






Bahasa
Halaman
Hukum


1
BAB I
PENDAHULUAN
A. Latar Belakang Masalah
Indonesia dengan keanekaragaman hayatinya yang terdiri dari
sekitar 30.000 jenis tumbuhan tinggi (Achmad, 2007), 381 jenis mamalia
(Heny, 2010) dan 12.000 jenis jamur (Anonim, 2010) potensial untuk
memproduksi metabolit primer (misalnya karbohidrat, lemak dan protein)
dan metabolit sekunder (misalnya senyawa antibiotik). Mikroba banyak
dikembangkan untuk memproduksi metabolit primer dan sekunder, karena
mikroba berkembang biak dengan cepat, tidak terganggu oleh musim,
dan dapat diproduksi sewaktu-waktu. Selain itu produksi enzim dari
mikroba dapat ditingkatkan pada skala besar melalui fermentor dan dapat
dikontrol lingkungan tempat tumbuhnya (Suhartono, 1989).
Enzim lipase dari mikroba termasuk salah satu enzim komersial
yang mempunyai nilai ekonomi tinggi dan masih diimpor. Harga enzim
lipase impor relatif mahal, misalnya harga enzim lipase dengan merek
dagang BIO-lipase dan Lipolase mencapai 25 juta rupiah per kg (Kao
Corporation, 2004). Menurut Kirk dkk (2002), penggunaan enzim lipase
untuk industri bioteknologi senilai U$ 1,5 milyar. Enzim lipase dapat pula
digunakan dalam industri makanan, deterjen, kimia, farmasi, dan lain-
lain (Gupta dkk., 2004).

2
Enzim lipase merupakan kelompok enzim yang berfungsi untuk
menghidrolisis triasilgliserol menjadi monoasilgliserol, diasilgliserol dan
asam lemak bebas. Monoasilgliserol dan diasilgliserol termasuk produk
diversifikasi minyak yang bernilai ekonomi relatif tinggi dan mempunyai
prospek pasar yang cukup cerah pada era pasar global. Krog (1990)
memprediksi kebutuhan monoasilgliserol dan diasilgliserol sebagai
pengemulsi pangan pada era pasar global sekitar 132.000 ton/tahun.
Kebutuhan monoasilgliserol dan diasilgliserol dalam negeri saat ini masih
impor. Monoasilgliserol dan diasilgliserol adalah ester gliserol dari
triasilgliserol yang digunakan sebagai bahan pengemulsi dan penstabil
produk makanan, kosmetika dan farmasetika (Ling dkk., 2007).
Hasanuddin (2001) melaporkan bahwa pada dasarnya
diasilgliserol dan monoasilgliserol terbentuk dari reaksi antara gliserol
dengan triasilgliserol. Reaksi ini dapat berlangsung dengan katalisator
alkali (gliserolisis secara kimia) maupun biokatalisator enzim lipase
(gliserolisis secara enzimatik). Enzim lipase sebagai biokatalisator
mempunyai beberapa kelebihan dibandingkan katalisator alkali
diantaranya: bekerja secara spesifik, aktivitas katalitik enzim yang tinggi
dan kemampuannya bekerja pada suhu yang relatif rendah sekitar 30 0C,
katalisator alkali bekerja pada suhu (220-250) 0C. Suhu yang tinggi
menghasilkan produk berwarna coklat (gelap) dan bau tidak diinginkan
(Noureddini dkk., 2004). Enzim lipase sebagai biokatalisator dapat
meningkatkan kualitas crude palm oil (CPO) yang lebih baik yaitu minyak

3
sehat (healthy oil). Healthy Econa Cooking Oil telah diproduksi massal
pada tahun 2001 oleh Kao Industries of Japan bekerjasama dengan
Novozymes, Co. Bahan utama yang terkandung di dalam minyak ini
adalah diasilgliserol dibuat secara enzimatik menggunakan minyak murni
(virgin olive oil). Minyak ini dalam jangka panjang mampu mencegah
peningkatan lemak tubuh, terutama lemak yang terdeposit dalam organ
internal (Kao Corporation, 2004).
Fungi (jamur) adalah mikroba yang 80% substratnya terdiri dari
makromolekul yang memiliki banyak rantai karbon (Putranto dkk., 2006).
Beberapa jenis fungi penghasil enzim lipase seperti Aspergillus, Mucor,
Rhizopus, Penicillium diketahui dapat tumbuh pada habitat mengandung
minyak (Sharma dkk., 2001).
Buah kelapa adalah bahan baku industri pembuatan minyak
nabati, dalam pengolahannya terlebih dahulu dibuat kopra. Pembuatan
kopra dilakukan secara tradisional yaitu dengan cara pengupasan
tempurung kelapa kemudian penjemuran sampai kering. Survei lapangan
menunjukkan bahwa dalam pengolahan kelapa menjadi kopra, ditemukan
(1-5)% kopra berjamur, sehingga potensial menjadi limbah. Kopra memiliki
kandungan lemak sebesar 67,7% (Woodroof, 1979) merupakan media
yang baik untuk pertumbuhan fungi (jamur) penghasil enzim lipase. Studi
pendahuluan menunjukkan bahwa fungi yang tumbuh pada kopra
berjamur teridentifikasi dari genus Aspergillus dan Penicillium, Aspergillus
menunjukkan aktivitas enzim lipase lebih besar (Dali dan Pirman, 2005).

4
Studi mengenai penelitian enzim lipase dari beberapa mikroba
(jamur, ragi dan bakteri) telah dilaporkan oleh (Nascimento dan Takaki,
1994; Schmid dkk., 1997, 1998). Pada umumnya enzim lipase mikroba
aktif pada pH basa, pH (7-9). Produksi enzim lipase dari mikroba dapat
ditingkatkan aktivitasnya dengan membuat formulasi media yang sesuai
dengan kebutuhan mikroba untuk pertumbuhannya. Media yang
digunakan untuk pertumbuhan mikroba harus mengandung komponen-
komponen yang dibutuhkan seperti air, karbon, nitrogen, mineral dan lain-
lain sesuai kebutuhan setiap macam mikroba (Darwis dan Sukara, 1990).
Enzim lipase dari mikroba (misalnya: Mucor miehei, Candida rugosa,
Aspergillus oryzae) merupakan enzim ekstraseluler yang disekresikan
melalui membran eksternal ke dalam media kultur. Optimasi media
fermentasi untuk produksi enzim lipase bergantung pada beberapa faktor
seperti suhu pertumbuhan, pH, komposisi nitrogen, sumber karbon dan
lipid, konsentrasi garam anorganik serta ketersediaan oksigen (Suzuki
dkk., 1998)
Penggunaan enzim lipase terlarut sebagai biokatalisator sangat
tidak ekonomis, dibandingkan dengan enzim lipase yang tidak terlarut
(lipase imobil). Enzim dalam bentuk terlarut relatif tidak stabil dan tidak
dapat digunakan secara berulang-ulang (reuseable). Penggunaan enzim
hanya terbatas satu kali pemakaian, sehingga setiap memulai pengolahan
harus menggunakan enzim baru, dan hal ini tidaklah efisien dan
membutuhkan biaya yang lebih mahal. Kekurangan-kekurangan ini, dapat

5
diatasi melalui metode imobilisasi enzim untuk meningkatkan
kestabilannya, yaitu metode imobilisasi secara adsorpsi menggunakan
silika gel. Pemakaian silika gel sebagai material pendukung, karena
kemampuan adsorpsi permukaan dan intramolekul yang baik, memiliki
ruang pengunci (interlocking cavasities) yang memberikan media dengan
luas permukaan yang besar (Wulan dkk., 2008). Berbagai material
pendukung telah dilaporkan dalam literatur untuk meningkatkan
kestabilan enzim lipase imobil. Kelompok peneliti Inggris (Murray dkk.,
1997) telah melaporkan imobilisasi enzim lipase pada material pendukung
polimer polistirena EP 400 untuk menghidrolisis minyak bunga matahari.
Aktivitas enzim setelah penggunaan berulang tetap stabil. Imobilisasi
enzim lipase dengan menggunakan material pendukung sepharosa dan
resin dilaporkan oleh (Dosanjh dan Kaur, 2002). Enzim lipase imobil ini
dapat digunakan untuk sintesis ester. Penelitian Wulan dkk (2008),
terhadap enzim lipase imobil dari Rhizopus oryzae pada material
pendukung kitin dapat digunakan sebanyak tiga kali pemakaian berulang
untuk menghidrolisis minyak zaitun.
Enzim imobil adalah suatu enzim yang baik secara fisik maupun
kimia tidak dapat bebas bergerak sehingga dapat dikendalikan atau diatur
kapan enzim tersebut harus kontak dengan substrat. Imobilisasi
mencegah difusi enzim ke dalam campuran reaksi dan mempermudah
memperoleh kembali enzim tersebut dari aliran produk dengan teknik

6
pemisahan padat-cair yang sederhana, sehingga enzim tersebut
memungkinkan untuk digunakan kembali.
B. Rumusan Masalah
Berdasarkan uraian pada latar belakang di atas, maka penelitian
ini dipusatkan kajian beberapa masalah yaitu:
1. Bagaimanakah kondisi optimum memproduksi enzim lipase dari
A. oryzae pada kopra berjamur?
2. Bagaimanakah kemurnian dan karakteristik enzim lipase dari
A. oryzae?
3. Dapatkah enzim lipase imobil dari A. oryzae digunakan sebagai
biokatalisator yang efektif dan efisien? .
4. Dapatkah enzim lipase imobil dari A. oryzae menghidrolisis minyak
kelapa menjadi diasilgliserol?
C. Tujuan Penelitian
Tujuan penelitian adalah sebagai berikut :
1. Menentukan kondisi optimum produksi enzim lipase dari A. oryzae
pada kopra berjamur.
2. Memurnikan dan mengkarakterisasi enzim lipase dari A. oryzae
3. Menentukan unjuk kerja enzim lipase imobil dari A. oryzae sebagai
biokatalisator yang efektif dan efisien

7
4. Menggunakan enzim lipase imobil dari A. oryzae untuk hidrolisis
minyak kelapa menjadi diasilgliserol
D. Kegunaan Penelitian
Dari hasil penelitian ini diharapkan dapat memberi manfaat
sebagai berikut:
1. Menambah informasi ilmiah tentang mikroba A. oryzae yang dapat
diisolasi dari bahan baku lokal (kopra berjamur) dan potensial
menghasilkan enzim lipase
2. Enzim lipase yang dihasilkan dari A. oryzae dapat diimobilisasi dan
digunakan sebagai biokatalisator untuk menghasilkan pengemulsi
diasilgliserol dari minyak kelapa

8
BAB II
TINJAUAN PUSTAKA
A. Kopra
Kelapa (Cocos mucifera. L) adalah tanaman yang bernilai
komoditi komersial, karena semua bagian kelapa dapat dimanfaatkan
untuk berbagai keperluan. Budidaya tanaman kelapa merupakan salah
satu alternatif yang sangat menguntungkan. Salah satu pemanfaatan
kelapa dalam industri adalah pembuatan minyak kelapa yang sebelumnya
diproses terlebih dahulu menjadi kopra.
Kopra adalah daging buah tanaman kelapa yang telah dikeringkan
secara tradisional dengan cara penjemuran, pengasapan atau
pengeringan mekanis lainnya. Cara ini akan mencegah tumbuhnya jamur
dan bakteri yang dapat memakan daging buah kelapa dan merusak
minyak kelapa (Anonim, 2007). Namun demikian pada pengolahan kopra
masih ditemukan adanya kopra yang rusak (berjamur), seperti pada
Gambar 1. Kopra mengandung asam lemak tidak jenuh. Asam lemak
adalah asam organik yang terdapat sebagai ester trigliserida atau lemak,
baik yang berasal dari tumbuhan maupun hewan.

9
Gambar 1. Kopra berjamur
B. Uraian Aspergillus oryzae
Aspergillus merupakan jamur, yaitu tumbuhan dari divisi
Thallophyta yang memiliki ciri utama tubuh yang berbentuk talus, yaitu
belum dapat dibedakan dalam tiga bagian tubuh utama tumbuhan yang
disebut akar, batang dan daun, dan termasuk subdivisi fungi karena tidak
mempunyai klorofil. Jamur termasuk fungi sejati yang merupakan
organisme heterotropik dimana mereka memerlukan senyawa organik
untuk nutrisinya.

10
1. Taksonomi Aspergillus oryzae
Regnum : Plantae
Divisio : Thallophyta
Subdivisio : Fungi
Classis : Eumycetes
Subclassis : Ascomycetes
Ordo : Moniliales
Familia : Aspergillaceae
Genus : Aspergillus
Species : Aspergillus oryzae
Gambar 2. Aspergillus oryzae (Gandjar dkk, 1999) a. Konidiofor; b.Vesikel; c. Metula; d. Fialid; e. Konidia
A. oryzae mempunyai kepala konidia berwarna kuning kecoklatan
dengan dinding yang tipis. Panjangnya bisa mencapai 1-2 n m, dengan

11
diameter 7-10 µm. Visekel yang berada pada ujung konidiophor berbentuk
sedikit lonjong dengan diameter kurang lebih 100 µm. Strigmata berwarna
cokelat yang tersusun dalam dua seri. Konidia yang sudah matang
berwarna hitam. Aspergillus ada dimana-mana, di daerah kutub maupun
di daerah tropik, dan hampir pada setiap substrat.
Beberapa Aspergillus yang digunakan dalam industri fermentasi
adalah A. flavus dan A. oryzae. Di Jepang A. oryzae digunakan dalam
pabrik-pabrik sake. Fungsi utama jamur ini adalah untuk proses
saccharifikasi zat pati beras, dan juga digunakan pada pabrik-pabrik
shoyu (kecap asin) dan miso (tauco). Berbagai galur A. oryzae digunakan
pula dalam produksi enzim dengan nama dagang seperti: takadiastase,
polyzyme, digestin, oryzyme dan kashiwagidiastase (Judoamijoyo, 1992).
2. Fisiologi Aspergillus oryzae
a. Suhu pertumbuhan
Kebanyakan jamur bersifat mesofilik, yaitu tumbuh baik pada suhu
kamar. Suhu optimum pertumbuhan kebanyakan jamur adalah sekitar
(25 –30) 0C, tetapi beberapa diantaranya dapat tumbuh pada suhu
antara (35–37) 0C misalnya Rhizopus stolonifer, Aspergillus atau suhu
lebih tinggi, yaitu 45 0C, misalnya Absidia ramosa (Ganjar dkk., 1999).
Pada umumnya jamur dapat tumbuh pada kisaran pH yang luas, yaitu pH
(2–8,5). Beberapa jamur bersifat psikrotrofik, yaitu dapat tumbuh baik
pada suhu dingin, dan bahkan masih dapat tumbuh lambat pada suhu

12
di bawah suhu pembekuan, misalnya pada suhu -5 0C sampai -10 0C.
Semua spesies jamur bersifat aerobik, yaitu membutuhkan oksigen untuk
pertumbuhannya.
b. Nutrisi
Pada umumnya jamur dapat menggunakan berbagai komponen
makanan, dari yang sederhana sampai yang kompleks untuk nutrisinya
dalam mensintesis semua komponen selnya. Walaupun demikian jamur
bersifat heterotrof (sumber karbon hanya dari senyawa karbon organik).
Berbeda dengan bakteri, jamur tidak dapat menggunakan senyawa
karbon anorganik, misalnya karbondioksida. Karbon harus berasal dari
sumber organik, misalnya glukosa.
Beberapa spesies dapat menggunakan nitrogen; itulah sebabnya
media biakan untuk jamur biasanya mengandung pepton, pati, pektin,
protein atau lipid. Untuk menumbuhkan dan mengaktifkan isolat
Aspergillus yang merupakan organisme heterotrofik, dapat digunakan
media yang diperkaya (enrichment medium), yaitu media yang
ditambahkan zat tertentu, misalnya serum, darah, ekstrak hewan dan
tumbuhan serta komponen selular esensial yang tidak dapat disintesis
sendiri oleh suatu organisme dari sumber karbon atau nitrogen dasar
berupa asam-asam amino dan vitamin (Pelczar, 1986).

