
The Ty1-copia group retrotransposons of Allium cepa are distributed throughout the chromosomes but...
8
Chromosome Research 1996, 4, 357-364 The Tyl-copia group retrotransposons of Allium cepa are distributed throughout the chromosomes but are enriched in the terminal heterochromatin Stephen R. Pearce, Uta Pich, Gill Harrison, Andrew J. Flavell, J. S. (Pat) Heslop-Harrison, Ingo Schubert & Amar Kumar Received 28 November 1995; received in revised form 27 March 1996; accepted for publication by H. C. Macgregor 12 April 1996 The genomic organization and diversity of the Tyl-co- pia group retrotransposons has been investigated in a monocotyledonous plant, Allium cepa. We used the polymerase chain reaction (PCR) to generate se- quences corresponding to a conserved domain of the reverse transcriptase gene of Tyl-copia retrotranspo- sons in this plant. Sequence analysis of 27 of these PCR products shows that they are a highly heteroge- neous population, a feature which is common in plants but not in yeast and Drosophila. Slot-blot analysis shows there are 100 000-200 000 copies of Tyl-copia group retrotransposons within the A. cepa genome (2C-31.7 pg), indicating that they are a significant component of the genome of th~s plant. In situ hybri- dization to metaphase chromosomes reveals that Tyl- copia retrotransposons are distributed throughout the euchromatin of all chromosomes of A. cepa but are enriched in the terminal heterochromatic regions, which contain tandem arrays of satellite sequences. This is the first clear evidence for the presence of Tyl- copia retrotransposons in the terminal heterochroma- tin of plants and contrasts with the distribution of these elements in other plant species. Key words: heterochromatin, onion, plants, retro- transposons, Tyl-copia Introduction Retrotransposons are the commonest class of eukaryo- tic transposable elements. They are distinct from other transposons by their ability to transpose via an RNA intermediate, which is converted into extrachromoso- mal DNA by reverse transcription before reinsertion (Kumar 1996). The retrotransposons comprise the long terminal repeats (LTRs) and non-LTR retrotransposons (Boeke & Corces 1989, Grandbastien 1992, Smyth 1993). The most studied group of LTR retrotransposons is the Tyl-copia group, named after the best characterized elements in Saccharomyces cerevisiae and Drosophila melanogaster. Tyl-copia group retrotransposons are present throughout the plant kingdom as highly heterogeneous populations of high copy number elements (Flavell et al. 1992a, b, Voytas et al. 1992, Hirochika et al. 1992, Hirochika & Hirochika 1993, VanderWeil et al. 1993, Leeton & Smith 1993, Pearce et al. 1996). This is in marked contrast to insect and fungal systems, where they are found in much lower copy number and as more homogeneous populations (Bingham & Zachar 1989, Boeke 1989). Several plant retrotransposons have been fully sequenced (Voytas & Ausubel 1988, Grandbastien et al. 1989, Camirand et al. 1990, Manninen & Schulman 1993). A growing body of data exists on the phyloge- netic relationships between the populations of plant Tyl-copia retrotransposons within and between plant species, but little is known of the distribution of these elements within plant genomes. In situ hybridization of Tyl-copia retrotransposon sequences to metaphase chro- mosomes in Beta vulgaris (Schmidt et al. 1995), Vicia faba (Pearce et al. 1996) and Hordeum vulgare (Suoniemi et al. 1996) has revealed that the Tyl-copia elements of these species are located throughout the euchromatin, but are less prevalent in certain heterochromatin regions. This is in marked contrast to the distribution of transposable elements in Drosophila, which predominate in the het- erochromatin (Pimpinelli et al. 1995, Carmena & Gon- zalez 1995). To extend this investigation in plants, Allium cepa was chosen for this study as its genome organization is comparatively well understood, and the heterochromatin has been studied at both molecular and cytological levels (Barnes et al. 1985, Irifune et al. 1995, Pich et al. 1996a). This paper reports on the organization of the Tyl-copia group retrotransposons in the A. cepa genome, and finds that these elements, although dispersed throughout the euchromatin of all chromosomes, have a preferential association with the terminal heterochromatin. S. R. Pearceand A. Kumar (corresponding author) are at the Cell and Molecular Genetics Department, Scottish Crop Research Institute, Invergowrie, Dundee DD2 5DA, UK. Tel: (+ 44) 1382 562731; Fax: (+ 44) 1382 562426. S. R. Pearce and A. J. Flavell are at the Department of Biochemistry, University of Dundee DD1 4HN, UK. U. Pich and I. Schubert are at the Institute of Plant Genetics and Crop Plant Research, Corrensstr 3 , D-06466 Gatersleben, Germany. G. Harrison and J. S. (Pat) Heslop-Harrison are at the Karyobiology Group, John Innes Centre, Colney Lane, Norwich NR4 7UH, UK 1996 Rapid Science Publishers Chromosome Research Vol 4 1996 357
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of The Ty1-copia group retrotransposons of Allium cepa are distributed throughout the chromosomes but...

Chromosome Research 1996, 4, 357-364
The Tyl-copia group retrotransposons of Allium cepa are distributed throughout the chromosomes but are enriched in the terminal heterochromatin
Stephen R. Pearce, Uta Pich, Gill Harrison, Andrew J. Flavell, J. S. (Pat) Heslop-Harrison, Ingo Schubert & Amar Kumar
Received 28 November 1995; received in revised form 27 March 1996; accepted for publication by H. C. Macgregor 12 April 1996
The genomic organization and diversity of the Tyl-co- pia group retrotransposons has been investigated in a monocotyledonous plant, Allium cepa. We used the polymerase chain reaction (PCR) to generate se- quences corresponding to a conserved domain of the reverse transcriptase gene of Tyl-copia retrotranspo- sons in this plant. Sequence analysis of 27 of these PCR products shows that they are a highly heteroge- neous population, a feature which is common in plants but not in yeast and Drosophila. Slot-blot analysis shows there are 100 000-200 000 copies of Tyl-copia group retrotransposons within the A. cepa genome ( 2 C - 3 1 . 7 pg), indicating that they are a significant component of the genome of th~s plant. In situ hybri- dization to metaphase chromosomes reveals that Tyl- copia retrotransposons are distributed throughout the euchromatin of all chromosomes of A. cepa but are enriched in the terminal heterochromatic regions, which contain tandem arrays of satellite sequences. This is the first clear evidence for the presence of Tyl- copia retrotransposons in the terminal heterochroma- tin of plants and contrasts with the distribution of these elements in other plant species.
