







Bahasa
Halaman
Hukum


1
A Schiff Base-Derived Copper (II) Complex is a potent inducer of apoptosis in
colon cancer cells by activating the intrinsic pathway
Maryam Hajrezaie,1,2 Mohammadjavad Paydar,3 Soheil Zorofchian Moghadamtousi,1
Pouya Hassandarvish,1 Nura Suleiman Gwaram,4 Maryam Zahedifard,1,2 Elham
Rouhollahi,1 Hamed Karimian,1 Chung Yeng Looi,3 Hapipah Mohd Ali,4 Nazia Abdul
Majid,1 Mahmood Ameen Abdulla, 1
1 Department of Biomedical Science, Faculty of Medicine, University of Malaya, 50603 Kuala Lumpur, Malaysia
2 Institute of Biological Science, Faculty of Science, University of Malaya, 50603 Kuala Lumpur, Malaysia
3 Department of Pharmacology, Faculty of Medicine, University of Malaya, 50603 Kuala Lumpur, Malaysia
4 Department of Chemistry, University of Malaya, 50603 Kuala Lumpur, Malaysia
Maryam Hajrezaie email address: [email protected]
Mohammadjavad Paydar email address: [email protected]
Soheil Zorofchian Moghadamtousi email address: [email protected]
Pouya Hassandarvish email address: [email protected]
Nura Suleiman Gwaram email address: [email protected]
Maryam Zahedifard email address: [email protected]
Elham Rouhollahi email address: [email protected]
Hamed Karimian email address: [email protected]
Chung Yeng Looi email address: [email protected]
Hapipah Mohd Ali email address: [email protected]
Nazia Abdul Majid email address: [email protected]
* Correspondence should be addressed to Mahmood Ameen Abdulla: [email protected]

2
ABSTRACT
Metal-based drugs with extensive clinical applications hold great promise for the development of
cancer chemotherapeutic agents. In the last few decades, Schiff bases and their complexes have
attracted significant attention in the field of coordination chemistry and have become well known
for their extensive biological potential. In the present study, we examined the antiproliferative
effect of a Copper (II) complex derived from N,N’dimethyl ethylene diamine and a 2-
hydroxyacetophenone Schiff base ligand on HT-29 colon cancer cells. This study also evaluated
the induction of apoptosis by this compound and possible mechanisms of action. The
Cu(BrHAP)2 Schiff base compound demonstrated a potent antiproliferative effect in HT-29 cells,
with an IC50 value of 2.87 µg/ml after 72 h of treatment. HT-29 cells treated with Cu (II)
complexes underwent apoptosis death, as exhibited by a progressive elevation in the proportion
of the G1 cell population. At a concentration of 6.25 µg/ml, the Cu(BrHAP)2 compound caused
significant elevation in ROS production following perturbation of mitochondrial membrane
potential and cytochrome c release, as assessed by the measurement of fluorescence intensity in
stained cells. Furthermore, the activation of caspases-3/7 and -9 was part of the Cu (II) complex-
induced apoptosis, which confirmed the involvement of mitochondrial-mediated apoptosis.
Meanwhile, there was no significant activation of caspase-8, possibly suggesting the lack of
involvement of the extrinsic pathway. Taken together, these results imply that the Cu(BrHAP)2
compound is a potential candidate for further in vivo and clinical colon cancer studies to develop
novel chemotherapeutic agents derived from metal-based agents.
Keywords: Schiff base compound; Colon cancer; HT-29 cells; Apoptosis; Cell cycle
1. Introduction

3
Cancer is a debilitating disease that afflicts a substantial portion of the world population in all
generations and is a major health problem of global concern [1]. Among the various types of
cancer, colorectal cancer is the second and third most prevalent cancer among males and females
in the United States, respectively. In spite of all the considerable progress in protective methods
and recent improvements in screening techniques and chemotherapy, the 1-year and 5-year
relative survival rates for patients suffering from colorectal cancer are 83.2% and 64.3%,
respectively [2]. In addition, due to bitter controversy over optimal methods for early detection,
full compliance of patients with screening recommendations remains a major hindrance for
diagnosis at the early stages of cancer development. Development of resistance to chemotherapy
also represents a critical issue for which simultaneous treatment with various classes of
therapeutics to reduce the resistance has yielded some success [3]. Moreover, the numerous side
effects of chemotherapeutic drugs on cancer patients, including hair loss, diarrhea, bleeding and
immunosuppression, have made the process of treatment more complicated [4]. The highly
regulated programmed cell death process of apoptosis is a matter of great interest in oncology
and cancer therapy and represents a common molecular pathway for drug resistance and
carcinogenesis [5].
Maintenance of a constant cell number in the colonic mucosa is highly regulated through the
balance between apoptosis and cell proliferation. The perturbation in this balance leads to an
escape from normal cell number homeostasis and is associated with the progression of cancer
cells [6, 7]. Thus, suppression of proliferation and elevation of apoptosis in these aberrant cells is
suggested to be the essential mechanism for the inhibition of colon cancer. Furthermore,
apoptosis and the factors involved in its mechanism of action also present a window that can be
exploited for the improvement of potential therapeutic agents with high effectiveness and less

4
adverse side effects [8]. Hence, screening for novel compounds capable of inducing apoptosis in
colon cancer cells that can be used alone or in combination with other chemotherapeutic drugs is
a significant need and represents a critical challenge in medicinal chemistry.
Metal complexes have been extensively utilized in clinics for centuries and have attracted
numerous inorganic chemists to analyze them, with the main focus being medical applications
[9, 10]. Copper, an essential trace element with an oxidative nature and bioessential activity in
human metabolism, does not exist in an ionic form in biological systems. Thus, measurement of
copper in the body is evaluated in the form of complexes with organic compounds [11]. Schiff
bases are a critical class of compounds in medical chemistry that have demonstrated significant
chemotherapeutic and antibacterial application [12, 13]. Schiff base Cu(II) complexes revealed
great potential for antiproliferative, antibacterial and gastroprotective activity [14-18].This study
evaluated the anticancer potential of a copper (II) complex derived from N,N’dimethyl ethylene
diamine and 2-hydroxyacetophenone Schiff base ligand, Cu(BrHAP)2. Furthermore, the possible
apoptotic mechanism underlying this activity was also examined.
2. Materials and methods
2.1.Reagents and chemicals
All chemicals were obtained from Sigma-Aldrich (St. Louis, MO, USA) unless otherwise
indicated. Stock solutions of tested compounds were prepared in dimethyl sulfoxide (DMSO)
and stored at -20°C, protected from the light.
2.2.Test material

