







Bahasa
Halaman
Hukum


i
PERBANDINGAN ANTARA LEGUMINOSA Stylosanthes
guianensis DAN Stylosanthes scabra TERHADAP
CEKAMAN KEKERINGAN DAN PEMBERIAN
HIDROGEN PEROKSIDA (H2O2)
SKRIPSI
SRI SUMARYANI
DEPARTEMEN ILMU NUTRISI DAN TEKNOLOGI PAKAN
FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR
2012

ii
RINGKASAN
SRI SUMARYANI. D24080356. 2012. Perbandingan antara Leguminosa
Stylosanthes guianensis dan Stylosanthes scabra terhadap Cekaman Kekeringan
dan Pemberian Hidrogen Peroksida (H2O2). Skripsi. Departemen Ilmu Nutrisi
dan Teknologi Pakan. Fakultas Peternakan. Institut Pertanian Bogor.
Pembimbing Utama : Dr. Ir. Panca Dewi M.H.K., M. S.
Pembimbing Anggota : Ir. Widya Hermana, M. Si.
Pakan merupakan faktor utama dalam peternakan yang mempengaruhi
produksi peternakan. Apabila ternak kekurangan pakan maka produksi ternak akan
menurun dan kualitas hasil produksi juga akan menurun. Salah satu pakan yang
sangat dibutuhkan oleh peternak adalah hijauan dan hijauan dapat mengalami
penurunan produksi yang dapat disebabkan oleh cekaman kekeringan. Tidak semua
tanaman tidak dapat hidup karena cekaman kekeringan, salah satu tanaman yang
dapat mentolerir cekaman kekeringan adalah leguminosa Stylosanthes. Penelitian ini
menggunakan S. guianensis dan S. scabra. Penelitian ini juga menggunakan H2O2
dimana H2O2 merupakan bahan yang dapat membuat tanaman mengalami cekaman
kekeringan. H2O2 digunakan pada penelitian ini karena cuaca sekitar tempat
penelitian tidak menentu sehingga penelitian ini menggunakan H2O2 yang dapat
menggantikan cekaman kekeringan selain cekaman kekeringan dari air.
Penelitian ini menggunakan 3 perlakuan dan 4 ulangan, dengan perlakuan: M0:
Kontrol (disiram setiap hari dan tidak diberikan H2O2); M1: stres kekeringan tunggal
(tidak diberikan H2O2 dan tidak disiram setiap hari); M2: stres kekeringan ganda
(diberikan H2O2 dan tidak disiram setiap hari). Penelitian ini dilakukan dengan
menggunakan Rancangan Acak Lengkap dan dilakukan Uji Jarak Duncan (Steel dan
Torrie, 1995). Pengambilan data untuk kadar air tanah, rataan pertambahan tinggi,
rataan pertambahan jumlah daun trifoliate, dan rataan RWC (Relative Water
Content) daun diambil pada hari ke-16 karena pada hari ke-16 leguminosa S.
guianensis dipanen pada hari ke-16.
Hasil dari penelitian ini bahwa leguminosa S. guianensis tidak tahan akan
kekeringan ganda, hal ini karena S. guianensis lebih awal dipanen karena S.
guianensis sudah terjadi masa pelayuan permanen. Kadar air tanah, rataan tinggi
tanaman, dan rataan bobot kering batang tidak memiliki perbedaan yang nyata
antara M1 dan M2 pada kedua tanaman, yang terjadi perbedaan yang nyata (P<0,05)
adalah antara M0 dengan M1 dan M2. Rataan pertambahan jumlah daun trifoliate
pada S. guianensis tidak memiliki perbedaan yang nyata pada semua perlakuan
sedangkan untuk S. scabra memiliki perbedaan yang nyata (P<0,05). Rataan RWC
pada S. guianensis memiliki perbedaan yang nyata (P<0,05) antara perlakuan M1
dengan M2 dengan nilai M2 yang lebih besar sedangkan pada S. scabra tidak
memiliki perbedaan yang nyata. Rataan bobot kering daun pada S. guianensis tidak
memiliki perbedaan yang nyata antara M1 dengan M2 sedangkan pada S. scabra
memiliki perbedaan yang nyata (P<0,05) dengan nilai M2 lebih besar dibandingkan
dengan M1.
Kata-kata kunci : Stylosanthes guianensis, Stylosanthes scabra, stres kekeringan
H2O2

iii
ABSTRACT
Comparison between Leguminose Stylosanthes guianensis and Stylosanthes
scabra Toward the Threat of Drought and the Distribution
of Hydrogen Peroxide (H2O2)
S. Sumaryani, P. D. M. H. Karti, and W. Hermana.
Feed is the major factor that greatly affects the livestock production animals. One
most needed in the livestock feed is forage, where forage consists of grass and
legume. Legume also has good amount of nutrition in it. This research used legume
Stylosanthes scabra and Stylosanthes guianensis that are drought resistant. This
research was given crop drought stress test for Stylosanthes scabra and Stylosanthes
guianensis, spraying treatment with H2O2 also given for their. The aim of this
research was to determined how resistant Stylosanthes guianensis and Stylosanthes
scabra to drought stress with H2O2 spraying. This research used a completely
randomized design with three treatment: M0 = with water and without sprayed H2O2
(control), M1 = drought stress and without sprayed H2O2 (drought stress), and M2 =
drought stress and sprayed H2O2 (double stress). Parameters observed were the soil
moisture content, the average height increament, the average of the number of
trifoliate leaves, the average relative water content of leaf, the average dry weight of
leaf production, the average dry weight of stem, and the average dry weight of roots.
The results of this research showed that M0 treatment produced the best products and
growth sompare with the other treatments and Stylosanthes guianensis had not
endured with double stress but for Stylosanthes scabra had endured with double
stress.
Keywords : Stylosanthes guianensis, Stylosanthes scabra, drought stress, H2O2

iv
PERBANDINGAN ANTARA LEGUMINOSA Stylosanthes
guianensis DAN Stylosanthes scabra TERHADAP
CEKAMAN KEKERINGAN DAN PEMBERIAN
HIDROGEN PEROKSIDA (H2O2)
SRI SUMARYANI
D24080356
Skripsi ini merupakan salah satu syarat untuk
memperoleh gelar Sarjana Peternakan pada
Fakultas Peternakan
Institut Pertanian Bogor
DEPARTEMEN ILMU NUTRISI DAN TEKNOLOGI PAKAN
FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR
2012

v
Judul : Perbandingan antara Leguminosa Stylosanthes Guianensis
dan Stylosanthes Scabra terhadap Cekaman Kekeringan dan Pemberian
Hidrogen Peroksida (H2O2)
Nama : Sri Sumaryani
NIM : D24080356
Menyetujui,
Pembimbing Utama, Pembimbing Anggota,
(Dr. Ir. Panca Dewi M.H.K., M. S) (Ir. Widya Hermana, M.Si.)
NIP : 19611025 198703 2 002 NIP :19680110 199203 2 001
Mengetahui,
Ketua Departemen
Ilmu Nutrisi dan Teknologi Pakan
(Dr. Ir. Idat Galih Permana, M.Sc. Agr.)
NIP. 19670506 199103 1 001
Tanggal Ujian : 10 Agustus 2012 Tanggal Lulus :

vi
RIWAYAT HIDUP
Penulis dilahirkan pada 1 Februari 1990 di Bekasi.
Penulis merupakan anak keempat dari lima bersaudara dari
pasangan Bapak Sudjono dan Ibu Suryati.
Studi pertama Penulis di SDN Pondok Kelapa 1
Bekasi Barat. Setelah lulus, Penulis melanjutkan studinya
di Sekolah Menengah Pertama Negeri 139 Jakarta Timur
dari tahun 2002 sampai 2005 dan pada tahun 2008 Penulis
menyelesaikan studinya di Sekolah Menengah Umum
Negeri 71 Jakarta Timur. Penulis diterima di Institit
Pertanian Bogor pada tahun 2008 melalui jalur Seleksi Nasional Masuk Perguruan
Tinggi Negeri (SNMPTN). Setelah melewati Tingkat Persiapan Bersama selama satu
tahun, Penulis masuk di Departemen Ilmu Nutrisi dan Teknologi Pakan, Fakultas
Peternakan, Institut Pertanian Bogor pada tahun 2009.
Selama menjalankan studinya di IPB, Penulis masuk di Badan Eksekutif
Mahasiswa Peternakan (BEM-D). Tahun pertama di BEM-D Penulis menjadi
Anggota Biro Kewirausahaan. Penulis juga mengikuti Ekstrakurikuler Teater
Kandang, Fakultas Peternakan, Institut Pertanian Bogor. Penulis juga sering
mengikuti acara-acara yang dilaksanakan oleh Badan Eksekutif Mahasiswa Fakultas
Peternakan sebagai panitia.
Bogor, Agustus 2012
Sri Sumaryani
D24080356

vii
KATA PENGANTAR
Bismillahirrahmanirrohim.
Alhamdulillahiraabil’alamin.
Penulis panjatkan puji syukur kehadirat Allah SWT yang telah memberikan
nikmat iman, islam, kesehatan serta karunia-Nya kepada Penulis sehingga Penulis
dapat menyelesaikan penulisan skripsi ini. Shalawat serta salam Penulis curahkan
kepada junjungan Nabi Muhammad SAW, keluarga, sahabat, dan pengikutnya yang
senantiasa istiqomah menjalankan sunnahnya.
Skripsi yang berjudul “Perbandingan antara Leguminosa Stylosanthes
guianensis dan Stylosanthes scabra terhadap Cekaman Kekeringan dan Pemberian
Hidrogen Peroksida (H2O2)” merupakan salah satu syarat untuk mendapatkan gelar
sarjana peternakan di Fakulas Peternakan, Institut Pertanian Bogor. Penulis berharap
semoga penulisan skripsi ini dapat memberikan tambahan ilmu bagi para pembaca
khususnya yang bergerak dibidang peternakan sehingga akan lebih berpengalaman.
Penulis menyadari masih banyak kekurangan dalam penulisan skripsi ini,
namun Penulis berharap tulisan ini dapat bermanfaat bagi para pembaca yang ingin
mengetahui tentang hal yang terkait dengan judul skripsi ini.
Bogor, Agustus 2012
Penulis

viii
DAFTAR ISI
Halaman
RINGKASAN .............................................................................................. ii
ABSTRACT ................................................................................................. iii
LEMBAR PERNYATAAN ......................................................................... iv
LEMBAR PENGESAHAN ......................................................................... v
RIWAYAT HIDUP ..................................................................................... vi
KATA PENGANTAR ................................................................................. vii
DAFTAR ISI ................................................................................................ viii
DAFTAR TABEL............ ............................................................................ x
DAFTAR GAMBAR ................................................................................... xi
DAFTAR LAMPIRAN........................................................... ..................... xii
PENDAHULUAN........................................................... ............................ 1
Latar Belakang.................................................................... ............ 1
Tujuan .............................................................................................. 2
TINJAUAN PUSTAKA .............................................................................. 3
Stylosanthes guianensis ................................................................... 3
Stylosanthes scabra ......................................................................... 4
Hidrogen Peroksida ......................................................................... 5
Peranan Air pada Tanaman ............................................................. 7
Pengaruh Cekaman Kekeringan pada Tanaman ............................. 8
Relative Water Content ................................................................... 9
MATERI DAN METODE ........................................................................... 11
Lokasi dan Waktu ............................................................................ 11
Materi ............................................................................................... 11
Bahan ................................................................................. 11
Alat ..................................................................................... 11
Prosedur .......................................................................................... 11
Larutan Peroksida ............................................................. 11
Pemilihan Jenis Leguminosa ............................................ 11
Persiapan median Tanam .................................................. 11
Penanaman ........................................................................ 12
Perlakuan Kekeringan ....................................................... 12
Pemeliharaan ..................................................................... 13
Pengamatan dan Pengambilan Data Pertumbuhan ........... 13
Pemanenan ........................................................................ 13
Rancangan dan Analisis Data ......................................................... 13
Perlakuan .......................................................................... 13

