
bazı 2-sübstitüe-1h-benzimidazol türevleri üzerinde çalışmalar

Zinc Binding in PeptideModels of Angiotensin-IConverting Enzyme ActiveSites Studied Through1H-NMR and Chemical ShiftPerturbation Mapping
Athanassios S. Galanis1
Georgios A. Spyroulias1
Roberta Pierattelli2
Andreas Tzakos3
Anastassios Troganis4
Ioannis P. Gerothanassis3
George Pairas1
Evy Manessi-Zoupa5
Paul Cordopatis1
1 Department of Pharmacy,University of Patras,
GR-26500, Patras, Greece
2 Department of Chemistryand Magnetic Resonance
Center,University of Florence,
1-50019 Sesto Fiorentino(Florence), Italy
3 Department of Chemistry,University of Ioannina,
GR-45110, Ioannina, Greece
4 Department of BiologicalApplications and Technology,
University of Ioannina,GR-45110, Ioannina, Greece
5 Department of Chemistry,University of Patras,
GR-26500, Patras, Greece
Received 5 August 2002;accepted 3 December 2002
Abstract: We report the design and synthesis through solid phase 9-flourenylmethoxycarbonyl(Fmoc) chemistry of the two angiotensin-I converting enzyme active sites possessing the generalsequence HEMGHX23EAIGDX3. Their zinc-binding properties were monitored in solution throughhigh-resolution 1H-NMR. The obtained data were analyzed in terms of chemical shift differences.The results indicate that zinc binds to the HEMGH and the EAIGD characteristic motifs, andsuggest possible coordination modes of zinc in the native enzyme. © 2003 Wiley Periodicals, Inc.Biopolymers 69: 244–252, 2003
Keywords: angiotensin-I converting enzyme; renin–angiotensin system; zinc binding motifs;NMR; chemical shift index
Correspondence to: Georgios A. Spyroulias; email:[email protected]
Contract grant sponsor: University of Patras (UOP), GRSTPENED-99 Program, and EC
Contract grant number: 2790 (UOP) and HPRI-CT-1999-00009(EC)Biopolymers, Vol. 69, 244–252 (2003)© 2003 Wiley Periodicals, Inc.
244

Angiotensin-I converting enzyme (ACE) (EC3.4.15.1) is among the principal components of therenin–angiotensin system.1 It is found in two dif-ferent isoforms, the somatic and the testis, possess-ing two and one zinc-containing active sites, re-spectively.2,3 ACE is a gluzincin zinc dipeptidylcarboxypeptidase I,4 – 6 of which thermolysin isconsidered the prototypical member.6,7 ACE catal-yses the hydrolysis of angiotensin-I peptide8 andplays a crucial role in the release of angiotensin-IIvasopressor peptide in blood.1,9 Somatic ACE(1306 AA, 150 kDa) has two homologous domains,one at each terminus, which contains the zinc cat-alytic sites.10 The testis isoform is composed of 732residues (83 kDa) with a 665-residue C-terminusthat is identical to the C-terminal domain of thesomatic form.11–14 Zinc is believed to bind to thepolypeptide frame that possesses two characteristicgluzincins’ HEXXH and EXIXD motifs separatedby a 23-residue spacer.3,15 It has been proposed3,6
that the two histidine residues in the former se-quence are two of the four zinc ligands, while thethird ligand is the glutamate residue of the lattersequence and the fourth is a water molecule. Theability of H2O to act as a zinc ligand has beenestablished for a large variety of zinc enzymesindependently by amino acid composition and rel-ative positioning of their binding motifs.16,17 Theaspartate in the EXIXD sequence is believed topossess a significant role in the catalytic activity ofACE18; for instance, in thermolysin the chargedside chain of this aspartate forms a critical salt linkwith the imidazole ring of the first zinc-ligatedhistidine.19,20
Although the primary sequence of both ACE iso-forms has been identified,14 no three-dimensional(3D) structures had been determined until recently,and structure–function relationship information couldonly be inferred by loss of enzyme activity whenspecific residues were replaced through site-directedmutagenesis.15,21–23 Such amino acid substitutionswere guided by comparison with homologous motifsof other zinc metallopeptidases with similar function.Information on active site conformation and zinc co-ordination geometry were extracted through compar-ison of ACE with known 3D structures of other mem-bers of the metallopeptidase superfamily.6,17–19,24,25
Thus, we designed and synthesized two 36-residuepeptides26,27,* of the HEMGH–X23–EAIGD–X3
(89% homology) amino acid sequence (X � nativeamino acids), which represent the primary sequenceof the two zinc-containing catalytic sites existing inthe larger, human somatic isoform. In this article wereport for the first time results concerning the engi-neering of ACE active sites and the zinc-bindingproperties of the synthesized model peptides deter-mined in solution through high-resolution 1H-NMRspectroscopy.† The results reported here constitute thefirst part of a broader study focused on structuralcharacterization and interaction of ACE active siteswith various ACE inhibitors. The 36-residue N-termi-nal ACE active site (residues 361–396) will be re-ferred to ACEN, whereas the C-terminal active site(residues 959–994) will be denoted as ACEC, accord-ing to the numbering of the somatic ACE primarysequence.