5
The previously described Copper (II) complex Cu(BrHAP)2 (Figure 1) was kindly supplied
by Prof. Dr. Hapipah Mohd Ali, Department of Chemistry, Faculty of Science, University of
Malaya, Kuala Lumpur, Malaysia [18].
Figure 1. Chemical structure of Cu(BrHAP)2
2.3.Cell culture and viability measurement
HT-29 human colon cancer cells and CCD 841 normal human colon epithelial cells were
purchased from the American Type Culture Collection (Manassas, VA, USA) and maintained
routinely in Dulbecco’s Modified Eagle Medium (DMEM, Life Technologies, Inc., Rockville,
MD) containing 10% fetal bovine serum, 100 µg/mL streptomycin and 100 U/mL penicillin G at
37°C in a humidified atmosphere of 5% CO2/95% air. The cells were plated at a fitting density in
tissue culture flasks (Corning, USA) according to each experimental scale. Cell viability was
measured by a conventional MTT [3-(4,5-dimethylthiazol-2-yl)-2,5,-diphenyltetrazolium

6
bromide] reduction assay. After 48 h exposure to six concentrations of Cu(BrHAP)2, cells were
treated with MTT solution (2 mg/ml) for 2 h. The dark formazan crystals formed in intact cells
were dissolved in DMSO, and the absorbance was measured at 570 nm and 650 nm as a
background using a microplate reader (Hidex, Turku, Finland). The IC50 value was determined
as the concentration of Cu(BrHAP)2 required to reduce the absorbance of treated cells to 50% of
the DMSO-treated control cells. All samples were prepared in triplicates.
2.4.LDH release assay
Measurement of lactate dehydrogenase (LDH) release is a biomarker for determining the
cytotoxicity of a compound. Briefly, HT-29 cells were treated with different concentrations of
Cu(BrHAP)2 and Triton X-100 (positive control) for 48 h, and the supernatants of the untreated
and treated cells were transferred to a new 96-well plate for LDH activity analysis. Next, 100 µl
of LDH reaction solution was added to each well, the plate was incubated at room temperature
for 30 min and the absorbance was read at 490 nm using a Tecan Infinite®200 Pro (Tecan,
Männedorf, Switzerland) microplate reader. The amount of formazan salt and intensity of red
color in treated and untreated samples were represented as the LDH activity of cells. The LDH
release level in cells treated with Cu(BrHAP)2 was expressed as a percentage of the positive
control.
2.5.Acridine orange/ Propidium iodide double staining
A propidium iodide (PI) and acridine orange (AO) double staining assay was carried out for
detection of apoptosis in the treated cells using a fluorescent microscope (Leica attached with Q-
Floro software) according to a standard procedure. HT-29 cells (5 × 104 cells/ml in a 25 ml
culture flask) were plated, treated with Cu(BrHAP)2 at the IC50 concentration and incubated for

7
24, 48 and 72 h. After harvesting the cells, they were stained with fluorescent dyes and observed
under a UV-fluorescent microscope (Olympus BX51) within 30 min.
2.6.Cell cycle analysis
In brief, HT-29 cells (1 × 104 cells/well in 96-well plate) were supplemented with
Cu(BrHAP)2 (2 µg/ml) or DMSO (negative control) for 24 h. The live cells were then incubated
with BrdU and Phospho-Histone H3 dyes for 30 min. After the cells were fixed and stained as
described by the manufacturer’s instructions, they were visualized and analyzed using the
Cellomics ArrayScan HCS reader (Thermo Scientific). The fluorescence intensities of the dyes
were measured using a target activation bioapplication module.
To confirm the result of the fluorescence cell cycle analysis, HT-29 cells (5×104 cells/ml)
were treated with Cu(BrHAP)2 for 24, 48 and 72 h for flow cytometry analysis. After incubation,
HT-29 cells were spun down at 1800 rpm for 5 min. Next, fixation of a cell population for flow
cytometry analysis was carried out to restore integrity. In brief, the cell pellets were fixed by
mixing them with 700 µl of cold ethanol (90%) and were then kept at 4°C overnight. Treated
HT-29 cells were spun down, and the ethanol was discarded. After washing and suspending the
cells in PBS, 25 µl of RNase A (10 mg/ml) and 50 µl of propidium iodide (PI) (1 mg/ml) were
added to the fixed cells for 1 h at 37 ˚C. The added RNase A limited the ability of PI to bind to
only DNA molecules. At the end, the DNA content of the cells was analyzed by a flow
cytometer (BD FACSCanto TM II).
2.7.ORAC assay
The oxygen radical antioxidant capacity (ORAC) assay was carried out based on the
protocols described in detail previously [20]. In brief, Cu(BrHAP)2 at the concentration of 100

8
µg/mL was used for this assay in a total reaction volume of 200 µL. The experiment was
performed in a black 96-well microplate with 25 µL of compound, blank (solvent/PBS), standard
(trolox) or positive control (quercetin). The plate was then supplemented with the working
fluorescein solution (150 µL), followed by a 5 min incubation at 37°. The total volume of 200
µL was made up by adding 25 µL of AAPH working solution. Fluorescence intensity was
measured at an excitation wavelength of 485 nm and an emission wavelength of 538 nm every 2
min for 2 h. The result was quantified by calculating the differences of area under the
fluorescence decay curve (AUC) of samples and blank. The values were Trolox equivalents
(TE).
2.8.Measurement of reactive oxygen species generation (ROS)
In brief, HT-29 cells (1× 104 cells/mL) were seeded in 96-well plates and treated with
different concentrations of Cu(BrHAP)2 and DMSO (negative control) for 24 h. After 30 min
treatment with dihydroethidium (DHE) dye, cells were fixed and washed with wash buffer as
described by the manufacturer’s instructions. In the presence of superoxides, DHE dye is
oxidized to ethidium. The fluorescence intensity was determined by a fluorescent plate reader at
an extension wavelength of 520 nm and an emission wavelength of 620 nm.
2.9. Multiple Cytotoxicity Assay
The critical factors for monitoring the cell health, namely, cell loss, changes in cell
permeability, cytochrome c release, mitochondrial membrane potential changes, nuclear size and
morphological changes, were studied using a Cellomics Multiparameter Cytotoxicity 3 Kit as
described in detail previously [19]. Plates with stained cells were analyzed using the ArrayScan
HCS system (Cellomics, PA, USA).