ix
Rancangan ......................................................................... 14
Peubah yang Diamati ........................................................ 14
Jumlah Daun Trifoliate ......................................... 14
Tinggi Vertikal ...................................................... 14
Kadar Air Tanah .................................................... 14
Relative Water Content ......................................... 15
Bobot Kering Akar ................................................ 15
Bobot Kering Daun ............................................... 15
Bobot Kering Batang ............................................ 15
HASIL DAN PEMBAHASAN ................................................................... 17
Respon Tanaman terhadap Cekaman Kekerigan ............................ 17
Pengaruh Perlakuan terhadap Rataan KA Tanah ............................ 18
Pengaruh Perlakuan terhadap Laju Pertumbuhan Tinggi Vertikal
Tanaman .......................................................................................... 19
Pengaruh Perlakuan terhadap Rataan Selisih Jumlah Daun
Trifoliate Tanaman .......................................................................... 20
Pengaruh Perlakuan terhadap Rataan RWC Daun Tanaman .......... 20
Pengaruh Perlakuan terhadap Rataan Bobot Kering Daun, Batang,
dan Akar .......................................................................................... 21
Pembahasan Umum ......................................................................... 23
PENUTUP .................................................................................................. 26
Kesimpulan ...................................................................................... 26
Saran ................................................................................................. 26
UCAPAN TERIMA KASIH ....................................................................... 27
DAFTAR PUSTAKA .................................................................................. 28
LAMPIRAN ................................................................................................ 32

x
DAFTAR TABEL
Nomor Halaman
1. Pengaruh H2O2 terhadap Kadar Air Tanah, Pertumbuhan,
dan Produksi Leguminosa Stylosanthes guianensis pada Hari ke-
16 .................................................................................................... 18
2. Pengaruh H2O2 terhadap Kadar Air Tanah, Pertumbuhan,
dan Produksi Leguminosa Stylosanthes scabra pada Hari ke- 16 .. 18
3. Pengaruh H2O2 terhadap Pertumbuhan dan Produksi
Leguminosa Stylosanthes guianensis .............................................. 22
4. Pengaruh H2O2 terhadap Pertumbuhan dan Produksi
Leguminosa Stylosanthes scabra ..................................................... 22

xi
DAFTAR GAMBAR
Nomor Halaman
1. Leguminosa Stylosanthes guianensis ................................................... 3
2. Leguminosa Stylosanthes scabra ......................................................... 4
3. Stek Batang dalam Baki ...................................................................... 12
4. Tanaman yang Permukaan Tanahnya Ditutup dengan Baki ............... 13
5. Tanaman Menggugurkan Daun dan Mati ............................................ 17

xii
DAFTAR LAMPIRAN
Nomor Halaman
1. Perhitungan H2O2 ................................................................................ 33
2. Sidik Ragam Rataan Kadar Air Tanah Stylosanthes guianensis
sampai Hari ke- 16 ............................................................................... 33
3. Sidik Ragam Rataan Kadar Air Tanah Stylosanthes scabra sampai
Hari ke- 16 ........................................................................................... 33
4. Sidik Ragam Rataan Pertambahan Tinggi Tanaman Stylosanthes
guianensis sampai Hari ke- 16 ............................................................ 34
5. Sidik Ragam Rataan Pertambahan Tinggi Tanaman Stylosanthes
scabra sampai Hari ke- 16 ................................................................... 34
6. Sidik Ragam Rataan Pertambahan Jumlah Daun Trifoliate Tanaman
Stylosanthes guianensis sampai Hari ke- 16 ....................................... 34
7. Sidik Ragam Rataan Pertambahan Jumlah Daun Trifoliate Tanaman
Stylosanthes scabra sampai Hari ke- 16 .............................................. 34
8. Sidik Ragam Rataan RWC Daun Tanaman Stylosanthes guianensis
sampai Hari ke- 16 ............................................................................... 34
9. Sidik Ragam Rataan RWC Daun Tanaman Stylosanthes scabra
sampai Hari ke- 16 ............................................................................... 35
10. Sidik Ragam Rataan Produksi Bobot Kering Daun Tanaman
Stylosanthes guianensis ....................................................................... 35
11. Sidik Ragam Rataan Produksi Bobot Kering Daun Tanaman
Stylosanthes scabra ............................................................................. 35
12. Sidik Ragam Rataan Produksi Bobot Kering Batang Tanaman
Stylosanthes guianensis ....................................................................... 35
13. Sidik Ragam Rataan Produksi Bobot Kering Batang Tanaman
Stylosanthes scabra .............................................................................. 36
14. Sidik Ragam Rataan Produksi Bobot Kering Akar Tanaman
Stylosanthes guianensis ........................................................................ 36
15. Sidik Ragam Rataan Produksi Bobot Kering Akar Tanaman
Stylosanthes scabra .............................................................................. 36

PENDAHULUAN
Latar Belakang
Pakan merupakan faktor utama dalam peternakan yang mempengaruhi
produksi peternakan. Ruminansia memerlukan pakan hijauan yang dapat dipenuhi
dari rumput dan leguminosa. Menurut Rekohadiprodjo (1985) pemberian rumput
yang dikombinasikan dengan leguminosa sangat dianjurkan karena leguminosa
mempunyai banyak kandungan nutrisi yang lebih tinggi dari rumput seperti
kandungan protein kasar yang mencapai 15%-25%. Selain sebagai pakan ternak
leguminosa juga dapat berfungsi sebagai tanaman penutup tanah. Pada musim
kemarau panjang banyak tanaman yang tidak tahan dengan kekeringan, hal inilah
yang membuat tanaman leguminosa sulit untuk tumbuh sehingga pakan ternak tidak
tercukupi. Tanaman yang mengalami cekaman kekeringan pertumbuhannya
terhambat, karena ketersediaan air dalam tanaman dan tanah mempengaruhi transport
hara tanah oleh akar tanaman.
Cekaman kekeringan membuat kadar air tanah menurun hal inilah yang
menyebabkan tanaman menjadi kering dan terjadilah plasmolisis atau keluarnya air
dari sel. Plasmolisis terjadi karena potensial air pada tanah menurun sedangkan
potensial pada akar tetap/normal sehingga air dari tanah tidak dapat diserap ke
tanaman karena sifat air adalah mengalir dari potensial air yang tinggi ke potensial
air yang lebih rendah (Gardber et al., 1991). Penelitian ini menggunakan leguminosa
Stylosanthes yang tanaman yang tahan akan kekeringan dan penelitian kali ini
menggunakan Stylosanthes guianensis dan Stylosanthes scabra.
Stres kekeringan dapat meningkatkan produksi Reactive Oxygen Species
(ROS) dalam kompartemen seluler seperti kloroplas, peroksisom, dan mitokondria.
Secara umum hal tersebut dapat diterima bahwa pemberian stres lingkungan seperti
kekeringan, dingin, panas, atau iradiasi cahaya yang tinggi, menimbulkan konsentrasi
ROS yang tinggi seperti superoksida, H2O2, singlet oxygen, dan radikal hidroksil
(Bowler et al., 1992; Foyer et al., 1994; Alscher et al., 1997; Shigeoka et al., 2002).
Penelitian ini menggunakan H2O2 dimana H2O2 merupakan salah satu cara untuk
membuat tanaman menjadi stres atau mensimulasi stres cahaya seperti yang
dijabarkan di atas sehingga dengan ini tanaman memiliki stres kekeringan ganda
selain stres kekeringan air juga dapat stres dari H2O2.

2
Tujuan
Penelitian ini bertujuan untuk mempelajari ketahanan pada leguminosa S.
guianensis dengan S. scabra terhadap perlakuan stres kekeringan air dengan stres
kekeringan air ditambah stres kekeringan karena pengaruh penambahan H2O2.

3
TINJAUAN PUSTAKA
Stylosanthes guianensis
Stylosanthes guianensis merupakan tanaman legum perenial, tingginya dapat
mencapai 1,2 m. Daunnya trifoliate dengan panjang 0,5-4,5 cm dan lebar 0,2-2 cm,
bunganya berwarna kuning sampai orange, benihnya berwarna coklat (bervariasi dari
kuning sampai agak kehitaman). Tanaman ini lebih dikenal dengan nama
stylosanthes yang digunakan untuk tanaman pakan pada lahan pastura
(penggembalaan maupun potongan), sebagai penutup tanah (mencegah erosi) pupuk
hijau, dan diolah menjadi hay atau pellet. Stylosanthes guianensis dapat tumbuh pada
tanah dengan drainase yang baik, dan pada tekstur tanah dari pasir sampai liat
(seperti pada tanah tropis latosol, liat, tanah berpasir, dan podsolik asam) (FAO,
2012).
Stylosanthes guianensis dapat tumbuh pada pH tanah berkisar 4,0-8,3 dan
toleran terhadap kandungan Al dan Mn yang tinggi namun tidak pada salinitas yang
tinggi. S. guianensis dapat memanfaatkan P pada tanah dengan kandungan P yang
rendah, namun dapat dengan baik merespon pemberian P, K, S, Ca, dan Cu pada
taraf yang rendah (FAO, 2012).
Gambar 1. Leguminosa Stylosanthes guianensis Sumber: www.informedfarmers.com [17 Juni 2012]
Menurut Mannetje dan Jones (1992), S. guianensis sangat responsif terhadap
pemupukan fosfor (P), dengan pemupukan fosfor dapat meningkatkan BK sebesar
0,06%. Produksi BK S. guianensis berkisar antara 5-10 ton/ha tergantung dari
penanaman, pertumbuhan, dan manajemen pemeliharaan, S. guianensis dapat
mencapai produksi tertinggi sebesar 20 ton/ha. Produksi benih berkisar antara 700-
1.350 kg/ha. Stylosanthes guienensis dapat dikembangkan secara generatif (benih)

4
dan vegetatif (pemotongan) namun pertumbuhannya lambat dan mahal. Produksi
benih sangat sulit didapatkan karena benih yang sudah matang lepas pada saat awal
berbunga, selain itu cairan lengket yang dikeluarkan pada kepala bunga membuat
proses pemanenan semakin sulit (Bogdan, 1977). Nilai nutrisi S. guianensis adalah
sebagai berikut PK 12-20%, kecernaan BKIV 52-60 %, P 0,2-0,6%, Ca 0.6-1.6%
(Chakraborty, 2004), konsentrasi nitrogen 1,5%-3,0%, berat kering yang dapat
dicerna dari tanaman yang muda 60%-70% (PROSEA, 1992).
Stylosanthes scabral
Suatu tanaman semak tahunan tinggi sampai 2 m, dengan akar tunggang yang
kuat dan dalam (sampai 4 m). Batang muda bervariasi dari warna hijau sampai
merah, tergantung dari tipe; biasanya dengan bulu-bulu yang padat dan kasar,
menjadi lebih berkayu seiring umur tanaman. Helai daun berbulu pada kedua
permukaan, berwarna hijau pucat sampai hijau tua dan hijau kebiruan, panjang 20-33
mm dan lebar 4-12 mm. Bunga berwarna kuning pucat sampai kuning tua. Buah
polong dengan 2 segmen, kedua segmen biasanya subur; segmen bagian atas panjang
4-5 mm dan segmen bagian bawah panjang 2 mm, coklat pucat sampai coklat muda.
400.000-500.000 biji dalam buah polong/kg dan 600.000-800.000 biji bersih/kg
(PROSEA, 1992).
Gambar 2. Leguminnosa Stylosanthes scabra Sumber: www. Wikipedia.org [17 Juni 2012]
Stylosanthes scabra biasanya digunakan sebagai padang gembala tahunan,
ditanam bersama dengan rumput unggul dan rumput alam. Digunakan sebagai
tanaman potong angkut pada beberapa negara. Tanaman muda mungkin cocok untuk