Key words: heterochromatin, onion, plants, retro- transposons, Tyl-copia
I n t r o d u c t i o n
Retrotransposons are the commonest class of eukaryo- tic transposable elements. They are distinct from other transposons by their ability to transpose via an RNA intermediate, which is converted into extrachromoso- mal DNA by reverse transcription before reinsertion (Kumar 1996). The retrotransposons comprise the long terminal repeats (LTRs) and non-LTR retrotransposons (Boeke & Corces 1989, Grandbastien 1992, Smyth 1993). The most studied group of LTR retrotransposons is the Tyl-copia group, named after the best characterized
elements in Saccharomyces cerevisiae and Drosophila melanogaster.
Tyl-copia group retrotransposons are present throughout the plant kingdom as highly heterogeneous populations of high copy number elements (Flavell et al. 1992a, b, Voytas et al. 1992, Hirochika et al. 1992, Hirochika & Hirochika 1993, VanderWeil et al. 1993, Leeton & Smith 1993, Pearce et al. 1996). This is in marked contrast to insect and fungal systems, where they are found in much lower copy number and as more homogeneous populations (Bingham & Zachar 1989, Boeke 1989). Several plant retrotransposons have been fully sequenced (Voytas & Ausubel 1988, Grandbastien et al. 1989, Camirand et al. 1990, Manninen & Schulman 1993). A growing body of data exists on the phyloge- netic relationships between the populations of plant Tyl-copia retrotransposons within and between plant species, but little is known of the distribution of these elements within plant genomes. In situ hybridization of Tyl-copia retrotransposon sequences to metaphase chro- mosomes in Beta vulgaris (Schmidt et al. 1995), Vicia faba (Pearce et al. 1996) and Hordeum vulgare (Suoniemi et al. 1996) has revealed that the Tyl-copia elements of these species are located throughout the euchromatin, but are less prevalent in certain heterochromatin regions. This is in marked contrast to the distribution of transposable elements in Drosophila, which predominate in the het- erochromatin (Pimpinelli et al. 1995, Carmena & Gon- zalez 1995). To extend this investigation in plants, Allium cepa was chosen for this study as its genome organization is comparatively well understood, and the heterochromatin has been studied at both molecular and cytological levels (Barnes et al. 1985, Irifune et al. 1995, Pich et al. 1996a). This paper reports on the organization of the Tyl-copia group retrotransposons in the A. cepa genome, and finds that these elements, although dispersed throughout the euchromatin of all chromosomes, have a preferential association with the terminal heterochromatin.
S. R. Pearce and A. Kumar (corresponding author) are at the Cell and Molecular Genetics Department, Scottish Crop Research Institute, Invergowrie, Dundee DD2 5DA, UK. Tel: (+ 44) 1382 562731; Fax: (+ 44) 1382 562426. S. R. Pearce and A. J. Flavell are at the Department of Biochemistry, University of Dundee DD1 4HN, UK. U. Pich and I. Schubert are at the Institute of Plant Genetics and Crop Plant Research, Corrensstr 3 , D-06466 Gatersleben, Germany. G. Harrison and J. S. (Pat) Heslop-Harrison are at the Karyobiology Group, John Innes Centre, Colney Lane, Norwich NR4 7UH, UK
1996 Rapid Science Publishers Chromosome Research Vol 4 1996 357

S. R. Pearce et al.
Mater ia ls and methods
Plant materials The seeds of A. cepa cv. Stuttgarter Riesen were from the Allium germplasm collections of the Institute of Plant Genetics and Crop protection, Gatersleben, Germany. Plants were grown from seed in an environmental ly controlled greenhouse.
Generation of retrotransposon sequences by PCR D N A w a s p r e p a r e d f r o m l eaves of A. cepa b y the m e t h o d of S a g h a i - M a r o o f et al. (1984). P r i m e r o l i g o n u - c l eo t ides ( s e q u e n c e s d e s c r i b e d in F lave l l et al. 1992a). PCR reac t ions , s u b c l o n i n g of PCR f r a g m e n t s a n d se- q u e n c e a n a l y s i s w e r e as d e s c r i b e d p r e v i o u s l y (Flavel l et al. 1992a).
Computer analysis Computing was carried out using the Seqnet facilities at the BBSRC computing centre, Daresbury, UK. DNA sequences were cross-compared using the TREE/PAPA package of pro- grams (Doolittle & Feng 1990, Feng & Doolittle 1990) and sequence identities were determined using the GAP program of the UWGCG package (Devereux et al. 1984). The aligned sequences generated by TREE were also subject to the PAPA3 analysis (nearest neighbour method). The tree with the lowest s tandard deviation resulting from the TREE analysis was adopted.
Southern slot blotting A. cepa DNA (0.15 #g) (4.28 x 103 genomes) was serially di- luted (see Figure 3), denatured in 0.2 M sodium hydroxide / sodium chloride for 5 rain, and transferred onto a Biodyne A nylon membrane using a vacuum slot blotter (BRL). Control double s t randed Tac 4 M13mp18 subclone (3.48 ng) (4.28 x 108 molecules) was serially diluted and transferred as above. PCR fragments (5/xg) were radiolabelled to approximately 109 c.p.m.//~g, using random-pr imed labelling (Sambrook et al. 1989) and hybridized to the filters 1 # g / m l in 5 x SSC at 65~ for 100 h. The filters were washed (0.1 x SSC, 65~ and viewed by autoradiography.