9
2.10. Measurement of Caspase Activities
Caspase-3/7, -8 and -9 activities were determined using the commercial Caspase-Glo® 3/7, 8
and 9 assay kit (Promega, Madison, WI). HT-29 cells (1.0 × 104 cells/well) were seeded
overnight in white-walled 96-well plates and treated with different concentrations of
Cu(BrHAP)2 for 24 h. According to the manufacturer’s protocol, the treated cells were
supplemented with caspase-Glo reagent (100 µl) and incubated at room temperature for 30 min.
The active caspases from apoptotic cells caused the cleavage of aminoluciferin-labeled synthetic
tetrapeptide, leading to the release of substrate for the luciferase enzyme. Caspase activities were
analyzed using a Tecan Infinite®200 Pro (Tecan, Männedorf, Switzerland) microplate reader.
2.11.Measurement of NF-κB activity
In brief, HT-29 cells (1.0 × 104 cells/well in a 96-well plate) were treated with different
concentrations of Cu(BrHAP)2 for 3 h, followed by stimulation with TNF-α (1 ng/ml) for 30
min. After discarding the medium, cells were fixed and stained using a Cellomics nucleus factor-
κB (NF-κB) activation kit (Thermo Scientific) according to the manufacturer's instructions. Next,
an Array Scan HCS Reader was used for evaluation of the plate. Cytoplasmic and nuclear NF-κB
intensity ratios were calculated using Cytoplasm to Nucleus Translocation Bioapplication
software. The average intensity of 200 cells/well was determined. The ratios for untreated,
treated and TNF-α-stimulated cells were compared.
2.12. Statistical Analysis
All the experiments were performed at least three times independently. The results were
presented as the mean ± standard deviation (SD) of the number of experiments shown in the

10
legends. An analysis of variance (ANOVA) was carried out using the prism statistical package
(GraphPad Software, USA). P < 0.05 was considered statistically significant.
3. Results
3.1.The effect of Cu(BrHAP)2 on human normal and cancer cells of the colon
Initially, the cytotoxicity of Cu(BrHAP)2 was tested on HT-29 and CCD 841 cell lines. The
IC50 values of the Schiff base compound were determined based on the result collected from
three independent MTT experiments. As indicated in Table 1, Cu(BrHAP)2 elicited a significant
cytotoxicity and cell inhibitory effect after 24, 48 and 72 h of treatment on HT-29 cell.
Table 1. Inhibitory effects of Cu(BrHAP)2 on the proliferation of human normal and cancer
cells. Cells were treated with various concentrations of Cu(BrHAP)2 for 24, 48 and 72 h. The
IC50 values were analyzed by non-linear regression analysis.
Cell line Classification IC50 (µg/ml)
24 h 48 h 72 h
HT-29 Colon cancer cells 2.87 ± 0.21 1.93 ± 0.37 1.44 ± 0.26
CCD 841 Normal colon cells 50 ˂ 50 ˂ 50 ˂
3.2.Cu(BrHAP)2 -induced LDH release
Lactate dehydrogenase (LDH) release in the medium is a marker that shows the loss of
membrane integrity, apoptosis or necrosis. The cytotoxicity of the Cu(BrHAP)2 compound, as
determined by the LDH release assay, was quantified on HT-29 cells treated with various
concentrations of the Schiff base compound for 48 h. Cu(BrHAP)2 induced a significant

11
elevation in LDH release, demonstrating cytotoxicity at the 6.25 and 12.5 µg/ml concentrations
compared to the control cells (Figure 2).
Figure 2. The LDH release assay revealed the significant cytotoxicity of the Cu(BrHAP)2
compound on HT-29 cells at the 6.25 and 12.5 µg/ml concentrations.
3.3.Quantification of apoptosis using phase-contrast microscopy and AO/PI double-
staining
Morphological changes in HT-29 cells treated with Cu(BrHAP)2 compound were observed
under a fluorescent microscope at 24, 48 and 72 h. The cells were scored under a fluorescent
microscope to analyze viable cells, early apoptosis and late apoptosis. Early apoptosis, defined as
intervening AO within the fragmented DNA, was observed under bright green fluorescence. At
the same time, control cells were visualized with a green intact nuclear structure. After 24 and 48
h of treatment with Cu(BrHAP)2, moderate apoptosis was observed in the form of blebbing and
nuclear chromatin condensation. Furthermore, in the late stage of apoptosis, changes, such as the

12
presence of a reddish-orange color due to binding of PI to denatured DNA, were observed after
72 h of treatment (Figure 3). The results showed that the Cu(BrHAP)2 compound induced
morphological features of apoptosis in a time-dependent manner.
Figure 3. (A) Untreated HT-29 cells after 72 h demonstrated normal structure without prominent
apoptosis or necrosis. Early apoptosis features, including blebbing and chromatin condensation,
were observed after (B) 24 and (C) 48 h of treatment with Cu(BrHAP)2. (D) Late apoptosis was
noticed after 72 h of Cu(BrHAP)2 treatment (magnification: 200×). VI: Viable cells; BL:
Blebbing of the cell membrane; CC: Chromatin condensation; LA: Late apoptosis.
3.4. Cu(BrHAP)2 -induced G1/G2 cell cycle arrest
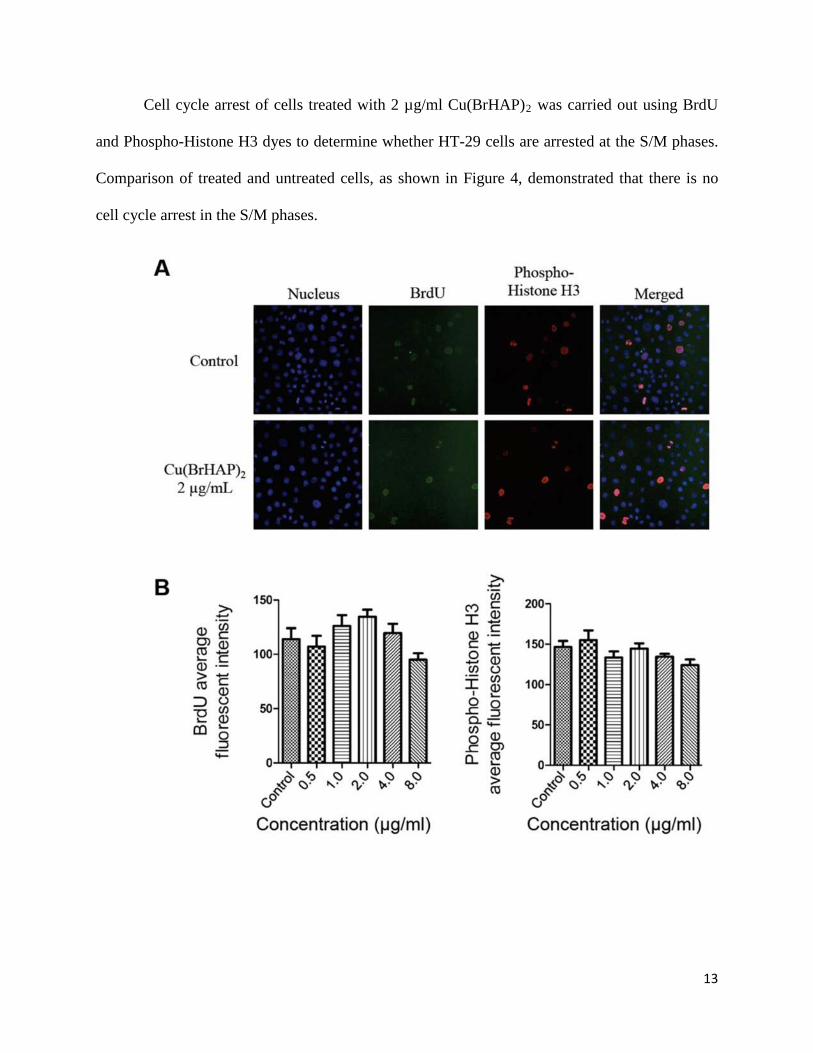
13
Cell cycle arrest of cells treated with 2 µg/ml Cu(BrHAP)2 was carried out using BrdU
and Phospho-Histone H3 dyes to determine whether HT-29 cells are arrested at the S/M phases.
Comparison of treated and untreated cells, as shown in Figure 4, demonstrated that there is no
cell cycle arrest in the S/M phases.

