
tese 10.12 - Universidade Federal do Ceará
172
UNIVERSIDADE FEDERAL DO CEARÁ FACULDADE DE MEDICINA DEPARTAMENTO DE CIRURGIA PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS MÉDICO-CIRÚRGICAS LUCIANA FUJIWARA AGUIAR RIBEIRO PRÉ-CONDICIONAMENTO NUTRACÊUTICO COM MISTURAS DE ÓLEOS ÔMEGA 3, 6 E 9 PARA ATENUAR AS CARACTERÍSTICAS NEUROINFLAMATÓRIAS DO TRANSTORNO DO ESPECTRO AUTISTA INDUZIDAS PELO ÁCIDO VALPRÓICO EM UM MODELO ANIMAL FORTALEZA 2020
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of tese 10.12 - Universidade Federal do Ceará

UNIVERSIDADE FEDERAL DO CEARÁ
FACULDADE DE MEDICINA
DEPARTAMENTO DE CIRURGIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS MÉDICO-CIRÚRGICAS
LUCIANA FUJIWARA AGUIAR RIBEIRO
PRÉ-CONDICIONAMENTO NUTRACÊUTICO COM MISTURAS DE ÓLEOS
ÔMEGA 3, 6 E 9 PARA ATENUAR AS CARACTERÍSTICAS
NEUROINFLAMATÓRIAS DO TRANSTORNO DO ESPECTRO AUTISTA
INDUZIDAS PELO ÁCIDO VALPRÓICO EM UM MODELO ANIMAL
FORTALEZA
2020

LUCIANA FUJIWARA AGUIAR RIBEIRO
PRÉ-CONDICIONAMENTO NUTRACÊUTICO COM MISTURAS DE ÓLEOS ÔMEGA
3, 6 E 9 PARA ATENUAR AS CARACTERÍSTICAS NEUROINFLAMATÓRIAS DO
TRANSTORNO DO ESPECTRO AUTISTA INDUZIDAS PELO ÁCIDO VALPRÓICO EM
UM MODELO ANIMAL
Tese submetida ao Programa de Pós- Graduação Stricto Sensu em cirurgia do Departamento de Cirurgia da Faculdade de Medicina da Universidade Federal do Ceará, como requisito parcial para a obtenção do grau de Doutor em Ciências Médico-Cirúrgicas. Orientador: Prof. Dr. Paulo Roberto Leitão Vasconcelos Co-orientador: Prof.ª Dra. Lissiana Magna Vasconcelos Aguiar
FORTALEZA
2020


LUCIANA FUJIWARA AGUIAR RIBEIRO
PRÉ-CONDICIONAMENTO NUTRACÊUTICO COM MISTURAS DE ÓLEOS ÔMEGA
3, 6 E 9 PARA ATENUAR AS CARACTERÍSTICAS NEUROINFLAMATÓRIAS DO
TRANSTORNO DO ESPECTRO AUTISTA INDUZIDAS PELO ÁCIDO VALPRÓICO EM
UM MODELO ANIMAL
Tese submetida ao Programa de Pós- Graduação Stricto Sensu em cirurgia do Departamento de Cirurgia da Faculdade de Medicina da Universidade Federal do Ceará, como requisito parcial para a obtenção do grau de Doutor em Ciências Médico-Cirúrgicas.
Aprovada em: ___/___/_____
BANCA EXAMINADORA
_____________________________________________
Prof. Dr. Paulo Roberto Leitão de Vasconcelos
Universidade Federal do Ceará- UFC
_____________________________________________
Profa. Dra. Lissiana Magna Vasconcelos Aguiar
Universidade Federal do Ceará - UFC
_____________________________________________
Profa. Dra. Cristiana Libardi Miranda Furtado
Universidade Federal do Ceará - UFC
_____________________________________________
Prof. Dr. Gislei Frota Aragão
Universidade Estadual do Ceará - UECE
_____________________________________________
Prof. Dr. Fábio Miyajima
Fundação FIOCRUZ- CEARÁ

À Deus, por sempre me ajudar acreditar que
podemos ter um mundo melhor onde as pessoas
possam se respeitar e amar mutuamente e que a
fé na busca de uma viória torna o impossível
acontecer.

Aos meus pais ERNANI INÁCIO DE
AGUIAR (In memoriam) e HELENA
FUJIWARA AGUIAR pelos ensinamentos e
credibilidade a mim concebidos. O que sou
devo a vocês.
Ao meu marido e companheiro FABIANO
CARNEIRO RIBEIRO, pelo apoio, dedicação,
compreensão e amor por todos os momentos
quando precisei estar ausente você se fez
presente com nossos filhos.
A minha filha e amiga ÁGATHA FUJIWARA
AGUIAR RIBEIRO pelo amor, carinho e
compreensão, um dia quero ser igual a você.
E a você que foi o grande responsável na
idealização deste estudo – BERNARDO
FUJIWARA AGUIAR RIBEIRO, meu filho,
meu anjo, minha inspiração.

AGRADECIMENTOS
Ao Prof. Dr PAULO ROBERTO LEITÃO DE VASCONCELOS, Coordenador do
Programa de Pós-Graduação Strictu Sensu do Departamento de Cirurgia da Faculdade de
Medicina da Universidade Federal do Ceará, meu orientador, pela paciência, compreensão e
oportunidade de realizar este trabalho que abrilhantou meus olhos e minha vida acadêmica.
À Drª LISSIANA MAGNA VASCONCELOS AGUIAR, Professora e Vice-
coordenadora do Curso de Medicina da Universidade Federal do Ceará Campus Sobral, minha
co-orientadora, agradeço pelo carinho e oportunidade de abrir as portas do Laboratório de
Fisiologia e Neurociência mostrando o caminho para finalizar minha pesquisa.
À Drª CRISTINA LIBARDI MIRANDA FURTADO, Programa de Pós-Graduação
Strictu Sensu do Departamento de Cirurgia da Faculdade de Medicina da Universidade Federal
do Ceará, pela oportunidade de ter realizado parte das análises no Laboratório de Oncologia
Experimental (LOE) do NPDM, por toda atenção, carinho e dedicação à minha pesquisa.
À Drª MIRNA MARQUES BEZERRA BRAYNER, professora do Curso de
Medicina Campus Sobral, por todo apoio, incentivo, carinho, amizade e colaboração na
correção da minha tese.
Ao Dr GISLEI FROTA ARAGÃO, professor Adjunto do Curso de Medicina da
Universidade Estadual do Ceará, pelos esclarecimentos e ajuda no manejo das análises
comportamentais, pela compreensão estatística dos dados e todo apoio durante à pesquisa.
Ao aluno e amigo MATEUS ARAGÃO ESMERALDO do Curso de Medicina
Campus Sobral, pela árdua dedicação à esta pesquisa, pela disponibilidade diária de domingo a
domingo sem medir esforços do início ao fim, pela participação e coerência nas fases mais
turbulentas onde se tornava possível a realização das etapas, sem sua ajuda acho que não teria
conseguido chegar ao fim.
Aos meus irmãos Ernani Inácio de Aguiar Junior, Thiago Rodrigo de Aguiar e
Jefferson Felipe de Aguiar pelo apoio, incentivo, que desde à infância acreditavam e me faziam
ser uma pessoa melhor.
Às secretárias do programa de Pós-Graduação em Cirurgia, MARIA LUCIENE
VIEIRA DE OLIVEIRA e MAGDA GOMES FONTENELE, secretárias do Programa de Pós-
Graduação Strictu Sensu do Departamento de Cirurgia da Faculdade de Medicina da
Universidade Federal do Ceará, por todo o apoio, atenção, estímulo que me foi concebido
durante todo este percurso.

Ao técnico do laboratório de Fisiologia e Neurociências FRANCISCO JOSÉ
GOMES, pela disponibilidade em ajudar em todas fases desta pesquisa, pelo apoio emocional
onde seu “vai dar certo” foi fundamental. Muito obrigada.
Aos alunos da Iniciação Científica, em especial a aluna SILVIA FLÁVIA ALVES
DE FREITAS e NICKOLAS SOUSA SILVA, pela dedicação, participação e amizade que
criamos durante toda a fase experimental e aos alunos que tiveram uma participação essencial:
Daniel Salmito Chaves, Kaio Júlio César de Sousa Nogueira, Mateus, Bárbara Barboza de
Alencar, Marília Rocha Silva,
A médica veterinária do Biotério da UFC/Sobral, ALANA GODINHO, por toda a
assistência prestada principalmente nos momentos de desespero, pelo carinho e incentivo que
tudo iria dar certo.
Ao técnico do Núcleo de Bioprospecção e Experimentação Molecular Aplicada
(UNINTA), ANTÔNIO MATEUS GOMES PEREIRA, pelo auxílio e paciência nas leituras
das análises neuroquímicas.
A pós-doutoranda CLEANE GOMES MOREIRA e doutoranda FRANCISCA
VALÉRIA BEZERRA SAMPAIO MARQUES que contribuíram nas atividades mais árduas
da pesquisa e através de suas lindas palavras acalentavam minhas aflições e horas de desespero.
A Universidade Federal do Ceará (UFC) pela oportunidade de estar afastada por 1
ano das atividades acadêmicas e tornar possível a realização de toda pesquisa em um curto
período de tempo.
Enfim, a todos que de alguma maneira estiveram presentes apoiando através de
ações, palavras ou atitudes.

“Do lado de fora, olhando para dentro, você
nunca poderá entendê-lo. Do lado de dentro,
olhando para fora, você jamais conseguirá
explicá-lo. Isso é autismo.” (Autism Topics)

RESUMO
Universidade Federal do Ceará. Pós-Graduação Stricto-Sensu do Departamento de Cirurgia, Faculdade de Medicina, (Grau de Doutor em Ciências Médico-Cirúrgicas). Nutracêutico com misturas de óleos ômega 3, 6 e 9 atenua as características neuroinflamatórias e anti-oxidantes do transtorno do espectro autista induzidas pelo ácido valpróico em modelo experimental. Luciana Fujiwara Aguiar Ribeiro. Março 2020.
O Transtorno do Espectro Autista (TEA) é uma desordem do desenvolvimento neurológico, com
anormalidades no sistema imunológico, nas funções metabólicas e comportamentais. Nos últimos 15
anos, um grande interesse tem sido dado ao potencial papel dos ácidos graxos poliinsaturados ômega-3
na compreensão da patogênese e tratamento de distúrbios do neurodesenvolvimento e psiquiátricos. No
presente estudo misturas de óleos ômega 3, 6 e 9, com diferentes fontes de ômega-3 foram usadas
conforme suas características estruturais: Mix-1 (ácido graxo linolênico–ALA), Mix-2 (ALA (35%) +
ácido graxo eicosapentaenoico (EPA–39%) + ácido graxo docosahexaenóico (DHA–26%) e Mix-3
(ALA-84% + DHA-16%) com objetivo de avaliar seus efeitos neuroinflamatórios, anti-oxidantes e
comportamentais, bem como a expressão gênica em ratos wistar macho e fêmea submetidos a indução
de autismo com uma única dose de ácido valpróico (AVP), 400mg/kg, via sub cutânea, no 14ª dia após
nascimento. Os mixs de óleos foram administrados por gavagem durante 25 dias contínuo a partir do
15ª dia de nascidos. Durante este período foram realizados os testes comportamentais – campo aberto;
imersão em cauda; performance da natação e por último o teste do rota rod. No 55ª dia pós-nascimento
foram dissecadas quatro áreas cerebrais (hipocampo, córtex pré-frontal, amígdalas e cerebelo) para
determinar o grau de peroxidação lipídica, concentração de nitrito e glutationa redutase.
Adicionalmente, foi avaliada a expressão gênica do hipocampo no grupo tratado que apresentou
melhores resultados (Mix-3) utilizando os genes Dnmt3a e Achd2. Os resultados mostraram que tanto
em animais machos como fêmeas, o grupo AVP demonstrou um aumento significante no tempo de
latência a sensibilidade nociceptiva comparado com o salina. Os grupos Mix-1, Mix-2 e Mix-3 com
indução de TEA por AVP diminuiram esta resposta nociceptiva. Regressão nas habilidades motoras
foram avaliadas na performance da natação nos grupos AVP de ambos os sexos, mas não foi revertido
por nenhum tratamento administrado. Na atividade locomotora, apenas as fêmeas do grupo AVP
sofreram uma redução significante, onde mais uma vez os mix ricos em ômega-3 (Mix-1 e Mix-2)
melhoraram sua coordenação motora. Aumento expressivo do estresse oxidativo se deu nas áreas
hipocampo e córtex pré-frontal nos machos, sendo reduzido com ação dos Mix-2 e Mix-3 dos grupos
AVP. A alta produção de óxido nítrico (NO) foi presenciada na área cerebelar do grupo AVP dos animais
machos, podendo representar um tipo de processo inflamatório. O tratamento com AVP nos machos não
representou aumento significante da expressão gênica para área hipocampo dos genes Dnmt3a e Achd2.
Os achados deste estudo sugerem que a mistura dos óleos ricos em ômega-3 foi favorável à redução do
estresse oxidativo (Mix-2 e Mix-3) e efeitos na redução de sintomas do TEA.
Palavras-chave: Autismo. Ácidos graxos. Ômega-3. Expressão gênica.

ABSTRACT
Nutraceutical with mixtures of omega 3, 6 and 9 oils attenuate neuroinflammatory and anti-oxidant characteristics of autism spectrum disorder induced by valproic acid in an experimental model. Federal University of Ceará Stricto Sensu post-graduation. Department of Surgery, Medical School, (Degree of Doctor of in Medical-surgery Sciences). Luciana Fujiwara Aguiar Ribeiro. March 2020.
Autism spectrum disorder (ASD) is a neurodevelopmental disorder with abnormalities in the immune
system, metabolism and behavioral functions. Over the past 15 years, great interest has been given to
the potential role of Omega-3 polyunsaturated fatty acids to understand the pathogenesis of some
diseases and to treat neurodevelopmental and psychiatric disorders. In the present study, mixtures of
omega 3, 6, and 9 oils with different types of Omega-3 were used according to their structural
characteristics: Mix-1 (linolenic fatty acid – ALA), Mix-2 (ALA – 35%) + eicosapentaenoic fatty acid
(EPA – 39%) + fatty acid docosahexaenoic acid (DHA – 26%) and Mix-3 (ALA – 84%; DHA – 16%)
to evaluate their neuroinflammatory and antioxidant effects in the animals’ behavior, as well as the gene
expression in male and female Wistar rats submitted to ASD induction with a single dose of valproic
acid (VPA) sc 400mg/kg on the 14th day after birth. The oil mixes were administered by gavage for 25
days continuously starting on the 15th day of birth; water and ration were made available
ad libitum. Then, behavioral tests were performed - open field; tail immersion; swimming performance
and rotarod. On the 55th day after birth, four brain areas (hippocampus, prefrontal cortex, amygdalae
and cerebellum) were dissected to determine the degree of lipid peroxidation, nitrite concentration and
glutathione reductase levels. Additionally, the hippocampus gene expression of the control 0.9% saline,
VPA, Mix-3 and Mix-3 + VPA groups was evaluated using the Dnmt3a and Achd2 genes. Both in male
and female groups VPA showed a significant increase in latency compared to saline, improving their
pain response after treatment with Mix-1, Mix 2 and Mix-3 in the VPA groups. Deficit in motor skills
integration was noted in swimming performance in VPA groups of both sexes. In locomotor activity,
only the females of the VPA group suffered a significant reduction, where once again the mixes rich
in Omega-3 (Mix-1 and Mix-2) were able to restore their locomotor activity. A significant increase in
oxidative stress occurred in the hippocampus and prefrontal cortex in males, being reduced with the
action of Mix-2 and Mix -3 in the VPA groups. No antioxidant response from glutathione reductase was
observed in this study. A high production of NO nitric oxide was noted in the cerebellum of the VPA
group of male animals, which may represent a consequence of inflammatory process. Treatment with
VPA in males did not represent a significant increase in gene expression for the hippocampal area of
the Dnmt3a and Achd2 genes. Our findings suggest that the mixture of oils rich in omega-
3 was favorable to the reduction of oxidative stress (Mix-2 and Mix-3) and effects in reducing
behavioral alterations of ASD in rats.
Keywords: Autism. Fatty acids. Omega 3. Gene expression.

LISTA DE FIGURAS
Figura 1 – Manifestações comportamentais do TEA concomitantes ao neurodesenvolvimento ................................................................................................................................ 23
Figura 2 – Representação esquemática de como a estimulação de mastócitos e micróglia pode levar a vários efeitos que contribuem para a inflamação cerebral, a patogênese e os sintomas do autismo. .............................................................................................. 39
Figura 3 – A história da genética do TEA de 1975 a 2015. ..................................................... 44 Figura 4 – Estrutura dos ácidos graxos insaturados. ................................................................ 54 Figura 5 – Processos de dessaturação e elongação do ômega-3 e ômega-6 e consequente
geração de compostos bioativos. ............................................................................ 55 Figura 6 – Animal com filhotes no biotério. ............................................................................ 62 Figura 7 – Produção dos Mix-1, Mix-2 e Mix-3. ..................................................................... 63 Figura 8 – Esquema do delineamento experimental ................................................................ 64 Figura 9 – Soluções e composição com as respectivas fontes de w3 (MIX 1) ........................ 65 Figura 10 – Soluções e composição com as respectivas fontes de w3 (MIX 2) ...................... 65 Figura 11 – Soluções e composição com as respectivas fontes de w3 (MIX 3) ...................... 66 Figura 12 – Quantidade de solução lipídica infundida diariamente ......................................... 68 Figura 13 – Análise comportamental dos animais pelo Teste Campo Aberto. ........................ 70 Figura 14 – Análise comportamental dos animais pelo método Imersão em Cauda. .............. 71 Figura 15 – Análise comportamental dos animais pelo método performance da natação. ...... 72 Figura 16 – Representação esquemática do teste comportamental Rotarod. Dois dias antes do
teste, os animais foram adaptados ao aparato por meio da permanência neste por 300 segundos com baixa velocidade inicial, gradualmente crescente (4 a 15 RPM). No dia do teste, foi utilizada a velocidade crescente de 4 a 40 RPM com duração máxima de 300 segundos. Foram avaliados os seguintes parâmetros: Tempo de Latência (tempo até a primeira queda) e Tempo ................................. 73
Figura 17 – Representação esquemática da determinação do estresse oxidativo por meio da obtenção de TBARS. ............................................................................................ 75
Figura 18 – Representação esquemática da determinação do estresse oxidativo por meio da obtenção de Nitrito. .............................................................................................. 76
Figura 19 – Representação esquemática da determinação do estresse oxidativo por meio da Concentração de Glutationa Reduzida ................................................................. 77
Figura 20 – Análise da atividade motora no Teste do Campo Aberto em ratos (machos) realizado no 24ª dia pós-natal, induzidos ao autismo tratados por 25 dias com mix de óleos e uma única dose de ácido valproico. .................................................... 82
Figura 21– Análise da atividade motora no Teste do Campo Aberto em ratos (fêmeas) realizado no 24ª dia pós-natal, induzidos ao autismo tratados por 25 dias com mix de óleos e uma única dose de Ácido Valproico. .................................................. 83

Figura 22 – Análise da imersão de cauda em ratos machos realizado no 30ª dia pós-natal, induzidos ao autismo tratados por 25 dias com Mix de óleos e uma única dose ácido valproico. .................................................................................................... 84
Figura 23 – Análise da atividade motora no Teste de Imersão em Cauda em ratos fêmeas realizado no 30ª dia pós-natal, induzidos ao autismo tratados por 25 dias com mix de óleos e uma única dose de Ácido valproico. ................................................... 85
Figura 24 – Análise da imersão de cauda em ratos machos realizado no 32ª dia pós-natal, induzidos ao autismo tratados por 25 dias com Mix de óleos e uma única dose ácido valproico ..................................................................................................... 86
Figura 25 – Análise da atividade motora no Teste de Imersão em Cauda em ratos fêmeas realizado no 32ª dia pós-natal, induzidos ao autismo tratados por 25 dias com mix de óleos e uma única dose de ácido valproico. .................................................... 87
Figura 26 – Análise da locomoção através do teste de RotaRod em ratos machos realizado no 50ª dia pós-natal, induzidos ao autismo tratados por 25 dias com Mix de óleos e única dose de ácido valproico. ............................................................................. 88
Figura 27 – Análise da atividade motora no Teste Rota Road em ratos fêmeas realizdo no 50ª dia pós-natal, induzidos ao autismo tratados por 25 dias com mix de óleos e uma única dose de ácido valpróico. ............................................................................. 89
Figura 28 – Níveis de Peroxidação Lipídica (TBARS) nas áreas cerebrais de ratos machos realizados no 55ª dia pós-mortem, tratados por 25 dias com Mix de óleos e uma única dose de Ácido Valproico. ........................................................................... 90
Figura 29 – Níveis de Peroxidação Lipídica (TBARS) nas áreas cerebrais de ratos fêmeas realizados no 55ª dia post-mortem, tratados por 25 dias com Mix de óleos e uma única dose de Ácido valpróico. ............................................................................ 91
Figura 30 – Concentrações de Nitrito/Nitrato nas áreas cerebrais de ratos machos realizados no 55ª dia pós-mortem, tratados por 25 dias com Mix de óleos e uma única dose de Ácido Valproico. ............................................................................................. 92
Figura 31 – Concentração de Nitrito (NO2) / Nitrato (NO3) nas áreas cerebrais de ratos machos realizados no 55ª dia pós-mortem, tratados por 25 dias com Mix de óleos e uma única dose de Ácido Valpróico. ................................................................. 93
Figura 32 – Concentrações de GSH (µ/g de tecido) nas áreas cerebrais de ratos machos realizados no 55ª pós-mortem, tratados por 25 dias com Mix de óleos e uma única dose de Ácido Valpróico. ..................................................................................... 94
Figura 33 – Níveis de GSH (µ/g de tecido) nas áreas cerebrais de ratos fêmeas realizados no 55ª dia pós-mortem, tratados por 25 dias com Mix de óleos e uma única dose de Ácido Valpróico. .................................................................................................. 95
Figura 34 – Padronização dos genes alvo e referência. A - eficiência dos primers, B - curva de melting. ................................................................................................................. 96
Figura 35 – Avaliação da expressão relativa diferencial dos genes Dnmt3a e Chd2. .............. 96

LISTA DE QUADROS
Quadro 1 – Classificação dos óleos ômega. ............................................................................. 54 Quadro 2 – Protocolo de Tratamento Experimental (FÊMAS - 96 animais) e MACHOS (121
animais), tipo de dosagens analisada. ................................................................... 67 Quadro 3 – Protocolo de Tratamento Experimental (Droga + Dieta) ...................................... 68

LISTA DE TABELAS
Tabela 1 - Predição da expressão gênica por tecido, sequência de primers utilizados, alvos e tamanhos dos produtos amplificados esperados. ..................................................... 79

LISTA DE ABREVIATURAS E SIGLAS
5-LOX 5-lipoxigenase
AA Ácido Graxo Araquidônico
ADDM Monitoramento de Autismo e Deficiências do Desenvolvimento
AG Ácidos graxos
AGE Ácidos Graxos Essenciais
AGPI Ácidos Graxos Poliinsaturados
AL Ácido Graxo Linoléico
ALA Ácido Graxo Linolênico
APA Associação Americana de Pediatria
AVA Ácido Valpróico Anticonvulsivante
AVP Ácido Valpróico
CDC Centro para Controle de Doenças
CEPA Comitê de Ética em Pesquisa Animal
CEUA Comitê de Ética no Uso de Animais
CID Classificação Internacional das Doenças
CNV Variação do Número de Cópias
COX Ciclo-oxigenase
CPF Cortex Pré-Frontal
DD Atraso no Desenvolvimento
DG Dia Gestacional
DHA Ácido Docosahexaenóico
DPN Dia Pós-Natal
DSM Manual do Diagnóstico e Estatístico de Transtornos Mentais
DZ Dizigóticos
EPA Ácido Eicosapentaenóico
GSH Glutationa
GWA Associação Genômica Ampla
HAC Acetilase de Histonas
HC Hipocampo
HDAC Desatilase de Histonas
ID Deficiência Intelectual
LC-PUFA Ácido Graxo Poliinsaturado de Cadeia Longa

LD Dificuldade de Aprendizagem
LOX Lisil Oxidase
MIA Ativação Imune Maternal
MZ Monozigóticos
NO Óxido Nítrico
OMS Organização Mundial da Saúde
PUFA Ácido Graxo Polinsaturado
ROS Espécie Reativa de Oxigênio
RPM Rotações Por Minuto
RT-PCR Reação em Cadeia da Polimerase com Transcrição Reversa
SBCAL Sociedade Brasileira de Ciência em Animais de Laboratório
SNC Sistema Nervoso Central
SNP Short Tandem Repeats
TBARS Substâncias Reativas ao Ácido Tiobarbitúrico
TDHA Transtorno do Déficit de Atenção com Hiperatividade
TEA Transtorno do Espectro Autista
TID Transtorno Invasivo do Desenvolvimento
VPA Ácido Valpróico

SUMÁRIO
1 INTRODUÇÃO ................................................................................................................ 19
1.1 Transtorno do Espectro Autista ................................................................................. 19
1.1.1 Conceito de TEA ................................................................................................ 21
1.1.2 Epidemiologia do TEA ...................................................................................... 24
1.1.3 Hipóteses Etiológicas ......................................................................................... 27
1.1.3.1 Teorias Psicogenéticas ................................................................................ 29
1.1.3.2 Mecanismos Epigenéticos ........................................................................... 31
1.1.4 Características/ Manifestações Clínicas ............................................................. 33
1.1.4.1 Déficit de Socialização ............................................................................... 33
1.1.4.2 Déficit de Comunicação/Linguagem .......................................................... 35
1.1.4.3 Padrões de Comportamentos ...................................................................... 36
1.1.4.3.1 Interesses Restritos e Estereotipados ....................................................... 36
1.1.5 Características Neuroquímicas ........................................................................... 37
1.1.5.1 Neuroinflamação ......................................................................................... 37
1.1.5.2 Mecanismos Moleculares e Genéticos ........................................................ 41
1.2 Ácido Valpróico ........................................................................................................ 46
1.2.1 Efeitos Teratogênicos ......................................................................................... 47
1.2.2 Mecanismo de Ação ........................................................................................... 48
1.2.3 Uso em Modelos Animais .................................................................................. 49
1.2.4 Áreas cerebrais estudadas ................................................................................... 52
1.3 Ácidos Graxos Poliinstaurados .................................................................................. 53
1.4 Testes Comportamentais em Modelo Animal versus Sinais do TEA ....................... 58
2 OBJETIVO ....................................................................................................................... 61
2.1 Objetivo Geral ........................................................................................................... 61
2.2 Objetivos Específicos ................................................................................................ 61
3 MÉTODO ......................................................................................................................... 62
3.1 Aspectos éticos e Animais ......................................................................................... 62
3.2 Drogas e soluções ...................................................................................................... 63
3.3 Delineamento Experimental ...................................................................................... 64
3.3.1 Distribuição dos Grupos de Estudo .................................................................... 64
3.3.2 Tratamentos dos grupos experimentais .............................................................. 65
3.3.3 Divisão dos Grupos de Estudo e Análises Experimentais .................................. 66
3.4 Testes Comportamentais ........................................................................................... 68
3.4.1 Atividade Locomotora (Campo Aberto) ............................................................ 69
3.4.2 Teste de imersão na cauda .................................................................................. 70

3.4.3 Teste de Performance de Natação ...................................................................... 71
3.4.4 Teste do rotarod .................................................................................................. 72
3.4.5 Dissecação das áreas cerebrais e descarte dos animais ...................................... 73
3.5 Avaliação Neuroquímica ........................................................................................... 74
3.5.1 Determinação da peroxidação lipídica por medição do conteúdo de substâncias reativas do ácido tiobarbitúrico (TBARS) ........................................................................ 74
3.5.2 Determinação da Concentração de Nitrito nas Áreas Cerebrais e no Sangue .... 75
3.5.3 Determinação da Concentração de Glutationa Reduzida GSH) ......................... 76
3.6 Análise de Expressão Gênica .................................................................................... 77
3.6.1 Desenho e análise dos primers ........................................................................... 77
3.6.2 Expressão Gênica do Hipocampo ...................................................................... 78
3.6.3 Extração de RNA total de hipocampo ................................................................ 78
3.6.4 Transcrição reversa obtenção de cDNA PCR quantitativo em Tempo Real (RT-qPCR) 79
3.7 Análises estatísticas (comportamentos, estresse oxidativo, GSH e Nitrito) .............. 80
3.7.1 Análise estatística (Expressão Gênica) .............................................................. 80
4 RESULTADOS ................................................................................................................. 81
4.1 Análises Comportamentais em MACHOS e FÊMEAS ............................................ 81
4.1.1 Avaliação da Atividade Locomotora - Teste do Campo Aberto ........................ 81
4.1.2 Avaliação da Atividade Locomotora - Teste de Imersão em Cauda .................. 83
4.1.3 Teste de Performance da Natação ...................................................................... 85
4.1.4 Avaliação da Atividade Locomotora -Teste de Rota-rod ................................... 87
4.2 Análises Neuroquímicas em MACHOS e FÊMEAS ................................................ 89
4.2.1 Determinação dos Níveis de Peroxidação Lipídica (TBARS) ........................... 89
4.2.2 Determinação da Concentração de Nitrito (NO2) / Nitrato (NO3) ..................... 91
4.2.3 Determinação das Concentrações de Glutationa Reduzida (GSH) .................... 93
4.3 Expressão Gênica por RT-qPCR ............................................................................... 95
5 DISCUSSÃO .................................................................................................................... 97
6 CONCLUSÃO ................................................................................................................ 110
REFERÊNCIAS ..................................................................................................................... 111
APÊNDICE A - VALORES BRUTOS REFERENTES AOS RESULTADOS DOS TESTES COMPORTAMENTAIS E TESTES NEUROQUÍMICOS .................................................... 141
APÊNDICE B - VALORES BRUTOS REFERENTES AOS RESULTADOS DA ANÁLISE PCR ........................................................................................................................................ 169
ANEXO A- PARECER DE APROVAÇÃO DO COMITÊ DE ÉTICA ................................ 171

19
1 INTRODUÇÃO
1.1 Transtorno do Espectro Autista
O transtorno do espectro do autismo (TEA) é um distúrbio generalizado prototípico
do desenvolvimento resultante do desenvolvimento cerebral anormal. O TEA constitui dois
sintomas comportamentais principais, incluindo prejuízo nas interações sociais e na
comunicação, e comportamentos repetitivos restritos, diagnosticados em idade precoce no
desenvolvimento (American Psychiatric Association, 2013). O distúrbio é frequentemente
acompanhado por anormalidades no processamento sensorial (Rogers et al., 2003), problemas
de sono (SCHRECK et al., 2004), ansiedade e depressão (STRANG et al., 2012), hiperatividade
(AMAN e LANGWORTHY, 2000), agressões ou comportamentos autolesivos (SINGH et al.,
2006), convulsões (VOLKMAR e NELSON, 1990) e problemas alimentares ou digestivos
(Martins et al., 2008), deficiência intelectual (DI), epilepsia, dificuldade de controle motor,
transtorno de deficit de atenção e hiperatividade (TDHA), tiques (GILLBERG 2010;
MORENO-DE-LUCA et al., 2013).
O TEA é heterogêneo e é comumente comórbido com outros distúrbios e do
desenvolvimento neurológico (LEYFER et al., 2006). Como se pode esperar, essa condição
causa grandes dificuldades às famílias acometidas, pois pode levar a aflições sociais,
ocupacionais e outras funcionais (American Psychiatric Association, 2013).
Nas últimas décadas pesquisas identificaram vários fatores de risco que levam ao
TEA, que podem ser classificados em anormalidades genéticas, alterações epigenéticas e
agressões ambientais (GEPNER; FERON, 2009). Os fatores de risco genéticos assumem a
forma de mutações monogênicas, polimorfismo de nucleotídeo único (SNP) e variantes de
número de cópias (CNVs). Não é de surpreender que muitos dos genes implicados tenham sido
associados à modulação do desenvolvimento cerebral e organização cortical, formação e função
de sinapses e neurotransmissão. Por outro lado, os insultos ambientais implicados no TEA
compreendem a exposição pré-natal a infecções ou produtos químicos virais, incluindo rubéola
(CHESS et al., 1978), citomegalovírus (YAMASHITA et al., 2003), timerosal (BERNARD et
al., 2002), talidomida (STROMLAND et al., 2002).
Com base nos fatores de risco identificados e na possível etiologia do TEA, foram
criados modelos animais para imitar e entender os mecanismos patológicos subjacentes às
anormalidades comportamentais desse distúrbio. Dois tipos principais de modelos animais, a
lesão e os modelos genéticos, tornaram-se muito usados. Os modelos de lesão incluem infecção

20
pré-natal (vírus da doença de Borna neonatal), lesão de amígdala neonatal e exposição pré-natal
ao ácido valpróico (AVP).
Uma alternativa para o tratamento adjuvante na tentativa de amenizar e/ou diminuir
os possíveis sintomas do TEA pode vir a ser a inclusão de alguns nutrientes isolados, ou
combinados, administrados em doses farmacológicas, classificados como nutracêuticos ou
fármaconutrientes. Estes nutrientes têm sido utilizados como modificadores da resposta
biológica exercendo ações antioxidantes (ácidos graxos monoinsaturados ω-9), ações anti-
inflamatórias (ácidos graxos poliinsaturados ω-3) e ações pró-inflamatórias (ácidos graxos
poliinsaturados ω-6). Quando a relação entre ω-9 e ω-6 é maior que 1 (ω-9/ω-6 > 1), ocorre
função anti-oxidante. Quando a relação entre ω-6 e ω-3 é menor que 4 (ω-6 /ω-3<4), uma função
anti-inflamatória é desencadeada (VASCONCELOS, 2013).
O termo nutracêutico, ou fármaconutriente, faz referência a um alimento, ou parte
de um alimento, capaz de proporcinar benefícios à saúde, incluindo prevenção e/ou tratamento
de doenças (ANDLAUER; FÜRST, 2002). Esses produtos podem incluir desde nutrientes
isolados, suplementos dietéticos na forma de suplementos orais e cápsulas, até produtos
beneficamente projetados, e alimentos processados tais como cereais, sopas e bebidas. Podem
ser classificados como nutracêuticos as fibras dietéticas, ácidos graxos poliinsaturados,
proteínas, peptídios, aminoácidos ou cetoácidos, minerais, vitaminas antioxidantes e outros
antioxidantes. Em suma, é uma substância de ocorrência natural, com evidente efeito benéfico
à saúde, que faz parte como ingrediente de alimentos específicos, alimentos funcionais ou
suplementos alimentares (MORAES; COLLA, 2006).
A busca de alternativas para o tratamento dos TEAs levantou a hipótese de que
anormalidades do metabolismo de ácidos graxos possam estar relacionadas a transtornos
mentais. Os ácidos graxos mais investigados são os w-3 por serem considerados componentes
essenciais das membranas das células nervosas e fundamentais para o desenvolvimento
neurológico (LEVANT et al., 2006).
Os ácidos graxos essenciais são ácidos graxos poliinsaturados que o ser humano
corpo não produz e é necessário para o desenvolvimento normal e funcionamento do cérebro e
do sistema imunológico (YEHUDA, 2005). Existem vários tipos principais de ácidos graxos
essenciais. Peixe e os frutos do mar são as principais fontes de ácido eicosapentaenóico (EPA)
e ácido docosahexaenóico (DHA), que são tipos de ácidos graxos ômega-3 de cadeia longa.
Linhaça, grãos e soja são fontes de ácidos alfa-linolênico (ALA), que são ácidos graxos ômega-
3 de cadeia curta. O ALA pode ser convertido no corpo em EPA e DHA, mas este é um processo
ineficiente; estima-se que menos de 5% do ALA em humanos seja convertido em EPA ou DHA

21
(BRENNA, 2002). Óleos vegetais são uma fonte de ácido linolênico (LA), que é um tipo de
ácido graxo ômega-6; gorduras animais também são uma fonte de ômega-6. O consumo ideal
de ômega-6 para ômega-3 deve ser de 1: 1 a 4: 1 (SIMOPOULOS, 2002). A maioria das dietas
modernas excede em muita essa proporção, que tem sido associado à vulnerabilidade a
deficiências e distúrbios de saúde mental (SIMOPOULOS, 2003).
A base de evidências da dose e tipo ideais de suplementação de ácidos graxos
ômega-3 para tratar TEA está atualmente ausente. Embora geralmente se acredite que a ingestão
diária de ômega-3 não deve exceder três gramas, são necessários estudos de dose (ranging) para
avaliar a ingestão ideal, o que pode diferir com base em características como peso, idade e
gravidade do distúrbio (RICHARDSON, 2006).
1.1.1 Conceito de TEA
A expressão “autismo” foi utilizada pela primeira vez por Eugene Bleuler em 1911,
para designar a perda de contato com a realidade com dificuldade ou impossibilidade de
comunicação, comportamento esse que foi, por ele, observado em pacientes diagnosticados
com quadro de esquizofrenia (AJURIAGUERRA et al., 1977). Durante as primeiras décadas
do século passado, autismo referia-se à tendência do esquizofrênico de se isolar do mundo
social e colocar-se em um mundo à parte (ABRAMOVITCH et al., 2001). Bleuler incluiu o
autismo dentro dos quatro critérios principais para a identificação desta afecção mental, que
ficaram conhecidos como os quatro ‘A(s)’ de Bleuler: alucinações, afeto incongruente,
ambivalência, autismo (BLEULER, 1991).
Somente entre os anos de 1950 e 1960 a causa biológica do autismo começou a ser
discutida. Até então, acreditava-se que a afecção era resultado da falta de proximidade dos pais
com seus filhos. Essa compreensão contribuiu para o surgimento do termo “mães-geladeira”,
que expressava a falta de afeto e a frieza dos pais (VERHOEFF et al., 2013).
O autismo começou a ser tratado de forma oficial em 1952, após a publicação da
primeira edição do Manual Diagnóstico e Estatístico de Transtornos Mentais (DSM- I)
(American Psychiatric Association, 1952), que reuniu a nomenclatura padrão e critérios para o
diagnóstico de transtornos mentais. Nas primeiras edições desse manual os sintomas
característicos do autismo foram classificados dentro da esquizofrenia infantil, sem o
diagnóstico diferenciado (American Psychiatric Association, 1952).
O autismo apareceu pela primeira vez somente na terceira edição do Manual
Diagnóstico e Estatístico de Transtornos Mentais (DSM-III-R), revisado em 1989 (NADON et

22
al., 2011). No ano de 1994, com a atualização do DSM, em sua quarta edição (DSM-IV, 1994)
e com a Classificação Estatística de Doenças e Problemas Relacionados à Saúde - 10a Revisão
(OMS, 1993) o autismo foi classificado como um Transtorno Invasivo do Desenvolvimento
(TID) cujo portador poderia atingir níveis mais graves no desempenho de atividades como:
redução do contato visual, restrição em mostrar, pegar ou utilizar objetos, atividades repetitivas
e comportamentos estereotipados, movimentação ou torção de mãos e dedos, agitação corporal
ou dificuldade/ausência de fala (DSM, 2000; NEMEROFF et al, 2013).
Na quinta edição do DSM, lançado em 2013, o autismo passou a ser definido como
um distúrbio neurocomportamental complexo, caracterizado por vários graus de deficiência
incluindo interação social, agilidade de pensamento, mudanças peculiares de linguagem e/ou
percepção atrasada, prejuízo na comunicação verbal e não verbal e inabilidade em responder a
certos estímulos apresentando um comportamento restrito e repetitivo (MCPARTLAND et al.,
2012). A partir desta atualização, o DSM-V substituiu o sistema multicategórico usado
anteriormente por uma única dimensão do diagnóstico – Transtorno do Espectro Autista (DSM,
2013; MCPARTLAND et al., 2012).
O diagnóstico no TEA ainda se concentra basicamente nas avaliações
comportamentais devido à ausência de marcadores biológicos para identificá-lo (ECKER;
ROCHA, 2010; WOLFERS, 2015; BUITELAAR, 2015). Mesmo com os avanços recentes na
capacidade de diagnosticar o transtorno precocemente, na maioria dos casos esse diagnóstico
acontece depois dos três anos de idade (CDCP, 2013). A Associação Americana de Pediatria
(APA) sugere realizar o acompanhamento do desenvolvimento de todas as crianças até 24
meses, pois é a partir do segundo ano de vida que os comportamentos característicos do TEA
surgem e esses são capazes de auxiliar no diagnóstico (SCHIEVE et al., 2012).
Estudos sobre a base neurológica do TEA antes dos três anos de idade é decisivo
para o tratamento, pois é nessa fase que ocorrem os cuidados primários da infância e é nesse
período que os resultados do tratamento podem ser melhores (SCHIEVE et al., 2012). As
manifestações comportamentais do autismo englobam uma ampla gama de alterações clínicas
e variabilidade na sintomatologia. A Figura 1 ilustra alguns dos sintomas que afetam indivíduos
com autismo. Com a falta de homogeneidade das manifestações comportamentais e a
consciência dos diferentes graus de acometimento e seguramente a interação entre múltiplos
fatores etiológicos, tornou-se justificável criar o termo “transtorno do espectro autista” (TEA)
(DSM-V, 2013).
É possível avaliar como critério de classificação o nível de gravidade e
comprometimento do TEA em diferentes estados (leve, moderado e grave) (LEE; THOMAS;

23
LEE, 2015). Para validar o diagnóstico, os indivíduos precisam apresentar no mínimo seis
características comportamentais (FRAZIER et al., 2012; TAHERI; PERRY, 2012), sendo pelo
menos dois sintomas na área de interação social, um na área de comunicação e um na área de
comportamentos estereotipados (SILVA; MULICK, 2009).
Figura 1 – Algumas das manifestações comportamentais do TEA concomitantes ao
neurodesenvolvimento
Fonte: https://autismo2.wordpress.com/caracteristicas-2/
Manifestações do TEA incluem prejuízos na comunicação e interação social,
anomalias sensoriais, comportamentos repetitivos e níveis variados de inteligência, invalidez

