
The Effects of Metaphoricity and Presentation Style on Brain Activation During Text Comprehension
16
This article was downloaded by: [University of Haifa Library] On: 26 January 2014, At: 12:35 Publisher: Routledge Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Metaphor and Symbol Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/hmet20 The Effects of Metaphoricity and Presentation Style on Brain Activation During Text Comprehension N. Mashal a & Miriam Faust a a Bar-Ilan University , Published online: 03 Feb 2010. To cite this article: N. Mashal & Miriam Faust (2010) The Effects of Metaphoricity and Presentation Style on Brain Activation During Text Comprehension, Metaphor and Symbol, 25:1, 19-33, DOI: 10.1080/10926480903538464 To link to this article: http://dx.doi.org/10.1080/10926480903538464 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms- and-conditions
Transcript of The Effects of Metaphoricity and Presentation Style on Brain Activation During Text Comprehension

This article was downloaded by: [University of Haifa Library]On: 26 January 2014, At: 12:35Publisher: RoutledgeInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Metaphor and SymbolPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/hmet20
The Effects of Metaphoricity andPresentation Style on Brain ActivationDuring Text ComprehensionN. Mashal a & Miriam Faust aa Bar-Ilan University ,Published online: 03 Feb 2010.
To cite this article: N. Mashal & Miriam Faust (2010) The Effects of Metaphoricity and PresentationStyle on Brain Activation During Text Comprehension, Metaphor and Symbol, 25:1, 19-33, DOI:10.1080/10926480903538464
To link to this article: http://dx.doi.org/10.1080/10926480903538464
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoever orhowsoever caused arising directly or indirectly in connection with, in relation to or arisingout of the use of the Content.
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Metaphor and Symbol, 25: 19–33, 2010Copyright © Taylor & Francis Group, LLCISSN: 1092-6488 print / 1532-7868 onlineDOI: 10.1080/10926480903538464
HMET1092-64881532-7868Metaphor and Symbol, Vol. 25, No. 1, Jan 2010: pp. 0–0Metaphor and Symbol
The Effects of Metaphoricity and Presentation Style on Brain Activation During Text Comprehension
Effects of Metaphoricity and Presentation StyleMashal and Faust N. Mashal and Miriam FaustBar-Ilan University
Evidence from neuroimaging studies suggests that the right hemisphere contributes significantly tothe processing of 2-word metaphoric expressions, whereas metaphoric sentences activate mainly lefthemispheric areas. Yet the pattern of brain activation during the processing of metaphors at the textlevel, beyond simple word pairs or sentences, is still unknown. We used functional magnetic reso-nance imaging to examine brain activation patterns while 15 healthy participants read either 4-linemetaphoric or nonmetaphorical texts. The findings indicate that both text types showed activation inthe left and right posterior and anterior superior temporal gyri. Surprisingly, metaphoric textsresulted in significantly lower activation than nonmetaphoric texts in the anterior superior temporalgyri. The results are interpreted as indicating that presentation style affects brain activation, and thattext processing may rely on a complicated interaction between metaphoricity (metaphoric, nonmeta-phoric) and text presentation style (poetic, prosaic).
An increasing number of studies focus on the neural substrate involved in metaphor as comparedto literal comprehension. For the past few decades, the distinction between the brain’s process-ing of metaphoric and literal language has been largely supported by findings from brain-damaged patients, suggesting that the right hemisphere (RH) contributes significantly to thecomprehension of metaphors (e.g., Winner & Gardner, 1977) and other aspects of figurative lan-guage (e.g., Brownell, Simpson, Bihrle, Potter, & Gardner, 1990; Brownell, Carroll, Rehak, &Wingfield, 1992; Burgess & Chiarello, 1996; Kaplan, Brownell, Jacobs, & Gardner, 1990;McDonald, 2000). This observation was further supported by a series of behavioral and brainimaging (functional Magnetic Resonance imaging [fMRI]) studies showing a unique RHinvolvement in the processing of two-word metaphoric expressions (Faust & Mashal, 2007;Mashal, Faust, Hendler, & Jung-Beeman, 2007, Mashal & Faust, 2008). However, this notion ofRH involvement did not remain unchallenged. Thus, recent brain imaging studies as well asstudies with brain-damaged persons revealed a more complicated picture suggesting predomi-nant left hemisphere (LH) involvement in metaphor processing (Giora, Zaidel, Soroker, Batori,& Kasher, 2000; Mashal, Faust, Hendler, & Jung-Beeman, 2009; Rapp, Leube, Erb, Grodd, &Kircher, 2007; Stringaris, Medford, Giampetro, Brammer, & David, 2007). Thus, the distinctionbetween metaphoric and literal language in terms of the associated brain bases is still underdebate.
Address correspondence to Dr. N. Mashal, School of Education, Bar-Ilan University, Ramat Gan, 52900, Israel.E-mail: mashaln@ mail.biu.ac.il
Dow
nloa
ded
by [
Uni
vers
ity o
f H
aifa
Lib
rary
] at
12:
35 2
6 Ja
nuar
y 20
14

20 MASHAL AND FAUST
More specifically, behavioral studies (e.g., Anaki, Faust, & Kravetz, 1998), studies withbrain-damaged patients (e.g., Rinaldi, Marangolo, & Baldassari, 2004; Winner & Gardner,1977), Positron Emission Tomography study with healthy participants (e.g., Bottini et al.,1994), recent fMRI studies (Mashal, Faust, & Hendler, 2005; Mashal et al., 2007), and a Tran-scranial Magnetic Stimulation (TMS) study (Pobric, Mashal, Faust, & Lavidor, 2008) sup-ported the involvement of the RH in metaphoric processing. In particular, a unique rightposterior superior temporal sulcus involvement was found when novel as compared to conven-tional two-word metaphoric expressions were compared (i.e., “autumn violins” vs. “ironfist”). These results were interpreted in term of the coarse semantic coding model (Beeman,1993, 1998). According to this view, the RH seems to weakly activate broad semantic fields,including peripheral, distant, and unusual semantic associations (coarse semantic coding),whereas the LH activates a selective range of closely related concepts and central aspects ofword meanings (fine semantic coding). The words in a metaphoric utterance (e.g., “autumnviolins”) are usually distantly related (“autumn” and “violins” are not close semantic associ-ates). According to the semantic coding model, RH activates broad semantic associates of theword autumn (e.g., wind, whistle, sounds, tune) that may overlap with words associated withviolin (e.g., sounds, tune). This overlap forms a new meaning (the background) of the meta-phoric utterance “autumn violins,” which could be interpreted as “the sounds of the violins area reminder of autumn’s wind” or alternatively, “the sounds of autumn’s wind are a reminder ofviolins.”