13
C. Media Pertumbuhan
Media yang digunakan untuk menumbuhkan mikroba di
laboratorium terbagi atas tiga jenis, yaitu : (a) media cair, (b) media padat,
dan (c) media semi padat. Media yang digunakan untuk pertumbuhan
mikroba harus mengandung komponen-komponen yang dibutuhkan
seperti : air, karbon, nitrogen, mineral dan lain-lain yang sesuai dengan
kebutuhan setiap jenis mikroba.
Menurut Somkuti dan Babel (1968), untuk memproduksi enzim
lipase dari Mucor pusillus digunakan media gandum wheat-bran dengan
pH 6,8, sedangkan untuk lipase dari Bacillus J33 digunakan media kaldu
(nutrient both) dengan pH 8,0. Nawani dkk (1998) dan Maia dkk (1999),
menggunakan media minyak zaitun-pepton untuk memproduksi enzim
lipase ekstraselluler dari fungi Fusarium solani.
Produksi enzim lipase dari Aspergillus carneus dan Penicillium sp,
Helisto dan Korpela (1998), menggunakan media modifikasi Czapek
dengan komposisi bahan yaitu: NaNO3, KHPO4, KCl, MgSO4.7H2O,
FeSO4, tepung kedelai, minyak zaitun dan agar serta pH media 7.
Menurut Liu dan Tsai (2003), n-heksadekana dan minyak zaitun berfungsi
sebagai sumber karbon untuk memproduksi enzim lipase alkali dari
A. radioresistens. Sumber nitrogen dalam media juga berpengaruh
terhadap produksi enzim lipase. Umumnya sumber nitrogen yang
digunakan adalah pepton dan ekstrak ragi, dapat meningkatkan produksi

14
enzim lipase dari Bacillus sp, Pseudomonas, dan Staphylococcus
haemolyticus (Gupta dkk., 2004).
D. Mikroba dan Kultur Fermentasi
Mikroba banyak digunakan dalam proses fermentasi di antaranya
adalah khamir, fungi (jamur) dan bakteri. Pada dasarnya tidak semua
khamir, fungi (jamur) dan bakteri dapat digunakan secara langsung tetapi
diperlukan seleksi dari masing-masing untuk menjamin berlangsungnya
proses fermentasi sesuai dengan tujuan. Mikroba bagi industri fermentasi
memang bersifat unik, karena persyaratan yang harus dipenuhi sangat
bervariasi dan sering kali sulit dipenuhi secara langsung. Suatu galur
khusus yang digunakan dalam memproduksi streptomisin, harus dapat
memproduksi zat tersebut cukup tinggi agar menurunkan biaya
operasionalnya. Selain itu tahan disimpan sebagai galur murni tanpa
penurunan sifat unggulnya. Persyaratan penting di luar daya produksi
suatu mikroba untuk industri, hendaknya tidak mengganggu kesehatan
manusia, hewan dan tumbuhan apalagi bersifat toksik. Apabila mikroba
tersebut bersifat toksik, hendaknya toksik yang dihasilkan mudah
dimatikan (Judoamijoyo, 1992).
E. Teknik Biakan Murni
Inokulasi adalah memindahkan bakteri atau fungi dari medium
yang lama ke medium yang baru. Inokulasi dapat dilakukan di dalam
suatu kotak berkaca yang disebut enkas untuk menghindari kontaminasi.

15
Bahan yang diinoklasikan pada medium disebut inokulum. Sel-sel dalam
inokulum akan terpisah sendiri-sendiri. Setelah inkubasi, sel-sel mikroba
individu ini memperbanyak diri sedemikian cepatnya sehingga dalam
waktu 18 sampai 24 jam terbentuklah massa sel yang dapat dilihat dan
dinamakan koloni. Setiap koloni yang berlainan dapat mewakili macam
organisme yang berbeda-beda, setiap koloni merupakan biakan murni
satu macam mikroorganisme.
F. Isolasi dan Pemurnian Enzim
Isolasi enzim perlu memperhatikan tujuan serta bentuk isolat
enzim yang diinginkan. Untuk keperluan penelitian dan analisis, hasil
isolasi enzim tidak diperlukan dalam jumlah banyak, tetapi mempunyai
tingkat kemurnian yang tinggi. Isolasinya memerlukan teknologi isolasi
enzim yang mempunyai sifat dan aktivitas maksimum (Suhartono, 1989).
1. Sentrifugasi
Sentrifugasi merupakan suatu cara pemisahan yang berdasarkan
kepada perbedaan kecepatan sedimentasi dari partikel-partikel molekul
yang disebabkan oleh adanya gaya sentrifrugal. Cara ini terutama
digunakan untuk memisahkan endapan yang sukar disaring dengan
saringan biasa (filter), dimana zat terlarut dapat dipisahkan dengan cepat
menuju pusat medan sentrifugal. Dalam hal ini partikel-partikel mula-mula
yang terdistribusi secara merata di dalam larutan, pada suatu kecepatan
perputaran tertentu akan bergerak meninggalkan larutan induknya dan

16
bila partikel-partikel terlarut tersebut lebih besar dari partikel pelarut, maka
akan memisah dan terjadi pengendapan. Sebaliknya partikel-partikel yang
memiliki berat jenis lebih kecil dari pelarutnya, akan terapung di
permukaan. Pada saat kesetimbangan tercapai, dimana konsentrasi zat
terlarut di bagian atas lebih kecil dari pada konsentrasi bagian bawahnya,
maka pada saat itulah terjadi pengendapan.
2. Fraksinasi
Hasil sentrifugasi diperoleh suatu larutan enzim kasar, selanjutnya
dilakukan metode pemurnian. Pemurnian enzim pada dasarnya
bergantung pada beberapa variabel diantaranya: pH, suhu, komposisi
pelarut dan sifat dari protein itu sendiri (ukuran, kelarutan, muatan dan
bentuknya). Fraksinasi protein dengan menggunakan garam, berdasarkan
atas kelarutan protein yang merupakan interaksi antara gugus polar
dengan air, interaksi ionik dengan garam dan daya tolak menolak protein
yang bermuatan sama. Dalam hal ini fenomena kelarutan protein ada dua
macam yaitu salting in dan salting out. Salting in adalah peristiwa dimana
dengan penambahan garam konsentrasi rendah pada larutan protein
dalam air, akan menurunkan koefisien aktivitas sehingga kelarutan protein
akan bertambah.
Bila konsentrasi garam dinaikkan sehingga kekuatan ion
bertambah besar, maka interaksi ion dari garam dengan air akan
bertambah, hal ini akan menyebabkan interaksi antara protein dengan air

17
menurun. Proses ini disebut salting out. Salting out sangat tergantung
pada hidrofobilitas protein. Peningkatan konsentrasi garam yang
ditambahkan secara bertahap, akan mengendapkan protein secara
bertahap pula, sehingga dapat digunakan untuk pemurnian protein. Pada
peristiwa salting out akan terjadi pengendapan protein, dimulai dari
kelarutan yang lebih kecil. Dalam larutan elektrolit kuat, penurunan
logaritma kelarutan suatu protein merupakan fungsi linier dari kenaikan
konsentrasi garam Colowick dan Kaplan (1957), yang dinyatakan
dalam persamaan:
Log s = β Ks µ (1)
dimana : S = Kelarutan protein dalam gram/liter
β = Konstanta intersep; juga merupakan kelarutan protein dalam
air murni
µ = Kekutan ion dari larutan
Ks = Konstanta kelarutan protein pada salting out
Hubungan antara kekuatan ion dengan konsentrasi larutan adalah :
µ = ½ Ci Zi2 (2)
dimana: µ = kekuatan ion
Ci = konsentrasi ion i
Zi = valensi ion i

18
Gambar 3. Kurva antara log s terhadap µ (Colowick dan Kaplan, 1957)
Amonium sulfat adalah salah satu jenis garam yang paling banyak
digunakan untuk mengendapkan protein enzim. Keuntungannya adalah
(1) dalam keadaan jenuh molaritasnya cukup tinggi sehingga dapat
mengendapkan sebagian besar protein; (2) panas pelarutannya rendah,
sehingga panas yang dihasilkannya mudah hilang; (3) bahkan pada
larutan jenuhnya (4,04 M pada 20 0C) memiliki kerapatan sekitar
1,235 gram per cm3, yang tidak cukup besar mengganggu sedimentasi
sebagian besar protein yang mengendap karena sentrifugasi; (4) larutan
amonium sulfat yang pekat mencegah atau membatasi pertumbuhan
bakteri, dan (5) dalam larutan amonium sulfat sebagian besar protein
terlindungi dari denaturasi. Berdasarkan keuntungan terakhir ini,
seringkali protein murni disimpan sebagai suspensi dalam larutan
amonium sulfat pekat (Englard dan Seiffer, 1990).
β
µ
Slope = - Ks
Log s

19
3. Dialisis
Dialisis dalam biokimia dikenal sebagai proses untuk
menghilangkan garam-garam dari larutan protein. Proses dialisis
diperlukan suatu membran yang bersifat semipermeabel, dapat menahan
zat-zat molekul besar tetapi melewatkan zat-zat molekul kecil. Adapun
membran yang dipakai pada umumnya adalah kantong selofan dengan
ukuran ketebalan dan panjang yang berbeda-beda.
Permeabilitas suatu kantong selofan tergantung pada ukuran dan
juga pada praperlakuan yang dilakukan. Pada dasarnya bahwa suatu
membran atau selaput itu permeabel, jika ia dapat melewatkan molekul-
molekul yang berberat molekul kurang dari 30.000 selama satu malam.
Permeabilitas suatu kantong membran tidak tergantung pada lamanya
dialisis. Jika suatu molekul tidak dapat melalui selaput maka selamanya ia
tidak akan lewat walaupun waktu dialisisnya diperpanjang. Kondisi lain
yang perlu diperhatikan dalam melakukan dialisis adalah pelarut. Secara
umum dikatakan bahwa kecepatan dialisis maksimal jika menggunakan
akuades. Pada hal suatu larutan ditentukan oleh pH dan kekuatan ionisasi
zat terlarut yang diperlukan untuk menstabilisasikan kondisi biomolekul
dalam larutan tersebut. Proses dialisis yang menyebabkan masuknya air
ke kantong membran adalah tekanan osmotik, oleh karena itu selalu
diusahakan supaya volume kantong selofan setelah tercapai
kesetimbangan masih normal, tidak mengalami kerusakan akibat tekanan
osmotik (Gambar 4). Sebaliknya dapat terjadi, misalnya jika biomolekul

20
yang keluar meninggalkan kantong selofan lebih cepat, sehingga kantong
selofan menjadi kempes. Sebagai contoh dari 200 mL isi kantong selofan
jika dibiarkan satu malam dapat bertambah volumenya menjadi beberapa
mililiter lagi.
awal dialisis kondisi saat kesetimbangan
Gambar 4. Pemisahan protein berdasarkan ukuran molekul dengan dialisis (Stryer, 1995).
Cara-cara untuk mempercepat pergerakan molekul:
a. Membuat luas permukaan membran sebesar mungkin, antara
lain dengan menambah panjang tabung (kantong).
b. Mengubah lapisan larutan yang berhubungan langsung dengan
membran secara terus menerus dengan mengaduk pelarut.
c. Mengganti pelarut pada selang waktu tertentu
kantong dialisis
larutan protein
bufer

21
4. Kromatografi kolom penukar ion
Prinsip yang mendasari fraksinasi protein dengan kolom penukar
ion, adalah adsorpsi molekul-molekul pada kolom penukar ion, dengan
cara merubah lingkungan ioniknya, molekul yang terikat akan dilepaskan
lagi. Protein dapat dipisahkan berdasarkan pada muatan netonya dengan
kromatografi penukar ion.
Jika suatu protein memiliki muatan positif pada pH 7, maka akan
terikat pada matrik kolom yang mengandung gugus karboksilat,
sebaliknya protein yang bermuatan negatif tidak akan terikat. Protein
bermuatan positif yang terikat pada suatu kolom, dapat dielusi dengan
gradien konsentrasi garam bufer pengelusi atau kalium klorida (gambar 5).
Ion kalium berkompetisi dengan gugus bermuatan positif protein untuk
terikat pada kolom. Protein yang memiliki densitas rendah muatan neto
positif akan terelusi yang pertama, kemudian diikuti protein yang memiliki
densitas muatan lebih tinggi. Selain faktor muatan neto, afinitas terhadap
matrik pendukung kolom, dapat mempengaruhi sifat-sifat protein pada
kolom penukar ion (Stryer, 1995).

22
Gambar 5. Kromatografi kolom penukar ion memisahkan protein berdasarkan muatan netonya (Steve, 2008)
5. Kromatografi kolom filtrasi gel
Prinsip yang mendasari kromatografi kolom filtrasi gel adalah
partisi ukuran molekul pada kolom yang mengandung butiran sangat kecil
berpori, yang terbuat dari polimer berhidrat tinggi gel. Jika molekul-
molekul protein dengan berbagai ukuran dielusikan lewat kolom yang
telah dicuci dan disetimbangkan dengan eluennya, maka molekul-molekul
yang lebih kecil dari ukuran pori gel akan masuk kedalam partikel-partikel
Protein bermuatan negatif mudah dipindahkan
Penambahan larutan KCl
Hanya protein bermuatan positif dipindahkan dengan kekuatan ionik yang tinggi

23
gel sebagai fasa stasioner. Sedangkan molekul-molekul yang lebih besar
dari pori gel akan ditolak oleh partikel gel dan masuk kedalam fasa mobil
diantara partikel gel (Gambar 6)
Gambar 6. Pemisahan protein berbeda ukuran dengan kromatografi filtrasi gel, protein besar keluar lebih awal daripada protein lebih kecil (Stryer, 1995)
Gaya gravitasi kedua fasa tersebut akan bergerak ke bawah
sepanjang kolom dan keluar sebagai eluat (Stryer, 1995). Matriks kolom
yang digunakan adalah berpori dan terbuat dari polimer yang tidak larut
namun bersifat hidratasi tinggi, seperti dextran atau agarosa yang
merupakan karbohidrat atau poliakrilamida. Nama dagang dikenal sebagai
sephadex, sepharosa dan bio-gel, yang memiliki diameter butiran 100 µm.
molekul kecil masuk ke pori
matriks polimer
molekul besar tidak dapat masuk ke pori

24
6. Elektroforesis
Umumnya biomolekul polimer bermuatan listrik, sehingga dapat
bergerak dalam medan listrik. Pergerakan partikel-partikel bermuatan
listrik oleh medan listrik melalui suatu pelarut disebut elektroforesis.
Fenomena ini dapat digunakan untuk karakterisasi molekul berdasarkan
kecepatan pergerakan dalam medan listrik. Sifat pergerakan tersebut
dapat digunakan untuk memisahkan protein dan makromolekul lain,
seperti DNA dan RNA (Stryer, 1995).
Kecepatan migrasi (V) suatu protein (atau molekul lain) didalam
suatu medan listrik adalah bergantung pada besarnya medan listrik (E),
muatan neto protein (z), serta koefisien gesekan (f):
V = EZ / f (3)
Gaya listrik (EZ ) yang menggerakkan molekul bermuatan kearah
elektroda dengan muatan berlawanan, adalah berlawanan dengan
viskositas, fv, yang timbul dari gesekan antara molekul yang bergerak
dengan medium. Koefisien gesekan (f) bergantung pada massa dan
bentuk molekul yang bermigrasi, serta viskositas (η) medium, untuk
bentuk partikel bulat dengan radius r, adalah : f = 6πηr
Pemisahan biomolekul secara elektroforesis umumnya dilakukan
di dalam gel atau medium pendukung padat seperti kertas dari pada di
dalam larutan bebas, dengan alasan gel bertindak sebagai penyaring
molekul yang mempertinggi proses pemisahan serta menekan naiknya
suhu yang dihasilkan arus listrik. Molekul yang lebih kecil dari poli gel

25
dengan mudah bergerak melalui gel, sebaliknya molekul yang lebih besar
tertahan pergerakannya.
Gambar 7. Pembentukan gel poliakrilamida. Ukuran pori dapat dikendalikan dengan cara mengatur konsentrasi akrilamida dan metilenbisakrilamida sebagai pembentuk ikatan silang (Stryer, 1995)
Gel poliakrilamida dipilih sebagai media pendukung untuk
elektroforesis, karena secara kimia inert dan mudah dibentuk dengan
polimerisasi akrilamida. Selain itu, ukuran porinya dapat dikontrol dengan
mengatur konsentrasi akrilamida dan metilenbisakrilamida (pereaksi
ikatan –silang) pada saat akan dilakukan reaksi polimerisasi (Gambar 7).