5
diawetkan. Tumbuh dengan baik pada tanah pasir tidak subur, asam dan mudah
menyusut atau keras; demikian pula tumbuh dengan baik pada tanah dengan tekstur
lebih berat, sedikit asam, dan tidak cocok sama sekali pada semua jenis tanah liat
berat (PROSEA, 1992).
Spesies yang sangat tahan kekeringan, tumbuh pada daerah dengan curah
hujan rendah sampai 350 mm/tahun. Dalam penanamannya, tanaman ini terutama
digunakan pada daerah dengan curah hujan tahunan sekitar 600 dan 2000 mm.
Musim kering yang panjang dapat menjadi faktor pembatas pada daerah dengan
curah hujan rendah dan tanah yang lebih dangkal, dimana tanaman semusim, atau
tanaman tahunan dengan kemampuan berperilaku sebagai tanaman semusim
(misalnya S. hamata ), biasanya lebih berhasil. Pertumbuhan bibit biasanya terlalu
lambat pada S. scabra karena tanaman ini berperilaku sebagai tanaman semusim.
Beberapa tipe tidak tahan terhadap penggenangan air (PROSEA, 1992)
Nilai nutrisi S. scabra menurun seiring umur tanaman, PK daun dari 20%
menjadi 10%, P dari 0,3% menjadi 0,1% dan Kecernaan bahan kering in vitro dari
70% menjadi 50%. Proporsi batang meningkat bersama umur, dari sekitar 20% pada
pertumbuhan awal menjadi 75% pada akhir musim (dan lebih tinggi pada padang
gembala yang digembalai) (Edye dan Toprark-Ngarm, 1992).
Hidrogen Peroksida (H2O2)
Hidrogen peroksida dengan rumus kimia H2O2 ditemukan oleh Louis Jacques
Thenard di tahun 1818. Senyawa ini merupakan bahan kimia anorganik yang
memiliki sifat oksidator kuat. Bahan baku pembuatan hidrogen peroksida adalah gas
hidrogen (H2) dan gas oksigen (O2). Teknologi yang banyak digunakan di dalam
industri hidrogen peroksida adalah auto oksidasi Anthraquinone (Patnaik, 2002)
Hidrogen peroksida tidak berwarna, berbau khas agak keasaman, dan larut
dengan baik dalam air. Dalam kondisi normal (kondisi ambient), hidrogen peroksida
sangat stabil dengan laju dekomposisi kira-kira kurang dari 1% per tahun. Mayoritas
pengunaan hidrogen peroksida adalah dengan memanfaatkan dan merekayasa reaksi
dekomposisinya, yang intinya menghasilkan oksigen. Pada tahap produksi hidrogen
peroksida, bahan stabilizer kimia biasanya ditambahkan dengan maksud untuk
menghambat laju dekomposisinya. Termasuk dekomposisi yang terjadi selama
produk hidrogen peroksida dalam penyimpanan. Selain menghasilkan oksigen, reaksi

6
dekomposisi hidrogen peroksida juga menghasilkan air (H2O) dan panas.
Penambahan H2O2 dapat meningkatkan konsentrasi oksigen terlarut, spesimen H2
memiliki sifat oksidator dan H+ memiliki sifat reduktor (Andayani dan Sumartono,
1999).
Stres kekeringan dapat meningkatkan produksi Reactive Oxygen Species
(ROS) dalam kompartemen seluler seperti kloroplas, peroksisom, dan mitokondria.
Secara umum hal tersebut dapat diterima bahwa pemberian stres lingkungan seperti
kekeringan, dingin, panas, atau iradiasi cahaya yang tinggi, menimbulkan konsentrasi
ROS yang tinggi seperti superoksida, H2O2, singlet oxygen, dan radikal hidroksil
(Bowler et al., 1992; Foyer et al., 1994; Alscher et al., 1997; Shigeoka et al., 2002).
Radikal bebas juga bisa berasal dari konsumsi oksigen, 2%-3% oksigen yang
dikonsumsi akan dikonversi menjadi oksigen radikal (O-) dan H2O2 dan peningkatan
konsumsi oksigen pada jaringan akan menghasilkan Reactive Oxygen Species (ROS)
pada mitokondria, demikan juga dengan peningkatan suhu akan menghasilkan ROS
yang akan menyebabkan kerusakan oksidatif (Abele et al., 1998).
Banyak faktor fisiologis yang terlibat dalam stres kekeringan ataupun panas
yang dapat membuat tanaman menjadi sakit. Dalam beberapa spesies, stres panas dan
kekeringan dapat menyebabkan stres oksidatif, yang dihasilkan dari produksi dan
akumulasi spesies oksigen beracun seperti radikal superoksida, hidrogen peroksida
(H2O2), dan radikal hidroksil (OH-) (Bowler et al., 1992; Inze dan Montagu, 1995).
Spesies oksidatif yang dihasilkan selama stres dapat merusak komponen sel,
termasuk lipid, protein, karbohidrat, dan asam nukleat (Monk et al., 1989). Stres
oksidatif dapat menyebabkan penghambatan proses fotosintesis dan respirasi,
termasuk pertumbuhan tanaman. Tanaman telah mengevolusi enzimatis dan bukan
enzimatis untuk menolak spesies oksigen aktif. Dalam sistem enzimatik misalnya,
superoxide dismutase (SOD) mengkatalisis dismutasi O-2 untuk H2O2 dan O2.
Kalatase (CAT) dan peroksida askorbat (AP) dapat memecah H2O2. Glutation
reduktase (GR) juga dapat menghapus H2O2 melalui siklus askorbat-glutathione
untuk mempertahankan tingkat askorbat yangg tinggi berkurang dalam kloroplas.
Namun fungsi dari enzim pengais ini dapat terganggu oleh stres kekeringan dan
panas, yang dapat meningkatnya peroksidasi lipid dan kerusakan membran

7
konsekuen (Chowdhury dan Choudhuri, 1985; Zhang dan Kirkham, 1994; Jagtap dan
Bhargava, 1995, Dat et al., 1998).
Peranan Air pada Tanaman
Harjadi dan Yahya (1988) menerangkan bahwa peranan air antara lain: (1) air
merupakan bagian yang esensial bagi protoplasma dan membentuk 80%-90% bobot
segar jaringan yang tumbuh aktif, (2) air adalah pelarut, di dalamnya terdapat gas-
gas, garam, dan zat-zat terlarut lainnya yang bergerak keluar, (3) air adalah pereaksi
dalam proses fotosintesis dan proses hidrolisis, (4) air esensial untuk menjaga
turgiditas diantaranya dalam pembesaran sel, pembukaan stomata, dan menyangga
bentuk (morfologi) daun-daun atau struktur lainnya yang berlignin sedikit.
Menurut Noggle dan Frizt (1983) menjelaskan fungsi air bagi tanaman yaitu
sebagai: (1) senyawa utama pembentuk protoplasma, (2) senyawa pelarut bagi
masuknya mineral-mineral dari larutan tanah ke tanaman dan sebagai pelarut mineral
nutrisi yang akan diangkut dari satu bagian sel ke bagian sel lain, (3) media
terjadinya reaksi-reaksi metabolik, (4) reaktan pada sejumlah reaksi metabolisme
seperti siklus asam trikarboksilat, (5) penghasil hidrogen pada proses fotosintesis, (6)
penjaga turgiditas sel dan berperan sebagai tenaga mekanik dalam pembesaran sel,
(7) pengatur mekanisme gerakan tanaman seperti membuka dan menutupnya
stomata, membuka dan menutupnya bunga, serta melipatnya daun-daun tanaman
tertentu, (8) berperan dalam perpanjangan sel, (9) bahan metabolisme dan produk
akhir respirasi, serta (10) digunakan dalam proses respirasi. Kebutuhan air pada
tanaman dipengaruhi oleh beberapa faktor antara lain jenis dan umur tanaman, kadar
air tanah dan kondisi cuaca.
Setiap gram pembentukan bahan organik penyusun tanaman, rata-rata
membutuhkan 500 g air yang diabsorbsi oleh akar ditranportasikan ke seluruh bagian
tanaman dan selanjutnya air akan hilang ke atmosfer. Setiap tanaman harus dapat
menyeimbangkan antara proses kehilangan air dan proses penyerapannya, bila proses
kehilangan air tidak diimbangi dengan penyerapan melalui akar maka akan terjadi
kekurangan air di dalam sel tanaman yang dapat menyebabkan berbagai kerusakan
pada banyak proses dalam sel tanaman (Taiz dan Zeiger, 2002).
Kadar air dalam tanah mendeskripsikan jumah sumber air yang tersedia,
dimana air dapat diserap untuk pertumbuhan, sedangkan kekeringan dapat

8
menyebabkan air tidak tersedia dan tanaman menjadi menderita dan layu (Karti,
2004). Karti et al. (2012) bahwa tanaman yang diberikan stres kekeringan dapat
menurunkan kadar air tanah dibandingkan dengan tanah yang disiram setiap hari.
Pengaruh Cekaman Kekeringan pada Tanaman
Menurut Mcllroy (1976), stres kering pada tanaman dapat menyebabkan
terjadinya peubahan struktur dan konfigurasi protein, sehingga aktifitas enzim dan
laju metabolism menurun. Pada daun sendiri stress kering dapat menyebabkan
stomata menutup, hal ini terjadi karena turgiditas sel-sel penyangga menurun.
Cekaman air pada tanaman terjadi karena ketersediaan air dalam media tidak cukup
dan transpirasi yang berlebihan (Islami dan Utomo, 1995).
Cekaman kekeringan terjadi ketika ketersediaan air tanah menurun dan
kondisi atmosfir menyebabkan kehilangan air terus menerus melalui transpirasi atau
evaporasi (Jaleel et al., 2009). Lebih lanjut dijelaskan bahwa cekaman kekeringan
ditandai dengan rendahnya kadar air, penurunan potensial air daun dan tekanan
turgor, penutupan stomata dan berkurangnya pembesaran dan pertumbuhan sel.
Reaksi tanaman terhadap cekaman kekeringan berbeda secara signifikan pada
berbagai tingkatan tergantung pada intensitas dan durasi dari cekaman itu sendiri,
dan juga species tanaman dan tingkatan pertumbuhannya (Chaves et al., 2002) .
Cekaman kekeringan sebelum berpengaruh terhadap pertumbuhan dan hasil
tanaman, terlebih dahulu mengakibatkan dehidrasi dan menurunkan tekanan turgor
sel tanaman, sehingga merangsang penutupan stomata, menghambat difusi CO2 dan
fotosintesis. Akar yang mengalami cekaman kekeringan akan membentuk asam
absisat lebih banyak dan diangkut melalui xylem menuju daun untuk menutup
stomata, yaitu dengan cara menghambat pompa proton yang kerjanya tergantung
pada ATP dan membran plasma sel penjaga (Salisbury, 1995).
Kekeringan dapat menyebabkan daun menjadi lebih sempit dan pendek,
demikian pula batang dan organ reproduktif yang terbentuk lebih kecil dari ukuran
normal (Teare dan Peet, 1983). Cekaman kekeringan dan panas dapat menyebabkan
menurunnya kualitas tanah, dengan menurunnya kualitas tanah maka dapat
menurunkan pertumbuhan pada akar, potensial air daun, stabilitas membran sel,
kecepatan fotosintesis, dan akumulasi karbohidrat (Howard dan Watschke, 1991;
Carrow, 1996; Perdomo et al., 1998; Huang dan Gao, 1999; Jiang dan Huang, 2000)

