycoprotein bycysteine-scanning mutagenesis and reaction withdibromobimane. J. Biol. Chem. 275, 39272–39278.
46. Guzman, L.-M., Belin, D., Carson, M. J. & Beckwith,J. (1995). Tight regulation, modulation, and high-level expression by vectors containing the arabinosepBAD promoter. J. Bacteriol. 177, 4121–4130.
47. Chang, G. & Roth, C. B. (2001). Structure of MsbAfrom E. coli: a homolog of the multidrug resistanceATP binding cassette (ABC) transporters. Science,293, 1793–1800.
48. Karpowich, N., Martsinkevich, O., Millen, L., Yuan,Y. R., Dai, P. L., MacVey, K. et al. (2001). Crystal struc-tures of the MJ1267 ATP binding cassette reveal aninduced-fit effect at the ATPase active site of anABC transporter. Structure, 9, 571–586.
49. Smith, P. C., Karpowich, N., Millen, L., Moody, J. E.,Rosen, J., Thomas, P. J. & Hunt, J. F. (2002). ATP bind-ing to the motor domain from an ABC transporterdrives formation of a nucleotide sandwich dimer.Mol. Cell, 10, 139–149.
50. Yuan, Y. R., Blecker, S., Martsinkevich, O., Millen, L.,Thomas, P. J. & Hunt, J. F. (2001). The crystal struc-ture of the MJ0796 ATP-binding cassette. Impli-cations for the structural consequences of ATPhydrolysis in the active site of an ABC transporter.J. Biol. Chem. 276, 32313–32321.
51. Locher, K. P., Lee, A. T. & Rees, D. C. (2002). TheE. coli BtuCD structure: a framework for ABC trans-porter architecture and mechanism. Science, 296,1091–1098.
Edited by W. Baumeister
(Received 28 November 2002; received in revised form 31 January 2003; accepted 3 February 2003)
SPR Analysis of the HlyA/HlyB-NBD Interaction 1179
Copyright © 2022 FDOKUMEN