26
Sebagian besar protein dapat dipisahkan berdasarkan massanya
secara elektroforesis gel poliakrilamida pada kondisi terdenaturasi.
Sampel protein yang akan dielektroforesis terlebih dahulu direduksi
dengan β-merkaptoetanol, kemudian direaksikan dengan sodium
dedosilsulfat. β-merkaptoetanol akan memecahkan semua ikatan disulfida
(-S-S-) dalam protein maupun subunit protein sehingga struktur
sekundernya hilang. Sedangkan sodium dodesilsulfat (SDS) akan
berikatan dengan semua rantai polipeptida dan membentuk kompleks
SDS- polipeptida yang sangat anionik. Perlakuan protein dengan cara ini
akan menghasilkan bentuk yang seragam, dengan rasio muatan anionik
terhadap massa yang tetap sehingga dapat mengatasi gesekan pada gel
dengan konsentrasi yang tinggi.
Konsentrasi gel yang tinggi berfungsi sebagai penyaring molekul,
dimana porositas dan viskositas gel menentukan mobilitas protein.
Selanjutnya rantai polipeptida ini dielektroforesis pada gel poliakrilamida
dalam lingkungan bufer yang mengandung SDS dan β-merkaptoetanol.
Dengan adanya arus listrik maka semua rantai polipeptida yang
bermuatan negatif akan bergerak menuju anoda. Mobilitas rantai
polipeptida merupakan fungsi ukuran molekul, sehingga akan terjadi
proses pemisahan rantai-rantai polipeptida dalam bentuk pita-pita pada
gel akrilamida. Untuk menentukan letak pergerakan pita-pita protein maka
dilakukan pewarnaan. Mobilitas relatif setiap rantai polipeptida anionik
yang denaturasi merupakan fungsi log bobot molekul polipeptida. Dengan

27
menghitung mobilitas relatif rantai polipeptida standar terhadap pewarna
pelacak kemudian dialurkan terhadap log bobot molekul standar, maka
akan diperoleh kurva bobot molekul protein standar. Dengan demikian
cara ini dapat menentukan bobot molekul suatu protein.
G. Imobilisasi enzim
Enzim dapat dipertahankan agar penggunaannya efektif dan
efisien melalui teknologi imobilisasi enzim. Dikenal beberapa teknik
imobilisasi enzim seperti ikatan silang, penjebakan, dan pengikatan
secara kovalen pada bahan pendukung. Bahan pendukung yang
digunakan untuk imobilisasi enzim antara lain : glutaraldehid, polifenol,
chip silikon, naflon, poli-o-diamino benzena dan poli-vinil piridin (Davis
dkk., 1995). Aktivitas enzim imobil ditentukan oleh bahan pendukung
yang digunakan. Silika gel merupakan sebuah polimer anorganik disusun
oleh siloksan (Si-O-Si) dan distribusi silanol (Si-OH) pada permukaan
senyawa. Modifikasi kimia dapat terjadi dengan adanya polimer dari
kelompok silanol (Lee dkk., 2006).
1. Adsorpsi enzim pada permukaan zat padat
Adsorpsi secara fisik merupakan metode yang relatif mudah
dilakukan. Adsorben yang umum digunakan dari berbagai bahan organik
dan anorganik seperti alumina (aminosilase, amilase), selulosa (selulase),
tanah liat (katalase), kaca (urease) hidroksilapatit (NAD pirofosforilase),
karbon dan berbagai macam bahan silika (amilase). Adsorpsi terjadi

28
karena gaya Van der Waals, ikatan hidrogen dan interaksi hidrofobik
diantara molekul enzim dan molekul penyangga dicampurkan pada
kondisi lingkungan yang sesuai, selama waktu tertentu. Enzim terikat oleh
bahan dengan ukuran fisik yang lebih besar sehingga dapat dipisahkan
dari enzim bebas dengan cara filtrasi dan sentrifugasi (Suhartono, 1989).
Pengikatan terhadap enzim bersifat reversibel, sehingga enzim yang
teradsorpsi mungkin mengalami desorpsi dengan adanya substrat atau
menaikkan kekuatan ion.
2. Sifat-sifat enzim imobil
Berbagai jenis perubahan kimia fisika enzim terjadi selama proses
imobilisasi dilakukan, bergantung pada jenis metode yang digunakan.
Akibat dari proses imobilisasi yang merugikan adalah turunnya aktivitas
spesifik enzim; sedangkan manfaat yang menyebabkan metode ini
berkembang adalah meningkatnya stabilitas enzim dan daya tahannya
terhadap kondisi lingkungan ekstrim seperti pH dan suhu. Imobilisasi
menyebabkan enzim yang telah berubah ini bersifat lebih tahan lama dan
dapat dipakai berulang-ulang. Pada umumnya masa aktif enzim imobil
melebihi masa aktif enzim terlarut. Apabila substrat atau produk yang
dihasilkan sensitif terhadap pH, maka teknologi imobilisasi
memungkinkan memilih polimer pengikat yang sesuai, sehingga kisaran
pH optimum enzim imobil sesuai dengan stabilitas substrat maupun
produknya. Perubahan konformasi molekul enzim yang menimbulkan

29
perubahan stereokimiawi dan muatan total pada sisi aktif enzim akan
mengubah daya katalitik enzim terhadap substratnya. Hal ini tercermin
pada perubahan beberapa parameter kinetika enzim. Konstanta kinetika
enzim imobil biasanya berubah dari bentuk aslinya. Imobilisasi dapat
meningkatkan atau menurunkan nilai Km enzim. Penurunan Km
mencerminkan reaksi yang lebih cepat dibandingkan dengan reaksi pada
enzim bebas. Penurunan Km enzim terjadi apabila muatan polimer
penyangga dan muatan substrat terjadi tarik menarik elektrostatik
diantara polimer penyangga dan substrat yang akan membantu
meningkatkan konsentrasi substrat di sekitar enzim imobil. Peningkatan
Km oleh proses imobilisasi berimplikasi bahwa konsentrasi substrat yang
lebih tinggi diperlukan untuk mencapai kecepatan reaksi yang sama pada
enzim bebas. Perubahan konformasi pada molekul enzim dapat
meningkatkan Km, karena menurunnya daya gabung antara enzim dan
substrat. Perubahan kimiawi pada proses imobilisasi yang diakibatkan
oleh teknik pengikatan kovalen biasanya menyebabkan peningkatan Km.
Kadang-kadang ukuran partikel enzim imobil yang terlalu besar dapat
meningkatkan harga Km enzim imobil. Hal ini berkaitan dengan pengaruh
difusi substrat (Suhartono, 1989).
H. Uraian enzim lipase
Enzim lipase (triasilgliserol hidrolase, EC.3.1.1.3) adalah enzim
yang aktif mengkatalisis hidrolisis ikatan ester trigliserida antar permukaan
air-lemak. Dalam kondisi tertentu, enzim lipase dapat mengkatalisis reaksi

30
sebaliknya (sintesis, reaksi esterifikasi) membentuk gliserida dari asam
lemak dan gliserol. Enzim lipase adalah suatu glikoprotein dengan berat
molekul 48 kDa dan dibuat serta disekresikan oleh kelenjar pankreas
(gambar 8). Enzim lipase bekerja terhadap senyawa yang tidak larut
dalam air dan hanya dapat mengolah lemak yang bersinggungan dengan
permukaan air.
Di alam enzim lipase terdapat dimana mana dan diproduksi oleh
beberapa tumbuhan, binatang dan mikroorganisme. Sebagian besar
enzim lipase diproduksi oleh mikroorganisme, yaitu bakteri dan jamur.
Enzim lipase adalah kelompok enzim yang secara luas penggunaannya
dalam bioteknologi dan kimia organik. Hal ini berhubungan dengan
kelebihan enzim ini, seperti aktivitas enzim pada temperatur dan cakupan
pH yang luas.
Gambar 8. Enzim lipase (Brown, 1997)

31
Studi biokimia enzim lipase termolabil maupun termostabil telah
dilaporkan. Enzim lipase dari mikroba termolabil berhasil dimurnikan dan
dikarakterisasi dari berbagai sumber antara lain: Pseudomonas fragi
CRDA 037, Aspergillus terreus, Mucor miemalis, dan Staphylococcus
haemolyticus (Kermasha dkk., 1997; Davidson dkk., 1998; Comeau dkk.,
1999).
Selain enzim lipase termolabil, berbagai laporan penelitian
mengungkapkan bahwa enzim lipase termostabil diproduksi dari bakteri
termofilik, misalnya Bacillus thermotenulatus (Schmid dkk.,1994), Bacillus
sp J33 (Kaur dkk., 1998), Bacillus sp THL 027 (Luchai dkk., 1999).
Bacillus thermotenulatus menghasilkan dua jenis lipase yaitu lipase BTL 1
dan BTL 2. Gen lipase BTL 2 telah diklon dan urutan nukleotida serta
sifat-sifat enzimnya telah diketahui (Schmid dkk., 1997; 1998).
Karakteristik enzim lipase termostabil dari kelompok Bacillus
termofilik telah diketahui, misalnya bobot molekul sekitar 69 kDa. Enzim
lipase dari Bacillus thermotenulatus dan Bacillus sp J33 stabil dalam
pelarut organik dan diaktifkan dengan Triton X-100, namun berbeda
kespesifikan terhadap substrat (Schmid dkk., 1997; Kaur dkk., 1998).
Enzim lipase dari Bacillus thermoleonovorans ID-1 mempunyai suhu
optimum 70 0C, sedangkan enzim lipase dari Bacillus sp THL 027 suhu
optimumnya 65 0C.
Enzim lipase yang diisolasi dari Bacillus sp J33 bersifat alkali, dan
kelarutan yang tinggi dalam pelarut organik sehingga dapat digunakan

32
sebagai katalis dalam sintesis senyawa ester dan imobilisasi enzim dalam
n-heksan (Nawani dkk., 1998). Demikian pula enzim lipase dari Candida
rugosa diketahui mempunyai kemampuan yang tinggi untuk
mengkonsentrasikan asam lemak omega-3 dalam campuran gliserida
dengan cara hidrolisis minyak ikan (Utami dkk., 2005). Karakteristik enzim
lipase dari beberapa mikroba lainnya pada Tabel 1.
Tabel 1. Karakteristik enzim lipase beberapa mikroba
Mikroba BM(KDa)
pH optimum
Suhu optimum (0C)
Aktivator
Yarrowia lipolitica (Mingrui Yu dkk., 2007)
38 8,0 40 Ca; Mg
P. aeruginosa MB 5001(Sharma dkk., 2001)
29 8,0 55 Ca
Bacillus cereus C71(Shaoxin dkk., 2007)
42 9,0 33 Ca; Mg;Na
Mucor sp ( Abbas dkk., 2002)
42 7,0 30 Ca; Co
Enzim lipase dapat mengkatalisis pembentukan monoasilgliserol
dan diasilgliserol melalui tiga cara (Gracia dkk., 1996). (1) esterifikasi dari
asam lemak bebas dan gliserol, (2) hidrolisis dari minyak, (3)
transesterifikasi yaitu reaksi transfer asil antara ester asam lemak/minyak
dengan alkohol seperti etanolisis atau gliserolisis.
Salah satu contoh reaksi yang dikatalisis oleh enzim lipase adalah
reaksi hidrolisis, yaitu hidrolisis triasilgliserol menjadi monoasilgliserol
dan diasilgliserol serta asam lemak bebas .

33
Gambar 9. Reaksi hidrolisis triasilgliserol oleh enzim lipase (Kuchel dan Ralston,1988)
Monoasilgliserol terdiri dari satu asam lemak dan dua gugus
hidroksil bebas yang terikat pada satu molekul gliserol. Bagian asam
lemaknya atau rantai asil lemaknya bersifat lipofilik dan dapat bercampur
dengan bahan-bahan yang berlemak, sedangkan gugus hidroksilnya
bersifat hidrofilik dapat bercampur dengan air (O’Brien, 1998). Bagian
asam lemak dapat teresterifikasikan ke satu, dua atau tiga gugus hidroksil
menjadi monoasilgliserol, diasilgliserol dan triasilgliserol.
Kespesifikan enzim lipase dalam menghidrolisis triasilgliserol
menjadi monoasilgliserol dan diasilgliserol pada beberapa penelitian yaitu,
lipase dari Rhizomucor miehei spesifik pada posisi sn 1,3- DAG, lipase
dari Candida antartica spesifik pada posisi sn -1(3) - MAG ( Blasi dkk.,
2007).

34
Diasilgliserol dan monoasilgliserol dalam industri pangan
digunakan sebagai pengemulsi pada pengolahan margarine, mentega
kacang (peanut butter), whitener, pudding, roti, biskuit dan kue-kue kering
berlemak lainnya (Malundo dan Resurreccion, 1994; Igoe dan Hui, 1996)
dalam Hasanuddin 2001.
Pada suatu reaksi transesterifikasi atau reaksi alkoholis satu mol
triasilgliserol bereaksi dengan tiga mol alkohol untuk membentuk satu mol
gliserol dan tiga mol alkohol ester asam lemak berikutnya. Proses tersebut
merupakan suatu rangkaian dari reaksi reversibel yang didalamnya
molekul triasilgliserol diubah satu tahap demi tahap, diasilgliserol,
monoasilgliserol , dan gliserol (Syah, 2005).
I. Kerangka konseptual
Mikroba asal Indonesia baru sebagian kecil diketahui dapat
memproduksi bahan-bahan bernilai ekonomi. Potensi alam tersebut
hingga saat ini belum dimanfaatkan secara optimal. Hingga saat ini
kebutuhan enzim lipase mikroba untuk keperluan industri masih diimpor.
Kelapa adalah bahan baku dalam pembuatan minyak nabati. Pengolahan
kelapa menjadi kopra didapatkan kopra yang rusak (berjamur). Survei
lapangan menunjukkan bahwa 1- 5% kelapa yang diolah menjadi kopra,
dapat berjamur sehingga potensial menjadi limbah. Kopra berjamur dapat
dimanfaatkan sebagai sumber mikroba penghasil enzim lipase. Kopra
berjamur mengandung berbagai macam mikroba, salah satu diantaranya

35
adalah A. oryzae. Jamur ini dapat memproduksi enzim lipase melalui
proses fermentasi.
Minyak nabati yang diproduksi dari kopra bernilai ekonomi rendah,
oleh karena itu minyak nabati dapat ditingkatkan nilai ekonominya dengan
mengolahnya menjadi diasilgliserol bernilai ekonomi tinggi, oleh enzim
lipase yang diproduksi dari A. oryzae pada kopra berjamur.
Gambar 10. Diagram kerangka konseptual
J. Hipotesis
Kopra berjamur
Minyak kelapa(nilai ekonomi rendah)
Kopra Kelapa
Limbah
Aspergillus oryzae
Diasilgliserol(nilai ekonomi tinggi)
Enzim lipase imobil
Enzim lipase
Proses fermentasi

36
1. Waktu fermentasi, konsentrasi media produksi dan kecepatan
pengadukan berpengaruh terhadap produksi enzim lipase dari
A. oryzae pada kopra berjamur.
2. Enzim lipase imobil efektif dan efisien sebagai biokatalisator.
3. Enzim lipase imobil dapat digunakan untuk menghidrolisis minyak
kelapa menjadi diasilgliserol.
BAB III

37
METODE PENELITIAN
A. Desain Penelitian
Penelitian ini dilakukan dengan menggunakan pola dan tahap-
tahap yang sesuai untuk memproduksi enzim dari mikroba (jamur).
Adapun pola dan tahap-tahap tersebut meliputi: pembuatan media (media
agar miring, media inokulum dan media produksi), isolasi mikroba
penghasil enzim lipase pada kopra berjamur, produksi, karakterisasi,
pemurnian dan imobilisasi serta aplikasi enzim lipase sebagai
biokatalisator untuk menghidrolisis minyak kelapa menjadi diasilgliserol.
B. Waktu dan lokasi penelitian
Penelitian ini dilakukan mulai pada bulan Agustus 2008 – Maret
2010 , lokasi penelitian dilakukan di Laboratorium Biokimia Jurusan Kimia
FMIPA, Laboratorium Bioteknologi dan Mikrobiologi Pangan Teknologi
Hasil Pertanian UNHAS, Laboratorium Bioteknologi dan Biokimia PAU
IPB, Laboratorium Mikrobiologi Fakultas Farmasi UMI dan Laboratorium
Kesehatan Makassar.
C. Bahan dan Alat yang Digunakan