in situ hybridization Seeds of A. cepa were germinated under running water for 7- 10 days and placed on filter paper soaked in 0.5% colchicine for 4 h, then fixed in 3:1 methanol-acet ic acid. Fixed root tips were washed in enzyme buffer (6 mM sodium citrate, 4 mM citric acid, pH4.6) followed by digestion with cellulase and pectinase (1.6% cellulase, Calbiochem; 0.4% cellulase; Ono- zuka, 20% pectinase; Sigma) at 37~ for 80 min. Squashes were made in 45% acetic acid and coverslips removed after freezing on dry ice. In situ hybridizat ion followed the technique of Schwarzacher et al. (1994). Briefly, chromosome preparat ions were pretreated with RNAase (100 mg/ml; 1 h, 37~ The hybridizat ion mixture consisted of 50% formamide, 10% dex- tran sulphate, 0.1-0.5% sodium dodecyl sulphate (SDS), auto- claved salmon sperm DNA (25-100 ng/1), 2 x SSC with 50 ng/s l ide of labelled probe. The probe was made by gel purification of the heterogeneous PCR fragment by agarose electrophoresis, and labelled by random primed oligonucleo-
tide labelling using biotin-ll-dUTP. The probe mix was de- natured at 70~ for 10 min and held on ice for 5 min before being added to the slides. Probe and root tip preparations were then denatured in a Hybaid Omnislide (Hybaid) at 80:C for 5 rain, before hybridizat ion overnight at 37~ Post-hy- bridization washes were carried out, the most stringent being 20% formamide in 0.1 x SSC at 37~ Sites of hybridization were detected using streptavidin-Cy3 conjugate (red emis- sion, Sigma). The chromosomes were counterstained with 2/~g/ml DAPI in McIlvaine's citric buffer pH 7 (blue emission) before examination by epifluorescence light microscopy.
Generation of satellite sequences by PCR Satellite DNA was generated by PCR using 5 ng of A. cepa DNA as template and the following primers used at a con- centration of 0.2 /xM:
UP42 5 ' - C C A C G T G A C G A A A A A A C G A A G G G T - 3 ' UP44 5 ' - C G G G A T C C C C G T G G C C G G T C T A T G - 3 '
PCR conditions were identical to those described by Flavell et aI. (1992a) except that the temperatures were 94~ for 1 min, 50~ for 1 rain and 72~ for 1 rain for 20 cycles. The PCR product was gel purified (Sambrook et al. 1989) before label- ling as above.
Southern blotting A. cepa DNA (1.5/xg), digested with BarnHI and EcoRI, was analysed by agarose gel electrophoresis using a 1% gel, and transferred to nitrocellulose membrane according to the meth- od of Sambrook et al. (1989). PCR fragments (0.25 #g) were radiolabelled to approximately 109 c.p.m.//lg, using random- primed labelling (Sambrook et al. 1989) and hybridized to the filters (0.05 /xg/ml in 5 x SSC at 65~ for 8 h). The filters were washed (0.1 x SSC, 65~ and viewed by autoradiography.
Results
Tyl-copia group retrotransposons in A. cepa are heterogeneous and present in high copy number Tyl-copia r e t r o t r a n s p o s o n s e q u e n c e s w e r e i so la t ed f rom A. cepa b y p o l y m e r a s e c h a i n r eac t i on (PCR) u s i n g de- g e n e r a t e p r i m e r s c o r r e s p o n d i n g to t w o c o n s e r v e d do- m a i n s of t he r eve r s e t r a n s c r i p t a s e gene , w h i c h h a v e b e e n u s e d e x t e n s i v e l y in the g e n e r a t i o n of s e q u e n c e s f r o m h i g h e r p l a n t s (Flavel l et al. 1992a, b, Pearce et al. 1996). The PCR p r o d u c t w a s s u b c l o n e d , a n d 27 ran- d o m l y c h o s e n i n se r t s s e q u e n c e d . The a m i n o acid se- q u e n c e s w e r e d e d u c e d , a n d these w e r e t h e n a l i gned (F igure 1). The A. cepa Tyl-copia r e t r o t r a n s p o s o n se- q u e n c e s s h o w a cha rac t e r i s t i c f ea tu re of the Tyl-copia
p l a n t s e q u e n c e , w h i c h is the c o n s e n s u s s e q u e n c e S L Y G L K Q A / S P / S R A / Q W (Flavel l et al. 1992a, b, Hir- o c h i k a & H i r o c h i k a 1993).