24
total. Juntamente com esses sintomas principais, distúrbios psiquiátricos ou neurológicos
concomitantes são comuns em pessoas com autismo, das quais hiperatividade distúrbios de
atenção (como déficit de atenção/ distúrbios de hiperatividade (TDHA), ansiedade, depressão
e epilepsia são bastantes prevalentes (LORD et al., 2020).
Várias formas de TEA foram descritas, como a síndrome de Asperger ou o autismo
do tipo Kanner (1949), revelando que o TEA é um distúrbio altamente heterogêneo,
provavelmente com várias causas subjacentes. Trabalho científico intenso foi realizado nos
últimos anos para entender a origem potencial do TEA, acreditando que esse distúrbio surge de
fatores genéticos e ambientais, especialmente aqueles que influenciam o desenvolvimento fetal
e o início da vida (LAI et al., 2013). Embora o TEA tenha se mostrado altamente herdável
(estimativas recentes de 38 a 54%), várias metanálises destacaram que existem causas pré-
natais não genéticas do TEA, abrindo a porta para novos estudos para investigar esses
mecanismos (HALLMAYER et al., 2011).
Aproximadamente 10% dos casos de TEA estão relacionados a distúrbios de
etiologia genética, como síndrome do X frágil, esclerose tuberosa e distúrbio de Rett. Apoiando
a idéia de heterogeneidade do TEA, as mutações genéticas únicas representam apenas 1-2%
dos casos de TEA (ABRAHAMS et al., 2008), com a maioria dos casos permanecendo
idiopáticos. Mutações identificadas por estudos genéticos revelaram que alguns genes afetados
estão envolvidos no desenvolvimento do cérebro desde o útero até a infância (WON et al.,
2013). Os genes envolvidos na formação de sinapses ou na conectividade cerebral (por
exemplo, fmr1, mecp2, shank3, tsc, neuroligin e cntnap2) têm sido repetidamente ligados ao
TEA (ODDI et al 2013; SHCHEGLOVITOV et al., 2013).
1.1.2 Epidemiologia do TEA
A prevalência de indivíduos com TEA sofreu um aumento substancial desde sua
descoberta. Por exemplo, nas décadas de 60 e 70 a frequência era de 4/10.000 indivíduos
diagnosticados com autismo em todo o mundo (FOMBONNE et al., 2005). Nas últimas duas
décadas está prevalência tem aumentado significativamente. Em 2000, a Rede de
Monitoramento de Autismo e Deficiências no Desenvolvimento (ADDM) do Centro de
Prevenção e Controle de Doenças (Center for Disease Control) estimou a incidência de TEA
em 1 em cada 150 crianças (SHARMA et al., 2018).
Em 2006, a incidência de TEA aumentou para 1 em 110 crianças e, em 2008,
aumentou novamente para 1 em 88 crianças. Em 2012, de acordo com o Centro de Prevenção

25
e Controle de Doenças, cerca de 1,5% (1/68 indivíduos) das crianças nos Estados Unidos
possuem diagnóstico de TEA (CHRISTENSEN et al., 2016), corroborando com Elsabbagh
(2012) onde cita a mesma incidência em uma proporção de 1:4 de homens e mulheres afetados.
Enquanto estudos realizados na Ásia e Europa identificaram uma prevalência populacional
média entre 1 e 2% de pessoas com o distúrbio (CHRISTENSEN et al., 2016).
O National Health Interview Survey (NHIS) nos EUA, divulgou sua mais recente
taxa de prevalência e relatou um novo recorde, citando que o TEA pode ser encontrado em até
1 em cada 36 crianças, em uma pesquisa que levantou dados de 2014 a 2016 (ZABLOTSKY;
BLACK; BLUMBERG, 2017). Pensa-se que esta proporção seja a mesma em todos os
contextos raciais, étnicos ou socioeconômicos, no entanto, existem variações de gênero.
Possivelmente o aumento no número de casos de autismo deva-se as melhores condições de
diagnóstico precoce, mudanças na prática de identificação da doença e incentivos financeiros
subsidiados pelo governo para realizar diagnósticos (BLUMBERG et al., 2013; LEE et al.,
2015).
Shriver (2005), relata que o autismo ocorre igualmente em diferentes grupos raciais,
étnicos e sociais, mas existem três grupos distintos que apresentam um risco mais elevado de
ocorrência de TEA: o sexo masculino, sendo cerca de quatro vezes mais afetado que o sexo
feminino, os irmãos de crianças com TEA, sendo que as famílias com uma criança autista
apresentam uma taxa recorrência 2 a 8%, e ainda as pessoas com outros distúrbios do
desenvolvimento como sejam o atraso mental, a Esclerose Tuberosa e o Síndrome do X Frágil.
Dados epidemiológicos e com base em estudos nas redes públicas, relatam que o
autismo é mais comum em homens do que nas mulheres, com taxas relatadas variando de 2:1 a
5:1, com uma estimativa de 4:1 no Global 2010 Estudo do ónus da doença (LOOMES et al.
2017; BRUGHA et al., 2016). Bedfor et al, (2016) também cita que o TEA é cerca de 4-5 vezes
mais comum em indivíduos do sexo masculino (1 em 42) do que no sexo feminino (1 em 89).
O gênero masculino indica um fator de risco poderoso e bem estabelecido para distúrbios do
espectro autista. Até o momento, definir a causa pela qual o autismo acomete mais indivíduos
do sexo masculino tem sido uma tarefa desafiadora (FRAZIER et al., 2014).
A Genética é um papel proeminente no desenvolvimento do TEA. Estudos que
avaliaram a prevalência de TEA descobriram que em gêmeos idênticos, se um gêmeo tiver
TEA, o outro terá 36% a 95% de chance de também ter TEA. Em gêmeos não idênticos, se uma
criança tem TEA, a chance de o outro gêmeo ter o mesmo distúrbio cai para 0 a 30%
(HALLMAYER et al., 2011; ROSENBERG et al., 2009). Os irmãos de crianças com TEA têm
um risco de 2 a 8% de desenvolver também o distúrbio, e isso aumenta para 12 a 20% se a

26
criança infligida apresentar déficits em um a dois dos três domínios prejudicados no autismo
(BOLTON et al., 1994).
TEA possui um grande componente herdável. Bebês nascidos de famílias com um
irmão mais velho afetado têm 20% de chance de desenvolver autismo (OZONOFF et al., 2011).
Concordância as taxas entre gêmeos monozigóticos, gêmeos dizigóticos e irmãos são de 30 a
99%, 0 a 65% e 3 a 30%, respectivamente, com uma herdabilidade global estimada de 0,7-0,8
(ROSENBERG et al., 2009; HALLMAYER et al., 2011; COLVERT et al., 2015). No entanto,
populações estudadas na Escandinávia mostraram uma herdabilidade estimada mais baixa de
0,5-0,6% (SANDIN et al., 2014).
Avaliando a literatura é possível observar que poucos estudos exploraram
alterações comportamentais nas fêmeas, em parte, por apresentarem sintomas e alterações
menos frequentes (comportamentais, neuroanatômicas e expressão molecular) evidentes do
autismo (KIRSTEN et al., 2012; LAI; LOMBARDO; BARON-COHEN et al., 2014;
SCHWARZ et al., 2011), ou ainda pelo fato de fêmeas possuírem manifestações clínicas e
comportamentais difícil de serem diagnosticados (MANDY; CHILVERS; COMORBIDITY,
2012). O comprometimento em pessoas do sexo feminino com autismo parece apresentar-se de
uma maneira muito menos grave do que o descrito no sexo masculino (MANDY; CHILVERS;
COMORBIDITY, 2012). Estudos têm mostrado que as fêmeas de várias espécies geralmente
apresentam aumento da resposta imune e da resistência à infecção em comparação aos machos
(SCHWARZ; BILBO, 2012). Estas diferenças têm sido atribuídas em grande parte, às ações
imunomoduladoras de hormônios esteróides sexuais.
Pesquisas que investigam a origem biológica da diferença entre os sexos no TEA
enfatizam os níveis circulantes de esteróides gonadais no útero (AUYEUNG et al., 2009,
BARON-COHEN et al., 2002) ou acúmulos de fatores de risco genéticos que tem valores
diferenciais em meninos versus meninas (JACQUEMONT et al., 2014; GOCKLEY et al.,
2015). A teoria do Cérebro Masculino Extremo postula que muitos traços autistas são a
masculinidade levada ao ponto de disfunção, ou seja, sistematização excessiva, baixa empatia,
falta de vínculo e falta de habilidades sociais (BARON-COHEN et al., 2002). Por outro lado,
vários estudos apoiam a afirmação de que as mulheres carregam uma carga maior de mutação
genética antes de sucumbir ao TEA, sugerindo que elas estão protegidas (JACQUEMONT et
al., 2014; ROBINSON et al., 2013; GOCKLEY et al., 2015; TURNER et al., 2015). Uma
terceira possibilidade atualmente não testada, é que as fêmeas são realmente mais sensíveis a
anomalias genéticas que afetam o desenvolvimento do cérebro e morrem desproporcionalmente
no útero (MARGARETH et al., 2016).

27
1.1.3 Hipóteses Etiológicas
A etiologia do TEA é bastante complexa e na maioria dos casos o processo
patológico é desconhecido. Estudos mostram que o TEA está diretamente ligado a fatores
genéticos, condições ambientais, neurobiológicas, neuroanatômicas, metabólicas e
imunológicas (SUPEKAR 2015; ONORE et al., 2014; KIM et al., 2015; KIRSTEN et al.,
2015).
O diagnóstico do TEA exige evidência de comunicação verbal e não verbal
prejudicada, compromete o interesse social, interação e comportamentos estreitos ou
repetitivos. Embora algumas citações possam definir as características desses distúrbios em
todo o espectro, existe muita diversidade entre os indivíduos diagnosticados. Essa variabilidade
é retratada na gravidade e variedade dos sintomas e na presença de outras características
adicionais, como comprometimento cognitivo, sintomas neuropsiquiátricos (emocionais e
comportamentais) e características físicas ou biomédicas (GABIS et al., 2014).
A taxa estimada da função cognitiva no TEA é de cerca de 50% subnormal. Ela é
medida como dois desvios padrão abaixo da média, que refletem uma pontuação <70 na maioria
das medidas de QI padronizadas (JOHNSON et al., 2007). Quanto às comorbidades comuns,
até 40% das crianças com autismo em contextos clínicos são diagnosticadas com epilepsia
(GAL et al., 2012), distúrbios neuropsiquiátricos como depressão, ansiedade, obsessões e
fobias (MATSON et al., 2007), transtorno de atenção / hiperatividade (GADOW, 2006) e
síndrome de Tourette (CANITANO et al., 2007). Além disso, características dismórficas estão
presentes em muitos indivíduos diagnosticados com TEA, algumas vezes indicando síndromes
genéticas conhecidas (KAPINOS-GORCZYCA; GORCZYCA; KAPINOS, et al., 2009;
MILES et al., 2000).
Segundo Gabi e colaboradores (2014) a apresentação clínica do TEA parece ter uma
heterogeneidade acentuada, o que sem dúvida obscureceu os esforços para identificar causas
simples ou específica dessa condição. Como resultado, a pesquisa etiológica atual está focada
em inúmeras e variadas vias patogenéticas que levam à manifestação do fenótipo TEA. Por
exemplo, características autísticas englobam uma série de característica fenotípica comum de
vários distúrbios do desenvolvimento neurológico determinados geneticamente, como
síndrome de Rett, síndrome do X frágil, síndrome de Angelman e esclerose tuberosa (MOSS et
al., 2009).
Observou-se que indivíduos diagnosticados com uma das síndromes acima
mencionadas, que também atendem aos critérios de diagnóstico para TEA, geralmente

28
apresentam habilidades cognitivas e adaptativas mais gravemente prejudicadas do que aqueles
que não apresentam TEA associado (CAPONE et al., 2005; JESTE et al., 2008; KAU et al.,
2004). Além disso, várias alterações fisiológicas tem sido associadas a uma incidência
significativamente maior de características autísticas incluindo baixo peso ao nascer
(INDREDAVIK et al., 2005), malformações congênitas (MAIMBURG et al., 2006),
sofrimento fetal (GLASSON et al., 2004), encefalopatia isquêmica hipóxica (BADAWI et al.,
2006), infecção por citomegalovírus congênito (YAMASHITA et al., 2003) fenilcetonúria
(BAIELI et al., 2003) e prematuridade (LIMPEROPOULOS et al., 2008). As evidências
sugerem que, às vezes, a sintomatologia autista pode resultar de insulto cerebral menos
específico, semelhante aos processos variados que causam comprometimento intelectual.
Assim, o perfil autístico pode se apresentar como uma característica primária com etiologia
desconhecida ou como secundária a uma síndrome específica ou distúrbio cerebral.
O TEA é amplamente considerado um distúrbio multifatorial resultante de fatores
de risco genéticos e não genéticos. Há evidências cumulativas para o envolvimento de fatores
genéticos na etiologia do TEA, uma vez que irmãos nascidos em famílias com TEA têm um
risco 35 a 40% maior de desenvolver TEA e com um aumento na taxa atual de
aproximadamente 1% a partir de uma taxa de 0,05% na década de 1970 (STUBBS et al., 2016).
Além disso, estudos genéticos revelaram que alterações nas vias de desenvolvimento de
estruturas neuronais e axonais fortemente envolvidas na sinaptogênese emergem de mutações
genéticas únicas (GESCHWIND et al., 2011; VOINEAGU et al., 2011; CHANG et al., 2015).
É provável que interações entre múltiplos genes e variabilidade na expressão de genes como
resultado de fatores epigenéticos e exposição a fatores ambientais sejam responsáveis por TEA
(MUHLE et al., 2004).
Em um estudo clínico envolvendo gêmeos, foi avaliado que o risco de desenvolver
TEA era de 35 a 40% devido à variabilidade genética, e os 60% restantes foram contribuídos
por fatores ambientais pré-natais, perinatais e pós-natais (HALLMAYER et al., 2011).
Consequentemente, fatores ambientais implicados no TEA incluem complicações pré-natais e
perinatais (GLASSON et al., 2004; MARAMARA et al., 2014), complicações de nascimento
e neonatais (GARDENER et al., 2011; GUINCHAT et al., 2012), infecção viral, doenças
autoimunes e exposição a teratógenos no pré-natal como o ácido valpróico (KERN; JONES,
2006; KOLEVZON et al., 2007). Portanto, uma maior compreensão da interface entre fatores
genéticos e ambientais na patogênese do TEA pode levar a uma estratégia terapêutica
otimizada.

29
A etiologia exata do TEA é desconhecida; é pensado em ter bases genéticas fortes
e complexas com modulação da expressão fenotípica por fatores ambientais (BAILEY et al.,
1996), as vacinas normais da infância foram falsamente implicado na causação do TEA, e uma
vasta pesquisa já comprova que não há nenhuma ligação entre a vacinação e TEA (SPENCER
et al., 2017).
1.1.3.1 Teorias Psicogenéticas
Kanner foi o primeiro a definir o termo Autismo e considerou-o uma perturbação
do desenvolvimento, surgindo a hipótese de uma componente genética. Defendia que, até ao
nascimento, as crianças apresentavam um desenvolvimento normal, mas que, devido aos fatores
familiares (frieza, pouca expressividade dos pais) era originado um déficit afetivo nas crianças,
surgindo desta forma o autismo. Nesta época, considerava-se o Autismo uma perturbação
emocional e não biológica. A criança autista apresentaria défices cognitivos e linguísticos
deficitários devido ao meio que a envolvia, o ambiente não carinhoso da mãe (KANNER et al.,
1943; ASSUMPÇÃO et al., 2007).
Teorias psicogenéticas no autismo fundamenta-se nas teorias psicanalíticas que
defendem que as crianças com autismo são normais no momento do nascimento, mas que,
devido a fatores familiares (pais frios e pouco expressivos), o desenvolvimento afetivo das
crianças fica afetado, provocando um quadro de autismo (BORGES et al., 2000; DUARTE et
al., 2001).
Surge assim, a teoria conhecida pelo termo “Mães Frigorifico” teoria desenvolvida
por Bettelheim em 1967, que assumiu grande importância na década de 70, dando origem a
novos estudos com crianças vítimas de maus-tratos, que desenvolviam autismo como uma
resposta negativa a um ambiente ameaçador e não carinhoso por parte da mãe (BORGES et al.,
2000). Esta teoria foi bastante criticada já que os investigadores se limitaram a observar as
relações da criança com os pais depois de a perturbação estar instalada, não apresentando
suporte empírico que tal justifique (MARQUES et al., 2000).
O estudo sobre o Autismo Infantil baseava-se sobretudo nas anomalias de interação
social, mas a partir da década de 60, a atenção recaiu nos défices cognitivos associados a esta
perturbação. Assim sendo, o déficit cognitivo começa a assumir um papel crucial na
determinação da gênese do autismo (ASSUMPÇÃO et al., 2007).
Alguns anos após a descrição do então denominado autismo, Kanner (1943) alega
ser esse quadro “[...] uma verdadeira psicose [...]”, quando relata que todos os exames clínicos

30
e laboratoriais (disponíveis à época) foram incapazes de fornecer dados consistentes no que se
refere à sua etiologia (EISENBERG; KANNER, 1956). Trinta anos após seu artigo inaugural,
o próprio Kanner questiona as discrepâncias na evolução das onze crianças por ele inicialmente
avaliadas e descritas, uma vez que algumas apresentaram franca deterioração, enquanto outras
evoluíram com discreto prejuízo no desempenho global, propondo que “[...] investigações
bioquímicas podem abrir novas perspectivas ao estudo do autismo infantil [...]” (KANNER et
al., 1971).
A partir dos primeiros relatos descartando a idéia de psicose e associando o autismo
à déficits cognitivos (RITVO et al., 1976), surge uma nova perspectiva para esse quadro,
considerando então um transtorno do desenvolvimento. Gillberg (1990) afirma ser “[...]
altamente improvável que existam casos de autismo não orgânico [...]” frisando que “[...] o
autismo é uma disfunção orgânica – e não um problema dos pais – isso não é matéria para
discussão. O novo modo de se ver o autismo é biológico [...]”
Difícil se torna sua diferenciação dos quadros de déficit cognitivo sobretudo em
função do comprometimento qualitativo no desenvolvimento das interações socias e nas
habilidades comunicacionais, o qual, ainda que passível de ser identificado, em geral não é
destaque no retardo mental (ASSUMPÇÃO et al., 2003).
Assim, passa-se a considerar o autismo uma síndrome comportamental com
etiologia ainda desconhecida, mas certamente orgânica (ASSUMPÇÃO et al., 2003). Pensando
desse modo, se nos dispomos a estudar esse campo extremamente complexo, temos que nos
lembrar dos muitos quadros de origem neurológica (e/ou genética) distintas, descritos em
associação ao autismo. Diante dessa perspectiva, as alterações cromossômicas são relatadas
com muita frequência, ainda que sejam de natureza inespecífica e variada (ASSUMPÇÃO et
al., 2007; KUCZYNSKI et al., 1996).
As manifestações do autismo incluem prejuízos na comunicação e interação sociais,
anomalias sensoriais, comportamentos repetitivos e níveis variáveis de incapacidade
intelectual. Juntamente com esses sintomas centrais, distúrbios psiquiátricos ou neurológicos
recorrentes são comuns em pessoas com autismo, dos quais distúrbios de hiperatividade e
atenção (como transtorno de déficit de atenção / hiperatividade (TDAH), ansiedade, depressão
e epilepsia são bastante prevalentes. Um diagnóstico de autismo é alcançado após a obtenção
de uma história detalhada do desenvolvimento, muitas vezes dos pais, e a observação do
indivíduo interagindo com os pais ou outros indivíduos (LORD et al., 2020).

31
1.1.3.2 Mecanismos Epigenéticos
Durante muito tempo, os cientistas tentaram descrever distúrbios associados às
doenças apenas por fatores genéticos ou ambientais. No entanto, o papel da epigenética nas
doenças humanas foi considerado desde meio século atrás. Na última década, esse assunto
atraiu muitos interesses, especialmente em distúrbios complicados, como plasticidade
comportamental, memória, câncer, doenças auto-imunes além de distúrbios neurodegenerativos
e psicológicos (MOOSAVI, 2016).
O termo epigenética significa “em adição à informação genética codificada no
DNA” (FEINBERG; TYCKO, 2004) e é genericamente utilizado para definir mudanças que
ocorrem na expressão gênica sem, no entanto, ocorrer nenhuma alteração na seqüência do
código genético (GIBBS, 2007; JIRTLE; SKINNER, 2007 WAGGONER, 2007). Assim, a
epigenética constitui uma camada “extra” de regulação da expressão gênica a nível
transcricional. Alguns mecanismos epigenéticos incluem a metilação do DNA, o imprinting,
mudanças na conformação da cromatina e o silenciamento mediado por RNA (MOSS;
WALLRATH, 2007; RODENHISER; MANN, 2006; SULEWSKA et al., 2007).
Nas últimas décadas, muitas investigações mostraram que os mecanismos
epigenéticos estão envolvidos na regulação de todo processo biológico no corpo, desde a
concepção até a morte. Esses mecanismos funcionais estão envolvidos na reorganização do
genoma, embriogênese e gametogênese precoces, bem como na diferenciação celular. A
interação entre a metilação do DNA e as alterações pós-traducionais das histonas, causadas por
proteínas reguladoras e RNAs não codificadores, são agentes epigenéticos essenciais para
reorganizar a cromatina em áreas como eucromatina, heterocromatina e compartimentação
nuclear. Os sinais epigenéticos podem ter impressões de longo prazo, por exemplo, na
aprendizagem e organização da memória ou predisposições para diferentes tipos de
câncer. Marcas epigenéticas incorretas podem resultar em defeitos congênitos, doenças da
infância, ou sintomas de doenças em outras áreas da vida. Por outro lado, algumas marcas
epigenéticas podem ser reversíveis, e esse fato incentivou muitos pesquisadores a se
concentrarem na terapia epigenética (BALI, 2011). Nos últimos anos, foi demonstrado que a
metilação do DNA, em alguns casos, pode ser irreversível (BALI, 2011; LU et al., 2006). Essa
característica pode ser útil em características complexas e doenças desafiadoras, como função
de memória, comportamentos e lesões psicológicas, dependência, câncer e outras doenças que
não poderiam ser explicadas apenas por fatores genéticos ou pelo ambiente.

32
O projeto do genoma humano (HGP) forneceu importantes informações sobre
estrutura, organização e previsão função de 2,85 bilhões de nucleotídeos na sequência de DNA,
que é a base da pesquisa biomédica, diagnóstico genético e genômica evolutiva (COLLINS,
2004). No entanto, a base genética de complexos distúrbios como diabetes, doenças
cardiovasculares, alzheimer, esquizofrenia, doenças auto-imunes e câncer, permanece incerto.
A natureza multifatorial dessas doenças envolve uma complexa rede de genes e influências
ambientais que reprogramam a genoma e dificulta o conhecimento dos mecanismos envolvidos
em sua fisiopatologia. O ambiente afeta a mecanismos epigenéticos que controlam a função do
gene, sem alterar a sequência do DNA, levando alteração da expressão gênica e à interrupção
de muitos processos biológicos. Ao contrário do genoma, o epigenoma pode ser reprogramado
e com grande plasticidade. Reprogramação epigenética é o principal evento durante a
diferenciação celular, conferindo diferenças funcionais e funcionais de um determinado tipo de
célula (fenótipo) herdável mitoticamente, garantindo uma expressão gênica específica de tecido
(FURTADO, 2019).
Os fenótipos comórbidos associados também incluem outras condições de
desenvolvimento neurológico, como distúrbios neurológicos como epilepsia, sinais de
alterações motoras neurológicos e condições clínicas não neurológicas, como problemas
gastrointestinais e cardíacos (GESCHWIND et al., 2009; ALDINGER et al., 2015).
Outro ponto de gatilho epigenético que vem sendo abordado é quanto a exposição
fetal à inseticidas como clorpirifós tem sido associada a uma redução no peso e comprimento
do corpo infantil, atraso no desenvolvimento psicomotor e um maior risco de TEA
(LANDRIGAN et al., 2010; RAUH et al., 2006). Além disso, estudos epidemiológicos recentes
fornecem evidências de que a exposição de mães grávidas, especialmente durante o 1º ou 2º
trimestre, a infecções virais ou bacterianas, promove a ativação imune materna (MIA) e
aumenta o risco de doenças neuropsiquiátricas, incluindo TEA em seus filhos (em 13%) em
comparação com filhos de mães grávidas não expostas (ESTES; MCALLISTER, 2016;
DEVERMAN, 2009). A MIA tem sido associada a aumentos de citocinas neuroinflamatórias,
bem como a anormalidades na expressão de proteínas sinápticas e a desenvolvimentos
aberrantes na conectividade sináptica, os quais podem estar subjacentes à fisiopatologia da TEA
(PENDYALA et al., 2017).
A exposição de gestantes a medicamentos psicotrópicos, principalmente durante o
primeiro trimestre, tem sido considerada um fator de risco para TEA (GARDENER;
SPIEGELMAN; BUKA, 2009). De fato, crianças expostas ao ácido valpróico no útero tem um
risco 8 vezes maior de desenvolver TEA (REVENDA et al., 2005). Estudos adicionais sugerem

33
que a prescrição de antidepressivos para gestantes aumenta consideravelmente o risco de TEA
(CROEN, et al., 2011; GIDAYA et al., 2014).
No entanto, ainda são necessários estudos clínicos adicionais para validar se existe
um aumento definitivo do risco de TEA em crianças nascidas de mulheres diagnosticadas com
depressão que não recebem tratamento ou tratamento antidepressivo durante a gravidez e
também para correlacionar esse risco com o efeito de medicamentos, tipos de antidepressivos
e dosagens durante a gravidez (SHARMA et al., 2018).
O papel significativo da epigenética no desenvolvimento e na doença cerebral é
devido aos seguintes fatores: 1) plasticidade da epigenética durante todos os períodos de
desenvolvimento e envelhecimento do cérebro, bem como regulação dinâmica nos neurônios,
2) organização desordenada da cromatina tanto na infância quanto na neurodegeneração adulta
e 3) aumento rápido dos fármacos modificadores da cromatina demonstrou ter potencial
terapêutico inesperado para distúrbios degenerativos e funcionais do sistema nervoso. Esses
fatores atraíram um grande interesse nos mecanismos associados a cromatina de doenças
neurológicas, e um novo campo de estudo chamado 'neuroepigenética (JAKOVCEVSKI, 2012)
foi estabelecido.
1.1.4 Manifestações Clínicas
1.1.4.1 Déficit de Socialização
Indivíduos com autismo exibem um amplo padrão de deficiências em situações de
interação social. Sob uma perspectiva de diagnóstico, estes déficits sociais são considerados
uma característica central do autismo (WHITMAN et al., 2015).
Embora crianças com autismo se apegue aos seus pais, utilizem ocasionalmente
habilidades não verbais como gestos para fazer solicitações; e possam mostrar habilidades de
revezamento durante as brincadeiras, elas, geralmente apresentam limitações acentuadas nessas
e em outras áreas da interação social, particularmente em situações que exigem atenção
conjunta, iniciação e reciprocidade dinâmica (WHITMAN et al., 2015).
As dificuldades na interação social em indivíduos com TEA podem manifestar- se
por um isolamento ou comportamento social impróprio, pobre contato visual, dificuldades em
participar de atividades em grupo, indiferença afetiva ou manifestações inapropriadas de afeto,
falta de empatia social ou emocional. À medida que crescem e entram na idade adulta, pode
haver uma melhoria quanto ao isolamento social, não obstante permanecer uma socialização

34
empobrecida, com dificuldade de estabelecer amizades persistentes (VOLKMAR,
MCPARTLAND, 2014).
De acordo com o Manual Diagnóstico e Estatístico de Transtornos Mentais (DSM
- 5), as pessoas com TEA tem dificuldade de comunicação e interação com outras pessoas,
interesses restritos e comportamentos e sintomas repetitivos que prejudicam sua capacidade de
se comunicar e manter a rotina adequadamente na escola, no trabalho e em outras áreas da vida
(DSM-V, 2013).
A. Déficits persistentes na comunicação social e na interação social em vários
contextos, conforme manifestado pelo seguinte, atualmente ou pela história:
1. Déficits na reciprocidade socioemocional, variando, por exemplo, na abordagem social
anormal e falha na conversa normal como interagir; para compartilhamento reduzido de
interesses, emoções ou afetos; falha ao iniciar ou responder a interações sociais.
2. Déficits nos comportamentos comunicativos não verbais usados para interação social,
variando, por exemplo, entre comunicação verbal e não verbal mal integrada; a
anormalidades no contato visual e na linguagem corporal ou déficits na compreensão e
uso de gestos; total falta de expressões faciais e comunicação não verbal.
3. Déficits no desenvolvimento, manutenção e compreensão de relacionamentos, variando,
por exemplo, entre dificuldades em ajustar o comportamento para atender a vários
contextos sociais; a dificuldades em compartilhar brincadeiras imaginativas ou em fazer
amigos; à falta de interesse pelos pares
B. Padrões restritos e repetitivos de comportamento, interesses ou atividades, manifestados por
pelo menos dois dos seguintes, atualmente ou pelo histórico
1. Movimentos motores estereotipados ou repetitivos, uso de objetos ou fala (por exemplo,
estereótipos motores simples, alinhando brinquedos ou lançando objetos, ecolalia, frases
idiossincráticas).
2. Insistência na mesmice, aderência inflexível às rotinas ou padrões ritualizados de
comportamento verbal ou não verbal (por exemplo, angústia extrema com pequenas
mudanças, dificuldades com as transições, padrões rígidos de pensamento, rituais de
saudação, precisam seguir a mesma rota ou comer a mesma comida todos os dias).

35
3. Interesses altamente restritos e fixos que são anormais em intensidade ou foco (por
exemplo, forte apego ou preocupação com objetos incomuns, interesses excessivamente
circunscritos ou perseverantes).
4. Hiper ou hipo reatividade a estímulos sensoriais ou interesse incomum em aspectos
sensoriais do ambiente (por exemplo, indiferença aparente à dor / temperatura, resposta
adversa a sons ou texturas específicas, cheiro ou toque excessivo de objetos, fascínio
visual por luzes ou movimento).
C. Os sintomas devem estar presentes no desenvolvimento inicial, mas podem não se manifestar
completamente de forma tardia ou podem ser mascarados na vida por estratégias aprendidas
D. Os sintomas causam comprometimento clinicamente significativo nas áreas sociais,
ocupacionais ou outras áreas importantes do funcionamento atual.
E. Esses distúrbios são melhor explicados por deficiência intelectual (distúrbio do
desenvolvimento intelectual) ou atraso global do desenvolvimento. A incapacidade intelectual
e o distúrbio do espectro do autismo co-ocorrem frequentemente para fazer diagnósticos
comórbidos de desordem do espectro do autismo e deficiência intelectual, a comunicação social
deve estar abaixo do esperado para o nível geral de desenvolvimento.
1.1.4.2 Déficit de Comunicação/Linguagem
Existe um relacionamento dinâmico entre a linguagem das crianças e seu
funcionamento social e cognitivo. Através da linguagem, elas expressam suas necessidades,
pedem aquilo que precisam, regulam o comportamento próprio e dos outros, comunicam seus
sentimentos, exploram e respondem ao seu ambiente social, entendem melhor a si mesmas e
imaginam mundos diferentes daqueles em que vivem (WHITMAN et al., 2015).
A comunicação intencional desenvolve-se cedo (entre 9 e 13 meses), seguidas pelas
primeiras palavras (entre 13 e 18 meses), combinações de palavras (18 e 30 meses), frases (30
a cinco anos) e gramática do discurso (entre cinco e oito anos) (PRIZANT; WETHERBY,
1993).
Deficiência na linguagem é uma característica diagnóstica central do autismo.
Estimativas sugerem que aproximadamente 50% das crianças com autismo não adquirem a
linguagem como um modo principal de comunicação (PRIZANT et al., 1996).

36
O prejuízo relativo à comunicação ocorre de forma variada, indo desde o atraso ou
ausência total de desenvolvimento da linguagem verbal até a presença de linguagem. Nos
indivíduos com linguagem verbal, o prejuízo é notório, por não terem capacidade de iniciar
uma conversa ou mantê-la, registrando-se o uso estereotipado, repetitivo e idiossincrático da
linguagem. Existem alguns indivíduos que desenvolvem uma linguagem imatura, caracterizada
por ecolalia, reversão do pronome, prosódia anormal e entonação monótona. Os que possuem
capacidade expressiva adequada podem ter dificuldades em iniciar ou manter uma conversa.
As dificuldades de linguagem e de comunicação podem perdurar até a vida adulta (COSTA et
al., 2012).
Aqueles que possuem habilidades verbais podem apresentar déficits em
compreender sutilezas de linguagem, piadas, assim como dificuldades em compreender ou
interpretar a linguagem corporal e as expressões faciais (VOLKMAR; MCPARTLAND, 2014).
1.1.4.3 Padrões de Comportamentos
1.1.4.3.1 Interesses Restritos e Estereotipados
Condutas estereotipadas foram encontradas em animais e no homem. Seu
surgimento está frequentemente relacionado ao mau funcionamento do sistema de controle de
conduta, impulsos e motivações no desempenho de algum movimento ou vocalização.
Conforme a incidência, a qualidade dos episódios e a idade da criança, de uma forma ou de
outra, as estereotipias comprometem o desenvolvimento motor, linguístico e as atividades
físicas, sociais, emocionais, cognitivas e educativas (HOFFMANN et al., 2015).
A Classificação Internacional das Doenças, em sua décima edição (CID-10), traz
as estereotipias motoras como uma categoria nosográfica caracterizada por movimentos
intencionais, repetitivos, estereotipados, ritmados, desprovidos de finalidade e sem relação a
um transtorno psiquiátrico ou neurológico identificado. Esses movimentos caracterizam-se por
balançar o corpo e/ou a cabeça, arrancar e/ou torcer os cabelos, estalar os dedos e bater as mãos.
Porém, podem ter algum componente automutilador, sendo reconhecidos por bater a cabeça,
esbofetear a face, colocar o dedo nos olhos, morder as mãos, os lábios ou outras partes do corpo
(HOFFMANN et al., 2015).
Laznik (2004) informa que as estereotipias e automutilações surgem compondo o
quadro do autismo no segundo ano de vida. No entanto, afirmamos que é comum observarmos
em bebês condutas repetitivas que compõem o quadro do desenvolvimento psicomotor infantil,

37
entre elas, o balanceio da cabeça buscando o equilíbrio em decúbito ventral, o balanceio de
braços e pernas em decúbito dorsal, o balanceio do corpo ao se levantar quando sentada e o
hábito de sugar.
Os indivíduos caracterizados pela ausência de espontaneidade compatível com o
nível de desenvolvimento, relativo a jogos ou brincadeiras de imitação social, demonstram
resistência às mudanças, insistências em determinadas rotinas, apego excessivo a determinados
objetos, como rodas ou hélices, usando-os mais para representação simbólica, o que faz parecer
que estejam brincando. Estereotipias motoras e verbais, como balançar, bater palmas
repetitivamente, andar em círculos ou repetir de algumas palavras, frases ou canções são
algumas das manifestações frequentes em indivíduos com TEA. Quando adultos, os autistas
apresentam uma melhor adaptação às mudanças, porém os interesses restritos persistem e
aqueles que demonstram habilidades cognitivas adequadas tem a tendência a concentrar seus
interesses de forma limitada, tal como os horários de trens, aviões, mapas ou fatos históricos,
isto é, elementos que dominam suas vidas (FIRST.; FRANCE; PINCUS, 2004).
1.1.5 Características Neuroquímicas
1.1.5.1 Neuroinflamação
A inflamação é uma reação de defesa contra diversas lesões, o papel da inflamação
nos transtornos neurodesenvolvimentais e neurodegenerativos tem, no entanto, uma explicação
muito mais complexa caracterizada pela ativação de vias de sinalização intracelulares
associadas à expressão de inúmeros mediadores inflamatórios (RIBEIRO et al., 2015).
A neuroinflamação é um componente-chave na indução e progressão de processos
degenerativos no SNC. Estudos demonstram que a neuroinflamação é um importante
mecanismo fisiopatológico das doenças neurodegenerativas no SNC, principalmente por meio
do do aumento da expressão de genes relacionados à produção de mediadores inflamatórios e
neurotóxicos, que contribuem para a cascata de eventos que levam a lesão e morte celular
(FRANK-CANNON et al., 2009; TAYLOR et al., 2013). O equilíbrio no sistema imune
consiste no reconhecimento padrão dos receptores, macrófagos e os sistemas em geral
desempenhando um papel fundamental na homeostase do SNC após a lesão e infecção. Os
componentes imunológicos inatos desequilibrados também podem contribuir para
neuroinflamação e neurodegeneração (SHASTRI; BONIFATI; KISHORE, 2013). Durante a

38
inflamação, ocorre a liberação de mediadores inflamatórios, como: a prostaglandina E2 (PGE2)
e citocinas, como a interleucina 1 beta (IL1β) e o fator de necrose tumoral alfa (TNFα).
Um dos principais indicadores de neuroinflamação no TEA é ativação das
microgliais - células imunes primárias da SNC e são muito semelhantes aos macrófagos
periféricos. Além disso, é relatado que a microglia defende o cérebro, limpando detritos
celulares e neurônios mortos do tecido nervoso através da fagocitose. No entanto, quando
ocorre ativação microglial de forma exagerada e sustentada, esse pode levar à danos colaterais
ou ao tecido saudável (RODRIGUEZ e KERN 2011). Por exemplo, ativação microglial em
excesso sustentada pode resultar na perda de conexões sinápticas saudáveis, por meio dos
desvios de suas funcionalidades normais, sendo consistentemente encontrada e explicada no
TEA (VARGAS et al., 2005). Inflamação em amostras cerebrais post-mortem de indivíduos
diagnosticados com TEA foram observados, especificamente, no cerebelo, giro cingular
anterior e as regiões médio-frontais do cérebro (VARGAS et al., 2005). Ativação neuroglial e
presença de concentrações aumentadas de citocinas inflamatórias como IFN-γ, IL-1β, IL-6,
TNF-α e quimiocinas CCL-2 foram encontradas no tecido cerebral de indivíduos
diagnosticados com TEA.
Anteriormente foi revelado que indivíduos diagnosticados com TEA em
comparação com os controles, apresentam uma resposta no efeito inflamatório elevada como
linfócitos e citocinas pró-inflamatórias, incluindo TNF-α e IFN-γ e em concentrações menores
a citocina anti-inflamatória IL-10 (TORRENTE et al., 2002, ASHWOOD et al., 2004,
NAKAGAWA e CHIBA, 2016), resultando entre os indivíduos diagnosticados com TEA,
inflamação crônica persistente.
Diversas linhas de pesquisa demonstram que tanto fatores genéticos como ambientais
influenciariam o desenvolvimento de anormalidades na circuitária neural que pode
desencadeiar as características do autismo. Os transtornos do espectro do autismo (TEA) são
condições ao longo da vida e muitas vezes devastadoras que afetam gravemente o
funcionamento social auto-suficiente, com um impacto muito negativo durante a vida inteira e
de toda família dos indivíduos afetados. Presumivelmente tem uma etiopatogenia multifatorial,
resultante de uma interação entre fatores genéticos e ambientais (LYALL, 2014; BÖLTE,
2014).
Em estudos realizados com roedores, a inflamação do cérebro e a ativação da
microglia foram prejudiciais para a sobrevivência dos neurônios do hipocampo mesmo após o
nascimento dos animais. Estímulos inflamatórios aplicados em culturas de tecido hipocampal,
tais como: interferon-α, fator de necrose tumoral , lipopolissacarídeo, vírus da imunodeficiência

39
humana-1 com revestimento proteico da glicoproteína 120, adenovírus e β-amiloide,
aumentaram a síntese de numerosas quimiocinas e citocinas do hipocampo, indicando que os
fatores secretados por sítios da neuroinflamação são atraentes para as células progenitoras
neuronais e sugerindo que as quimiocinas desempenham um papel importante neste processo
(BELMADANI et al., 2006; EKDAHL; KOKAIA; LIDVALL, 2009).
Todos os tipos de células neurais e não neurais que fazem parte do SNC em
desenvolvimento, como micróglia e células endoteliais, usam citocinas para sinalizações
parácrinas e autócrinas, e, pelo fato de muitas dessas mesmas citocinas também servirem como
moduladores imunes nos processos de desenvolvimento do SNC, podem estar susceptíveis à
distúrbios por desequilíbrio imunológico, como por exemplo, a partir de infecções maternas
(DEVERMAN et al., 2009).
Na Figura 2 pode ser observado um exemplo de lesão microglial após
neuroinflamação: em resposta à um desequilíbrio imunológico e dano neuronal, um estado de
neuroinflamação desenvolvido no interior do cérebro ativa principalmente a microglia. A
microglia ramificada é transformada em microglia ativada e libera várias moléculas
neuroinflamatórias que levam a danos na barreira hematoencefálica. Esse dano promove a
infiltração macrofágica que mais tarde exagera o influxo de citocinas inflamatórias, agravando
o estado neuroinflamatório existente no SNC, causando dano neuronal secundário e
subsequente ativação microglial.
Figura 2 – Representação esquemática de como a estimulação de mastócitos e micróglia pode
levar a vários efeitos que contribuem para a inflamação cerebral, a patogênese e os sintomas
do autismo.