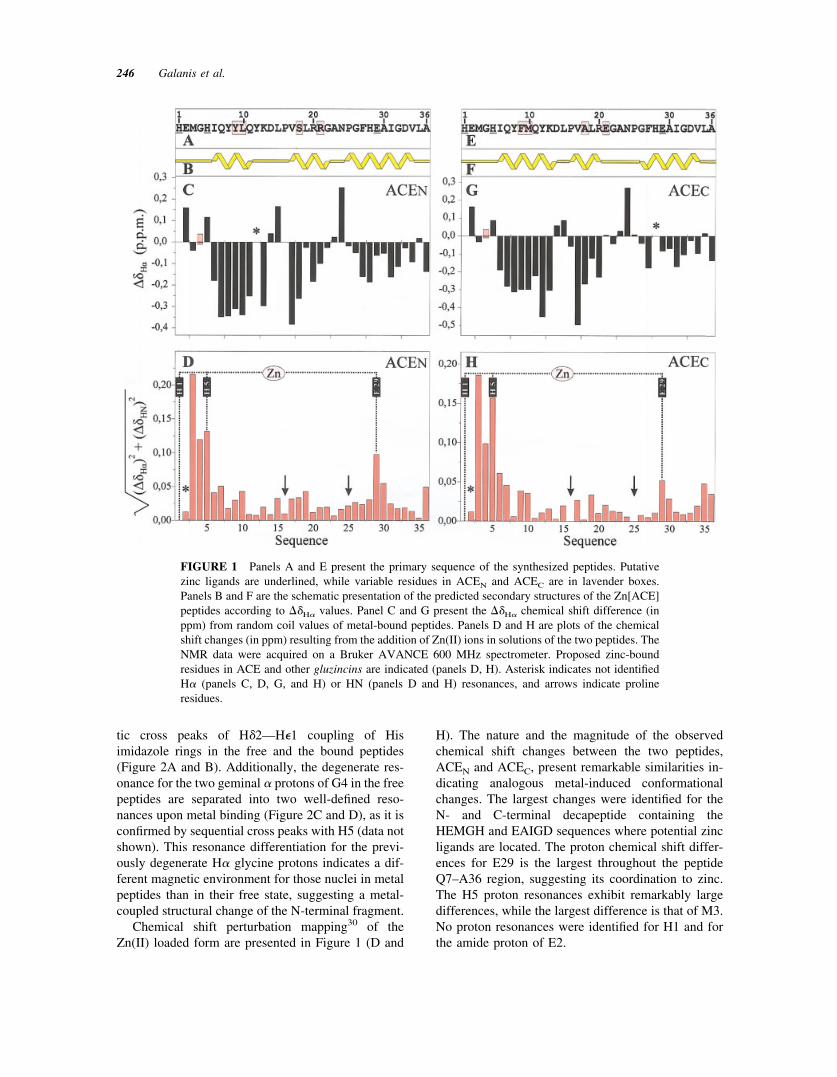
In Figure 1 (A and E) the sequences of the synthe-sized peptides are presented. Analysis of the chemicalshift values of H� and HN proton resonance‡ andtheir deviation from random coil values28,29 for thefree peptides indicates that under the present experi-mental conditions the two peptides fold in a unique,rather helical, structure (data not shown). Similar el-ements of secondary structure were observed in thepresence of Zn(II). Indeed, the majority of residues(23 residues for ACEN and 22 for ACEC) exhibitnegative deviation (larger than 0.05 ppm) from ran-dom coil values (Figure 1, panels C and G). In bothsequences these residues define three regions wherethe criteria set for prediction of helical secondarystructure by Wishart et al.28,29 are fulfilled (Figure 1,panels B and F). These three fragments are defined by7 (in ACEN) and 8 (in ACEC) residues close to thepeptide N-terminus (I6-K13), by 6 residues in themiddle of the sequence (V17–R/E21) and by 10 and 8residues close to the peptide C-terminus (G26–V34)in ACEN and ACEC, respectively.