9
Cekaman kekeringan, panas maupun keduanya dapat menurunkan kualitas
tanah, Relative Water Content (RWC), dan kandungan klorofil daun. Stres ganda
efeknya lebih dapat merusak dibandingkan dengan hanya stres kering saja maupun
stres panas saja. Kualitas tanah menurun secara dramatis dibawah cekaman
kekeringan, sedangkan cekaman ganda dapat menurunkan nilai RWC selama
perlakuan stres (Turner et al., 1966; Nobel, 1988).
Relative Water Content (RWC)
Relative Water Content dapat digunakan dalam seleksi langsung terhadap
resisten kekeringan (Yuniaty, 1998; Reynolds et al., 1999; Chandrasekar et al.,
2000). Relative Water Content adalah ukuran status tanaman air yang mencerminkan
juga bervariasi dalam potensial air, potensi turgor dari penyediaan osmotik (Blum,
1999). Relative Water Content daun dapat dengan mudah untuk ditentukan.
Mempertahankan Relative Water Content tetap tinggi dapat dilakukan melalui tiga
mekanisme (1) kapasitas untuk mempertahankan potensi air yang tinggi dengan
ekstraksi kelembaban tanah yang mendalam, (2) kapasitas penyesuaian osmotik,
yang dapat memungkinkan mempertahankan turgor RWC dan potensi air yang lebih
rendah, dan (3) penutupan stomata dalam menanggapi pengeringan daun dan
mengangkut sinyal hormon yang diproduksi oleh akar dan respon terhadap
pengeringan akar. Yuniaty (1998) juga menyatakan bahwa RWC daun memiliki
asosiasi tingkat tertinggi dengan hasil biji kedelai dengan berbagai parameter yang
diamati (tingkat transpirasi, difusif resistensis, dan suhu daun). Relative Water
Content daun kurang dipengaruhi oleh faktor-faktor lingkungan sejak pengukuran
yang dilakukan pada pagi hari dimana variasi atmosfer kurang diucapkan. Relative
Water Content berkaitan erat dengan volume sel, mungkin lebih mencerminkan
keseimbangan antara pasokan air ke daun dan laju transpirasi (Schonfeld et al.,
1998). Hal ini dapat mempengaruhi waktu tanaman tua pulih dari stres dan akibatnya
dapat mempengaruhi stabilitas hasil (Lilley dan Ludlow, 1996).
Relative Water Content merupakan indikator yang yang sangat penting untuk
keadaan keseimbangan air pada tanaman, RWC menerangkan jumlah absolut air,
dimana tanaman membutuhkannya untuk mengetahui kandungan turgor air.
(González dan González-Vilar, 2001). Relative Water Content mengekspresikan

10
dalam persentase kandungan air pada waktu tertentu dan jaringan yang terkait
dengan kandungan air turgor (Slatyer, 1967).

11
MATERI DAN METODE
Lokasi dan Waktu
Penelitian ini dilaksanakan pada bulan November 2011 sampai bulan Maret
2012, bertempat di Laboratorium Lapang Agrostologi, Laboratorium Ilmu Nutrisi
Ternak Perah, Laboratorium Agrostologi, Fakultas Peternakan, Institut Pertanian
Bogor.
Materi
Bahan
Bahan-bahan penelitian yang digunakan adalah tanaman leguminosa
Stylosanthes guianensis, Stylosanthes scabra, tanah, pupuk NPK mutiara, H2O2 30%,
dan aquadest.
Alat
Peralatan yang digunakan untuk penelitian ini adalah sekop, timbangan
kapasitas 5 kg, plastik, polybag 2 kg, gunting, timbangan digital, penggaris, sprayer,
oven, kertas untuk mengoven/amplop, saringan, pinset, pipet, cup, kertas label,
wadah plastik, plastik, dan spidol.
Prosedur
Larutan Peroksida
Pembuatan larutan peroksida 1mM menggunakan peroksida sebanyak 0,245
ml dan aquadest sebanyak 2,4 liter. Pemberian hidrogen peroksida sebanyak 100 ml
untuk setiap tanaman.
Pemilihan Jenis Leguminosa
Jenis leguminosa yang dipakai pada penelitian ini adalah jenis legum yang
merupakan jenis tanaman pakan yang biasa dimanfaatkan sebagai pakan ternak yaitu
Stylosanthes guianensis dan Stylosanthes scabra.
Persiapan Media Tanam
Media tanam yang digunakan adalah tanah sebanyak 2 kg yang dicampurkan
dengan pupuk NPK mutiara sebanyak 2 gram. Sebelum dimasukkan ke dalam
polybag 2 kg, persiapan tanam ini melakukan stek batang (Gambar. 3) untuk kedua

12
tanaman terlebih dahulu di dalam baki yang tertutup rapat dengan plastik setelah
tumbuh akar dan tumbuh cukup baik tanaman dimasukkan ke dalam polybag 2 kg.
Gambar 3. Stek Batang dalam Baki Sumber: Dokumentasi Penelitian
Penanaman
Stylosanthes guianensis dan Stylosanthes scabra masing-masing ditanam di
dalam polybag berkapasitas 2 kg tanah. Penanaman dilakukan dengan cara
memasukkan satu tanaman legum ke dalam polybag yang sudah tersedia yang
sebelumnya sudah disiram terlebih dahulu. Tanaman ditumbuhkan selama satu bulan.
Setelah ditumbuhkan selama satu bulan maka dapat dimulai perlakuan.
Perlakuan Kekeringan
Sebelum perlakuan kekeringan dimulai, semua polybag disiram terlebih
dahulu sampai tercipta kondisi jenuh. Polybag untuk perlakuan M1 dan M2 ditutup
permukaan tanahnya dengan menggunakan plastik dengan rapat (gambar. 4),
sehingga tidak ada air dari luar yang masuk juga penguapan air. Perlakuan dimulai
pada keesokan harinya dan dihitung sebagai hari ke-0 (H0), untuk polybag perlakuan
M0 (kontrol) yang tidak disemprotkan dengan H2O2 dilakukan penyiraman setiap
pagi, untuk perlakuan M1 (stres kekeringan tunggal) yang tidak diberikan
penyemprotan dengan H2O2 dan tidak dilakukan penyiram, sedangkan untuk
perlakuan M2 (stres kekeringan ganda) yang disemprotkan H2O2 dan tidak disiram
setiap hari.

13
Gambar 4. Tanaman yang Permukaan Tanah Ditutup dengan Plastik Sumber: Dokumentasi Penelitian
Pemeliharaan
Pemeliharaan tanaman pada penelitian ini adalah dengan dilakukan
penyiraman dan pencabutan gulma. Penyiraman dilakukan satu kali sehari yaitu pada
pagi hari. Pembersihan gulma dilakukan secara manual yaitu dengan cara mencabut
gulma setiap minggu.
Pengamatan dan Pengambilan Data Pertumbuhan
Pengamatan dan pengambilan data pertumbuhan setiap delapan hari sekali
dengan mengukur tinggi tanaman, jumlah daun trifoliate, mengukur jumlah daun,
mengukur Relative Water Content (RWC), daun dan bobot kering daun serta
pengambilan sampel tanah untuk mengukur kadar air tanah.
Pemanenan
Pemanenan dilakukan pada tanaman yang sudah kering dan mati setelah
diberi perlakuan kekeringan, kemudian dilakukan pengamatan dan pengambilan
sampel untuk analisa beberapa peubah.
Rancangan dan Analisis Data
Perlakuan
Perlakuan yang diberikan pada penelitian ini adalah:
M0 : kontrol
M1 : stres kekeringan tunggal
M2 : stres kekeringan ganda

14
Rancangan
Rancangan percobaan yang digunakan dalam penelitian ini adalah Rancangan
Acak Lengkap (RAL) dengan empat kali ulangan.
Model analisis menggunakan model matematik sebagai berikut:
Yij = µ + τi + εij
Keterangan :
Yij : Pengamatan pada perlakuan ke-i dan ulangan ke-j
µ : Nilai rataan umum dari pengamatan
τi : Pengaruh perlakuan ke-i
εij : Pengaruh acak pada perlakuan ke-i dan ulangan ke-j
Analisis Data
Data yang terkumpul di analisis dengan sidik ragam ANOVA (Steel dan
Torrie, 1995) dan dilanjutkan dengan uji Duncan.
Peubah yang Diamati
Peubah yang diamati dalam penelitian ini antara lain yaitu jumlah daun
trifoliate, tinggi vertikal, kadar air tanah, RWC, bobot kering daun, batang, dan akar.
Jumlah Daun Trifoliate. Penghitungan jumlah daun trifoliate dilakukan dengan
menghitung jumlah daun trifoliate yang masih utuh. Penghitungan jumlah daun
trifoliate dimulai sejak perlakuan dan diamati setiap delapan hari sekali selama
penanaman hingga pemanenan.
Tinggi Vertikal. Setiap perlakuan tanaman leguminosa diukur tinggi vertikalnya
diukur dari permukaan tanah hingga pucuk daun tertinggi.
Kadar Air Tanah (KA Tanah). Sampel tanah diambil pada masing-masing
tanaman yang diambil dari tengah tanah dengan menggunakan spatula kecil
kemudian sampel tanah lalu ditimbang beratnya. Sampel tanah tersebut kemudian
dimasukkan ke dalam oven 105ºC selama 24 jam. Setelah itu timbang berat sampel
setelah dioven. Kadar air didapat dari berat sampel sebelum dimasukkan ke oven
dikurangi berat sampel setelah dioven kemudian dibagi berat sampel sebelum
dimasukkan ke oven dan dikalikan 100%. Pengukuran dilakukan setiap delapan hari
sekali (0, 8, 16, dan seterusnya).

15
Perhitungan kadar air tanah adalah sebagai berikut :
KA tanah (%) = W0 – Wt x 100%
W0
Keterangan :
W0 = berat sampel tanah basah
Wt = berat sampel tanah kering oven
Relative Water Content. Hal pertama yang harus dilakukan untuk mengetahui
nilai RWC daun adalah dengan mengumpulkan potongan daun dari 3-4 daun lalu
ditimbang berat daun tersebut dinamakan berat segar, setelah itu tanaman direndam
kedalam cup lalu ditutup rapat. Daun dalam cup setelah 1 x 24 jam langsung
ditiriskan lalu ditimbang sebagai berat turgor, setelah itu daun dimasukkan kedalam
amplop dan dikeringkan kedalam oven 60oC selama 3 x 24 jam atau selama tiga hari.
daun yang telah di oven selama tiga hari lalu ditimbang sebagai berat kering.
Perhitungan RWC adalah sebagai berikut :
RWC = (FW-DW) x 100%
(TW-DW)
Keterangan :
RWC : Relative Water Content
FW : berat segar
DW : berat kering
TW : berat turgor
Bobot Kering Daun. Sampel daun diambil lalu ditimbang sebagai berat segar
selanjutnya daun dikeringudarakan selama satu hari kemudian dioven 60oC selama
3x24 jam lalu ditimbang beratnya sebagai bobot kering.
Bobott Kering Batang. Sampel batang diambil pada saat panen lalu ditimbang
untuk dicari berat segar setelah itu dikeringudarakan selama sehari kemudian dioven
60°C selama 3x24 jam untuk mendapatkan bobot kering batang.
Bobot Kering Akar. Bobot kering akar diukur pada saat panen. Akar dipisahkan
dari tanah yang menempel dengan cara dibilas dengan air bersih, setelah itu
ditiriskan hingga tidak ada air yang menetes, kemudian akar ditimbang sehingga

16
didapat berat segar akar. Setelah itu akar dikering udarakan selama satu hari
kemudian di oven 60°C selama 3x24 jam setelah itu ditimbang beratnya sebagai
bobot kering.