Consistent with this view, understanding metaphors might rely on the activation of abroader range of word meanings and thus involve right posterior superior temporal regions.However, results inconsistent with this view were obtained from more recent fMRI studies,showing that the processing of metaphoric sentences shows no RH involvement when bothnovel and conventional metaphoric sentences are compared with literal sentences (Rapp,Leube, Erb, Grodd, & Kircher, 2004; Mashal et al., 2009; Shibata, Abe, Terao, & Miyamoto,2007; Stringaris et al., 2007). An exception was found in a study reporting some evidence ofbilateral activation for processing of metaphoric sentences (Ahrens et al., 2007). Also, abehavioral study Faust and Weisper (2000) demonstrated that the metaphoric meaning ofmetaphoric sentences is available to both hemispheres, albeit more so in the LH than the RH.So, although the preponderance of evidence suggests only LH activity for metaphoriclanguage at the sentence level, usually when participants respond to target words presentedat the end of sentences, very little is known about hemispheric involvement in processingof metaphoric language at the supra-sentential levels (i.e., text level). Furthermore, very lit-tle is known about the neural correlates of texts that require cross-domain mappings withinthe text.
The majority of work investigating the brain bases of text processing has focused on theprocessing of isolated sentences, which involves the integration of semantic elements withinsentences. Text comprehension, on the other hand, depends on integrating the meaning of indi-vidual sentences into the context constructed from previous sentences in the narrative (Hough,1990) and constructing a coherent representation of the narrative. The exact neural substrateassociated with text integration is still unclear, given the different manipulations that have beenapplied in neuroimaging studies of discourse processing (e.g., consistent vs. inconsistent dis-course, coherent vs. incoherent discourse, intelligible vs. unintelligible speech, sentences vs.discourse, etc.). Thus, to elicit specific brain regions implicated in sentence integration, some
Dow
nloa
ded
by [
Uni
vers
ity o
f H
aifa
Lib
rary
] at
12:
35 2
6 Ja
nuar
y 20
14

EFFECTS OF METAPHORICITY AND PRESENTATION STYLE 21
studies used the inconsistent paradigm (i.e., coherent texts are compared to incoherent texts).For example, in one fMRI study of speech comprehension, participants listened to simplenarrative speech (Crinion, Lambon-Ralph, Warburton, Howard, & Wise, 2003). A direct com-parison between the narratives and their reverse versions, (which contained the same acousticfeatures as the speech but were unintelligible) resulted in increased activation in both temporallobes, with a robust activation at left anterolateral and ventral temporal areas. In another fMRIstudy (Ferstl, Rinck, & von Cramon, 2005) participants listened to short stories, half of whichcontained an (chronologically or emotionally) inconsistent word. Increased activation wasobserved in right anterior temporal lobe when participants detected the inconsistent word. Theauthors acknowledge the contribution of the left and right anterior temporal region to languageprocessing at the text level without being able to specify the exact role of the right anterior tem-poral lobe to text-level processing.
Jung-Beeman (2005) proposed a text integration network in bilateral anterior superiortemporal areas. A text integration network facilitates recognition of higher-order semantic rela-tions, thus supporting the linkage between two seemingly unrelated concepts and another singleconcept within the narrative. Accordingly, “‘Semantic integration’ supports message-levelinterpretation by computing the degree of semantic overlap among multiple semantic fields”(Jung-Beeman, 2005, p. 515). Take for example the stanza:
“And for a sick sun fields were cushionedand it feels softIt’s yellow tongue will elongateagainst the river’s scales.”
The utterance yellow tongue comprises two unrelated concepts (yellow, tongue). However,the previous text (And for a sick sun . . .) primes semantic fields related to sun (shine, yellow,hot, rays, etc.) that overlap with the following utterance yellow tongue. Thus, text integrationoccurs because the semantic fields of sun and yellow tongue overlap, supporting the emergenceof a message-level interpretation (i.e., “the ray of the sun is like a yellow tongue”). Furthermore,when the reader reads the last line (against the river’s scales) he or she is able to integrate itsmeaning with the previous context, that is, the surface of the river shines like scales due to thesun rays.
In addition to the findings from neuroimaging studies, evidence obtained from studies withbrain-injured patients suggests that the RH is also involved in discourse comprehension. Patientswith RH brain damage (RBD) are often described as having difficulties in processing at the dis-course level although they do not exhibit difficulties in recognizing words or comprehendingsentences. For example, Huber & Gleber (1982) reported that patients with RBD demonstrateddifficulty in comprehending and integrating sentences into a coherent narrative, that is, in gener-ating an overall narrative theme, especially when the theme was presented at the end of the para-graph. Delis, Wapner, Gardner, & Moses (1983) reported that patients with RBD showeddifficulty arranging five or six sentences into a coherent narrative story. Patients with RBDshowed difficulties in maintaining the topic of a conversation (Brownell & Martino, 1998).Patients with RH lesions in the right homologue of Wernicke’s area exhibited difficulties indrawing inferences in order to integrate sentences and maintain coherence (Beeman, 1993;Brownell, Potter, Birle, & Gardner, 1986). Patients with RBD perform worse in the integrationof narratives as compared to aphasic patients and controls. Also, participants with anterior RH
Dow
nloa
ded
by [
Uni
vers
ity o
f H
aifa
Lib
rary
] at
12:
35 2
6 Ja
nuar
y 20
14

22 MASHAL AND FAUST
brain damage confabulated and embellished story interpretations more often than other RBDgroups (Wapner, Hamby, & Gardner, 1981).
This study used unfamiliar stanzas, a grouping of lines within a poem, containing metaphoricexpressions. The juxtaposition of words within the metaphoric text differs from everyday prosaiclanguage and contains novel metaphoric uses. The stanzas were four-line poems displayed infour lines, one above the other, (as the stanzas appeared in their original poem book, from wherethey were taken). These texts were syntactically matched with newly created texts that did notcontain metaphoric use of language. Thus, the metaphoric text,
“The windows of my eyesare wide opento the streetand my interior is here chilly and dark”
was matched with a nonmetaphoric text,
The shutters of my houseare wide opento the streetand my home is here quiet and lighted.
This study addresses an issue that was not studied before: what are the brain bases of meta-phor comprehension at the text level. More specifically, although hemisphere involvement inprocessing metaphoric language at the word (two-word expressions) level and sentence level hasbeen relatively well studied (RH vs. LH, involvement, respectively), the processing of meta-phoric texts as compared to literal texts is still unclear.
We anticipated bilateral superior temporal lobe involvement in text comprehension regard-less of type of text (e.g., Friederici, Meyer, von Cramon, 2000; Xu, Kemeny, Park, Frattali, &Braun, 2005), with more pronounced activation in the LH than in the RH (e.g., Crinion et al.,2003; Ferstl, & von Cramon, 2001; Robertson et al., 2000). Based on the literature, posteriorparts were expected to be involved in more basic processes of text comprehension (e.g., lexicalaccess, syntax parsing; Just, Carpenter, Keller, Eddy, & Thulborn, 1996; Friederici et al., 2000;Robertson et al., 2000), whereas anterior parts should be engaged in text integration (Jung-Bee-man, 2005; Mason & Just, 2006). Because both types of texts have the same syntactic structure,we hypothesized that the anterior parts of the superior temporal cortices would show increasedactivation when integration demand was increased. One source of integration demand can be theuse of novel metaphoric expressions appearing in the metaphoric texts (“The windows of myeyes”), which involve the construction of distant semantic relations (Jung-Beeman, 2005).