P h y l o g e n e t i c a n a l y s i s of the a b o v e s e q u e n c e s re- v e a l e d a h i g h d e g r e e of s e q u e n c e h e t e r o g e n e i t y (Figure
2), w h i c h is cha rac t e r i s t i c of p l a n t LTR r e t r o t r a n s p o - sons . N o n e of the s e q u e n c e s are iden t ica l , s o m e h o w -
eve r are m o s t c lose ly re la ted , a n d f o r m s u b g r o u p s , for
358 Chromosome Research Vol 4 1996

TIC29
Tac29
Tac41
T & r
Tac24
Tac4
Tac31
Tac26
Tat42
Tac42
Tac33
Tact8
Tacll
Tac8
Tacl5
Tac28
TIC7
Tac7
Tac16
Tac23
Tac25
Tac35
Tac46
Un~rouped
Tac49
Tac47
Tacl
Tacl0
Tac43
Tac6
Tac34
Tac36
Tyl-copia retrotransposons in Allium cepa l u b g r o u ] ; )
TA F L H G DLAEEVYMDEP PGFVAQR - EHG- - - KVCKLK~GLYGLKQ S PRAWFGKFSQA I EKFGSLQ SK - ADHSVF - -Y-NVKGTL ILLVVYVDDM
- LARGVY -GAP PGFVAQG - GYG- - - KVCKLF/<SLYGLKQR PRAWFGKF SQAI EKFGLLQS K - ADHLVF - -YKNMKGTLILLIV
subgroul)
HL~SSLY- DQ PMGFRNKN- YLN-- -HVCLLKKSLYGLKQAPRAWYQRFADFVAT IGFSHST - SDHSLLS - TRMV-- LI LYI LL
HLNEAVYNHQ PMGFHAV~/- YPN - - - HVFLLKKFLYGLKQAPRAWYQRFANFVAT I GF SH ST - SDHSLF I - YKNGSDI -TYI LL
H INEQVY I DQPMGFRNKN-YPN- - - HVFLLKKS LYGLKQAPRAWYQRFADFVAS I RFSHTT - SDHS * F I - YKNGSDI L I FILL
HLNKPFYKHQPMGFLNKS -YPD- - -NGCL * EKSLYGLKHAPRAWYQRFADFFATIGFSHSTFRS S ILFI -YNNGSDI -AYILL
Iubgroup
DLDGH LH - EAARR I FCRW - KRKLGEE I - - - EES LYGLK~S PKQWYKRFDSFMLS LGFDRRS - K~ACVYYK - HLKNDRWI YL LL
DFGDH LH - EQLEEFRCR * - ERKLGFKL - - - KKS LYGLKQS PRQWYKMFDS FMLSFGF~HS I - KDACVYFK - HLKNGRWVYL LL
DL DET I YMKQ I ERF FV - D * KENWVCKL - - - KKFLYGLKRRLDNGTRGLTLL~FELGFDRS I - KEACVYFK - NLKDGRWVYLLL
DL DET I Y I KQ LE I F F I DG - KVNWVCKL - - - KKS LYGLKQM PR * WSKRFDFFMLSLDFDRS I - KDVIrv-YF K - HLKNGRWVI PTH
DLDE I I YMKS QKKFVVYG - KEKWVYKL - - - KKS LYGLKSRLDNGTRDLTFFM I S LGFDRS I - KDACVYFK - HLKNGKLVYL LL
I E I E I I YKKQLKGFFVDG- KVNWLCKS-- - KKFLYGLKQSP* -WYKRFDL~M- SLSFDCS I - KDAYVYLK - YLKNVR * TYVVL
AWGSHLY- EQLEGFVI SG- KEKCFCML - - - KKSLYGLKQ S PRQWYKRFS SFMLS LGFDS S I - KDACVFLN - NL E SGSWVYLLL
Iubgroup
DLDEKTLHGVTGGFVVDG - KLNL I CKL - - - KKS LYGI KQ STRQWYKRFDYFML E~IDFDR S I - KDVF I YFK - KLRNDCWLYLLV
DLDQDLYME QLEGFVMDG - K SNLVCKL - - - NKLLYGLKK S LRQWYKRVDHFMLENGFDRN I -KDACIFFK-KLRNDC .... GV
DLDEDIYMVQPEVFVVDG- KSNLVCKL -- - KKS LYGLKQS PRQWYKRFDHFMLENGFDRST - NDACVYFK - KLRNDCWVYL LL
DLDEDIYMEQPGGEVVDG - KSNLVCKL - - - EKSLYGL *AS PRQWYKRFDHFMLENGFERS I - KDACVYFK - KLKNNCWMYLLL
DLNEDVYMEQHEGFSMI)G - KSNLVCKQ - - - }(KS LYGLMQ S PRQWYKRFDHFMLDNG F DR SN - KDACFYFK - KLKNDYWVYL LL
T S DEHIqYMEQ PEGFVVDG - K SKLVCKL - - - KKSLYVLKQS PRSWYKRFDHFMLENSFDERS-N I LCL - FQ - EVENDVGC I SLL
I I (ZLII ~"I C I I
HK PSARYLEE~FTYS SRGI HRRQAHHVCKLNKSLYGLKQS PR *WYKKFDKFMV* YGYSRS- FRGQCVYFK- KCNEE I I IYLLL
DLEEE IYME * PEGFKVPG- KEDI'VC RLCRLTKS LYGLKQAPRQWYKKLQAFMAEHDFKKME - SNTCVS SQ - SDCDGDFYYL - -
HLEE-VYMSQ PKAF I I EG- QKD- - - MVCKLKKS IYGLKQASRQWYLKFHEI ITAFDFTENI-VDPC IYK- LKFR * SKF I FL I L
D I EET IYMVQPENFVVED - PKN-- -MVCKLKKS I YGLKQASRQWYHKFHNVI TSFGFETNI- I DNCVYH - - KFNVSKF I FL I L
DLKEEVYMEQPPGFVARG-~NG - - - LVCRLKKTLYGLKHS PRAWFGKFSQTIGNFGLKKSK- EDHSVF- -YKNS ENG I I LLVV
ELQQEVYMQQPPGYVKAS- DKK - - - MVCKLHKAIYGLKQAPRVWYEKLKSTL I SLECKPTR- SDNSLY - - I KIND I GATYI LV
ELNEDVYVEQPLGYV * KG - EEE - - - KVYKLKKALYGLK* APRAWYGRI EAYFMKVSFSD - R - HDYTLF I K I EESKSFL IVNFY
ELKEEVYVEQPQGFE I EG - QEG - - - KVYKLKKALYGLKQAS RAWYNN I DTYFMEKGFEKS K - NE PTLYVNDKSMDD I L I I AL -
Figure 1. Aligned putative peptide sequences of Tyl-copia reverse transcriptase fragments from A. cepa. Sequences were aligned using CLUSTALV at the BBSRC computing centre Daresbury. , Frameshift mutations; * nonsense codons.
I
Tac29 --Tac41
Tac43 Tac6
r_rl Tac24 Tac4 Tac31
Tac26 Tac34
Tac36 Tac42 Tac33
Tac18 Tac I I Tac8
Tac15
Tac28
i~ Tac7 Tacl6 Tac23
TaC25Tac35
Tac46 Tac49 Tac47 Tacl
Tac10
i i
i i i i i i
0 10 20 30 40 50 Branch length
Figure 2. Phylogenetic analysis of nucleotide sequences representing A. cepa reverse transcriptase fragment. Nucleotide sequences corresponding to the fragments shown in Figure 1 were subjected to phylogenetic analysis, using the TREE/PAPA set of programmes (Feng & Doolittle 1990, Doolittle & Feng 1990). Vertical branch lengths are of no significance. Divergences are indicated by horizontal branch lengths. Because the trees are unrooted, there is no significance to the relative lengths of the two horizontal sections of the arms joining the deepest branches of the tree. However, the total of these two branch lengths is equal to the distance separating these from the other sequences.