17
HASIL DAN PEMBAHASAN
Respon Tanaman terhadap Cekaman Kekeringan
Dua jenis legum yang digunakan pada penelitian ini setelah diberikan
perlakuan atau cekaman kekeringan menyebabkan terjadinya banyak perubahan
morfologi. Pengaruh morfologi akibat cekaman kekeringan biasanya sangat
tergantung pada faktor waktu terjadinya cekaman dan besarnya perlakuan cekaman
(Keles dan Oncel, 2002). Salah satu perubahan morfologi cekaman kekeringan
adalah tanaman menjadi layu dan kering, tanaman menggugurkan daunnya, serta
mati, seperti yang diperlihatkan pada Gambar 5.
Gambar 5. Tanaman Mengugurkan Daun dan Mati Sumber: Dokumentasi Peneitian
Reaksi tanaman terhadap cekaman kekeringan berbeda secara signifikan pada
berbagai tingkatan tergantung pada intensitas dan durasi dari cekaman itu sendiri,
dan juga species tanaman dan tingkatan pertumbuhannya (Chaves et al., 2002).
Ketika jumlah absorbsi air mulai terbatas, maka tanaman memiliki mekanisme untuk
mencegah kehilangan air dengan melakukan penutupan stomata (Taiz dan Zeiger,
2002).
Kedua tanaman memiliki respon yang berbeda pada penelitian ini setelah
diberikan perlakuan. Kedua tanaman yang mendapatkan perlakuan stres kekeringan
tunggal dan stres kekeringan ganda menunjukkan respon dengan adanya daun yang
gugur, kelayuan pada tanaman hingga kondisi tanaman mencapai titik layu permanen
dibandingkan dengan tanaman yang disiram setiap hari.
Penelitian ini menunjukkan bahwa leguminosa S. guianensis dipanen pada
hari ke-16 karena tanaman sudah memasuki titik layu permanen. Pemanenan legum
S. scabra dilakukan pada hari ke-24 dimana tanaman yang diberikan perlakuan

18
mengalami titik layu permanen pada hari ke-24. Hal ini dapat dikatakan bahwa S.
scabra lebih tahan terhadap kekeringan dan stres kekeringan ganda dibandingkan
dengan S. guianensis.
Parameter kadar air tanah, pertambahan tinggi vertikal, pertambahan jumah
daun, dan Relaive Water content dilakukan pengkajian pada pengamatan di hari ke-
16, kerena kedua tanaman masih lengkap atau belum dipanen, namun sudah
merespon stres kekeringan.
Tabel 1. Pengaruh Cekaman Kekeringan dan Pemberian H2O2 terhadap Kadar Air
(KA) Tanah, Pertumbuhan, dan Produksi Leguminosa Stylosanthes
guianensis Hari ke-16
Perlakuan M0 M1 M2
KA Tanah (%) 35,369 ± 0,561a 28,467 ± 2,840
b 29,316 ± 2,129
b
Rataan Pertambahan Tinggi
(cm/ hari) 0,702 ± 0,089a 0,119 ± 0,092
b 0,088 ± 0,051
b
Rataan Pertambahan Jumlah
daun trifoliate (unit/hari) 5,188 ± 1,452a 1,031 ± 1,192
b 1,063 ± 0,650
b
Rataan RWC Daun (%) 60,108 ± 1,614a 29,988 ± 6,723
b 22,077 ± 1,127
c
Keterangan : Huruf kecil yang berbeda pada baris yang sama menunjukkan pengaruh yang berbeda
nyata (P<0,05) (M0: tanpa H2O2 dan disiram; M1: tanpa H2O2 dan tidak disiram; M2:
dengan H2O2 dan tidak disiram)
Tabel 2. Pengaruh Cekaman Kekeringan dan Pemberian H2O2 terhadap KA Tanah,
Pertumbuhan, dan Produksi Leguminosa Stylosanthes scabra Hari ke-16
Perlakuan M0 M1 M2
KA Tanah (%) 35,065 ± 1,191a 29,608 ± 0,960
b 30,292 ± 0,558
b
Rataan Pertambahan Tinggi
(cm/ hari) 0,948 ± 0,306a 0,383 ± 0,119
b 0,379 ± 0,144
b
Rataan Pertambahan Jumlah
daun trifoliate (tunit/hari) 11,000 ± 4,150a 0,500 ± 0,25
c 1,219, ± 0,329
b
Rataan RWC Daun (%) 50,705 ± 2,588a 14,048 ± 1,325
b 13,896 ± 0,715
b
Keterangan : Huruf kecil yang berbeda pada baris yang sama menunjukkan pengaruh yang berbeda
nyata (P<0,05) (M0: tanpa H2O2 dan disiram; M1: tanpa H2O2 dan tidak disiram; M2:
dengan H2O2 dan tidak disiram)
Pengaruh Perlakuan terhadap Rataan Kadar Air Tanah
Ketersediaan air tanah merupakan suatu faktor dalam kemampuan bertahan
hidup dan distribusi spesies tanaman (Lakitan, 2002). Menurut Noggle dan Frizt
bahwa salah satu fungsi air merupakan senyawa pelarut bagi masuknya mineral-
mineral dari larutan tanah ke tanaman sebagai pelarut mineral nutrisi yang akan

19
diangkut dari satu bagian sel ke bagian sel lain. Tabel 1 dan 2 menunjukkan bahwa
tanaman S. guianensis maupun S. scabra setelah diberikan perlakuan cekaman
kekeringan atau tidak disiram (M1 dan M2) menunjukkan bahwa KA tanah pada
kedua tanaman berpengaruh nyata (P<0,05) terhadap perlakuan yang disiram setiap
hari (M0). Persentase penurunan kadar air yang disiram setiap hari dengan tanaman
yang mendapatkan stres kering dan stres kekeringan ganda pada kedua tanaman
sebesar ±19%. Kadar air dalam tanah mendeskripsikan jumah sumber air yang
tersedia, dimana air dapat diserap untuk pertumbuhan, sedangkan kekeringan dapat
menyebabkan air tidak tersedia, dan tanaman menjadi menderita dan layu (Karti,
2004). Tanaman yang diberikan air atau perlakuan M0 terlihat bahwa
pertumbuhannya sangat baik dan berbeda nyata (P<0,05) dengan kedua perlakuan
yang lain, sedangkan untuk perlakuan M1 dan M2 tidak memiliki perbedaan yang
nyata hal ini terjadi karena memang pada perlakuan keduanya tidak disiram setiap
hari. Menurut Karti et al (2012) bahwa tanaman yang diberikan stres kekeringan
dapat menurunkan kadar air tanah dibandingkan dengan tanah yang disiram setiap
hari.
Pengaruh Perlakuan terhadap Laju Pertumbuhan Tinggi Vertikal Tanaman
Terjadinya pertambahan tinggi menurut Hermawan (2004) merupakan hasil
dari pembelahan sel dan pembesaran jaringan sel tanaman. Leguminosa S. guianensis
dan S. scabra setelah diberikan perlakuan memiliki berbagai macam perubahan
seperti yang tercantum pada Tabel 1 dan 2 Perlakuan yang diberikan pada tanaman
leguminosa S. scabra dan S. guianensis menghasilkan perbedaan yang nyata
(P<0,05) untuk laju pertumbuhan tinggi vertikal tanaman antara perlakuan tanpa stres
atau M0 dengan perlakuan stres (M1 dan M2), sedangkan untuk M1 dan M2 tidak
memiliki perbedaan yang nyata. Hal ini menunjukkan bahwa kekeringan
memberikan pengaruh terhadap laju pertumbuhan tinggi vertikal namun tidak
berpengaruh pada tanaman yang disemprotkan dengan H2O2 atau mengalami stres
kekeringan ganda. Hal ini diduga karena terlalu berbedanya ketersediaan air yang
ada pada tanaman antara tanaman perlakuan M0 dengan tanaman perlakuan M1 dan
M2 dimana air sangat berguna untuk proses pertumbuhan tanaman. Pemberian stres
kekeringan ganda terhadap kedua tanaman tidak menyebabkan tanaman lebih buruk

20
pertambahan tingginya dibandingkan dengan tanaman yang hanya diberikan stres
kekeringan tunggal.
Pengaruh Perlakuan terhadap Rataan Selisih Jumlah Daun Trifoliate Tanaman
Rataan selisih jumlah daun trifoliate seperti yang ditunjukkan pada Tabel 1
dan 2 bahwa pada tanaman S. guianensis tidak memiliki perbedaan yang nyata antara
M1 dan M2. Lain halnya dengan tanaman S. scabra dimana pada tanaman ini semua
perlakuan memiliki perbedaan yang nyata, dimana pada perlakuan M0 memiliki
perbedaan yang sangat mencolok terhadap kedua perlakuan yang lain dimana nilai
rataan pertumbuhan jumlah daun trifoliate M0 sebesar 11,000 ± 4,150 sedangkan
perlakuan M1 sebesar 0,500 ± 0,2500 dan M2 sebesar 1,219 ± 0,329 hal ini terjadi
karena perlakuan M0 tidak diberikan stres kekeringan sedangkan M1 dan M2
diberikan stres kekeringan.
Perlakuan yang hanya mengalami stres kekeringan tunggal atau M1 pada
tanaman S. scabra ternyata memiliki nilai rataan pertumbuhan jumlah daun trifoliate
yang lebih kecil dibandingkan dengan perlakuan yang diberikan stres kekeringan
ganda (M2), hal ini menunjukkan bahwa tanaman S. scabra tahan akan kekeringan
bahkan setelah diberikan double stress. Berbeda dengan S. guianensis yang setelah
diberikan dengan H2O2 bahkan terjadinya penurunan nilai dimana nilai M1 lebih
besar dibandingkan dengan M2, hal ini terjadi karena perlakuan M2 adalah perlakuan
yang mengalami stres kekeringan ganda dan tanaman S. guianensis tidak tahan akan
kekeringan seperti yang sudah dijelaskan diatas dimana tanaman S. guianensis
memiliki hari titik pelayuan yang lebih cepat, jadi nilai rataan M2 lebih kecil
dibandingkan dengan perlakauan M1.
Pengaruh Perlakuan terhadap Rataan RWC Daun Tanaman
Pengaruh perlakuan pada S. guianensis yang dapat dilihat pada Tabel 1 dan 2,
bahwa terjadi perbedaan yang nyata pada semua perlakuan dimana antara perlakuan
M0, M1, dan M2 memiliki perbedaan yang nyata (P<0,05) dengan nilai rataan M2
lebih kecil dari M1. Menurut Jiang dan Huang (1999) bahwa stres ganda antara stres
kekeringan dan stres panas dapat menurunkan nilai RWC, hal inilah yang
menyebabkan S. guianensis mengalami penurunan pada perlakuan M2 dimana
perlakuan M2 adalah stres kekeringan ganda yaitu stres kekeringan karena tidak
disiram dan stres kekeringan yang dihasilkan oleh pemberian H2O2.