METHOD
Participants
Fifteen neurologically normal participants (nine male), right-handed (yielding a laterality quo-tient of at least +80 on the Edinburgh Inventory (Oldfield, 1971), all native Hebrew speakers,participated in the study. Mean age was 25 (SD = 2.3, range = 22–28) years. All participants
Dow
nloa
ded
by [
Uni
vers
ity o
f H
aifa
Lib
rary
] at
12:
35 2
6 Ja
nuar
y 20
14

EFFECTS OF METAPHORICITY AND PRESENTATION STYLE 23
received a detailed explanation of the study and gave informed consent under a protocolapproved by the Tel-Aviv Medical Center Human Science Ethics Committee.
Stimuli
Stimuli included 28 text segments. Half of the text segments were stanzas (i.e., unit of sentenceswithin the poem) taken from Hebrew poetry and half were nonmetaphorical text segments. Thecriteria for selecting the metaphoric texts were the number of lines, length of line, the metaphor-icity of the text, and the unfamiliarity of the stanzas. Thus, metaphoric text segments wereselected if they consisted of four short lines (each line consisting of 2–5 words) without rhymes,containing at least one metaphoric phrase.
The nonmetaphorical text segments were constructed in a way that matched the metaphorictext in syntax, number of words in each line, word frequency, and part of speech of each word.The metaphoric expressions of the metaphoric text were replaced with literal expressions in thenonmetaphoric text.
Metaphoric phrase means an expression for which no literal meaning could be derived(e.g., “my past emerge”; “fields were cushioned”, in Table 1). Metaphoricity, semantic rela-tions, and familiarity of the metaphoric phrases and the texts were determined following a seriesof pretests.
Metaphoricity of the text was determined by two independent judges (one of them expert inliterature and poetry). The number of metaphoric phrases embedded in each text was counted:the mean of metaphoric phrases was 2.4 for the metaphoric texts (ranging from 1–4) and 0 forthe nonmetaphoric texts.
To determine the distant nature of the semantic relations of the metaphoric phrases, five judgeswere introduced to a list of word pairs that comprised the metaphoric phrases and asked to ratethe degree to which the words are semantically related on a 5-point scale ranging from 1 (closelyrelated) to 5 (distantly related). The mean score of the semantic relatedness was 4.2, indicating
TABLE 1Example of Four-Line Metaphoric Text Taken From Hebrew Poetry and a Matching
Nonmetaphoric Text
Metaphoric Text Nonmetaphoric Text
I look at him I look at himAnd see my past emerge And examine his young faceFrom his countenance From his photo albumThat refuses to grow up That is next to me
Target Past Face
And for a sick sun fields were cushioned And for a sick child pillows were paddedAnd it feels soft And he feels softIt’s yellow tongue will elongate His red tongue will elongateAgainst the river’s scales In front of the doctor’s eyes
Target Dance Boat
Note. Each text was followed by a probe word that could be related (e.g., “past,” “face”) to thetext segment or not (e.g., “dance,” “boat”).
Dow
nloa
ded
by [
Uni
vers
ity o
f H
aifa
Lib
rary
] at
12:
35 2
6 Ja
nuar
y 20
14

24 MASHAL AND FAUST
that the metaphoric phrases were composed of distantly related concepts (e.g., past and emergefor the expression “my past emerge”). These results suggest that the metaphoric stimuli, consist-ing of two semantically distinct domains, are likely to engage coarse semantic coding.
Seven additional judges rated the familiarity level of the metaphoric phrases on a 5-pointscale ranging from 1 (highly unfamiliar) to 5 (highly familiar). The mean score for familiaritylevel was 1.4, indicating that the metaphoric phrases were unfamiliar and therefore do not havepreestablished linkage as in conceptual metaphors (e.g., “LIFE IS A JOURNEY”; Lakoff &Johnson, 1980).
To assess the familiarity level of the entire text, eight judges were presented with the stanzasand were asked to rate the degree of familiarity of the text segments on a 3-point scale ranging 1(unfamiliar) to 3 (familiar). All judges indicated that all of the stanzas were unknown to them(M = 1 for the familiarity scale).
An additional group of nine judges who did not participate in the fMRI study rated the textsegments on a 7-point understandability scale ranging 1 (very easy to understand) to 7 (verydifficult to understand). Each of the judges was presented with the list of the metaphoric andnonmetaphoric text segments (order of segments was randomized) and asked to rate “how muchthe text is difficult to understand.” Metaphoric text segments were rated on average as lessunderstandable than nonmetaphoric text segments (metaphoric M = 3.15, SD = 1.15; nonmeta-phoric M = 1.49, SD = 0.49, respectively) on the understandability scale, t(13) = 4.8, p < .001).However, both types of text segments were rated under 4 (the middle point on the understand-ability scale), indicating that both were understandable.
Each text segment was followed by a probe word. Half of the words were taken from the textand represented the topic. Half of the text segments were followed by a probe word that had notappeared in the text and that did not represent the theme of the metaphoric or the nonmetaphorictext (for examples, see Table 1). Different probe words were used for each text and for themetaphoric and the nonmetaphoric versions of the text.
Task
Each participant was presented with both the metaphoric and the nonmetaphoric versions ofeach text and was asked to read and understand them silently and to decide as quickly as possi-ble whether the following probe word had appeared or had not appeared in the text by pressingone of two response buttons. They were told that the probes were words that best described thetheme of the text.
As this study focused on patterns of brain activation during text comprehension, the studywas designed in a way to allow for the analysis of brain activation during text reading but notduring the probe task.
Procedure
Each line of the four-line text segment was presented for 2100 ms, followed by 400 ms of blankscreen; thus each text segment was presented for 10 sec. At the end of each text segment afixation appeared for 2000 ms, followed by a probe word that was presented for 2000 ms. At theoffset of the probe word, a fixation point appeared for 12 sec (baseline). The experiment startedwith a 2000-ms instruction screen. Thus, each block (duration 10s) contained one text segment
Dow
nloa
ded
by [
Uni
vers
ity o
f H
aifa
Lib
rary
] at
12:
35 2
6 Ja
nuar
y 20
14

EFFECTS OF METAPHORICITY AND PRESENTATION STYLE 25
(metaphoric or nonmetaphoric) resulting in 28 blocks (14 blocks for each condition). Block (textsegment) and probe onset presentation was time locked to beginning of a time repetition (TR).Metaphoric and nonmetaphoric texts were presented in a mixed order such that correspondingmetaphoric and nonmetaphoric text were not presented back to back. Text segments were pre-sented using Presentation software (Neurobehavioral Systems, Inc.; http://nbs.neuro-bs.com)and projected via an LCD projector (EPSON MP 7200) onto a translucent tangent screen locatedon the head coil in front of the participant’s eyes. All text segments were presented in blackuppercase Arial font on a gray background.