40
Fonte: Adaptado: Theoharides (2013). Legenda: MCP= proteína quimiotática de monócitos
O papel da inflamação para a neurogênese tem, no entanto, uma explicação muito
mais complexa. Evidências experimentais indicam que a microglia, em determinadas
circunstâncias, pode ser benéfica e apoiar as diferentes etapas na neurogênese, desde a
proliferação de células progenitoras, sobrevivência, migração e até a diferenciação. Sua
ativação, como indicador de inflamação, pode ou não ser antineurogênico, mas o resultado final
é dependente do equilíbrio entre moléculas secretadas com ação anti e pró-inflamatória. A
inflamação é uma reação de defesa contra diversos insultos, destinada a eliminar os agentes
nocivos sendo capaz de inibir os seus efeitos prejudiciais. Ela consiste de uma deslumbrante
variedade de mecanismos moleculares e celulares e uma intrincada rede de trabalho para mantê-
los sob controle (EKDAHL; KOKAIA; LIDVAL, 2009; WYSS- CORAY; MUCKE et al.,
2002).
Até que ponto a inflamação é benéfica para o tecido cerebral lesionado, e como ele
contribui para os danos cerebrais secundários e perda neuronal progressiva? Existe uma dupla
evidência, tanto um efeito benéfico, como um papel deletério da neuroinflamação após lesão
cerebral traumática. A estimulação dos neurônios sensoriais produz vasodilatação,
extravasamento de plasma e hipersensibilidade. Progressos foram feitos na compreensão da
etiologia da inflamação neurogênica, estudos tem-se centrado em grande parte no papel do
péptido relacionado com o gene neuropeptídeo, a substância P e a calcitonina, que são liberados
na periferia por ativação de neurônios sensoriais de pequeno diâmetro (MORGANTI-
KOSSMANN et al., 2002; RICHARDSON; VASKO, 2002).
Além da imunidade sistêmica alterada (GOINES et al., 2010; MEAD et al., 2015),
neuroinflamação foi observada no cérebro de pacientes com TEA. A presença de microglia
ativada foi relatada no córtex pré-frontal dorsolateral de pacientes autistas (MORGAN et al.,
2010). Estudos post-mortem de indivíduos com TEA também demonstraram a ativação da
microglia, bem como um aumento na densidade (MORGAN et al., 2010; TETREAULT et al.,
2012; VARGAS et al., 2005). Reforçando a ideia de disfunção imunológica no TEA (CAO et
al., 2012; ASHWOOD et al., 2011; MOLLY et al., 2006), essa ativação da microglia é
acompanhada pelo aumento da expressão de fatores pró-inflamatórios, como citocinas e
quimiocinas, no cérebro e no líquido cefalorraquidiano de indivíduos com TEA (MORGAN
et al., 2010; VARGAS et al., 2005).
Apesar da falta de evidências em humanos de que a neuroinflamação desempenha
um papel direto na patogênese do autismo, pesquisas em modelos animais sugerem fortemente

41
que esse seja o caso. Demonstrou-se que os déficits na atividade microglial durante o
desenvolvimento do cérebro são prejudiciais à formação de sinapses maduras, levando a um
aumento de sinapses imaturas que podem ser responsáveis por déficits cognitivos e
comportamentais do tipo TEA (PAOLICELLI et al., 2011; ZHAN et al., 2014). Portanto, além
dos fatores de risco genéticos para inflamação, fatores ambientais que levam a eventos
neuroinflamatórios estão recebendo mais escrutínio na etiologia do autismo.
1.1.5.2 Mecanismos Moleculares e Genéticos
Atualmente, existem fortes evidências que apontam para um forte componente
genético na maioria das doenças psiquiátricas, entre elas o TEA (LEVY; MANDELL;
SCHULTZ; 2009). Desse modo, estudos buscam demonstrar a relação entre o quadro clínico
do TEA e a influência genética na expressão de alguns sintomas (TAMANAHA;
PERISSINOTO; CHIARI, 2008).
Da mesma forma, encontrados nas literaturas em estudo com roedores, o desafio
materno com AVP induz fenótipos do tipo autista em descendentes masculinos e femininos
(KATAOKA et al., 2013; HARA et al., 2017), não sendo diferente os resultados nas
anormalidades comportamentais e celulares observadas entre os sexos.
As expressões genéticas específicas dos alelos dos pais ao longo de domínios
impressos são produzidas por elementos de sequência especializados chamados 'regiões de
controle de impressão' (ICRs). Os ICRs estão localizados apenas em uma das cópias dos pais,
cuja função é regular as expressões gênicas por meio de um alelo específico. Embora a
metilação do DNA seja a alteração epigenética mais bem investigada nos ICRs, a metilação e
acetilação das histonas nos ICRs também foram relatadas (KANEDA, 2004). Para colocar
impressões de metilação nos ICRs, o Dnmt3a é essencial. Após a fertilização, através da
manutenção somática, as alterações de metilação alélica são conservadas durante o
desenvolvimento. Esse processo é complicado devido à sua ligação ao ciclo celular e requer
funções proporcionais de complexos multi-enzimáticos que podem ser afetados por agentes
intrínsecos e extrínsecos (DELAVAL, 2006). Outras importantes modificações de metilação
foram reconhecidas em genes envolvidos no desenvolvimento da doença de Alzheimer e
esquizofrenia. Além disso, foi confirmado um papel para modificações epigenéticas em
doenças psiquiátricas, como síndrome e dependência de Rubinstein-Taybi, doença de
Huntington e síndrome do X frágil. Atualmente, é amplamente aceito que, para a função normal
e as características do neurodesenvolvimento do cérebro, a constância da metilação do DNA e

42
das modificações das histonas é essencial, e sua desregulação pode resultar em doenças
fenótipas (PTAK, 2010).
O status de metilação do DNA tem alta estabilidade e serve como uma memória
epigenética especial de células específicas em todos os períodos do ciclo celular. Também pode
regular a expressão e a atividade dos códigos de histonas. A aceleração da metilação do DNA
nos locais CpG é mediada por enzimas DNA metiltransferase, como Dnmt1, Dnmt3a e
Dnmt3b. Dentro das células, a S-adenosilmetionina atua como um importante doador do grupo
metil. Nesse sentido, o ácido fólico e a vitamina B12 desempenham papéis determinantes na
re-metilação ou na atração da forma desmetilada da S-adenosilmetionina por meio de
mecanismos passivos e ativos (ABDOLMALEKY, 2008; KIM, 2005). Vários estudos
demonstraram que esses fatores efetivos podem alterar os padrões de metilação do DNA e
alterar os níveis de expressão gênica (COONEY, 2002; WATERLAND, 2006). Estudos
também confirmaram que o estado nutricional nos primeiros anos de vida pode afetar o padrão
de metilação do DNA e os níveis de expressão gênica na idade adulta (WEAVER, 2004). Além
disso, os padrões de metilação nas seqüências de CpG nos resíduos de citosina podem ser
herdados e atuar como características específicas de tecidos e espécies. É interessante que 70%
a 80% do DNA humano nas seqüências de CpG sejam geralmente metilados (LU, 2006;
TAYLOR, 2007). e foram relatadas correlações entre a metilação e os níveis de expressão
em cis e trans (VAN, 2014).
Cinco membros da família de proteínas de ligação o DNA helicase cromodomínio (Chd)
- (Chd1, Chd2, Chd5, Chd7 e Chd8) têm sido implicados no TEA e NDD (desordens do
desenvolvimento). No entanto, apenas Chd2, no cromossomo 15q26.1, e Chd8, no cromossomo
14q11.2, estão incluídos na lista de genes com fortes evidências estatísticas baseadas sobre os
estudos recentes de WES whole-genome sequencing (sequenciamento completo do genoma)
em grande escala no TEA e nas NDDs (STUDY, 2017; IOSSIFOV et al., 2014). Mesmo antes
dos estudos de WES em grande escala, havia evidências para o denvolvimento de Chd2 em
NDDs por meio de relatórios de microdeleções que afetam o 15q26.1 englobando o gene
(BHAKTA et al., 2005). Esses relatórios foram seguidos por uma descrição detalhada de seis
indivíduos com DNVs em Chd2 encontrados entre uma amostra de 500 (1,2%) com
encefalopatias epilépticas (CARVILL et al., 2013). Este estudo relatou que cinco dos seis
indivíduos (83%) com DNVs que afetam Chd2 apresentaram regressão e dois indivíduos
também foram diagnosticados com TEA. Atualmente, é conhecido que 32 indivíduos têm
NDDs relacionados a Chd2 e o fenótipo consiste em epilepsia de início precoce encefalopatia

43
seguida por desaceleração cognitiva ou regressão associada com a atividade epileptiforme
contínua (CARVILL et al., 2015).
Alguns pesquisadores vêm tentado desvendar os fatores genéticos associados a esse
transtorno. Causas como convulsões, deficiência mental, diminuição de neurônios e sinapses
na amígdala, sugerem um forte componente genético, bem como o tamanho aumentado do
encéfalo e concentração aumentada de serotonina circulante (LEVY; MANDELL; SCHULTZ;
2009).
Cerca de 15 anos atrás, estimava-se que aproximadamente 15 genes pudessem estar
envolvidos nesse transtorno. Não obstante, os estudos genéticos na atualidade veem sinalizando
que vários loci no genoma tem sido associado a esse transtorno, embora poucos genes
específicos tenham sido identificados (WILLSEY; STATE, 2013).
Interessa destacar que apesar de não existir um consenso, entre os pesquisadores na
identificação dos genes relacionados com o TEA, ocorre em contrapartida unanimidade na
comunidade cientifica o reconhecimento que na maioria dos indivíduos com TEA, os múltiplos
loci interagem entre si, contribuindo de forma significativa para expressão do transtorno
(TORDJMAN et al., 2014).
No que concerne aos estudos neurobiológicos algumas anomalias em estruturas
cerebrais, encontradas em pelo menos um subgrupo, vem sendo relacionadas as idiossincrasias
comportamentais do TEA (LEVY; MANDEL; SCHULTZ; 2009). Sob esse enfoque cabe
destacar, a presença de fatores de riscos pré-natais, perinatais e pós-natais tais como: a
prematuridade, a pós-maturidade, a hipóxia neonatal, rubéola, sífilis congênitas, toxoplasmose,
citomegalovírus e as infecções pelo vírus Herpes Simplex, os quais representam eventos
importantes para o desfecho do transtorno (TORDJMAN et al., 2014).
Os anos de 1960 e 1970 foram críticos para o papel dos fatores genéticos na gênese
do autismo (ASSUMPÇÃO et al., 2015). As evidências replicadas tanto em estudos com
gêmeos quanto em famílias durante as décadas de 1970 e 1980 reforçam a hipótese genética e
levantaram a probabilidade de que se tratava de um fenótipo autístico muito amplo do que a
categoria de diagnóstico mais convencional. Os achados clínicos e cromossômicos também
apontaram para a heterogeneidade genética dessa condição (RUTTER et al., 2000).
Satterstrom et al., (2020) apresentaram o maior estudo de sequenciamento de
exoma do TEA até o momento contemplando um número de 35.584 amostras no total, 11.986
com TEA. Usaram uma estrutura analítica aprimorada para integrar variação rara de novo e
controle de caso, identificando 102 genes de risco a uma taxa de descoberta falsa de 0,1 ou
menos. Desses genes, 49 mostraram frequências mais altas de variantes disruptivas de novo em

44
indivíduos que apresentaram um atraso grave no desenvolvimento, enquanto 53 mostraram
freqüências mais altas em indivíduos que tinham TEA; comparar casos de TEA com mutações
nesses grupos revela diferenças fenotípicas. Expressas precocemente no desenvolvimento do
cérebro, a maioria dos genes de risco tem papel na regulação da expressão gênica ou
comunicação neuronal (isto é, mutações afetam alterações neurodesenvolvimentais e
neurofisiológicas) e 13 caem em loci atingidos recorrentemente por variantes de número de
cópias (SATTERSTROM et al., 2020).
O aumento nos genes identificados associados ao TEA (SFARI, 2019) observados
na Figura 3 é representado juntamente com a prevalência de TEA (dados extraídos do Centro
de Controle e Prevenção de Doenças), as diferentes versões do Manual Estatístico de
Diagnóstico (do DSM II ao DSM 5.0) e o avanço da tecnologia genética (Adaptado de
HUGUET; BOURGERON, 2016).
Figura 3 – A história da genética do TEA de 1975 a 2015.
Fonte: Adaptado de Huguet e Bourgeron, 2016.
Com base em mais de 13 estudos com gêmeos publicados entre 1977 e 2015, os
pesquisadores estimaram a contribuição genética e ambiental para o TEA (HUGUET et al.,
2016). Em 1977, o primeiro estudo sobre autismo por Folstein e Rutter (1977) relatou uma
coorte de 11 gêmeos MZ e 10 gêmeos DZ. Este estudo mostrou que os gêmeos MZ eram mais
concordantes com o autismo - 36% (4/11) - em comparação com 0% (0/10) para os gêmeos DZ.
Quando um "fenótipo de autismo mais amplo" foi usado, a concordância aumentou para 92%
para os gêmeos MZ e para 10% dos gêmeos DZ (BAILEY et al. 1995). Desde este primeiro
estudo de pequena escala, estudos com gêmeos relataram constantemente uma maior
concordância para TEA em MZ em comparação com DZ (RITVO et al., 1985;
STEFFENBURG et al., 1989; BAILEY et al., 1996).

45
Entre 2005 e 2009, três estudos de gêmeos com grupos relativamente grandes de
gêmeos (285-3419) relataram altas concordâncias para TEA em gêmeos MZ (77-95%) em
comparação com gêmeos DZ (31%), (RONALD et al., 2005; TANIAI et al., 2008;
ROSENBERG et al., 2009). Notavelmente, as concordâncias MZ foram semelhantes às
relatadas nos estudos anteriores, mas as concordâncias DZ foram maiores. Lichtenstein et al.,
(2010) relataram uma concordância relativamente baixa para TEA em 39% dos gêmeos MZ em
comparação com outros estudos (a concordância para gêmeos DZ neste estudo foi de 15%). No
entanto, como indicado anteriormente por estudos usando o "fenótipo de autismo mais amplo",
todos os gêmeos MZ discordantes dessa coorte apresentavam sintomas de ESSENCE (por
exemplo, ID, TDAH, atraso no idioma etc.).
Uma proporção significativa da contribuição genética para o TEA mostrou-se
compartilhada com outros distúrbios do desenvolvimento neurológico, como TDAH (> 50%) e
dificuldade de aprendizagem (> 40%), (LICHTENSTEIN et al., 2010; RONALD et al., 2010;
LUNDSTROM et al., 2011; RONALD; HOEKSTRA, 2011). Em resumo, quando todos os
estudos com gêmeos são levados em consideração, a concordância para TEA é de
aproximadamente 45% para gêmeos MZ e 16% para gêmeos DZ (RITVO et al., 1985;
STEFFENBURG et al., 1989; BAILEY et al., 1995).
Além disso, cerca de 10% dos casos estão associados as doenças de herança
monogênica como esclerose tuberosa (relata em estudo na década de 1990) segundo Smalley
(1998), síndrome do X-frágil e neurofibromatose (FOLSTEIN; ROSEN-SHEIDLEY, 2001), e
em aproximadamente 3% dos casos, os indivíduos possuem alguma anomalia cromossômica
detectável por cariótipo (VORSTMAN et al., 2006).
Os primeiros estudos usando técnicas clássicas de mapeamento genético, tais como
ligação, triagem de genes candidatos e associação de variantes comuns, forneceram resultados
modestos, principalmente porque foram insuficientes para detectar variantes causais à luz da
alta heterogeneidade genética no TEA. Na última década, os avanços nas tecnologias de
sequenciamento de DNA, juntamente com a coleta de grandes grupos de pacientes e o amplo
compartilhamento de dados, permitiram um enorme progresso na identificação de genes
implicados no TEA (GESCHWIND; STATE, 2015).
Uma abordagem que tem se mostrado muito interessante para identificação de
genes e vias relacionados a doenças psiquiátricas são os estudos de expressão gênica,
especialmente após o desenvolvimento da técnica de microarranjos de DNA (ou em inglês,
DNA microarray). Esta técnica passou a permitir a avaliação da expressão de um grande
número de transcritos em um único experimento, possibilitando uma análise ampla em um

46
determinado tecido e momento particular e, portanto, abrindo perspectivas para a identificação
de vias importantes para os TEA (OLIVEIRA et al., 2011).
Houve um grande progresso na última década na compreensão da genética do TEA,
estabelecendo novas mutações, incluindo variantes de número de cópias (CNVs) e mutações
pontuais que provavelmente atacam os genes codificadores de proteínas, como causas
importantes do TEA (SANDERS et al., 2012; IOSSIFOV et al., 2014). No entanto, quando
combinados, todos os genes conhecidos associados ao TEA explicam apenas uma pequena
fração de novos casos e estima-se que, de maneira geral, novas mutações em genes que
codificam proteínas (incluindo CNVs) contribuam para não mais do que 30% de casos de TEA
(IOSSIFOV et al., 2014; YUEN et al., 2017). A grande maioria das novas mutações
identificadas está localizada em regiões intrônicas e intergênicas; no entanto, pouco se sabe
sobre sua contribuição para a arquitetura genética de TEA ou para qualquer outra doença
complexa (ZHOU et al., 2019).
1.2 Ácido Valpróico
O valproato é um medicamento antiepilético amplamente aceito que também é
utilizado para o gerenciamento do transtorno bipolar (NATASHA et al., 2008) e para a
profilaxia da dor de cabeça da enxaqueca (LAGACE et al., 2004).
O ácido valpróico (AVP) é comercializado e utilizado no mercado como
anticonvulsivante desde 1974 e é usado em muitos países devido à sua eficiência contra vários
tipos de epilepsias nos últimos 40 anos, se mostrando eficiente no controle de crises convulsivas
em adultos e especialmente na infância (NALIVAEVA et al., 2009), e como um estabilizador
do humor. Uma das suas principais ações é o aumento no nível do ácido gama aminobutírico
(GABA) no cérebro (ORNOY et al., 2009).
Do ponto de vista farmacológico, o AVP pertence à classe de inibidores da enzima
histona desacetilase (HDAC) com ação também sobre canais iônicos e transportadores de
GABA e pode atuar alterando expressão gênica através de controle epigenético. A regulação da
acetilação de lisinas na porção C-terminal das moléculas de histonas presentes nos
nucleossomos altera o estado de compactação e a estrutura da cromatina (ORNOY et al., 2009).
O aumento da acetilação de histonas produz descompactação do DNA, aumentando
o acesso da RNA polimerase II e proteínas acessórias necessárias à transcrição. O efeito é
observado através do aumento da expressão de genes comumente silenciados. Assim, o
equilíbrio entre as funções das enzimas HDAC e histona acetilase (HAC), além das DNA metil-

47
transferases, que controlam o grau reestruturação da cromatina e o nível de transcrição de
regiões cromosômicas específicas, fica comprometido (GÖTTLICHER et al, 2001).
A exposição de uma droga com alto poder de regulação da expressão gênica durante
o período de neurogênese pode levar então a uma modificação nos padrões de expressão de
genes relacionados ao neurodesenvolvimento. A inibição das proteínas histonas desacetilases
induzidas pelo AVP tem se mostrado importante na neuroproteção de cultura de neurônios
corticais do AVP, aumentando a viabilidade dessas células in vitro (JEONG et al, 2003).
1.2.1 Efeitos Teratogênicos
A exposição ambiental pré ou pós-natal a fatores pró-oxidantes, como ácido AVP,
mercúrio, chumbo, vírus, poluentes do ar, toxinas, talidomida e ácido retinóico pode servir
como fatores desencadeantes do estresse oxidativo no autismo (CHAUHAN; CHAUHAN,
2006; KERN; JONES, 2006; SWEETEN et al., 2004; TONG et al., 2003; ZOROGLU et al.,
2004).
Muitos relatos sugerem que o AVP pode produzir anormalidades comportamentais
(VOORHEES, 1987; YOCHUM et al., 2010), dificuldades no aprendizado e múltiplos defeitos
congênitos (SCHNEIDER; TURCZAK; PRZEWŁOCKI, 2005), devido aos quais foi
empregado para manifestar sintomas de autismo em animais. O AVP é capaz de induzir
anomalias nos neurodesenvolvimentos e isso tem sido atribuído ao seu status de "teratogênico"
(COLLINS et al., 1991; EHLERS et al., 1992). Acredita-se que o autismo induzido por AVP
ocorra devido a déficits no desenvolvimento neuronal no cerebelo, sistema límbico e tronco
cerebral, causando interrupções na conectividade sináptica (BAILEY et al., 1998; KEMPER;
BAUMAN, 1993; RINALDI et al., 2007).
Logo após o AVP ser introduzido no mercado para uso clínico como
anticonvulsivante surgiram casos de alterações no tubo neural, particularmente de espinha
bífida, entre os filhos expostos a esse medicamento no início de gestação (KOREN et al., 2006).
Estas alterações podem afetar o desenvolvimento da linguagem e habilidades de aprendizagem
(ORNOY et al., 2009). Além disso, alterações nos sistemas cardiovascular, genitourinário e
endócrino também foram associadas ao uso de AV (ALSDORF; WYSZYNSKI 2005).
Mulheres em idade fértil, tratadas com este fármaco, devem usar contraceptivos e
interromper a medicação antes de qualquer gravidez planejada. Se não houver nenhuma outra
alternativa para o tratamento com ácido valpróico, a gravidez deve ser considerada de alto risco

48
e deve ter o acompanhamento adequado incluindo o diagnóstico pré-natal (ORNOY et al.
2009).
1.2.2 Mecanismo de Ação
Foi relatado que a propensão teratogênica de drogas antiepilépticas ocorre devido
ao envolvimento do estresse oxidativo (DEFOORT et al., 2006). Achados experimentais
sugerem que o valproato e seus produtos metabólicos podem desencadear a liberação abundante
de espécies reativas de oxigênio, como peróxido de hidrogênio e radicais hidroxila. Esses
radicais livres podem atuar em vários locais do corpo, resultando na formação de biomarcadores
de estresse oxidativo, que são substâncias reativas ao ácido tiobarbitúrico e 15-F-isoprostano
(VERROTTI et al., 2008).
A geração de radicais livres constitui, por excelência, um processo contínuo e
fisiológico, cumprindo funções biológicas relevantes. Durante os processos metabólicos, esses
radicais atuam como mediadores para a transferência de elétrons nas várias reações
bioquímicas. Sua produção, em proporções adequadas, possibilita a geração de ATP (energia),
por meio da cadeia transportadora de elétrons; fertilização do óvulo; ativação de genes; e
participação de mecanismos de defesa durante o processo de infecção. Porém, a produção
excessiva pode conduzir a danos oxidativos (SHAMI 2004; FERREIRA, 1997).
A instalação do processo de estresse oxidativo decorre da existência de um
desequilíbrio entre compostos oxidantes e antioxidantes, em favor da geração excessiva de
radicais livres ou em detrimento da velocidade de remoção desses. Tal processo conduz à
oxidação de biomoléculas com consequente perda de suas funções biológicas e/ou desequilíbrio
homeostático, cuja manifestação é o dano oxidativo potencial contra células e tecidos
(HALLIWELL, 2004). A cronicidade do processo em questão tem relevantes implicações sobre
o processo etiológico de numerosas enfermidades crônicas não transmissíveis, entre elas a
aterosclerose, diabetes, obesidade, transtornos neurodegenerativos e câncer (GREEN, 2004).
Ferrari (2004), em estudo de revisão, ratifica que a geração de radicais livres desencadeia
eventos patológicos que, por sua vez, estão envolvidos nos processos cardiovasculares,
carcinogênicos e neurodegenerativos.
O AVP atravessa a placenta e está presente em uma concentração mais alta no feto
do que na mãe, embora não esteja claro se a concentração sérica fetal é maior durante
embriogênese (DICKINSON et al., 1997). Vários estudos sugeriram que a permanência dos
níveis total do ácido valpróico caem, mas essas frações livres aumentam durante a gravidez.

49
Achados sugerem que o metabolismo real do AVP não é alterado pela gravidez e que as
alterações da depuração plasmática se devem principalmente à diminuição da ligação às
proteínas (JOHANNESSEN et al., 1992).
A farmacocinética clínica e a farmacodinâmica do AVP foram revisadas
extensivamente (LOSCHER et al., 1999; GUGLER et al., 1980). A taxa de concentração sérica
terapêutica foi estimada em 50 - 100 mg/l (GIBALDI et al., 1991; BOURGEOIS et al., 1997)
ou 40-150 mg/l (DEAN et al., 1997). Recomenda-se evitar doses grandes e únicas em mulheres
grávidas, pois elas podem causar altos picos sérico com ação agressiva e irreversível, pondo em
risco o feto (DELGADO et al., 1992). É preferível tomar doses múltiplas/fracionadas de AVP
nos intervalos ao longo do dia para evitar picos séricos elevados durante a gestação (ASDORF
et al., 2005).
1.2.3 Uso em Modelos Animais
Os ratos têm sido amplamente utilizados em pesquisas sobre autismo por muitas
razões, principalmente por causa da tecnologia disponível para manipular o genoma do rato.
Isso permitiu o teste de muitas hipóteses sobre os genes que se pensa estarem envolvidos na
patogênese do TEA. Existem excelentes revisões com críticas sobre os modelos de
camundongos com mutações monogênicas (KAZDOBA et al., 2016). Esses modelos são
informativos para a biologia específica para mutações genéticas únicas, como síndrome do X
frágil, síndrome de Rett e esclerose tuberosa, da qual menos de 10% dos casos também são
diagnosticados com TEA (CHADMAN et al., 2017; ELLENBROEK et al., 2016).
Dos agentes ambientais, epigenéticos ligados ao TEA, o AVP é o que está sendo
estudado mais extensivamente. Um dos modelos baseia-se na exposição de ratas prenhas ao
ácido valpróico no nono dia gestacional (SCHNEIDER et al., 2006), afetando a migração de
neurônios como os serotoninérgicos, ou no 12,5º dia de gestação (RODIER et al., 1996),
podendo afetar a diferenciação neural. Essa exposição não produz anormalidades
neuroanatômicas, no entanto os filhotes apresentam distúrbios comportamentais, como o
aumento de tempo gasto em atividades estereotipadas semelhantes aos pacientes autistas
(SCHNEIDER et al., 2006). Esse período é equivalente ao dia 21 da gestação em humanos,
quando o tubo neural fecha, um passo importante no neurodesenvolvimento. Ratos expostos ao
AVP no pré-natal apresentam menor exploração social e aumento de estereotipias locomotoras
(STANTON et al., 2007).

50
No entanto, vários artigos citaram utilizar especificamente o AVP no período pós-
natal e apresentaram resultados semelhantes aos demais quando se referem às alterações
neuroquímicas e comportamentais observadas no TEA (YOCHUM et al., 2010; YAOCHUM
et al., 2008; YADAV et al., 2016), sendo um passo interessante nas pesquisas com autismo
regressivo, ou seja, perda das habilidades adquiridas desde o nascimento.
Modelos animais de autismo podem ser divididos em quatro grupos (BELZUNG et
al., 2005):
(i) animais com certas deficiências de receptores neuro-peptídeos, em particular
os de vasopressina (cepa de ratos e camundongos Brattleboro com uma mutação
zero dos subtipos de receptores de vasopressina (VAN et al., 1982), ocitocina
(camundongos knockout com a ausência de receptores de ocitocina) (POBBE et
al., 2012) e opióides (camundongos com uma exclusão direcionada de receptores
μ-opioides (WÖHR et al., 2011);
(ii) modelos com a reprodução de estados patológicos que aumentam o risco de
autismo nas pessoas; efeitos pré-natais de anticonvulsivantes, embriopatia
induzida por talidomida (NARITA et al., 2010) e distúrbios dos mecanismos de
síntese de serotonina no decorrer do desenvolvimento pré-natal (KAHNE et al.,
2002);
(iii) modelos com comprometimentos neonatais das zonas cerebrais, cujas
anomalias são conhecidas por estarem presentes em pessoas com autismo; as
estruturas estudadas são o cerebelo, o complexo amigdaliano ou o córtex pré-
frontal medial (TSAI et al., 2012);
(iv) modelos genéticos de doenças humanas associadas ao autismo, como a
síndrome de um frágil cromossomo X (MINES et al., 2010).
No entanto, há uma pergunta bastante natural: quão adequados são os modelos
acima? Nesses modelos, apenas certos aspectos da etiologia dos respectivos distúrbios são
reproduzidos; ao mesmo tempo, o autismo é um estado multifatorial. Em modelos animais,
observa-se manifestações associadas a distúrbios autistísticos "humanos", mas é difícil
identificar se essas manifestações são as razões desses distúrbios, suas consequências,
diferentes consequências de razões idênticas ou se essas manifestações estão interconectadas
por outras relações. Por exemplo, distúrbios no funcionamento do cerebelo se correlacionam
com a ocorrência de TEA, mas não há relação causal neste caso (MINSHEW et al., 1999). Além
disso, alguns fatores que induzem sintomas de autismo em animais estão ausentes em seres
humanos. Em particular, a exclusão do gene do receptor V1a proposto como um dos modelos

51
de autismo em roedores não foi encontrada em pessoas que sofrem de TEA (WASSINK et al.,
2004).
Belzung et al., (2005) mencionaram sete modelos nos quais se manifestam duas
características principais do TEA (distúrbios das inter-relações sociais e estereotipicidade /
rigidez cognitiva): modelos baseados nos efeitos pré-natais da 5-metoxitriptamina (5-MT)
(AZMITIA et al., 1999) e do ácido valpróico (FOLEY et al., 2012), três modelos de distúrbios
pós-natais iniciais, porquinhos-da-índia da linha GS e ratos infectados neonatalmente pelo vírus
Bourne (PLETNIKOV et al., 2002). Oito outros modelos foram qualificados na mesma
publicação que os válidos para modelar sintomas específicos do autismo. A terceira
característica da tríade autista (distúrbios da comunicação social) é difícil de modelar porque a
especificidade do fenômeno da fala, típico apenas dos seres humanos, cria dificuldades cruciais
definidas (CRAWLEY et al., 2000).
Modelos de lesões anatômicas são há muito utilizados para isolar regiões cerebrais
envolvidas nas vias patológicas de uma série de distúrbios neurológicos. No caso do autismo,
no entanto, danos cerebrais induzidos por uma lesão química ou eletrolítica grossa de regiões
específicas não foram capazes nem remotamente replicar TEA humano. A complexidade do
desenvolvimento humano e a heterogeneidade encontrada nos fenótipos do TEA poderia talvez
ser duas das principais razões que dificultam muito a recapitulação do TEA neste tipo de lesão
de modelos pós-natal. Além disso, as deficiências encontradas no TEA não são prováveis estar
enraizada apenas em um único circuito neural ou região cerebral (KIM et al., 2016).
Existem dois tipos de modelos animais produzidos pela exposição perinatal de
roedores ao ácido valpróico (AVP). Roedores expostos ao AV no dia embrionário (E) 12 têm
sido usados como modelo animal para o autismo, caracterizado por comportamento social
prejudicado, ansiedade aumentada e diminuição da sensibilidade à dor após a maturidade
(MARKRAM et al., 2008; SCHNEIDER et al., 2007, 2008). Por outro lado, roedores expostos
ao AVP no 14ª dia pós-nascimento têm sido usados como modelo animal para o autismo
regressivo, que mostra comportamentos sociais prejudicados, como modelos animais para o
autismo, mas acompanhados pela perda de algumas habilidades adquiridas (YOCHUM et al.,
2008, 2010; WAGNER et al., 2006). O modelo de autismo regressivo também mostrou
ansiedade aumentada em nosso estudo preliminar (ATSUKO et al., 2013).
Wang et al., (2016) citam em sua pesquisa que roedores que receberam AVP no 14ª
dia pós-natal mostraram comportamentos relacionados à ansiedade e menos interações sociais
(OGUCHI et al., 2013; YOCHUM et al., 2008). Foi relatado que os filhotes de ratos que
receberam injeções diárias de AVP no 6ª dia embrionário por um período máximo de 2 semanas

52
exibiram um comportamento social reduzido, juntamente com hipertrofia cortical e
propriedades eletrofisiológicas do tipo adulto em 40% dos neurônios localizados no córtex de
associação temporal (CHOMIAK et al., 2010). Dado que os déficits induzidos pela exposição
pré-natal ou pós-natal ao AVP em roedores são semelhantes aos sintomas observados em
crianças com exposição ao AVP no útero ou com autismo, a exposição de roedores ao AVP no
momento crítico do desenvolvimento do cérebro é considerada o modelo animal com o uso de
valproato fetal (WILLIAMS et al., 2001) e autismo regressivo (OGUCHI et al., 2013). No
entanto, pouca informação está disponível sobre a associação de alterações moleculares e
celulares no hipocampo e na amígdala com o desenvolvimento de distúrbios comportamentais,
quando experiências adversas de vida precoce ocorrem no momento do pico do crescimento
cerebral que pode ser encontrado ao nascimento em humanos e 7ª dia embrionário em ratos
(SEMPLE et al., 2013).
1.2.4 Áreas cerebrais estudadas
O cérebro de uma pessoa com autismo apresenta falha de comunicação entre os
neurônios, dificultando o processamento de informações. Apresenta alterações principalmente
no corpo caloso, que é responsável por facilitar a comunicação entre os dois hemisférios do
cérebro, a amígdala, responsável pelo comportamento social e emocional e o cerebelo, que está
envolvido com as atividades motoras, como o equilíbrio e a coordenação.
Moraes et al., (2014) descrevem as principais alterações no cérebro autista da
seguinte forma:
Hipocampo: é uma estrutura alinhada a formação da memória. É uma área de
armazenamento e formação de memórias de longo prazo (STEPHAN et al., 1983). Crianças
com autismo, com ou sem retardo mental, apresentam um maior volume do hipocampo direito
do que não autistas (NACEWICZ et al., 2006 apud MORAES et al., 2014).
Amígdala: é uma estrutura complexa adjacente ao hipocampo, envolvida no
processamento de emoções e do medo, coordena respostas fisiológicas com base nas
informações cognitivas, isso porque ela liga áreas do córtex com o sistema hipotalâmico e os
sistemas do tronco encefálico que coordenam respostas metabólicas (MAREN et al., 2005). A
amígdala em crianças com autismo é inicialmente maior, entretanto, não continua a crescer com
o desenvolvimento da idade, como é com os não autistas. Foi encontrado também correlação
entre o volume da amígdala e a gravidade do quadro clínico (NACEWICZ, et al 2006; POLŠEK
et al., 2011 apud MORAES, 2014).

53
Córtex pré-frontal: é uma área importante para diversas funções. É uma parte
crítica do sistema executivo que se refere à capacidade de planejar, raciocinar e julgar. Está
envolvido também no desenvolvimento da personalidade, nas emoções e na capacidade de
exercer avaliação e controle adequado dos comportamentos sociais (FUSTER et al., 2008). Os
padrões de maturação do córtex pré-frontal em crianças autistas é mais lento, o que é consistente
com o desempenho cognitivo dos mesmos (SIQUEIRA et al., 2016).
Cerebelo: Anormalidades anatômicas do cerebelo foram amplamente observados
em pacientes com autismo (BAUMAN; KEMPER, 1985). As anormalidades relatadas incluem
hipoplasia dos lóbulos verticais cerebelares VI e VII (COURCHESNE et al., 1994), que foram
negativamente correlacionados com o aumento de comportamentos repetitivos e
comportamento exploratório diminuído em crianças com autismo (PIERCE; COURCHESNE,
2001). Uma ampla revisão foi publicada explicando os fatores patológicos envolvimento do
cerebelo no desenvolvimento do autismo, onde foi destacado que em algumas pessoas com
TEA, existem anormalidades cerebelares que apresentam defeitos anatômicos, inflamação,
estresse oxidativo, anormal neurotransmissores e níveis de proteínas e relacionados ao cerebelo
comprometimentos motores e cognitivos (FATEMI et al., 2012).
1.3 Ácidos Graxos Poliinstaurados
Óleos: Ômega 9 (oléico), Ômega 6 (linoléico) e Ômega 3 (docosahexaenóico,
eicosapentaenóico e linolênico)
Esses óleos são ácidos graxos (AG) que são inicialmente isolados de fontes naturais,
principalmente de lipídios, e são ácidos carboxílicos monocarboxílicos que podem ser
representados pela forma RCOOH (na maioria das vezes o agrupamento R é uma cadeia
carbônica) e são classificados de acordo com: o tamanho da cadeia hidrocarbônica (curta – até
quatro átomos de carbono; média – de seis a dez átomos de carbono; e longa – mais de dez
átomos de carbono); presença de insaturações - ligações duplas entre átomos da cadeia R -
(saturados – não possuem insaturações na cadeia hidrocarbônica; insaturados – possuem
insaturações) e quantidade de insaturações (monoinsaturadas – apenas uma ligação dupla;
poliinsaturadas – duas ou mais ligações duplas) (DAHELE et al., 2006). Sendo assim os óleos
ômega em estudo estão classificados conforme o Quadro 1.

54
Classe Nome sistemático Nome comum Carbonos* Duplas ligações**
Ômega-3 Docosahexaenóico DHA 22 6
Ômega-3 Eicosapentaenóico EPA 20 5
Ômega-3 Octadecatrienóico Linolênico 18 3
Ômega-6 Octadecadienóico Linoleico 18 2
Ômega-9 Octadecenóico Oleico 18 1*
Quadro 1 – Classificação dos óleos ômega.
Fonte: Campelo (2018). Legenda: * Número de Carbonos, ** Número de Duplas ligações, # Monoinsaturado.
Os ácidos graxos poliinsaturados (PUFA) são ácidos graxos categorizados em
vários grupos de acordo com a estrutura química e a posição da primeira dupla ligação contada
a partir do terminal metil; ômega-3 (n-3), n-6, n-7 e n-9 (BERG et al., 2007; GÓMEZ et al.,
2011; ANDERSON et al., 2009) (Figura 4). Os componentes mais importantes são os ácidos
graxos essenciais: ácido linoléico, ômega-6 (w-6 – LA) e ácido alfa-linolênico, ômega-3 (w-3
– ALA) que devem ser ingeridos obrigatoriamente por meio da dieta ou suplementação por não
poderem ser sintetizados pelo organismo (BERG et al., 2007; SURETTE et al., 2008; GÓMEZ
et al., 2011). Estes últimos compartilham das mesmas vias metabólicas e oxidativas; no entanto,
os seus produtos metabólicos finais têm efeitos fisiológicos antagônicos (COOK et al., 2004).
Figura 4 – Estrutura dos ácidos graxos insaturados.
Fonte: Gowdak, et al. 2015
Os ácidos graxos essenciais (AGEs) são obtidos de plantas ou outros organismos
que possuem vias enzimáticas para sua síntese. Os AGEs são, portanto, nutrientes dietéticos
essenciais para a sobrevivência de humanos e outros mamíferos devido à ausência das enzimas

55
dessaturase (∆) 12 e ∆ 15 necessárias para inserir uma ligação dupla na posição n ω-6 ou -3 de
uma cadeia de carbono de ácidos graxos (INNIS et al., 2003).
Após serem consumidos exogenamente (dieta ou suplementação), o LA e o ALA
são metabolizados no fígado pelas enzimas ∆6 e ∆5 dessaturases para gerar PUFAs de cadeia
longa (LCPUFAs) como ácido araquidônico (AA, 20:4, ω-6), ácido eicosapentaenóico (EPA,
20:5, ω-3) e ácido docosahexaenóico (DHA, 22:6, ω-3). Os produtos metabólicos enzimáticos
de 20 LCPUFAs de carbono são chamados eicosanóides e incluem as prostaglandinas,
tromboxanos e leucotrienos (Figura 5): em geral, os eicosanóides derivados do AA têm
principalmente propriedades pró-inflamatórias, enquanto os eicosanóides derivados do EPA
são anti-inflamatórios (KIECOLT et al., 2003).
Figura 5 – Processos de dessaturação e elongação do ômega-3 e ômega-6 e consequente geração de compostos bioativos. Fonte: Adaptado: Verma, O.P. Flaxseed - Miraculous Anti-ageing Divine Food M.B.B.S.,M.R.S.H. (London) President, Flax Awareness Society 7-B-43, Mahaveer Nagar III, Kota (Raj.) Published in Dr India April, 2013.
Além disso, outros autores identificaram um novo grupo de mediadores
denominados resolvinas das séries E e D, formados a partir de EPA e DHA, respectivamente.
Juntamente com a neuroprotectina D1 formada a partir de DHA por várias reações, esses
mediadores parecem exercer fortes efeitos de resolução da inflamação (KIECOLT et al., 2003).
Isso sugere que os LCPUFAs formam precursores de moléculas pró e anti-inflamatórias, e o

56
equilíbrio entre esses compostos mutuamente antagônicos pode determinar o resultado final do
processo da doença (DAS et al., 2006).
O ácido ômega-3 docosahexaenóico (DHA) e o ácido eicosapentaenóico (EPA),
além do ácido ômega-6 araquidônico (AA), são três LC-PUFAs de particular importância para
o desenvolvimento e funcionamento ideal do cérebro, com efeitos independentes e
compartilhados (DYALL et al., 2015), são incorporados aos fosfolipídios da membrana e a
incorporação do DHA ocorre em níveis excepcionalmente altos no sistema nervoso central
(LAURITZEN et al., 2016).
Esse aumento no cérebro inicia-se durante o surto de crescimento cerebral no
estágio intra-uterino, particularmente no último trimestre do desenvolvimento fetal e continua
até os dois anos de idade (CARVER et al., 2001), quando o cérebro da criança sofre um rápido
aumento no crescimento e proliferação celular, atingindo 80% do seu tamanho adulto
(CLAYOTN et al., 2001). Durante esse período, conhecido como "surto de crescimento
cerebral", há um aumento de aproximadamente 30 vezes no conteúdo de DHA (LAURITZEN
et al., 2001). Durante a infância, o DHA continua a se acumular no cérebro, em associação com
a maturação da substância cinzenta do córtex frontal (McNAMARA et al., 2015).
O suprimento intrauterino de PUFAs ocorre através de ácidos graxos polinsaturados
não esterificado, derivado principalmente da circulação materna através da placenta
(HERRERA et al., 2002). O aumento pós-natal de LCPUFAs nos tecidos infantis é suportado
pela transferência materna de LCPUFAs através do leite materno, e os níveis sanguíneos de
LCPUFAs em bebês amamentados permanecem mais elevados do que os níveis maternos por
algum tempo após o nascimento (IORGENSEN et al., 2006).
Comparado ao DHA, a concentração de EPA nos fosfolipídios cerebrais é
tipicamente 250 a 300 vezes menor (DYALL et al., 2015). No entanto, o EPA, mas não o DHA,
é um precursor dos eicosanóides das séries 3 e 5, que são mediadores lipídicos capazes de
promover efeitos benéficos, como anti-inflamação, vasodilatação e anti-agregação. A
biossíntese de eicosanóides derivados do EPA requer as enzimas ciclooxigenases (COX) e
lipoxigenases (LOX). Essas mesmas enzimas também estão envolvidas na formação dos
eicosanóides das séries 2 e 4 derivados do AA que tem principalmente efeitos pró-inflamatórios
e pró-agregadores (BAZINET et al., 2014; SCHUCHARDT et al., 2010). Consequentemente,
à medida que mais EPA está presente nas membranas celulares, menos quantidade de enzima
estará disponível para converter o AA em eicosanóides pró-inflamatórios. A razão AA / EPA
é, portanto, considerada um importante indicador de inflamação celular e a EPA pode atuar
como modulador da inflamação nos tecidos cerebrais (SONG et al., 2016). Por outro lado,

57
devido ao seu maior tamanho espacial, o DHA é um substrato pobre para as enzimas COX e
LOX (SONG et al., 2016). O DHA, no entanto, também pode exercer efeitos anti-inflamatórios
através da produção dos 'Mediadores Especializados em Pro-resolução', uma classe
recentemente identificada de mediadores lipídicos que evitam a inflamação excessiva e
promovem a resolução ativa das respostas inflamatórias (MOZAFFTARIAN et al., 2012).
Metabolizados pela ciclooxigenase (COX) e 5-lipoxigenase (5-LOX), os
eicosanoides NBR 6023:2018 (prostaglandinas, tromboxanas e leucotrienos) medeiam reações
inflamatórias (TASSONI et al., 2008). Os PUFAS disponibilizam precursores para a síntese de
prostaglandinas e modulam e expressão de enzimas fundamentais para o metabolismo destes
eicosanóides, bem como de esteróides (TRAN et al., 2016). De acordo com Xu et al., (2017),
uma razão DHA/EPA baixa na dieta, ocasiona efeitos inibitórios sobre a estereoidogênese
gonadal.
O DHA é o PUFA mais abundante nas membranas celulares fotorreceptoras da
retina e na substância cinzenta do cérebro, representando 15-20% da composição total de ácidos
graxos no córtex frontal (FLIESLER et al., 1983; MCNAMARA et al., 2015). Assim, o DHA
provavelmente está envolvido em funções executivas, como planejamento, atenção concentrada
e regulação comportamental, conhecidas por estarem localizadas principalmente no lobo frontal
(TESEI et al., 2016).
O AG ômega-9 (ácido oléico) confere proteção contra a peroxidação lipídica
diferente dos ácidos graxos polinsaturados como linolênico, EPA, DHA que contêm 4, 5 e 6
duplas ligações respectivamente são muito menos estáveis (CURI et al., 2002). O uso de
formulações ricas em lipídios na forma monoinsaturada, comparado ao uso de poliinsaturados
evidenciou menor resposta inflamatória e menor produção de radicais livres com fórmula rica
em monoinsaturados (CURI et al., 2002).
Sabe-se que existe uma comorbidade significativa dentre os que apresentam
Transtorno do espectro do autismo (TEA), distúrbio de déficit de atenção / hiperatividade
(TDAH), dificuldade de aprendizagem (LD) e deficiência intelectual (DI). Dentre os distúrbios
neurodesenvolvimentais mais comuns nestes indivíduos acometidos, e que todos eles têm um
componente herdável, foi levantada a hipótese de que distúrbios neurodesenvolvimentais e
psiquiátricos podem compartilhar fatores de risco no nível de predisposição biológica
(RICHARDSON et al., 2006).
Estudar alterações na composição dos PUFAs e a relação dos seus mediadores pode
ser uma estratégia importante para entender a fisiopatologia dos distúrbios do espectro do
autismo (TEA) (WONG et al., 2014). Especificamente, nos últimos 15 anos, foi dado

58
considerável interesse ao papel potencial das deficiências de ácidos graxos poliinsaturados
(PUFA) para a compreensão patogênica desses distúrbios e, posteriormente, ao possível papel
dos PUFAs como adjuvantes ao tratamento farmacológico (TESEI et al., 2017).
Devido aos seus efeitos antioxidantes e anti-inflamatórios, o ômega-3 foi estudado
nessas doenças. Atua na formação da bainha de mielina, no desempenho cognitivo e na
comunicação entre células neurais. Também está presente na ação vasodilatadora e, portanto,
aumenta o suprimento de oxigênio e nutrientes ao sistema nervoso central (SNC) (MANKAD
et al., 2015; EL-ANSARY et al., 2014).
Considerando a semelhança entre epilepsia e TEA, a suplementação de PUFA
ômega-3 na dieta pode levar a uma melhora dos sintomas dessa síndrome (JAMES et al., 2011).
A suplementação com PUFA ômega-3 foi estudada no tratamento de distúrbios neurológicos
como o TEA, que não é considerado uma doença única, mas está associado a várias etiologias
e graus variados de gravidade. O TEA é caracterizado por alterações comportamentais, déficits
de linguagem e cognição, deficiências intelectuais em 70% dos casos e convulsões em 30%
(VALVO et al., 2013). O estudo em modelos animais de epilepsia com suplementação de EPA
e DHA permitiu que as doses de tratamento farmacológico fossem reduzidas na fase de
manutenção da doença, reduzindo as convulsões do hipotámo (MUSTO et al., 2011).
As membranas neuronais, constituídas principalente por AGPIs ômega-3, modulam
inúmeras vias e mecanismos biológicos, dentre os quais a fluidez de membrana da
neuroinflamação, apoptose e senescência, expressão gênica, função e plasticidade sináptica
(ABEDI; SAHARI, 2014).
1.4 Testes Comportamentais em Modelo Animal versus Sinais do TEA
A tentativa de modelar distúrbios neuropsiquiátricos, como o TEA, em roedores é
extremamente desafiadora, porque os comportamentos equivalentes aos dos roedores como
comunicação social e dos fenômenos mentais que caracterizam o TEA são muito limitados e
não existem biomarcadores validados para roedores com TEA. Por exemplo, fenômenos como
ecolalia, reversão de pronome, fala metafórica e comprometimentos pragmáticos não têm
equivalência com os roedores. Da mesma forma, deficiências na comunicação não verbal e
fenômenos mentais como reconhecimento de emoções e 'mentalização' não são passíveis de
modelagem. Além disso, embora modelos animais possam exibir vários fenômenos em testes
comportamentais que são pensados para se assemelhar aos recursos do autismo, eles não são
necessariamente equivalentes às deficiências relatadas em humanos (TAKUMI et al., 2019).