38
1. Bahan
Bakto agar (Merck), pepton (Merck), natrium karbonat (Merck),
natrium klorida (Merck), KH2PO4 (Merck), FeSO47H2O (Merck), Na2S2O3
(Merck), AgNO3 (Merck), minyak zaitun, p-nitrofenilbutirat (Sigma),
p-nitrofenol (Sigma), glisin (Merck), bufer Tris-HCl (Merck), DMSO
(Merck), bufer borat (Merck), Lowry A (Merck), Folin cioucalteu (Merck),
Lowry B (Merck), bovin serum albumin (BSA) (Merck), β-merkaptoetanol
(Merck), gliserol (Merck), amonium sulfat (Merck), metanol (Merck), asam
asetat glasial (Merck), formalin (Merck), etanol (Merck), kalium hidroksida
(Merck), nitrogen dioksida (Merck), TEMED (Merck), Q sepharosa FF
(Sigma), kantong selofan, sephadex G-75 (Sigma), SDS-PAGE 10%
dengan marker: posforilase-b (116 kDa); BSA (66,2 kDa); ovalbumin (45
kDa); karbonik anhidrase (35 kDa); rease BSP 981 (25 kDa); β-
laktoglobulin (18,4 kDa); lisozim (14,4 kDa) (Pharmacia, Biotech),
polietilen-glikol (Merck), silika gel (Merck), 1,2-DAG; 1,3 - DAG dan MAG
(Sigma), lipase komersial (porcine pancreatin) (Sigma), minyak kelapa,
tauge, kopra berjamur, A. oryzae.
2. Alat
Alat yang digunakan dalam penelitian ini adalah: cawanpetri dan
alat-alat gelas yang umum digunakan di laboratorium, mikropipet,
mikroskop elektron (Nevo), autoklaf (Napco Model 8000-DSE), inkubator
(Memmert), sentrifuse 5000 rpm suhu 4 0C (Universal 320 R), shaker
incubator (BL Barnstead/Lab-line Max Q 4000) , spektronik 20 D+

39
(Thermo), spektrofotometer UV-Vis (Farmacia), sentrifuse 10.000 rpm
suhu 4 0C (Sorval Super T 21), kromatografi kolom penukar ion,
kromatografi kolom filtrasi gel, seperangkat alat elektroforesis (Apelex), pH
meter, kromatografi gas (G 1800 C GCD System, Hewlett Packerd).
D. Prosedur Kerja
Penelitian dilakukan dalam beberapa tahap kerja, sebagai berikut:
1. Pengambilan sampel
Sampel berupa kopra berjamur diperoleh dari Campalagian
Kabupaten Polewali Mandar.
2. Pembuatan media
Pembuatan media meliputi: media agar miring, media inokulum
dan media produksi menggunakan metode Pirman dan Aryanta (2003).
a. Media agar miring/media cawan petri
Media agar miring terdiri dari bahan-bahan sebagai berikut:
pepton 0,25%; KH2PO4 0,1%; FeSO47H2O 0,001%; bakto agar 1,5%;
minyak zaitun 0,5%, kemudian semua bahan dilarutkan dalam ekstrak
tauge 100 mL (tauge 10 gram direbus dengan akuades sebanyak 100 mL)
dan pH media diatur sekitar 7. Selanjutnya larutan dipanaskan di atas hot
plate. Setelah dipanaskan media agar dituang ke dalam tabung reaksi dan
ditutup dengan kapas, kemudian disterilisasi dalam autoklaf selama

40
15 menit pada suhu 121 0C dan tekanan 1 atm. Setelah sterilisasi, media
di dinginkan pada suhu kamar dengan posisi dimiringkan sampai
memadat, kemudian disimpan pada suhu kamar. Untuk media agar di
cawan petri, media dan cawan petri disterilkan secara terpisah. Media
agar steril dituang ke dalam cawan petri, kemudian disimpan pada suhu
kamar.
b. Media inokulum
Penyiapan media inokulum menggunakan bahan-bahan sebagai
berikut: pepton 0,25%; KH2PO4 0,1%; FeSO47H2O 0,001%; minyak zaitun
0,5%, kemudian semua bahan dilarutkan dalam 100 mL akuades dan pH
media diatur sekitar 7. Selanjutnya larutan dipanaskan di atas hot plate.
Setelah dipanaskan media dituang ke dalam Erlenmeyer 250 mL,
kemudian disterilisasi dalam autoklaf selama 15 menit pada suhu 121 0C
dan tekanan 1 atm. Setelah sterilisasi, didinginkan pada suhu kamar.
c. Media produksi
Penyiapan media produksi menggunakan bahan-bahan sebagai
berikut: pepton 0,5%; KH2PO4 0,1%; FeSO47H2O 0,001%; minyak
zaitun 1%, kemudian semua bahan dilarutkan dalam 100 mL akuades
dan pH media diatur sekitar 7. Selanjutnya larutan dipanaskan di atas hot
plate. Setelah dipanaskan dituang ke dalam Erlenmeyer 250 mL,
kemudian disterilisasi dalam autoklaf selama 15 menit pada suhu 121 0C
dan tekanan 1atm. Setelah sterilisasi, didinginkan pada suhu kamar.
3. Isolasi mikroba penghasil enzim lipase pada kopra berjamur

41
Mikroba pada kopra berjamur dipindahkan dengan menggunakan
ose ke dalam tabung reaksi yang mengandung NaCl 0,9% sebanyak
10 mL. Selanjutnya dilakukan pengenceran secara berseri dari 10-1
sampai dengan 10-9. Setiap tabung reaksi yang mengandung biakan
diambil 1 mL, kemudian dituang ke dalam cawan petri steril lalu
ditambahkan media agar yang telah disterilkan. Media agar cawan petri
yang mengandung biakan diinkubasi pada suhu 37 0C selama 4 hari.
Mikroba yang tumbuh pada media agar cawan petri dipindahkan lagi ke
media agar cawan petri yang lain, dan selanjutnya ke media agar miring
untuk mendapatkan kultur murni. Kultur yang diperoleh diidentifikasi
secara makroskopis dan mikroskopis.
4. Produksi enzim lipase
a. Isolasi enzim lipase dan penentuan waktu produksi optimum
Biakan mikroba hasil isolasi dimasukkan sebanyak 5 ose ke
dalam 100 mL media cair (media inokulum) dan dikocok dengan shaker
incubator selama 4 hari dengan kecepatan 150 rpm pada suhu 37 0C.
Biakan dari media inokulum diambil sebanyak 5 mL kemudian
ditumbuhkan ke dalam media produksi sebanyak 100 mL untuk setiap
Erlenmeyer (digunakan 10 Erlenmeyer) dan dikocok dengan shaker
incubator kecepatan 150 rpm pada suhu 37 0C. Setiap hari media
produksi pada Erlenmeyer diambil, dipisahkan antar sel dan supernatan
dengan sentrifuse pada kecepatan 3500 rpm selama 30 menit pada suhu
4 0C, kemudian disaring dengan kertas wathman. Supernatan yang

O∥ + HO – C – (CH2) 2CH 3
Lipase
OH
NO2 NO2
42
diperoleh merupakan ekstrak kasar enzim lipase, selanjutnya diuji
aktivitas enzim dan kadar proteinnya. Pengukuran aktivitas enzim dan
kadar protein dilakukan setiap hari untuk menentukan pada hari keberapa
produksi enzim lipase diproduksi secara maksimal. Massa sel ditentukan
berdasarkan berat sel kering. Penentuan berat sel kering dilakukan
dengan menimbang cawan petri kosong, cawan petri berisi sel, dan
cawan petri berisi sel yang telah dipanaskan dalam oven pada suhu
80 0C, dilakukan sampai diperoleh berat konstan.
b. Uji aktivitas enzim lipase
Aktivitas enzim lipase ekstrasel ditentukan dengan menggunakan
substrat p-nitrofenilbutirat seperti metode yang digunakan Vorderwulbecke
dkk (1992), yang dimodifikasi dengan prosedur sebagai berikut: sebanyak
0,3 mL larutan enzim lipase atau blanko ditambahkan ke dalam bufer
2,67 mL yang mengandung Tris-HCl 0,05 M pH 7,0. Reaksi diawali
dengan penambahan 0,03 mL substrat p-nitrofenilbutirat 0,1 M (pelarut
dimetilsulfoksida) dan dikocok secepatnya lalu campuran reaksi diinkubasi
selama 10 menit pada suhu 37 0C, kemudian campuran reaksi diukur
serapannya dengan spektronik 20 D+ pada panjang gelombang 410 n m.
Aktivitas enzim lipase dihitung berdasarkan p-nitrofenol yang terbentuk
dari hasil hidrolisis enzim lipase terhadap substrat p-nitrofenilbutirat
(Shaoxin Chen dkk., 2007).
O∥O – C – (CH2) 2CH 3)

H2O
43
Gambar 11. Reaksi hidrolisis enzim lipase terhadap p-nitrofenilbutirat
c. Penentuan kadar protein enzim
Kadar protein enzim lipase ditentukan dengan menggunakan
metode Lowry dkk (1951), dengan prosedur sebagai berikut: sebanyak
4 mL larutan enzim lipase direaksikan dengan 5,5 mL Lowry B, kemudian
dikocok dan dibiarkan selama 10 menit. Campuran reaksi ditambahkan
dengan 0,5 mL Lowry A, dikocok dan dibiarkan selama 30 menit kemudian
serapan protein diukur dengan spektronik 20 D+ pada panjang gelombang
710 n m. Sebagai blanko, dipipet 4 mL air suling perlakuannya sama
dengan prosedur uji kadar protein enzim lipase. Untuk kurva standar
digunakan protein standar bovine serum albumin (BSA) pada berbagai
konsentrasi, (yaitu: 0,02; 0,04; 0,06; 0,08; 0,1; 0,12; 0,14; 0,16; 0,18 dan
0,20) mg/mL.
Lowry A : Folin cioucalteu : H2O = 1: 1
Lowry B : 100 mL Na2CO3 2% dalam NaOH 0,1 N ditambahkan dengan
1 mL CuSO45H2O 1% dan 1 mL natrium kalium tartrat 2%
5. Optimasi produksi enzim lipase
p-nitrofenilbutirat (bening)
p-nitrofenol (kuning)
Asam butirat

44
Optimasi produksi enzim lipase dilakukan dengan membuat media
inokulum dan media produksi (seperti pada bagian 2.b dan 2.c). Pada
pembuatan media produksi dilakukan variasi konsentrasi pepton, (yaitu:
0,5; 0,75; 1,0; 1,25; dan 1,50 )%. Konsentrasi pepton optimum digunakan
untuk memvariasikan konsentrasi minyak zaitun (yaitu: 1; 2; 3; 4 dan 5)%.
Konsentrasi minyak zaitun optimum digunakan untuk memvariasikan
kecepatan pengadukan (yaitu: 50; 100; 150; 200 dan 250) rpm, sehingga
akan diperoleh komposisi media produksi dan kecepatan pengadukan
optimum untuk memproduksi enzim lipase dari A. oryzae.
6. Pemurnian enzim lipase
Setelah diperoleh komposisi media produksi dan kecepatan
pengadukan optimum, maka komposisi media ini digunakan untuk
memproduksi enzim lipase seperti pada percobaan sebelumnya. Enzim
lipase yang diperoleh dimurnikan dalam beberapa tahap sebagai berikut:
a. Fraksinasi dengan amonium sulfat
Tahap fraksinasi dimulai dengan menentukan dahulu berapa
jumlah amonium sulfat harus ditambahkan pada setiap tingkat kejenuhan
dengan memperhatikan volume awal ekstrak kasar enzim yang diperoleh.
Kedalam larutan ekstrak kasar enzim ditambahkan sedikit demi sedikit
padatan amonium sulfat sambil diaduk dengan pengocok magnetik
dengan kecepatan lambat, hingga mencapai tingkat kejenuhan tertentu
dari (0-100)%. Selanjutnya endapan dibiarkan selama satu malam untuk
mencapai kesetimbangannya. Setelah itu di sentrifugasi pada suhu 4 0C

45
dengan kecepatan 10.000 rpm selama 20 menit . Pada tabung sentrifuse
akan terpisah antara supernatan dan endapan. Supernatan yang
diperoleh dilanjutkan untuk tahap fraksinasi selanjutnya, sedangkan
endapan dilarutkan dengan bufer borat 0,2 M pH 8,2. Fraksinasi amonium
sulfat tertinggi aktivitas enzimnya didialisis.
b. Proses dialisis
Hasil fraksinasi diperoleh pada tingkat kejenuhan amonium sulfat
dari (60-80)% dengan endapan sebanyak 0,8000 gram. Selanjutnya
dilarutkan ke dalam 3 mL larutan bufer borat 0,2 M pH 8,2. Dari larutan ini
diambil 1,5 mL ditambahkan dengan 1,5 mL bufer borat, sehingga volume
larutan menjadi 3 mL. Larutan dimasukkan ke dalam kantong selofan
kemudian didialisis dengan bufer borat, konsentrasi 0,05 M, diaduk
dengan pengaduk magnetik stirer selama satu malam pada suhu 5 0C.
Setiap 3 jam dilakukan penggantian bufer.
c. Kromatografi kolom penukar ion
Pemurnian enzim lipase dengan menggunakan kromatografi kolom
penukar ion dilakukan berdasarkan metode Mingrui Yu dkk ( 2007), yang
dimodifikasi.
Cara kerja kromatografi kolom penukar ion
Enzim lipase hasil dialisis sebanyak 3 mL dimasukkan ke dalam
kolom yang telah diisi matriks Q sepharosa FF (panjang kolom 14,5 cm
dan diameter 1 cm) sebelumnya telah disetimbangkan dengan bufer borat
0,05 M pH 8,2 selama satu malam. Setelah semua sampel masuk ke

46
dalam matriks, gradien-mixer dan fraction collector dioperasikan. Gradien-
mixer tersebut berisi bufer borat 0,05 M, pH 8,2 dan NaCl (0-04 M)
sebagai larutan pengelusi. Volume tiap fraksi 3,0 mL dan setiap fraksi
diukur serapan protein dan aktivitas enzimnya. Fraksi aktif dengan
aktivitas enzim yang tinggi dikumpulkan dan digunakan untuk kromatografi
kolom filtrasi gel.
d. Kromatografi kolom filtrasi gel
Fraksi aktif yang diperoleh dari kromatografi kolom penukar ion,
dimasukkan ke dalam kolom dengan matriks sephadex G-75 (panjang
kolom 35 cm dan diameter 1 cm) sebelumnya telah disetimbangkan
dengan bufer borat 0,05 M pH 8,2 selama satu malam. Selanjutnya dielusi
dengan bufer yang sama. Volume tiap fraksi sebanyak 3,0 mL dan setiap
fraksi diukur serapan protein dan aktivitas enzimnya. Fraksi aktif dengan
aktivitas enzim tertinggi digunakan untuk elektroforesis.
e. Elektroforesis dan pewarnaan perak
Elektroforesis dan pewarnaan perak dilakukan untuk mendeteksi
kemurnian enzim paling tinggi dari fraksi hasil kolom filtrasi gel.
Elektoforesis menggunakan gel poliakrilamida natrium dedosil sulfat
(SDS-PAGE), dengan konsentrasi gel pemisah poliakrilamida 10% dan
gel penahan poliakrilamida 4% (Bollag dan Edelstein,1991). Sampel
fraksi hasil pemurnian enzim sejumlah 30 µL terlebih dahulu diinkubasi
selama 3 menit pada suhu 100 0C. Selanjutnya dilakukan loading pada
setiap sumur gel, kemudian proses elektroforesis dilakukan dengan

47
kondisi tegangan listrik 70 Volt, arus sebesar 100 mA. Setelah selesai,
gel difiksasi dengan larutan fiksasi, yaitu: (125 mL metanol ditambahkan
dengan 25 mL asam asetat glacial 10%, 0,125 mL formalin dan 99,85 mL
akuabides) selama semalam. Selanjutnya perendaman diganti dengan
larutan enhancer yaitu: (0,05 gram Na2S2O3 dalam 250 mL akuabides)
selama 2 menit. Setelah gel dicuci dengan akuabides sebanyak 2 kali
masing-masing selama 20 detik, gel diwarnai dengan larutan perak nitrat
yaitu: (0,1 gram AgNO3 dalam 50 mL akuabides dan 0,038 mL formalin),
dilakukan selama 20 menit. Terakhir larutan perendaman gel diganti
dengan larutan natrium karbonat yaitu: (3 gram Na2CO3 dalam 50 mL
akuabides ditambahkan dengan 0,025 mL formalin dan 1 mL Na2S2O3
0,004%) selama 10 menit, kemudian dilakukan penghentian reaksi
dengan larutan fiksasi.
7. Karakterisasi enzim lipase murni
Karakterisasi enzim lipase murni meliputi penentuan suhu dan pH
optimum.
a. Penentuan suhu optimum