Participants were trained prior to performing the fMRI experiment with text segments notincluded in the online study. Participants were sited in front of a computer and asked to readthree text segments (one was nonmetaphoric and two were metaphoric texts). Each segment wasfollowed by a probe word. Participants were asked to read silently and to understand the seg-ment and then to decide whether the following probe word had appeared or had not appeared inthe text. They were told that the word that appeared in the text is the word that best describes thetheme of the text.
It should be noted that brain activation was analyzed only while participants read the textsand not while they performed the task, in order to be consistent with more ecologically valid lan-guage use. In addition, participants were not told that some of the text segments were stanzastaken from poetry; thus, they were naïve to the type of text they read. On the other hand, to makeparticipants engaged with the task, we used a semantic judgment task (rather than a more super-ficial task, such as lexical decision). Deciding whether the probe word best described the themeof the text required that the participants read and understand the text at a level that encouragedthe integration of the lines into a coherent representation.
Image Acquisition
Imaging measurements were performed on a 3T GE G3 scanner (GE, Milwaukee, WI, USA)located at the Wohl Institute for Advanced Imaging in the Tel-Aviv Sourasky Medical Center.All images were acquired using standard quadrature head coil. A 3D T1 SPGR sequence wasperformed in order to allow for a volume statistical analysis of signal changes during the experi-ment and to facilitate later coordinate determinations. Acquisition parameters for the SpoiledGradient Recalled (SPGR) were TR/TE (time repetition/Echo Time) = 30/3 ms, flip angle of 12°with resolution of 0.97 × 0.97 × 1 mm3.
The functional T2*-weighted images were acquired using gradient echo echo-planar imagingpulse sequence (TR/TE/flip angle = 2000/35/90°) with field of view (FOV) of 200 × 200 mm2,and acquisition matrix dimensions of 64 × 64. Twenty-nine contiguous axial slices with 3.5-mmthickness and 0-mm gap were prescribed, based on a sagittal localizer, covering the whole brain.
Data Analysis
fMRI data were processed using the BrainVoyager software package (version 4.9; Brain Innova-tion, Maastricht, The Netherlands). Prior to statistical tests, raw data were examined for motionand signal artifacts. Preprocessing included 3D motion correction, linear trend removal, andhigh-pass filtering (filtering out the lowest three frequencies). The first nine functional volumes(i.e., 18 seconds), before signal stabilization, were excluded from the analysis. Functional
Dow
nloa
ded
by [
Uni
vers
ity o
f H
aifa
Lib
rary
] at
12:
35 2
6 Ja
nuar
y 20
14

26 MASHAL AND FAUST
images were then superimposed on 2D anatomical images and incorporated into 3D data setsthrough trilinear interpolation. The complete data set was transformed into Talairach space(Talairach & Tournoux, 1988).
Functional analysis was performed for each participant using a general linear model (Fristonet al., 1995). Task-related activation was defined in correlation to the two predictors of the usedprotocol (metaphoric, nonmetaphoric texts). Thus, the entire 10-sec segment presentation timewas included in the analysis (but, as pointed out previously, the probe presentation and the fol-lowing response were not included in the analysis). To account for a hemodynamic response,predictors were shifted with a lag of 6 sec from the onset of the text. This shift was selected tomaximize correlation between individual blood oxygen level–dependent signal intensity timecourse and predictors. Voxel-wise t-maps were constructed for each of the participants and thenwhole-brain random-effects analysis was applied to the general linear model of the group of 15participants. There were no significant clusters (greater than 500 mm3 at p < .001) demonstrat-ing different blood oxygen level–dependent signals for the metaphoric versus nonmetaphoricstimuli.
Region of interest (ROI) analysis in bilateral temporal and prefrontal areas that were involvedin processing both metaphoric and nonmetaphoric text, as compared to fixation blocks, was thenperformed. Our approach was to select the “best” similarly sized ROIs within each area (rightand left anterior and posterior temporal areas, left and right prefrontal areas), rather than pickingone area of strong activation and contrasting it with its direct homologue (because the two hemi-spheres are not exactly homologous). Our ROIs were functionally selected by calculating 3Dstatistical parametric maps for the group of participants using a general linear model in whichmetaphoric and nonmetaphoric text were positive predictors and the blank screen was a negativepredictor (with an expected lag of 6 seconds). Then time courses of statistically significant vox-els were collected such that cluster size of each of the following ROIs implicated in text integra-tion was approximately 220 mm3: left posterior superior temporal gyrus (PSTG; Talairachcoordinates: −51, –41, 6; p < .00001); right PSTG (Talairach coordinates: 57, –48, 9; p < .001);left anterior superior/middle temporal gyrus (ASTG; Talairach coordinates: −49, –10, −8); p <.00001); right ASTG (Talairach coordinates: 57, −1, −5); p < .0001; Figure 1); left inferior/mid-dle frontal gyrus (IFG; Talairach coordinates: −42, 10, 48), p < .0001), and right IFG (Talairachcoordinates: 45, 18, 27), p < .0001) . It should be noted that voxels in the left PSTG and leftASTG were collected at a higher level of p < .00001 because setting the threshold to a lowerlevel would have resulted in a cluster volume larger than 220 mm3, which could bias the effect atthis ROI. The right homologous clusters did not lie exactly symmetrically to their left ROIshomologues, probably due to morphological asymmetries within the temporal lobes. Thenbeta weights derived from each of these ROIs were entered into an analysis of variance(ANOVA).
RESULTS
Behavioral Results
Participants’ behavioral performance was recorded via Presentation. Accuracy of responses did notdiffer between the nonmetaphoric and the metaphoric texts (95.40% and 93.57%, respectively).