Upon Zn(II) addition a significant number of pro-ton resonances in the NH region are shifted consid-erably while their half-width, are noticeably in-creased. These features are illustrated by the diagnos-
* Both 36-residue peptides were synthesized by the solid-phasemethod using 9-flourenylmethoxycarbonyl (Fmoc) chemistry26 andwere purified by reverse phase high performance liquid chroma-
tography (HPLC) using a linear gradient H2O/CH3CN buffer sys-tem on a C-18 column.27
† The metal binding studies for both ACEC and ACEN wereperformed on a 2 mM peptide solution in TFE/H2O (2:1, v/v),containing 0.05M Tris HCl and 0.2M NaCl, at which 1.1 eq ofZnCl2 solution prepared in an identical buffer was added. TFE isused in order to avoid the aggregation/precipitation of free andmetal bound peptide observed in aqueous solutions (at pH valuesabove 5.0 and 3.8, respectively). The free peptides samples wereprepared under the same experimental conditions.
‡ No HN and H� proton resonances were identified for H1,while only the H� resonance was identified for Q2.
Zinc Binding in Peptide Models 245
tic cross peaks of H�2—H�1 coupling of Hisimidazole rings in the free and the bound peptides(Figure 2A and B). Additionally, the degenerate res-onance for the two geminal � protons of G4 in the freepeptides are separated into two well-defined reso-nances upon metal binding (Figure 2C and D), as it isconfirmed by sequential cross peaks with H5 (data notshown). This resonance differentiation for the previ-ously degenerate H� glycine protons indicates a dif-ferent magnetic environment for those nuclei in metalpeptides than in their free state, suggesting a metal-coupled structural change of the N-terminal fragment.
Chemical shift perturbation mapping30 of theZn(II) loaded form are presented in Figure 1 (D and
H). The nature and the magnitude of the observedchemical shift changes between the two peptides,ACEN and ACEC, present remarkable similarities in-dicating analogous metal-induced conformationalchanges. The largest changes were identified for theN- and C-terminal decapeptide containing theHEMGH and EAIGD sequences where potential zincligands are located. The proton chemical shift differ-ences for E29 is the largest throughout the peptideQ7–A36 region, suggesting its coordination to zinc.The H5 proton resonances exhibit remarkably largedifferences, while the largest difference is that of M3.No proton resonances were identified for H1 and forthe amide proton of E2.
FIGURE 1 Panels A and E present the primary sequence of the synthesized peptides. Putativezinc ligands are underlined, while variable residues in ACEN and ACEC are in lavender boxes.Panels B and F are the schematic presentation of the predicted secondary structures of the Zn[ACE]peptides according to ��H� values. Panel C and G present the ��H� chemical shift difference (inppm) from random coil values of metal-bound peptides. Panels D and H are plots of the chemicalshift changes (in ppm) resulting from the addition of Zn(II) ions in solutions of the two peptides. TheNMR data were acquired on a Bruker AVANCE 600 MHz spectrometer. Proposed zinc-boundresidues in ACE and other gluzincins are indicated (panels D, H). Asterisk indicates not identifiedH� (panels C, D, G, and H) or HN (panels D and H) resonances, and arrows indicate prolineresidues.