21
Berbeda halnya dengan S. guianensis bahwa pengaruh perlakuan yang
dialami oleh S. scabra memiliki perbedaan yang nyata (P<0,05) antara M0 dengan
perlakuan yang lain, namun tidak berbeda nyata antara perlakuan M1 dengan M2.
Perlakuan M2 dikatakan ganda karena selain mendapat cekaman kekeringan tanpa
disiram juga ditambah dengan penyemprotan H2O2 dimana H2O2 dapat menyebabkan
stres kekeringan juga. Hal tersebut terjadi karena tanaman S. scabra merupakan
tanaman yang tahan akan kekeringan sehingga masih dapat bertahan bila diberikan
stres kekeringan ganda. Menurut Ishibashi et al. (2011) bahwa H2O2 dapat
meningkatkan level mRNA dari D-myo-inositol, 3-phosphate synthase 2 (GmMIPS2)
dan galacticol synthase (GolS) yang mengkodekan kunci enzim untuk biosintesis
oligosakarida yang dikenal dapat membantu tanaman untuk tetap bertahan pada
kondisi cekaman kekeringan. Menurut Karti et al. (2012) bahwa mekanisme
toleransi kekeringan pada S. seabrana melalui akumulasi prolin dan gula terlarut,
mungkin hal itu pula yang dilakukan oleh S. scabra agar dapat tetap bertahan pada
kondisi kekeringan. Menurut Castillo (1996) bahwa ketika RWC turun mencapai
20% dibawah cekaman kekeringan maupun stres ganda maka dapat mencegah
aktivitas SOD (Superoxide dismutase) secara signifikan. Tidak berubahnya aktivitas
SOD dibawah stres kering dapat menyesuaikan diri dari kekeringan dan dapat
berkontribusi dalam mentolerir panas.
Pengaruh Perlakuan terhadap Rataan Bobot Kering Daun, Batang, dan Akar
Tabel 1 dan 2 menunjukkan bahwa pada perlakuan yang disiram setiap hari
atau M0 memiliki nilai yang lebih tinggi dibandingkan dengan perlakuan yang
lainnya baik pada S. guianensis maupun S. scabra, hal ini terjadi karena kekeringan
dapat menyebabkan daun menjadi lebih sempit dan pendek (Teare dan Peet, 1983).
Rataan bobot kering daun pada S. guianensis terjadi perbedaan yang nyata (P<0,05)
antara M0 dengan M1 dan M2 namun tidak terjadi perbedaan yang nyata antara
perlakuan M1 dan M2, sedangkan untuk S. scabra terjadi perbedaan yang nyata pada
semua perlakuan. Perlakuan M2 pada S. scabra memiliki nilai yang lebih tinggi
dibandingkan dengan M1 hal ini dapat dikatakan bahwa S. scabra tahan akan stres
kekeringan ganda bahkan lebih baik daripada hanya diberikan stres kekeringan
tunggal, hal ini menunjukkan bahwa H2O2 memiliki pengaruh yang nyata pada
leguminosa S. scabra untuk mempertahankan produksi bobot kering daun yang lebih

22
baik melalui suatu mekanisme tertentu terhadap kekeringan. Pengaruh meningkatnya
nilai bobot kering daun pada leguminosa S. scabra adalah bahwa H2O2 dapat
meningkatkan level mRNA dari D-myo-inositol, 3-phosphate synthase 2 (GmMIPS2)
dan galacticol synthase (GolS) yang mengkodekan kunci enzim untuk biosintesis
oligosakarida yang dikenal dapat membantu tanaman untuk mentolerir cekaman
kekeringan (Ishibashi et al., 2011).
Tabel 3. Pengaruh Cekaman Kekeringan dan Pemberian H2O2 terhadap
Pertumbuhan dan Produksi Leguminosa Stylosanthes guianensis
Perlakuan M0 M1 M2
Rataan Produksi Bobot Kering
Daun (gram) 2,531 ± 0,239a 0,641 ± 0,155
b 0,633 ± 0,207
b
Rataan Bobot Kering Batang
(gram) 4,979 ± 0,643a 3,456 ± 0,885
b 3,125 ± 0,478
b
Rataan Bobot Kering Akar
(gram) 2,006 ± 0,822 1,535 ± 0,849 1,243 ± 0,367
Keterangan : Huruf kecil yang berbeda pada baris yang sama menunjukkan pengaruh yang berbeda
nyata (P<0,05) (M0: tanpa H2O2 dan disiram; M1: tanpa H2O2 dan tidak disiram; M2:
dengan H2O2 dan tidak disiram)
Tabel 4. Pengaruh Cekaman Kekeringan dan Pemberian H2O2 terhadap
Pertumbuhan dan Produksi Leguminosa Stylosanthes scabra
Perlakuan M0 M1 M2
Rataan Produksi Bobot Kering
Daun (gram) 2,803 ± 0,589a 0,661 ± 0,126
c 0,971 ± 0,175
b
Rataan Bobot Kering Batang
(gram) 6,377 ± 1,612a 3,066 ± 0,486
b 3,011 ± 0,418
b
Rataan Bobot Kering Akar
(gram) 0,385 ± 0,192 0,334 ± 0,116 0,323 ± 0,142
Keterangan : Huruf kecil yang berbeda pada baris yang sama menunjukkan pengaruh yang
berbeda nyata (P<0,05) (M0: tanpa H2O2 dan disiram; M1: tanpa H2O2 dan tidak
disiram; M2: dengan H2O2 dan tidak disiram)
Bukan hanya daun saja yang mengalami penurunan ketika mengalami
kekeringan produksi batang dan organ reproduktifpun juga terbentuk lebih kecil dari
ukuran normal (Teare dan Peet, 1983). Rataan bobot kering batang untuk kedua
leguminosa pada perlakuan M0 memiliki perbedaan yang nyata dengan perlakuan
yang lain. S. guianensis dan S. scabra pada perlakuan M1 dan M2 tidak memiliki
perbedaan yang nyata.

23
Rataan bobot kering akar baik pada S. guianensis maupun S. scabra tidak
mengalami perbedaan yang nyata pada semua perlakuan. Teare dan Peet (1983)
menyatakan bahwa terdapat mekanisme cara menghindari ketika tanaman mengalami
kekeringan yaitu dengan memperluas sistem perakaran dan pertumbuhan memanjang
ke dalam akar, hal tersebutlah yang membuat bobot kering pada kedua tanaman yang
mengalami perlakuan stres kekeringan tidak memiliki perbedaan yang nyata dengan
perlakuan yang selalu disiram setiap hari. S. scabra juga memiliki perakaran yang
dalam sepanjang 4 m sehingga tanaman ini tahan akan kekeringan.
Penjelasan tersebut dapat diketahui bahwa tanaman S. guianensis merupakan
tanaman yang tidak tahan terhadap stres kekeringan dengan penambahan H2O2 atau
stres kekeringan ganda. Hal tersebut terjadi karena tanaman S. guianensis memang
merupakan tanaman yang kurang tahan akan kekeringan, sehingga setelah diberikan
H2O2 yang merupakan salah satu dari stres kekeringan tidak mampu menampilkan
hasil yang baik. Lain halnya dengan tanaman S. scabra yang tahan akan stres
kekeringan.
Pembahasan Umum
Pengaruh morfologi akibat cekaman kekeringan umumnya sangat tergantung
pada faktor waktu terjadinya cekaman dan besarnya perlakuan cekaman (Keles dan
Oncel, 2002). Kadar air dalam tanah mendeskripsikan jumlah sumber air yang
tersedia, dimana air dapat diserap untuk pertumbuhan, sedangkan kekeringan dapat
menyebabkan air tidak tersedia, dan tanaman menjadi menderita dan layu (Karti,
2004). Karti et al. (2012) menyatakan bahwa tanaman yang diberikan stres
kekeringan dapat menurunkan kadar air tanah dibandingkan dengan tanah yang
disiram setiap hari. Penelitian ini memberikan stres kekeringan terhadap S.
guianensis dan S. scabra, kedua tanaman tersebut memiliki respon terhadap cekaman
kekeringan dengan cara mengugurkan daunnya, menghambat pertumbuhannya
hingga mengalami kelayuan yang permanen. Waktu titik layu permanen pada kedua
tanaman ini berbeda untuk S. guianensis pada hari ke-16 setelah tanaman tersebut
diberikan perlakuan sedangkan untuk S. scabra pada hari ke-24, hal ini dapat
diketahui bahwa S. scabra lebih tahan akan kerkeringan dibandingkan dengan S.
guianenesis.

24
Cekaman kekeringan, panas maupun keduanya dapat menurunkan kualitas
tanah, RWC, dan kandungan klorofil daun. Stres ganda efeknya lebih dapat merusak
dibandingkan dengan hanya stres kering saja maupun stres panas saja. Kualitas tanah
menurun secara drastis dibawah cekaman kekeringan, sedangkan cekaman ganda
dapat menurunkan nilai RWC selama perlakuan stres (Turner et al., 1966; Nobel,
1988). Stylosanthes guianensis tidak tahan akan kekeringan ganda karena setelah
dapat dilihat pada hasil penelitian dan penjabaran diatas. Stylosanthes guianensis
selain dilihat pada hari titik pelayuannya dapat dilihat pula dari nilai rataan RWC,
karena RWC merupakan salah satu cara untuk dapat menseleksi langsung tanaman
terhadap resisten kekeringan (Yuniaty, 1998; Reynolds et al., 1999; Chandrasekar et
al., 2000). Leguminosa S. guianensis untuk rataan jumlah daun trifoliate tidak
memiliki perbedaan yang nyata perlakuan M1 dan M2, namun untuk nilai RWC
berbeda nyata hal inilah yang menandakan bahwa S. guianensis tidak tahan akan
kekeringan ganda dan tidak efisien dalam penggunaan airnya. Leguminosa S.
guianensis juga tidak mampu menghasilkan akumulasi prolin dan gula terlarut
seperti yang dapat dilakukan oleh S. seabrana (Karti et al., 2012).
Berbeda halnya dengan S. guianensis, S. scabra dapat tahan terhadap stres
kekeringan ganda hal ini dapat dilihat dari nilai RWC dimana antara perlakuan M1
dan M2 tidak memiliki perbedaan yang nyata, meskipun pada pertambahan jumlah
daun M1 dan M2 memiliki perbedaan yang nyata (P<0,05) hal ini dapat dikatakan
bahwa S. scabra dapat lebih efisien dalam penggunaan airnya. Hal tersebut juga
dapat terjadi karena menurut Karti et al. (2012) bahwa mekanisme toleransi
kekeringan pada S. seabrana melalui akumulasi prolin dan gula terlarut, mungkin hal
itu pula yang dilakukan oleh S. scabra agar dapat tetap bertahan pada kondisi
kekeringan. Akumulasi prolin dan gula prolin merupakan mekanisme toleransi
kekeringan, hal ini terjadi karena pada saat tanaman mengalami kekeringan kadar air
tanah pada tanaman mengalami penurunan atau potensial airnya menurun.
Menurunnya potensial air pada kadar air tanah maka akar tanaman tidak dapat
mengalirkan persediaan air dari tanah keseluruh bagian tanaman karena salah satu
sifat dari air adalah mengalir dari potensial air tinggi ke potensial air yang lebih
rendah, untuk dapat mengalirkan air keseluruh bagian tanaman maka tanaman yang
lebih tahan akan kekeringan akan menghasilkan akumulasi prolin dan gula terlarut

25
sehingga dapat membuat konsentrasi tanaman tinggi atau menurunkan potensial air
pada tanaman, sehingga potensial air tanaman menjadi lebih rendah dari tanah
sehingga tanaman dapat menyerap air untuk hidupnya.
Menurut Ishibashi et al. (2011) pula bahwa H2O2 dapat meningkatkan level
mRNA dari D-myo-inositol, 3-phosphate synthase 2 (GmMIPS2) dan galacticol
synthase (GolS) yang mengkodekan kunci enzim untuk biosintesis oligosakarida
yang dikenal dapat membantu tanaman untuk tetap bertahan pada kondisi cekaman
kekeringan apabila tanaman tersebut toleran, hal ini pula yang dilakukan oleh S.
scabra untuk mentolerir kekeringan. Rataan produksi bobot kering daun S. scabra
memiliki perbedaan yang nyata (P<0,05) antara M1 dengan M2 hasil tersebut sejalan
dengan rataan pertambahan jumlah daun trifoliate pada S. scabra yang juga berbeda
nyata (P<0,05) dengan keduanya. Begitu pula dengan S. guianensis yang tidak
memiliki perbedaan yang nyata antara M1 dengan M2 pada rataan pertambahan
jumlah daun trifoliate dan rataan produksi bobot kering daun.