14
Figure 4. (A) Effect of Cu(BrHAP)2 on cell cycle arrest in the S/M phase. After incubation with
DMSO or Cu(BrHAP)2 at 2 µg/ml for 24 h, HT-29 cells were collected, stained with BrdU and
Phospho-Histone H3 and subjected to the Cellomics ArrayScan HCS reader for cell cycle
analysis. (B) Representative bar charts indicating that Cu(BrHAP)2 treatment caused no
significant changes in BrdU or Phospho-Histone H3 fluorescence intensities in treated HT-29
cells. Cu(BrHAP)2 induced no cell cycle arrest at the S/M phases in treated HT-29 cells. Data
were expressed as the mean ± SD of fluorescence intensity readings for three independent
experiments.
The lack of cell cycle arrest in the S/M phases suggested possible cell cycle arrest in the
G1/G2 phases. To determine the exact arrested phase, treated HT-29 cells were analyzed for cell
cycle progression using flow cytometry. As expected, there was no significant arrest in the S/M
phases. Meanwhile, significant cell cycle arrest in the G1 phase was observed for HT-29 cells
after 24 and 48 h of treatment (Figure 5).

15
Figure 5. Effect of Cu(BrHAP)2 on cell cycle progression in HT-29 cells as assessed by flow
cytometry. After incubation with Cu(BrHAP)2 for 24 and 48 h, significant cell cycle arrest in the
G1 phase were observed.
3.5.ORAC anti-oxidant activity assay
Antioxidant capacity was measured by ORAC assay, which is the only assay that involves
the use of peroxyl radical as a pro-oxidant and quantifies activity via the area under the curve
(AUC) technique. In our experiment, quercetin was used as a positive control. The result
demonstrated that Cu(BrHAP)2 exhibited low to moderate antioxidant activity compared to
quercetin (Table 2).
16
Table 2. Antioxidant capacity of the compound was determined by the ORAC method.
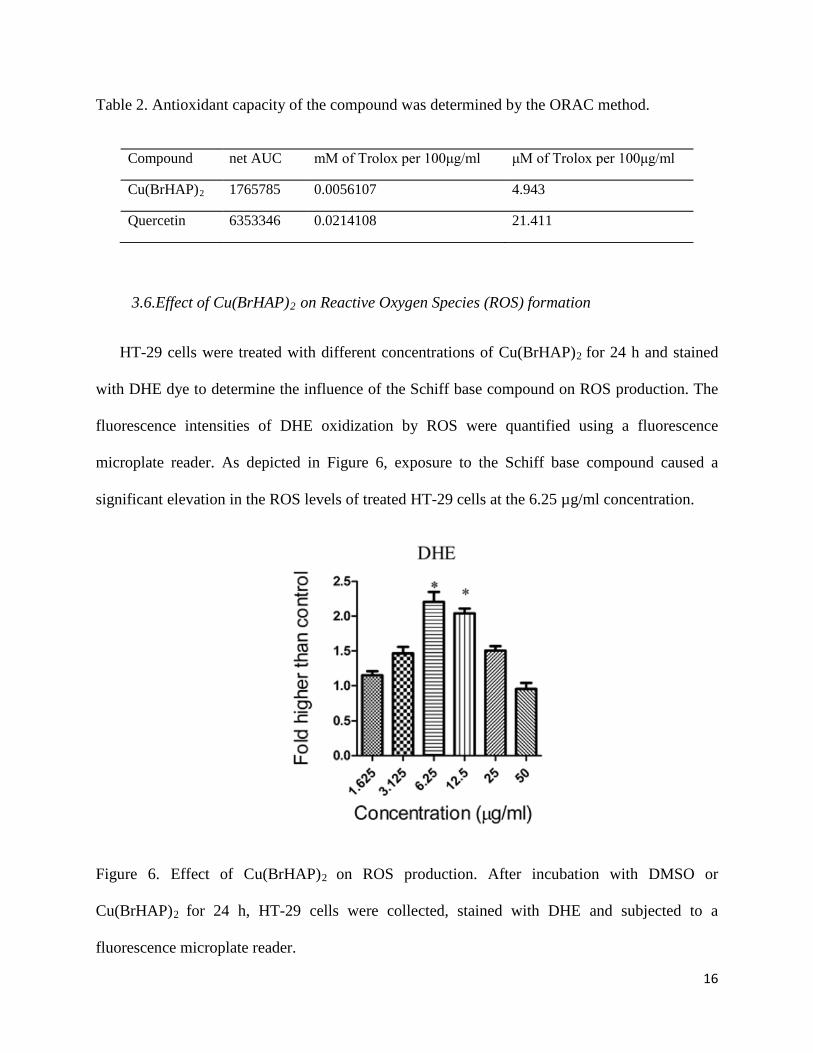
3.6.Effect of Cu(BrHAP)2 on Reactive Oxygen Species (ROS) formation
HT-29 cells were treated with different concentrations of Cu(BrHAP)2 for 24 h and stained
with DHE dye to determine the influence of the Schiff base compound on ROS production. The
fluorescence intensities of DHE oxidization by ROS were quantified using a fluorescence
microplate reader. As depicted in Figure 6, exposure to the Schiff base compound caused a
significant elevation in the ROS levels of treated HT-29 cells at the 6.25 µg/ml concentration.
Figure 6. Effect of Cu(BrHAP)2 on ROS production. After incubation with DMSO or
Cu(BrHAP)2 for 24 h, HT-29 cells were collected, stained with DHE and subjected to a
fluorescence microplate reader.
Compound net AUC mM of Trolox per 100μg/ml μM of Trolox per 100μg/ml
Cu(BrHAP)2 1765785 0.0056107 4.943
Quercetin 6353346 0.0214108 21.411