246 Galanis et al.

The obtained data suggest that in the zinc-boundpeptides the second putative binding motif (EAIGD)is part of a helix, while for the two histidyl motif nodefinite conclusion could be extracted. Two (E2, H5)out of four observed H� proton resonance for theHEMGH sequence exhibit a positive deviation fromrandom coil structure larger than, or close to, 0.1 ppm.These findings, even if they cannot be unambiguouslyinterpreted in terms of extended conformation, sug-gest the presence of a nonhelical character for thatmotif. Despite the fact that the chemical shift differ-ences for this N-terminal pentapeptide should be an-alyzed with care, due to the restriction of its confor-
mational flexibility in the zinc-bound form, this resultis somewhat surprising. Indeed, this motif has beenfound to be part of a helix in the crystal structure ofthermolysin.31,32 However, helix-diagnostic d�N(i� 3) type connectivity between G4 and Q7, present inthe nuclear Overhauser effect spectroscopy (NOESY)spectrum of free peptides, disappears when Zn(II) isadded (Figure 2C and D). Nevertheless, characteristichelix-type d�N (i � 4) and d�N (i � 3) in free peptidesbetween I6–M10 and Q7–M10, respectively, are stillobservable in the zinc-bound peptides. These datasuggest that the HEMGH motif conformation mightundergo a structural transition from a helix-like (or
FIGURE 2 Downfield regions of 600 MHz NOESY spectra (T � 298 K) with characteristic Hisimidazole ring and H�—HN cross peaks of ACEC peptide before (panel A and C, respectively) andafter the Zn(II) addition (panel B and D, respectively). Intraresidue H�—HN cross peaks of selectedN-terminal residues are illustrated in coral and black, while interesidue d�N(i � 1,3,4) cross peaksare illustrated in light coral. The NOESY experiments were performed in the time proportionalphase incrementation (TPPI)37 mode; 2048 � 1024 data points were acquired, with 16–32 transientsper increment, over a 6000 Hz spectral width, by using a 1.5 s relaxation delay and �m � 200 ms.
Zinc Binding in Peptide Models 247

partially helix) to a nonhelical conformation whenzinc is bound, while the helical structure for thefragment I6–K13 remains intact. This is likely due tothe fact that the first peptide zinc ligand, H1, is alsothe first residue in the peptide sequence. This confersthe H1 backbone and side chain a remarkably highdegree of conformational freedom. Thus, on zincbinding, the pentapeptide-binding sequence HEMGHcould be accommodated in an unstructured conforma-tion. This is reflected by the absence of typical helixsequential NOEs (see Figure 3). Preliminary distancegeometry calculations for the ACEN 36 residue areconsistent with this interpretation.
On the other hand, no changes for the fragmentI6–K13 are implied by NMR data analysis. Morespecifically, the helix-diagnostic sequential NOE hasbeen detected for this fragment and preliminary struc-ture calculation indicate that the HEMGH in ACEN
follows a helical fragment extended from I6 to Y12/K13. Another difference between free and zinc-boundpeptides, which has been identified through sequentialconnectivities diagram (Figure 3), is the absence ofthe helical fragment in the area S18–R21 in ACEN
zinc peptide. The majority of NH—NH and H�—NHsequential connectivities identified in free ACEN havenot been detected after zinc addition. This could also
FIGURE 3 Schematic representation of the sequential and medium range NOEs involving HN,H�, and H� protons for ACEN free (upper panel) and ACEN zinc-bound peptide (lower panel).Lavender boxes indicate the residues, which are different in the two ACE active sites.