Dow
nloa
ded
by [
Uni
vers
ity o
f H
aifa
Lib
rary
] at
12:
35 2
6 Ja
nuar
y 20
14
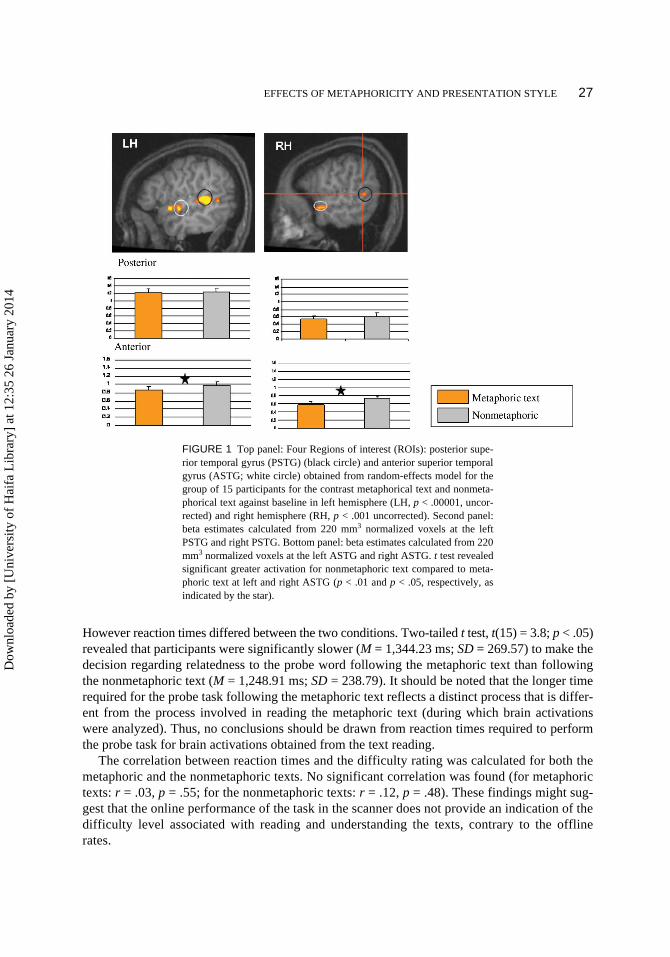
EFFECTS OF METAPHORICITY AND PRESENTATION STYLE 27
However reaction times differed between the two conditions. Two-tailed t test, t(15) = 3.8; p < .05)revealed that participants were significantly slower (M = 1,344.23 ms; SD = 269.57) to make thedecision regarding relatedness to the probe word following the metaphoric text than followingthe nonmetaphoric text (M = 1,248.91 ms; SD = 238.79). It should be noted that the longer timerequired for the probe task following the metaphoric text reflects a distinct process that is differ-ent from the process involved in reading the metaphoric text (during which brain activationswere analyzed). Thus, no conclusions should be drawn from reaction times required to performthe probe task for brain activations obtained from the text reading.
The correlation between reaction times and the difficulty rating was calculated for both themetaphoric and the nonmetaphoric texts. No significant correlation was found (for metaphorictexts: r = .03, p = .55; for the nonmetaphoric texts: r = .12, p = .48). These findings might sug-gest that the online performance of the task in the scanner does not provide an indication of thedifficulty level associated with reading and understanding the texts, contrary to the offlinerates.
FIGURE 1 Top panel: Four Regions of interest (ROIs): posterior supe-rior temporal gyrus (PSTG) (black circle) and anterior superior temporalgyrus (ASTG; white circle) obtained from random-effects model for thegroup of 15 participants for the contrast metaphorical text and nonmeta-phorical text against baseline in left hemisphere (LH, p < .00001, uncor-rected) and right hemisphere (RH, p < .001 uncorrected). Second panel:beta estimates calculated from 220 mm3 normalized voxels at the leftPSTG and right PSTG. Bottom panel: beta estimates calculated from 220mm3 normalized voxels at the left ASTG and right ASTG. t test revealedsignificant greater activation for nonmetaphoric text compared to meta-phoric text at left and right ASTG (p < .01 and p < .05, respectively, asindicated by the star).
Dow
nloa
ded
by [
Uni
vers
ity o
f H
aifa
Lib
rary
] at
12:
35 2
6 Ja
nuar
y 20
14
28 MASHAL AND FAUST
fMRI Results
Whole-brain analysis. A direct comparison between the two types of texts (metaphoricand nonmetaphoric) all over the brain did not reveal a cluster of activation more robust for onecondition over the other (greater than 500 mm3 at p < .001, uncorrected). Table 2 representsthe activation patterns associated with both types of texts vs. baseline (fixation), thresholded atp < .001 (corrected). As can be seen, a robust activation is observed for both types of texts overmultiple left-brain regions.
ROI analysis. Parameter estimates were analyzed by three-way repeated-measuresANOVA for the 15 participants with Region (anterior STG, posterior STG), Hemisphere (LH/RH), and Type of Text (metaphoric text, nonmetaphoric text) as factors. Results revealed a maineffect of Hemisphere, F(1, 14) = 42.65, p < .0001); and Type of Text, F(1, 14) = 7.27, p < .05),indicating higher activation in the LH than in the RH (p < .001) and stronger activation for non-metaphoric than metaphoric texts (p < .05). The Region × Hemisphere interaction was significant,F(1, 14) = 10.18, p < .001). Tukey’s least significant difference post hoc comparisons revealedstatistically significant higher activation in the left than the right ASTG (p < .05) and higher acti-vation in the left than the right PSTG (p < .001). The Region × Type of Text interaction was alsosignificant, F(1, 14) = 6.45, p < .05). Tukey’s least significant difference post hoc comparisonsrevealed statistically significant higher activation for the nonmetaphoric than the metaphoric textsonly for the ASTG (p < .001). The Hemisphere × Text interaction was not significant, F(1, 14) =.77; p = .39), nor was the Hemisphere × Type of Text × Region three-way interaction, F(1, 14) =.05; p =. 81).
TABLE 2All Regions, the Center of the Activation in Talairach Coordinates, t Value, and Number of Voxels
(p < .0001, Corrected) Derived From a Random-Effects Model, Showing Greater Activation for Reading Metaphoric and Nonmetaphoric Texts (vs. Fixation)
Region Hemisphere #voxels t-value Talaraich
Inferior frontal gyrus LH 246 6.8 −46,21,14Inferior/middile frontal gyrus LH 594 7.6 −42,11,28Inferior/middile frontal gyrus RH 471 8.3 43,18,27Dorsal Premotor cortex LH 765 7.8 −42,10,45Caudate nucleus LH 541 8.1 −14,8,10Caudate nucleus RH 742 7.3 14,12,7Thalamus (dorsomedial) LH 957 13.8 −5,−14,7Thalamus (dorsomedial) RH 889 7.6 11,−16,15Posterior superior/middle temporal gyrus LH 1219 16.9 −50,−41,4Posterior superior/middle temporal gyrus RH 163 6.4 8,−36,11Anterior superior temporal gyrus LH 924 8.5 −50,−9,−8Parahippocampal LH 950 8.6 −37,−38,−18Parahippocampal RH 304 7.5 32,−36,−16Occipital lobe LH >2000 16.9 −32,−84,−5Occipital lobe RH >2000 9.9 27,−87,−7Anterior insula RH 359 9.73 29,26,11SMA medial 565 9.3 1,0,56
Dow
nloa
ded
by [
Uni
vers
ity o
f H
aifa
Lib
rary
] at
12:
35 2
6 Ja
nuar
y 20
14

EFFECTS OF METAPHORICITY AND PRESENTATION STYLE 29
A second two-way repeated-measure ANOVA with Hemisphere (LH/ RH), and Type of Text(metaphoric text, nonmetaphoric text) as factors was performed in the left and right IFGs.Results indicated that the main effects of Hemisphere and Type of Text were not significant,F(1, 14) = 0.18, p = .67, and F(1, 14) = 0.22, p = .64, respectively. The Hemisphere × Type ofText interaction was also not significant, F(1, 14) = 0.02, p = .88).