17
3.7.Effects of Cu(BrHAP)2 on nuclear morphology, membrane permeability, mitochondrial
membrane potential (MMP) and cytochrome c release
To investigate the induction of apoptosis by Cu(BrHAP)2, nuclear morphological changes in
HT-29 cells were analyzed by detection of nuclear condensation. As shown in Figure 7, Hoechst
33342 staining demonstrated that nuclear condensation, which is directly related to apoptotic
chromatin changes, emerged in some cells after treatment with Cu(BrHAP)2. Meanwhile, the
permeability of treated cells was also elevated. Mitochondria are the main source for the
production of ROS and adenosine triphosphate (ATP) and are critical in controlling the death and
survival of cells. The reduction in fluorescence intensity depicted in Figure 6 reflects the
significant decrease of MMP in the cells treated with the Schiff base compound. Meanwhile,
Cu(BrHAP)2 triggered the translocation of cytochrome c from mitochondria into the cytosol
during apoptosis in HT-29 cells.

18
Figure 7. (A) Representative images of HT-29 cells treated with medium alone and 2 µg/ml of
Cu(BrHAP)2 and stained with Hoechst 33342 for nuclear, cytochrome c, membrane
permeability, and mitochondrial membrane potential dyes. Cu(BrHAP)2 induced a noteworthy

19
elevation in membrane permeability and cytochrome c release and a marked reduction in
mitochondrial membrane potential (magnification: 200×). (B) Representative bar charts
indicating dose-dependent increased cell permeability, reduced MMP and increased cytochrome
c release in treated HT-29 cells.
3.8.Caspase activation
The elevation in ROS production associated with a collapse in MMP may lead to the
activation of the caspase cascade. To investigate caspase activation, the bioluminescent
intensities representing caspase-3/7, -8, -9 activities were quantified in HT-29 cells treated with
different concentrations of Cu(BrHAP)2 for 24 h. As shown in Figure 8, significant elevation in
the activity of caspase -3/7 at the 6.25 µg/ml concentration and caspase -9 at the 6.25 and 12.5
µg/ml concentrations was observed in Cu(BrHAP)2-treated cells, while no significant change in
the activity of caspase-8 was detected between treated and untreated HT-29 cells. Thus, the
apoptosis induced by the Schiff base compound in HT-29 cells is possibly mediated via
the intrinsic pathway, but not the extrinsic pathway.

20
Figure 8. Relative luminescence expression of Caspases 3/7, 8 and 9 in HT-29 cells treated with
various concentrations of Cu(BrHAP)2.
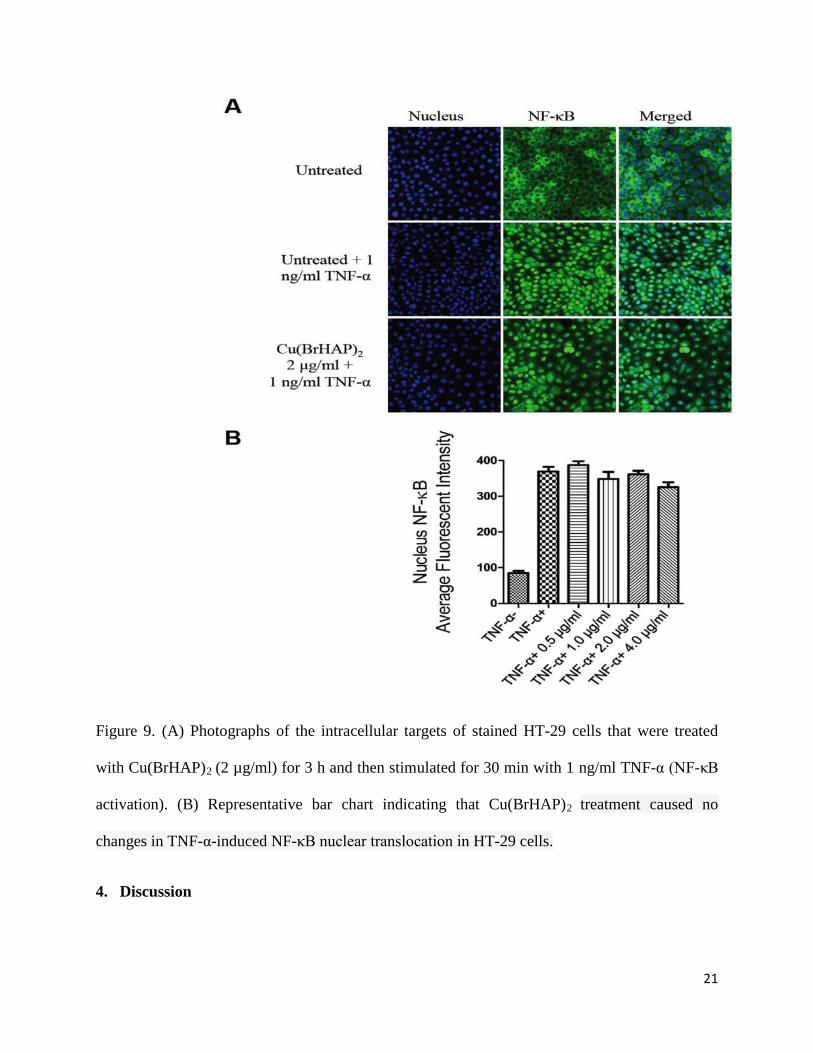
3.9.NF-κB Translocation
Nuclear factor kappa B (NF-κB) is a transcription factor that has a critical role in cytokine
gene expression. NF-κB activation and translocation to the nucleus to enable DNA-binding
activity and facilitate target gene expression is mediated by inflammatory cytokines such as
tumor necrosis factor-α (TNF-α). The Cu(BrHAP)2 Schiff base compound did not exhibit any
inhibitory effect on translocation of TNF-α-stimulated NF-κB in HT-29 treated cells, and TNF-
α-stimulation led to NF-κB translocation from the cytoplasm to the nucleus (Figure 9).
21
Figure 9. (A) Photographs of the intracellular targets of stained HT-29 cells that were treated
with Cu(BrHAP)2 (2 µg/ml) for 3 h and then stimulated for 30 min with 1 ng/ml TNF-α (NF-κB
activation). (B) Representative bar chart indicating that Cu(BrHAP)2 treatment caused no
changes in TNF-α-induced NF-κB nuclear translocation in HT-29 cells.
4. Discussion