248 Galanis et al.
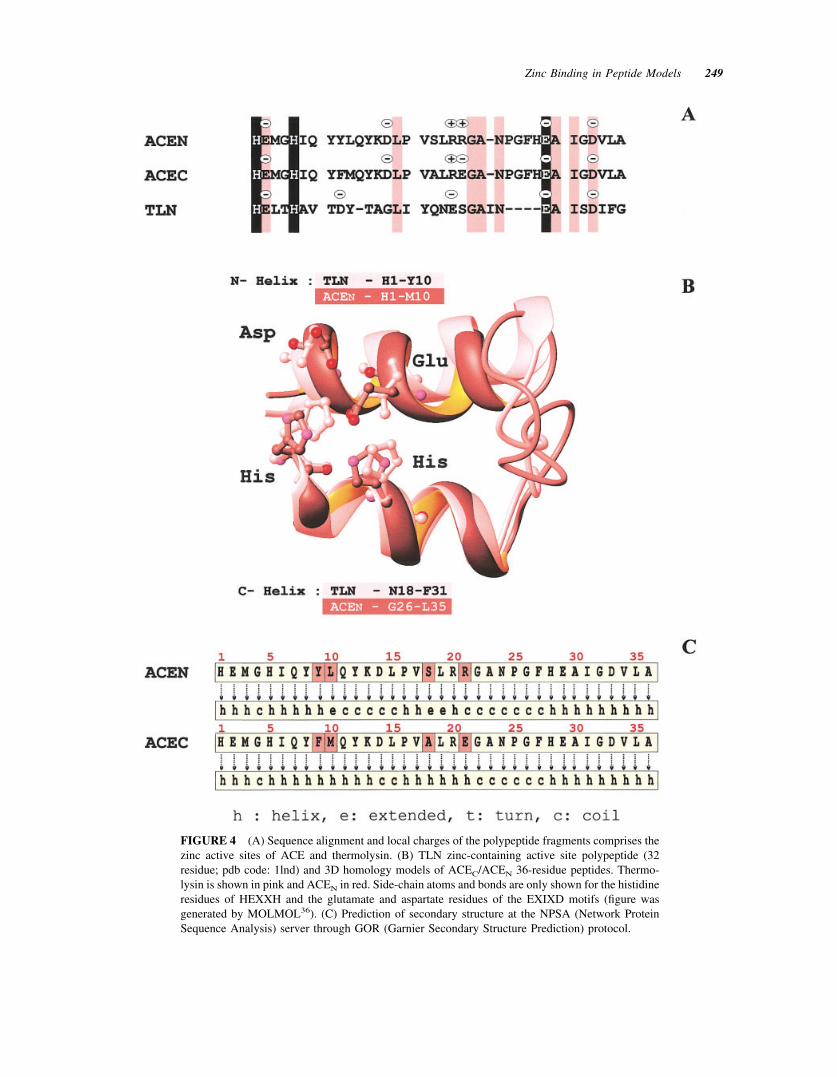
FIGURE 4 (A) Sequence alignment and local charges of the polypeptide fragments comprises thezinc active sites of ACE and thermolysin. (B) TLN zinc-containing active site polypeptide (32residue; pdb code: 1lnd) and 3D homology models of ACEC/ACEN 36-residue peptides. Thermo-lysin is shown in pink and ACEN in red. Side-chain atoms and bonds are only shown for the histidineresidues of HEXXH and the glutamate and aspartate residues of the EXIXD motifs (figure wasgenerated by MOLMOL36). (C) Prediction of secondary structure at the NPSA (Network ProteinSequence Analysis) server through GOR (Garnier Secondary Structure Prediction) protocol.
Zinc Binding in Peptide Models 249
be related to the fact that many of these protons havebeen found to resonate in similar spectral regions withalmost identical chemical shift values.