C h r o m o s o m e Research Vol 4 1996 359

S. R. Pearce et al.
1 2 3 4 4.28 x 108 control DNA molecules (Tac4, slot 5). This indicates that there are about 100 000-200 000 copies of
Control (Tar 4) Tyl-copia re t ro t ransposon sequences per 2C genome in A. cepa.
Genomic DNA
Figure 3. Copy number of Tyl-copia retrotransposons in A. cepa. Row Tac4, contains cloned reverse transcriptase fragment of Tac4 sequences corresponding to 4.28 x 106 molecules (slot 1), 2.14 x 10 7 molecules (slot 2), 4.28 x 107 molecules (slot 3), 2.14 x 108 molecules (slot 4), 4.28 x 108 molecules (slot 5). Row Genomic DNA, contains 4.28 x 101 genomes (slot 1), 2.14 x 102 gen- omes (slot 2), 4.28 x 102 genomes (slot 3), 2.14 x 103 genomes (slot 4), 4.28 x 103 genomes (slot 5). The filter was probed with a heterogeneous PCR probe containing the complete population of Tyl-copia reverse transcriptase sequences from A. cepa.
example the Tac4 subgroup consists of Tac4, Tac24, Tac31 and Tac26. Within this subgroup all sequences are at least 95% identical at the nucleot ide level. We have also identif ied three other subgroups Tac29, Tac8 and Tac7 (named after the lowest integer subgroup sequence) wi th in A. cepa that are at least 90% identical at the nucleot ide level wi th in each subgroup. For eight se- quences no close relat ives exist (ungrouped sequences), for instance the closest relat ive to Tac6 is Tac43, which is only 66.5% identical at amino acid level.
To de termine the copy number Tyl-copia retrotran- sposon sequences in A. cepa, the complete heteroge- neous popu la t ion of sequences generated by PCR was hybr id ized to known amounts of genomic DNA, and also to control re t ro t ransposon DNA (Figure 3). As the genome size of A. cepa is known (2C = 31.7 pg, Bennett & Smith 1976), compar ison be tween these two hybri- d iza t ion signals enables the copy number of the Tyl- copia re t ro t ransposon popu la t ion to be de termined. This technique has been shown to be s imilar in accuracy to de te rmina t ions based on genomic l ibrary screening (Pearce et al. 1996). The intensi ty of hybr id iza t ion of the heterogeneous Tyl-copia probe to 4.14 x 103 A. cepa genomes (genomic DNA, slot 4) is the same as that of
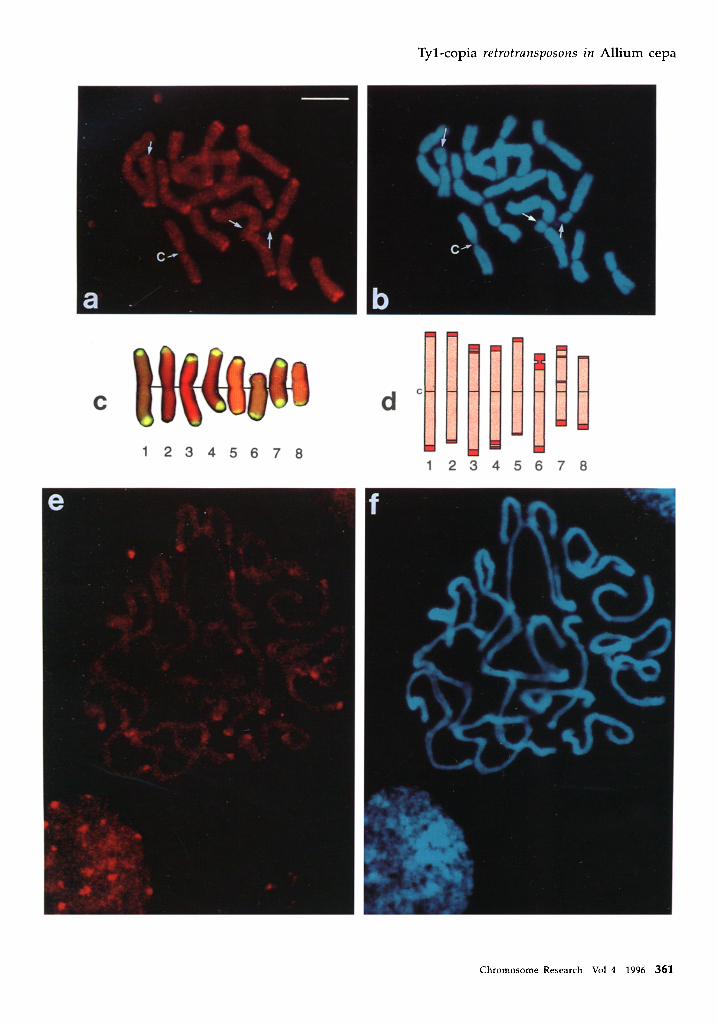
Distribution of -I-yl-copia retrotransposons in the genome The chromosomal localizat ion of Tyl-copia elements in A. cepa was de te rmined by hybr id iz ing biot in-labelled heterogeneous re t ro t ransposon probe DNA to meta- phase chromosome spreads (Figure 4a), and to pro- phase and in terphase cells (Figure 4e). Figure 4b & f show the same chromosomes as those in Figure 4a & e respectively, s tained with DAPI. The posi t ion of the Tyl-copia re t ro t ransposons is shown by red fluorescence (Figure 4a & e). The elements are present throughout the chromosomes (also shown by the hybr id iza t ion of the probe to p rophase chromosomes, Figure 4e) but are p redominan t ly located in discrete regions at the ends of most chromosomes (Figure 4a & e). These regions correspond to areas containing tandem arrays of highly repet i t ive sequences that are s t rongly s tained by Giem- sa (Figure 4c & d, Barnes et al. (1985), Irifune et al. (1995), Pich et al. (1996a)). The terminal region of the short arm of chromosome 6 (Figure 4c), where hetero- chromat in consists of condensed r ibosomal DNA, re- vealed no detectable signal (Figure 4a arrows). Also, clear signals were missing at chromosome ends with smal ler amounts of heterochromat in (e.g. the short arm of chromosome 8; Figure 4a, a r rowhead , c & d). It is interest ing that Tyl-copia sequences are present in the terminal heterochromat in that does not decondense dur ing in terphase (Figure 4e & f), a p roper ty of hetero- chromat in as defined by Heitz (1932).