59
Para que modelos animais em doenças humanas sejam considerados precisos, deve
haver uma avaliação de três tipos de validade (BAKER et al., 2011; SILVERMAN et al., 2010).
Primeiro, a validade de construção exige que tudo o que causa sintomas no animal seja o mesmo
que caracterize a doença em humanos. Segundo, a validade da face requer que os sintomas em
um modelo animal espelhem os do paciente humano. Terceiro, a validade preditiva avalia se o
animal responde às drogas de uma maneira que corresponde às reações humanas; no entanto,
para o autismo, não existem drogas eficazes que sirvam como controle positivo.
Para estimativa de manifestações externas de distúrbios em modelos animais de
autismo, certas técnicas de teste de comportamento são utilizadas. Testes clássicos universais
(teste de campo aberto, morris labirinto de água, labirinto tipo T, labirinto radial, de Skinner
câmara) são métodos para a detecção de características dos distúrbios autistas. Frequentemente,
o comportamento do animal é observado em condições em que esses animais são mantidos em
ambientes que possam se adapatar, imediatamente nas gaiolas onde eles geralmente vivem.
Combinação e modificação de certas técnicas são convenientes; permitindo que os avaliadores
possam fornecer uma completa avaliação detalhada das manifestações fenotípicas nos
respectivos modelos (HRABOVSKA et al., 2016).
Em crianças, a exposição pré-natal ao ácido valpróico anticonvulsivante (AVP)
pode aumentar em até 10 vezes o risco de desenvolver TEA (RASALAM et al., 2005). Apoiada
por essa evidência clínica, a administração pré-natal de AVP no 12,5ª dia gestacional, e 14ª dia
pós-natal tem sido amplamente utilizada com um modelo de roedor de TEA (RIBEIRO et al.,
2020).
Os animais com modelo de autismo mostram uma redução na sociabilidade
(LUCCHINA; DEPINO, 2014) e comportamentos estereotipados aumentados (SCHNEIDER
et al., 2008), além disso, apresentam várias alterações celulares e moleculares também
observadas em indivíduos com TEA (LUCCHINA; DEPINDO, 2014). Curiosamente,
diminuição da sociabilidade, aumento de comportamentos repetitivos e comportamentos
relacionados à ansiedade foram observados em machos expostos ao AVP (KIM et al., 2013;
LUCCHINA; DEPINO, 2014; SCHNEIDER et al., 2008), mas não em fêmeas expostas no pré-
natal com AVP (KATAOKA et al., 2013; KIM et al., 2013; SCHNEIDER et al., 2008).
Pode ser observado que vários autores procuram demonstrar através de seus estudos
respostas que possam elucidar em modelos animais, em ratos ou camundongos machos e/ou
fêmeas, os parâmetros clínicos, sinais e sintomas comportamentais mais parecidos com as
possíveis características no indivíduo humano com TEA descritas no DSM V (2013), e desta

60
forma, encontrar meios de amenizar e/ou diminuir as respostas negativas que impossibilitam
estes indivíduos de terem uma independência familiar no futuro.
Tendo em vista a importância do uso de nutracêuticos, estudos que os utilizem
podem vir a ser realizados com o intuito de elucidar formas alternativas ao tratamento para
amenizar os possíveis efeitos da neuroinflamação, do estresse oxidativo, e alguns fatores
epigenéticos do TEA com dietas e/ou suplementos ricos em ácidos graxos poli-insaturados
(AGPI), (PINHEIRO et al., 2011) visando atenuar processos inflamatórios e pró já em curso
(CAMPELO, 2012).
No presente estudo foram utilizadas misturas de óleos em concentrações
nutracêuticas, com alta razão de w-9/w-6 com ação anti-oxidante e baixa razão de w-6/w-3 que
favorece uma ação anti-inflamatória, conforme estudos apresentados em outras situações
biológicas (ALLAYEE; ROTH; RODIS, 2009; SIMOPOULOS 2002, 2008). O uso destas
misturas, por via orogástrica, tem sido implementado após a instalação da lesão (CAMPELO,
2012) ou como pré-condicionante, ou seja, antes da geração do evento, seja ele trauma
(NAKAMURA et al., 2005), isquemia/reperfusão (PINHEIRO et al., 2011) ou indução de
inflamação crônica (VASCONCELOS, 2013).

61
2 OBJETIVO
2.1 Objetivo Geral
Analisar o efeito do nutracêutico de misturas de óleos ômega 3, 6 e 9 para atenuar
as características comportamentais e neuroquímicas do transtorno do espectro autista induzidas
pelo ácido valpróico em modelo experimental.
2.2 Objetivos Específicos
a) Avaliar o efeito dos Mix-1, Mix-2 e Mix-3 nas alterações comportamentais e
locomotoras por meio dos testes de Campo Aberto, Imersão de Cauda, Performance da Natação
e Rota Rod.
b) Analisar o efeito dos Mix-1, Mix-2 e Mix-3 sobre o estresse oxidativo (TBARS), da
relação nitrito/nitrato, e nas alterações da atividade enzimática dos sistemas antioxidantes
(GSH) no hipocampo, córtex pré-frontal, amígdalas e cerebelo de ratos.
c) Investigar o efeito do Mix-3 sobre a expressão dos genes Dnmt3 e Chd2 em amostras
do hipocampo de ratos, através da biologia molecular RT-qPCR.

62
3 MÉTODO
3.1 Aspectos éticos e Animais
Foram utilizados ratos da linhagem Wistar machos (120 animais) e fêmeas (96
animais) pesando entre 160-260g, provenientes do Biotério Central da Universidade Federal do
Ceará - UFC. Os animais acasalados foram mantidos em caixas de polipropileno condicionado
um casal por caixa, e após o nascimento os filhotes foram randomizados de diferentes ninhadas
para aumentar a variabilidade genética, permanecendo com as mães até o desmame com 21 dias
de nascidos (Figura 6). Nesta idade os animais foram separados por sexo e colocados em caixas
com no máximo 07 animais, à temperatura média de 24 ± 2°C em ciclos de alternância
claro/escuro de 12 horas, recebendo ração padrão e água ad libitum. Durante todo o
experimento, os animais permaneceram em padrão sanitário controlado, onde foram mantidos
em caixas de polipropileno (50 x 50 cm) forradas com cama de maravalha. As caixas eram
trocadas todas as 2ª e 5ª feiras, bem como o recipiente de água e feito reposição de ração. O
protocolo experimental foi aprovado pelo Comitê de Ética em Pesquisa Animal (CEPA) da
Universidade Federal do Ceará (UFC) gerando o protocolo registrado com n°16/18. Cada um
dos grupos teve em média seis filhotes selecionados a partir da data do parto, os mesmos
passaram pelo processo de indução do TEA ainda filhotes no 14ª dia pós-nascimento, haja vista
a casuísta da enfermidade dar-se a partir da mais tenra infância.
Figura 6 – Animal com filhotes no biotério.
Fonte: Autora.

63
O protocolo experimental foi elaborado de acordo com o “Guia para Cuidados e
Uso de Animais de Laboratório” da Sociedade Brasileira de Ciência em Animais de Laboratório
(SBCAL) e previamente submetido ao Comitê de Ética no Uso de Animais (CEUA) da UFC
Campus Sobral. Todos os esforços foram feitos para minimizar o número e o sofrimento dos
animais utilizados.
3.2 Drogas e soluções
A suplementação anti-inflamatória e antioxidante foi preparada e armazenada
dentro do laboratório de neurofisiologia da Faculdade de Medicina Campus de Sobral. Os óleos
para elaboração: Mix-1 (ácido graxo linolênico–ALA), Mix-2 (ALA (35%) + ácido graxo
eicosapentaenoico (EPA–39%) + ácido graxo docosahexaenóico (DHA–26%) e Mix-3 (ALA-
84% + DHA-16%). A mistura destes Mixs foi desenvolvida e patenteada a partir de uma junção
de gorduras com características antioxidantes e antinflamatória estudadas em pesquisas com
ratos e camundongos sob orientação do pesquisador (VASCONCELOS, 2013). Os óelos foram
adquiridos no mercado local “lojas de suplementação e nutrição”. Para preparação destes mixes,
os óleos foram pesados utilizando uma balança analítica da marca METTLER TOLEDO 10 mg
a 52g, d = 0,0001g com calibração automática, e adicionados em um vidro escuro de 25 ml com
tampa e imediatamente armazenados sob refrigeração 20 a 24 graus (Figura 7).
Gorduras insaturadas
Figura 7 – Produção dos Mix-1, Mix-2 e Mix-3.
Fonte: Autora.
Mix-1 Mix-2 Mix-3
Balanças
Mixs prepara
Beckerrrr
Armazenamento dos Mixs

64
3.3 Delineamento Experimental
3.3.1 Distribuição dos Grupos de Estudo
O modelo de autismo foi induzido por injeção s.c. de AVP no dia 14ª dia pós-
nascimento seguido de terapia de suplementação de gordura ricas em ácidos graxos insaturados
distribuídos em Mix-1 (óleo de canola, óleo de girassol, óleo de linhaça – ALA (ácido graxo
linolênico), Mix-2 (óleo de canola, óleo de girassol, óleo de peixe – ALA (35%) + EPA (ácido
graxo eixopentaenóico – 39%) e Mix-3 (óleo de canola, óleo de girassol, óleo puro de algas
(DHA – ácido graxo docosahexaenóico – 16%) e ALA (84%). Os filhotes foram avaliados e
acompanhados quanto ao risco e/ou aparecimento de enfermidades pós-natal e alterações
comportamentais até 50ª dia pós-nascimento (Figura 8). No 55ª dia pós-nascimento, os animais
foram eutanasiados e as áreas cerebrais (hipocampo, córtex pré-frontal, amígdalas e cerebelo)
extraídas sem danos. Os tecidos cerebrais coletados foram utilizados para as determinações de
alterações bioquímicas, marcadores de estresse oxidativo, marcadores inflamatórios e
expressão gênica.
A droga AVP (Valproato de Sódio 250 ml) “DEPAKENE” foi adquirida no
mercado local (farmácias) e armazenada sob refrigeração. Abaixo podemos analisar as etapas
do delineamento experimental em várias etapas.
Figura 8 – Esquema do delineamento experimental
Fonte: Autora. Legenda: DPN (dia pós-natal); VPA (ácido valpróico).
Nascimento
Dia 1
Indução do modelo com VPA
14 DPN 15 DPN 40 DPN
50 DPN
55 DPN
Suplementação com mix (durante 25 dias)
30 DPN
32 DPN
Teste da imersão da
calda
Teste da natação
Teste do rotarod
24 DPN
Teste do campo aberto
Coleta de sangue

65
3.3.2 Tratamentos dos grupos experimentais
Os Mix-1, Mix-2 e Mix-3 (Figura 09, 10 e 11 - respectivamente), foram
administrados no 15ª dia pós-nascimento por um período de 25 dias na dose de 1,2 ml /kg p.o
consecutivos (conforme cronograma experimental – Figura 8) - tendo em vista que essa é a
dosagem nutracêutica já estabelecida na literatura em pesquisas prévias (MELO et al., 2014;
MELO et al., 2017), sempre no mesmo horário 24/24h por via orogástrica com ajuda de uma
cânula de inox (Figura 12). Essas doses diárias de 1,2g /kg/dia estavam distribuídas nos mixs
na proporção de 1,4:1 de ω-6/ω-3 (efeito anti-inflamatório) e de 3,7:1 de ω-9/ω-6 (efeito
antioxidante). O que diferencia os grupos tratados são as distintas fontes de ω-3: ALA, EPA e
DHA, provenientes de óleo de linhaça, óleo de peixe e algas, conforme esquematizados abaixo:
Figura 9 – Soluções e composição com as respectivas fontes de w3 (MIX 1)
Fonte: Autora. Legenda: ALA = ácido α-linolênico;
Figura 10 – Soluções e composição com as respectivas fontes de w3 (MIX 2)
Fonte: Autora. Legenda: ALA = ácido α-linolênico; EPA = ácido eicosapentaenoico; DHA =
ácido docosahexaenóico.
Fonte de W-3
Proporção
Composição
Solução MIX 1
Óleo de canola
Óleo de girassol
w-6: w-3 = 1,4:1
w-9: w-6= 3,7:1
ALA
Óleo de linhaça
Fonte de W-3
Proporção
Composição
Solução MIX 2
Óleo de canola
Óleo de girassol
w-6: w-3 = 1,4:1
w-9: w-6 = 3,7:1
ALA (35%) EPA (39%) DHA (26%)
Óleo de peixe

66
Figura 11 – Soluções e composição com as respectivas fontes de w3 (MIX 3)
Fonte: Autora. Legenda: ALA = ácido α-linolênico; DHA = ácido docosahexaenóico.
3.3.3 Divisão dos Grupos de Estudo e Análises Experimentais
Os grupos serão separados e selecionadas da seguinte forma:
• Grupo I (Controle Sham 0,9% Salina normal p.o.)
• Grupo II (Controlo Positivo, VPA 400 mg / kg s.c)
• Grupo III (MIX-1, 1,2 ml / kg p.o.)
• Grupo IV (MIX-2, 1,2 ml / kg p.o.)
• Grupo V (MIX-3, 1,2 ml / kg p.o)
• Grupo VI (VPA + MIX-1 400 mg / kg + 1,2 ml / kg p.o.)
• Grupo VII (VPA + MIX-2 400 mg / kg + 1,2 ml / kg p.o.)
• Grupo VIII (VPA + MIX-3 400 mg/ kg + 1,2 ml/kg p.o.)
Abaixo encontra descriminado o protocolo do tratamento experimental para
compreender a distribuição do número de animais por sexo utilizado em cada etapa:
comportamental, parâmetros neuroquímicos e da expressão gênica utilizado neste estudo
(Quadro 2). Em seguida (Quadro 3), um esboço esquematizando a divisão dos grupos
experimentais, tipo de tratamento recebido durante 25 dias, bem como a via de administração
das gorduras antioxidantes e anti-inflamatórias por meio dos mixs e dose do “Ácido Valpróico
- AVP” no modelo do animal com autismo.
Fonte de W-3
Proporção
Composição
Solução MIX 3
Óleo de canola
Óleo de girassol
w-6: w-3 = 1,4:1
w-9: w-6 = 3,7:1
ALA (84%) DHA (16%)
DHA puro de algas

67
Sham VPA MIX-1 MIX-2 MIX-3 MIX-1 +VPA
MIX-2 +VPA
MIX-3 +VPA
Dosagens Total
6 animais
6 animais
6 animais
6 animais
6 animais
6 animais
6 animais
6 Animais
TBARS NITRITO
48
6 animais
6 animais
6 animais
6 animais
6 animais
6 animais
6 animais
6 animais GSH
48
TOTAL DE MACHOS E FÊMEAS 192
7 animais
6 animais
- - 6
animais - -
6 animais
Chd2 Dnmt3a
25
TOTAL DE RATOS NO ESTUDO 217
Quadro 2 – Protocolo de Tratamento Experimental (FÊMAS - 96 animais) e MACHOS (121
animais), tipo de dosagens analisada.
Fonte: Autora.
GRUPOS EXPERIMENTAIS
M E F TRATAMENTO
DOSE (/KG)
VIA DE
ADM.
DURAÇÃO DO TRATAMENTO
(DIAS)
SHAM 0,9% Salina 0,9% 0 Oral 25 dias
AVP Ácido Valpróico (DEPAKENE)
400 mg s.c. 1 dia
MIX-1
Ω 9 + Ω 6 + Ω 3 Óleo de canola Óleo de girassol rico em oleico Óleo de linhaça
1,2 mg Oral 25 dias
MIX-2
Ω9 + Ω6 + Ω3 Óleo de canola Óleo de girassol rico em oleico Óleo de peixe
1,2 mg Oral 25 dias
MIX-3
Ω-9 + Ω-6 + Ω-3 Óleo de canola Óleo de girassol rico em oleico DHA puro de algas
1,2 mg Oral 25 dias
MIX-1 + AVP MIX-1 + Ácido Valpróico 1,2mg + 400mg
s.c oral
25 dias
Continua
Conclusão

68
GRUPOS EXPERIMENTAIS
M E F TRATAMENTO
DOSE (/KG)
VIA DE
ADM.
DURAÇÃO DO TRATAMENTO
(DIAS)
MIX-2 + AVP MIX-2 + Ácido Valpróico 1,2mg + 400mg
s.c oral 25 dias
MIX-3 + AVP MIX-3 + Ácido Valpróico 1,2mg + 400mg
s.c oral
25 dias
Quadro 3 – Protocolo de Tratamento Experimental (Droga + Dieta)
Fonte: Autora.
peso do animal
MIX 1/ MIX2 / MIX 3 X = P.1,2g / 968
volume administrado
Figura 12 – Quantidade de solução lipídica infundida diariamente
Fonte: Autora.
3.4 Testes Comportamentais
Os testes comportamentais foram realizados conforme esquema citado a seguir.
Antes de qualquer um dos procedimentos de avaliação comportamental, os animais eram
levados em suas caixas ao laboratório de fisiologia com 30 minutos de antecedência –
representando o período de habituação para que não houvesse interferências do ambiente. A
sala de comportamento ficava dentro dos acessos deste laboratório, onde antes de iniciar cada
teste, os animais eram deslocados individualemente. Todos os equipamentos e utensílios dos
testes comportamentais (Campo aberto e Rota road) foram limpos com álcool 70% após o teste
de cada animal, impedindo a permanência de odores.
Peso do Animal
Volume Administrado

69
• TESTE DE CAMPO ABERTO = 24ª dia pós-nascimento
• TESTE DE IMERSÃO EM CAUDA = 30ª dia pós-nascimento
• TESTE DE PERFORMANCE = 32ª dia pós-nascimento
• TESTE ROTA ROAD = 50ª dia pós-nascimento
3.4.1 Atividade Locomotora (Campo Aberto)
Este experimento foi realizado a fim de testar a atividade exploratória do animal
baseado na metodologia validada por Archer (1973) no 24ª dia pós-nascimento (Figura 13). Os
ratos foram colocados individualmente após o período de habituação, em um campo aberto com
área de 50 x 50 cm e em ambiente iluminado por uma luz vermelha. Este campo era
confeccionado em madeira com fundo preto, dividida em nove quadrantes iguais. Os animais
foram previamente habituados durante 1 minuto e, posteriormente, observados por 5 minutos
onde estava sendo observados e avaliado:
Locomoção: foram avaliados a frequência de cruzamentos (número de vezes eu o
animal cruzou as divisões marcadas no chão), uma medida de atividade exploratória.
Tempo parado nos quadrados: foi avaliado o tempo de permanência que o animal
ficou imóvel, um parâmetro de ansiedade.
Atividade de levantar (rearings): foi avaliada a frequência (número total de vezes
que o animal levantou sobre as patas dianteira e movimentou a cabeça numa postura típica de
orientação em um ambiente novo – Figura 13) (SILVEIRA et al., 2005).
Atividade de limpeza (grooming): é a atividade de esfregar o corpo com as patas ou
boca e esfregar a cabeça com as patas, num comportamento característico de limpeza (Figura
13). Foi contabilizado a quantidade de vezes que o animal ficou realizando esse comportamento
(HSIAO et al., 2012).

70
Figura 13 – Análise comportamental dos animais pelo Teste Campo Aberto.
Fonte: Autora. Figura criada na plataforma "Mind the Graph", disponível em: www.mindthegraph.com.
3.4.2 Teste de imersão na cauda
Neste modelo, um estímulo térmico entre 50º a 55ºC aplicado à cauda de ratos
desencadeia o reflexo da retirada da mesma por flexão predominantemente em direção rostral.
Etapa conhecida como efeito antinocicepção, sendo realizado no 30ª dia pós-nascimento
(Figura 14).
A avaliação das respostas é feita por meio da medida do lapso de tempo entre a
aplicação do estímulo térmico e o momento de afastamento da cauda da fonte de calor
(CARTENS et al., 1993). Esse modelo permite obter informações sobre o mecanismo e o local
da atividade antinociceptiva detectada, uma vez que o parâmetro avaliado, ou seja, o reflexo de
retirada da cauda, é de integração medular.
O teste foi realizado por meio da imersão do terço inferior da cauda dos animais em
água aquecida à 55°C, seguida da anotação do tempo de reação ao estímulo, através da
filmagem em câmera de celular. Este teste comportamental se baseia na premissa que a imersão
da cauda do animal na água provoca um movimento abrupto da cauda e, por vezes, o recuo de
todo o corpo; sendo o tempo de reação monitorado (SOUZA et al., 2014).

71
Figura 14 – Análise comportamental dos animais pelo método Imersão em Cauda.
Fonte: Autora. Figura criada na plataforma "Mind the Graph", disponível em: www.mindthegraph.com.
3.4.3 Teste de Performance de Natação
Em um recipiente cheio de água (28–29°C) os animais foram submetidos ao teste
de performance no 32ª dia pós-nascimento (Figura 15). Cada animal foi colocado no centro do
recipiente e avaliado por 10 segundos. A natação e o desempenho foram avaliados de acordo
com a posição do nariz e cabeça (ângulo) na superfície da água.
O ângulo da natação foi caracterizado da seguinte forma: 0 - cabeça e nariz abaixo
da superfície, 1- o nariz abaixo da superfície, 2- o nariz e topo da cabeça ou acima da superfície,
mas as orelhas ainda abaixo da superfície, 3- o mesmo que dois, exceto que a linha de água
esteja em nível do ouvido médio, e 4 o mesmo que três, exceto que a linha de água esteja no
fundo das orelhas. A natação avalia o desenvolvimento motor e consolidação de respostas de
reflexos coordenados (SCHAPIRO et al., 1970).

72
Figura 15 – Análise comportamental dos animais pelo método performance da natação. Fonte: Autora. Figura criada na plataforma "Mind the Graph", disponível em: www.mindthegraph.com.
3.4.4 Teste do rotarod
Este teste foi realizado no 50ª dia pós-nascimento para avaliar os aspectos de
coordenação e equilíbrio do desempenho motor em todos os grupos estudados. O aparelho
consiste em uma barra giratória com 3 cm de diâmetro presa a um motor com a velocidade
ajustada para 40 rpm, esquematizado na Figura 16. Os filhotes foram colocados
individualmente na barra giratória e observado o tempo gasto para permanecer sem cair bem
como a velocidade medida em RPM na barra giratória, tendo 300 segundos como o tempo de
corte (BHOSALE et al., 2011).
Para realização deste teste foi utilizado um período de adaptação (pré-teste) 48
horas antes do dia 50ª, seguindo o mesmo protocolo.

73
Figura 16 – Representação esquemática do teste comportamental Rotarod. Dois dias antes do
teste, os animais foram adaptados ao aparato por meio da permanência neste por 300 segundos
com baixa velocidade inicial, gradualmente crescente (4 a 15 RPM). No dia do teste, foi
utilizada a velocidade crescente de 4 a 40 RPM com duração máxima de 300 segundos. Foram
avaliados os seguintes parâmetros: Tempo de Latência (tempo até a primeira queda) e Tempo
Fonte: Autora. Figura criada na plataforma "Mind the Graph", disponível em: www.mindthegraph.com.
3.4.5 Dissecação das áreas cerebrais e descarte dos animais
Os animais após todas as determinações comportamentais no 55ª dia pós-
nascimento foram eutanasiados por guilhotina dentro do laboratório de
Fisiologia/Neurociência. Em seguida, os encéfalos foram rapidamente removidos e colocados
sobre papel alumínio em uma placa de Petri contendo gelo, seguido de dissecação das áreas
cerebrais a serem estudadas.
O hipocampo (HC), córtex pré-frontal (CPF), amígdalas e cerebelo, foram isolados
das demais áreas do encéfalo por divulsionamento com auxílio de uma pinça de
microdissecação. Cada área foi pesada, armazenada e identificadas em papel alumínio e
conservada a -80ºC até sua análise. As carcaças dos animais foram acondicionadas em sacos
plásticos, armazenadas em freezer e posteriormente o descarte ocorreu sob responsabilidade do
Biotério da Universidade Federal do Ceará/Campus Sobral.

74
3.5 Avaliação Neuroquímica
3.5.1 Determinação da peroxidação lipídica por medição do conteúdo de substâncias
reativas do ácido tiobarbitúrico (TBARS)
O grau de peroxidação lipídica foi determinado pelo método de Draper e Hadley
(1990). A determinação das concentrações de substâncias reativas do ácido tiobarbitúrico
(TBARS) foi avaliada nas áreas cerebrais (HC, CPF, amígdalas e cerebelo). Foram preparados
homogenatos das áreas cerebrais supracitadas a 10% em solução de tampão fosfato 150 mM.
Uma alíquota de 63 µL do homogeneizado foi retirado e precipitado com 100 µL de ácido
perclórico 35% dentro de um eppendorff, e posteriormente centrifugado à 5000 rpm por 10
minutos a 4 °C. Após centrifugação, foi separado 150 uL do sobrenadante e adicionado 50 µL
de ácido tiobarbitúrico 1,2 %. Posteriormente a mistura foi agitada e mantida em banho de
água fervente (95-100 ºC) por 30 minutos. Após isso, foi transferido 200 μL da amostra para os
poços de uma placa de poliestireno de 96 poços, a determinação do conteúdo de TBARS foi
realizada por espectrofotômetro a 535 nanômetros (nm) em leitor de microplacas
(MOLECULAR DEVICES –Spectra Max Paradigm Multi-Mode). Os resultados foram
expressos em micromol (μmol) de malonildialdeído (MDA) por miligrama de proteína. A curva
padrão de MDA foi preparada a partir de uma solução padrão nas concentrações 0,627; 1,247;
2,463; 4,8; 9,16 e 16,77 μmol/L e o branco foi feito com água destilada (Figura 17).

75
Figura 17 – Representação esquemática da determinação do estresse oxidativo por meio da
obtenção de TBARS.
Fonte: Autora.
3.5.2 Determinação da Concentração de Nitrito nas Áreas Cerebrais e no Sangue
Para a determinação da produção de nitrito, primeiramente foi preparado uma curva
padrão. Para isso, foram pesados 6,9 mg de NaNO2 e dissolvidos em 10 mL de água bidestilada
(estoque-10mM) e feitas as diluições em série (10 e 20 x), ficando 1 μM, 100 μM, 10 μM, 5
μM, 2,5 μM, 1,25 μM, 0,625 μM, 0,312 μM. Foi realizada uma equação da reta para o cálculo
das concentrações do teste (GREEN et al.,1981). Para a determinação da concentração de nitrito
em cada tecido, foram preparados homogenatos das áreas cerebrais a 10% (p/v) em solução
tampão fosfato 150 mM. Após a centrifugação (11000 rpm/15 min A 4ºC) os sobrenadantes
foram coletados e a produção de óxido nítrico (NO) determinada através da reação de Griess.
Uma alíquota de 50 μL do sobrenadante foi incubada com 50 μL do reagente de Griess
[(sulfanilamida 1% em H3PO4/ naftiletilenodiamino-bicloridrato 0,1%/, H3PO4 1%/diluído
em água, na proporção de (1:1)] e foram distribuídos em uma placa de poliestireno de 96 poços
sob a temperatura ambiente por 10 minutos. Após 10 minutos de reação a absorbância foi
medida em espectrofotômetro a 570 nm em leitor de microplacas (MOLECULAR DEVICES –
Spectra Max Paradigm Multi-Mode). A concentração de nitrito (μM) foi determinada a partir
de uma curva padrão de NaNO2 e os resultados foram expressos em μM de nitrito-nitrato por
grama de tecido (Figura 18).
8 grupos de animais M e F
Hipocampo Cortex Pré-frontal
Amígdala Cerebelo
Peroxidação Lipídica (Tbars) mMol/g de tecido
Dissecados
Placa Elisa
Leitura Elisa Banho-maria
Homogenatos

76
Figura 18 – Representação esquemática da determinação do estresse oxidativo por meio da
obtenção de Nitrito.
Fonte: Autora.
3.5.3 Determinação da Concentração de Glutationa Reduzida GSH)
A atividade antioxidante foi determinada pela concentração de GSH, que se baseia
na reação do reagente de Ellman (5,5'-ditiobis-ácido 2-nitrobenzóico) (DTNB) com o tiol livre
originando um dissulfeto misto mais ácido 2-nitro-5-tiobenzóico. Inicialmente foi preparado o
homogenato a 10% em EDTA 0,02M das áreas cerebrais em estudo, em seguida foi retirado 40
μL desse homogenato e adicionado 50 μL de água destilada e 10 μL de ácido tricloracético a
50%. O material foi agitado e centrifugado a 5000 rpm por 15 minutos a 4 ºC. Posteriormente,
foi recolhido 60 μL do sobrenadante e acrescido 102 μL de tampão Tris-HCL 0,4 M, pH 8,9 +
DTNB 0,01M e distribuídos na placa de poliestireno de 96 poços, após 1 minuto da reação foi
feito a leitura da coloração em 412 nm em leitor de microplacas (MOLECULAR DEVICES –
Spectra Max Paradigm Multi-Mode), (Figura 19). A concentração de glutationa reduzida foi
expressa em nanogramas de GSH/g de tecido, tendo por base uma curva padrão. A curva padrão
foi construída a partir da solução padrão de GSH (1mg/mL), a qual foi preparada em triplicata
de soluções a 6,25; 12,5; 25; 50 e 100 μg/mL. O branco foi feito com água destilada (4 mL) e
a cada tubo das soluções de GSH foi acrescentado de 4 mL de tampão Tris HCl 0,4M (pH 8,9).
Para a determinação da equação da curva padrão de GSH, foi adicionado ainda a cada tubo 0,1
mL de DTNB (SELDLAK; LINDSAY, 1988).
Dissecados
Hipocampo Cortex Pré-
frontal Amígdala Cerebelo
100 µL do reagente de Griess
(Sulfanilamida 0,1% em ac.
fosfórico 5% e Água destilada +
100 µL do homogenato
NITRITO mMol/g de tecido
Placa Elisa
Homogenatos
Leitura
Elisa
8 grupos de animais M e F

77
Figura 19 – Representação esquemática da determinação do estresse oxidativo por meio da
Concentração de Glutationa Reduzida
Fonte: Autora.
3.6 Análise de Expressão Gênica
3.6.1 Desenho e análise dos primers
O planejamento dos primers foi baseado nas sequências genéticas codificantes
identificadas tanto para genes de referência (Actb e Gapdh), quanto para genes alvos de
avaliação a resposta comportamental utilizando o banco de dados do NCBI -
https://www.ncbi.nlm.nih.gov). O desenho dos primers deu-se através do software Primer 3,
onde foram validadas in silico temperaturas de anelamento em torno de 60ºC, com tamanhos
de amplicons variáveis entre 84 pb e 147 pb para as sequencias-alvo estudadas. As sequências
dos primers selecionados encontram-se apresentadas na Tabela 1. Esse material foi analisado
quanto às probabilidades de formação de hairpins omodímeros e heterodímeros, estruturas que
podem prejudicar a eficiência de amplificação durante as reações de qPCR. Os níveis de
expressão dos genes foram avaliados a fim de escolher os melhores tecidos para desenho das
curvas-padrão.
Hipocampo
Cortex Pré-frontal Amígdala
Cerebelo
Centrifugar 5000 rpm por 15 min 4 °C
GSH mMol/g de
tecido
Dissecados
40 °µldohomogenato+50µldeáguadestilada+10µldeTCA50%
Tris EDTA + DTNB
Leitura Elisa
Homogenatos
8 grupos de animais M e F

78
3.6.2 Expressão Gênica do Hipocampo
Diversos trabalhos têm estudado os efeitos da administração de ácidos graxos w-3
sobre o hipocampo de ratos, desde experimentos in vitro, verificando sua atividade anti-
inflamatória em fatias de hipocampo (MOREIRA et al., 2010), até sua ação protetora in vivo
com inibição de atividade epileptiforme (YONG et al., 2000). Puskás et al., (2003)
demonstraram que a administração oral de óleo de peixe em ratos idosos acarreta um aumento
do DHA e uma drástica redução do AA na composição das membranas fosfolipídicas dos
neurônios do hipocampo. De forma similar, Lamaziere et al., (2011) evidenciaram que a
provisão exógena de ácidos graxos w-3, por meio da administração orogáastrica de óleo em
ratos, resulta em sua incorporação diferenciada nas membranas fosfolipídicas dos neurônios,
principalmente em determinadas regiões do cérebro, como o córtex e o hipocampo. Desta
forma, neste trabalho a área escolhida para o estudo da expressão gênica foi o hipocampo de
ratos em 4 grupos citados a seguir.
Em uma outra série de experimentos, grupos de animais tiveram o hipocampo
coletado após o tratamento para análise da expressão gênica. O material do hipocampo foi
coletado ao final de tratamento de quatro grupos experimentais: SHAM (n=7), AVP (n=6),
Mix-3 (n=6) e AVP + Mix-3 (n=6). O grupo do Mix-3 foi escolhido para realização da
expressão gênica na fase da pesquisa em a maioria dos resultados obtidos apresentavam-se
significativamente favoráveis aos achados esperados nesta pesquisa. Ao final da
experimentação os animais foram eutanasiados, conforme descrito na sessão “4.5.4 Dissecação
das áreas cerebrais e descarte dos animais”, o hipocampo foi dissecado, lavado duas vezes em
solução salina e transferido para tubo previamente identificado contendo solução de RNA later
(ThermoFisher Scientific), onde permaneceram submersos por 1h a temperatura ambiente, em
seguida esse material foi congelado a -30 ºC, transportado para unidade de realização
experimental e mantido a -30 ºC até o momento da extração de RNA.
3.6.3 Extração de RNA total de hipocampo
Todos os procedimentos que envolveram a manipulação de RNA foram realizados
utilizando materiais dedicados exclusivamente a esta finalidade. A superfície das bancadas, os
equipamentos e as micropipetas utilizadas foram higienizados e tratados com o spray “RNAse”
conforme recomendado pelo fabricante. Utilizando a extração do RNA total foi realizada o
reagente Trizol® (ThermoFisher Scientific) de acordo com as recomendações do fabricante. A

79
ressuspensão do RNA total foi realizada com 50 µL de água mili-Q DEPC nuclease free. O
RNA total foi avaliado e quantificado equipamento NanoDrop 1000 (Thermo scientific®) e as
amostras foram armazenadas em freezer -80ºC até sua utilização.
Rattus novergicus
Código NCBI Gene Primers Produto (pb)
NM_001107523.1 Chd2* F: GGAAAGGGGAGAGAGAATGG
R: ACTTTGGCAGGAAGGGATTT 104
NM_001003957.1 Dnmt3a* F: GAACGGCTCGTGTATGAGGT
R: TGAAGAGTGGGTGCTCCAG 101
NM_031144.3 Actb# F: CCTGAACCCTAAGGCCAACC
R: GCCATGTACGTAGCCATCCA 84
Tabela 1 - Predição da expressão gênica por tecido, sequência de primers utilizados, alvos e
tamanhos dos produtos amplificados esperados.
Fonte: Autora. Legenda: A = Alta; MB = Muito Baixa; B = Baixa; M = Média. *Primer3; **NCBI; #Primer Mus musculo com homologia previamente verificada in sílico
3.6.4 Transcrição reversa obtenção de cDNA PCR quantitativo em Tempo Real (RT-
qPCR)
Ensaios de RT-qPCR foram realizados para a quantificação relativa dos níveis
transcricionais dos genes Dnmt3a e Chd2. As etapas de transcrição reversa e de amplificação
foram realizadas separadamente (Método Two-Step). As amostras de RNA total extraídas foram
submetidas à reação de transcrição reversa com o uso do kit “High Capacity cDNA Master
Mix” ThermoFisher Scientific A estratégia obtenção cDNA a partir do RNA baseia-se no
anelamento de oligonucleotídeos de sequências aleatórias. A cada reação foram adicionados 10
µL de “RT Buffer 2X”, 1 µL de “RT Enzyme Mix”, cDNA, 5 µL de amostra de RNA total e
água mili-Q livre de RNases em um volume final de 20 µL. Para todos os grupos foram
utilizados 5 uL da solução de RNA total. As amostras foram incubadas à 37 ºC durante 60 min
e depois a 95 ºC por 5 min em termociclador “CFX96 Touch Real-Time PCR Detection System
(Biorad). As amostras contendo o DNA complementar (cDNA) gerado foram quantificadas,
diluídas para concentração de 50 ng/uL e mantidas em freezer -20 ºC até sua utilização.
As reações de qPCR para os genes alvo foram realizadas em triplicata utilizando
como método de detecção o reagente intercalante de DNA Sybr Green utilizando o “PowerUp
SYBR® Green PCR Master Mix” de acordo com as recomendações do fabricante. A eficiência

80
dos primers foi avaliada através de uma curva de diluição seriada do cDNA, os quais
apresentaram eficiência entre 95 e 105%. A ciclagem de temperatura seguiu com um
aquecimento inicial a 95 ºC por 5 min e 60ºC durante 60 min, seguidas da curva de melting a
60oC por 1 minuto seguida de 95oC por 15 segundos (+0.3 oC). Os ensaios de qPCR foram
realizados replicata biológicas para confirmação dos resultados. Os resultados foram avaliados
utilizado o método de 2ˆddCq de acordo com Livak and Schmittgen (2001)
3.7 Análises estatísticas (comportamentos, estresse oxidativo, GSH e Nitrito)
O teste de Shapiro-Wilk foi utilizado para verificar se os dados de cada variável
analisada apresentam distribuição normal, de modo que os valores outliers foram retirados (p-
valor > 0,05). Para identificação dos valores atípicos (outliers) foi utilizado o método de ROUT
com Q-value de 10%. Os dados paramétricos foram analisados através do teste t de Student,
para a comparação da média entre dois grupos, e de ANOVA para a comparação das médias
em variáveis com mais de dois grupos. Em caso de homogeneidade de variâncias (p-valor ≥
0,05), foi definido como pós-teste para a ANOVA o teste de Tukey. Os dados não-paramétricos
foram expressos como mediana, seguida de seus valores extremos, e analisados pelo teste de
Kruskal-Wallis.
Para os dados paramétricos e não-paramétricos o nível de significância estatística
utilizado será de p<0,05 e as análises estatísticas serão realizadas utilizando os softwares SPSS
20.0 (SPSS Inc., Chicago, IL, EUA) e GraphPad PrismÒ versão 6.0 (La Jolla, CA, EUA).
3.7.1 Análise estatística (Expressão Gênica)
Os resultados de dois experimentos indepententes, realizados em triplicatas foram
expressos em Cq ± desvio padrão por animal. A estatística descritiva foi apresentada média ±
desvio padrão por grupo. O teste de normalidade KS foi realizado. As medianas dos grupos não
normais foram submetidas ao teste não paramétrico Mann-Whitney para a comparação pareada
entre os grupos experimentais, comparado ao controle SHAM.
As médias dos grupos foram submetidas ao teste t não pareado para a comparação
paramétrica entre a média dos grupos experimentais, comparados ao controle SHAM.

81
4 RESULTADOS
4.1 Análises Comportamentais em MACHOS e FÊMEAS
4.1.1 Avaliação da Atividade Locomotora - Teste do Campo Aberto
Esta tarefa foi utilizada como um parâmetro de avaliação do comportamento do tipo
ansioso e da atividade locomotora dos animais, assim como da memória de habituação para este
ambiente. Durante 5 minutos foram mensurados os seguintes parâmetros: número de
cruzamentos (crossings), respostas de orientação, ou atividade de levantar (rearings), tempo
parado nos quadrados (ansiedade), tempo de auto-limpeza (groomings).
No 24ª dia pós-nascimento foi realizado o primeiro teste de comportamento para
avaliar atividade locomotora. Não foram constatados nenhuma diferença estatisticamente
significativa entre os grupos AVP e grupo SHAM dos animais machos como podemos observar
na Figura 20.