Other significant chemical shift differences areidentified in the region covered by L15–G22, whichconstitutes the intermediate fragment of a 23-residuespacer between the two proposed binding motifs. Thisfragment is susceptible to exhibiting a differentiationin HN and H� proton shift values since zinc coordi-nation requires a bent peptide skeleton in order for thedonor atoms of the potential peptide ligands in N- andC-terminal sequences to achieve a distorted-tetrahe-dral or trigonal-bipyramidal coordination geometry.These types of geometries are the most frequentlyencountered in other members of the zinc metallopep-tidase family. Furthermore, bent polypeptide frag-ments have been identified in other gluzincs, such asthermolysin, which presents similar topological (seeFigure 4A) and conformational features for the
polypeptide fragment of its catalytic site. Homologymodeling based on thermolysin binding motifs(HELTH and EAISD separated by a 19-residuespacer), using its x-ray structure (EC 3.4.24.27; pdbcode: 1lnd; Ref. 24), suggest that the ACE peptidebackbone atoms could adopt a helical conformation atthe N- and C-terminal (Figure 4B). The helical N- andC-terminus fragments are similar in magnitude withthe fragments for which sequential NOEs typical forhelix conformation have been identified (Figures 3and 4B). Helical structure for the two termini as wellas for the intermediate fragment covered by P16–R/E21 was also predicted through GOR protocols33,34ona NPSA server35 (see Figure 4C). Thus, theoreticaland modeling data for peptide secondary structureprediction are in great agreement with the elements ofsecondary structure suggested by NMR data, such asNOEs, chemical shifts, etc. The only variation be-tween helix-like in free and nonhelical in Zn(II)-
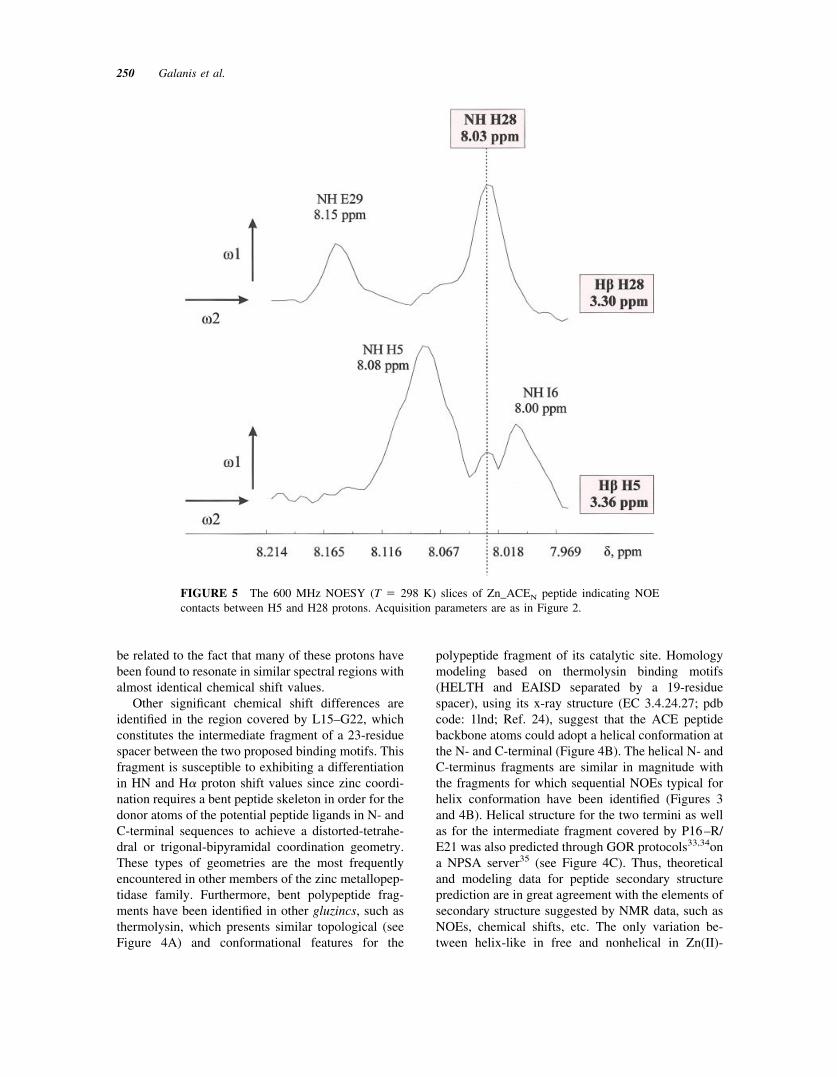
FIGURE 5 The 600 MHz NOESY (T � 298 K) slices of Zn_ACEN peptide indicating NOEcontacts between H5 and H28 protons. Acquisition parameters are as in Figure 2.