DISCUSSION
This research focused on the role of the superior temporal cortices in reading and compre-hending four-line metaphoric and nonmetaphoric texts. The metaphoric texts were taken fromHebrew poetry and the nonmetaphoric texts were constructed to match the metaphoric texts insyntactic structure, lexical features of words, and number of words in each line. Both types oftexts elicited brain activation in bilateral brain regions, including PSTG and IFG, which wasmore pronounced in the LH than the RH (Table 2). The whole brain analysis revealed no sig-nificant difference in any brain region when the two types of texts were compared. A possibleexplanation for this finding could be that these brain areas supply the infrastructure for themore basic processes of text comprehension at the word level (e.g., lexical access and seman-tic activation) and at the sentence level (e.g., syntactic processing, semantic integration),which are common to both type of texts. According to Jung-Beeman (2005), semantic activa-tion takes place in the PSTG and middle temporal gyrus, whereas semantic integration isextended to bilateral anterior parts of the temporal cortex. Semantic activation refers to theinitial process of accessing semantic representations and first-order associations of the inputword, whereas semantic integration refers to the process that supports message-level interpre-tation (beyond the word level) and that detects, elaborates, and refines higher-order semanticrelations.
Furthermore, whereas it has been shown that when compared to literal expressions, the com-prehension of two-word novel metaphoric expressions involves RH brain regions (Faust &Mashal, 2007; Mashal et al., 2005; Mashal et al., 2007; Pobric et al., 2008) and also that whencompared with literal sentences, the processing of metaphoric sentences relies on LH brainmechanism (Mashal et al., 2009; Rapp et al., 2007; Shibata et al., 2007; Stringaris et al., 2007),our findings suggest that metaphoric language at the discourse level relies on both the LH andthe RH. These results are consistent with other brain-imaging studies that found common neuralsubstrates in bilateral superior temporal cortices and left IFG when listening to sentences(Humpheries et al., 2001), reading narrative (e.g., Just et al., 1996; Xu et al., 2005), or process-ing syntactic information during speech comprehension (e.g., Friederici et al., 2000). This resultis also consistent with some cognitive theories postulating that the same cognitive mechanismsunderlie the processing of both literal and nonliteral language (Glucksberg, 2001; Glucksberg &Keysar, 1990). However the lack of unique RH involvement in the processing of the metaphorictexts might result from the poor temporal resolution of the fMRI technique applied in this study.Future studies using fine temporal resolution techniques (e.g., event-related potential (ERP)magnetoencephalography) on the order of milliseconds may be more suitable for detecting dif-ferences between the two hemispheres during metaphoric text processing.
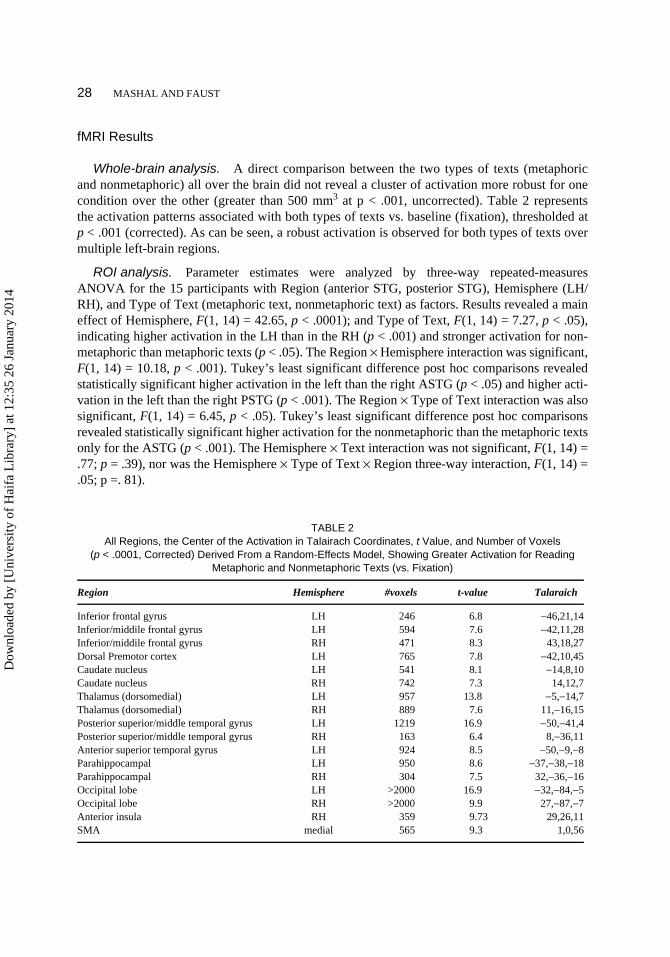
A second result emerging from the ROI analysis revealed stronger activation for the process-ing of nonmetaphoric texts as compared with metaphoric texts in both the left and right ASTGs
Dow
nloa
ded
by [
Uni
vers
ity o
f H
aifa
Lib
rary
] at
12:
35 2
6 Ja
nuar
y 20
14

30 MASHAL AND FAUST
(Figures 1 and 2). This finding suggests that the metaphoricity of the text did not exert extrademand on brain activity. This finding was somewhat surprising because increased brain activa-tion is usually correlated with increased task demand, which was expected for the metaphorictexts more than the nonmetaphoric texts. However, some comments made by the participantsfollowing the fMRI session suggest that a different source of difficulty may lie in the nonmeta-phoric texts. Following the fMRI session, some of the participants commented that they havenoticed that some of the texts seemed to them prosaic although they were displayed in four lines(poetic form), suggesting that they found the nonmetaphoric texts artificially divided into fourlines instead of being displayed naturally as a single, possibly longer, sentence. Thus, the pro-saic content displayed in a poetic form may have resulted in extra brain activation. To test thispossibility, we conducted a posttest to determine the extent of form−content compatibility forboth the metaphoric and the nonmetaphoric texts, which were both presented in four-line stan-zas. Thus, a group of 16 judges who did not participate in the fMRI study rated the form bywhich they preferred to read the texts: either as one long sentence (prosaic form) or as four shortsuccessive lines (poetic form). The judges were presented with pairs of each text (i.e., 28 × 2 =56), one above the other, in which the same text was presented both as a four-line text segmentand as a single line. They were asked to rate which form they prefer on a 6-point scale rangingfrom 1 (four lines, poetic form) to 6 (one line, prosaic form). The results indicated that meta-phoric texts were indeed preferred to be read in the poetic form (M = 2.8, SD = .8), whereas thenonmetaphoric texts were preferred to be read in the prosaic form (M = 3.7, SD = .0.6), t(13) =3.04, p < .01. These results suggest that although the nonmetaphoric texts were presented in a
FIGURE 2 Time course of percentage of signal change for readingthe text segments (Time Repetition [TRs] 1–5) followed by blank screen(TR 6), presentation of the probe word and making a decision (TR 7), andfixation period (rest) for nonmetaphoric texts (squares) and metaphorictexts (triangles). ASTG, anterior superior temporal gyrus; PSTG, poste-rior superior temporal gyrus.