82
Figura 20 – Análise da atividade motora no Teste do Campo Aberto em ratos (machos)
realizado no 24ª dia pós-natal, induzidos ao autismo tratados por 25 dias com mix de óleos e
uma única dose de ácido valpróico.
Fonte: Autora. Legenda: A: Número de cruzamentos; B: Tempo parado; C: Grooming; D: Rearing. Os valores representam a média ± SEM. (n= 13 – 23 animais por grupo experimental). Para análises estatísticas foram utilizados o teste One-way ANOVA seguido por Tukey. ‘a’ mostra diferença significativa em relação ao grupo SHAM. ‘b’ mostra diferença significativa em relação ao grupo Ácido valpróico (AVP). ‘c’ mostra diferença significativa em relação ao Mix 1. ‘d’ mostra diferença significativa em relação ao Mix3 + AVP. Valores significativos: aaaa=p<0,005, b=p<0,05, c=p<0,05 e d=p<0,05.
Para os grupos dos animais fêmeas, ao analisar o número de cruzamentos,
comparando os animais tratados com o SHAM (31,5±2,05; p<0,05), o grupo ácido valpróico se
mostrou diferente significativamente (46,44±2,69; p<0,05), com uma elevação exploratória
maior. Para variável tempo parado, os animais tratados com AVP (72,22±3,59; p<0,05) e Mix
3 (57,08±12,11; p<0,01) reduziram significativamente em relação ao grupo SHAM, podendo
caracterizar um comportamento de ansiedade (Figura 21).
Quando analisado os valores de autolimpeza (Grooming), nenhum grupo
experimental demonstrou diferença estatística quando comparado com o SHAM ou com o
grupo tratado somente com ácido valpróico. O último parâmetro analisado neste teste se refere
a atividade exploratória vertical (rearing). Somente apresentaram diferença significativa em

83
relação ao grupo SHAM, os animais tratados com AVP (18,2±1,88; p<0,05). Não foi constatado
nenhuma diferença significativa em relação ao grupo de Ácido valpróico.
Figura 21– Análise da atividade motora no Teste do Campo Aberto em ratos (fêmeas) realizado
no 24ª dia pós-natal, induzidos ao autismo tratados por 25 dias com mix de óleos e uma única
dose de Ácido Valpróico.
Fonte: Autora. Legenda: A: Número de cruzamentos; B: Tempo parado; C: Grooming; D: Rearing. Os valores representam a média ± SEM. (n= 10 – 18 animais por grupo experimental). Para análises estatísticas foram utilizados o teste One-way ANOVA seguido por Tukey. ‘a’ mostra diferença significativa em relação ao grupo SHAM. ‘b’ mostra diferença significativa em relação ao grupo Ácido valpróico (AVP). ‘c’ mostra diferença significativa em relação ao grupo Mix1. ‘d’ mostra diferença significativa em relação ao grupo Mix2+AVP; ‘e’ mostra diferença significativa em relação ao grupo Mix3+AVP; Valores significativos: a=p<0,05; b=p<0,05; c=p<0,05; ee=p<0,01.
4.1.2 Avaliação da Atividade Locomotora - Teste de Imersão em Cauda
Para o teste de imersão em cauda, realizado no 30ª dia pós-nascimento, foi avaliado
o tempo em milissegundos no qual o animal realiza a total retirada da cauda quando imerso em
água.
Ao comparar os grupos controles do experimento nos animais machos, foi possível
constatar uma diferença significativa entre eles, onde o grupo AVP (565,7±13,87) aumentou o
tempo em relação ao grupo SHAM (369,6±8,84; p<0,0001). Ainda em relação ao grupo SHAM,

84
este se mostrou diferente do grupo Mix-1(462,1±18,55; p<0,01), onde o tratamento elevou o
tempo de retirada conforme a Figura 22.
Quando comparado com o Grupo AVP (565,7±13,87), todos os grupos
experimentais associados ao ácido valpróico se mostraram estatisticamente diferentes,
conseguindo reduzir o tempo de retirada da cauda [ (Mix1+AVP:377,5±19,66; p<0,0001);
(AVP + Mix-2: 349,1±10,27; p<0,0001) e (AVP + Mix-3: 429,3±12,77; p<0,0001) ].
O Mix-1 (462,1±18,55) se mostrou deferente significativamente em relação aos
grupos tratados com Mix-2 (383,8±16,52; p<0,01) e Mix-3 (390,5±16,52; p<0,01), bem como
com AVP + Mix-1 (377,5±19,66; p<0,01). Já o AVP + Mix-2 (349,1±10,27) se mostrou
diferente significativamente do AVP + Mix-3(429,3±12,77; p<0,001), aumentando o tempo de
retirada da cauda.
Figura 22 – Análise da imersão de cauda em ratos machos realizado no 30ª dia pós-natal,
induzidos ao autismo tratados por 25 dias com Mix de óleos e uma única dose ácido valpróico.
Fonte: Autora. Legenda: AVP: Ácido valpróico. Os valores representam a média ± SEM. (n= 13 – 23 animais por grupo experimental). Para análises estatísticas foram utilizados o teste One-way ANOVA seguido por Tukey. ‘a’ mostra diferença significativa em relação ao grupo SHAM. ‘b’ mostra diferença significativa em relação ao grupo Ácido valpróico (AVP). ‘c’ mostra diferença significativa em relação ao Mix-1. ‘d’ mostra diferença significativa em relação ao Mix2+AVP. Valores significativos: aaaa=p<0,0001; bbbb=p<0,0001; cc=p<0,01, ddd=p<0,001.
Na Figura 23, ao ser comparado os grupos experimentais fêmeas, foi possível
constatar a diferença entre os grupos controle SHAM (379,7±12,59) e AVP (671,9±67,08;
p<0,005), onde os animais tratados com ácido valpróico necessitaram de maior tempo para
desempenhar a função. Os demais grupos experimentais não se mostraram significativamente
diferente do grupo SHAM.
Quando comparado com o Grupo AVP, todos os grupos experimentais se
mostraram estatisticamente diferentes, conseguindo reduzir o tempo de retirada da cauda.

85
(Mix1+AVP: 440,9±10,64; p<0,01); (Mix2+AVP: 397,8±11,19; p<0,0001) e (341±21,82;
p<0,0001)].
Figura 23 – Análise da atividade motora no Teste de Imersão em Cauda em ratos fêmeas
realizado no 30ª dia pós-natal, induzidos ao autismo tratados por 25 dias com mix de óleos e
uma única dose de Ácido valpróico.
Fonte: Autora. Legenda: AVP: Ácido Valpróico. Os valores representam a média ± SEM. (n= 10 – 15 animais por grupo experimental). Para análises estatísticas foram utilizados o teste One-way ANOVA seguido por Tukey. ‘a’ mostra diferença significativa em relação ao grupo SHAM. ‘b’ mostra diferença significativa em relação ao grupo AVP. Valores significativos: aaa=p<0,005; bb=p<0,05 e ‘bbbb= p<0,0001.
4.1.3 Teste de Performance da Natação
Para o teste de natação realizado no 32ª dia pós-nascimento, foi avaliado os scores
de acordo com o ângulo de inclinação da cabeça dos animais na superfície da água de 1 a 4
(Figura 24).
Ao comparar os grupos controles machos, foi possível constatar a diferença
significativa entre o grupo SHAM (3,9±0,09) e o grupo AVP (3±0,17; p<0,005), onde houve
uma redução da média dos scores. Os demais grupos experimentais, foi possível observar
alterações significativas somente em relação aos grupos Mix-1 (3,92±0,07) e o Grupo AVP +
Mix-1 (3,167±0,16; p<0,01), onde foi possível observar a diminuição do desempenho motor.

86
Figura 24 – Análise da imersão de cauda em ratos machos realizado no 32ª dia pós-natal,
induzidos ao autismo tratados por 25 dias com Mix de óleos e uma única dose ácido valpróico
Fonte: Autora. Legenda: AVP: Ácido valpróico. Os valores representam a média ± SEM. (n= 12 – 20 animais por grupo experimental). Para análises estatísticas foram utilizados o teste One-way ANOVA seguido por Tukey. ‘a’ mostra diferença significativa em relação ao grupo SHAM. ‘c’ mostra diferença significativa em relação ao Mix 1. Valores significativos: aaa=p<0,005 e cc=p<0,01.
Resultados parecidos foram encontrados nos grupos dos animais fêmeas como pode
ser observado na Figura 25. Para os grupos controles, foi possível constatar a diferença
significativa entre o grupo SHAM (3,84±0,10) e o grupo AVP (3,33±0,11; p<0,05), onde houve
uma redução da média dos scores. Entre os grupos tratados, foi possível observar alterações
significativas somente em relação aos grupos Mix 3 (4,0±0) e o Grupo Mix3+AVP (3,11±0,11;
p<0,005), apresentando uma redução do score no grupo associado ao Ácido Valpróico.

87
Figura 25 – Análise da atividade motora no Teste de Imersão em Cauda em ratos fêmeas
realizado no 32ª dia pós-natal, induzidos ao autismo tratados por 25 dias com mix de óleos e
uma única dose de ácido valpróico.
Fonte: Autora. Legenda: AVP: Ácido valpróico. Os valores representam a média ± SEM. (n= 12 – 20 animais por grupo experimental). Para análises estatísticas foram utilizados o teste One-way ANOVA seguido por Tukey. ‘a’ mostra diferença significativa em relação ao grupo SHAM. ‘e’ mostra diferença significativa em relação ao Mix 3. Valores significativos: a=p<0,05 e eee=p<0,005.
4.1.4 Avaliação da Atividade Locomotora -Teste de Rota-rod
Para o teste de Rota-rod, foram avaliadas quatro variáveis relacionadas a atividade
motora do animal, a saber: primeira queda e tempo final (coordenação motora e equilíbrio) e
RPM para a primeira queda e RPM final (equilíbrio e força muscular). Os parâmetros avaliados
no teste de Rotarod incluem: tempo de latência (ou de resistência para queda), distância total
percorrida (dependente da freguência de rotação e do tempo de latência).
Na Figura 26, ao ser analisado os parâmetros no teste rota rod para os grupos de
animais machos, não foi possível observar resultados significativos entre o grupo AVP em
comparação ao grupo salina (sham).

88
Figura 26 – Análise da locomoção através do teste de RotaRod em ratos machos realizado no
50ª dia pós-natal, induzidos ao autismo tratados por 25 dias com Mix de óleos e única dose de
ácido valpróico.
Fonte: Autora. Legenda: A: Tempo final; B: RPM final; C: Primeira queda; D: RPM para a primeira queda. Os valores representam a média ± SEM. (n= 13 – 23 animais por grupo experimental). Para análises estatísticas foram utilizados o teste One-way ANOVA seguido por Tukey. ‘b’ mostra diferença significativa em relação ao grupo Ácido valpróico (AVP). ‘c’ mostra diferença significativa em relação ao Mix 1. ‘d’ mostra diferença significativa em relação ao Mix2. Valores significativos: b=p<0,05, cc=p<0,01, d=p<0,05 e dddd=p<0,0001.
Nos grupos dos animais fêmeas, a avaliação do tempo final representado na Figura
27, os grupos AVP (125±12,08; p<0,005) e Mix-3 (114±9,2; p<0,0001) reduziram
estatisticamente o tempo em relação ao SHAM (201,8±7,78). Quando comparado em relação
ao grupo tratado somente com Ácido Valpróico (125,0±12,08), Mix-1+AVP (208,8±12,9;
p<0,0001) e Mix-2+AVP (264,8±8,71; p<0,0001) apresentaram diferença significativa,
levando ao aumento do tempo.
Ao avaliar o tempo necessário para a ocorrência da primeira queda, os únicos
grupos que se mostraram diferentes do grupo SHAM foram o AVP (49,5±6,72; p<0,005) e Mix-
3 (48,77±5,93; p<0,01), diminuindo o tempo.
O último parâmetro avaliado foi o RPM necessário para que ocorresse a primeira
queda onde, em relação ao grupo SHAM (18,54±1,6), AVP (10,06±0,79; p<0,005) e Mix-3
(9,92±0,68; p<0,01) se mostraram diferentes.

89
Figura 27 – Análise da atividade motora no Teste Rota Road em ratos fêmeas realizdo no 50ª
dia pós-natal, induzidos ao autismo tratados por 25 dias com mix de óleos e uma única dose de
ácido valpróico.
Fonte: Autora. Legenda: A: Tempo final; B: RPM final; C: Primeira queda; D: RPM para a primeira queda. AVP: Ácido valpróico. Os valores representam a média ± SEM. (n= 13 – 18 animais por grupo experimental). Para análises estatísticas foram utilizados o teste One-way ANOVA seguido por Tukey. ‘a’ mostra diferença significativa em relação ao grupo SHAM. ‘b’ mostra diferença significativa em relação ao grupo Ácido valpróico (AVP). ‘c’ mostra diferença significativa em relação ao Mix 1. ‘d’ mostra diferença significativa em relação ao Mix1+AVP. ‘e’ mostra diferença significativa em relação ao Mix2+AVP. ‘f’ mostra diferença significativa em relação ao Mix3+AVP Valores significativos: aa=p<0,01; aaa=p<0,005; aaaa=p<0,0001 b=p<0,05; bbb=p<0,005; bbbb=p<0,0001; cc=p<0,01; ccc=p<0,005; cccc=p<0,0001 e d=p<0,05; dd=p<0,01.; ddd=p<0,005; . ee=p<0,01; eee=p<0,005 e ffff.=p<0,0001.
4.2 Análises Neuroquímicas em MACHOS e FÊMEAS
4.2.1 Determinação dos Níveis de Peroxidação Lipídica (TBARS)
De acordo com os resultados obtidos (Figura 28), quando analisados os níveis de
Peroxidação Lipídicas no Hipocampo, os animais tratados com ácido valpróico (1488,0±433,1;
p<0,05) elevaram o grau de peroxidação lipídica em relação ao grupo SHAM (777,5±112,3),
sendo restaurado com o mix-3 no grupo mix-3 + AVP. Ao comparar os grupos experimentais
com o grupo tratado somente com Ácido Valpróico (AVP), AVP + Mix-2 (863,0±37,52;
p<0,05) e AVP + Mix-3 (737,6±40,88; p<0,01) demostraram diferença significativa,
conseguindo reduzir os níveis de TBARS das amostras.
No Córtex Pré-Frontal, foi possível constatar diferença significativa entre os grupos
controles, onde AVP (1358,0±311,0) aumentou significativamente os níveis de MDA em

90
relação ao SHAM (514,8±100,3 p<0,05). Também corroborando com o resultado do
hipocampo, restaurado com o mix-3 do grupo Mix-3+AVP (575,0±49,58).
Nas amígdalas, nenhuma dose testada dos mixs foi diferente significativamente do
grupo SHAM ou do ácido valpróico para os níveis de peroxidação lipídica. Por fim, no
Cerebelo, constatou-se diferença significativa entre os grupos controles AVP e SHAM, onde
apresentou uma redução dos níveis de TBARS em animais tratados com AVP (1097,0±176;
p<0,05) em relação ao grupo SHAM (1858,0±197,7), não sendo esperado neste estudo.
Figura 28 – Níveis de Peroxidação Lipídica (TBARS) nas áreas cerebrais de ratos machos
realizados no 55ª dia pós-mortem, tratados por 25 dias com Mix de óleos e uma única dose de
Ácido Valpróico.
Fonte: Autora. Legenda: A: Hipocampo; B: Córtex pré-frontal; C: Amigdalas; D: Cerebelo. Os valores representam a média ± SEM dos níveis de MDA (malondialdeído) medidos em µg/g de tecido. (n= 6 – 8 animais por grupo experimental). Para análises estatísticas foram utilizados o teste One-way ANOVA seguido por Tukey. ‘a’ mostra diferença significativa em relação ao grupo SHAM. ‘b’ mostra diferença significativa em relação ao grupo Ácido valpróico (AVP). ‘c’ mostra diferença significativa em relação ao Mix 1. ‘d’ mostra diferença significativa em relação ao Mix1+AVP. Valores significativos: a=p<0,05; b=p<0,05; bb=p<0,01; c=p<0,05; cc=p<0,01, d=p<0,05; dd=p<0,01 e dddd=p<0,0001.
De acordo com os resultados obtidos na Figura 29, para as áreas analisadas
hipocampo, córtex pré-frontal, amígdalas e cerebelo o estresse oxidativo não esteve presente
no grupo AVP quando comparado estatisticamente com o grupo controle salina para os animais
fêmeas.

91
Figura 29 – Níveis de Peroxidação Lipídica (TBARS) nas áreas cerebrais de ratos fêmeas
realizados no 55ª dia post-mortem, tratados por 25 dias com Mix de óleos e uma única dose de
Ácido valpróico.
Fonte: Autora. Legenda: A: Hipocampo; B: Córtex pré-frontal; C: Amigdalas; D: Cerebelo. Os valores representam a média ± SEM dos níveis de MDA (malondialdeído) medidos em µg/g de tecido. (n= 6 – 8 animais por grupo experimental). Para análises estatísticas foram utilizados o teste One-way ANOVA seguido por Tukey. ‘a’ mostra diferença significativa em relação ao grupo SHAM; ‘b’ mostra diferença significativa em relação ao grupo Ácido valpróico (AVP); ‘c’ mostra diferença significativa em relação ao Mix 1; ‘d’ mostra diferença significativa em relação ao Mix 2; ‘e’ mostra diferença significativa em relação ao Mix 3. Valores significativos: a=p<0,05; b=p<0,05; bb=p<0,01; bbbb=p<0,0001; c=p<0,05; cccc=p<0,0001, d=p<0,05; dddd=p<0,0001 e eee=p<0,005.
4.2.2 Determinação da Concentração de Nitrito (NO2) / Nitrato (NO3)
Avaliação da determinação de nitrito no estudo com o uso de ácido valpróico
espera-se expressar uma elevação dos níveis de Espécies Reativas de Nitrogênio (ROS)
caracterizando um tipo de processo inflamatório. Na Figura 30, para os animais machos, as
áreas hipocampo, córtex pré-frontal e amígdalas não foram observadas diferenças estatísticas
entre os grupos AVP e SHAM, no entanto, ao analisar o Cerebelo, os grupos AVP (5,32±1,03)
demonstrou diferença significativa em relação ao grupo SHAM (1,33±0,24; p<0,0001),
aumentando os níveis de nitrito/nitrato nas amostras cerebrais, sendo expressivamente
restaurada após o uso dos mix-1, mix-2 e mix-3 dos animais AVP, conseguindo diminuir os
níveis de NO2 e NO3 no tecido cerebral em estudo (Figura 30).

92
Figura 30 – Concentrações de Nitrito/Nitrato nas áreas cerebrais de ratos machos realizados no
55ª dia pós-mortem, tratados por 25 dias com Mix de óleos e uma única dose de Ácido
Valpróico.
Fonte: Autora. Legenda: A: Hipocampo; B: Córtex pré-frontal; C: Amigdalas; D: Cerebelo. Os valores representam a média ± SEM dos níveis de Nitrito/nitrato (NO2/NO3), medidos em mMol/g de tecido. (n= 5 – 8 animais por grupo experimental). Para análises estatísticas foram utilizados o teste One-way ANOVA seguido por Tukey. ‘a’ mostra diferença significativa em relação ao grupo SHAM. ‘b’ mostra diferença significativa em relação ao grupo Ácido valpróico (AVP). ‘c’ mostra diferença significativa em relação ao Mix 1. ‘d’ mostra diferença significativa em relação ao Mix1+AVP. Valores significativos: a=p<0,05; aaa=p<0,001; aaaa=p<0,0001 b=p<0,05; bb=p<0,01; bbbb=p<0,0001; c=p<0,05; cc=p<0,01; cccc=p<0,0001 e dd=p<0,01.
Representando os resultados da determinação de Nitrito no grupo das fêmeas, é
possível constatar na Figura 31 que em nenhuma das áreas cerebrais analisadas houve um
aumento significativo de níveis de NO2 e NO3 no tecido cerebral.

93
Figura 31 – Concentração de Nitrito (NO2) / Nitrato (NO3) nas áreas cerebrais de ratos machos
realizados no 55ª dia pós-mortem, tratados por 25 dias com Mix de óleos e uma única dose de
Ácido Valpróico.
Fonte: Autora. Legenda: A: Hipocampo; B: Córtex pré-frontal; C: Amigdalas; D: Cerebelo. Os valores representam a média ± SEM dos níveis de Nitrito/nitrato (NO2/NO3), medidos em mMol/g de tecido. (n= 5 – 8 animais por grupo experimental). Para análises estatísticas foram utilizados o teste One-way ANOVA seguido por Tukey ‘b’ mostra diferença significativa em relação ao grupo Ácido valpróico (AVP). ‘c’ mostra diferença significativa em relação ao Mix 1. ‘d’ mostra diferença significativa em relação ao Mix1+AVP. Valores significativos: b=p<0,05; bbbb=p<0,0001 cccc=p<0,0001; d=p<0,05; dd=p<0,01 e dddd=p<0,0001.
4.2.3 Determinação das Concentrações de Glutationa Reduzida (GSH)
A glutationa (GSH) faz parte de importante sistema enzimático de defesa contra
radicais livres, sendo um marcador da saúde celular e sua queda é indicativa de lesão oxidante.
Foi realizado nas áreas cerebrais: hipocampo, córtex pré-frontal, amígdalas e cerebelo a
dosagem dos níveis de glutationa reduzida onde esperava-se uma diminuição nos grupos com
ácido valpróico em comparação ao SHAM. Analisando as Figuras 32 e 33, para machos e
fêmeas respectivamente, não houve diferença estatística em nenhum dos grupos experimentais.

94
Figura 32 – Concentrações de GSH (µ/g de tecido) nas áreas cerebrais de ratos machos
realizados no 55ª pós-mortem, tratados por 25 dias com Mix de óleos e uma única dose de Ácido
Valpróico.
Fonte: Autora. Legenda: A: Hipocampo; B: Córtex pré-frontal; C: Amigdalas; D: Cerebelo. Os valores representam a média ± SEM dos níveis de Glutationa reduzida (GSH), medidos em uMol/g de tecido. (n= 6 – 11 animais por grupo experimental). Para análises estatísticas foram utilizados o teste One-way ANOVA seguido por Tukey. ‘c’ mostra diferença significativa em relação ao Mix 1. ‘d’ mostra diferença significativa em relação ao Mix2+AVP. Valores significativos:c=p<0,05; cc=p<0,01 e dd=p<0,01.
Abaixo podem ser analisados os resultados para GSH em animais fêmeas e sua
respectivas áreas cerebrais avaliadas.

95
Figura 33 – Níveis de GSH (µ/g de tecido) nas áreas cerebrais de ratos fêmeas realizados no 55ª
dia pós-mortem, tratados por 25 dias com Mix de óleos e uma única dose de Ácido Valpróico.
Fonte: Autor. Legenda: A: Hipocampo; B: Córtex pré-frontal; C: Amigdalas; D: Cerebelo. Os valores representam a média ± SEM dos níveis de Glutationa reduzida (GSH), medidos em uMol/g de tecido. (n= 6 – 9 animais por grupo experimental). Para análises estatísticas foram utilizados o teste One-way ANOVA seguido por Tukey. ‘a’ mostra diferença significativa em relação ao grupo SHAM. ‘c’ mostra diferença significativa em relação ao grupo MIX 1.‘d’ mostra diferença significativa em relação ao grupo MIX 2. ‘e’ mostra diferença significativa em relação ao grupo MIX 1+AVP. ‘f’ mostra diferença significativa em relação ao grupo MIX 3+AVP. ‘g’ mostra diferença significativa em relação ao grupo MIX 2+AVP. ‘h’ mostra diferença significativa em relação ao grupo MIX 3. Valores significativos: a=p<0,05, cc=p<0,01; dd= p<0,01; e=p<0,05; f=<0,05; gg=<0,01; h=p<0,05
4.3 Expressão Gênica por RT-qPCR
Os resultados do Figura 34 mostram as curvas-padrão realizadas para os genes alvos
(Dnmt3a e Chd2) e o gene referência (Actb). Todas as curvas apresentadas apresentam
eficiência similar (Figura 34A a 34C) (Actb, 1,11; Dnmt3a, 1,08 e Chd2 1,19 respectivamente).
A similaridade das eficiências permite a análise de quantificação da expressão relativa realizada
pelo médoto 2-ddCt. Observa-se que após curva de melting, todas as amplificações foram
específicas de acordo com as curvas de melting apresentadas a baico (Figura 34D a 34F).

96
Figura 34 – Padronização dos genes alvo e referência. A - eficiência dos primers, B - curva de
melting.
Fonte: Autora. Legenda: Curvas padrão dos genes Bactin (A/D), Dnmt3a (B/E) e Chd2 (C/F) e suas respectivas curvas de melting.
Figura 35 – Avaliação da expressão relativa diferencial dos genes Dnmt3a e Chd2.
Fonte: Autora. Legenda: A: Hipocampo. O teste de normalidade KS foi realizado.
1As medianas dos grupos não normais foram submetidas ao teste não paramétrico Mann-Whitney para a
comparação pareada entre os grupos AVP, Mix-3, AVP + Mix-3 comparado ao controle SHAM.
2As médias dos grupos foram submetidas ao teste t não pareado para a comparação paramétrica entre a média dos
grupos experimentais, comparados ao controle SHAM.
A B C
D E F

97
5 DISCUSSÃO
Foi avaliado o estresse oxidativo em 4 áreas cerebrais mais investigadas e
acometidas pelo TEA: hipocampo, córtex pré-frontal, amígdalas e cerebelo, segunda a
literatura.
Estudos em modelos animais fornecem grandes possibilidades de investigações que
não são possíveis em humanos, incluindo análises imunológicas e do desenvolvimento,
plasticidade e sinalização neural. O autismo é uma condição particularmente difícil de ser
mimetizada em animais. A empatia e a capacidade de intuir os sentimentos e intenções de
terceiros parecem ser impossíveis de ser medidas em ratos ou camundongos (MILLER et al.,
2006). Por outro lado, os roedores são, em geral, espécies que têm altos índices de interação e
comunicação social (CRAWLEY et al., 2003). Essa característica faz com que mudanças no
comportamento social dos roedores sejam relacionadas a fatores ambientais ou genéticos
(CRAWLEY et al., 2007; INGRAM et al.,2000).
A vantagem de utilizar ratos nesses estudos é baseado no relatório de que o AVP
atingiria a mesma concentração no cérebro em seres humanos do que em camundongos,
extrapolando assim os dados mais relevantes para o homem (JOHANNESSEN;
JOHANNESSEN, 2003). Exposição no 14ª dia pós-nascimento é selecionada como janela
crítica, na migração e diferenciação de células granulares cerebelares e hipocampais. Devido às
anomalias cerebrais produzidas pelo VPA, há perda de atividade motora, cognição,
comportamentos intrusivos, sensibilidade alterada a estímulos dolorosos e déficit pertinente
interação social (MARKRAM et al., 2008).
O AVP induz regressões comportamentais em camundongos na exposição pós-natal
idêntica àquele visto em indivíduos com autismo (MING et al., 2008).
O teste de campo aberto é comumente usado para avaliar a atividade locomotora
geral, explorar o comportamento, e comportamento relacionado à ansiedade. A distância total
percorrida e o tempo gasto no centro do campo são medidos para avaliar a atividade dos
animais. Além disso, a duração total da auto-limpeza e a atividade de levantar as patas dianteiras
são medidas para o comportamento (SEGAL-GAVISH et al., 2015; SEIBENHENER et al.,
2015; BASSO et al., 1995; KUNIISHI et al.,2017).
A exploração restrita é uma das principais características dos casos clínicos e pré-
clínicos de autismo (OLEXOVÁ et al.,2013). Esta leitura muitas vezes pode ser realizada pelo
teste do campo aberto através do número de cruzamentos e rearing. A diminuição da atividade
exploratória e locomotora neste estudo não foi percebida para os animais machos como

98
mencionado por Wagner et al., (2006) onde demonstraram que comportamentos críticos
mediados pelo cerebelo aparecem pela primeira vez no 14ª dia pós-natal. E que, uma única
exposição no 14ª dia pós-natal de 400 mg/kg de ácido valpróico foi eficaz na produção de
deficits cognitivos e motores induzidos por VPA que imitavam a regressão autística resultando
em deficits sociais nos camundongos em desenvolvimento.
No entanto, as fêmeas apresentaram resultados antagônicos a estes bem como no
esperado de efeito teratogênico com ácido valpróico. Um aumento significante da atividade
locomotora e exploratória podem ser observadas estatisticamente no grupo com autismo AVP
em relação ao Sham (Figura 21). O parâmetro “número de cruzamentos” é considerado uma
medida simultânea de comportamento ansioso e de atividade locomotora. Um alto valor deste
parâmetro sugere exploração e locomoção aumentadas e, consequentemente, menor nível de
ansiedade (ZIMCIKOVA et al., 2017). É possível verificar estes resultados também entre os
grupos, onde Mix-3 + AVP e Mix-1 + AVP, apresentaram valores significantemente maiores
em relação ao neurotípicos do mesmo grupo. Quanto à análise dos rearings, o grupo AVP
fêmeas apresentou uma quantidade significativamente maior deste comportamento em relação
ao grupo Salina. O rearing é interpretado tanto como um índice de medição de comportamento
exploratório como de comportamento que expressa ansiedade, apesar de haver controvérsias na
literatura acerca do tema (SEIBENHENER et al., 2015). O resultado obtido no presente estudo
foi um aumento significante na quantidade de rearings no grupo AVP em relação ao Salina,
sem diferenças significantes entre os demais grupos, o que pode ser interpretado como um
aumento da atividade exploratória que se deve ao AVP, relacionando-se negativamente com
estados ansiosos e de medo (SESTAKOVAL et al., 2013), observado na atividade de “tempo
parado” onde as fêmeas do grupo Salina, estiveram significativamente menos interesse em se
locomover quando comparados com o grupo AVP e o grupo Mix-3. Mais estudos devem ser
encorajados para explicar o aumento desta exploração com uso do ácido graxo DHA no animal
AVP, “seus efeitos neuroprotetores” como podemos encontrar na Figura 21 para o grupo Mix-
3 + AVP em relação ao grupo Mix-2 + AVP, bem como para os grupos neurotípicos tratados
dos animais machos, onde Mix-1 teve um aumento significante quando comparados aos grupos
SHAM e Mix-3.
Um estudo clínico recente relatou que a relação entre os PUFA ômega-3 e o AA é
antagônica à manutenção da homeostase em altas concentrações de EPA e DHA. A
incorporação de PUFAs ômega-3 reduz a concentração de AA (JANSSEN et al., 2014;
MORBTATEN et al., 2013), regulando a síntese de eicosanóides, mediadores da sinalização de
AA na membrana celular (MORBTATEN et al., 2013). Os PUFAs ômega-3 na dieta podem

99
atenuar os níveis de AA do tecido e a formação de eicosanóides (WHELAN et al., 1996). O
antagonismo da sinalização de AA pode ter efeitos importantes na sinalização celular no
sistema nervoso central (OBAJIMA et al., 2005; MORI 2004) modificando os efeitos
biomédicos e comportamentais (OBAJIMA et al., 2005). Essa interação competitiva entre os
PUFA ômega-3 e o AA reflete um aumento nas proporções ômega-3 / AA no plasma
(SCHMITZ et al., 2008; LUXWOLDA et al., 2011; JANSSEN et al., 2014). Podendo ser uma
luz nas interpretações destes dados acima neste estudo.
Modelos de animais de ansiedade que utilizam diversos comportamentos de defesa,
investigam comportamentos que não precisam ser aprendidos, são preferências de grande parte
da comunidade cientifica, na medida em que não envolvem situações de treino, apresentam
baixo custo e têm sessões de curta duração. O grooming de roedores, apesar de negligenciado
pela maioria da comunidade científica, se enquadra nesta categoria de comportamentos e vem
sendo investigado por alguns pesquisadores (VELOSO et al., 2012; ESTANISLAU et al., 2012;
KALUEFF; TOUHIMMA 2005a; KALUEFF; TOUHIMMA, 2005b), como um possível
marcador de ansiedade. Nosso estudo englobou ambos os sexos no 24ª dia pós-natal, período
da ontogenia destes animais, já tendo sido investigado por (YADAV et al., 2016) aplicando o
mesmo protocolo, porém com óleos de ácidos graxos diferentes (ALA e GLA). Embora o AVP
possa induzir anormalidades cerebrais em ratos semelhantes aos de indivíduos com autismo
(BAUMAN et al., 1985; BAUMAN et al., 2005; CZYŻ et al., 2001), como a ansiedade, neste
estudo não foi possível observar estas mudanças em ambos os sexos.
O teste de imersão de cauda é um teste largamente utilizado em modelos animais
para aferir a nocicepção, i.e., a recepção de estímulos aversivos, transmissão, modulação
e percepção de tais estímulos. Em relação ao TEA, a literatura é conflitante acerca das
alterações na sensibilidade dolorosa nos indivíduos afetados, entretanto é consenso que crianças
com TEA reagem de forma diferente à dor (BARONIO et al, 2015). Em geral, nos modelos
animais de TEA induzidos por AVP, há uma redução da nocicepção supraespinhal e espinhal,
segundo revisão de Schneider, Przewłocki, similar aos resultados encontrados no presente
estudo na avaliação da nocicepção espinhal, visto que tanto os animais machos quanto fêmeas,
quando induzidos ao modelo de TEA pelo AVP, demonstraram um aumento significativo e
quantitativamente expressivo no tempo de latência no teste de imersão de cauda (SCHNEIDER;
PRZEWŁOCKI, 2005). Tal achado sugere o sucesso na indução do TEA pelo modelo de
indução no 14ª dia pós-natal por AVP no quesito de sensibilidade à dor, que encontra-se
diminuída ou abolida em casos humanos, sendo este um paralelo fisiológico entre o atraso para

100
a retirada da cauda pelo animal mediante estímulo doloroso e a hipossensibilidade nociceptiva
observada em humanos com TEA (MILITERNI et al, 2000).
Quanto às gorduras ricas em ômega testadas, Mix-1, Mix-2 e Mix-3, todas foram
capazes de normalizar os tempos de latências nos animais induzidos ao modelo de TEA,
levando o tempo de reação em geral para valores semelhantes ao grupo SHAM em ambos os
sexos. Tal resultado corrobora com achados de outros estudos, como no de YADAV et al, 2016,
que também obteve uma normalização do tempo de reação de retirada de cauda à estímulo
doloroso nos grupos tratados com os ácidos graxos poli-insaturados ácido alfa-
linolênico (ALA) e ácido gama-linolênico (GLA) em um modelo de TEA induzido por AVP.
Resultados semelhantes também foram obtidos com o uso de outras substâncias como
tratamento, como no caso de Huang et al em que a Betaína também foi capaz de fazer com que
o tempo de latência dos animais tratados no modelo de TEA induzido por AV retornasse a
valores semelhantes ao grupo controle. No caso da Betaína, o mecanismo sugerido para tal
melhora foi o aumento do metabolismo da homocisteína e consequente redução em sua
concentração (HUANG et al., 2019). Já no caso dos ácidos graxos poli-insaturados, o
mecanismo ainda não se encontra bem estabelecido no caso da melhora de reflexos medulares
como no caso do teste de imersão em cauda, mas postula-se que haja relação com a prevenção
de uma mielinização inadequada de regiões centrais e periféricas do sistema nervoso
(WYSOCZAŃSKI et al., 2016).
O teste de performance de natação foi criado por Schapiro et al, em 1970, com o
objetivo de desenvolver um teste para avaliar o desenvolvimento motor geral do rato e seus
possíveis atrasos em diferentes modelos através do padrão de nado, desde o menos
desenvolvido (0) ao mais completo (4), presente no animal adulto (SCHAPIRO et al, 1970).
Este teste foi validado para o uso em modelo animal de TEA induzido pelo AVP, no qual os
ratos induzidos ao modelo tendem a apresentar um atraso em tal desenvolvimento
(SCHNEIDER et al., 2005). Por mais que o teste tenha sido idealizado em 1970 para ser
realizado em torno dos dias pós-nascimento 6 a 16, há estudos mais recentes que utilizam o
teste em períodos mais tardios, como Yadav et al que utilizou no dia pós-nascimento 32, e que
têm obtido resultados úteis para a avaliação de repercussões mais tardias no desenvolvimento
motor em modelo de TEA induzido por AVP e para testar o efeito de substâncias sobre tais
repercussões.
No presente estudo, foi possível perceber que, em ambos os sexos, o grupo AVP
apresentou em média um padrão de nado significantemente mais atrasado do que o grupo
SHAM no dia 32ª dia pós-nascimento, o que sugere um prejuízo no desenvolvimento motor

101
causado pelo AVP com repercussões duradouras, classificando a maioria dos animais de tal
grupo no padrão 3 de nado (água a nível da orelha na maior parte do teste). No grupo dos
machos, os grupos Mix-2+AVP e Mix-3+AVP apresentaram uma média de escore de natação
maior do que o grupo AVP e não apresentaram diferença significante em relação aos respectivos
grupos Mix-2 e Mix-3. No grupo de animais fêmeas, os grupos Mix-1+AVP e Mix-2+AVP
obtiveram escores em média maiores que o do grupo AVP, sem diferença significante em
relação aos grupos Mix-1 e Mix-2, respectivamente. Tais resultados em conjunto sugerem que
a melhora do desempenho motor está relacionada ao tratamento com os ácidos graxos poli-
insaturados administrados, visto que estão implicados na regulação do bom funcionamento da
membrana celular neuronal e na mielinização adequada de axônios, essencial à formação e
integração de circuitos motores complexos no animal, recrutados durante o nado (VAN ELST
et al., 2014). Importante observar que o Mix-2 foi o único capaz de mitigar significativamente
a repercussão negativa causada pela administração do AVP ao desenvolvimento motor do rato
em ambos os sexos. Tal efeito positivo pode ocorrer devido ao fato de que as fontes de ω-3 do
Mix2 são mais variadas (ALA, EPA e DHA), o que pode levar a uma melhor distribuição no
sistema nervoso central e periférico ao longo do período de tratamento, haja vista que o teste
avalia desde funções motoras periféricas até sua integração, regulação e modulação central em
uma atividade motora integrada única, o nado (SCHAPIRO et al., 1970).
O teste de Rotarod é um teste comportamental que atualmente tem ganho grande
importância na avaliação de modelos animais de TEA por ser um dos mais clássicos testes para
avaliação da coordenação motora em ratos, função primordial do cerebelo, estrutura cada vez
mais implicada no TEA segundo a literatura, visto que há evidência crescente de que alterações
cerebelares como depleção de células de Purkinje, elevação do estresse oxidativo e disfunção
gabaérgica geram repercussões motoras, mnésicas e linguísticas observáveis nos indivíduos
com TEA (MORAKOTSRIWAN et al., 2016).
No presente estudo, o grupo AVP de fêmeas sofreu uma redução significante em
relação ao grupo SHAM, tanto no tempo de latência até a primeira queda quanto no tempo final,
o que condiz com os resultados da maioria dos estudos que avaliam a coordenação motora de
modelos animais de TEA no teste de Rotarod, pois é esperada uma piora da coordenação motora
nos testes que avaliam tal função neural nos modelos de TEA (MAIN; KULESZA, 2017).
Ainda em relação aos ratos fêmeas, o valor de tempo final significantemente mais alto dos
grupos Mix-1+AVP e Mix-2+AVP em relação ao grupo AVP demonstra que possivelmente a
administração dos Mix-1 e Mix-2 estão implicados no melhor desempenho motor no teste. Em
um paralelo com humanos, tal resultado condiz com conclusões da metanálise de estudos

102
clínicos controlados e randomizados realizada por Cheng et al, que conclui que a suplementação
com ácidos graxos da família ω-3 pode estar implicada na melhora de escores relacionados à
atividade locomotora (hiperatividade, estereotipias e letargia) em indivíduos com TEA
(CHENG et al., 2017).
Na análise dos resultados dos ratos machos, com exceção da discrepância entre o
grupo Mix-2+AVP em relação ao grupo Mix-2, não houve diferença significante nas variáveis
avaliadas em relação ao grupo SHAM e ao grupo AVP, limitando assim a interpretação dos
resultados, não sendo encontrada melhora significante estatisticamente no desempenho motor
que possa ser atribuída às substâncias testadas. Há várias hipóteses para explicar essa diferença
expressiva dos resultados entre sexos. Evidências recentes mostram alterações diferentes nos
circuitos neurais de ratos machos em relação a ratos fêmeas quando induzidos ao TEA por AVP,
como redução de conexões entre córtex visual e cerebelo, reduzidas de forma bem mais nítida
em machos, além de circuitos que envolvem a medula e o córtex motor, que encontram-se
aumentados nos machos em relação às fêmeas (CHO et al., 2017). Ademais, as concentrações
de corticoesterona nos ratos machos induzidos ao modelo de TEA por AVP são mais elevados
do que os das fêmeas, além de diferença expressiva na maturação do sistema nervoso central
nos vastos aspectos que possuem envolvimento com hormônios sexuais, fatos que podem levar
à uma grande variância em testes de desempenho motor (ASHWOOD et al., 2006).
O estresse oxidativo pode ser caraterizado por um desequilíbrio metabólico entre
espécies pró-oxidantes e antioxidantes. Os agentes pró-oxidantes podem ser representados por
espécies reativas de oxigênio (ROS) e nitrogênio (RNS), já os anti-oxidantes são sistemas de
defesa constituídos por enzimas como, por exemplo: catalase (CAT), superóxido dismutase
(SOD) e glutationa peroxidase (GSH) (PISOCHI et al., 2015).
Ao serem analisados os resultados na Figura 28 no animal macho, os valores para
o hipocampo contemplado no grupo tratado com AVP em comparação ao grupo controle
SHAM, apresentou um aumento significante quando considerados aos marcadores de
peroxidação lipídica para TBARS; não sendo representado igualmente na Figura 29 para as
fêmeas na mesma área.
Quanto à análise inter-grupos, (Mix-2 + AVP e Mix-3 + AVP), tiveram uma
redução significante dos valores do estresse oxidativo ao serem comparados com o grupo Mix1,
ou seja, os mixes ricos em EPA e DHA sendo caracterizados com alto poder anti-inflamatório
e antioxidante, foram capazes de atenuar a concentração dos valores da peroxidação lipídica,
validando uma proteção notável nos filhotes tratados e com autismo. Estes resultados refletem
claramente a exacerbação do estresse oxidativo pelo AVP e demonstração na eficácia destas

103
gorduras EPA/DHA em proteger contra o estresse oxidativo quando comparados aos demais
grupos.
Corroborando com o presente estudo (YADAV et al., 2016) ao serem analisados os
efeitos do ALA e o GLA nas características comportamentais e neuroquímicas em um modelo
animal com autismo no 14ª dia pós-nascimento, comprovou-se que administração de AVP
proporcionou aumento nos níveis de TBARs, e que foi significantemente restaurado por ômega
3 rico em ALA e GLA.
Córtex pré-frontal é uma parte crítica do sistema executivo que se refere à
capacidade de planejar, desempenho cognitivo e raciocínio. Nesta pesquisa foi possível
observar na Figura 28, diferença estatística no grupo ácido valpróico elevando a peroxidação
lipídica no córtex pré-frontal de animais machos, comprometendo não somente as funções do
hipocampo “mencionado anteriormente” como também do córtex pré-frontal. Uma avaliação
entre os grupos AVP tratados, a suplementação de ômega 3 à base de algas (Mix-3 + AVP =
ALA + DHA), foi capaz de impedir estatisticamente quando comparada ao grupo Mix-1 + AVP
rico em ALA, a geração de TBARs, constituindo uma defesa enzimática para combater a
perioxidação lipídica (AL-GADANI et al., 2009; ZOROGLU et al., 2004), e melhorar o
desempenho cognitivo destes animais ocasionado pelo ácido valpróico. Al-Amin et al estudou
o efeito da astaxantina, um poderoso antioxidante, no modelo animal de AVP, encontrando uma
correlação sólida entre o aumento dos níveis de TBARS no córtex pré-frontal no grupo AVP e
o pior desempenho em testes comportamentais de funções sociais (teste de interação social).
Tal déficit comportamental foi normalizado após o tratamento anti-oxidante com a astaxantina,
que reduziu significativamente os níveis de TBARS no córtex dos animais tratados (AL-AMIN
et al, 2015). Este e inúmeros outros resultados presentes na literatura corroboram com estudos
em humanos que demonstram que alterações estruturais e o aumento do estresse oxidativo em
áreas como o córtex pré-frontal orbitofrontal e dorsolateral são centrais na fisiopatologia da
doença, e a redução do estresse oxidativo nestas áreas, especialmente em testes em ratos, tem
se mostrado constantemente ligada a um melhor desempenho nos testes comportamentais
sociais e executivos (SAJDEL-SULKOWSKA et al, 2011).
Estes resultados podem consolidar os valores apresentados neste estudo para
aumento significativo de TBARS nas amígdalas dos animais machos do grupo Mix-1 + AVP
quando comparados ao grupo Mix-3 + AVP e Mix-1 onde foram capazes de expressar valores
inferiores, e que ao correlacionar com os valores inversos para NO nas amígdalas na Figura 31,
o Mix-1 apresentou valores superiores comparado com Mix-1 + AVP.