48
Penentuan suhu optimum enzim lipase dilakukan sesuai dengan
prosedur penentuan aktivitas enzim lipase, dengan memvariasikan suhu
(yaitu : 20 ; 25 ; 30 ; 35 ; 40 ; 45 ; dan 50) 0C.
b. Penentuan pH optimum
Penentuan pH optimum enzim lipase dilakukan sesuai dengan
prosedur penentuan aktivitas enzim lipase, dengan memvariasikan pH
pada suhu optimum menggunakan bufer borat 0,2 M yaitu pH: 7,0; 7,2;
7,4; 7,6; 7,8; 8,0; 8,2; 8,4; 8,6; 8,8 dan 9,0.
c. Penetapan konstante kinetika Km dan Vmaks
Penetapan konstante kinetika Km dan Vmaks dilakukan sesuai
dengan prosedur penentuan aktivitas enzim lipase, dengan
memvariasikan konsentrasi substrat pada suhu dan pH optimum berturut-
turut: ( 0,05; 0,10; 0,15 dan 0,20) M.
8. Imobilisasi enzim lipase murni
Enzim lipase murni diimobilisasi dengan metode adsorpsi
menggunakan silika gel (Nawani dkk., 2006).
Sebanyak 2,5 mL enzim lipase murni ditambahkan dengan
0,5 gram silika gel, lalu campuran di shaker selama waktu tertentu
(0 – 120) menit pada suhu kamar. Selanjutnya disentrifugasi selama
5 menit dengan kecepatan 1000 rpm. Jumlah enzim lipase yang tidak
terimobilisasi pada supernatan ditentukan dengan menggunakan
spektronik 20 D+ pada panjang gelombang 410 n m untuk menentukan

49
aktivitas enzim lipase dan pada panjang gelombang 710 n m untuk
menentukan kadar protein.
Jumlah enzim lipase imobil dihitung dengan rumus:
CE = C0 - Ct (4)
dimana : CE = jumlah enzim terimobilisasi (U/mL)
C0 = jumlah enzim sebelum imobilisasi (U/mL)
Ct = jumlah enzim pada waktu t (U/mL)
9. Karakterisasi enzim lipase imobil
Karakterisasi enzim lipase imobil meliputi: penentuan pH, suhu
optimum, kestabilan termal dan operasional enzim lipase imobil.
a. Penentuan pH dan suhu optimum enzim lipase imobil
Sebanyak 1,5 mL enzim lipase murni ditambahkan dengan
0,4 gram silika gel. pH larutan dibuat bervariasi yaitu pH : 7,0; 7,2; 7,4;
7,6; 7,8; 8,0; 8,2; 8,4; 8,6; 8,8 dan 9,0 menggunakan bufer borat 0,2 M.
Larutan tersebut di shaker selama 90 menit (waktu optimum imobilisasi)
pada suhu kamar, selanjutnya disentrifugasi selama 5 menit dengan
kecepatan 1000 rpm. Jumlah enzim lipase yang tidak terimobilisasi pada
supernatan ditentukan dengan menggunakan spektronik 20 D+ pada
panjang gelombang 410 n m.
Cara kerja penentuan suhu optimum enzim lipase imobil
dilakukan dengan membuat variasi suhu (yaitu: 20; 25; 30; 35; 40; 45;
50; dan 55) 0C. Pengaturan suhu dilakukan dengan menginkubasi larutan

50
dalam shaker inkubator selama 90 menit, selanjutnya disentrifugasi
selama 5 menit dengan kecepatan 1000 rpm. Jumlah enzim lipase yang
tidak terimobilisasi pada supernatan ditentukan dengan menggunakan
spektronik 20 D+ pada panjang gelombang 410 n m.
b. Penentuan kestabilan termal enzim lipase imobil
Uji kestabilan termal ditentukan dengan cara enzim lipase
bebas dipaparkan pada suhu optimum selama waktu tertentu, enzim
lipase bebas diuji aktivitas enzimatiknya pada kondisi optimumnya. Hal
yang sama dilakukan terhadap enzim lipase imobil (Cahyaningrum dkk.,
2008).
c. Penentuan kestabilan operasional enzim lipase imobil
Uji kestabilan lipase imobil ditentukan berdasarkan metode
Sigurgisladittor dkk (1993), sebagai berikut: sebanyak 3 mL campuran
reaksi yang mengandung substrat p-nitrofenilbutirat (berdasarkan
penentuan aktivitas enzim lipase) ditambahkan kedalam enzim imobil
kemudian di shaker selama 90 menit pada suhu 45 0C (suhu optimum
enzim lipase imobil). Selanjutnya disentrifugasi dengan kecepatan
1000 rpm selama 5 menit. Supernatan yang dihasilkan diuji aktivitas
enzimnya. Endapan dicuci dengan bufer borat pH 8,2, konsentrasi 0,2 M
selanjutnya digunakan untuk penentuan aktivitas enzim lipase berikutnya.
10. Penggunaan enzim lipase imobil untuk hidrolisis minyak kelapa menjadi 1,3-DAG

51
Penggunaan enzim lipase imobil dilakukan berdasarkan metode
(Blasi dkk., 2007) sebagai berikut:
a. Reaksi etanolisis
Seratus miligram minyak kelapa ditambahkan dengan 0,5 mL
etanol 96%. Selanjutnya ditambahkan enzim lipase imobil sebanyak
50 mg. Campuran reaksi disimpan selama satu malam dengan
menggunakan magnetik stirer pada suhu 45 0C kecepatan 100 rpm, lalu
disaring .Filtrat enzim yang diperoleh dicuci tiga kali dengan campuran
kloroform/metanol (1:1, v/v). Evaporasi pelarut, dialiri dengan gas
nitrogen.
b. Preparasi sampel hasil etanolisis
Sebanyak 10 mg sampel hasil etanolisis dilarutkan ke dalam 3 mL
n-heksan, kemudian ditambahkan 0,5 mL KOH metanolik 2 N, setelah
3 menit, ditambahkan akuades 3 mL. Fase organik pada lapisan atas
dikeringkan dengan Na2SO4, konsentrat yang dihasilkan dialirkan dengan
gas nitrogen. Hal yang sama dilakukan juga untuk standar. Sampel dan
standar siap untuk dianalisis dengan kromatografi gas yang dilengkapi
dengan detektor EID, integrator chromatopac C-R 6A dan kolom kapiler
HP-5 (30 m x 0,25 mm). Kondisi kromatografi gas : suhu injektor dan
detektor 250 0C, spliting injektor 70:1, suhu awal kolom 130 0C,
penambahan suhu kolom 3 0C / menit hingga mencapai suhu 250 0C,
dipertahankan selama 10 menit, dan gas pembawa He 1 mL/menit.

52
BAB IV
HASIL DAN PEMBAHASAN

53
1. Isolasi mikroba penghasil enzim lipase pada kopra berjamur
Hasil isolasi mikroba penghasil enzim lipase pada kopra berjamur,
menunjukkan bahwa pada pengenceran 10-1 terdapat satu macam warna
spora pada cawan petri yaitu putih kekuning-kuningan mulai hari pertama
sampai hari kelima. Setelah hari kelima warna spora berubah menjadi
hijau pucat kekuning-kuningan. Hasil identifikasi menunjukkan bahwa
mikroba tersebut adalah A. oryzae, dan sesuai dengan petunjuk
identifikasi pada literatur Identification of Pathogenic Fungi (Campbell
dkk., 1996) dan Pengenalan Kapang Tropik Umum (Ganjar dkk., 1999).
2. Produksi enzim lipase
Produksi enzim lipase dilakukan dengan menumbuhkan mikroba
(A. oryzae) dalam media inokulum. Selanjutnya media inokulum
ditumbuhkan dalam Erlenmeyer menggunakan shaker incubator selama
empat hari. Hari pertama dan kedua terjadi kekeruhan pada media dan
belum terlihat adanya butiran-butiran kecil, tetapi pada hari ketiga sudah
nampak ada butiran pada Erlenmeyer dan pada hari keempat butiran-
butiran berwarna putih kekuning-kuningan memenuhi wadah Erlenmeyer.
Hal ini menunjukkan bahwa media inokulum dapat digunakan untuk
memproduksi enzim lipase pada media produksi. Seperti halnya media
inokulum, media produksi dibuat dalam Erlenmeyer menggunakan
shaker incubator. Setiap hari dilakukan pengambilan sampel. Selanjutnya
disentrifugasi pada kecepatan 3500 rpm suhu 4 0C selama 30 menit, filtrat

54
yang diperoleh merupakan ekstrak kasar enzim lipase ekstraselluler.
Ekstrak kasar diuji aktivitas enzim dan kadar protein, massa sel ditentukan
berdasarkan berat sel kering.
.
1 2 3 4 5 6 7 8 9 10
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0
20
40
60
80
100
120
140
berat sel ker-ing
kadar protein
aktivi-tas enzim
Waktu fermentasi (hari)
Bera
t se
l ker
ing
(mg/
mL)
, Kad
ar
prot
ein
(mg/
mL)
Akti
vita
s en
zim
(U/m
L)
Gambar 12. Penentuan waktu fermentasi optimum terhadap aktivitas enzim lipase dari A. oryzae.
Dari uji aktivitas enzim lipase diperoleh bahwa enzim lipase
diproduksi maksimal pada hari kedelapan melalui proses fermentasi.
Grafik pada Gambar 12 menunjukkan bahwa aktivitas enzim lipase, kadar
protein dan berat sel kering meningkat seiring dengan bertambahnya
waktu fermentasi sampai pada hari kedelapan, setelah itu terjadi
penurunan aktivitas enzim lipase dan kadar protein. Hal ini disebabkan
karena terjadinya kerapatan sel dan ketersediaan nutrisi dalam media
produksi mulai berkurang, sehingga produksi enzim lipase menurun. Dari
x 103

55
berat sel kering diketahui bahwa produksi maksimal enzim lipase terjadi
pada akhir fase logaritmik (awal fase stasioner). Produksi maksimal enzim
lipase berhubungan dengan siklus pertumbuhan mikroorganisme
penghasil enzim lipase. Menurut Andersson (1980), aktivitas enzim lipase
selama pertumbuhan Pseudomonas flourescens mencapai maksimum
pada awal fase stasioner. Hasil penelitian Tripanji dkk (2008),
menunjukkan bahwa enzim lipase dari Neurospora sitophila diproduksi
maksimal pada hari ketiga.
3. Optimasi produksi enzim lipase
a. Variasi konsentrasi pepton
Pada Gambar 13 menunjukkan bahwa aktivitas relatif enzim lipase
tertinggi diperoleh pada konsentrasi pepton 1,0%, setelah itu menurun
sampai pada konsentrasi 1,5%. Penurunan aktivitas enzim lipase pada
konsentrasi pepton diatas 1,0%, karena pada media fermentasi terjadi
kelebihan pepton yang menyebabkan komposisi nutrien tidak seimbang
dalam pertumbuhan mikroba, sehingga produksi enzim lipase mengalami
penurunan. Menurut Suhartono (1989) komposisi nutrien yang seimbang
berpengaruh untuk pertumbuhan setiap mikroorganisme, jamur berfilamen
membutuhkan (10–25)% protein (pepton)/berat kering untuk
pertumbuhannya. Pepton adalah komponen pada media produksi
berfungsi sebagai sumber nitrogen, berperan dalam pembentukan

56
protoplasma dan dinding sel. Pepton diperlukan oleh mikroba (A. oryzae)
untuk pertumbuhannya dalam memproduksi enzim lipase.
0.5 0.8 1.0 1.3 1.50
20
40
60
80
100
120
Konsentrasi pepton (%)
Akt
ivit
as r
elat
if (
%)
Gambar 13. Pengaruh konsentrasi pepton terhadap aktivitas enzim lipase dari A. oryzae (suhu 37 0C; pH 7,0; [E] = 10%;
[S] = 0,1 M)
b. Variasi konsentrasi minyak zaitun
Pada Gambar 14 menunjukkan bahwa aktivitas relatif enzim lipase
tertinggi diperoleh pada konsentrasi minyak zaitun 3%, setelah itu aktivitas
enzim lipase menurun. Penurunan aktivitas enzim lipase pada konsentrasi
minyak zaitun diatas 3%, karena pada media produksi terjadi kelebihan
minyak zaitun yang menyebabkan komposisi nutrien tidak seimbang
dalam pertumbuhan mikroba, sehingga produksi enzim lipase mengalami
penurunan. Menurut Suhartono (1989) komposisi nutrien yang seimbang

57
berpengaruh untuk pertumbuhan setiap mikroorganisme, jamur berfilamen
membutuhkan (2–50)% lipida (minyak zaitun)/berat kering untuk
pertumbuhannya. Minyak zaitun adalah komponen pada media produksi
berfungsi sebagai sumber karbon untuk pembentukan sel dan sumber
energi, juga sebagai induser yaitu zat penginduksi pada sintesis enzim
lipase dari A. oryzae. Penambahan senyawa penginduksi pada
konsentrasi yang tepat akan membawa hasil yang maksimal pada enzim
yang terinduksi.
1 2 3 4 50
20
40
60
80
100
120
Konsentrasi minyak zaitun (%)
Akt
ivit
as r
elat
if (
%)
Gambar 14. Pengaruh konsentrasi minyak zaitun terhadap aktivitas enzim lipase dari A. oryzae (suhu 37 0C; pH 7,0; [E] = 10% [S] = 0,1 M)
c. Variasi kecepatan pengadukan
Pada Gambar 15 menunjukkan bahwa aktivitas enzim lipase
tertinggi pada kecepatan pengadukan 150 rpm. Kecepatan pengadukan

58
diatas 150 rpm, aktivitas enzim lipase mengalami penurunan cukup besar,
karena pada media fermentasi terdapat busa cukup banyak. Busa dapat
memberikan efek buruk, karena gelembung udara terkurung dalam busa.
Keberadaan busa pada media fermentasi, menambah waktu tinggal
gelembung udara, sehingga terjadi kekurangan oksigen (Tarigan, 1991).
Penurunan oksigen terlarut menyebabkan penurunan dalam laju
pertumbuhan sel-sel di dalam media fermentasi, sehingga produksi enzim
juga menurun (Praweda, 2004).
50 100 150 200 2500
20
40
60
80
100
120
Akt
ivit
as r
elat
if (
%)
Kecepatan pengadukan (rpm)
Gambar 15. Pengaruh kecepatan pengadukan terhadap aktivitas enzim lipase dari A. oryzae (suhu 37 0C; pH 7,0; [E] = 10%;
[S] = 0,1 M) Menurut Chander dkk (1980), aktivitas enzim lipase pada media
produksi yang teraduk 50% lebih tinggi dibandingkan dengan tidak
teraduk. Kecepatan pengadukan berfungsi untuk mendispersi udara di

59
dalam larutan nutrien dan menyeragamkan suhu serta konsentrasi
nutrien di dalam proses fermentasi. Selama proses fermentasi
berlangsung, diperlukan pengocokan (pengadukan) terus menerus agar
konsumsi oksigen selalu ada. Oksigen adalah gas yang sedikit larut dalam
air (10 mg/liter), sehingga perlu ditransfer terus-menerus ke dalam media
fermentasi.
Optimasi produksi enzim lipase sebelum variasi komposisi media
produksi dan kecepatan pengadukan dapat dilihat pada Tabel 2, 3 dan 4.
Tabel 2. Aktivitas enzim lipase dari A. oryzae sebelum variasi komposisi media produksi dan kecepatan pengadukan (suhu 37 0C; pH 7,0; [S] = 0,1 M; [E] = 10%)
V. media Kom. media Kecepatan Serapan Aktivitas produksi produksi (%) pengadukan (410 n m) (U/mL) (mL) (rpm)
Pepton 0,5 100 Minyak zaitun 1 150 0,126 13.222
Tabel 3. Aktivitas enzim lipase dari A. oryzae setelah variasi komposisi media produksi dan kecepatan pengadukan (suhu 37 0C;
pH 7,0; [S = 0,1 M]; [E = 10%]

60
V. media Kom. media Kecepatan Serapan Aktivitas produksi produksi (%) pengadukan (410 n m) (U/mL) (mL) (rpm)
100 Pepton 1 150 0,140 14.777 Minyak zaitun 3
Tabel 4. Aktivitas enzim lipase dari Aspergillus oryzae setelah variasi komposisi media produksi dan kecepatan pengadukan pada kondisi optimum (suhu 35 0C; pH 8,2; [S] = 0,2 M; [E] = 45%)
V. media Kom. media Kecepatan Serapan Aktivitas produksi produksi (%) pengadukan (410 n m) (U/mL) (mL) (rpm)
100 Pepton 1 150 0,230 18.888 Minyak zaitun 3
Pada Tabel 2, 3 dan 4 menunjukkan bahwa komposisi media
produksi dan kecepatan pengadukan pada kondisi optimum dapat
meningkatkan aktivitas enzim lipase dari 13.222 U/mL menjadi 18.888
U/mL .
4. Pemurnian enzim lipase
a. Fraksinasi dengan amonium sulfat