250 Galanis et al.

bound peptide conformations is focused on the N-terminal, first, binding motif. However, an orderednonhelical conformation for the HEMGH sequence ofthese model peptides could not be excluded since thisfragment comprises the N-terminal peptide fragment.This assumption is fully consistent with the observedelements of secondary structure obtained throughchemical shift analysis.
Moreover, according to thermolysin-based homol-ogy model of the ACEN peptide (Figure 4B) the twozinc binding motifs should be in spatial proximity,with protons of N- and C-terminal residues close toeach other. Indeed, analysis of Zn–ACEN NOESYmaps revealed long-range NOEs between the residuepairs G4–L35, H5–H28 (Figure 5), and Y8-F27, allconsistent with the ACEN model, confirming the closeproximity of the two “active site helices,” the zincbinding geometry and the proposed folding for the36-residue peptide.
Summarizing, the overall evaluation of NMR dataled to the conclusion that ACE active site peptides areable to bind zinc in a native-like mode, through thecharacteristic HEMGH and EAIGD sequences.Metal-induced changes in secondary structure havebeen identified for the mobile N-terminal pentapep-tide, which is the binding sequence, and backboneconformational rearrangement of the L15–G22 regionis also suggested. Such changes are expected when apeptide binds the metal and folds in such a way thatthe two motifs of the 36-residue peptides are broughtinto a parallel “face-to-face” geometry (Figure 4B).Furthermore, the metal binding affinity of these pep-tides shows that model peptides can be designed inorder to study the structural properties of metalloen-zymes with unknown structures. Indeed, we are cur-rently synthesizing a series of peptides with substitu-tions of residues considered to be important for struc-ture and function. Studies on these peptides willunambiguously identify the coordinating residues ofzinc in the ACE-model peptides prior to calculatingtheir solution structure.
The authors acknowledge the following: Univer-sity of Patras (UOP) K. Karatheodoris Research Grant(no. 2790, to PC and GAS), GRST PENED-99 Pro-gram (to ASG, IPG, EM-Z, and GP), and EC Accessto Research Infrastructures Action of the ImprovingHuman Potential Programme (no. HPRI-CT-1999-00009), MIUR Cofin 2001 (to RP).
Note added in proof: While this paper was in press,the crystal structures of human testicular38 and Dro-sophila39 ACE were reported.
REFERENCES
1. Burell, L. M.; Johnston, C. I. In Drugs, Enzymes, andReceptors of the Renin-Angiotensin System: Celebrat-ing a Century of Discovery; Husain, A., Graham, R. M.,Eds.; Harwood Academic Publishers, The Netherlands,2000; pp 191–200.
2. Corvol, P.; Williams T. A. In Handbook of ProteolyticEnzymes; Barrett, A. J., Rawlings, N. D., Woessner,J. F., Eds.; Academic Press: New York, 1998; pp1066–1076.
3. Sourbier, F.; Alhenc-Gelas, F.; Hubert, C.; Allegrini, J.;John, M.; Tregear, G.; Corvol, P. Proc Natl Acad SciUSA 1988, 85, 9386–9390.
4. Piquilloud, Y.; Reinharz, A.; Roth, M. Biochim Bio-phys Acta 1970, 206, 136–142.
5. Soffer, R. L. Annu Rev Biochem 1976, 45, 73–94.6. Hooper, N. M. FEBS Lett 1994, 354, 1–6.7. Barrett, A. J.; Rawlings, N. D.; Woessner, J. F. In
Handbook of Proteolytic Enzymes; Academic Press:New York, 1998; pp 992–993.