104
A mistura de óleos do Mix-1 é caracterizada por ter baixa relação w-6/w-3, devendo
apresentar efeitos anti-inflamatórios a nível de membrana celular. Sabemos que o ácido graxo
ômega 6 (LA) - 18: 2, ω-6) é convertido em ácido gama-linolênico (GLA) (18:3, ω-6) no corpo,
que é posteriormente decomposto em AA (20: 4, ω-6). Em divergência com o fato de o GLA
ser uma das moléculas intermediárias para a síntese de AA (20: 4, ω-6), a literatura sugere que
o GLA desempenha um papel importante na alocação de inflamação em todo o corpo. Assim,
mesmo o grupo Mix-1 + AV recebendo uma única dose elevada de ácido valpróico, não justifica
ser diferente estatisticamente dos outros grupos autistas tratados, até porque o grupo Mix3 +
AV apresentou um poder de proteção na peroxidação lipídica maior quando comparado a estes
resultados. Portanto, existe um dilema científico para definir o potencial / mecanismo anti-
inflamatório do ácido graxo ω-3 e / ou ω-6 (RUSSO et al., 2009; KAPOOR et al., 2006).
Resultado antagônico foi observado para os machos na área cerebelar neste estudo,
onde o grupo Sham gerou um estresse oxidativo quando comparado com o grupo autista, não
sendo desta forma consistente com os artigos científicos mencionados na literatura acadêmica
descritos abaixo.
O malonyldialdeído (MDA) é gerado pela peroxidação de ácidos graxos
poliinsaturados e ésteres relacionados e é usado como marcador de LPO (ESSA et al. 2012).
Um estudo mostrou que o conteúdo de MDA no plasma era maior em 13 de 15 (87%) dos
indivíduos autistas, em comparação com indivíduos normais. Zoroglu e cols. (2004) relataram
níveis aumentados de espécies reativas ao ácido tiobarbitúrico no soro de pacientes
autistas. Analisando os tecidos cerebrais (amígdalas e cerebelo) neste estudo e em ambos os
sexos, não foi possível comprovar o aumento de MDA nestas áreas no grupo AV, porém o
grupo Mix 1 elevou significativamente o estresse oxidativo em comparação ao Mix-1 + AV.
Para os animais fêmeas, o grupo Mix2 nos tecidos amígdalas e cerebelo Vários estudos clínicos
realizados, sugerem que suplementar lactentes com AA, EPA e DHA melhorou
significativamente seu desenvolvimento cognitivo e memória (LUXWOLDA et al., 2011;
VRABLIK, 2013). Este benefício poderia se estender sendo iniciado a partir do estágio da
gravidez até o crescimento cerebral onde o desenvolvimento está completo (DAS, 2013). Além
disso, os PUFAs inibiram produção de citocinas neurotóxicas TNF-alfa IL-1 e IL-6; e aumentou
a síntese do óxido nítrico (NO), evento que impedem a apopotose neuronal e facilitam melhoria
e consolidação de memória (RUSSO, 2006; AL-GADANI, 2009; CROEN, 2002; LYALL,
2013).
No teste do Rotarod, no qual se observou um déficit sensório-motor nos ratos
fêmeas do grupo AVP e desempenho normal nos grupos tratados com Mix 1 e Mix 2, é possível

105
postular que tal desempenho esteja associado à redução dos níveis de TBARS nos grupos AVP
fêmeas tratados com Mix 1 e Mix 2, visto que Morakotsriwan et al demonstrou que o aumento
do estresse oxidativo e de alterações histopatológicas como rarefação de células de Purkinje no
cerebelo estão provavelmente ligadas ao pior desempenho motor dos animais do grupo AVP
no teste de Rotarod e, por raciocínio análogo, a melhora de tal desempenho estaria associada à
redução dos níveis de radicais livres e ao mantimento da histopatologia normal do cerebelo,
efeito promovido por certas substâncias com poder antioxidante e neuroprotetor, como os
componentes dos Mixes utilizados no presente estudo (MORAKOTSRIWAN, et al., 2016). O
melhor desempenho no teste de performance de natação, observado nos animais dos grupos
AVP tratados com Mix 2 e Mix 3, pode também ser explicado parcialmente pelo mesmo
mecanismo cerebelar, visto que o cerebelo está implicado na integração motora e na ontogênese
da motricidade coordenada (FATEMI, S. H. et al, 2012).
A glutationa (GSH) é o antioxidante primário responsável por manter o
microambiente intracelular redutor, essencial para a função e viabilidade celular normal. É um
indicador confiável na redução do status celular, e na sua redução crônica o GSH / GSSG reflete
uma capacidade antioxidante reduzida e maior vulnerabilidade a danos oxidativos (JONES et
al., 2006). Recentemente, um aumento na proteína oxidativa e no dano ao DNA foi associado
à diminuição do GSH / GSSG plasmático e intracelular em crianças com autismo, sugerindo
que a capacidade reduzida de defesa antioxidante nessas crianças pode ter consequências
funcionais manifestadas em termos de dano oxidativo (MELNYK et al., 2012).
Com o intuito de avaliar possíveis mecanismos associados aos efeitos preventivos
dos diferentes tipos de ômega-3, buscou-se avaliar parâmetros neuro-oxidativos. Neste
contexto, a glutationa reduzida (GSH) é considerada um antioxidante não enzimático que regula
o equilíbrio redox e o estresse oxidativo (LU et al., 2013; PASTORE et al., 2001), estando
fortemente implicada no TEA, dada sua interação com neurotransmissão glutamatérgica. Os
resultados do presente estudo revelaram que os níveis de GSH não diferiram significativamente
entre o grupo AVP e o Salina em ambos os sexos e nem com os demais grupos estudados.
Yadav et al (2016), relata efeitos positivos no aumento de glutationa ao realizar um
protocolo equivalente ao que foi utilizado pela nossa equipe: indução com AVP no 14ª dia pós-
natal com uma dose de 400mg/kg/p, e tratamento com administração dos PUFAs por 25 dias
de ácido α linoléico (ALA) ou γ ácido linoléico (GLA), sendo esta a diferença do nosso estudo,
onde os grupos autistas e neurotípicos receberam apenas 1 tipo de ácido graxo insaturado.
Os PUFAs e seus produtos metabólicos têm sido implicados no TEA através de
seus papéis na estrutura e função cerebral, neurotransmissão, estrutura da membrana celular e

106
organização dos microdomínios, inflamação, imunidade e estresse oxidativo (MCNAMARA et
al., 2010; WITTE et al., 2013; PU et al., 2013; TANG et al., 2016).
As concentrações de ácidos graxos poliinsaturados no sangue (plasma, soro,
glóbulos vermelhos e sangue total) são consideradas biomarcadores confiáveis (SERRA-
MAJEM et al., 2012). Foi relatada níveis anormais sanguíneos de n -3 LCPUFA em distúrbios
psiquiátricos, incluindo, entre outros, transtorno do déficit de atenção e hiperatividade (TDAH)
e TEA (SPAHIS et al., 2008; KHAN et al., 2002; BRIGANDI et al., 2015). Sugeriu-se que as
explicações para tais anormalidades sejam uma menor ingestão alimentar de n -3 LCPUFAs, e
distúrbios no metabolismo e na incorporação destes ácidos graxos nas membranas celulares de
populações autistas em comparação com controles saudáveis (KHAN et al.2002; BRIGANDI
et al., 2015). Um grande número de relatórios indica diferenças nas proporções de n -3
PUFAs, n -6 PUFAs e / ou n -6 para n -3 PUFA entre populações com autismo e controles
saudáveis (MOSTAFA et al., 2015), mas alguns também não mostraram diferenças (BELL et
al., 2010). O motivo de tais discrepâncias não é bem examinado e não houve tentativas de
comparar sistematicamente esses estudos.
A evidência inicial de óxidos de nitrogênio no metabolismo veio de experimentos
que demonstraram produção de nitratos em camundongos germ-free no início da década de 80
(GREEN et al., 1981). Em 1985 demonstrou-se que macrófagos ativados por lipopossacárides
bacterianos eram capazes de levar à produção de nitritos e nitratos (STUEHR 1985). Na
seqüência evidenciou-se que a L-arginina era o substrato e a L-citrulina era formada como co-
produto (HIBBS et al., 1987). Em 1988, Marletta identificou o óxido nítrico como o produto
da reação de oxi-redução da L-arginina (MARLETTA et al., 1988). Quase que
simultaneamente, Furchgott investigava um fator vasodilatador associado ao endotélio vascular
(endothelium-derived relaxing fator) – EDRF (FURCHGOTT et al., 1984) e poucos anos mais
tarde concluiu-se ser o NO responsável pela atividade biológica do EDRF (IGNARRO et al.,
1987). Durante o final da década de 80 e início de 90, a comunidade científica aprofundou
pesquisas nesta direção, adicionando importantes conhecimentos sobre o NO como mensageiro
(ou sinalizador inter e intracelular) e como toxina, atuando em inúmeros processos patológicos.
O óxido nítrico (NO), produzido a partir de L-arginina pela família de enzimas
óxido nítrico sintetase (NOS), é uma importante molécula de segundo mensageiro que regula
várias funções fisiológicas. Nas células endoteliais, relaxa o músculo liso, o que diminui a
pressão sanguínea. As células de macrófagos produzem NO como um sistema de defesa
imunológica para destruir patógenos e microorganismos. Nas células neuronais, o NO controla
a liberação de neurotransmissores e está envolvido na sinaptogênese, plasticidade sináptica,

107
função de memória e secreção neuroendócrina. Acredita-se que o NO seja um radical livre que
geralmente contribui para o dano oxidativo e a destruição de moléculas e tecidos e, portanto, é
um tanto surpreendente que ele tenha tantos efeitos fisiológicos benéficos significativos.
Óxido nítrico sintase (NOS) é uma família de síntase catalítica, incluindo a NOS
neuronal (nNOS ou NOS1), NOS endotelial (eNOS ou NOS3) e NOS indutível (iNOS ou
NOS2), que sintetizam a produção de NO, uma molécula de sinalização de gás (ALDERTON
et al., 2001). A isoforma endotelial, eNOS, é constitutivamente expressa nas células endoteliais,
enquanto a isoforma do macrófago, iNOS, não é expressa constitutivamente como mencionado
anteriormente, mas é induzida por citocinAS (HEVEL et al., 1991), como corroborando pelo
nosso estudo no grupo AVP na área cerebelar dos animais machos.
A alta produção de NO nos distúrbios do autismo é um fenômeno bem relatado e o
mesmo foi evidente nos achados de YADAV et al., (2016) após a administração do AVP. A
administração concomitante de GLA (ácido graxo gama-linolênico) foi credenciada com menor
produção de NO, fundamentais para os seres humanos devido à sua ampla participação nos
papéis do sistema fisiológico, incluindo cérebro em desenvolvimento.
Investigações realizadas com doenças neurodegenerativas (OZYURT et al., 2014),
cita que mediante as alterações oxidativas na esquizofrenia, aponta-se que níveis elevados de
metabólitos do óxido nítrico e associados a outros radicais livres, provavelmente contribuem
para a neurobiologia do transtorno.
Outro estudo com modelo animal de esquizofrenia realizado com camundongos
machos e fêmeas, apresentaram níveis elevados de nitrito no CPF e HC nas fêmeas, sendo
amenizado com uso de ômega-3 no 30ª dia no pós-nascimento por 20 dias ininterruptos nas
áreas do CPF, HC e CE.
Como já abordado, o NO esteve alterado em todos os estudos onde houve
estimulação através de metabólitos tóxicos responsáveis pelos processos patológicos (iNOS),
não sendo obtido a mesma resposta áreas cerebrais dos grupos AVP de animais machos e
fêmeas utilizados neste estudo.
Como mencionado anteriormente, o ácido ômega-3 docosahexaenóico (DHA) e o
ácido eicosapentaenóico (EPA), além do ácido ômega-6 araquidônico (AA), são três LC-
PUFAs de particular importância para o desenvolvimento e funcionamento ideal do cérebro,
com efeitos independentes e compartilhados (DYALL et al., 2015), são incorporados aos
fosfolipídios da membrana e a incorporação do DHA ocorre em níveis excepcionalmente altos
no sistema nervoso central (LAURITZEN et al., 2016).

108
Os avanços recentes no campo da genética como triagem e investigações do genoma
de indivíduos e parentes afetados pelos TEA, promoveram uma explosão de informações sobre
a fisiopatologia desses transtornos (LISIK et al., 2014).
Apesar de anomalias de quase todos os cromossomos já terem sido associadas ao
autismo (COUTINHO et al., 2015) esses avanços permitiram detectar bases genéticas
heterogêneas e complexas de deficiência e, parcialmente, a compreensão de processos que
resultam diferentes fenótipos (LISIK et al., 2014).
O Gene Chd2 pertence a uma família de proteínas de remodelação da cromatina
dependente de ATP (LAMAR; CARVILL, 2018) tem demonstrado mutações em membros
dessa família que tem sido associadas a distúrbios do neurodesenvolvimento, como transtorno
do espectro do autismo, deficiência intelectual e epilepsia (CARVILL et al., 2013, Allen et al.,
2013). Mutações encontradas no gene Chd2 incluem deleção parcial ou completa do gene,
todas são descritas como de novo e um alelo mutante é suficiente para causar a doença
(CARVILL et al., 2013, ALLEN et al., 2013) Apesar dessa correlação entre mutações
no Chd2 e doenças neurológicas, pouco se sabe sobre o papel desse gene no desenvolvimento
do cérebro.
Estudos em camundongos demonstraram que as deleções homozigóticas
de Chd2 produzem um atraso geral no crescimento e letalidade perinatal, enquanto as deleções
afetam a viabilidade neonatal e lesões não neoplásicas na maioria dos órgãos primários
(MARFELLA et al., 2006), mais recentemente, foi descrito por knockdown que
Chd2 promove uma diferenciação neuronal prematura durante o desenvolvimento cortical do
camundongos embrionários devido a uma diminuição nas células progenitoras neurais Pax6 + e
um aumento nas células progenitoras intermediárias Tbr2 + (SHEN et al., 2015).
Para identificar novas causas de crescimento humano, foi realizado um
sequenciamento do exoma em dez trios probandos-pais e detectado duas mutações Dnmt3a de
novo. Neste estudo foi possível detectar 11 mutações de novo adicionais sequenciados pelo gene
Dnmt3a em mais 142 indivíduos com crescimento excessivo. Estas mutações alteram resíduos
nos domínios funcionais do Dnmt3a e a modelagem de proteínas sugere que elas interferem nas
interações domínio-domínio e na ligação de histonas. Assim, o Dnmt3a se une a um grupo
emergente de genes epigenéticos modificadores de DNA e histona associados a distúrbios do
crescimento do desenvolvimento e doenças malignas hematológicas (TATTON-BROWN et
al., 2014).
Estudo realizado na identificação de novas mutações utilizando exoma completo do
gene Dnmt3a que causam a síndrome de West observou que certos genes de alta confiança para

109
epilepsia foram compartilhados entre as redes de co-expressão e interação genética
de Dnmt3a, Cdkl5 e Kmt2a. Além disso, todos os genes candidatos à epilepsia na rede de co-
expressão do Dnmt3a foram significativamente enriquecidos nos estágios iniciais de
desenvolvimento do cérebro, em particular, relataram que os DNMs do Dnmt3a foram
compartilhados entre epilepsia, transtorno do espectro autista (TEA) e deficiência intelectual
(DI) e ocorreram principalmente no domínio funcional do Dnmt3a (LIU et al., 2018).
Além das variáveis do estresse oxidativo, nitrito e ação antioxidante proveniente do
GSH, no presente estudo, os grupos controles da área HC do sexo macho: SHAM, AVP e os
grupos que no momento da coleta de material expressaram melhor desempenho nos resultados
comportamentais: grupos Mix-3 e Mix-3 + AVP a análise da expressão gênica dos gens Chd2
e Dnmt3a como possíveis mediadores genéticos expressivos como responsáveis pelo
desenvolvimento do TEA. No entanto, a expressão dos genes neste estudo não apresentou
resultados significantes nos grupos e áreas analisadas.
O principal achado do presente estudo foram as respostas e alterações
comportamentais em períodos diferentes, nos animais machos e fêmeas a longo prazo induzidos
pelo desafio do TEA com uso de Ácido Valpróico no 14ª dia pós-nascimento e sua mimetização
com a infusão de ácidos graxos ômega-3, ômega-6 e ômega-9.
Futuramente, novos experimentos poderão ser realizados em animais ratos machos
e fêmeas induzidos ao TEA com ácido valpróico, acrescentando outros testes comportamentais
tais como: interação social, reconhecimento de objetos. Estudos futuros para elucidar possíveis
ações e mecanismos das misturas de óleos, em outros tempos distintos tais como fase
considerada adolescência e adulta em ratos. Estuda outra vari’veis tais como atividade
enzimática COX e LOX para medir ação inflamatória. Avaliar outros fatores epigenéticos como
algumas citocinas estudadas no autismo (IL-1, IL-6, Il-8 e TNF- alfa) e correlacionar aos
resultados e morfologia neuronal pelo método Golgi Cox.

110
6 CONCLUSÃO
1. O modelo experimental induzido por Ácido Valpróico no 14ª dia pós-nascimento
desencadeia alterações comportamentais e neurobioquímicas semelhantes às que ocorrrem
em portadores de transtorno do especto autista.
2. Os mixes de óleos estudados promovem melhora nas alterações comportamentais dos
animais submetidos à indução de modelo de autismo pelo ácido valpróico.
3. Os mixes de óleos reduzem a perioxidação lipídica mas não conferem proteção anti-
oxidante pela GSH.
4. Neste modelo a relação nitrito/nitrato no tecido cerebelar nos animais machos aumenta
significantemente. Os mixes estudados reduzem esta relação para as mesmas apresentadas
pelo grupo controle.
5. Não há impacto sobre a expressão dos genes Dmnt3a e Chd2 na área cerebral hipocampo
com o uso dos mixes de óleos.

111
REFERÊNCIAS
ABEDI, Elahe; SAHARI, Mohammad Ali. Long-chain polyunsaturated fatty acid sources and evaluation of their nutritional and functional properties. Food science & nutrition, v. 2, n. 5, p. 443-463, 2014. ABDOLMALEKY, Hamid Mostafavi et al. Epigenetic and pharmacoepigenomic studies of major psychoses and potentials for therapeutics. 2008. ABRAHAMS, B. S.; GESCHWIND, D. H. Advances in autism genetics: on the threshold of a new neurobiology. Nature reviews genetics, v. 9, n. 5, p. 341-355, 2008. ABRAMOVITCH, S. Correntes teóricas sobre o autismo infantil precoce. Psicanálise e psiquiatria: controvérsias e convergências. In: QUINET, Antonio (org.). Psicanálise e psiquiatria: controvérsias e convergências. Rio de Janeiro: Rios Ambiciosos, p. 255-260, 2001. AJURIAGUERRA, Julián; LÓPEZ-ZEA, Aurelio. Manual de psiquiatría infantil. Masson, 1977. ALDERTON, Wendy K.; COOPER, Chris E.; KNOWLES, Richard G. Nitric oxide synthases: structure, function and inhibition. Biochemical journal, v. 357, n. 3, p. 593-615, 2001. ALDINGER, Kimberly A. et al. Patterns of risk for multiple co-occurring medical conditions replicate across distinct cohorts of children with autism spectrum disorder. Autism Research, v. 8, n. 6, p. 771-781, 2015. AL-GADANI, Y1 et al. Metabolic biomarkers related to oxidative stress and antioxidant status in Saudi autistic children. Clinical biochemistry, v. 42, n. 10-11, p. 1032-1040, 2009. ALLAYEE, Hooman; ROTH, Nitzan; HODIS, Howard N. Polyunsaturated fatty acids and cardiovascular disease: implications for nutrigenetics. Lifestyle Genomics, v. 2, n. 3, p. 140-148, 2009. ALLEN, Andrew S. et al. De novo mutations in epileptic encephalopathies. Nature, v. 501, n. 7466, p. 217-221, 2013. ALSDORF, Rachel; WYSZYNSKI, Diego F. Teratogenicity of sodium valproate. Expert opinion on drug safety, v. 4, n. 2, p. 345-353, 2005. AMAN, Michael G.; LANGWORTHY, Kristen S. Pharmacotherapy for hyperactivity in children with autism and other pervasive developmental disorders. Journal of Autism and Developmental Disorders, v. 30, n. 5, p. 451-459, 2000. AMERICAN PSYCHIATRIC ASSOCIATION et al. Manual diagnóstico e estatístico de transtornos mentais IV-TR. Porto Alegre: ARTMED, 2004. AMERICAN PSYCHIATRIC ASSOCIATION et al. Diagnostic and statistical manual of mental disorders (DSM-5®). American Psychiatric Pub, 2013.

112
AMERICAN PSYCHIATRIC ASSOCIATION et al. Diagnostic and statistical manual: mental disorders. American Psychiatric Association, 1952. AMERICAN PSYCHIATRIC ASSOCIATION. Manual de Diagnóstico e Estatística de Transtornos Mentais DSM-IV. 4 ed. São Paulo: Manole, 1994. AMERICAN PSYCHIATRIC ASSOCIATION. Manual Diagnóstico e Estatístico de Transtornos Mentais (DSM-IV-TR). 4 ed. Porto Alegre: Artes Médicas, 2000. ANDERSON, B. M.; MA, David WL. Are all n-3 polyunsaturated fatty acids created equal?. Lipids in health and Disease, v. 8, n. 1, p. 33, 2009. ANDLAUER, Wilfried; FÜRST, Peter. Nutraceuticals: a piece of history, present status and outlook. Food Research International, v. 35, n. 2-3, p. 171-176, 2002. ARCHER, John. Tests for emotionality in rats and mice: a review. Animal behaviour, v. 21, n. 2, p. 205-235, 1973. ASHWOOD, P. et al. Elevated plasma cytokines in autism spectrum disorders provide evidence of immune dysfunction and are associated with impaired behavioral outcome. Brain, behavior, and immunity, v. 25, n. 1, p. 40-45, 2011. ASHWOOD, P.; WILLS, S.; VAN DE WATER, Judy. The immune response in autism: a new frontier for autism research. Journal of leukocyte biology, v. 80, n. 1, p. 1-15, 2006. ASSUMPÇÃO JUNIOR, F. B.; KUCZYNSKI, E. Anormalidades genéticas e autismo infantil. Anormalidades genéticas e autismo infantil, 2015. ASSUMPÇÃO JUNIOR, F. B.; KUCZYNSKI, E. Autismo infantil: novas tendências e perspectivas. Atheneu, 2007. ASSUMPÇÃO JUNIOR, F. B.; Transtornos abrangentes do desenvolvimento. Tratado de psiquiatria da infância e adolescência, p. 265-280, 2003. AUYEUNG, B. et al. Fetal testosterone and autistic traits. British journal of psychology, v. 100, n. 1, p. 1-22, 2009. BADAWI, N. et al. Autism following a history of newborn encephalopathy: more than a coincidence?. Developmental medicine and child neurology, v. 48, n. 2, p. 85-89, 2006. BAIELI, S. et al. Autism and phenylketonuria. Journal of autism and developmental disorders, v. 33, n. 2, p. 201-204, 2003. BAILEY, A. et al. Autism as a strongly genetic disorder: evidence from a British twin study. Psychological medicine, v. 25, n. 1, p. 63-77, 1995. BAILEY, A.; PHILLIPS, W.; RUTTER, M. Autism: towards an integration of clinical, genetic, neuropsychological, and neurobiological perspectives. Journal of child psychology and psychiatry, v. 37, n. 1, p. 89-126, 1996.

113
BAILEY, Aea et al. A clinicopathological study of autism. Brain: a journal of neurology, v. 121, n. 5, p. 889-905, 1998. BAKER, M. Inside the minds of mice and men. Nature, v. 475, n. 7354, p. 123-128, 2011. BALI, Purva; IM, Heh-In; KENNY, Paul J. Methylation, memory and addiction. Epigenetics, v. 6, n. 6, p. 671-674, 2011. BARON-COHEN, S. Empathizing, systemizing, and the extreme male brain theory of autism. In: Progress in brain research. Elsevier, p. 167-175. 2010 BARONIO, Diego et al. Effects of an H3R antagonist on the animal model of autism induced by prenatal exposure to valproic acid. PLoS One, v. 10, n. 1, 2015. BASSO, D. Michele; BEATTIE, Michael S.; BRESNAHAN, Jacqueline C. A sensitive and reliable locomotor rating scale for open field testing in rats. Journal of neurotrauma, v. 12, n. 1, p. 1-21, 1995. BAUMAN, Margaret L.; KEMPER, Thomas L. Neuroanatomic observations of the brain in autism: a review and future directions. International journal of developmental neuroscience, v. 23, n. 2-3, p. 183-187, 2005. BAUMAN, Margaret; KEMPER, Thomas L. Histoanatomic observations of the brain in early infantile autism. Neurology, v. 35, n. 6, p. 866-866, 1985. BAZINET, R.P.; LAYÉ, S. Polyunsaturated fatty acids and their metabolites in brain function and disease. Nature Reviews Neuroscience, v. 15, n. 12, p. 771-785, 2014. BEDFORD, Rachael et al. Sex differences in the association between infant markers and later autistic traits. Molecular autism, v. 7, n. 1, p. 21, 2016. BELL, John Gordon et al. The fatty acid compositions of erythrocyte and plasma polar lipids in children with autism, developmental delay or typically developing controls and the effect of fish oil intake. British journal of nutrition, v. 103, n. 8, p. 1160-1167, 2010. BELMADANI, Abdelhak et al. Chemokines regulate the migration of neural progenitors to sites of neuroinflammation. Journal of Neuroscience, v. 26, n. 12, p. 3182-3191, 2006. BELZUNG, Catherine et al. Rodent models for autism: a critical review. Drug discovery today: Disease models, v. 2, n. 2, p. 93-101, 2005. BERG, J.M., Tymoczko, J.L., Stryer, L. Biochemistry.W.H. Freeman and Company, New York. 2007. BERNARD, S. et al. The role of mercury in the pathogenesis of autism. Molecular Psychiatry, v. 7, n. 2, p. S42-S43, 2002. BETTELHEIM, Bruno. Empty fortress. Simon and Schuster, 1972.

114
BLEULER E. Dementia Praecox oder die Gruppe der Schizophrenien. In: KAUFMANN P. Dicionário enciclopédico de psicanálise. Rio de Janeiro: Jorge Zahar Editor, 1996. BLUMBERG, Stephen J. et al. Changes in prevalence of parent-reported autism spectrum disorder in school-aged US children: 2007 to 2011-2012. US Department of Health and Human Services, Centers for Disease Control and Prevention, National Center for Health Statistics, 2013. BÖLTE S. Is autism curable? Dev Med Child Neurol. 2014; 56:927-31. BOLTON, Patrick et al. A case-control family history study of autism. Journal of child Psychology and Psychiatry, v. 35, n. 5, p. 877-900, 1994. BORGES, Maria. Autismo-um silêncio ruidoso: perspectiva empírica sobre o autismo no sistema regular de ensino. Almada: Escola Superior de Educação Jean Piaget de Almada, 2000. BOURGEOIS B.F.D.: Pharmacokinetics and pharmacodynamics in clinical practice. In: WYLLIE E. The treatment of epilepsy: principles and practice. Baltimore: Williams & Wilkins, 728-737, 1997. BOURGERON, Thomas. From the genetic architecture to synaptic plasticity in autism spectrum disorder. Nature Reviews Neuroscience, v. 16, n. 9, p. 551-563, 2015. BRENNA, J. Thomas. Efficiency of conversion of α-linolenic acid to long chain n-3 fatty acids in man. Current Opinion in Clinical Nutrition & Metabolic Care, v. 5, n. 2, p. 127-132, 2002. BRIGANDI, Sarah A. et al. Autistic children exhibit decreased levels of essential fatty acids in red blood cells. International journal of molecular sciences, v. 16, n. 5, p. 10061-10076, 2015.
BHAKTA, K.Y., Marlin, S.J., Shen, J.J., Fernandes, C.J., 2005. Terminal deletion of chromosome 15q26.1: case report and brief literature review. Journal of perinatology : official journal of the California Perinatal Association 25, 429-432.
BRUGHA, Traolach S. et al. Epidemiology of autism in adults across age groups and ability levels. The British Journal of Psychiatry, v. 209, n. 6, p. 498-503, 2016. CAMPELO, Ana Paula Bomfim Soares. Óleos ômega 9, 6 e 3 em pele de ratos submetidos a queimadura térmica. 2012. 79 f. Dissertação (Mestrado em Cirurgia) - Faculdade de Medicina. Universidade Federal do Ceará, Fortaleza, 2012. CAMPELO, A. P. B. S; Efeitos nutracêuticos do leite enriquecido com ômega 9, 6 e 3 em ratos submetidos à queimaduras térmica por condução direta. Dissertação (Doutorado em Ciências Médicas Cirúrgicas) – Faculdade de Medicina, Universidade de Fortaleza. Ceará. p50, 2018. CANITANO, Roberto; VIVANTI, Giacomo. Tics and Tourette syndrome in autism spectrum disorders. Autism, v. 11, n. 1, p. 19-28, 2007.

115
CARVILL, G.L., Heavin, S.B., Yendle, S.C., McMahon, J.M., O'Roak, B.J., Cook, J., Khan, A., Dorschner, M.O., Weaver, M., Calvert, S., Malone, S., Wallace, G., Stanley, T., Bye, A.M., Bleasel, A., Howell, K.B., Kivity, S., Mackay, M.T., Rodriguez-Casero, V., Webster, R., Korczyn, A., Afawi, Z., Zelnick, N., Lerman-Sagie, T., Lev, D., Moller, R.S., Gill, D., Andrade, D.M., Freeman, J.L., Sadleir, L.G., Shendure, J., Berkovic, S.F., Scheffer, I.E., Mefford, H.C., 2013. Targeted resequencing in epileptic encephalopathies identifies de novo mutations in CHD2 and SYNGAP1. Nature genetics 45, 825-830.
CARVILL, G., Helbig, I., Mefford, H., 2015. CHD2-Related Neurodevelopmental Disorders, in: Adam, M.P., Ardinger, H.H., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Stephens, K., Amemiya, A. (Eds.), GeneReviews((R)). University of Washington, Seattle University of Washington, Seattle. GeneReviews is a registered trademark of the University of Washington, Seattle. All rights reserved., Seattle (WA).
CAO, Tuoxin et al. Morphological and genetic activation of microglia after diffuse traumatic brain injury in the rat. Neuroscience, v. 225, p. 65-75, 2012. CAPONE, George T. et al. Down syndrome and comorbid autism-spectrum disorder: Characterization using the aberrant behavior checklist. American journal of medical genetics Part A, v. 134, n. 4, p. 373-380, 2005. CARSTENS, Earl; WILSON, C. Rat tail flick reflex: magnitude measurement of stimulus-response function, suppression by morphine and habituation. Journal of neurophysiology, v. 70, n. 2, p. 630-639, 1993. CARVER, Jane D. et al. The relationship between age and the fatty acid composition of cerebral cortex and erythrocytes in human subjects. Brain research bulletin, v. 56, n. 2, p. 79-85, 2001. CARVILL, Gemma L. et al. Targeted resequencing in epileptic encephalopathies identifies de novo mutations in CHD2 and SYNGAP1. Nature genetics, v. 45, n. 7, p. 825, 2013. CENTERS FOR DISEASE CONTROL AND PREVENTION. State and Local Area Integrated Telephone Survey. 2011–2012 National Survey of Children’s Health frequently asked questions. March 2013. Disponível: < http://www.cdc.gov/nchs/slaits/ nsch.htm > CHADMAN, Kathryn K. Animal models for autism in 2017 and the consequential implications to drug discovery. Expert opinion on drug discovery, v. 12, n. 12, p. 1187-1194, 2017. CHANG, Jonathan et al. Genotype to phenotype relationships in autism spectrum disorders. Nature neuroscience, v. 18, n. 2, p. 191-198, 2015. CHAUHAN, Abha; CHAUHAN, Ved. Oxidative stress in autism. Pathophysiology, v. 13, n. 3, p. 171-181, 2006. CHENG, Yu-Shian et al. Supplementation of omega 3 fatty acids may improve hyperactivity, lethargy, and stereotypy in children with autism spectrum disorders: a meta-analysis of randomized controlled trials. Neuropsychiatric disease and treatment, v. 13, p. 2531, 2017. CHESS, Stella; FERNANDEZ, Paulina; KORN, Sam. Behavioral consequences of congenital rubella. The Journal of pediatrics, v. 93, n. 4, p. 699-703, 1978.

116
CHO, Hojin et al. Changes in brain metabolic connectivity underlie autistic-like social deficits in a rat model of autism spectrum disorder. Scientific reports, v. 7, n. 1, p. 1-16, 2017. CHOMIAK, Taylor et al. Altering the trajectory of early postnatal cortical development can lead to structural and behavioural features of autism. BMC neuroscience, v. 11, n. 1, p. 102, 2010. CHRISTENSEN, Deborah L. et al. Prevalence and characteristics of autism spectrum disorder among children aged 8 years—autism and developmental disabilities monitoring network, 11 sites, United States, 2012. MMWR Surveillance Summaries, v. 65, n. 13, p. 1, 2016. CLAYTON, P. E. Normal growth and its endocrine control. Clinical Pediatric Endocrinology, v. 4, p. 95-114, 2001. COLLINS FS, LANDER ES, ROGERS J, WATERSTON RH, CONSO IHGS (2004) Finishing the euchromatic sequence of the human genome. Nature 431(7011): 931-945. COLVERT, Emma et al. Heritability of autism spectrum disorder in a UK population-based twin sample. JAMA psychiatry, v. 72, n. 5, p. 415-423, 2015. COOK H, McMaster C. Fatty acid desaturation and chain elongation in eukaryotes. In: Vance DE, Vance JE, editors.Biochemistry of lipids, lipoproteins and membranes. 4th edition ed.Amsterdam: Elsevier. p. 181-204. 2004. COONEY, Craig A.; DAVE, Apurva A.; WOLFF, George L. Maternal methyl supplements in mice affect epigenetic variation and DNA methylation of offspring. The Journal of nutrition, v. 132, n. 8, p. 2393S-2400S, 2002. COURCHESNE, E. et al. Abnormality of cerebellar vermian lobules VI and VII in patients with infantile autism: identification of hypoplastic and hyperplastic subgroups with MR imaging. AJR. American journal of roentgenology, v. 162, n. 1, p. 123-130, 1994. COUTINHO, J. V. S. C.; BOSSO, Rosa Maria do Vale. Autismo e genética: uma revisão de literatura. Revista Científica do ITPAC, v. 8, n. 1, p. 1-14, 2015. CRAWLEY, Jacqueline N. Behavioral phenotyping of rodents. Comparative medicine, v. 53, n. 2, p. 140-146, 2003. CRAWLEY, Jacqueline N. Mouse behavioral assays relevant to the symptoms of autism. Brain pathology, v. 17, n. 4, p. 448-459, 2007a. CRAWLEY, Jacqueline N. What's wrong with my mouse?: Behavioral phenotyping of transgenic and knockout mice. John Wiley & Sons, 2007b. CROEN, Lisa A. et al. Antidepressant use during pregnancy and childhood autism spectrum disorders. Archives of general psychiatry, v. 68, n. 11, p. 1104-1112, 2011. CROEN, Lisa A. et al. The changing prevalence of autism in California. Journal of autism and developmental disorders, v. 32, n. 3, p. 207-215, 2002.

117
CZYŻ, Katarzyna et al. Omega-3 fatty acids and their role in central nervous system-a review. Current medicinal chemistry, v. 23, n. 8, p. 816-831, 2016. DAHELE, M.; FEARON, K. Imunomodulação Lipídica e Câncer. Dieta, Nutrição e Câncer, 2004. DAS, U. N. Biological significance of essential fatty acids. Journal-Association Of Physicians of India, v. 54, n. R, p. 309, 2006. DAS, Undurti N. Autism as a disorder of deficiency of brain-derived neurotrophic factor and altered metabolism of polyunsaturated fatty acids. Nutrition, v. 29, n. 10, p. 1175-1185, 2013. DEAN, JCh. Valproate. The treatment of epilepsy: Principles and practices, p. 915-923, 1993. DEFOORT, Ericka N.; KIM, Perry M.; WINN, Louise M. Valproic acid increases conservative homologous recombination frequency and reactive oxygen species formation: a potential mechanism for valproic acid-induced neural tube defects. Molecular pharmacology, v. 69, n. 4, p. 1304-1310, 2006. DELAVAL, Katia; WAGSCHAL, Alexandre; FEIL, Robert. Epigenetic deregulation of imprinting in congenital diseases of aberrant growth. Bioessays, v. 28, n. 5, p. 453-459, 2006. DELGADO-ESCUETA, Antonio V.; JANZ, Dieter. Consensus guidelines: preconception counseling, management, and care of the pregnant woman with epilepsy. Neurology, v. 42, n. 4 Suppl 5, p. 149-160, 1992. DEVERMAN, Benjamin E.; PATTERSON, Paul H. Cytokines and CNS development. Neuron, v. 64, n. 1, p. 61-78, 2009. DICKINSON, Ronald G. et al. Transmission of valproic acid (Depakene) across theplacenta: Half-life of the drug in mother and baby. The Journal of pediatrics, v. 94, n. 5, p. 832-835, 1979. DIGUISEPPI, Carolyn et al. Screening for autism spectrum disorders in children with Down syndrome: population prevalence and screening test characteristics. Journal of developmental and behavioral pediatrics: JDBP, v. 31, n. 3, p. 181, 2010. DUARTE, C. S.; BORDIN, I. A. S.; JENSEN, Peter. A abordagem clínica das mães de crianças autistas. Psiquiatria na Prática Médica, v. 34, n. 2, p. 3-5, 2001. DURKIN, Maureen S. et al. Advanced parental age and the risk of autism spectrum disorder. American journal of epidemiology, v. 168, n. 11, p. 1268-1276, 2008. DYALL, Simon C. Long-chain omega-3 fatty acids and the brain: a review of the independent and shared effects of EPA, DPA and DHA. Frontiers in aging neuroscience, v. 7, p. 52, 2015. ECKER, Christine et al. Investigating the predictive value of whole-brain structural MR scans in autism: a pattern classification approach. Neuroimage, v. 49, n. 1, p. 44-56, 2010.

118
EHLERS, C. L. et al. Corticotropin releasing factor (CRF): studies in alcohol preferring and non-preferring rats. Psychopharmacology, v. 106, n. 3, p. 359-364, 1992. EISENBERG, Leon; KANNER, Leo. Childhood schizophrenia: Symposium, 1955: 6. Early infantile autism, 1943–55. American Journal of Orthopsychiatry, v. 26, n. 3, p. 556, 1956. EKDAHL, C. T.; KOKAIA, Zaal; LINDVALL, OLLE. Brain inflammation and adult neurogenesis: the dual role of microglia. Neuroscience, v. 158, n. 3, p. 1021-1029, 2009. EL-ANSARY, Afaf; AL-AYADHI, Laila. Relative abundance of short chain and polyunsaturated fatty acids in propionic acid-induced autistic features in rat pups as potential markers in autism. Lipids in health and disease, v. 13, n. 1, p. 140, 2014. ELLENBROEK, Bart; YOUN, Jiun. Rodent models in neuroscience research: is it a rat race?. Disease models & mechanisms, v. 9, n. 10, p. 1079-1087, 2016. ELSABBAGH, Mayada et al. Global prevalence of autism and other pervasive developmental disorders. Autism research, v. 5, n. 3, p. 160-179, 2012. EUGEN BLEULER; McGlashan TH.: centennial anniversary of his 1911 publication of Dementia Praecox or the group of schizophrenias. Schizophr Bull. 2011;37(6):1101–1103. ESSA, Musthafa M. et al. Increased markers of oxidative stress in autistic children of the Sultanate of Oman. Biological trace element research, v. 147, n. 1-3, p. 25-27, 2012. ESTANISLAU, Celio. Cues to the usefulness of grooming behavior in the evaluation of anxiety in the elevated plus-maze. Psychology & Neuroscience, v. 5, n. 1, p. 105-112, 2012. ESTES, Myka L.; MCALLISTER, A. Kimberley. Maternal immune activation: Implications for neuropsychiatric disorders. Science, v. 353, n. 6301, p. 772-777, 2016. ESTES, Myka L.; MCALLISTER, A. Kimberley. Maternal TH17 cells take a toll on baby's brain. Science, v. 351, n. 6276, p. 919-920, 2016. FATEMI, S. Hossein et al. Consensus paper: pathological role of the cerebellum in autism. The Cerebellum, v. 11, n. 3, p. 777-807, 2012. FEINBERG, Andrew P.; TYCKO, Benjamin. The history of cancer epigenetics. Nature Reviews Cancer, v. 4, n. 2, p. 143-153, 2004. FERRARI, Carlos KB. Functional foods, herbs and nutraceuticals: towards biochemical mechanisms of healthy aging. Biogerontology, v. 5, n. 5, p. 275-289, 2004. FERREIRA, A. L. A.; MATSUBARA, L. S. Radicais livres: conceitos, doenças relacionadas, sistema de defesa e estresse oxidativo. Revista da associação médica brasileira, v. 43, n. 1, p. 61-68, 1997. FIRST, Michael B.; FRANCE, Allen; PINCUS, Harold Alan. DSM-IV-TR guidebook. American Psychiatric Publishing, Inc., 2004.