As the d is t r ibut ion of the Tyl-copia and satellite sequences at the chromosome ends are so similar, it was impor tan t to exclude the possibi l i ty that these sequences cross-hybridize. Sequence comparisons of the 27 Tyl-copia sequences (Figure 1) and the 380-bp satelli te sequences (Barnes et al. 1985, Irifune et al. 1995) show they have no significant sequence ident i ty (data not shown). This was confirmed by hybr id iz ing the heterogeneous Tyl-copia probe to Southern blots con- ta ining a l adder of satelli te repeats (Figure 5a). No hybr id iza t ion of the Tyl-copia probe to the satellite
Figure 4. In situ hybridization using biotinylated Tyl-copia retrotransposon reverse transcriptase fragments to metaphase, prophase and interphase chromosomes of A. cepa. a A. cepa metaphase chromosomes hybridized with heterogeneous Tyl-copia probe. Hybridization is present throughout the length of all chromosomes but the strongest hybridization signals appear at the terminal heterochromatin regions, except at the nucleolus organizer in the short arm end of chromosome 6 (arrows). The signal intensity at individual chromosome ends (for example at chromosome 8, arrowheads) corresponds to that obtained by in situ hybridization with the 380-bp satellite sequence (4c) for comparison, b Same A. cepa metaphase chromosomes stained with DAPI. C, centromere. Scale bar = 10 #m. r Hybridization of 380 bp satellite sequence (yellow) to A. cepa metaphase chromosomes (Pich et al. 1996a). d A diagrammatic representation of A. cepa metaphase chromosomes after Giemsa banding. C, centromere, e & f A. Cepa interphase nucleus and prophase chromosomes of A. cepa probed as a and stained as b respectively.
360 Chromosome Research Vol 4 1996
Tyl-copia retrotransposons in Allium cepa
Chromosome Research Vol 4 1996 361

S. R. Pearce et al.
ladder was observed, although the retrotransposon probe does hybridize to a range of high molecular weight fragments (Figure 5b). Therefore, the preferen- tial hybridization of the Tyl-copia sequences to terminal heterochromatin is not a consequence of cross-hybri- dization between the Tyl-copia probe and terminal satellite sequence, but reflects the true distribution of Tyl-copia retrotransposons in the genome of A. cepa.
Discussion
This study has used molecular and in situ hybridization techniques to study the organization of Tyl-copia group retrotransposons within the A. cepa genome. The Tyl- copia retrotransposons of A. cepa are a heterogeneous population of between 100 000 and 200 000 sequences. Plant Tyl-copia retrotransposons are generally at least five kb in length (Grandbastein 1992). If all the A. cepa sequences we have identified belong to full-size ele- ments, this would correspond to approximately 109 bp, which is roughly 3% of the A. cepa genome. This (Figure 1) and other studies have shown that the majority of retrotransposons within plant species are defective and incomplete owing to the presence of frameshift and nonsense codons (Voytas & Ausubel 1988, Konieczny et al. 1991, Flavell et al. 1992a, Voytas et al. 1992, Hirochika & Hirochika 1993, White et al. 1994). We therefore cannot be certain of the exact percentage of the A. cepa genome that is occupied by Tyl-copia retrotransposons, but it is probably a significant proportion of the genome of this plant. High copy number and sequence hetero- geneity (Figures 1 & 2) is typical for this type of retro- transposon in plant species with large genomes (Flavell et al. 1994). For example, we have recently observed a heterogeneous population of high copy number of Tyl- copia retrotransposons in three diploid Vicia species, with one (V. faba) containing about 1 million copies of these elements (Pearce et al. 1996).
Our in situ hybridization data show that the Tyl-copia retrotransposons of A. cepa are present on all chromo- somes, throughout their entire length, but are enriched in the terminal heterochromatin (Figure 4a & e). The terminal heterochromatin of A. cepa has been studied in some detail and is at least partly composed of a 380 bp tandemly repeated satellite sequence (Barnes et al. 1985, Irifune et al. 1995, Pich et al. 1996a). Based on our in situ hybridization data, the Tyl-copia retrotransposon and satellite sequences are located within the same terminal heterochromatic regions of A. cepa chromosomes (Fig- ure 4a, c & e). The only other plant species for which the chromosomal distributions of Tyl-copia retrotranspo- sons are known are B. vulgaris (Schmidt et al. 1995), V. faba (Pearce et al. 1996) and H. vulgare (Suoniemi et al. 1996). The Tyl-copia retrotransposon sequences in these species are associated primarily with euchromatin, and are less prevalent in heterochromatic regions. For in- stance, in Vicia faba, they are absent from the interstitial heterochromatin (Pearce et al. 1996).
The reasons for the different distribution patterns of the Tyl-copia retrotransposons in these different plant species are at present unclear. One possibility could be that these retrotransposons have a preference for some sequences as targets for insertions. This has some sup- port in yeast and Drosophila, where certain LTR retro- transposons such as the Ty3 and 17.6 elements have been shown to have insertion site specificity (Inouye et al. 1984, Kirchner et al. 1995). There are differences between the heterochromatic structure of V. faba and A. cepa, and this may influence the distribution of Tyl- copia retrotransposons in these plants. In V. faba, het- erochromatic regions are located interstitially (Dobel et al. 1978), are AT rich and have a high affinity for DAPI (Kononowich, 1986, Pearce et al. 1996). The A. cepa heterochromatic regions in contrast are mainly located at the ends of chromosomes, are GC rich and therefore do not bind DAPI strongly (Figure 4b) but bind Giemsa more readily (Vosa 1976).