26
KESIMPULAN DAN SARAN
Kesimpulan
Stylosanthes scabra merupakan tanaman yang dapat tahan terhadap stres
kekeringan dan stres kekeringan karena H2O2 sedangkan S. guianensis hanya tolerir
tehadap stres kekeringgan saja.
Saran
Diperlukan pengujian terhadap leguminosa lokal Indonesia yang tahan akan
kekeringan saja ataupun stres kekeringan ganda.

27
UCAPAN TERIMA KASIH
Puji Syukur Penulis panjatkan kehadirat Allah SWT. yang telah
memberikan banyak kenikmatan dan karunia terutama kesehatan sehingga Penulis
dapat menyelesaikan penulisan skripsi ini dan shalawat serta salam Penulis curahkan
kepada Nabi Muhammad SAW. Penulis menyampaikan terima kasih kepada Dr. Ir.
Panca Dewi, M.H.K., M.S. selaku pembimbing utama yang telah memberikan
masukan-masukan yang sangat bermanfaat. Penulis juga mengucapkan banyak
terima kasih kepada Ir. Widya Hermana M.Si. selaku pembimbing anggota sekaligus
pembimbing akademik yang telah membantu Penulis melewati masa studi serta
memberikan saran-saran yang membangun. Penuis mengucakan terima kasih juga
kepada Iwan Prihantoro. S.Pt. selaku dosen penguji seminar yang teah memberikan
masukan untuk penulisan skripsi ini. Terima kasih pula Penulis ucapkan kepada Dr.
Ir. Salundik. M.S., Ir. Asep Tata Permana, MSc., dan Dr. Ir. Rita Mutia, M.Agr.
selaku dosen penguji dan panitia sidang yang telah memberikan saran dalam
penyelesaian skripsi ini. Terima kasih juga Penulis sampaikan kepada staf
Laboratorium Lapang Agrostologi dan Laboratorium Ilmu Nutrisi Ternak Perah dan
seluruh dosen yang telah mengamalkan ilmunya.
Penulis tak lupa menyampaikan terima kasih kepada Ayahanda Sudjono
dan Ibunda Suryati selaku orang tua serta kakanda Ako, Dwi, dan Nia serta adinda
Dian yang telah memberikan motivasi baik materi maupun non materi hingga Penulis
dapat menyelesaikan studi. Terima kasih juga kepada Beasiswa BUMN. Kepada Nur
Herati Akhadia selaku teman sepenelitian Penulis yang selalu bersama dan
membantu Penulis dalam melaksanakan penelitian ini, serta keluarga besar Penulis
yang telah memberikan motivasi untuk menyelesaikan penulisan skripsi ini.
Penulis sampaikan terima kasih kepada teman-teman Kecebonk,
GENETIC45 dan seluruh mahasiswa INTP yang telah menemani Penulis selama
studi ini terutama kepada Silvi, Kiki, Hera, Didi, Maha, Vivi, Nurus, Fitri, dan Rima
juga kepada teman-teman wisma nabila yang telah memberikan motivasi dan
menemani Penulis dalam penulisan skripsi.
Bogor, Agustus 2012
Penulis

28
DAFTAR PUSTAKAs
Abele, D, B. Burlando, A. Viarengo, & H. O. Portner. 1998. Exposure to elevated
temperatures and hydrogen peroxide elicits oxidative stress and antioxidant
renponse in the Antarctic intertidal limpet Nucella concinna. Comp Biochem
Physiol Part B 120:425-435.
Alscher, R. G, J. L. Donahue, & C. Cramer. 1997. Reactive oxygen species and
antioxidants: relationships in green cells. Physiol Plant. 100 : 224-233.
Andayani, W & A. Sumartono. 1999. Aplikasi radiasi pengion dalam penguraian
limbah industri I. Radiolisis larutan standar zat warna reaktif cibacron violet
2r. Pusat Aplikasi Isotop dan Radiasi-Batan. Majalah Batan Vol. XXXII No
1/2.
Blum, A. 1999. Toward standard assays of drought resistance in crop plants. P. 29-
35. In J. M. Ribaut and D. Poland. Molecuar approaches for the genetic
improvement of cereals for stable production in water-limited enviroment. A
strategic planning workshop held at CCIMMYT, 21-25 June 1999.
Bogdan, A.V. 1977. Tropical Pasture and Fodder Plants. Longman Inc., New York.
pp. 397-402.
Bowler, C, M.V. Montagu, & D. Inzé. 1992. Superoxide dismutase and stress
tolerance. Annu Rev Plant Physiol Plant Mol Biol, 43 : 83-116.
Carrow, R.N. 1996. Drought avoidance characteristics of diverse tall fescue cultivars.
Crop Sci. 36:371–377.
Castillo, F. J. 1996. Antioxidative protection in the inducible CAM plant Sedum
album L. Following imposition of severe water stress and recovery.
Oecologia 107: 469-477.
Chakraborty S. 2004 High-yielding anthracnose-resistant Stylosanthes for
agricultural systems. ACIAR Monograph No 111. (Australian Centre for
International Agricultural Research, Canberra).
Chandrasekar, V., R. K. Sairam, & G. C. Srivastava. 2000. Physiological and
biochemical respones of hexaploid and tetraploid wheat to drought stress. J.
Agron. Crop Sci. 185: 219-227.
Chaves, M. M, J. S. Pereira, J. Maroco, M. L. Rodrigues, C. P. Ricardo, M. L.
Osorio, I. Carvalho, T. Faria, C. Pinheiro. 2002. How plants cope with water
stress in the field. Photosynthesis and growth. Annals. Bot. 89: 907–916.
Chowdhury, S. R., & M. A. Choudhuri. 1985. Hydrogen peroxide metabolism as an
index of water stress tolerance in jute. Physiol. Plant. 65:503–507.
Dat, J.F., H. Lopez-Delgado, C.H. Foyer, & I.M., Scott. 1998. Parallel changes in
H2O2 and catalase during thermotolerance induced by salicylic acid or heat
acclimation in mustard seedlings. Plant Physiol. 116:1351–1357.
Edye, L.A. & A. Topark-Ngarm. (1992) Stylosanthes scabra Vogel. In: 't Mannetje,
L. and Jones, R.M. (eds) Plant Resources of South-East Asia No. 4. Forages.
pp. 219–221. (Pudoc Scientific Publishers, Wageningen, the Netherlands).

29
FAO. 2012. Stylosanthes guianensis (Aub) Sw.var. intermedia (Vog.) Hassler.
http://www.fao.org/ag/AGP/AGPC/doc/GBASE/data/pf000071.htm. [25
Januari 2012]
Foyer, C. H, M. Lelandais, & K. J. Kunert. 1994. Photooxidative stress in plants.
Physiol Plant. 92 : 696-717.
Gardber, F. P, R. B. Pearce, & R. L. Mitchell. 1991. Fisiologi Tanaman Budidaya. UI
Press. Jakarta.
Gonzalez & Gonzalez-Vilar. 2001. Determination of relative water content. In
Handbook of plant ecophysiology techniques. M. I. Reigosa Roger, Kluwer
Pubishers. pp. 207-212.
Harjadi, S. S. & S. Yahya. 1988. Fisiologi Stres Lingkungan. PAU Bioteknologi.
Institut Pertanian Bogor, Bogor.
Hermawan, B. 2004. Penetapan kadar air tanah melalui pengukuran sifat dielektrik
pada berbagai tingkat kepadatan. Jurnal Ilmu-ilmu Pertanian Indonesia 6 (2) :
66-74.
Heyne, K. 1987. Tumbuhan Berguna Indonesia. Jilid I (Terjemahan). Badan
Penelitian dan Pengembangan Kehutanan. Jakarta.
Howard, H., & T.L. Watschke. 1991. Variable high-temperature tolerance among
Kentucky bluegrass cultivars. Agron. J. 83:689–693.
Huang, B., & H. Gao. 1999. Physiological responses of diverse tall fescue cultivars
to drought stress. Hort Science 34:897–901.
Huang, B., X. Liu, and J.D. Fry. 1998. Shoot physiological responses of two
bentgrass cultivars to high temperature and poor soil aeration. Crop Sci.
38:1219–1214.
Inze, D, M. Van Montagu. 1995. Oxidative stress in plants. Current Opinion in
Biotechnology 6: 153-158.
Ishibashi, Y, H. Yamaguchi, T. Yuasa, M. Iwaya-Inoue, S. Arima, & SH. Zheng.
2011. Hydrogen peroxide spraying alleviates drought stress in soybean plants.
Journal of Plants Physiology 168 : 1562-1567
Islami, T. & W. H. Utomo. 1995. Hubungan Tanah, Air dan Tanaman. IKIP
Semarang Press, Semarang.
Jagtap, V. & S. Bhargava. 1995. Variation in antioxidant metabolism of drought
tolerant and drought susceptible varieties of Sorghum bicolor (L.) Moench.
Exsposed to high light, high temperature stress. J. Plant Physiol. 145:195-
197.
Jaleel, C. A., P. Manivannan, Wahid, A. Farooq, M. Al-Juburi, H. J. Somasundaram,
R. Pannerselvam. 2009. Drought stress in plant: A review on morphological
characteristics and pigments composition. International Journal of Agriculture
& Biology.
Jiang, Y & B. Huang, 1999. Drought and heat stress injury to two cool-season
turfgrasses in relation to antioxidant metabolism and lipid peroxidation. No.
00-227-J . Kansas Agric. Exp. Stn.