22
Carcinogenesis is a multistage process in which unregulated cell proliferation as well as a
reduction in apoptosis incidence serve as initial characterizations for its progression [21]. One of
the defense procedures in multicellular organisms is the destruction of undesirable cell
development, which is defined as programmed cell death. Apoptosis is the most noticed
programmed cell death mechanism and is characterized by distinct morphological changes such
as membrane permeability, cell shrinkage, disruption of the mitochondrial membrane and
chromatin condensation [22, 23]. The disruption of cellular homeostasis between cell death and
cell proliferation leads to cancer incidence [24], and agents that can induce apoptosis are known
to have potential anticancer effects [25, 26]. Apoptosis pathways are effective targets for cancer
therapy as well as chemoprevention. Numerous chemopreventive drugs have been determined to
regulate key events or molecules in apoptosis-inducing signal transduction pathways [27]. In the
present study, the Cu(BrHAP)2 Schiff base compound was evaluated for its ability to inhibit the
growth of HT-29 cells using an MTT assay. HT-29 cells have recently been characterized as a
suitable model for colon cancer studies [28-30]. The Cu (II) complex used in this study exhibited
significant inhibition of HT-29 cell growth, with the IC50 values ranging from 2.87 at 24 h to
1.44 at 72 h. The results indicated that the Schiff base complex induced an antiproliferative
effect on human colon cancer cells in a time- and dose-dependent manner. Meanwhile, the non-
tumorigenic colon cell line (CCD 841) showed no cytotoxicity after treatment with the
compound. The cytotoxic effect of the Cu(II) compound was also confirmed by measuring the
level of LDH release from treated cells. Considerably elevated LDH release showed that the
cytotoxicity of the Cu(BrHAP)2 compound possibly occurred via the loss of membrane integrity,
whether through activation of apoptosis or the necrosis pathway [31].

23
The observation of early apoptosis and late apoptosis by fluorescent microscopy analysis and
AO/PI double-staining following treatment of HT-29 cells with the compound included some
signs of apoptosis, namely, cytoplasmic shrinkage, membrane blebbing and DNA fragmentation
[32, 33]. We found that the number of cells with early apoptosis features was higher at earlier
stages of treatment. However, when treatment time increased to 72 h, late apoptosis or necrosis
characterizations were dominant among treated HT-29 cells. Concurrent detection of late
apoptosis or necrosis is scientifically possible because treated HT-29 cells undergoing apoptosis
may have progressed into necrosis due to the prolonged incubation with the Schiff base
compound.
To elucidate the mechanisms underlying the observed antiproliferative effect of the Cu(II)
complex on cancer cells, cell cycle distribution was analyzed using BrdU and Phospho-Histone
H3 staining along with flow cytometry [34-36]. BrdU dye can attach to the synthesized DNA of
replicating cells during the S phase of the cell cycle, while Phospho-Histone H3 dye stains cells
in different mitotic stages. The cell cycle results from the BrdU and Phospho-Histone H3 double
staining assay indicated that there were no significant changes in the number of cells in the S/M
phases after the exposure of HT-29 cells to the Schiff base compound. This result suggests the
possibility that the cells were arrested in the G1 or G2 phase of the cell cycle. Thus, the flow
cytometry analysis of the cell cycle was performed to determine the exact arrested phase, and the
results demonstrated significant cell cycle arrest at G1 after 24 and 48 h of treatment, suggesting
proliferative suppression via induction of apoptosis [37, 38].
Perturbation of mitochondrial membrane potential is one of the earliest intracellular events
that occurs following the induction of apoptosis [39]. As the main source of cellular ROS and
adenosine triphosphate (ATP), mitochondria are the key regulators of mechanisms controlling

24
the survival or death of cells. After confirming that the Cu(BrHAP)2 Schiff base compound did
not have significant antioxidant capacity in HT-29 cancer cells using the ORAC assay, the
induction of ROS production in treated cells was analyzed. According to our study, after
exposing the Cu (II) compound to HT-29 cells and analyzing the levels of ROS, it was
demonstrated that the level of ROS in treated HT-29 cells was significantly elevated at a
compound concentration of 6.25 µg/ml.
In metal-induced apoptosis, the mitochondria have the crucial role in mediating apoptosis
through metal-induced ROS [40]. The intrinsic or mitochondrial-dependent signaling pathway
involves different factors of non-receptor-mediated stimuli that induce intracellular signals.
These signals, mainly through the p53 protein, act on the mitochondrial-initiated events.
Excessive ROS production is a negative signal that can result in the failure of suppression of
antiapoptotic factors, thereby triggering apoptosis. Therefore, we used mitochondrial membrane
potential (MMP) fluorescent probes to examine the effect of elevated ROS production on the
function of mitochondria in treated HT-29 cells. As shown in Figure 7, changes in MMP after
treatment with the Cu(BrHAP)2 Schiff base compound leading to the membrane depolarization
of the mitochondria were demonstrated by Rhodamine 123 release to the cytoplasm from the
mitochondria matrix. The result implies that the induction of apoptosis by Cu (II) Schiff base
complexes may be associated with the mitochondrial pathway [26, 41, 42]. One of the important
signals to initiate the procedure of apoptosis is cytosolic cytochrome c. The release of
cytochrome c into the cytosol and reduction of its levels in the mitochondria have been shown to
occur as a result of changes in MMP [30]. As the result illustrated, the synthetic Schiff base
compound also led to an increase in the level of cytochrome c in the cytosol compared to the
control.

25
The excessive production of ROS from mitochondria and the collapse of MMP may activate
the downstream caspase molecules and consequently lead to apoptotic cell death. After the
binding of cytochrome c to apoptotic activating factor-1, caspase-9 is activated via apoptosome
formation, which leads to active caspase-3/7, the most effective caspase with many cellular
targets [43]. In the extrinsic pathway, apoptosis is mediated by death receptors. As an example,
FAS ligand interacts with the FAS receptor, leading to the activation of caspase-8 [44]. Caspase-
8 activation cleaves and activates downstream executioner caspases such as caspase-3/7 [45, 46].
In our study, the Cu(BrHAP)2 schiff base compound induced significant elevation in the
caspase-3/7 and -9 activities compared to the control. Meanwhile, there was no activation of
caspase-8, suggesting that the apoptosis induced in HT-29 cells was mediated via the intrinsic
mitochondrial pathway but not the extrinsic, death receptor-linked caspase-8 pathway.
The supporting evidence of LDH release, ROS production, MMP suppression, elevation in
the level of cytochrome c and activation of caspase-3/7 and -9 demonstrated the promising
anticancer activity of the Cu(BrHAP)2 Schiff base compound against the HT-29 colon cancer
cell line via the intrinsic mitochondrial pathway.
Acknowledgments
The authors would like to thank the University of Malaya for supporting this project
through the PV069-2012A grant and the Ministry of Higher Education Malaysia for their support
through the HIR-MOHE (E0045-20001) grant.
References
[1] S.E. Atawodi, “Nigerian food stuffs with prostate cancer chemopreventive polyphenols,”
Infectious agents and cancer, vol .6, no. S9, 2011.