Evolutionary differences in the origin of heterochro- matin between V. faba and A. cepa may be another factor that influences the distribution of the Tyl-copia retro- transposons. For instance, it may be that original amplification of the sequences in the A. cepa hetero- chromatin also happened to carry along Tyl-copia group retrotransposons whereas the V. faba heterochromatin did not. Also, as the V. faba interstitial heterochromatin is relatively young in evolutionary terms (Fuchs et al. 1994) its consequence may be that the Tyl-copia retro- transposons have not yet had the chance to colonize the recently evolved interstitial heterochromatin.
The distribution pattern of Tyl-copia retrotransposons in the A. cepa genome has some similarity with the distribution of the copia elements in D. melanogaster (Kumar 1996). In D. melanogaster, the elements are not only associated with the terminal heterochromatin but are also associated with centromeric heterochromatin (Pimpinelli et al. 1995, Carmena & Gonzalez 1995). Furthermore, the long interspersed repeat elements (LINE)-like retrotransposons HeT-A and TART are the most terminal sequences of Drosophila chromosomes and have been shown to play a role in chromosome stability (Biessmann & Mason 1992, Levis et al. 1993). In most eukaryotes, chromosome ends consist of short, simple sequences that are synthesized by the enzyme telomerase (Zakian 1989). D. melanogaster however lacks these telomeric repeat sequences. The terminal nucleotide loss due to incomplete DNA replication is counterbalanced in Drosophila by transposition of HeT- A and TART sequences at high frequency to the termini, where they form tandem arrays with their oligo(A) tails facing towards the centromere (Biessman & Mason 1992, Levis et al. 1993). Recently, it has been shown that Allium species also lack tandemly repeated TTTAGGG sequences. Instead their chromosome ends are composed of highly repetitive satellite and/or rDNA sequences (Pich et al. 1996b). The results pre- sented in this paper clearly show that Tyl-copia retro-
362 Chromosome Research Vol 4 1996

T y l - c o p i a r e t r o t r a n s p o s o n s in A l l i u m c e p a
Gran t PG94/534 to A.J.F. & A.K. Part of this w o r k was also s u p p o r t e d by a g ran t f rom the K u l t u s m i n i s t e r i u m of the Land Sachsen -Anha l t to I.S.
Figure 5. Ty 1-copia reverse transcriptase fragments do not hybridize to satellite sequences, a Southern blot showing 380 bp satellite sequences (Barnes et al. 1995) hybridized to A. cepa DNA digested with EcoRV (lane 1) and BamHI (lane 2). The ladder of satellite sequences is clearly visible as indicated, b The same blot probed with a heterogeneous probe representing the entire population of A. cepa Tyl- copia reverse transcriptase sequences.
t r ansposons are also assoc ia ted wi th the t e rmina l het- e roch roma t in in A. cepa. At present , the s igni f icance of the associa t ion of the Tyl-copia g r o u p r e t ro t r ansposons and satell i te sequences w i th in the t e rmina l he te rochro- mat in of A. cepa is unclear. We an t ic ipa te that a de ta i l ed analysis of the o rgan iza t ion of Tyl-copia r e t ro t ranspo- sons and satel l i te sequences w i t h i n the t e rmina l het- e roch roma t in m a y reveal ins ights into the e v o l u t i o n of these sequences and their role, if any, in the s tabi l iza- t ion of c h r o m o s o m e s in A. cepa.
Acknowledgements
This w o r k was s u p p o r t e d by the BBSRC PMBII init ia- t ive and the SOAEFD. S.R.P. is s u p p o r t e d by BBSRC
References
Barnes SR, James AM, Jamieson G. (1985) The organisation, nucleotide sequence, and chromosomal distribution of a satellite DNA from Allium cepa. Chromosoma 92: 185-192.
Beissmann H, Mason JM (1992) Genetics and molecular biol- ogy of telomeres. Adv Genet 30: 185-249.
Bennett MD, Smith JB (1976) Nuclear DNA amounts in angiosperms. Phil Trans Royal Soc Lond B 274: 227-274.
Bingham PM, Zachar Z (1989) Retrotransposons and FB ele- ments from Drosophila melanogaster. In: Berg DE, Howe MM, eds. Mobile DNA. Washington DC: American Society for Microbiology, pp 485-502.
Boeke JD (1989) Transposable elements in Saccharomyces cerevisiae. In: Berg DE, Howe MM, eds. Mobile DNA. Wa- shington DC: American Society for Microbiology, pp 335- 374.
Boeke JD, Corces VG (1989) Transcription and reverse-tran- scription of retrotransposons. Annu Rev Microbiol 43: 403- 434.
Camirand A, St Pierre B, Martineau C, Brisson N (1990) Occurrence of a Copia-like transposable element in one of the introns of the potato starch phosphorylase gene. Mol Gen Genet 224: 33-39.
Carmena M, Gonzalez C (1995) Transposable elements map in a conserved pattern of distribution extending from Beta- heterochromatin to centromeres in Drosophila. Chromosoma 103: 676-684.
Devereux J, Haeberli P, Smithies O (1984) A comprehensive set of sequence analysis programmes for the VAX. Nucleic Acids Res 12: 387-395.
Dobel P, Schubert I, Rieger R (1978) Distribution of hetero- chromatin in a reconstructed karyotype of Vicia faba as identified by banding and DNA-late replication patterns. Chromosoma 69: 193-209.
Doolittle RF, Feng D (1990) Nearest neighbour procedure for relating progressively aligned amino acid sequences. In: Doolittle RF, ed., Methods in Enzymology, Vol. 183, 659-669. New York: Academic Press.
Feng DF, Doolittle RF (1990) Progressive alignment and phy- logenetic tree construction of protein sequences. In: Doo- little RF, ed., Methods in Enzymology, Vol. 183, 375-387. New York: Academic Press.
Flavell AJ, Smith D, Kumar A. (1992a) Copia-Ty family retro- transposon heterogeneity in plants. Mol Gen Genet 231: 233- 242.