Dow
nloa
ded
by [
Uni
vers
ity o
f H
aifa
Lib
rary
] at
12:
35 2
6 Ja
nuar
y 20
14

EFFECTS OF METAPHORICITY AND PRESENTATION STYLE 31
poetic form they were preferred to be read as one sentence, thus potentially exerting a source ofincompatibility between their presentation form and their content.
Thus, a possible explanation could be that—as opposed to the metaphoric texts, which wereconstructed so that the length of each line is compatible with the pattern of its sounds and hencewith its content—the nonmetaphoric texts were constructed in a content–form incompatiblemanner. Indeed, the pattern of the sounds frames the length of a line in a poem. In turn, thisdivision of lines organizes the poem into stanzas that lead to the construction of content units(Harshev, 2008). It might be that the nonmetaphoric texts were less compatible with theircontent. This preference toward a prosaic form (rather than a poetic form) might indicate that theorganization of the nonmetaphoric text into four lines is less expected and as a result can poten-tially alter the flow of reading. According to this explanation, linking together (integrating)successive lines to form a coherent representation might be relatively more demanding in thecase of the nonmetaphoric texts.
We thus suggest that the source of discrepancy between form and content during the process-ing of nonmetaphoric but not metaphoric texts may have resulted in extra activation in theASTG. This finding is in line with our prediction that increased integration demand results inincreased activation in the ASTG. The implications of the anterior temporal area for text com-prehension were found in fMRI studies that used different text manipulations. When writtensentences were compared to word lists, greater activation was observed in the anterior superiortemporal cortex for the sentences than for the word lists (Stowe et al., 1999). Convergent evi-dence from the auditory modality was also obtained when participants listened to sentences andword lists (Friederici et al., 2000). These two studies emphasize the selective role of the anteriortemporal cortex in the comprehension of sentence-level speech but not in the processing ofunstructured meaningful speech (word lists). These results, together with this study’s findings,suggest that the anterior superior temporal cortex plays an important role in sentence-level com-prehension. Future studies should test whether metaphoric texts displayed in an incompatibleform (one long sentence) would also result in an increased activation as compared with nonmet-aphoric texts displayed in the more common form (long sentences) in order to assess whetherthe form–content incompatibility effect revealed in the bilateral ASTG for the nonmetaphorictexts is the main source of enhanced brain activation.
In conclusion, our results suggest that bilateral posterior superior temporal areas are engagedwith basic stages of discourse processing regardless the type and the difficulty level of the text.In addition, nonmetaphoric texts resulted in stronger brain activation in the bilateral ASTG thandid metaphoric texts, probably due to increased integration demand resulting from the inconsis-tency between the poetic form of the text (four short lines) and its prosaic content. Thus, textprocessing may rely on a complicated interaction between type of context (metaphoric, nonmet-aphoric) and text presentation style (poetic, prosaic).
ACKNOWLEDGMENTS
Support for this work was provided by a grant from the U.S.–Israel Binational Science Foundation(grant 2003317) to Miriam Faust. We would like to thank Chen Kleinman for useful assistancein preparing the linguistic stimuli.
Dow
nloa
ded
by [
Uni
vers
ity o
f H
aifa
Lib
rary
] at
12:
35 2
6 Ja
nuar
y 20
14

32 MASHAL AND FAUST
REFERENCES
Ahrens, K., Liu, H., Lee, C., Gong, S., Fang, S., & Hsu, Y. (2007). Functional MRI of conventional and anomalousmetaphors in Mandarin Chinese. Brain and Language, 100, 163–171.
Anaki, D., Faust, M., & Kravetz S. (1998). Cerebral hemispheric asymmetries in processing lexical metaphors. Neuro-psychologia, 36, 353–362.
Beeman, M. (1993). Semantic processing in the right hemisphere may contribute to drawing inferences from discourse.Brain and Language, 44, 80–120.
Beeman, M. (1998). Coarse semantic coding and discourse comprehension. In M. Beeman & C. Chiarello (Eds.), Righthemisphere language comprehension: Perspectives from cognitive neuroscience (pp. 255–284). Mahwah, NJ:Lawrence Erlbaum Associates, Inc.
Bottini, G., Corcoran, R., Sterzi, R., Paulesu, E. S. P., Scarpa, P., Frackoviak, R. S. J., et al. (1994). The role of the righthemisphere in the interpretation of the figurative aspects of language: A positron emission tomography activationstudy. Brain, 117, 1241–1253.
Brownell, H. H., & Martino, G. (1998). Deficits in inference and social cognition: The effects of right hemisphere braindamage on discourse. In M. Beeman & C. Chiarello (Eds.), Right hemisphere language comprehension: Perspec-tives from cognitive neuroscience (pp. 309–328). Mahwah, NJ: Erlbaum.
Brownell, H. H., Carroll, J. J., Rehak, A., & Wingfield, A. (1992). The use of pronoum anaphora and speaker modein the interpretation of conversational utterances by right hemisphere damaged patients. Brain and Language, 43,121–147.
Brownell, H. H., Potter, H. H., Birle, A. M., & Gardner, H. (1986). Inference deficits in right brain-damaged patients.Brain and Language, 27, 310–321.
Brownell, H. H., Simpson, T. L., Bihrle, A. M., Potter, H. H., & Gardner, H. (1990). Appreciation of metaphoric alterna-tive word meanings by left and right brain damaged patients. Neuropsychologia, 28, 375–383.
Burgess, C., & Chiarello, C. (1996). Neurocognitive mechanisms underlying metaphor comprehension and other figura-tive language. Metaphor and Symbolic Activity, 11, 67–84.
Crinion, J. T., Lambon-Ralph, M. A., Warburton, E. A., Howard, D., & Wise, R. J. S. (2003). Temporal lobe regionsengaged during normal speech comprehension. Brain, 126, 1193–1201.
Delis, D. C., Wapner, W., Gardner, H., & Moses, J. A., Jr. (1983). The contribution of the right hemisphere to theorganization of paragraphs. Cortex, 19, 43–50.
Faust, M., & Mashal, N. (2007). RH advantage in processing novel metaphoric expressions: Behavioral data. Neuro-psychologia, 45, 860–870.
Faust, M., & Weisper, S. (2000). Understanding metaphoric sentences in the two cerebral hemispheres. Brain andCognition, 43, 186–191.