61
Ekstrak kasar enzim lipase (supernatan) diendapkan dengan
menambahkan garam amonium sulfat pada berbagai tingkat kejenuhan
(0-100)%. Penambahan garam amonium sulfat dilakukan sedikit demi
sedikit pada suhu rendah (5 0C) sambil diaduk dengan magnetik stirer
selama satu malam. Hasil pengendapan protein dengan amonium sulfat
terlihat pada Tabel 5.
Tabel 5. Fraksinasi enzim lipase menggunakan amonium sulfat
No. Fraksi (%) Volume tiap fraksi (mL)
Jumlah amonium sulfat
(gr)
Aktivitas enzim (U/mL)
Kadar protein (mg/mL)
1
2
3
4
5
0 – 20
20 – 40
40 – 60
60 – 80
80 - 100
500
514
520
544
572
53,000
58,082
62,400
70,176
79,510
11.691
13.914
15.067
26.511
7.975
17.994
24.679
49.120
57.176
5.738
Pada Tabel 5 menunjukkan bahwa fraksinasi amonium sulfat
tertinggi diperoleh pada tingkat kejenuhan ( 60- 80)% , dengan aktivitas
enzim tertinggi sebesar 26.511 U/mL dan kadar protein 57.176 mg/mL.
Amonium sulfat pada konsentrasi tinggi akan mengendapkan komponen
protein (termasuk enzim) sehingga diperoleh enzim bebas dari komponen
non protein. Amonium sulfat banyak digunakan untuk mengendapkan
protein karena kelarutannya tinggi, harga murah, dan umumnya tidak

62
mempengaruhi struktur protein (Suhartono, 1989). Shaoxin Chen (2007),
memperoleh fraksinasi amonium sulfat pada tingkat kejenuhan antara
(50-70)% terhadap enzim lipase dari Bacillus cereus C71. Enzim kasar
yang diperoleh pada tingkat kejenuhan amonium sulfat antara (60-80)%
didialisis dengan bufer borat selama semalam pada suhu 4 0C. Enzim
yang diperoleh pada tahap ini adalah enzim semi murni.
b. Kromatografi kolom penukar ion
Enzim semi murni (hasil pengendapan amonium sulfat dan
dialisis) dimurnikan lebih lanjut dengan teknik kromatografi kolom penukar
ion, hasil kromatogram pada Gambar 16.
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 850
0.02
0.04
0.06
0.08
0.1
0.12
0
5000
10000
15000
20000
25000
Absorban 280 nm
Gambar 16. Kromatogram hasil pemurnian enzim lipase dari A. oryzae pada kromatografi kolom penukar ion Q sepharosa FF. volume enzim 3 mL, kec. alir 30 tetes/menit dan suhu 16 0C.
Pada Gambar 16 menunjukkan bahwa dari hasil pemurnian
kromatografi kolom penukar ion, terdapat puncak- puncak protein dan
tiga puncak aktivitas enzim lipase. Aktivitas enzim lipase tertinggi
Nomor fraksi
S
erap
an p
rote
in (
280
n
m)
Ak
tivi
tas
enzi
m (
U/m
L)Serapan Aktivitas

63
terdapat pada fraksi 74-77, dengan aktivitas enzim masing-masing fraksi
74 = 19.777 U/mL, fraksi 75 = 20.111 U/mL, fraksi 76 = 21.777 U/mL, dan
fraksi 77= 20.222 U/mL. Fraksi ini disatukan sehingga diperoleh
sebanyak 12 mL kemudian dimasukkan dalam kantong selofan,
dipekatkan dengan kristal polietilen-glikol diperoleh volume 6 mL dengan
aktivitas spesifik sebesar 27,50 U/mg protein. Dari hasil pemurnian dapat
disimpulkan bahwa proses pemurnian dengan kromatografi kolom
penukar ion dapat meningkatkan kemurnian enzim lipase sebesar 12,85
kali dibandingkan dengan fraksi enzim ekstrak kasarnya
c. Kromatografi kolom filtrasi gel
Hasil pemekatan yang diperoleh dari kromatografi kolom penukar
ion, dimurnikan lebih lanjut dengan kromatografi kolom filtrasi gel.
Kromatogram hasil pemurnian enzim lipase pada Gambar 17,
menunjukkan bahwa pemurnian dengan kromatografi kolom filtrasi gel
pada matriks sephadex G-75 menghasilkan puncak-puncak protein dan
satu puncak aktivitas enzim lipase tertinggi, yaitu fraksi 39 dengan
aktivitas spesifik sebesar 43,76 U/mg protein. Dengan demikian pada
tahap pemurnian ini, dapat meningkatkan kemurnian enzim lipase sebesar
20,25 kali dibandingkan dengan fraksi enzim ekstrak kasarnya. Tahapan
pemurnian enzim lipase ditunjukkan pada Tabel 5.

64
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 900
0.01
0.02
0.03
0.04
0.05
0.06
0.07
0.08
0
5000
10000
15000
20000
25000Absorban 280 nm
Gambar 17. Kromatogram hasil pemurnian enzim lipase dari A. oryzae pada kromatografi kolom filtrasi gel sephadex G-75, volume enzim 3 mL, kec. alir 6 tetes/menit dan suhu 16 0C.
Tabel 5. Tahap-tahap proses pemurnian enzim lipase dari A. oryzae
Tahap-tahap Volume Protein Aktivitas Enzim Recovery FP Pemurnian (mL) total (%) (X) (mg) Total Spesifik
(unit) (U/mg prot)
Ekstrak 1820 33.670 72.800 2,16 100 1 kasar
(NH4)2SO4 10 5.559 43.753 7,87 60,10 3,64 60-80%
Q Sepharosa 6 399,75 11.022 27,50 15,14 12,85 FF
Sephadex 3 137,37 6.012 43,76 8,25 20,25 G-75
Keterangan: FP (Faktor/tingkat pemurnian)
Nomor fraksi
S
erap
an p
rote
in (
280
n m
)
A
kti
vita
s en
zim
(U
/mL
)
Serapan Aktivitas

65
Beberapa hasil penelitian mengenai tingkat kemurnian enzim
lipase dari mikroba, yaitu Yarrowia lipolitica, Bacillus licheniformis B42
masing-masing dengan tingkat kemurnian 26,50 dan 27,33 kali
dibandingkan dengan fraksi enzim ekstrak kasarnya (Mingrui Yu dkk.,
2007) dan (Bayoumi dkk., 2007).
d. Uji kemurnian dan penentuan berat molekul enzim lipase
Kemurnian enzim lipase dan penentuan berat molekul diuji
dengan elektroforesis SDS-PAGE, diperoleh hasil seperti ditunjukkan
pada Gambar 18. Fraksi enzim ekstrak kasar (kolom 2) dan fraksi
amonium sulfat (kolom 3), kemurnian enzim lipase relatif rendah ditandai
dengan munculnya beberapa pita pada gel elektroforesis. Pada fraksi
kromatografi kolom matriks Q sepharosa FF(kolom 4) terlihat tiga pita
yang muncul sedangkan fraksi kromatografi kolom matriks sephadex G-75
(kolom 5) hanya terlihat dua pita, sehingga dapat disimpulkan bahwa
tahap pemurnian dengan kromatografi kolom matriks sephadex G-75
menghasilkan enzim lipase dengan kemurnian relatif tinggi dibandingkan
dengan enzim ekstrak kasar, pengendapan amonium sulfat dan
kromatografi kolom matriks Q sepharosa FF. Berdasarkan marker (protein
standar) pada Gambar 18 dan Tabel penentuan berat molekul pada
Lampiran 17 diperoleh berat molekul enzim lipase sebesar 40,7 kDa.
Adapun pita yang muncul pada penunjuk berat molekul 19,6 kDa, itu
karena terjadi denaturasi pada molekul protein. Berat molekul enzim
lipase dari beberapa mikroba yaitu, Mucor sp 42 kDa (Abbas dkk.,

66
2002), Bacillus cereus C71 42 kDa (Shaoxin Chen dkk., 2007), dan
Yarrowia lipolytica 38 kDa (Mingrui Yu dkk., 2007).
Gambar 18. Elektroforegram hasil elektroforesis gel SDS-PAGE 10% hasil pemurnian enzim lipase dari A. oryzae
Keterangan: Kolom 1 protein standar : Posforilase-b (116 kDa), BSA
(66,2 kDa), ovalbumin (45 kDa), karbonik anhidrase (35 kDa), rease BSP 981 (25 kDa), β-laktoglobulin (18,4 kDa), lisozim (14,4 kDa); kolom 2, ekstrak kasar enzim lipase; kolom 3, fraksi amonium sulfat (60-80)%; kolom 4, hasil pemurnian dengan matriks Q sepharosa FF; kolom 5, hasil pemurnian dengan matriks sephadex G-75.
116
66,2
45
35
25
18,4 14,4
1 2 3 4 5
40,7
19,6
kDakDa

67
5. Karakterisasi enzim lipase
a. Penentuan suhu optimum
Pada Gambar 19 menunjukkan bahwa aktivitas enzim lipase
mengalami peningkatan sampai pada suhu 35 0C (suhu optimum) dengan
aktivitas relatif 100%. Setelah melewati suhu optimum, terjadi penururan
aktivitas enzim pada suhu 40 0C dengan aktivitas relatif 61,20%. Kenaikan
aktivitas enzim di bawah suhu optimum disebabkan karena kenaikan
energi kinetika molekul-molekul yang bereaksi. Akan tetapi apabila suhu
tetap dinaikkan terus, energi kinetika molekul-molekul enzim menjadi
besar sehingga memecahkan ikatan-ikatan sekunder yang
mempertahankan enzim dalam bentuk aslinya, akibatnya struktur
sekunder dan tersier hilang sehingga aktivitas enzim menurun.
20 25 30 35 40 45 500
20
40
60
80
100
120
Akt
ivit
as r
elat
if (
%)
Gambar 19. Pengaruh suhu terhadap aktivitas enzim lipase dari A. oryzae [S] = 0,2 M; [E] = 45%; pH 7,0
Suhu (0C)

68
Adanya perubahan suhu juga akan mempengaruhi ikatan hidrogen
atau interaksi hidrofobik yang berperanan dalam menjaga konformasi
molekul enzim. Perubahan konformasi akan mempengaruhi sisi aktif dari
enzim, kondisi panas tertentu menyebabkan ikatan hidrogen tersebut akan
putus. Putusnya satu ikatan hidrogen akan menyebabkan mudahnya
pemutusan ikatan hidrogen selanjutnya dalam rantai polipeptida tersebut,
sehingga protein enzim mengalami denaturasi (Whitaker, 1972).
Penelitian yang dilakukan oleh Mingrui Yu ( 2007) terhadap enzim lipase
dari Yarrowia lipolytica diperoleh suhu optimum 40 0C.
b. Penentuan pH optimum
Variasi pH dilakukan pada suhu optimum. Kondisi pH berpengaruh
terhadap aktivitas enzim. Pada Gambar 20 menunjukkan bahwa aktivitas
enzim lipase mengalami peningkatan sampai pada pH 8,2 (pH optimum)
dengan aktivitas relatif sebesar 100%, kemudian mulai menurun pada pH
8,4 dengan aktivitas relatif 87,79%. Untuk enzim, perubahan muatan
dapat mempengaruhi aktivitas, baik dengan perubahan struktur maupun
dengan perubahan muatan pada residu asam amino yang berfungsi
mengikat substrat. Misalkan suatu enzim bermuatan negatif bereaksi
dengan suatu substrat bermuatan positif membentuk kompleks enzim-
substrat, kemudian pada nilai pH rendah enzim akan diprotonasi dan
kehilangan muatan negatifnya. Hal yang sama pada pH tinggi substrat
akan terionisasi dan kehilangan muatan positifnya (Murray dkk., 2009).
Perubahan ion H+ yang ada dalam larutan enzim memberikan efek pada

69
bagian katalitik dan konformasi enzim. Kondisi pH terlalu rendah atau
terlalu tinggi menyebabkan terjadinya perubahan konformasi enzim,
sehingga aktivitas enzim menurun (Gambar 20). Penelitian yang dilakukan
oleh Mingrui Yu dkk (2007) terhadap enzim lipase dari Yarrowia lipolytica
diperoleh pH optimum 8,0.
7.0 7.2 7.4 7.6 7.8 8.0 8.2 8.4 8.6 8.8 9.00
20
40
60
80
100
120
Akt
ivit
as r
elat
if (
%)
pH
Gambar 20. Pengaruh pH terhadap aktivitas enzim lipase dari A. oryzae [S] = 0,2 M ; [E] = 45%; suhu 35 0C
c. Penentuan Km dan Vmaks
Karakter utama yang ditentukan dalam mempelajari sifat kinetik
enzim adalah kecepatan katalitik maksimum (Vmaks) dan konsentrasi
substrat pada saat kecepatan katalitik mencapai setengah maksimum
(Km). Uji aktivitas enzim lipase menggunakan substrat p-nitrofenilbutirat
pada interval (0-0,05) M dan menggunakan p-nitrofenol sebagai standar.
Konstanta Vmaks dan Km ditentukan dengan metode Lineweaver-Burk.

70
Grafik hubungan antara (1/v) sebagai sumbu Y terhadap (1/S) sebagai
sumbu X diperlihatkan pada Gambar 21. Selanjutnya data-data yang
diperoleh dibuat regresi liniernya dan diperoleh persamaan garis linier,
yaitu: Y = 0,024 X + 0,519. Lereng regresi linier dimasukkan kedalam
persamaan Lineweaver-Burk 1/V = (Km/Vmaks)(1/S) +( 1/V maks), maka akan
diperoleh nilai Km = 0,046 M dan Vmaks adalah 1,926 µmol mL-1 menit-1 .
Samsumaharto (2007) memperoleh nilai Km = 0,011 M dan nilai Vmaks =
11,63 µmol menit-1 terhadap substrat triolein hasil katalisis enzim lipase
yang diisolasi dari Theobroma cacao. L. Nilai Km yang tinggi
menunjukkan afinitas terhadap substrat yang rendah. Semakin kecil nilai
Km semakin tinggi afinitasnya terhadap substrat, sehingga semakin
rendah konsentrasi substrat yang dibutuhkan untuk mencapai kecepatan
reaksi katalitik maksimumnya (Vmaks).
-25 -20 -15 -10 -5 0 5 10 15 20 250
0.2
0.4
0.6
0.8
1
1.2
f(x) = 0.0244060024811527 x + 0.519582307472092
1/[s] M
1/v
(um
ol/m
enit
Gambar 21. Kurva Lineweaver-Burk hubungan antara 1/substrat dan 1/kecepatan katalitik
-1/Km
1/Vmaks

71
6. Imobilisasi enzim lipase
a. Penentuan waktu imobilisasi
Waktu imobilisasi ditentukan dengan memvariasikan 8 waktu yang
berbeda untuk proses imobilisasi. Pada Gambar 22 menunjukkan bahwa
waktu mempunyai pengaruh terhadap proses imobilisasi enzim lipase
pada matriks silika gel.
0 15 30 45 60 75 90 105 1200
102030405060708090
Waktu imobilisasi (menit)
Jum
lah
lipas
e te
rimob
ilisa
si (%
)
Gambar 22. Penentuan waktu imobilisasi optimum enzim lipase murni dari A. oryzae
Pada waktu 15-90 menit jumlah enzim lipase terimobil bertambah
seiring kenaikan waktu imobilisasi. Setelah 90 menit jumlah enzim lipase
yang terimobil relatif konstan atau terjadi kenaikan tidak signifikan sampai
waktu 120 menit. Pada keadaan tersebut dapat dikatakan bahwa mulai
menit ke-90 situs aktif matriks telah jenuh oleh enzim lipase dan mencapai
kesetimbangan imobilisasi. Wulan dkk (2008) memperoleh waktu

72
imobilisasi maksimal 3 jam terhadap enzim lipase dari Rhyzopus oryzae
menggunakan kitin sebagai adsorben.
b. Karakterisasi enzim lipase imobil
Karakterisasi enzim lipase imobil meliputi penentuan pH dan suhu
optimum. Pada Gambar 23 menunjukkan bahwa aktivitas enzim lipase
bebas pada pH 7,0 rendah, kemudian terjadi peningkatan sampai pH 8,2.
Pada pH 8,2, aktivitas enzim lipase bebas memberikan aktivitas
maksimum, dimana pada kondisi ini aktivitas relatif enzim lipase bebas
mencapai 100%. Pada pH 8,4 aktivitas enzim lipase mulai menurun
sampai pada pH 9,0. Hal yang sama terjadi pada enzim lipase imobil,
sehingga baik enzim lipase bebas maupun imobil mempunyai pH optimum
sama yaitu 8,2. Aktivitas enzim lipase bebas sedikit lebih besar dari
imobil, ini disebabkan karena enzim lipase imobil terikat pada matriks
silika gel sehingga kecepatan untuk kontak dengan substrat lebih kecil.
Penelitian Dosanjh dan Kaur (2002) menunjukkan bahwa enzim
lipase imobil pada matriks HP-20, pH optimum 8,0 dan suhu optimum
54 0C. Enzim lipase imobil dari Rhizopus oryzae pada matriks kitin pH
optimum 7,0 dan suhu optimum 37 0C (Wulan dkk., 2008).