119
FOLEY, Andrew G. et al. Class I histone deacetylase inhibition ameliorates social cognition and cell adhesion molecule plasticity deficits in a rodent model of autism spectrum disorder. Neuropharmacology, v. 63, n. 4, p. 750-760, 2012. FOLSTEIN, Marshal F.; MAIBERGER, Richard; MCHUGH, Paul R. Mood disorder as a specific complication of stroke. Journal of Neurology, Neurosurgery & Psychiatry, v. 40, n. 10, p. 1018-1020, 1977. FOLSTEIN, Susan E.; ROSEN-SHEIDLEY, Beth. Genetics of austim: complex aetiology for a heterogeneous disorder. Nature Reviews Genetics, v. 2, n. 12, p. 943-955, 2001. FOMBONNE, E. Epidemiology of Autistic Disorder and Other Pervasive Developmental Disorders. Journal Clinical Psychiatry, v. 66, p. 3–8, 2005 FOMBONNE, E. Epidemiology of pervasive developmental disorders. Pediatric Research, v. 65, n. 6, p. 591-598, 2009. FOMBONNE, Eric. The epidemiology of autism: a review. Psychological medicine, v. 29, n. 4, p. 769-786, 1999. FRAZIER, T. W. et al. Behavioral and cognitive characteristics of females and males with autism in the Simons Simplex Collection. Journal of the American Academy of Child and Adolescent Psychiatry, v. 53, n. 3, p. 329-340, 2014. FRAZIER, T. W.; YOUNGSTROM, E. A.; SPEER, L.; EMBACHER, R.; LAW, P.; CONSTANTINO, J.; FINDLING, R. L.; HARDAN, A. Y.; ENG, C. Validation of proposed DSM-5 criteria for autism spectrum disorder. Journal of the American Academy of Child and Adolescent Psychiatry, v. 51, n. 1, p. 28–40, 2012. FURCHGOTT, R. F. et al. Endothelial cells as mediators of vasodilation of arteries. Journal of cardiovascular pharmacology, v. 6, p. S336-43, 1984. FURTADO, Cristiana Libardi Miranda. Epigenetic Markers in Human Diseases. Am J Biomed Sci & Res. 2019 - 6(2). AJBSR. MS.ID.001008. DOI: 10.34297/AJBSR.2019.06.001008. FUSTER, Joaquin. The prefrontal cortex. Academic Press, 2015. GABIS, Lidia V.; POMEROY, John. An Etiologic Classification Autism Spectrum Disorders. Sat, v. 11, p. 20, 2014. GADIA, Carlos A. et al. Autism and pervasive developmental disorders. J Pediatr (Rio J), v. 80, n. 2, p. S83-S94, 2004. GADIA, Carlos A.; TUCHMAN, Roberto; ROTTA, Newra T. Autismo e doenças invasivas de desenvolvimento. Jornal de pediatria, v. 80, n. 2, p. 83-94, 2004. GADOW, Kenneth D.; DEVINCENT, Carla J.; POMEROY, John. ADHD symptom subtypes in children with pervasive developmental disorder. Journal of autism and developmental disorders, v. 36, n. 2, p. 271-283, 2006.

120
GAL, Gilad et al. Time trends in reported autism spectrum disorders in Israel, 1986–2005. Journal of autism and developmental disorders, v. 42, n. 3, p. 428-431, 2012. GARDENER, Hannah; SPIEGELMAN, Donna; BUKA, Stephen L. Perinatal and neonatal risk factors for autism: a comprehensive meta-analysis. Pediatrics, v. 128, n. 2, p. 344-355, 2011. GARDENER, Hannah; SPIEGELMAN, Donna; BUKA, Stephen L. Prenatal risk factors for autism: comprehensive meta-analysis. The British journal of psychiatry, v. 195, n. 1, p. 7-14, 2009. GAUGLER, Trent et al. Most genetic risk for autism resides with common variation. Nature genetics, v. 46, n. 8, p. 881, 2014. GEPNER, Bruno; FÉRON, François. Autism: a world changing too fast for a mis-wired brain?. Neuroscience & Biobehavioral Reviews, v. 33, n. 8, p. 1227-1242, 2009. GESCHWIND, Daniel H. Advances in autism. Annual review of medicine, v. 60, p. 367-380, 2009. GESCHWIND, Daniel H. Genetics of autism spectrum disorders. Trends in cognitive sciences, v. 15, n. 9, p. 409-416, 2011. GESCHWIND, Daniel H.; STATE, Matthew W. Gene hunting in autism spectrum disorder: on the path to precision medicine. The Lancet Neurology, v. 14, n. 11, p. 1109-1120, 2015. GIBALDI, Milo. Biopharmaceutics and clinical pharmacokinetics. 1991. GIBBS, W. W. Além do DNA. Scientific American Brasil, São Paulo, n. 16, Edição Especial, Genoma: o código da vida, p. 44-51, 2007. GIDAYA, Nicole B. et al. In utero exposure to selective serotonin reuptake inhibitors and risk for autism spectrum disorder. Journal of autism and developmental disorders, v. 44, n. 10, p. 2558-2567, 2014. GILLBERG, Christopher. Autism and pervasive developmental disorders. Journal of Child Psychology and Psychiatry, v. 31, n. 1, p. 99-119, 1990. GLASSON, Emma J. et al. Perinatal factors and the development of autism: a population study. Archives of general Psychiatry, v. 61, n. 6, p. 618-627, 2004. GOCKLEY, Jake et al. The female protective effect in autism spectrum disorder is not mediated by a single genetic locus. Molecular autism, v. 6, n. 1, p. 25, 2015. GOINES, Paula; VAN DE WATER, Judy. The immune system’s role in the biology of autism. Current opinion in neurology, v. 23, n. 2, p. 111, 2010. GÓMEZ-GONZÁLEZ, Belén et al. Genome-wide function of THO/TREX in active genes prevents R-loop-dependent replication obstacles. The EMBO journal, v. 30, n. 15, p. 3106-3119, 2011.

121
GÖTTLICHER, Martin et al. Valproic acid defines a novel class of HDAC inhibitors inducing differentiation of transformed cells. The EMBO journal, v. 20, n. 24, p. 6969-6978, 2001. GOWDAK, Marcia Maria Godoy et al. Ácidos graxos ômega-3 e hipertensão arterial sistêmica: Evidências científicas baseadas em ensaios clínicos controlados. Revista Hipertensão, v. 18, n. 3, p. 73-84, 2015. GREEN, Katherine; BRAND, Martin D.; MURPHY, Michael P. Prevention of mitochondrial oxidative damage as a therapeutic strategy in diabetes. Diabetes, v. 53, n. suppl 1, p. S110-S118, 2004. GREEN, Laura C. et al. Nitrate synthesis in the germfree and conventional rat. Science, v. 212, n. 4490, p. 56-58, 1981. GUGLER, R.; VON UNRUH, G. E. Clinical pharmacokinetics of valproic acid. Clinical pharmacokinetics, v. 5, n. 1, p. 67-83, 1980. GUINCHAT, Vincent et al. Pre-, peri-and neonatal risk factors for autism. Acta obstetricia et gynecologica Scandinavica, v. 91, n. 3, p. 287-300, 2012. HALL, Scott S.; LIGHTBODY, Amy A.; REISS, Allan L. Compulsive, self-injurious, and autistic behavior in children and adolescents with fragile X syndrome. American Journal on Mental Retardation, v. 113, n. 1, p. 44-53, 2008. HALLIWELL, Barry; WHITEMAN, Matthew. Measuring reactive species and oxidative damage in vivo and in cell culture: how should you do it and what do the results mean?. British journal of pharmacology, v. 142, n. 2, p. 231-255, 2004. HALLMAYER, Joachim et al. Genetic heritability and shared environmental factors among twin pairs with autism. Archives of general psychiatry, v. 68, n. 11, p. 1095-1102, 2011. HARA, Yuta et al. Oxytocin attenuates deficits in social interaction but not recognition memory in a prenatal valproic acid-induced mouse model of autism. Hormones and behavior, v. 96, p. 130-136, 2017. HERRERA, Emilio. Implications of dietary fatty acids during pregnancy on placental, fetal and postnatal development—a review. Placenta, v. 23, p. S9-S19, 2002. HEVEL, Joan M.; WHITE, Kimberly A.; MARLETTA, Michael A. Purification of the inducible murine macrophage nitric oxide synthase. Identification as a flavoprotein. Journal of biological chemistry, v. 266, n. 34, p. 22789-22791, 1991. HIBBS, John B.; TAINTOR, Read R.; VAVRIN, Zdenek. Macrophage cytotoxicity: role for L-arginine deiminase and imino nitrogen oxidation to nitrite. Science, v. 235, n. 4787, p. 473-476, 1987. HOFFMANN, S. B. Estereotipias na infância. Porto Alegre – RS, 1996. HRABOVSKA, S. V.; SALYHA, Yu T. Animal models of autism spectrum disorders and behavioral techniques of their examination. Neurophysiology, v. 48, n. 5, p. 380-388, 2016.

122
HSIAO, Elaine Y. et al. Modeling an autism risk factor in mice leads to permanent immune dysregulation. Proceedings of the National Academy of Sciences, v. 109, n. 31, p. 12776-12781, 2012. HUANG, Fei et al. Betaine ameliorates prenatal valproic-acid-induced autism-like behavioral abnormalities in mice by promoting homocysteine metabolism. Psychiatry and clinical neurosciences, v. 73, n. 6, p. 317-322, 2019. HUGUET, Guillaume; BENABOU, Marion; BOURGERON, Thomas. The genetics of autism spectrum disorders. In: A time for metabolism and hormones. Springer, Cham, p. 101-129. 2016 HUGUET, Guillaume; BOURGERON, Thomas. Genetic causes of autism spectrum disorders. In: Neuronal and synaptic dysfunction in autism spectrum disorder and intellectual disability. Academic Press, 2016. p. 13-24. HUGUET, Guillaume; EY, Elodie; BOURGERON, Thomas. The genetic landscapes of autism spectrum disorders. Annual review of genomics and human genetics, v. 14, p. 191-213, 2013. HVIID, Anders; MELBYE, Mads; PASTERNAK, Björn. Use of selective serotonin reuptake inhibitors during pregnancy and risk of autism. New England Journal of Medicine, v. 369, n. 25, p. 2406-2415, 2013. IGNARRO, Louis J. et al. Endothelium-derived relaxing factor produced and released from artery and vein is nitric oxide. Proceedings of the National Academy of Sciences, v. 84, n. 24, p. 9265-9269, 1987. INDREDAVIK, Marit S. et al. Low-birth-weight adolescents: psychiatric symptoms and cerebral MRI abnormalities. Pediatric neurology, v. 33, n. 4, p. 259-266, 2005. INGRAM, Jennifer L. et al. Prenatal exposure of rats to valproic acid reproduces the cerebellar anomalies associated with autism. Neurotoxicology and teratology, v. 22, n. 3, p. 319-324, 2000. INNIS, Sheila M. Perinatal biochemistry and physiology of long-chain polyunsaturated fatty acids. The Journal of pediatrics, v. 143, n. 4, p. 1-8, 2003. IOSSIFOV, Ivan et al. The contribution of de novo coding mutations to autism spectrum disorder. Nature, v. 515, n. 7526, p. 216-221, 2014.
IOSSIFOV, I., O'Roak, B.J., Sanders, S.J., Ronemus, M., Krumm, N., Levy, D., Stessman, H.A., Witherspoon, K.T., Vives, L., Patterson, K.E., Smith, J.D., Paeper, B., Nickerson, D.A., Dea, J., Dong, S., Gonzalez, L.E., Mandell, J.D., Mane, S.M., Murtha, M.T., Sullivan, C.A., Walker, M.F., Waqar, Z., Wei, L., Willsey, A.J., Yamrom, B., Lee, Y.H., Grabowska, E., Dalkic, E., Wang, Z., Marks, S., Andrews, P., Leotta, A., Kendall, J., Hakker, I., Rosenbaum, J., Ma, B., Rodgers, L., Troge, J., Narzisi, G., Yoon, S., Schatz, M.C., Ye, K., McCombie, W.R., Shendure, J., Eichler, E.E., State, M.W., Wigler, M., 2014. The contribution of de novo coding mutations to autism spectrum disorder. Nature 515, 216-221.

123
JACQUEMONT, Sébastien et al. A higher mutational burden in females supports a “female protective model” in neurodevelopmental disorders. The American Journal of Human Genetics, v. 94, n. 3, p. 415-425, 2014. JAKOVCEVSKI M, AKBARIAN S. Epigenetic mechanisms in neurodevelopmental and neurodegenerative disease. Nat Med. 2012;18:1194-204. JAMES, Stephen; MONTGOMERY, Paul; WILLIAMS, Katrina. Omega-3 fatty acids supplementation for autism spectrum disorders (ASD). Cochrane Database of Systematic Reviews, n. 11, 2011. JANSSEN, Carola IF; KILIAAN, Amanda J. Long-chain polyunsaturated fatty acids (LCPUFA) from genesis to senescence: the influence of LCPUFA on neural development, aging, and neurodegeneration. Progress in lipid research, v. 53, p. 1-17, 2014. JEONG, Mi Ra et al. Valproic acid, a mood stabilizer and anticonvulsant, protects rat cerebral cortical neurons from spontaneous cell death: a role of histone deacetylase inhibition. FEBS letters, v. 542, n. 1-3, p. 74-78, 2003. JESTE, Shafali S. et al. Characterization of autism in young children with tuberous sclerosis complex. Journal of child neurology, v. 23, n. 5, p. 520-525, 2008. JIRTLE, Randy L.; SKINNER, Michael K. Environmental epigenomics and disease susceptibility. Nature reviews genetics, v. 8, n. 4, p. 253-262, 2007. JOHANNESSEN, Cecilie U.; JOHANNESSEN, Svein I. Valproate: past, present, and future. CNS drug reviews, v. 9, n. 2, p. 199-216, 2003. JOHANNESSEN, Svein I. Pharmacokinetics of valproate in pregnancy: mother-foetus-newborn. Pharmaceutisch Weekblad, v. 14, n. 3, p. 114-117, 1992. JOHNSON C.P.; Myers S.M. Identification and evaluation of children with autism spectrum disorders. Pediatrics, v. 120, n. 5, p. 1183-1215, 2007. JOKIRANTA, Elina et al. Parental psychiatric disorders and autism spectrum disorders. Psychiatry research, v. 207, n. 3, p. 203-211, 2013. JØRGENSEN, Jakob Ploug et al. Adrm1, a putative cell adhesion regulating protein, is a novel proteasome-associated factor. Journal of molecular biology, v. 360, n. 5, p. 1043-1052, 2006. KAHNE, David et al. Behavioral and magnetic resonance spectroscopic studies in the rat hyperserotonemic model of autism. Physiology & behavior, v. 75, n. 3, p. 403-410, 2002. KALUEFF, Allan V.; TUOHIMAA, Pentti. Mouse grooming microstructure is a reliable anxiety marker bidirectionally sensitive to GABAergic drugs. European journal of pharmacology, v. 508, n. 1-3, p. 147-153, 2005a. KALUEFF, Allan V.; TUOHIMAA, Pentti. The grooming analysis algorithm discriminates between different levels of anxiety in rats: potential utility for neurobehavioural stress research. Journal of neuroscience methods, v. 143, n. 2, p. 169-177, 2005b.

124
KANEDA, Masahiro et al. Essential role for de novo DNA methyltransferase Dnmt3a in paternal and maternal imprinting. Nature, v. 429, n. 6994, p. 900-903, 2004. KANNER, L. Autistic disturbances of affective contact. The Nervous Child, v. 2, p. 217–250, 1943. KANNER, Leo. Follow-up study of eleven autistic children originally reported in 1943. Journal of autism and childhood schizophrenia, v. 1, n. 2, p. 119-145, 1971. KANNER, Leo. Problems of nosology and psychodynamics of early infantile autism. American journal of Orthopsychiatry, v. 19, n. 3, p. 416, 1949. KAPINOS-GORCZYCA, A.; GORCZYCA, P.; KAPINOS, M. P02-240 Dysmorphic features and development of children with infantile autism. European Psychiatry, v. 24, p. S930, 2009. KAPOOR, Rakesh; HUANG, Yung-Sheng. Gamma linolenic acid: an antiinflammatory omega-6 fatty acid. Current pharmaceutical biotechnology, v. 7, n. 6, p. 531-534, 2006. KAREN, Weidenheim. Neurobiology of autism: An update. Salud Mental, v. 24, n. 3, p. 3-9, 2001. KATAOKA, Shunsuke et al. Autism-like behaviours with transient histone hyperacetylation in mice treated prenatally with valproic acid. International journal of neuropsychopharmacology, v. 16, n. 1, p. 91-103, 2013. KAU, Alice SM et al. Social behavior profile in young males with fragile X syndrome: characteristics and specificity. American Journal of Medical Genetics Part A, v. 126, n. 1, p. 9-17, 2004. KAZDOBA, Tatiana M. et al. Translational mouse models of autism: advancing toward pharmacological therapeutics. In: Translational Neuropsychopharmacology. Springer, Cham, 2015. p. 1-52. KEMPER, Thomas L.; BAUMAN, Margaret L. The contribution of neuropathologic studies to the understanding of autism. Neurologic clinics, v. 11, n. 1, p. 175-187, 1993. KERN, Janet K.; JONES, Anne M. Evidence of toxicity, oxidative stress, and neuronal insult in autism. Journal of Toxicology and Environmental Health, Part B, v. 9, n. 6, p. 485-499, 2006. KHAN, Mohammad M. et al. Reduced erythrocyte membrane essential fatty acids and increased lipid peroxides in schizophrenia at the never-medicated first-episode of psychosis and after years of treatment with antipsychotics. Schizophrenia research, v. 58, n. 1, p. 1-10, 2002. KIECOLT-GLASER, Janice K. et al. Chronic stress and age-related increases in the proinflammatory cytokine IL-6. Proceedings of the national Academy of Sciences, v. 100, n. 15, p. 9090-9095, 2003.

125
KIM, Ki Chan et al. Clinical and neurobiological relevance of current animal models of autism spectrum disorders. Biomolecules & therapeutics, v. 24, n. 3, p. 207, 2016. KIM, Ki Chan et al. Male-specific alteration in excitatory post-synaptic development and social interaction in pre-natal valproic acid exposure model of autism spectrum disorder. Journal of neurochemistry, v. 124, n. 6, p. 832-843, 2013. KIM, Sun-Young et al. Abnormal activation of the social brain network in children with autism spectrum disorder: an FMRI study. Psychiatry investigation, v. 12, n. 1, p. 37, 2015. KIM, Young-In. Nutritional epigenetics: impact of folate deficiency on DNA methylation and colon cancer susceptibility. The Journal of nutrition, v. 135, n. 11, p. 2703-2709, 2005. KIRSTEN, T. B.; et al. Hypoactivity of the central dopaminergic system and autistic-like behavior induced by a single early prenatal exposure to lipopolysaccharide. Journal of Neuroscience Research, v. 90, n. 10, p. 1903–12, 2012. KIRSTEN, Thiago Berti et al. Lipopolysaccharide exposure induces maternal hypozincemia, and prenatal zinc treatment prevents autistic-like behaviors and disturbances in the striatal dopaminergic and mTOR systems of offspring. PloS one, v. 10, n. 7, 2015. KOLEVZON, Alexander; GROSS, Raz; REICHENBERG, Abraham. Prenatal and perinatal risk factors for autism: a review and integration of findings. Archives of pediatrics & adolescent medicine, v. 161, n. 4, p. 326-333, 2007. KU, C. S. et al. A new paradigm emerges from the study of de novo mutations in the context of neurodevelopmental disease. Molecular psychiatry, v. 18, n. 2, p. 141-153, 2013. KUCZYNSKI, Evelyn. Anormalidades cromossômicas esporádicas associadas à síndrome autística. Revista de Neuropsiquiatria da Infância e Adolescência, v. 4, p. 26-36, 1996. KUNIISHI, Hiroshi et al. Early deprivation increases high-leaning behavior, a novel anxiety-like behavior, in the open field test in rats. Neuroscience research, v. 123, p. 27-35, 2017. LAGACE, Diane C. et al. Valproic acid: how it works. Or not. Clinical Neuroscience Research, v. 4, n. 3-4, p. 215-225, 2004. LAI, M.; LOMBARDO, M. V; BARON-COHEN, S. Autism. Lancet v. 383, n. 8, p. 896–910, 2014. LAI, Meng-Chuan et al. Biological sex affects the neurobiology of autism. Brain, v. 136, n. 9, p. 2799-2815, 2013. LAMAR, Kay-Marie J.; CARVILL, Gemma L. Chromatin remodeling proteins in epilepsy: lessons from CHD2-associated epilepsy. Frontiers in molecular neuroscience, v. 11, p. 208, 2018. LAMAZIERE, A.; Richard, D.; Barbe, U.; Kefi, K.; Bausero, P.; Wolf, C.; Visioli, F. Differential distribuition of DHA-phospholipids in rat brain after feeding a lipidomic approach. Prostaglandins Leukot. Essent. Fatty Acids, v. 84, p. 7-11, 2011.

126
LANDRIGAN, Philip J. What causes autism? Exploring the environmental contribution. Current opinion in pediatrics, v. 22, n. 2, p. 219-225, 2010. LAURITZEN, Lotte et al. DHA effects in brain development and function. Nutrients, v. 8, n. 1, p. 6, 2016. LAURITZEN, Lotte. The essentiality of long chain n-3 fatty acids in relation to development and function of the brain and retina. Prog Lipid Res, v. 40, p. 1-94, 2001. LAZNIK, Marie Christine. A voz da sereia: o autismo e os impasses na constituição do sujeito. Salvador: Ágalma, p. 27-31, 2004. LEE, Patrick F.; THOMAS, Roger E.; LEE, Patricia A. Approach to autism spectrum disorder: Using the new DSM-V diagnostic criteria and the CanMEDS-FM framework. Canadian Family Physician, v. 61, n. 5, p. 421-424, 2015. LEVANT, Beth; RADEL, Jeffery D.; CARLSON, Susan E. Reduced brain DHA content after a single reproductive cycle in female rats fed a diet deficient in N-3 polyunsaturated fatty acids. Biological psychiatry, v. 60, n. 9, p. 987-990, 2006. LEVY, E. S.; MANDELL, D. S.; SCHULTZ, R. Autism. Lancet. London, v. 374, n.9701, p.1627-1638, nov.2009. LEYFER, Ovsanna T. et al. Comorbid psychiatric disorders in children with autism: interview development and rates of disorders. Journal of autism and developmental disorders, v. 36, n. 7, p. 849-861, 2006. LYALL K, Schmidt RJ, Hertz-Picciotto I. Maternal lifestyle and environmental risk factors for autism spectrum disorders. Int J Epidemiol. 2014; 43:443-64. LICHTENSTEIN, Paul et al. The genetics of autism spectrum disorders and related neuropsychiatric disorders in childhood. American Journal of Psychiatry, v. 167, n. 11, p. 1357-1363, 2010. LIMA, M. M.; et al. Ácidos Graxos e Câncer. In: CURI, R. Entendendo a Gordura: os ácidos graxos. São Paulo: Manole, 2002. cap. 37, p. 523-538. LIMPEROPOULOS, Catherine et al. Positive screening for autism in ex-preterm infants: prevalence and risk factors. Pediatrics, v. 121, n. 4, p. 758-765, 2008. LISIK, Małgorzata Z. Molecular aspects of autism spectrum disorders. Psychiatr. Pol, v. 48, n. 4, p. 689-700, 2014. LIU, Zhenwei et al. Identification of de novo DNMT3A mutations that cause west syndrome by using whole-exome sequencing. Molecular neurobiology, v. 55, n. 3, p. 2483-2493, 2018. LOOMES, Rachel; HULL, Laura; MANDY, William Polmear Locke. What is the male-to-female ratio in autism spectrum disorder? A systematic review and meta-analysis. Journal of the American Academy of Child & Adolescent Psychiatry, v. 56, n. 6, p. 466-474, 2017.

127
LORD, C. et al. Autism spectrum disorder. Nature Reviews Disease Primers, v. 6, n. 1, p. 1-23, 2020. LÖSCHER, Wolfgang. Valproate: a reappraisal of its pharmacodynamic properties and mechanisms of action. Progress in neurobiology, v. 58, n. 1, p. 31-59, 1999. LU, Q. et al. Epigenetics, disease, and therapeutic interventions. Ageing research reviews, v. 5, n. 4, p. 449-467, 2006. LUCCHINA, Luciana; DEPINO, Amaicha Mara. Altered peripheral and central inflammatory responses in a mouse model of autism. Autism Research, v. 7, n. 2, p. 273-289, 2014. LUNDSTRÖM, Sebastian et al. Autistic-like traits and their association with mental health problems in two nationwide twin cohorts of children and adults. Psychological medicine, v. 41, n. 11, p. 2423-2433, 2011. LUXWOLDA, Martine F. et al. The relation between the omega-3 index and arachidonic acid is bell shaped: synergistic at low EPA+ DHA status and antagonistic at high EPA+ DHA status. Prostaglandins, leukotrienes and essential fatty acids, v. 85, n. 3-4, p. 171-178, 2011. LYALL, Kristen et al. Maternal dietary fat intake in association with autism spectrum disorders. American journal of epidemiology, v. 178, n. 2, p. 209-220, 2013. MAIMBURG, Rikke Damkjær; VÆTH, Michael. Perinatal risk factors and infantile autism. Acta Psychiatrica Scandinavica, v. 114, n. 4, p. 257-264, 2006. MAIN, Stacey L.; KULESZA, Randy J. Repeated prenatal exposure to valproic acid results in cerebellar hypoplasia and ataxia. Neuroscience, v. 340, p. 34-47, 2017. MANDY, W.; CHILVERS, R.; COMORBIDITY, H. Á. Sex Differences in Autism Spectrum Disorder: Evidence from a Large Sample of Children and Adolescents. Journal of Autism and Developmental Disorders v. 42, n. 7, p. 1304–1313, 2012. MANKAD, Deepali et al. A randomized, placebo controlled trial of omega-3 fatty acids in the treatment of young children with autism. Molecular autism, v. 6, n. 1, p. 18, 2015. MARAMARA, Lauren A.; HE, Wenzhuan; MING, Xue. Pre-and perinatal risk factors for autism spectrum disorder in a New Jersey cohort. Journal of child neurology, v. 29, n. 12, p. 1645-1651, 2014. MAREN, Stephen. Building and burying fear memories in the brain. The Neuroscientist, v. 11, n. 1, p. 89-99, 2005. MARFELLA, Concetta GA et al. Mutation of the SNF2 family member Chd2 affects mouse development and survival. Journal of cellular physiology, v. 209, n. 1, p. 162-171, 2006. MARKRAM, Kamila et al. Abnormal fear conditioning and amygdala processing in an animal model of autism. Neuropsychopharmacology, v. 33, n. 4, p. 901-912, 2008.

128
MARLETTA, Michael A. et al. Macrophage oxidation of L-arginine to nitrite and nitrate: nitric oxide is an intermediate. Biochemistry, v. 27, n. 24, p. 8706-8711, 1988. MARQUES, Carla Elsa Correia Castanheira. Perturbações do espectro do autismo: ensaio de uma intervenção construtivista e desenvolvimentalista com mães. 2000. MARTINS, Yolanda; YOUNG, Robyn L.; ROBSON, Danielle C. Feeding and eating behaviors in children with autism and typically developing children. Journal of autism and developmental disorders, v. 38, n. 10, p. 1878-1887, 2008. MATSON, Johnny L.; NEBEL-SCHWALM, Marie S. Comorbid psychopathology with autism spectrum disorder in children: An overview. Research in developmental disabilities, v. 28, n. 4, p. 341-352, 2007. MCNAMARA, Robert K. et al. Docosahexaenoic acid supplementation increases prefrontal cortex activation during sustained attention in healthy boys: a placebo-controlled, dose-ranging, functional magnetic resonance imaging study. The American journal of clinical nutrition, v. 91, n. 4, p. 1060-1067, 2010. MCNAMARA, Robert K.; VANNEST, Jennifer J.; VALENTINE, Christina J. Role of perinatal long-chain omega-3 fatty acids in cortical circuit maturation: Mechanisms and implications for psychopathology. World journal of psychiatry, v. 5, n. 1, p. 15, 2015. MCPARTLAND, James C.; REICHOW, Brian; VOLKMAR, Fred R. Sensitivity and specificity of proposed DSM-5 diagnostic criteria for autism spectrum disorder. Journal of the American Academy of Child & Adolescent Psychiatry, v. 51, n. 4, p. 368-383, 2012. MEAD, Jennifer; ASHWOOD, Paul. Evidence supporting an altered immune response in ASD. Immunology letters, v. 163, n. 1, p. 49-55, 2015.
MELO, Radamés Bezerra. et al. Antiperoxidative properties of oil mixes of high ratio omega- 9:omega-6 and low ratio omega-6:omega-3 after molar extraction in rats. Acta cir. Bras., v. 29, n. 6, p. 371-375, june, 2014. Disponível em: <http://dx.doi.org/10.1590/s0102- 86502014000600004>. 69
MELO, Radamés Bezerra et al. Anti-inflammatory effect of a fatty acid mixture with high ω-9:ω-6 ratio and low ω-6:ω-3 ratio on rats submitted to dental extraction. Archives of Oral Biology, v. 74, p. 63-68, feb. 2017. Disponível em: <https://www.sciencedirect.com/science/article/pii/S0003996916303326?via%3Dihub>. 70 MESQUITA, T. R. M. et al. Anti-inflammatory effect of dietary supplementation with omega-3 fatty acids in rats. Revista Dor, v. 12, n. 4, p. 337-341, 2011.
MILES, Judith H.; HILLMAN, Richard E. Value of a clinical morphology examination in autism. American journal of medical genetics, v. 91, n. 4, p. 245-253, 2000. MILITERNI, Roberto et al. Pain reactivity in children with autistic disorder. The Journal of Headache and Pain, v. 1, n. 1, p. 53-56, 2000. MILLER, Greg. Signs of empathy seen in mice. 2006.

129
MINES, Marjelo A. et al. GSK3 influences social preference and anxiety-related behaviors during social interaction in a mouse model of fragile X syndrome and autism. PloS one, v. 5, n. 3, 2010. MING, Xue et al. Evidence of oxidative stress in autism derived from animal models. American Journal of Biochemistry and Biotechnology, v. 4, n. 2, p. 218-225, 2008. MINSHEW, Nancy J.; LUNA, Beatriz; SWEENEY, John A. Oculomotor evidence for neocortical systems but not cerebellar dysfunction in autism. Neurology, v. 52, n. 5, p. 917-917, 1999. MOBRATEN, Kaia et al. Omega-3 and omega-6 PUFAs induce the same GPR120-mediated signalling events, but with different kinetics and intensity in Caco-2 cells. Lipids in health and disease, v. 12, n. 1, p. 101, 2013. MOLLOY, Cynthia A. et al. Elevated cytokine levels in children with autism spectrum disorder. Journal of neuroimmunology, v. 172, n. 1-2, p. 198-205, 2006. MOOSAVI, Azam; ARDEKANI, Ali Motevalizadeh. Role of epigenetics in biology and human diseases. Iranian biomedical journal, v. 20, n. 5, p. 246, 2016. MORAKOTSRIWAN, Nartnutda et al. Autistic-like behaviors, oxidative stress status, and histopathological changes in cerebellum of valproic acid rat model of autism are improved by the combined extract of purple rice and silkworm pupae. Oxidative medicine and cellular longevity, v. 2016, 2016. MORALES ALPÍZAR, Catalina; SALAS HERRERA, Isaías. Manejo farmacológico del dolor en el paciente oncológico. Acta Médica Costarricense, v. 46, n. 3, p. 114-124, 2004. MOREIRA, J.D.; Knorr, L.; Thomazi, A. P.; Battú, C.; Oses, J.P.; Gottfried, C.; Wofchuk, S.; Salbego, C.; Souza, D.O.; Perry, M. L.; Vinaê, L.; Dietary ômega-3 fatty acids altenuate damage after hippocampal ischemic insult em adult rats. J. Nutr. Biochem., v. 21, n.4 p. 351-356, 2010. MORGAN, John T. et al. Microglial activation and increased microglial density observed in the dorsolateral prefrontal cortex in autism. Biological psychiatry, v. 68, n. 4, p. 368-376, 2010. MORGANTI-KOSSMANN, Maria Cristina et al. Inflammatory response in acute traumatic brain injury: a double-edged sword. Current opinion in critical care, v. 8, n. 2, p. 101-105, 2002. MORI, Trevor A.; BEILIN, Lawrence J. Omega-3 fatty acids and inflammation. Current atherosclerosis reports, v. 6, n. 6, p. 461-467, 2004. MOSS, J.; HOWLIN, P. Autism spectrum disorders in genetic syndromes: implications for diagnosis, intervention and understanding the wider autism spectrum disorder population. Journal of Intellectual Disability Research, v. 53, n. 10, p. 852-873, 2009. MOSS, Timothy J.; WALLRATH, Lori L. Connections between epigenetic gene silencing and human disease. Mutation Research/Fundamental and Molecular Mechanisms of Mutagenesis, v. 618, n. 1-2, p. 163-174, 2007.

130
MOSTAFA, Gehan A.; AL-AYADHI, Laila Y. Reduced levels of plasma polyunsaturated fatty acids and serum carnitine in autistic children: relation to gastrointestinal manifestations. Behavioral and Brain Functions, v. 11, n. 1, p. 4, 2015. MOZAFFARIAN, Dariush; WU, Jason HY. (n-3) fatty acids and cardiovascular health: are effects of EPA and DHA shared or complementary? The Journal of nutrition, v. 142, n. 3, p. 614S-625S, 2012. MUSTO, Alberto E.; GJORSTRUP, Per; BAZAN, Nicolas G. The omega-3 fatty acid–derived neuroprotectin D1 limits hippocampal hyperexcitability and seizure susceptibility in kindling epileptogenesis. Epilepsia, v. 52, n. 9, p. 1601-1608, 2011. NADON, Geneviève et al. Association of sensory processing and eating problems in children with autism spectrum disorders. Autism research and treatment, v. 2011, 2011. NAKAMURAK, KARIYAZONO H, KOMOKATA T, HAMADA N, SAKATA R, YAMADA K. Influence of preoperative administration of w-3 fatty acid-enriched supplement on inflammatory and immune responses in patients undergoing major surgery for cancer. Nutrition. V.21, p:639-649, 2005. NALIVAEVA, Natalia N.; BELYAEV, Nikolai D.; TURNER, Anthony J. Sodium valproate: an old drug with new roles. Trends in pharmacological sciences, v. 30, n. 10, p. 509-514, 2009. NARITA, Masaaki et al. Nonexploratory movement and behavioral alterations in a thalidomide or valproic acid-induced autism model rat. Neuroscience research, v. 66, n. 1, p. 2-6, 2010. NATASHA, Gemma et al. Valproate and spinal muscular atrophy. Molecular medicine reports, v. 1, n. 2, p. 161-165, 2008. NEMEROFF, Charles B. et al. DSM-5: a collection of psychiatrist views on the changes, controversies, and future directions. BMC medicine, v. 11, n. 1, p. 202, 2013. OBAJIMI, Oluwakemi et al. Differential effects of eicosapentaenoic and docosahexaenoic acids upon oxidant-stimulated release and uptake of arachidonic acid in human lymphoma U937 cells. Pharmacological research, v. 52, n. 2, p. 183-191, 2005. ODDI, D. et al. Monogenic mouse models of social dysfunction: implications for autism. Behavioural brain research, v. 251, p. 75-84, 2013. OGUCHI-KATAYAMA, A. et al. Comparative gene expression analysis of the amygdala in autistic rat models produced by pre-and post-natal exposures to valproic acid. The Journal of toxicological sciences, v. 38, n. 3, p. 391-402, 2013. OLEXOVÁ, Lucia et al. Habituation of exploratory behaviour in VPA rats: animal model of autism. Interdisciplinary toxicology, v. 6, n. 4, p. 222-227, 2013. OLIVEIRA, Karina Griesi. Identificação de genes e vias associadas aos transtornos do espectro autista. Tese de Doutorado. Universidade de São Paulo. 2011.

131
ONORE, Charity E. et al. Maternal immune activation leads to activated inflammatory macrophages in offspring. Brain, behavior, and immunity, v. 38, p. 220-226, 2014 ORGANIZAÇÃO MUNDIAL DA SAÚDE. Classificação de transtornos mentais e de comportamento da CID-10: descrições clínicas e diretrizes diagnósticas. 1993. ORNOY, Asher. Valproic acid in pregnancy: how much are we endangering the embryo and fetus? Reproductive toxicology, v. 28, n. 1, p. 1-10, 2009. OZONOFF, Sally et al. Recurrence risk for autism spectrum disorders: a Baby Siblings Research Consortium study. Pediatrics, v. 128, n. 3, p. e488-e495, 2011. PAOLICELLI, Rosa C. et al. Synaptic pruning by microglia is necessary for normal brain development. science, v. 333, n. 6048, p. 1456-1458, 2011. PARDO, Carlos A.; VARGAS, Diana L.; ZIMMERMAN, Andrew W. Immunity, neuroglia and neuroinflammation in autism. International review of psychiatry, v. 17, n. 6, p. 485-495, 2005. PENDYALA, Gurudutt et al. Maternal immune activation causes behavioral impairments and altered cerebellar cytokine and synaptic protein expression. Neuropsychopharmacology, v. 42, n. 7, p. 1435-1446, 2017.. PINHEIRO, Petrúcia Maria Antero et al. Pré-condicionamento com misturas de óleos com Ômega-9: Ômega-6 (alta relação) e Ômega-6: Ômega-3 (baixa relação) em ratos submetidos à isquemia/reperfusão cerebral. Acta Cirurgica Brasileira, v. 26, p. 32-37, 2011. PINTO, Dalila et al. Convergence of genes and cellular pathways dysregulated in autism spectrum disorders. The American Journal of Human Genetics, v. 94, n. 5, p. 677-694, 2014. PLETNIKOV, Mikhail V.; MORAN, Timothy H.; CARBONE, Kathryn M. Borna disease virus infection of the neonatal rat: developmental brain injury model of autism spectrum disorders. Front Biosci, v. 7, p. d593-d607, 2002. POBBE, Roger LH et al. Oxytocin receptor knockout mice display deficits in the expression of autism-related behaviors. Hormones and behavior, v. 61, n. 3, p. 436-444, 2012. PRIZANT, Barry M. Brief report: communication, language, social, and emotional development. Journal of Autism and Developmental Disorders, v. 26, n. 2, p. 173-78, 1996. PRIZANT, Barry M.; WETHERBY, Amy M. Communication in preschool autistic children. In: Preschool issues in autism. Springer, Boston, MA, p. 95-128. 1993 PTAK, Carolyn; PETRONIS, Arturas. Epigenetic approaches to psychiatric disorders. Dialogues in clinical neuroscience, v. 12, n. 1, p. 25, 2010. PU, Hongjian et al. Omega-3 polyunsaturated fatty acid supplementation improves neurologic recovery and attenuates white matter injury after experimental traumatic brain injury. Journal of Cerebral Blood Flow & Metabolism, v. 33, n. 9, p. 1474-1484, 2013.

132
PUSKÁS, L.G.; Kitaika, K.; Nyakas, C.; Barcelo-Coblun, G.; Farkas, T. Short-tem administration of omega-3 fatty acids from fish oil results in increased transthyretin transcription in old rat hippocampus. Proc. Natl. Acad. Sci. USA, v. 100, n. 4, p. 1580-1585, 2003. RASALAM, A. D. et al. Characteristics of fetal anticonvulsant syndrome associated autistic disorder. Developmental medicine and child neurology, v. 47, n. 8, p. 551-555, 2005. RAUH, Virginia A. et al. Impact of prenatal chlorpyrifos exposure on neurodevelopment in the first 3 years of life among inner-city children. Pediatrics, v. 118, n. 6, p. e1845-e1859. Relevance for susceptibility and resilience in autism, Psychoneuroendocrinology, 2006 RIBEIRO, B. Estudo dos efeitos neuroprotetores do ômega-3, liraglutide, risperidona e clozapina em um modelo neuroinflamatório induzido por poly i:c em cultura primária de células hipocampais. Tese Doutorado em Farmacologia – Faculdade de Medicina, Universidade Federal do Ceará. Fortaleza, p.20, 2015. RICHARDSON, Alexandra J. Omega-3 fatty acids in ADHD and related neurodevelopmental disorders. International review of psychiatry, v. 18, n. 2, p. 155-172, 2006. RICHARDSON, JENNELLE DURNETT; VASKO, MICHAEL R. Cellular mechanisms of neurogenic inflammation.Journal of Pharmacology and Experimental Therapeutics, v. 302, n. 3, p. 839-845, 2002. RITVO, Edward. Autism--diagnosis, current research, and management. SP Books Division of Spectrum Publications: distributed by Halsted Press, 1976. RODENHISER, David; MANN, Mellissa. Epigenetics and human disease: translating basic biology into clinical applications. Cmaj, v. 174, n. 3, p. 341-348, 2006. RODIER, Patricia M. et al. Embryological origin for autism: developmental anomalies of the cranial nerve motor nuclei. Journal of Comparative Neurology, v. 370, n. 2, p. 247-261, 1996. ROGERS, Sally J.; HEPBURN, Susan; WEHNER, Elizabeth. Parent reports of sensory symptoms in toddlers with autism and those with other developmental disorders. Journal of autism and developmental disorders, v. 33, n. 6, p. 631-642, 2003. RONALD, Angelica; HAPPÉ, Francesca; PLOMIN, Robert. The genetic relationship between individual differences in social and nonsocial behaviours characteristic of autism. Developmental science, v. 8, n. 5, p. 444-458, 2005. RONALD, Angelica; HOEKSTRA, Rosa A. Autism spectrum disorders and autistic traits: a decade of new twin studies. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, v. 156, n. 3, p. 255-274, 2011. RONEMUS, Michael et al. The role of de novo mutations in the genetics of autism spectrum disorders. Nature Reviews Genetics, v. 15, n. 2, p. 133-141, 2014.

133
ROSENBERG, Rebecca E. et al. Characteristics and concordance of autism spectrum disorders among 277 twin pairs. Archives of pediatrics & adolescent medicine, v. 163, n. 10, p. 907-914, 2009. RUSSO, Gian Luigi. Dietary n− 6 and n− 3 polyunsaturated fatty acids: from biochemistry to clinical implications in cardiovascular prevention. Biochemical pharmacology, v. 77, n. 6, p. 937-946, 2009. RUTTER, Michael. Children in substitute care: Some conceptual considerations and research implications. Children and Youth Services Review, v. 22, n. 9-10, p. 685-703, 2000. SAJDEL-SULKOWSKA, Elizabeth M.; ZABIELSKI, Romuald. Gut microbiome and brain-gut axis in autism—aberrant development of gut-brain communication and reward circuitry. In: Recent Advances in Autism Spectrum Disorders-Volume I. IntechOpen, 2013. SANDERS, S. J. et al. De novo mutations revealed by whole-exome sequencing are strongly associated with autism. Nature, v. 485, n. 7397, p. 237-241, 2012. SANDIN, Sven et al. The familial risk of autism. Jama, v. 311, n. 17, p. 1770-1777, 2014. SATTERSTROM, F. K.; Jack A. KOSMICKI, J.A.; WANG, J.; BREEN, M. S. et al. Large-Scale Exome Sequencing Study Implicates Both Developmental and Functional Changes in the Neurobiology of Autism, Cell, Volume 180, Issue 3, 2020, Pages 568-584.e23, ISSN 0092-8674. SCHAPIRO, Shawn; SALAS, Manuel; VUKOVICH, Katherine. Hormonal effects on ontogeny of swimming ability in the rat: assessment of central nervous system development. Science, v. 168, n. 3927, p. 147-151, 1970. SCHENDEL, Diana; BHASIN, Tanya Karapurkar. Birth weight and gestational age characteristics of children with autism, including a comparison with other developmental disabilities. Pediatrics, v. 121, n. 6, p. 1155-1164, 2008. SCHIEVE, Laura A. et al. Parent-reported prevalence of autism spectrum disorders in US-born children: an assessment of changes within birth cohorts from the 2003 to the 2007 National Survey of Children’s Health. Maternal and child health journal, v. 16, n. 1, p. 151-157, 2012. SCHMITZ, Gerd; ECKER, Josef. The opposing effects of n− 3 and n− 6 fatty acids. Progress in lipid research, v. 47, n. 2, p. 147-155, 2008. SCHNEIDER, Tomasz et al. Gender-specific behavioral and immunological alterations in an animal model of autism induced by prenatal exposure to valproic acid. Psychoneuroendocrinology, v. 33, n. 6, p. 728-740, 2008. SCHNEIDER, Tomasz; PRZEWŁOCKI, Ryszard. Behavioral alterations in rats prenatally exposed to valproic acid: animal model of autism. Neuropsychopharmacology, v. 30, n. 1, p. 80-89, 2005.