Ferstl, E. C., & von Cramon, D. Y. (2001). The role of coherence and cohesion in text comprehension: An event-relatedfMRI study. Cognitive Brain Research, 11, 325–340.
Ferstl, E. C., Rinck, M., & von Cramon, D. Y. (2005). Emotional and temporal aspects of situation model processingduring text comprehension: An event related fMRI study. Journal of Cognitive Neuroscience, 17, 724–729.
Friederici, A. D., Meyer, M., & von Cramon, D. Y. (2000). Auditory language comprehension: An event-related fMRIstudy on the processing of syntactic and lexical information. Brain and Language, 74, 289–300.
Friston, K., Holmes, A. P., Worsley, K., Poline, J. B., Frith, C., & Farckowiak, R. S. J. (1995). Statistical parametricmaps in functional imaging: A general linear approach. Human Brain Mapping, 2, 189–210.
Giora, R., Zaidel, E., Soroker, N., Batori, G., & Kasher, A. (2000). Differential effects of right-and left-hemisphere dam-age on understanding sarcasm and metaphor. Metaphor and Symbol, 15, 63–83.
Glucksberg, S. (2001). Understanding figurative language: From metaphors to idioms. Oxford, UK: Oxford UniversityPress.
Glucksberg, S., & Keysar, B. (1990). Understanding metaphorical comparison: Beyond similarity. PsychologicalReview, 97(1), 3–18.
Harshev, B. (2008).The history of Hebrew versification [in Hebrew]. Ramat-Gan, Israel: Bar-Ilan University Press.Hough, M. S. (1990). Narrative comprehension in adults with right and left hemisphere brain-damage: Theme organiza-
tion. Brain and Language, 38, 253–277.Huber, W., & Gleber, J. (1982). Linguistic and nonlinguistic processing of narratives in aphasia. Brain and Language,
16, 1–18.
Dow
nloa
ded
by [
Uni
vers
ity o
f H
aifa
Lib
rary
] at
12:
35 2
6 Ja
nuar
y 20
14

EFFECTS OF METAPHORICITY AND PRESENTATION STYLE 33
Humphries, C., Willard, K., Buchsbaum, B., & Hickok, G. (2001). Role of anterior temporal cortex in auditory sentencecomprehension: An fMRI study. Neuroreport, 12, 1749–1752.
Jung-Beeman, M. (2005). Bilateral brain processes for comprehending natural language. Trends in Cognitives Sciences,9, 512–518.
Just, M. A., Carpenter, P. A., Keller, T. A., Eddy, W. F., & Thulborn, K. R. (1996). Brain activation modulated bysentence comprehension. Science, 274, 114–116.
Kaplan, J. A., Brownell, H. H., Jacobs, J. R., & Gardner, H. (1990). The effects of right hemisphere damage on the prag-matics interpretation of conventional remarks. Brain and Language, 38(2), 315–333.
Lakoff, G., & Johnson, M. (1980). Metaphors we live by. Chicago: Chicago University Press.Mashal, N., & Faust, M. (2008). Right hemisphere sensitivity to novel metaphoric relations: Application of the Signal
Detection Theory. Brain and Language, 104(2), 103–112.Mashal, N., Faust, M., & Hendler, T. (2005). The role of the right hemisphere in processing nonsalient metaphorical
meanings: Application of principal components analysis to fMRI data. Neuropsychologia, 43, 2084–2100.Mashal, N., Faust, M., Hendler, T., & Jung-Beeman, M. (2007). An fMRI investigation of the neural correlates underly-
ing the processing of novel metaphoric expressions. Brain and Language, 100, 115–126.Mashal, N., Faust, M., Hendler, T., & Jung-Beeman, M. (2009). An fMRI study of processing novel metaphoric
sentences. Laterality, 14(1), 30–54.Mason, R. A., & Just, M. A. (2006). Neuroimaging contributions to the understanding of discourse processes. In
M. Traxler & M. A. Gernsbacher (Eds.), Handbook of psycholinguistics (pp. 765–799). Amsterdam: Elsevier.Oldfield, D. J. (1971). The assessment of handedness: The Edinburgh Inventory. Neuropsychologia, 9, 97–113.Pobric, G., Mashal, N., Faust, M., & Lavidor, M. (2008). The causal role of the right cerebral hemisphere in processing
novel metaphoric expressions taken from poetry: A TMS study. Journal of Cognitive Neuroscience, 20, 170–181.Rapp, A. M., Leube, D. T., Erb, M., Grodd, W., & Kircher, T. T. J. (2004). Neural correlates of metaphor processing.
Cognitive Brain Research, 20, 395–402.Rapp, A. M., Leube, D. T., Erb, M., Grodd, W., & Kircher, T. T. (2007). Laterality in metaphor processing: Lack of
evidence from functional magnetic resonance imaging for the right hemisphere theory. Brain and Language, 100,142–149.
Rinaldi, M. C., Marangolo, P., & Baldassari, F. (2004). Metaphor comprehension in right brain-damaged patients withvisuo-verbal and verbal material: A dissociation (re)considered. Cortex, 40, 479–490.
Robertson, D. A., Gernsbacher, M. A., Guidotti, S. J., Robertson, R. R. W., Irwin, W., Mock, B. J., et al. (2000).Functional neuroanatomy of the cognitive process of mapping during discourse comprehension. PsychologicalScience, 11, 255–260.
Shibata, M., Abe, J., Terao, A., & Miyamoto, T. (2007). Neural mechanisms involved in the comprehension of meta-phoric and literal sentences: An fMRI study. Brain Research, 1166, 92–102.
Stowe, L. A., Paans, A. M. J., Wijers, A. A., Zwartz, F., Mulder, G., & Vaalburg, W. (1999). Sentence comprehensionand word repetition: A positron emission tomography investigation. Psychophysiology, 36, 786–801.
Stringaris, A. K., Medford, N. C., Giampetro, V., Brammer, M. J., & David, A. S. (2007). Deriving meaning: Distinctneural mechanisms for metaphoric, literal and non-meaningful sentences. Brain and Language, 100, 150–162.
Talairach, J., & Tournoux, P. (1988). Co-planar stereotaxic atlas of the human brain. Stuttgart, Germany: TheimeVerlag.
Wapner, W., Hamby, S., & Gardner, H. 1981. The role of the right hemisphere in the apprehension of complex linguisticmaterials. Brain and Language, 14, 15–33.
Winner, E., & Gardner, H. (1977). The comprehension of metaphor in brain damaged patients. Brain, 100, 717–729.Xu, J., Kemeny, S., Park, G., Frattali, C., & Braun, A. (2005). Language in context: Emergent features of word,
sentence, and narrative comprehension. NeuroImage, 25, 1002–1015.
Dow
nloa
ded
by [
Uni
vers
ity o
f H
aifa
Lib
rary
] at
12:
35 2
6 Ja
nuar
y 20
14




