73
7 7.2 7.4 7.6 7.8 8 8.2 8.4 8.6 8.8 90
20
40
60
80
100
120
Enzim lipase murni bebas enzim lipase imobil
pH
Aktiv
itas r
elati
f (%
)
Gambar 23. pH optimum enzim lipase bebas dan imobil dari A. oryzae (suhu 35 0C; [S]= 0,2 M; dan [E]= 45%)
Pada Gambar 24 menunjukkan bahwa aktivitas enzim lipase
bebas mengalami kenaikan seiring dengan kenaikan suhu dari
(20 – 55 0C). Pada suhu diatas 35 0C aktivitas enzim lipase bebas
mengalami penurunan. Aktivitas tertinggi terjadi pada suhu 35 0C dengan
aktivitas relatif 100%. Peningkatan suhu sampai pada suhu optimum akan
meningkatkan laju reaksi enzimatik, tetapi peningkatan suhu diatas suhu
optimum akan menurunkan laju reaksi enzimatik. Uji aktivitas enzim lipase
imobil mempunyai suhu optimum 45 0C, sedangkan enzim lipase bebas
35 0C. Hal tersebut menunjukkan bahwa matriks silika gel mampu
melindungi enzim lipase imobil terhadap panas sehingga enzim lipase
imobil mampu bertahan pada suhu yang lebih tinggi dibandingkan dengan
enzim lipase bebas.
Enzim lipase bebas Enzim lipase imobil

74
20 25 30 35 40 45 50 550
20
40
60
80
100
120
enzim lipase murni bebas enzim lipase imobil
Suhu (˚C)
Aktiv
itas r
elati
f (%
)
Gambar 24. Suhu optimum enzim lipase bebas dan imobil dari A.oryzae (pH = 8,2; [S] = 0,2 M; dan [E] = 45%)
c. Kestabilan termal enzim lipase imobil
Hasil uji kestabilan termal enzim lipase bebas dan imobil
ditunjukkan pada Gambar 25. Enzim lipase bebas pada suhu optimumnya
yaitu 35 0C hanya dapat bertahan sampai dengan pemanasan selama 60
menit dengan aktivitas relatif tinggal 20%. Penurunan aktivitas mulai
terjadi pada pemanasan selama 40 menit sampai 100 menit dimana
aktivitas relatif tinggal 5%. Enzim lipase imobil pada suhu optimumnya
45 0C mampu bertahan dengan pemanasan selama 80 menit dan jumlah
aktivitas yang tersisa relatif konstan sampai pemanasan 120 menit.
Penurunan aktivitas mulai terjadi pada waktu inkubasi 40 menit sampai 80
menit dimana aktivitas relatif tinggal 38%. Bila dibandingkan dengan
enzim lipase bebas maka enzim lipase imobil lebih baik stabilitas
termalnya.
Suhu (0C)Enzim lipase bebas Enzim lipase imobil

75
20 40 60 80 100 1200
20
40
60
80
100
120
enzim lipase murni bebas enzim lipase imobil
Gambar 25. Kestabilan termal enzim lipase bebas (suhu 35 0C) dan enzim lipase imobil (suhu 45 0C) dari A. oryzae pH = 8,2; [S] = 0,2 M; dan [E] = 45%
d. Kestabilan operasional enzim lipase imobil
Penggunaan berulang enzim lipase imobil pada adsorben silika
gel, terdapat penurunan aktivitas enzim lipase seperti yang terlihat pada
Gambar 26.
Ak
tivi
tas
rela
tif
(%)
Waktu pemaparan (menit)Enzim lipase bebas Enzim lipase imobil

76
1 2 3 4 5 60
20
40
60
80
100
120
Gambar 26. Penggunaan berulang enzim lipase imobil dari A. oryzae (suhu 45 0C, pH = 8,2; [S] = 0,2 M; dan [E] = 45%)
Pada Gambar 26 menunjukkan bahwa enzim lipase imobil dapat
digunakan berulangkali . Semakin banyak penggunaan berulang aktivitas
enzim lipase semakin menurun. Enzim lipase imobil mampu digunakan
sebanyak enam kali pemakaian berulang, dan pada penggunaan keenam
kali, aktivitas katalitiknya masih sekitar 37,50%. Enzim lipase pancreatin
imobil pada matriks karbon aktif efektif lima kali penggunaan berulang,
(Silva dkk., 2007), enzim lipase imobil dari Rhyzopus oryzae pada matriks
kitin efektif tiga kali penggunaan berulang (Wulan dkk., 2008). Enzim
lipase bebas hanya dapat digunakan satu kali, karena enzim lipase bebas
akan tercampur dengan produk reaksi sehingga harus dilakukan proses
perusakan untuk memisahkan enzim lipase dari produk reaksi. Pada
aplikasinya hal tersebut sangat tidak ekonomis mengingat enzim lipase
mahal harganya. Penurunan aktivitas setelah penggunaan berulang
Jumlah penggunaan berulang (waktu)
Ak
tivi
tas
rela
tif
(%)

77
disebabkan karena lemahnya ikatan antara enzim lipase dengan adsorben
pendukung, karena hanya di dukung oleh ikatan Van der Waals, ikatan
hidrogen dan interaksi hidrofobik (Suhartono, 1989). Jika terdapat
gangguan seperti halnya penggunaan berulang, ikatan ini dapat rusak
menyebabkan enzim terlepas dari adsorben.
7. Penggunaan enzim lipase imobil untuk hidrolisis minyak kelapa menjadi 1,3-diasilgliserol (1,3-DAG)
Penggunaan enzim lipase imobil hasil isolasi dari A. oryzae pada
kopra berjamur untuk hidrolisis minyak kelapa menjadi 1,3-DAG, dilakukan
analisis kromatografi gas. Melalui uji kromatografi gas dapat diketahui
persentase senyawa yang terkandung di dalam larutan sampel hasil
reaksi. Dari analisis kromatografi gas akan terlihat peak yang merupakan
identitas dari senyawa tertentu. Hasil analisis sampel tersebut diketahui
bahwa produk hasil reaksi menghasilkan 1,3-DAG sebesar 26,49%
dengan waktu retensi 21,99 seperti yang ditunjukkan pada Gambar 27,
dan sebagai pembanding digunakan kromatogram standar 1,3-DAG
dengan waktu retensi 22,04 pada Gambar 28. Penelitian Blasi dkk (2007)
terhadap enzim lipase dari Rhizomucor miehei pada substrat virgin olive
oil spesifik menghasilkan 1,3-DAG. Bornscheuer (1995) meneliti enzim
lipase dari Rhizomucor delemar spesifik menghasilkan monoasilgliserol
(MAG). Untuk lebih meyakinkan bahwa enzim lipase hasil isolasi
menghasilkan 1,3-DAG, maka dilakukan pula analisis terhadap sampel
minyak kelapa tanpa penambahan enzim lipase, hasilnya dapat
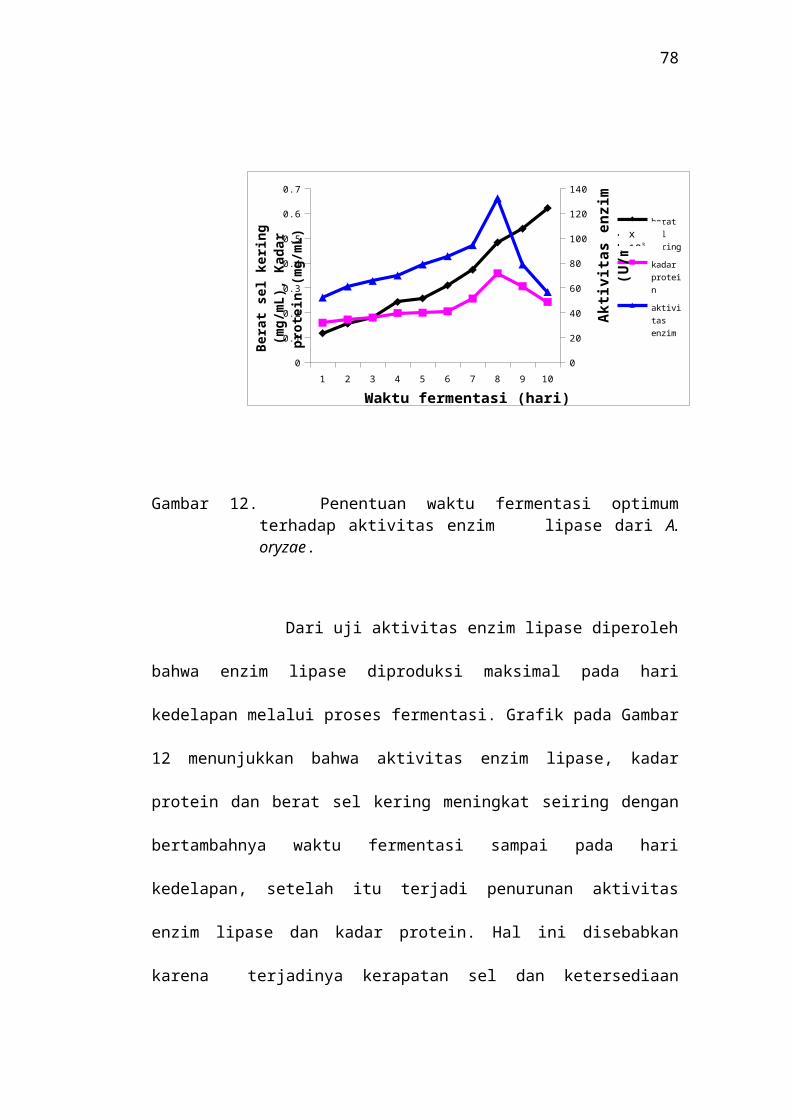
78
dijelaskan bahwa sampel minyak kelapa yang digunakan tidak
mengandung 1,3-DAG, seperti yang terlihat pada Gambar 29.
Waktu retensi (menit)
Gambar 27. Kromatogram hasil katalisis enzim lipase imobil dari Aspergillus oryzae pada minyak kelapa
Waktu retensi (menit)
Gambar 28. Kromatogram standar dari campuran 1,3-DAG dengan MAG
MAG
1,3-DAG

79
Waktu retensi (menit)
Gambar 29. Kromatogram minyak kelapa tanpa penambahan enzim lipase
Gambar 30. Kromatogram hasil katalisis oleh enzim lipase imobil komersial pada minyak kelapa
Waktu retensi (menit)

80
Pada kromatogram minyak kelapa hasil katalisis enzim lipase hasil
isolasi (Gambar 29) dan enzim lipase komersial (Gambar 30), terlihat
banyak muncul peak-peak selain 1,3-DAG, ini disebabkan karena
banyaknya senyawa yang terdapat pada minyak kelapa. Minyak kelapa
mengandung 90% asam lemak jenuh, dan dari 79 jenis triasilgliserol
penyusunnya, 84% tersusun atas tiga asam lemak jenuh. Menurut
Sontag (1979), triasilgliserol yang banyak menyusun minyak kelapa
antara lain lauromiristopalmitin, trilaurin, dilauromiristin, kaprolauromiristin
dan kaprodilaurin. Hasil penelitian Soeka dkk (2008) menunjukkan bahwa
kandungan asam lemak minyak kelapa hasil fermentasi: asam laurat
46,70%; asam stearat 19,73%; asam meristat 13,94% dan palmitat
5,97%.
Waktu retensi (menit)

81
BAB VI
KESIMPULAN DAN SARAN
A. Kesimpulan
1. Kondisi optimum produksi enzim lipase dari A. oryzae pada kopra
berjamur, waktu fermentasi 8 hari, konsentrasi pepton 1%, konsentrasi
minyak zaitun 3% dan kecepatan pengadukan 150 rpm.
2. Tingkat kemurnian enzim lipase dari A. oryzae 20,25 kali terhadap
ekstrak kasar, dengan karakteristik suhu optimum 35 0C, pH optimum
8,2, Km 0,046 M, dan Vmaks 1,926 µmol/mL.menit serta berat
molekul 40,7 kDa.
3. Enzim lipase imobil dari A. oryzae dapat digunakan sebagai
biokatalisator yang efektif dan efisien 6 kali pemakaian berulang.
4. Enzim lipase imobil dari A. oryzae dapat menghidrolisis minyak
kelapa menjadi 1,3–DAG.

82
B. Saran
1. Perlu dilakukan penelitian lebih lanjut terhadap sifat fisik dan
karakteristik dari 1,3- DAG yang dihasilkan dari minyak kelapa.
2. Aplikasi 1,3-DAG yang dihasilkan sebagai senyawa pengemulsi untuk
produk makanan dan kosmetik.

83
DAFTAR PUSTAKA
Abbas, H. Hiol, A. Deyris, V. and Comeau, L. 2002. Isolation and Characterization of an Extracellular Lipase from Mucor sp Strain Isolated from Palm Fruit. Enzyme and Microbial Technology. 31: 968-975
Achmad, S.A. 2007. Keanekaragaman Hayati dalam Pembelajaran Ilmu Kimia. Makalah Disajikan Dalam Pertemuan Peran Pendidikan Kimia Dalam Era Globalisasi.
Anderssonn, R.E. 1980. Lipase Production, lipolysis and Formation of Volatile Compounds by Pseudomonas flourescens in Fat Containing Media. J. Food. Sci. 45:1694-1701
Anonim. 2007. =com Cara Pembuatan Kopra. (Online), (Ic.bppt.go.id/iptek/index.php?option content&task=view&id=22&itemid=26&limit=1&li….59k, diakses 1 januari 2007).
Anonim. 2010. Mikroorganisme.(Online), (id.wikipedia.org.wiki, diakses 4 Juli 2010).
Bayoumi, R.A., El-Louboudey, S.S., Sidkey, N.M., and Rahman, A.E.l. M.A. 2007. Production, Purification and Characterization of Thermoalkalophilic Lipase for Application in Bio-detergent Industry. J.Applied Sciences Research. 3(12) 1752-1765.
Blasi, F. Cossignani, L. Simonetti, M.S., and Damiani, P. 2007. Biocatalysed Synthesis of sn-1,3- Diacylglycerol Oil from Extra Virgin Olive Oil. Enzim and Mikrobial Technology. 41: 727-732
Bollag, D.M., and Edelstein, S.J. 1991. Protein Methodes, 2nd. John Wiley & Sons, Inc., New York.
Bornscheuer, U.T. 1995. Lipase-Catalyzed Synthesis of Monoacylglycerols. Enzyme and Microbial Technology. 17: 578-586
Brown, K. 1997. (http://www.tracy.k12.ca.us/thsadvbio/images/Hydrop.gif, diakses 28 Mei 2008)

84
Cahyaningrum, S.E., Narsito, Santoso, S. J., and Agustini, R. 2008. Immobilization of Papain on Chitosan. Indo. J. Chem. 8 (3) 372-376.
Campbell, C.K., Johson, C.M., Philpot, D.W., and Warnock. 1996. Identification of Pathogenic Fungi. Public Health Laboratory Service, London.
Chander, H. V.K., Batish, S.S., Sannabhadti, and Srinivasan, R.A. 1980. Factors Affecting Lipase Production in Aspergillus wentii. J. Food Sci. 45 : 598-600
Colowick, S.P., and Kaplan, N.O. 1957. Methods in Enzymology. Academic Press. Inc. New York, 3.107
Comeau, L.D., Druet, M.D., Jonso, and Hiol, A. 1999. Producing, Purification and Characterization of an Extracellular Lipase from Mucor hiemalis. Enzyme. Microb. Technol. 25 : 80-87.
Cottrell, M.T., Moore, J.A., and Kircman, D.L. 1999. Chitinase from Unculture Marine Microorganism. Appl. and Enviromental Microbiology. 65 (6) 2553-2557.
Dali, S. dan Pirman. 2005. Eksplorasi dan Isolasi Enzim Lipase dari Fungi Inperfekti (genus Aspergillus dan Penicillium) Indigenus. Lembaga Penelitian UNHAS
Darwis, A.A., dan Sukara, E. 1990. Isolasi, Purifikasi, Karakterisasi Enzim. PAU Bioteknologi IPB Bogor.
Davidson, W.S., Gupta, R. Saxena, R.K., and Yadav, R.P. 1998. Purification and Characterization of Regiospecific Lipase from A. terreus. Biotehnol. Appl. Biochem. 28 : 243-249.
Davis, J. Vaughan, D.H., and Cordasi, M.F. 1995. Element of Biosensor Construction. Enzyme and Microbial Technology. 17 : 1030-1035.
Dosanjh, N.S., and Kaur, J. 2002. Immobilization, Stabillity and Esterification Studies of Lipase from a Bacillus sp. Biotechnol. Appl. Biochem. 36: 7-12
Englard, S. and Seiffer, S. 1990. Precipitation Techniques In Guide to Protein Purification. (Deutscher., M.P. eds), Academic Press, Inc., San Diego. p. 285-300.