26
[2] R. Siegel, C. DeSantis, K. Virgo, K. Stein, A Mariotto, and T Smith et al.,” Cancer
treatment and survivorship statistics ,“ a cancer journal for clinicians, vol. 62 , pp. 220-41 ,
2012.
[3] H. Li, Wu. WKK, Z. Zheng, CT. Che, L. Yu, ZJ. Li et al,” 2, 3′, 4, 4′, 5′-Pentamethoxy-
trans-stilbene, a resveratrol derivative, is a potent inducer of apoptosis in colon cancer cells via
targeting microtubules,“ Biochemical pharmacology, vol. 78, pp. 1224-32 , 2009.
[4] D. Kranz and M. Dobbelstein,” A killer promoting survival: p53 as a selective means to
avoid side effects of chemotherapy,“ Cell Cycle, vol.11, pp.2053-4., 2012.
[5] D. Hanahan and RA, Weinberg” The hallmarks of cancer, “cell, vol. 100, pp. 57-70,
2000.
[6] X. Hao, M. Du, AE. Bishop, and I. Talbot,” Imbalance between proliferation and
apoptosis in the development of colorectal carcinoma,“ Virchows Archiv, vol. 433 , pp. 523-7,
1998.
[7] JF. Whitfield,” Calcium, calcium-sensing receptor and colon cancer,“ Cancer letters,
vol. 275, pp. 9-16, 2009.
[8] CK. Chan, BH. Goh, MNA. Kamarudin, and HA. Kadir,” Aqueous Fraction of
Nephelium ramboutan-ake Rind Induces Mitochondrial-Mediated Apoptosis in HT-29 Human
Colorectal Adenocarcinoma Cells,“ Molecules, vol. 17, pp. 6633-57, 2012.
[9] I. Kostova,” Gold coordination complexes as anticancer agents. Anti-Cancer Agents in
Medicinal Chemistry,“, vol. 6, pp.19-32, 2006.

27
[10] V. Milacic, D. Chen, L. Ronconi, KR. Landis-Piwowar, D. Fregona, and QP. Dou,” A
novel anticancer gold (III) dithiocarbamate compound inhibits the activity of a purified 20S
proteasome and 26S proteasome in human breast cancer cell cultures and xenografts,“
Cancer research, vol. 66, no. , pp. 10478-86, 2006.
[11] H. Dollwet, and J. Sorenson,” Historic uses of copper compounds in medicine,“ Trace
elements in medicine, vol. 2, no. , pp. 80-7, 1985.
[12] AW. Tai, EJ. Lien, MM. Lai, and TA. Khwaja ,” Novel N-hydroxyguanidine derivatives
as anticancer and antiviral agents,“ Journal of medicinal chemistry, vol. 27, no. , pp. 236-8,
1984.
[13] PH. Wang, JG. Keck, EJ. Lien, and MM. Lai.,” Design, synthesis, testing, and
quantitative structure-activity relationship analysis of substituted salicylaldehyde Schiff bases of
1-amino-3-hydroxyguanidine tosylate as new antiviral agents against coronavirus,“ Journal of
medicinal chemistry, vol. 33, pp. 608-14, 1990.
[14] J. Sorensen,” Copper Complexes offer a physiological approach to treatment of chronic
leases,“ Prog Med Chem, vol. 26, pp. 437-568, 1989.
[15] C. Bolos, GS. Nikolov, L. Ekateriniadou, A, Kortsaris, D. Kyriakidis and ,” Structure-
activity relationships for some Diamine, Triamine and Schiff Base Derivatives and their copper
(II) complexes,“ Metal based drugs, vol. 5 , pp. 323-32, 1998.
[16] AT. Chaviara, P. Cox, K. Repana, A Pantazaki, K. Papazisis, A. Kortsaris et al,”
The unexpected formation of biologically active Cu (II) Schiff mono-base complexes with 2-

28
thiophene-carboxaldehyde and dipropylenetriamine: crystal and molecular structure of
CudptaSCl2,“ Journal of Inorganic Biochemistry, vol. 99, pp. 467-76, 2005.
[17] AT. Chaviara, P. Cox, K. Repana, R . Papi, K. Papazisis, D. Zambouli et al,” Copper
(II) Schiff base coordination compounds of dien with heterocyclic aldehydes and 2-amino-5-
methyl-thiazole: synthesis, characterization, antiproliferative and antibacterial studies. Crystal
structure of CudienOOCl2 ,“ Journal of Inorganic Biochemistry, vol. 98, pp.1271-83, 2004.
[18] M. Hajrezaie, S. Golbabapour, P. Hassandarvish, NS. Gwaram, AHA. Hadi, HM. Ali et
al ,” Acute toxicity and gastroprotection studies of a new schiff base derived copper (II)
complex against ethanol-induced acute gastric lesions in rats,“ PloS one, vol.7, pp. e51537,
2012.
[19] S-C. Cheah, DR. Appleton, S-T. Lee, M-L. Lam, AHA. Hadi, and MR. Mustafa,”
Panduratin a inhibits the growth of A549 cells through induction of apoptosis and inhibition of
NF-KappaB translocation, “Molecules, vol. 16, pp. 2583-98, 2011.
[20] Thomson, MJ. Paydar, Y. Li Wong et al ,” Antibacterial, Antioxidant activity and
Phytochemical studies of Crossandra infundibuliformis leaf extracts,“ Pakistan Journal of
Biological Sciences , vol. 16, pp. 1212-5, 2013.
[21] MR. Patel, GJ. Dehmer, JW. Hirshfeld, et al,” A Report by the American College of
Cardiology Foundation Appropriateness Criteria Task Force, Society for Cardiovascular
Angiography and Interventions, Society of Thoracic Surgeons, American Association for
Thoracic Surgery, American Heart Association, and the American Society of Nuclear Cardiology
Endorsed by the American Society of Echocardiography, the Heart Failure Society of America,