30
Karti, P. D. M. H, D. A. Astuti, & S. Nofyangtri. 2012. The rolle of arbuscular
mycorrhizal fungi in enhancing productivity, nutritional quality, and drought
tolerance mechanism of Stylosanthes seabrana. Media Peternakan. 35(1): 67-
72.
Karti, P. D. M. H. 2004. Effect of arbuscular mycorhizal fungi on growth and
production of Setaria splendida stapf in drought stress. Media Peternakan.
27(2): 63-68.
Keles, Y & I. Oncel. 2002. Response of antioxidative defence system to temperature
and water stress combinations in wheat seedlings. Plant Sci. 163: 783790.
Lakitan B. 2002. Dasar-dasar Klimatologi. Raja Grafindo Persada. Jakarta.
Lilley, J. M. & M. M. Ludlow. 1996. Expression of osmotic adjusment and
dehydration tolerance in diverse rice lines. Fields Crop Res. 48: 185-197.
Mannetje, L & R. M. Jones. 1992. Plant Resources of South-East Asia No 4.
Forages. Pudoc-DLO, Wangeningen, the Netherlands. Pp. 211-213.
McIlroy, R.J. 1976. Pengantar Budidaya Padang Rumput Tropika. Terjemahan: S.
Susetyo, Soedarmadi, I. Kismono dan S. Harini, I. Penerbit Pradnya Paramita,
Jakarta.
Monk, L.S., K.V. Fagerstedt, & R.M.M. Crawford. 1989. Oxygen toxicity and
superoxide dismutase as an antioxidant in physiological stress. Physiol. Plant.
76:456–459.
Nobel, P.S. 1988. Principles underlying the prediction of temperature in plants, with
special reference to desert succulents. In Symposia of the Society for
Experimental Biology. Number XLII. Eds. S.P. Long and F.I. Woodward.
Noggle, G. R, G. J. Fritz. 1983. Introductory plant physiology. Prentice-Hall, Inc.
Englewood Cliffs. New Jersey. 627p.
Patnaik, D & Khurana. 2002. Wheat biotechnology a miniteview, EJB Electrinic J.
Biotechnol. 4: 74-102.
Perdomo, P, J.A. Murphy, & G.A. Berkowitz. 1996. Physiological changes
associated with performance of Kentucky bluegrass cultivars during summer
stress. HortScience 31:1182–1186.
PROSEA. 1992. Plant Resources of South-East. Asia. Forages 4th
Edition. Bogor.
Reksohadiprodjo, S. 1985. Produksi Tanaman Hijauan Makanan Ternak Tropik.
BPFE, Yogyakarta.
Reynolds, M., B. Skovmand, R. Trethowan, & W. Pfeiffer. 1999. Evaluating a
conceptual model for drought tolerance. P. 49-53. In J. M. Ribaut & D.
Poland. Molecular approaches for the genetic improvement of cereals for
stable production in water-limited environment. A strategic planning
workshop held at CIMMYT, 21-25 June, 1999.
Salisbury, F. B. & C. W. Ross. 1995. Fisiologi Tumbuhan Jilid 3. Terjemahan: Diah
R Lukman dan Sumaryono. Institut Teknologi Bandung, Bandung.

31
Sharp, R. E., T. C. HSIO, W. K. SILK. 1990. Growth of the maize primary root at
low water potentials. II. Role of growth and deposition of hexose and
potassium in osmotic adlustment. Plant Physiol 93: 1337-1348.
Shigeoka, S, T. Ishikawa, M. Tamoi, Y. Miyagawa, T. Takeda, Y. Yabuta & K.
Yoshimura. 2002. Regulation and function of ascorbate peroxidase
isoenzymes. J Exp Bot. 54 : 1305-1319.
Slatyer, R. O. 1967. Plant-water relationships, Chapters 6 and 9. Academic Press,
New York.
Steel R.G.D. & J.H. Torrie. 1995. Prinsip dan Prosedur Statistika. Diterjemahkan
oleh Bambang Sumantri. PT. Gramedia Pustaka Utama. Jakarta
Taiz, L & E. Zeiger. 2002. Plant Physiology. 3rd
Edition. Sinauer Associates,
Sunderland, MA.
Teare, I. D. & M. M. Peet. 1983. Crop Water Relations. John Wiley & Sons. Canada.
211p.
Turner, R.M., S.M. Alcorn, S. Olin, & S.A. Booth. 1966. The influence of shade, soil
and water on saguaro seedlings establishment. Bot. Gaz. 127:95–102.
Yuniaty, A. 1998. Screening for drought resistance with the use of some
morphophysiological characters in soybean. M. S. Thesis, University of the
Philippines at Los Banos, Manila.
Zhang, J & M.B. Kirkham. 1994. Drought-stress-induced changes in activities of
superoxide dismutase, catalase, and peroxidase in wheat species. Plant Cell
Physiol. 35:785–791.

32
LAMPIRAN

33
Lampiran 1. Perhitungan H2O2 :
Botol bertanda:
1L : 1,11 kg H2O2 30%
M (mol/liter) = (30/100) x (1,11 x 1000)
34
= 9,794 M
= 9794 mmol/liter
H2O2 1mM 2,4 L = 2400 ml
Vol H2O2 = 2400/9794 = 0,24 ml
Dipipet 0,24 ml dari bobot H2O2 30% ditambahkan sampai 2,4 L
Lampiran 2. Sidik Ragam Rataan Kadar Air Tanah Stylosanthes guianensis sampai
Hari ke-16
SK JK db KT F Sig.
Perlakuan 70,693 2 35,347 39,968 0,000
Galat 7,959 9 0,884
Total 78,653 11
Lampiran 3. Sidik Ragam Rataan Kadar Air Tanah Stylosanthes scabra sampai Hari
ke-16
SK JK Db KT F Sig.
Perlakuan 109,909 2 54,954 12,659 0,002
Galat 39,071 9 4,341
Total 148,979 11
34
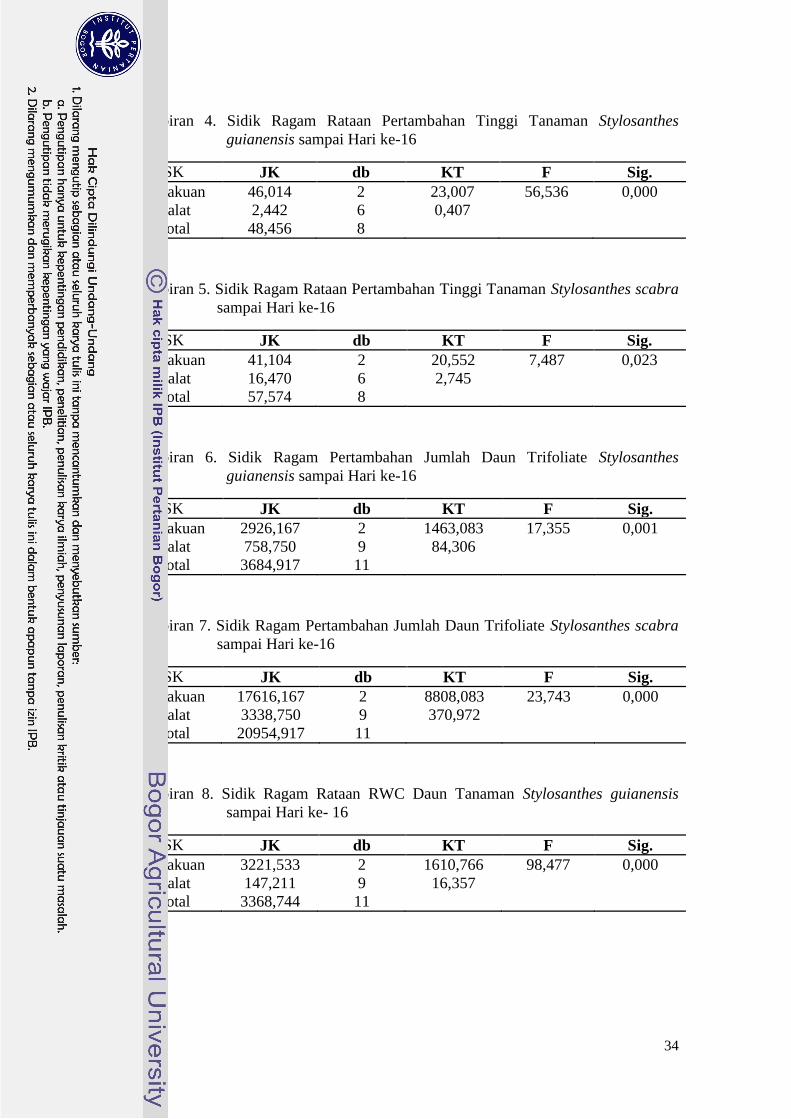
Lampiran 4. Sidik Ragam Rataan Pertambahan Tinggi Tanaman Stylosanthes
guianensis sampai Hari ke-16
SK JK db KT F Sig.
Perlakuan 46,014 2 23,007 56,536 0,000
Galat 2,442 6 0,407
Total 48,456 8
Lampiran 5. Sidik Ragam Rataan Pertambahan Tinggi Tanaman Stylosanthes scabra
sampai Hari ke-16
SK JK db KT F Sig.
Perlakuan 41,104 2 20,552 7,487 0,023
Galat 16,470 6 2,745
Total 57,574 8
Lampiran 6. Sidik Ragam Pertambahan Jumlah Daun Trifoliate Stylosanthes
guianensis sampai Hari ke-16
SK JK db KT F Sig.
Perlakuan 2926,167 2 1463,083 17,355 0,001
Galat 758,750 9 84,306
Total 3684,917 11
Lampiran 7. Sidik Ragam Pertambahan Jumlah Daun Trifoliate Stylosanthes scabra
sampai Hari ke-16
SK JK db KT F Sig.
Perlakuan 17616,167 2 8808,083 23,743 0,000
Galat 3338,750 9 370,972
Total 20954,917 11
Lampiran 8. Sidik Ragam Rataan RWC Daun Tanaman Stylosanthes guianensis
sampai Hari ke- 16
SK JK db KT F Sig.
Perlakuan 3221,533 2 1610,766 98,477 0,000
Galat 147,211 9 16,357
Total 3368,744 11

35
Lampiran 9. Sidik Ragam Rataan RWC Daun Tanaman Stylosanthes guianensis
sampai Hari ke- 16
SK JK db KT F Sig.
Perlakuan 3598,233 2 1799,117 602,206 0,000
Galat 26,888 9 2,988
Total 3625,121 11
Lampiran 10. Sidik Ragam Rataan Produksi Bobot Kering Daun Tanaman
Stylosanthes guianensis
SK JK db KT F Sig.
Perlakuan 9,561 2 4,781 115,471 0,000
Galat 0,373 9 0,041
Total 9,934 11
Lampiran 11. Sidik Ragam Rataan Produksi Bobot Kering Daun Tanaman
Stylosanthes scabra
SK JK db KT F Sig.
Perlakuan 10,720 2 5,360 40,922 0,000
Galat 1,179 9 0,131
Total 11,898 11
Lampiran 12. Sidik Ragam Rataan Produksi Bobot Kering Batang Tanaman
Stylosanthes guianensis
SK JK db KT F Sig.
Perlakuan 7,827 2 3,914 8,244 0,009
Galat 4,273 9 0,476
Total 12,100 11
Lampiran 13. Sidik Ragam Rataan Produksi Bobot Kering Batang Tanaman
Stylosanthes scabra
SK JK db KT F Sig.
Perlakuan 29,737 2 14,868 14,827 0,001
Galat 9,025 9 1,003
Total 38,761 11

36
Lampiran 14. Sidik Ragam Rataan Produksi Bobot Kering Akar Tanaman
Stylosanthes guianensis
SK JK db KT F Sig.
Perlakuan 1,187 2 0,593 1,162 0,356
Galat 4,594 9 0,510
Total 5,780 11
Lampiran 15. Sidik Ragam Rataan Produksi Bobot Kering Akar Tanaman
Stylosanthes scabra
SK JK db KT F Sig.
Perlakuan 0,037 2 0,08 0,892 0,443
Galat 0,185 9 0,021
Total 0,221 11
Top Related
Copyright © 2022 FDOKUMEN