8. Yang, H. Y. Y.; Erdos, E. G.; Levin, V. A. BiochimBiophys Acta 1970, 214, 374–376.
9. Skeggs, L. T.; Kahn, J. R.; Shumway, N. P. J Exp Med1956, 103, 295–299.
10. Bunning, P.; Riordan, J. F. Biochemistry 1983, 22,110–116.
11. Ehlers, M. R. W.; Fox, E. A.; Strydom, D. J.; Riordan,J. F. Proc Natl Acad Sci USA 1989, 86, 7741–7745.
12. Lattion, A. L.; Soubrier, F.; Allegrini, J.; Hubert, C.;Corvol, P.; Alhenc-Gelas, F. FEBS Lett 1989, 252,99–104.
13. Ehlers, M. R.; Riordan, J. F. Biochemistry 1991, 30,7118–7126.
14. Rieder, M. J., Taylor, S. L.; Clark, A. G.; Nickerson,D. A. Nat Genet 1999, 22, 59–62.
15. Wei, L.; Alhenc-Gelas, F.; Corvol, P.; Clauser, E. J BiolChem 1991, 266, 9002–9008.
16. Valle, B. L.; Auld, D. S. Proc Natl Acad Sci USA 1990,87, 220–224.
17. Auld, D. S. In Handbook on Metalloproteins; Bertini, I.,Sigel, A., Sigel, H., Eds.; Marcel Dekker: New York,Basel, 2001; pp 881–959.
18. Williams, T. A.; Corvol, P.; Soubrier, F. J Biol Chem1994, 269, 29430–29434.
19. Matthews, B. W. Acc Chem Res 1988, 21, 333–340.20. Colman, P. M.; Jansonius, J. N.; Matthews, B. W. J Mol
Biol 1972, 70, 701–724.21. Michaud, A.; Chauvet, M. T.; Corvol, P. Biochem
Pharmacol 1999, 57, 611–618.22. Marcic, B.; Deddish, P. A.; Jackman, H. L.; Erdos,
E. G. Hypertension 1999, 33, 835–843.23. Araujo, M. C.; Melo, R. L.; Cesari, M. H.; Juliano,
M. A.; Juliano, L.; Carmona, A. K. Biochemistry 2000,39, 8519–8525.
Zinc Binding in Peptide Models 251

24. Holland, D. R.; Hausrath, A. C.; Juers, D.; Matthews,B. W. Protein Sci 1995, 4, 1955–1965.
25. Pelmenschikov, V.; Blomberg, M. R. A.; Siegbahn,P. E. M. J Biol Inorg Chem 2002, 7, 284–298.
26. Papazacharias, S.; Pairas, G.; Kouimtzoglou, E.; Der-mitzaki, E.; Manessi-Zoupa, E.; Gravanis, A.; Cordopa-tis, P. In Peptides 2000; Martinez, J., Fehrentz, J.-A.,Eds.; EDK: Paris, 2001; pp 933–934.
27. King, D. S.; Fields, C. G.; Fields, G. B. Int J PeptProtein Res 1990, 36, 255–266.
28. Wishart, D., Sykes, B.; Richards, F. J Mol Biol 1991,222, 311–333.
29. Wishart, D., Sykes, B.; Richards, F. Biochemistry1992, 31, 1647–1651.
30. Zuiderweg E. R. P. Biochemistry 2002, 41, 1–7.31. Matthews, B. W.; Weaver, L. H.; Kester, W. R. J Biol
Chem 249, 1974, 8030–8044.
32. Holmes, M. A.; Matthews, B. W. J Mol Biol 1982, 160,623–639.
33. Garnier, J.; Osguthorpe, D. J.; Robson, B. J Mol Biol1978, 120, 97–120.
34. Gibrat, J. F.; Garnier, J.; Robson, B. J Mol Biol 1987,198, 425–443.
35. Combet, C.; Blanchet, C.; Geourjon, C.; Deleage, G.TIBS 2000, 25, 147–150.
36. Koradi, R.; Billeter, M.; Wuthrich, K. J Mol Graph1996, 14, 51–55.
37. Marion, D.; Wuthrich, K. Biochem Biophys Res Com-mun 1983, 113, 967–974.
38. Natesh, R.; Schwager, S. L. U.; Sturrock, E. D.;Acharya, K. R. Nature 2003, 421, 551–554.
39. Kim, H. M.; Shin, D. R.; Yoo, O. J.; Lee, H.; Lee, J. O.FEBS Lett 2003, 538, 65–70.
252 Galanis et al.
Copyright © 2022 FDOKUMEN




