29
and the Society of Cardiovascular Computed Tomography ,“ Journal of the American College of
Cardiology, vol. 53, pp. 530-53, 2009.
[22] R. Scatena ,” Mitochondria and cancer: a growing role in apoptosis, cancer cell
metabolism and dedifferentiation. Advances in Mitochondrial Medicine ,“ Springer, pp. 287-
308, 2012.
[23] T. Verfaillie, AD. Garg and P. Agostinis,” Targeting ER stress induced apoptosis and
inflammation in cancer ,“ Cancer letters, vol. 332, pp. 249-64, 2013.
[24] MJ. Lee, AS. Ye, AK. Gardino, AM. Heijink et al ,” Sequential application of
anticancer drugs enhances cell death by rewiring apoptotic signaling networks,“ Cell, vol. 149,
pp. 780-94, 2012.
[25] A. Chakraborty, P. Kumar, K. Ghosh and P. Roy,” Evaluation of a Schiff base copper
complex compound as potent anticancer molecule with multiple targets of action,“
European journal of pharmacology, vol.647, pp.1-12, 2010.
[26] NA. Rey, A. Neves, PP. Silva et al,” A synthetic dinuclear copper (II) hydrolase and its
potential as antitumoral: Cytotoxicity, cellular uptake, and DNA cleavage, “Journal of inorganic
biochemistry, vol.103, pp. 1323-30, 2009.
[27] R. Suzuki, Y. Yasui, H. Kohno et al ,” Catalpa seed oil rich in 9t, 11t, 13c-conjugated
linolenic acid suppresses the development of colonic aberrant crypt foci induced by
azoxymethane in rats,“ Oncology reports, vol.16, pp. 989-96, 2006.

30
[28] T-I. Jeon, C-H. Jung, J-Y Cho, DK. Park and J-H. Moon,” Identification of an anticancer
compound against HT-29 cells from Phellinus linteus grown on germinated brown rice,“ Asian
Pacific Journal of Tropical Biomedicine,vol. 3pp.785-9, 2013.
[29] S. Asciutti, E. Camaioni, P. Mong et al ,” Therapeutic Potential of a Novel Poly (ADP-
ribose) Polymerase Inhibitor, Hydamtiq, in Human Pancreatic and Colon Cancers,“
Gastroenterology, vol.144, pp. S-875, 2013.
[30] RA. Schneider, KG. Eckles, VC. Kelty et al,” Celecoxib induces apoptosis by the
intrinsic pathway in HT-29 colon carcinoma and A375 melanoma cells,“ Faseb Journal,
pp.20814-3998, 2013.
[31] EJ. Choi, JI. Lee and G-H. Kim,” Evaluation of the anticancer activities of thioflavanone
and thioflavone in human breast cancer cell lines ,“ International Journal of Molecular
Medicine, pp. 29:252, 2012.
[32] J. Ly, D. Grubb and A. Lawen,” The mitochondrial membrane potential (Δψm) in
apoptosis; an update,“ Apoptosis, vol. 8, pp. 115-28, 2003.
[33] W. Hu and JJ. Kavanagh,” Anticancer therapy targeting the apoptotic pathway ,“ The
lancet oncology, vol. 4, pp. 721-9, 2003.
[34] M. Aarts, R. Sharpe, I Garcia-Murillas et al ,”Forced mitotic entry of S-phase cells as a
therapeutic strategy induced by inhibition of WEE1,“ Cancer discovery, vol. 2, pp. 524-39,
2012.

31
[35] L. Wang, Y. Xu, L Fu and L. Lou,” (5R)-5-hydroxytriptolide (LLDT-8), a novel
immunosuppressant in clinical trials, exhibits potent antitumor activity via transcription
inhibition,“ Cancer letters, vol. 324, pp.75-82, 2012.
[36] AR. Tentner, MJ. Lee, GJ. Ostheimer et al ,” Combined experimental and computational
analysis of DNA damage signaling reveals context-dependent roles for Erk in apoptosis and
G1/S arrest after genotoxic stress,“ Molecular systems biology, vol. 8, 2012.
[37] F. Gasparri, P. Cappella, and A. Galvani,” Multiparametric cell cycle analysis by
automated microscopy,“ Journal of biomolecular screening, vol. 11, pp. 586-98, 2006.
[38] WL. See, JP. Miller, M. Squatrito, E. Holland, MD. Resh, and A. Koff,” Defective
DNA double-strand break repair underlies enhanced tumorigenesis and chromosomal instability
in p27-deficient mice with growth factor-induced oligodendrogliomas,“ Oncogene, vol. 29, pp.
1720-31, 2010.
[39] Y-J. Liu, Z-H. Liang, X-L. Hong et al ,” Synthesis, characterization, cytotoxicity,
apoptotic inducing activity, cellular uptake, interaction of DNA binding and antioxidant activity
studies of ruthenium (II) complexes,“ Inorganica Chimica Acta, vol. 387, pp.117-24, 2012.
[40] F. Chen, V. Vallyathan, V. Castranova, and X. Shi,” Cell apoptosis induced by
carcinogenic metals,“ Molecular and Cellular Biochemistry, vol. 222, pp.183-8, 2001.
[41] Y. Zhang, X. Wang, W. Fang et al ,” Synthesis and in vitro antitumor activity of two
mixed-ligand oxovanadium (IV) complexes of Schiff base and phenanthroline,“ Bioinorganic
chemistry and applications, 2013.

32
[42] A. Bishayee and AS. Darvesh ,” Pomegranate-derived constituents as inducers of cell
death: implications in cancer prevention and therapy,“ Natural compounds as inducers of cell
death: Springer, pp. 33-47, 2012.
[43] L. Li, X. Luo, and X. Wang,” Endonuclease G is an apoptotic DNase when released
from mitochondria,“ Nature, vol. 412, pp. 95-9, 2001.
[44] R. El-Ghany, N. Sharaf, L. Kassem, L. Mahran, and O. Heikal,” Thymoquinone
triggers anti-apoptotic signaling targeting death ligand and apoptotic regulators in a model of
hepatic ischemia reperfusion injury,“ Drug Discov Ther, vol. 3, pp. 296-306, 2009.
[45] M. Hyer, R. Shi, M. Krajewska, C Meyer et al,”Apoptotic activity and mechanism of 2-
cyano-3, 12-dioxoolean-1, 9-dien-28-oic-acid and related synthetic triterpenoids in prostate
cancer ,“ Cancer Research, vol. 68, pp. 2927-33, 2008.
[46] F. Qi, A. Li, Y. Inagaki et al,” Induction of apoptosis by cinobufacini preparation
through mitochondria-and Fas-mediated caspase-dependent pathways in human hepatocellular
carcinoma cells ,“ Food and Chemical Toxicology, vol. 50, pp. 295-302, 2012.
Top Related
Copyright © 2022 FDOKUMEN