85
Gandjar, I. Samson, R. A., Tweel-Vermeulen, K. Van den, Utari, A. dan Santoso I. 1999. Pengenalan Kapang Tropik Umum. Yayasan Obor Indonesia, Jakarta.
Garcia, H.S., Yang, B. Parkin, K.L. 1996. Continuous Reactor for Enzymatic Glycerolysis of Butter Oil in The Absence of Solvent. Food Research International. 8(6) 605-609
Gupta, R. Gupta, N. and Rathi, P. 2004. Bacterial Lipases: an Overview of Production, Purification and Biochemical Properties. Appl. Microbial Biotechnol. 64: 263-781.
Hasanuddin, A. 2001. Kajian Teknologi Pengolahan Minyak Sawit Mentah untuk Produksi Emusifier Mono-Diasilgliserol dan Konsentrat Karotenoid, (Online), (http//tumoutou.net/3_sem1-012/asriani_h.htm, diakses 8 April 2008).
Helisto, P. and Korpela, T. 1998. Effects of Detergents on Activity, of Microbial Lipase as Measured by The Nitrophenyl Alkanoate Esters Method. Enzyme and Microbial Technology. 23: 113-117.
Heny, K.S. 2010. Fauna Indonesia. (Online), (id.wikipedia.org/wiki/fauna, diakses 4 Juli 2010).
Houby. 2010. (http:// www.vscht.cz/kch/galeric/fungi.htm,diakses 4 juni 2010).
Judoamidjoyo, M. Darwis, A.A., dan Sa’id, E.G. 1992. Teknologi Fermentasi. Diterbitkan Atas Kerja Sama dengan PAU – Bioteknologi Institut Pertanian Bogor.
Kao Corporation. 2004. General Properties and Cooking Characteristic of Diacylglycerol as an Adible Oil in Y. Katsuragi (Ed). Diacylglycerol Oil. Chapter 19-22, AOCS Press.P. 197-252
Kaur, J. Dosanjh, N.S., and Nawawi, N. 1998. A Novel Thermostable Lipase from a Thermophilic Bacillus sp . Characterization and Esterification Studies. Biotechnol. Lett. 20. 10: 997-1000.
Kermasha, S.C., Schvepp, M.C., and Michalski, A. Morin. 1997. Production, Partial Purification and Characterization of Lipase from Pseudomonas fragi CRDA 037. Process Biochemistry. 32, 3: 225 - 232.
Kirk, O. Borchert, T.V., Fuglsang, C.C. 2002. Industrial Enzyme Applications. Curn Opm Biotechnol.13 : 345-351.

86
Krog, N.J. 1990. Food Emulsifier and Their Chemical and Physical Properties In Food Emulsion, (Ed) K. Larsson and S.E. Friberg, P. Marcel Dekker, New York.
Kuchel, P.W., and Ralston, G.B. 1988. Theory and Problem of Biochemistry. Departement of Biochemistry The University of Sydney, Australia. P. 357-368
Lee, D.H., Park, C.H., Yeo, J.M., and Kim, S.W. 2006. Lipase Immobilization on Silica Gel Using a Cross-linking Method. J. Ind. Eng. Chem. 12 (5) 777-782
Ling-Zhi Cheong, Chin-Ping Tan, Long, K. Yusoff, M.S.A., Arifin, N. Seong-Koon Lo, and Oi-Ming Lai. 2007. Production of a Diacylgliserol-Enriched Palm Olein Using Lipase-Catalyzed Partial Hydrolysis: Optimization Using Response Surface Methodology. Food Chemistry. 105 :1614-1622
Liu, IL. and Tsai S.W. 2003. Improvements in Lipase Production and Recovery from Acinobacter radioresistens in Presence. Appl Biochem Biotechnol. 104: 129-140
Lowry, O.H., Rossbrough, N.J., Farr, A.L., and Randall, R. J. 1951. Protein Measurement With The Folin Phenol Reagent. Journal of Biol. Chem. 193: 265-275.
Luchai, S. and Dharmsthiti, S. 1999. Production, Purification and Characterization of Thermophilic Lipase from Bacillus sp. THL 027. FEMS Microbiol. Lett. 179, 2 : 241-246.
Maia, M.M.D., Morais, M.M.C., Morais, M.A., Melo, E.H.M., and Filho, J.L.L. 1999. Production of Extracellular Lipase by The Phytopathogenic Fungus Fusarium solani FSI, Rev. Microbial. 30.
Mingrui Yu, Shaowei Qin, Tianwei Tan. 2007. Purification and Characterization of The Extracellular Lipase Lip2 from Yarrowia lipoliytica. Process Biochemistry. 42 :384-391
Murray, M. Rooney, D. Van Neikerk, M. Montenegro, A. and Weatherley L.R. 1997. Immobilization of Lipase Onto Lipophilic Polymer Particle and Application to Oil Hydrolysis. Process Biochemistry. 32, 6 : 479-486.
Murray, R.K., Granner, D.K., and Victor, R.W. 2009. Biokimia Harper. EGC Penerbit Buku Kedokteran Jakarta. 68-69.

87
Nascimento, A.E., and Takaki, G.M.C. 1994. Effect of Sodium Dodecyl Sulfate on Lipase of Candida lipolytica. Appl. Biochem. Biotechnol. 49 : 93-99.
Nawani, N. Dosanjh, N.S., and Kaur, J. 1998. A Novel Thermostable Lipase from A Thermofilic Bacillus sp: Characterization and Esterification Studies. Biotechnology Letters. 20 : 997-1000.
Nawani, N. Singh, R. and Kaur, J. 2006. Immobilization and Stability Studies of Lipase from Thermophilic Bacillus sp: The Effect of Process Parameters on Immobilization of Enzyme. Electronic Journal of Biotechnology. 9 : 559-565.
Noureddini, H. 2004. A Continuous Process for The Glycerolysis of Soybean Oil. Journal of American Oil Chemistry Society. vol 81 no 2 : 203-207.
O’Brien, R.D.1998. Fats and Oils: Formulating and Processing for Applications. Technomic Publ. Co. Inc. Lancaster. Basel.
Pelczar Jr, M. J. 1986. Dasar-dasar Mikrobiologi. Universitas Indonesia Press, Jakarta
Pirman, dan Aryantha, I.Ny.P. 2003. Eksplorasi dan Isolasi Enzim Glukosa Oksidase dari Fungi Genus Aspergillus dan Penicillium Galur Indigenus. Laporan Hasil Penelitian Dasar Lembaga Penelitian ITB, Bandung.
Praweda. 2004. Penerapan Teknologi Enzim, (Online),(http://free.vlsm.org/v12/sponsor/sponsor_pendamping/praweda/biologi.htm,diakses 16 september 2004).
Putranto, A.R., Santoso, D. Tri-Panji, Suharyanto, dan Budiani, A. 2006. Karakterisasi Gen Penyandi Lipase dari Kapang Rhizopus oryzae dan Absidia corymbifera. Menara Perkebunan. 74(1) 23-32.
Ritzmann, M. 1991. Metodologi Isolasi Enzim dan Aktivitasnya. Pusat Antar Universitas Bidang Ilmu Hayati-Institut Teknologi Bandung (PAU BIH-ITB). 5-23.
Samsumaharto, R.A. 2008. Partial Characterization of Lipase From Cocoa Beans (Theobroma cacao, L.) of Clone PBC 159. Indo. J. Chem. (3) 448-453.

88
Schmid, R.D., Schmid-Dannert, C. Sztajer, H. W., Stocklein, and Menge, U. 1994. Screening, Purification and Properties of a Thermophilic Lipase from Bacillus thermocatenulatus. Biochem. Biophys.Acta. 1214 : 43-53.
Schmid, R.D., Schmid-Dannert, C. Rua, M.L., Wahl, S. and Sprauer, A. 1997. Thermoalkalophilic Lipase of Bacillus Thermocatenulatus. Large Scale Production, Purification and Properties: Aggregation Behaviour and its Effect on Activity. J. of. Biotechnol. 56 : 89-102.
Schmid, R.D., Minning, S. and Schmid-Dannert, C. 1998. Functional Expression of Rhizopus oryzae Lipase in Pichia Pastoris : High-Level Production and Some Properties. J. of. Biotechnol. 66 : 147-156.
Shaoxin Chen, Lili Qian, and Bingzhao Shi. 2007. Purification and Properties of Enantioselective Lipase From a Newly Isolated Bacillus cereus C71. Process Biochemistry. 42 (988-944).
Sharma, R. Chisti, Y. and Banerjee, U.C. 2001. Production, Purification, Characterization, and Application of Lipase. Biotechnology Advances. 19: 627-662.
Sigurgisladottir, S. Konraosdottir, M. Jonsson, A. Kristjansson, J. K., and Matthiasson, E. 1993. Lipase Activity of Thermophilic Bacteria from Icelandic Hot Springs. Biotechnology Letters. vol. 15, no.4, p. 361-366.
Silva, V. D. M., De Marco, L.M., Afonso, W.O., Lopes, D.C.F., and Silvestre M.P.C. 2007. Comparative Study of the Immobilization of Pancreatin and Papain on Activated Carbon and Alumina, Using Whey as Protein Substrate. World Applied Sciences Journal. 2 (3):175-183.
Soeka, Y.S., Sulistyo, J. dan Naiola, E. 2008. Analisis Biokimia Minyak Kelapa Hasil Ekstraksi Secara Fermentasi. Biodiversitas. vol.9, no. 2: 91-95.
Somkuti, G.A., and Babel, F.J. 1968. Lipase Activity of Mucor pusillus, Appl. Microbiol. 16: 617-619.
Sonntag, N.O.V., Formo, W.M., Jugerman, E. Noris, F.A. 1979. Bailey’s Industrial Oil and Fat Product. John Willey and Sons, New York.

89
Steve. 2008. (Online), (http://www.steve.gb.com/images/science/ion_exchange_chromatography.png, diakses 28 Mei 2008).
Stryer, L. 1995. Biochemistry. 4 th ed., W.H. Freeman and Company, New York. p. 45-47, 911-915.
Suhartono, M.T. 1989. Enzim dan Bioteknologi. Direktorat Jenderal Pendidikan Tinggi Antar Universitas Bioteknologi, Institut Pertanian Bogor. 172-220.
Suzuki, T. Mushiga, Y. Yamane, S. and Shimizu .1998. Mass Production of Lipase by Fedbatch Culture of Pseudomonas fluorescens. Appl. Microbiol. 27 : 417-422.
Syah, A. 2005. Biodiesel Jarak Pagar Bahan bakar Alternatif yang Ramah Lingkungan. Agro Media Pustaka, Jakarta.
Tarigan, P. 1991. Teknik Aerasi dan Agitasi. Pusat Antar Universitas Bioteknologi Institut Teknlogi Bandung. 25-45.
Tri-panji, Suharyanto, dan Arini, N. 2008. Lipase Spesifik 1,3-gliserida dari Fungi Lokal untuk Biokonversi CPO Menjadi Diasilgliserol. Menara Perkebunan. 76(1):11-22.
Trudel, J. and Asselin, A. 1989. Detection of Chitinase After Polyacrylamide Gel Electrophoresis. Analytical Biochemistry. 178: 362-366.
Utami, T. Indrati, R. Hastuti, P. dan Kusumawati, I. 2001. Amobilisasi Lipase dari Candida rugosa Secara Adsorpsi Menggunakan Zeolit dan Polipropilena. (online),(http://www.dikti.org/p3m/abstrakHB/abstrakHB03.pdf, diakses 9 Januari 2005).
Vorderwulbecke, T. Kieslich, K . and Erdmann, H. 1992. Comparison of Lipases by Different Assay. Enzyme. Microb. Technol. 14 : 631-639.
Wang, D.I.C., Cooney, C.L., Demain, A.L., Dunhill, P. Humprey, A.E., and Lily, M.D. 1979. Fermentation and Enzyme Technology. John Willey and Sons, New York.

90
Whitaker, R.J. 1972. Principle of Enzymology for the Food Science, Mergel Dekker inc, New York. 561-570.
Woodrof, J.G. 1979. Coconut: Production, Processing, Products. 2nd ed. AVI Publishing Co., Inc, Weaport, Conneticut.
Wu, X.Y., Jaaskelainen, J. and Linko, Y. 1996. Enzyme, Microb. Technol. 19: 226-231.
Wulan, P.P.D.K., Rejoso, M. T., dan Hermansyah, H. 2008. Reaksi Hidrolisis Minyak Zaitun Menggunakan Lipase Rhizopus oryzae yang Diimobilisasi Melalui Metode Adsorpsi. Departemen Teknik Kimia Fakultas Teknik Universitas Indonesia, Depok.

91
Lampiran 1. Skema kerja isolasi dan identifikasi A. oryzae pada kopra berjamur
Kopra berjamur
Diambil 2 ose, dimasukkan ke dalam tabung reaksi yang berisi 10 mL NaCl 0,9%Dilakukan pengenceran secara berseri dari 10-1- 10-9, dimasukkan masing-masing ke dalam 9 buah cawan petri kemudian dituangkan media agar.
Cawan petri ditumbuhi jamur
Cawan petri berisi biakan
Cawan petri ditumbuhi jamur dengan warna yang seragam
Diinkubasi pada suhu 37 0C selama 4 hari
Dipindahkan ke media cawan petri yang lain dengan metode gores kuadran
Diidentifikasi secara mikroskopik, teridentifikasi sebagai A. oryzae

92
Lampiran 2. Skema kerja produksi enzim lipase dari A. oryzae
A. oryzae
Media produksi mengandung enzim lipase
Dimasukkan ke dalam erlenmeyer yang mengandung media inokulumDikocok selama 4 hari pada suhu 37 0C dengan kecepatan 150 rpm
Media inokulum mengandung A. oryzae
Dimasukkan ke dalam erlenmeyer yang mengandung media produksiDikocok pada suhu 37 0C dengan kecepatan 150 rpm
Setiap hari sampel diambil dan disentrifugasi pada suhu 4 0C dengan kecepatan 3500 rpm, selama 30 menit untuk menentukan waktu produksi enzim lipase secara maksimal
Supernatan Sel
- Uji aktivitas enzim lipase- Uji kadar protein
Penentuan berat sel kering

93
Lampiran 3. Skema kerja optimasi produksi enzim lipase dari A. oryzae
A. oryzae
Media inokulum mengandung biakan A. oryzae
Media produksi mengandung enzim lipase
Supernatan
Sel
Dimasukkan ke dalam erlenmeyer yang mengandung media inokulumDikocok selama 4 hari pada suhu 37 0C dengan kecepatan 150 rpm.
Dimasukkan ke dalam erlenmeyer yang mengandung media produksi, dengan variasi konsentrasi pepton (0,5; 0,75; 1,0; 1,25; 1,50) %Dikocok selama 8 hari pada suhu 37 0C dengan kecepatan 150 rpm
Disentrifugasi pada suhu 4 0C dengan kecepatan 3500 rpm, selama 30 menit
Hal yang sama dilakukan untuk variasi konsentrasi minyak zaitun (1, 2, 3, 4, 5) % dan kecepatan pengadukan (50, 100, 150, 200, 250) rpm

94
Lampiran 4. Skema kerja pemurnian dan imobilisasi enzim lipase dari A. oryzae serta aplikasinya sebagai biokatalisator untuk hidrolisis minyak kelapa menjadi diasilgliserol
Karakterisasi enzim lipase imobil - Penentuan suhu optimum - Penentuan pH optimum - Kestabilan termal - Kestabilan operasional - Unjuk kerja enzim lipase sebagai
biokatalisator untuk hidrolisis minyak kelapa menjadi diasilgliserol
Enzim Lipase murni
Enzim Lipase imobil
TAG (DAG)
Uji aktivitas enzim
Supernatan
Tahap pemurnian - Fraksinasi dengan amonium sulfat
(0-100) %- Dialisis- Kromatografi kolom penukar ion- Kromatografi kolom filtrasi gel- Elektroforesis
Karakterisasi enzim lipase murni- Penentuan suhu optimum- Penentuan pH optimum- Imobilisasi enzim lipase metode adsorpsi

95
Top Related
Copyright © 2022 FDOKUMEN