Flavell AJ, Dumbar E, Anderson R et al. (1992b) Tyl-copia group retrotransposons are ubiquitous and heterogeneous in higher plants. Nucleic Acids Res 20: 3639-3644.
Flavell AJ, Pearce SR, Kumar A (1994) Plant trans- posable elements and the genome. Curr Opin Genet Dev 4: 838-844.
Fuchs J, Pich U, Meister I, Schubert I (1994) Differentiation of field bean heterochromatin by in situ hybridisation with a repeated Fok 1 Sequence. Chrom Res 2: 25-28.
Grandbastien M-A (1992) Retroelements in higher plants. Trends Genet 8: 103-108.
Grandbastien M-A, Spielman A, Caboche M (1989) Tntl: a mobile retroviral-like transposable element of tobacco iso- lated by plant cell genetics. Nature 337: 376-380.
Chromosome Research Vol 4 I996 363

S. R. Pearce et al.
Heitz E (1932) Die Herkunft der chromozentren. Planta 18: 571-636.
Hirochika H, Hirochika R (1993) Tyl-copia retrotransposons as ubiquitous components of plant genomes. Jpn J Genet 68: 35-46.
Hirochika H, Fukuchi A, Kikuchi F (1992) Retrotransposon families in rice. Mol Gen Genet 233: 209-216.
Inouye S, Yuki S, Saigo K (1984) Sequence-specific insertion of the Drosophila t ransposable genetic element 17.6. Nature 310: 332-333.
Irifune K, Hirai K, Zheng J, Tanaka R, Morikawa H (1995) Nucleotide sequence of a highly repeated DNA sequence and its chromosomal localization in Allium fistulosum. Theor Appl Genet 90: 312-316.
Kirchner J, Connolly CM, Sandmeyer SB (1995) Requirement of RNA polymerase III transcription factor for in vitro position-specific integration of a retrovirus-like element. Science 267: 1488-1491.
Konieczny A, Voytas DF, Cummings MP, Ausubel FM (1991) A superfamily of Arabidopsis thaliana retrotransposons. Genetics 127: 801-809.
Kononowicz AK (1986) Cytofluorometric analysis of changes of heterochromatin fraction dur ing differentiation of root cells in two subspecies of Vicia faba. Bio Zentralbl 105: 69-83.
Kumar A (1996) The adventures of Tyl-copia retrotransposons in plants. Trends Genet 12: 41-43.
Leeton PJ, Smyth DR (1993) An abundant LINE like element amplified in the genome of Lilium speciosum. Mol Gen Genet 237: 97-104.
Levis RW, Ganesan R, Houtchens K (1993) Transposons in place of telomeric repeats at Drosophila telomere. Cell 75:
1083-1093. Manninen I, Schulman AH (1993) BARE-1 a copia-like retro-
element in barley (Hordeum vulgare). Plant Mol Biol 22: 829- 864.
Pearce SR, Harrison G, Li D et al. (1996) The Tyl-copia group retrotransposons in Vicia species: copy number, sequence heterogeneity and chromosomal localisation. Mol Gen Genet 250: 305-315.
Pich U, Fritsch R, Schubert I (1996a) Closely related Allium species share a very similar satellite sequence. Plant Syst Evol (in press).
Pich U, Fuchs, J, Schubert I (1996b) How do Alliaceae stabilise their chromosome ends in the absence of TTTAGGG-se- quences? Chrom Res 4: 207-213.
Pimpinelli S, Berloco M, Fanti L e t al. (1995) Transposable elements are stable structural components of Drosophila rnelanogaster heterochromatin. Proc Natl Acad Sci USA 92: 3804-3808.
Saghai-Maroof MA, Soliman KM, Jorgensoen RA, Allard RW (1984) Ribosomal DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosomal location, and populat ion dynamics. Proc Natl Acad Sci USA 81: 8014-8018.
Sambrook J, Fritsch E, Maniatis T (1989) Molecular Cloning: a Laboratory Manual. Cold Spring Harbor Laboratory Press: Cold Spring Harbor, New York.
Schmidt T, Kubis S, Heslop-Harrison JS (1995) Analysis and chromosomal localisation of retrotransposons in sugarbeet (Beta vulgaris L.): LINEs and Tyl-Copia-like elements as major components of the genome. Chrom Res 3: 335-345.
Schwarzacher T, Leitch AR, Heslop-Harrison JS (1994) DNA in situ hybridizat ion-methods for light microscopy. In: Harris N, Oparka KJ, eds. Plant Cell Biology: A Practical Approach. Oxford: Oxford University Press, pp 127-155.
Smyth DR (1993) Plant Retrotransposons. In: Verma DPS, ed. Control of Gene Expression. London: CRC Press, ,pp 1-15.
Suoniemi A, Anamthawat-Jonsson K, Arna T, Schulman AH (1996) Retrotransposons BARE-1 is a major, dispersed com- ponent of the barley (Hordeum vulgare L.) genome. Plant Mol Biol (in press).
VanderWeil PL, Voytas DF, Wendel JF (1993) Copia-like retro- transposable elements evolution in diploid and polyploid cotton (Gossypium L.) J Mol Evol 36: 429-447.
Vosa GC (1976) Heterochromatic patterns in Allium: L. The relationship between the species of the cepa group and its allies. Heredity 36: 383-392.
Voytas DF, Ausubel FM (1988) A copia-like transposable ele- ment family in Arabidopsis thaliana. Nature (London) 336:
342-344. Voytas DF, Cummings MP, Konieczny A, Ausubel FM,
Rodermel SR (1992) Copia-like retrotransposons are ubi- quitous among plants Proc Natl Acad Sci USA 89:
7124-7128. White SE, Hebara L, Wessler SR (1994) Retrotransposons in
the flanking regions of normal plant genes: a role for copia- like elements in the evolution of gene structure and expression. Proc Natl Acad Sci USA 91: 11792-11796.
Zakian VA (1989) Structure and function of telomeres. Annu Rev Genet 23: 579-604.
364 Chromosome Research Vol 4 1996




