134
SCHNEIDER, Tomasz; TURCZAK, Joanna; PRZEWŁOCKI, Ryszard. Environmental enrichment reverses behavioral alterations in rats prenatally exposed to valproic acid: issues for a therapeutic approach in autism. Neuropsychopharmacology, v. 31, n. 1, p. 36-46, 2006. SCHRECK, Kimberly A.; MULICK, James A.; SMITH, Angela F. Sleep problems as possible predictors of intensified symptoms of autism. Research in developmental disabilities, v. 25, n. 1, p. 57-66, 2004. SCHUCHARDT, Jan Philipp et al. Significance of long-chain polyunsaturated fatty acids (PUFAs) for the development and behaviour of children. European journal of pediatrics, v. 169, n. 2, p. 149-164, 2010. SCHWARZ, Emanuel et al. Sex-specific serum biomarker patterns in adults with Asperger's syndrome. Molecular psychiatry, v. 16, n. 12, p. 1213-1220, 2011. SCHWARZ, Jaclyn M.; BILBO, Staci D. Sex, glia, and development: interactions in health and disease. Hormones and behavior, v. 62, n. 3, p. 243-253, 2012. SEGAL-GAVISH, Hadar et al. Mesenchymal stem cell transplantation promotes neurogenesis and ameliorates autism related behaviors in BTBR mice. Autism Research, v. 9, n. 1, p. 17-32, 2016. SEIBENHENER, Michael L.; WOOTEN, Michael C. Use of the open field maze to measure locomotor and anxiety-like behavior in mice. JoVE (Journal of Visualized Experiments), n. 96, p. e52434, 2015. SEMPLE, Bridgette D. et al. Brain development in rodents and humans: Identifying benchmarks of maturation and vulnerability to injury across species. Progress in neurobiology, v. 106, p. 1-16, 2013. SERRA-MAJEM, Lluis et al. Dietary methods and biomarkers of omega 3 fatty acids: a systematic review. British journal of nutrition, v. 107, n. S2, p. S64-S76, 2012. SFARI, Gene Website, http://gene.sfari.org/ (2019) SHAMI, Najua Juma Ismail Esh; MOREIRA, Emília Addison Machado. Licopeno como agente antioxidante. Revista de Nutrição, v. 17, n. 2, p. 227-236, 2004. SHARMA, S. R.; GONDA, Xenia; TARAZI, Frank I. Autism spectrum disorder: classification, diagnosis and therapy. Pharmacology & therapeutics, v. 190, p. 91-104, 2018. SHASTRI, A.; BONIFATI, D. M.; KISHORE, U. Innate immunity and neuroinflammation. Mediators of inflammation, v. 2013, 2013. SHCHEGLOVITOV, A. O. et al., SHANK3 and IGF1 restore synaptic de cits in neurons from 22q13 deletion syndrome patients. Nature, vol. 503, no. 7475, pp. 267– 271, 2013. SHEN, Tianjin et al. CHD2 is required for embryonic neurogenesis in the developing cerebral cortex. Stem cells, v. 33, n. 6, p. 1794-1806, 2015.

135
SHRIVER, Eunice Kennedy. Autism overview: What we know. National Institute of Child Health and Human Development, v. 5, p. 1-16, 2005. SILVA, Micheline; MULICK, James A. Diagnosticando el trastorno autista: aspectos fundamentales y consideraciones prácticas. Psicologia: ciência e profissão, v. 29, n. 1, p. 116-131, 2009. SILVERMAN, J. L. et al. Behavioural phenotyping assays for mouse models of autism. Nature Reviews Neuroscience, v. 11, n. 7, p. 490-502, 2010. SIMOPOULOS, Artemis P. Importance of the ratio of omega-6/omega-3 essential fatty acids: evolutionary aspects. In: Omega-6/omega-3 essential fatty acid ratio: The scientific evidence. Karger Publishers, 2003. p. 1-22. SIMOPOULOS, Artemis P. Omega-3 fatty acids in inflammation and autoimmune diseases. Journal of the American College of nutrition, v. 21, n. 6, p. 495-505, 2002. SIMOPOULOS, Artemis P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomedicine & pharmacotherapy, v. 56, n. 8, p. 365-379, 2002. SINGH, Nirbhay N. et al. Mindful parenting decreases aggression, noncompliance, and self-injury in children with autism. Journal of Emotional and Behavioral Disorders, v. 14, n. 3, p. 169-177, 2006. SIQUEIRA, Carolina Carvalho et al. 221 O CÉREBRO AUTISTA: A BIOLOGIA DA MENTE E SUA IMPLICAÇÃO NO COMPROMETIMENTO SOCIAL. Revista Transformar, v. 8, n. 8, p. 221-237, 2016. SMALLEY, Susan L. Autism and tuberous sclerosis. Journal of autism and developmental disorders, v. 28, n. 5, p. 407-414, 1998. SOLÍS-AÑEZ, Ernesto; DELGADO-LUENGO, Wilmer; HERNÁNDEZ, María Luisa. Autismo, cromosoma 15 y la hipótesis de disfunción GABAérgica. Revisión. Investigación Clínica, v. 48, n. 4, p. 529-541, 2007. SONG, Cai et al. The role of omega-3 polyunsaturated fatty acids eicosapentaenoic and docosahexaenoic acids in the treatment of major depression and Alzheimer's disease: Acting separately or synergistically? Progress in lipid research, v. 62, p. 41-54, 2016. SØRENSEN, Merete Juul et al. Antidepressant exposure in pregnancy and risk of autism spectrum disorders. Clinical epidemiology, v. 5, p. 449, 2013. SPAHIS, Schohraya et al. Lipid profile, fatty acid composition and pro-and anti-oxidant status in pediatric patients with attention-deficit/hyperactivity disorder. Prostaglandins, leukotrienes and essential fatty acids, v. 79, n. 1-2, p. 47-53, 2008. SPENCER, Jeanne P.; PAWLOWSKI, Ruth H. Trondsen; THOMAS, Stephanie. Vaccine adverse events: separating myth from reality. American family physician, v. 95, n. 12, p. 786-794, 2017.

136
STANTON, Mark E. et al. Discrimination learning and reversal of the conditioned eyeblink reflex in a rodent model of autism. Behavioural brain research, v. 176, n. 1, p. 133-140, 2007. STEFFENBURG, Suzanne et al. A twin study of autism in Denmark, Finland, Iceland, Norway and Sweden. Journal of Child Psychology and Psychiatry, v. 30, n. 3, p. 405-416, 1989. STRANG, John F. et al. Depression and anxiety symptoms in children and adolescents with autism spectrum disorders without intellectual disability. Research in Autism Spectrum Disorders, v. 6, n. 1, p. 406-412, 2012. STRÖMLAND, Kerstin; PHILIPSON, Eva; ANDERSSON GRÖNLUND, Marita. Offspring of male and female parents with thalidomide embryopathy: birth defects and functional anomalies. Teratology, v. 66, n. 3, p. 115-121, 2002. STUBBS, G.; HENLEY, K.; GREEN, J. Autism: will vitamin D supplementation during pregnancy and early childhood reduce the recurrence rate of autism in newborn siblings? Medical hypotheses, v. 88, p. 74-78, 2016. STUDY, D. D. D. et al. Deciphering Developmental Disorders et al. Prevalence and architecture of de novo mutations in developmental disorders. Nature, v. 542, n. 7642, p. 433-438, 2017. STUEHR, Dennis J.; MARLETTA, Michael A. Mammalian nitrate biosynthesis: mouse macrophages produce nitrite and nitrate in response to Escherichia coli lipopolysaccharide. Proceedings of the National Academy of Sciences, v. 82, n. 22, p. 7738-7742, 1985. SULEWSKA, Anetta et al. DNA methylation in states of cell physiology and pathology. Folia histochemica et cytobiologica, v. 45, n. 3, p. 149-158, 2007. SUPEKAR, Kaustubh; MENON, Vinod. Sex differences in structural organization of motor systems and their dissociable links with repetitive/restricted behaviors in children with autism. Molecular Autism, v. 6, n. 1, p. 50, 2015. SURETTE, Marc E. The science behind dietary omega-3 fatty acids. Cmaj, v. 178, n. 2, p. 177-180, 2008. SWEETEN, Thayne L. et al. High nitric oxide production in autistic disorder: a possible role for interferon-γ. Biological Psychiatry, v. 55, n. 4, p. 434-437, 2004. TAHERI, A.; PERRY, A. Exploring the proposed DSM-5 criteria in a clinical sample. Journal of Autism and Developmental Disorders, v. 42, n. 9, p. 1810–7, set. 2012. TAMANAHA, Ana Carina; PERISSINOTO, Jacy; CHIARI, Brasilia Maria. Uma breve revisão histórica sobre a construção dos conceitos do Autismo Infantil e da síndrome de Asperger. Revista da Sociedade Brasileira de Fonoaudiologia, v. 13, n. 3, p. 296-299, 2008. TANG, Mimi et al. Maternal diet of polyunsaturated fatty acid altered the cell proliferation in the dentate gyrus of hippocampus and influenced glutamatergic and serotoninergic systems of neonatal female rats. Lipids in health and disease, v. 15, n. 1, p. 71, 2016.

137
TANIAI, Hiroko et al. Genetic influences on the broad spectrum of autism: Study of proband-ascertained twins. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics, v. 147, n. 6, p. 844-849, 2008. TASSONI, Danni et al. The role of eicosanoids in the brain. Asia Pacific journal of clinical nutrition, v. 17, n. S1, p. 220-228, 2008. TATTON-BROWN, Katrina et al. Mutations in the DNA methyltransferase gene DNMT3A cause an overgrowth syndrome with intellectual disability. Nature genetics, v. 46, n. 4, p. 385-388, 2014. TAYLOR, Kristen H. et al. Ultradeep bisulfite sequencing analysis of DNA methylation patterns in multiple gene promoters by 454 sequencing. Cancer research, v. 67, n. 18, p. 8511-8518, 2007. TESEI, Alessandra et al. The potential relevance of docosahexaenoic acid and eicosapentaenoic acid to the etiopathogenesis of childhood neuropsychiatric disorders. European child & adolescent psychiatry, v. 26, n. 9, p. 1011-1030, 2017. TETREAULT, Nicole A. et al. Microglia in the cerebral cortex in autism. Journal of autism and developmental disorders, v. 42, n. 12, p. 2569-2584, 2012. THEOHARIDES, Theoharis C.; ASADI, Shahrzad; PATEL, Arti B. Focal brain inflammation and autism. Journal of neuroinflammation, v. 10, n. 1, p. 815, 2013. TONG, Vincent et al. The effect of valproic acid on hepatic and plasma levels of 15-F2t-isoprostane in rats. Free Radical Biology and Medicine, v. 34, n. 11, p. 1435-1446, 2003. TORDJMAN, S. et al. Gene× Environment interactions in autism spectrum disorders: Role of epigenetic mechanisms. Frontiers in psychiatry, v. 5, 2014. TRAN, L. V. et al. Effect of omega-3 and omega-6 polyunsaturated fatty acid enriched diet on plasma IGF-1 and testosterone concentration, puberty and semen quality in male buffalo. Animal reproduction science, v. 173, p. 63-72, 2016. TSAI, Peter T. et al. Autistic-like behaviour and cerebellar dysfunction in Purkinje cell Tsc1 mutant mice. Nature, v. 488, n. 7413, p. 647-651, 2012. TURNER, Tychele N. et al. Loss of δ-catenin function in severe autism. Nature, v. 520, n. 7545, p. 51-56, 2015. VALVO, Giulia et al. Somatic overgrowth predisposes to seizures in autism spectrum disorders. PLoS One, v. 8, n. 9, 2013. VAN EIJK, Kristel R. Quantitative studies of DNA methylation and gene expression in neuropsychiatric traits. 2014. Tese de Doutorado. Utrecht University.

138
VAN ELST, Kim et al. Food for thought: dietary changes in essential fatty acid ratios and the increase in autism spectrum disorders. Neuroscience & Biobehavioral Reviews, v. 45, p. 369-378, 2014. VAN WIMERSMA GREIDANUS, Tj B. Disturbed behavior and memory of the Brattleboro rat. Annals of the New York Academy of Sciences, v. 394, n. 1, p. 655-662, 1982. VARGAS, Diana L. et al. Erratum: Neuroglial activation and neuroinflammation in the brain of patients with autism (Annals of Neurology (2005) 57 (67-81)). Annals of Neurology, v. 57, n. 2, p. 304, 2005. VASCONCELOS, C. P. P. Pré-condicionamento e tratamento com misturas de óleos de ômega-3, 6 e 9 sobre a inflamação e a reabsorção óssea na doença periodontal experimental. 2013 77 f. Dissertação (Mestrado em Ciências Médico-Cirúrgicas), Faculdade de Medicina, Universidade Federal do Ceará, Fortaleza, 2013. VELOSO, André Wilson Nazareth. Curso temporal e distribuição regional da autolimpeza de ratos em resposta a diferentes estressores ambientais. 2012. Tese de Doutorado. Dissertação apresentada como requisito para o título de mestre em análise do comportamento pela Universidade Estadual de Londrina. VERHOEFF, Berend. The autism puzzle: challenging a mechanistic model on conceptual and historical grounds. Philosophy, Ethics, and Humanities in Medicine, v. 8, n. 1, p. 17, 2013. VERROTTI, Alberto et al. Increased oxidative stress in epileptic children treated with valproic acid. Epilepsy research, v. 78, n. 2-3, p. 171-177, 2008. VIOLANTE, Inês Ribeiro. The neurobiological basis of Neurofibromatosis type I: new insights into brain structure, function and neurochemistry. 2012. Tese de Doutorado. VOINEAGU, Irina et al. Transcriptomic analysis of autistic brain reveals convergent molecular pathology. Nature, v. 474, n. 7351, p. 380-384, 2011. VOLKMAR, Fred R.; MCPARTLAND, James C. From Kanner to DSM-5: autism as an evolving diagnostic concept. Annual review of clinical psychology, v. 10, p. 193-212, 2014. VOLKMAR, Fred R.; NELSON, Douglas S. Seizure disorders in autism. Journal of the American Academy of Child & Adolescent Psychiatry, v. 29, n. 1, p. 127-129, 1990. VORHEES, C. V. Behavioral teratogenicity of valproic acid: selective effects on behavior after prenatal exposure to rats. Psychopharmacology, v. 92, n. 2, p. 173-179, 1987. VORSTMAN, J. A. S. et al. Identification of novel autism candidate regions through analysis of reported cytogenetic abnormalities associated with autism. Molecular psychiatry, v. 11, n. 1, p. 18-28, 2006. VRABLIK, Tracy L.; WATTS, Jennifer L. Polyunsaturated fatty acid derived signaling in reproduction and development: insights from Caenorhabditis elegans and Drosophila melanogaster. Molecular reproduction and development, v. 80, n. 4, p. 244-259, 2013.

139
WAGGONER, Darrel. Mechanisms of disease: epigenesis. In: Seminars in pediatric neurology. WB Saunders, 2007. p. 7-14. WAGNER, George C. et al. A new neurobehavioral model of autism in mice: pre-and postnatal exposure to sodium valproate. Journal of autism and developmental disorders, v. 36, n. 6, p. 779-793, 2006. WANG, Chih-Yen et al. Postnatal stress induced by injection with valproate leads to developing emotional disorders along with molecular and cellular changes in the hippocampus and amygdala. Molecular neurobiology, v. 53, n. 10, p. 6774-6785, 2016. WASSINK, T. H. et al. Examination of AVPR1a as an autism susceptibility gene. Molecular psychiatry, v. 9, n. 10, p. 968-972, 2004. WATERLAND, Robert A. et al. Post-weaning diet affects genomic imprinting at the insulin-like growth factor 2 (Igf2) locus. Human molecular genetics, v. 15, n. 5, p. 705-716, 2006. WEAVER, Ian CG et al. Epigenetic programming by maternal behavior. Nature neuroscience, v. 7, n. 8, p. 847-854, 2004. WHELAN, Jay. Antagonistic effects of dietary arachidonic acid and n-3 polyunsaturated fatty acids. The Journal of nutrition, v. 126, n. suppl_4, p. 1086S-1091S, 1996. WHITMAN, Thomas L. O desenvolvimento do autismo. M. Books Editora, 2019. WILLIAMS, Gail et al. Fetal valproate syndrome and autism: additional evidence of an association. Developmental medicine and child neurology, v. 43, n. 3, p. 202-206, 2001. WILLSEY, A. Jeremy et al. Coexpression networks implicate human midfetal deep cortical projection neurons in the pathogenesis of autism. Cell, v. 155, n. 5, p. 997-1007, 2013. WITTE, A. Veronica et al. Long-chain omega-3 fatty acids improve brain function and structure in older adults. Cerebral cortex, v. 24, n. 11, p. 3059-3068, 2014. WOLFERS, Thomas et al. From estimating activation locality to predicting disorder: a review of pattern recognition for neuroimaging-based psychiatric diagnostics. Neuroscience & Biobehavioral Reviews, v. 57, p. 328-349, 2015. WON, Hyejung; MAH, Won; KIM, Eunjoon. Autism spectrum disorder causes, mechanisms, and treatments: focus on neuronal synapses. Frontiers in molecular neuroscience, v. 6, p. 19, 2013. WONG, Christine; CRAWFORD, Dorota Anna. Lipid signalling in the pathology of autism spectrum disorders. Comprehensive guide to autism, v. 18, p. 1259-1283, 2014. WYSS-CORAY, T.; MUCKE, L. Inflammation in neurodegenerative disease a double-edged sword. Neuron, v. 35, n. 3, p. 419-432, 2002.

140
XU, H. et al. Effects of different dietary DHA: EPA ratios on gonadal steroidogenesis in the marine teleost, tongue sole (Cynoglossus semilaevis). British Journal of Nutrition, v. 118, n. 3, p. 179-188, 2017. YADAV, Sneha et al. Comparative efficacy of alpha-linolenic acid and gamma-linolenic acid to attenuate valproic acid-induced autism-like features. Journal of physiology and biochemistry, v. 73, n. 2, p. 187-198, 2017. YAMASHITA, Yushiro et al. Possible association between congenital cytomegalovirus infection and autistic disorder. Journal of autism and developmental disorders, v. 33, n. 4, p. 455-459, 2003. YEHUDA, S.; RABINOVITZ, S.; MOSTOFSKY, D. I. Essential fatty acids and the brain: from infancy to aging. Neurobiology of aging, v. 26, n. 1, p. 98-102, 2005. YOCHUM, C. L. et al. Animal model of autism using GSTM1 knockout mice and early post-natal sodium valproate treatment. Behavioural brain research, v. 210, n. 2, p. 202-210, 2010. YOCHUM, Carrie L. et al. VPA-induced apoptosis and behavioral deficits in neonatal mice. Brain research, v. 1203, p. 126-132, 2008. YONG, C.; Gean, P. W.; Chiou, L.C.; shen, Y.Z. Docosahexacnoic acid inhibit synaptic transmission and epileptiform activity in the rat hippocampus. Synapse, v.37, n. 2, p.90-94, 2000. YUEN, Ryan KC et al. Whole genome sequencing resource identifies 18 new candidate genes for autism spectrum disorder. Nature neuroscience, v. 20, n. 4, p. 602, 2017. ZABLOTSKY, B.; BLACK, L. I.; BLUMBERG, S. J. Estimated prevalence of children with diagnosed developmental disabilities in the United States, 2014-2016. US Department of Health and Human Services, Centers for Disease Control and Prevention, National Center for Health Statistics, 2017. ZEISEL, Steven H. Regulation of" nutraceuticals". v. 285, p. 1853-55,1999. ZHAN, Yang et al. Deficient neuron-microglia signaling results in impaired functional brain connectivity and social behavior. Nature neuroscience, v. 17, n. 3, p. 400-406, 2014. ZHOU, Jian et al. Whole-genome deep-learning analysis identifies contribution of noncoding mutations to autism risk. Nature genetics, v. 51, n. 6, p. 973-980, 2019. ZOROGLU, S. Salih et al. Increased oxidative stress and altered activities of erythrocyte free radical scavenging enzymes in autism. European archives of psychiatry and clinical neuroscience, v. 254, n. 3, p. 143-147, 2004.
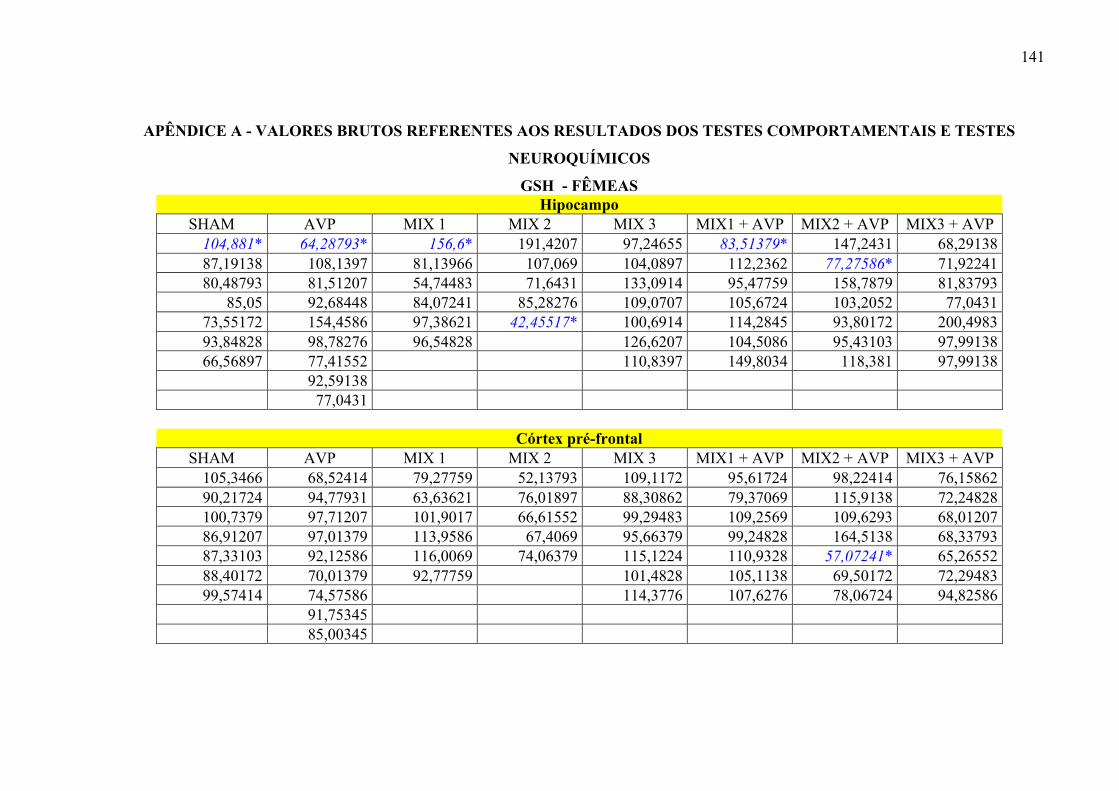
141
APÊNDICE A - VALORES BRUTOS REFERENTES AOS RESULTADOS DOS TESTES COMPORTAMENTAIS E TESTES
NEUROQUÍMICOS
GSH - FÊMEAS Hipocampo
SHAM AVP MIX 1 MIX 2 MIX 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP 104,881* 64,28793* 156,6* 191,4207 97,24655 83,51379* 147,2431 68,29138 87,19138 108,1397 81,13966 107,069 104,0897 112,2362 77,27586* 71,92241 80,48793 81,51207 54,74483 71,6431 133,0914 95,47759 158,7879 81,83793
85,05 92,68448 84,07241 85,28276 109,0707 105,6724 103,2052 77,0431 73,55172 154,4586 97,38621 42,45517* 100,6914 114,2845 93,80172 200,4983 93,84828 98,78276 96,54828 126,6207 104,5086 95,43103 97,99138 66,56897 77,41552 110,8397 149,8034 118,381 97,99138
92,59138 77,0431
Córtex pré-frontal SHAM AVP MIX 1 MIX 2 MIX 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
105,3466 68,52414 79,27759 52,13793 109,1172 95,61724 98,22414 76,15862 90,21724 94,77931 63,63621 76,01897 88,30862 79,37069 115,9138 72,24828 100,7379 97,71207 101,9017 66,61552 99,29483 109,2569 109,6293 68,01207 86,91207 97,01379 113,9586 67,4069 95,66379 99,24828 164,5138 68,33793 87,33103 92,12586 116,0069 74,06379 115,1224 110,9328 57,07241* 65,26552 88,40172 70,01379 92,77759 101,4828 105,1138 69,50172 72,29483 99,57414 74,57586 114,3776 107,6276 78,06724 94,82586
91,75345 85,00345

142
Amigdalas SHAM AVP MIX 1 MIX 2 MIX 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
80,76724 118,1483 79,88276* 87,37759 99,66724 115,7741 90,68276 97,61897 75,22759 99,52759 109,8155 80,72069 106,7431 112,2828 83,51379 73,78448 74,62241 87,42414 112,8879 77,83448 112,2362 124,8052 68,98966 104,9741 92,73103 119,4052 106,9293 55,53621 94,22069 91,33448 112,3293 88,58793 75,5069 88,40172 100,9707 65,73103 94,03448 103,0655 94,73276 102,0414
113,4931* 86,21379 113,2603 115,6345 117,869 95,43103 84,81724 101,2966 82,35* 72,85345* 117,45 97,71207 92,87069
97,94483 82,86207*
Cerebelo SHAM AVP MIX 1 MIX 2 MIX 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
77,1362 184,6707 166,3293 98,2707 116,1 188,3948* 109,4431 89,9845 105,3 111,3052 121,5 97,3397 122,0121 113,9121 118,3345 104,881
99,3879 109,8155 125,5034 93,6621 116,1 131,7879 92,8241 87,7034 112,4224 116,2397 125,4569 103,1586 127,7845 108,6983 96,4086 106,7431
147,4293* 105,0672 128,7621 92,8707 113,6793 105,6724 106,65 114,8431 88,2155 87,9362 35,4724* 111,2586 116,9845 95,6172 115,3552
175,7328 102,1345 118,2414 117,1707 97,9914 125,131 93,0103 32,1672*
GSH - MACHOS Hipocampo
SHAM AVP MIX 1 MIX 2 MIX 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP

143
83,6069 66,28966 66,84828 28,35 63,26379 58,98103 58,00345 56,00172 69,36207 73,64483 97,15345* 33,42414 73,78448 59,21379 68,9431 60,6569 70,1069 77,22931 76,99655 27,32586 183,4138* 60,93621 89,05345* 54,0931
115,4017* 138,5379* 66,33621 33,05172 30,44483 68,05862 87,19138 82,02414 86,26034* 54,88448 60,6569 28,95517 45,48103 72,52759* 66,66207 46,73793 79,23103 66,10345 66,42931 27,88448 29,7 65,12586 41,52414 42,96724 34,54138 34,30862 34,58793 43,15345 29,04828 30,77069 97,57241* 35,0069 32,53966 43,24655
33,98276 34,68103 29,00172
Córtex pré-frontal SHAM AVP MIX 1 MIX 2 MIX 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
58,93448 48,92586* 68,10517 32,4931 84,58448 48,18103 78,34655 55,58276 61,58793 55,62931 76,67069 70,61897* 46,22586 54,32586 87,19138 67,36034 53,9069 60,70345 66,38276 29,93276 87,00517 56,00172 72,10862 69,59483
65,82414* 59,07414 66,52241 29,00172 50,97414 51,62586 61,40172 51,53276 63,96207 59,07414 59,95862 34,16897 28,07069 49,90345 70,24655 72,52759 58,93448 140,4466 74,06379 33,47069 30,77069 47,66897 41,38448 52,09138 28,67586 33,19138 28,81552 44,50345 31,56207 29,65345 85,79483 30,53793 27,69828 43,33966 28,81552 35,47241
37,52069
Amigdalas SHAM AVP MIX 1 MIX 2 MIX 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
64,10172 56,7 64,42759 47,25 87,6569* 58,51552 64,93966 39,15 68,66379 59,86552 76,71724 30,58448 73,08621 66,28966 58,79483 38,31207

144
58,84138 65,54483 43,15345 56,00172* 64,8 49,90345 70,2 98,68966* 91,28793 66,56897 69,03621 31,37586 29,6069 77,78793* 86,67931* 51,25345 68,38448 67,12759 86,63276* 42,22241 28,90862 60,37759 72,94655 29,23448 64,19483 67,63966 58,42241 47,80862 36,12414 50,88103 37,66034 57,07241 31,42241 41,85 41,98966 84,35172 32,86552 31,84138 67,54655
29,7* 37,8931 123,5483* 34,02931 36,26379
30,49138
Cerebelo SHAM AVP MIX 1 MIX 2 MIX 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
73,59828 63,3569 58,93448 63,3103 81,65172 91,00862 56,7931 56,04828 69,40862 22,8569* 100,0397* 249,0052* 76,62414 74,66897 67,08103 51,11379
139,2828* 51,53276 85,32931 62,4259 66,56897 94,91897 53,25517 71,22414 90,3569 65,31207 51,85862 32,5397 31,6086* 69,36207 81,0931* 51,2069
75,55345 69,78103 61,35517 30,3983 77,1828 62,61207 63,72931 56,65345 62,56552 89,75172 65,21897 33,7966 36,5431 68,9431 31,4224 57,9569 44,7362 32,7724 30,9569 50,92759
31,1431* 28,769 70,85172 32,5862 41,7569 63,68276 47,6224 26,9069
27,7914
NITRITO - FÊMEAS Hipocampo
SHAM AVP MIX 1 MIX 2 MIX 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP 1,049382716 2,329218107 1,374485597 20,7037037* 1,728395062 1,255144033 6,716049383 1,057613169
5,238683128* 2,069958848 1,732510288 0,679012346 1,456790123 1,913580247 19,20164609* 0,888888889

145
1,037037037 2,617283951 2,020576132 0,757201646 1,530864198 1,950617284 1,053497942 2,201646091 1,345679012 1,872427984 2,563786008 0,716049383 1,082304527 2,24691358 1,016460905 5,24691358 1,164609053 1,440329218 0,810699588 1,049382716 1,325102881 3,9218107 8,37037037 1,728395062 0,761316872 2,218106996 1,041152263 1,197530864 1,938271605 7,024691358 3,604938272 6,613168724
0,711934156 5,917695473 11,85185185* 0,790123457 10,6090535*
córtex pré-frontal SHAM AVP MIX 1 MIX 2 MIX 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
1,473251029* 1,843621399 0,888888889 0,839506173 1,349794239 8,172839506 4,316872428 1,913580247 0,971193416 2,687242798 0,95473251 0,724279835 1,839506173 12,2345679 5,975308642 0,880658436 0,757201646 2,057613169 3,300411523* 0,695473251 6 11,40740741 9,522633745 1,316872428 0,769547325 0,975308642 0,823045267 0,769547325 6,580246914 8,823045267 5,827160494 0,991769547 0,773662551 1,156378601 0,888888889 0,765432099 5,555555556 11,83539095 4,802469136 1,395061728 0,909465021 1,288065844 0,864197531 0,773662551 13,16872428 4,349794239 0,794238683* 0,930041152
1,152263374 1,090534979* 0,884773663 1,222222222 4,382716049*
Amigdalas SHAM AVP MIX 1 MIX 2 MIX 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
0,765432099 0,806584362 3,024271845 2,87654321 2,736625514 0,868312757 2,230452675 1,16872428 0,823045267 5,082304527 3,742718447 2,600823045 16,10699588 1,358024691 1,432098765 0,917695473 1,456790123 0,720164609 1,757281553 4,469135802 1,131687243 1,366255144 1,065843621 0,827160494 0,757201646 3,888888889 2,689320388 4,160493827 0,9218107 4,259259259 2,588477366 1,604938272 0,83127572 2,699588477 2,640776699 3,720164609 6,382716049 3,823045267 10,43621399* 1,304526749
1,106995885 0,810699588 2,019417476 6,209876543 14,70781893 4,70781893 1,465020576 2,950617284 2,181069959 1,127572016 10,4526749* 1,205761317 2,823045267
Cerebelo

146
SHAM AVP MIX 1 MIX 2 MIX 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP 2,362139918 2,905349794 5,325242718 4,650205761 6,386831276 11,8436214 3,020576132 4,127572016
4,691358025* 3,378600823 2,033980583 5,699588477 8,732510288 8,04526749 0,868312757 4,62962963 2,106995885 1,226337449 5,655339806 6,50617284 10,45679012 9,288065844 3,181069959 5,152263374 1,469135802 4,131687243 2,029126214 9,847736626* 19,13168724 14,55144033 1,827160494 4,201646091 2,279835391 1,283950617 2,281553398 6,037037037 14,10288066 16,30452675 1,695473251 4,205761317 1,670781893 1,0781893 1,839805825 5,534979424 9,670781893 8,024691358 2,732510288 2,481481481
1,930041152 1,78600823 1,131687243 1,654320988
NITRITO - MACHOS Hipocampo
SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP 0,880658436 1,251028807 2,057613169 3,004115226 1,304526749 1,004115226 2,205761317 2,325102881 0,798353909 1,16872428 2,913580247 2,979423868 1,683127572 0,757201646 1,884773663 2,452674897 1,127572016 0,757201646* 1,329218107 1,893004115 6,160493827* 1,29218107 2,094650206 2,547325103 1,29218107 0,880658436* 2,004115226 2,893004115 3,555555556 1,04526749 2,481481481 2,666666667
9,893004115* 1,004115226 1,156378601 2,724279835 3,58436214 1,004115226 2,230452675 4,0781893 1,625514403 2,703703704 4,851851852
córtex pré-frontal SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
0,757201646 1,78600823 3,502057613 4,518518519 5,181069959 0,880658436 2,144032922 1,983539095 1,004115226 1,251028807 6,995884774* 4,205761317 2,967078189 0,633744856 3,279835391 1,473251029 0,880658436 1,333333333 1,641975309 6,530864198* 2,967078189 0,9218107 2,942386831 2,238683128 1,127572016 1,456790123 2,827160494 3,222222222 3,78600823 0,839506173 1,444444444 3,625514403 1,16872428 0,757201646* 8,917695473* 1,341563786 2,814814815 0,757201646 1,316872428 1,740740741

147
1,04526749 2,279835391 2,518518519 1,275720165 0,716049383 2,732510288 0,333333333 4,094650206*
Amigdalas SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
1,127572016 1,16872428 5,097087379 3,333333333 3,201646091 0,880658436 2,255144033 4,329218107 1,04526749 0,757201646 3,990291262 1,950617284 4,95473251 2,979423868 1,362139918 1,279835391
1,004115226 1,086419753 5,004854369 4,395061728 2,864197531 1,04526749 1,164609053 3,004115226 0,757201646 0,962962963 2,218446602 3,333333333 2,703703704 0,839506173 1,839506173 11,19341564* 1,16872428 1,04526749 9,86407767* 1,275720165 4,193415638 1,04526749 1,555555556 1,238683128
2,358024691 2,316872428 2,650205761
Cerebelo SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
19,02880658* 5,489711934 5,019417476 3,296296296 3,086419753 6,271604938* 1,724279835 5,481481481* -1,028806584* 8,12345679 5,427184466 2,950617284 4,58436214 0,962962963 1,950617284 3,395061728
1,580246914 3,308641975 4,54368932 9,156378601* 3,028806584 0,9218107 3,880658436 3,650205761 1,086419753 0,839506173* 4,009708738 2,855967078 3,127572016 0,9218107 6,70781893* 1,991769547 0,798353909 1,04526749* 4,640776699 3,736625514 2,139917695 0,962962963 2,724279835 2,50617284 1,868312757 4,378600823 3,485436893 3,106995885 1,41563786 4,407407407*
1,958847737 2,115226337
TBARS - FÊMEAS Hipocampo
SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP

148
1056 792 1571 839 790 1096 998 1085 1047 1237 1180 1005 3034* 681 1589 1125 1053 808 1666 933 748 729 805 885 1122 1424 1540 1272 888 727 1845 1100 802 1008 1396 844 717 778 1117 1180
1501 1335 1565 1372 893 790 923 888 1649 843 979 1049
Córtex pré-frontal SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
992 1179 1326 844 1164 874 680 1601 1677 1731 1454 1372 892 787 656 844 1161 1397 1301 900 957 801 768 880 1001 1059 2007 892 913 770 614 1161 1539 1047 1796 1221 1032 788 717 1018 1751 1414 1911 754 856 727 867 1116
1477 1094 620 1221
Amigdalas SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
992 1179 1326 1846 790 1167 1256 1271 1677 1731 1454 2108 753 889 1021 1031 1161 1397 1301 1552 926 626 673 1301 1001 1059 2007 1911 1028 724 696 915 1539 1047 1796 1744 1069 713 214 825 1751 1414 1911 2283 791 645 647 1036
1477 -256 582 911 912

149
Cerebelo
SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP 1770 1455 1310 1310 1014 634 469 114 1486 1173 1696 1696 713 574 465 127 1451 1281 1578 1578 842 633 679 117 1227 1316 873 873 526 607 789 110 986 1413 1466 1466 1044 1670 1266 116
1340 1092 1441 1441 764 786 1210 126 1227 1079 143 1267 150
TBARS - MACHOS Hipocampo
SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP 831 1196 1042 836 951 1811 881 671 840 -135* 850 856 1090 1446 824 738
2541* 510* 1075 814 1002 1802 1002 875 982 795 796 956 1058 368* 808 637
-125* 2755 778 877 1124 386* 800 956 457 1205 778 968 1846 664
694 666
Córtex pré-frontal SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
2354* 875 988 988 1443 1846 846 647 403 795 1245 1245 1292 742 930 392 573 973 956 956 1086 288* 872 549

150
769 1748 1038 1038 964 261* 835 658 1499* 2399 524* 524* 976 1650 427 629
314 4340* 1124 1124 1490
Amigdalas SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
1143 911 1445* 742 823 831 1185* 1020 2176* 1303 704 821 712 1597 640 343
466 519* 802 835 899 920 833 626 920 1214 427 812 612 2140 818 1213*
1784 1757 2129* 455 605 2906* 862 581 608 795* 645 283* 1909 371
Cerebelo
SHAM AVP MIX 1 MIX 2 MIX 3 MIX1 MIX2 MIX3 350* 778 1087 1001 1347 546 1155 824
2532* 1232 1345 1592 1407 573 754 814 2042 822 1551* 1159 1517 1063 1154 857 1463 261* 1035 1250 1136 1276 909 954 2069 3948* 1092 1605 1109 973 1226 889 546* 1517 1433 1141 1757 1014
TESTE CAMPO ABERTO - FÊMEAS Tempo parado
SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP 107 66 84 139 20 82 132 32 131 71 113 72 8 125 152 15 128 48 111 85 12 128 188 21 137 67 91 79 15 80 107 5

151
152 74 86 92 12 68 85 6 152 53 90 77 14 139 65 6 77 58 76 53 78 53 70 48 80 58 83 33 78 41 34 71 77 107 92 45 97 50 51 44 80 85 48 106 45 142 58 47 65 121 80 55 34 40
125 70 98 101 55 33 56 144 72 158 121 46 32
91 131 74 99 74 68
Cruzamentos SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
37 39 27 29 38 59 23 54 22 33 28 45 38 39 22 72 23 46 50 47 31 38 19 64 34 38 34 42 45 59 46 63 26 36 35 35 45 65 67 44 26 30 19 44 56 44 74 63
45* 37 37 56 31 44 52 54 41 41 25 51 39 43 67 54 39 46 28 57 35 48 49 60 25 67 52 20 46 6* 30 42 74 42 28 44 45 37 31 52 40 24 54 55 40 32 44 39 16 50 62
51 38

152
58 48 44 52
Rearing SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
7 23 8 8 21 26 16 20 13 15 9 14 15 15 14 9* 13 36 30 14 15 14 3 21 11 7 8 28 27 24 3 29 13 7 18 7 29 16 5 23 12 22 9 13 24 17 5 34
22* 16 14 11 10 23 21 26 11 17 8 16 13 14 17 18 5 3* 8 31 5 13 15 26 4 5* 28 4 15 5 33
20* 5* 7 17 13 19 22 6 21 13 7 2* 26 8* 8 22 5 9 12 10 20 4 18 12 24 13
Grooming SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
3 5 4 2 4 6 2 1 2 3 3 3 3 5 2 3

153
2 1 2 4 6 5 1 6 2 2 6 1 2 6 6 3 1 4 5 10* 4 6 2 7 0 2 5 3 4 1 4 4 3 2 4 1 5 3 5 5 4 4 4 4 2 4 3 2 2 6 9 0 2 3 3 3 7 2 3 0 1 2 1 3 4 5 3 2 2 1 1 5 7 1 2 0 2 3 3 2 5 3 4 4 3 3 1 2 3
TESTE CAMPO ABERTO - MACHOS Cruzamentos
SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP 25 39 40 36 27 22 67 46 40 50 41 35 33 30 71 33 64 50 48 43 44 13 81 56 58 24 48 42 48 50 62 62 37 58 31 51 42 74 29 59 54 46 28 42 36 81 46 32 30 43 20 46 27 49 31 29 34 35 40 44 25 41 28 48 33 34 36 73 28 36 24 57

154
33 24 46 34 46 18 32 46 35 41 51 34 21 33 16 52 33 28 50 31 30 18 22 48 53 30 27 24 32 29 41 56 34 24 30 98 61 51 33 36 35 27 45 33 18 30 29 35 37 29 27 33 34 51 47 31 32 28 36 61 27 50 48
Tempo parado
SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP 146* 70 100 118 143 171 51* 92
72 126 116 103 113 124 28* 109 86 113 142 69 135 118 23* 91 84 161 106 69 105 89 90 94 72 107 116 77 140 53* 10* 88
109 99 85 108 94 116 6* 109 68 92 103 51* 135 45* 134 131 42 105 74 82 90 76* 131 112 58 142 82 85 88* 102 129 109 68 115 85 126 77 140 119 105 89 80 87 118 111 90 176 106
118* 68 62 126 101 154 106 90 100 42* 84 145 97 108 107 77*

155
95 58 140 93 90 74 68 142 117 97 98 89 125 104 84 118 102 90
105 106 98 122* 101 100 100 105 46 64 50
Rearing
SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP 7 17 26 6 11 29 23 7*
13 14 28 15 5* 15 44 10 14 32 32 28 13 22 30 42 14 10 16 23 15 18 30 10 5 10 25 24 13 35 13 9 5 15 15 16 10 22 33 33 4 9 20 20 11 10 8* 9
13 12 19 3* 9 6* 6* 10 11 5* 12 13 9 3* 19 2* 11 10 25 12 7* 18 10 8 23 27 9 14 9* 7* 8 10 25 17 9 0 8
15* 4* 8* 5* 21 9 7* 11 13 10 12 10 9* 12
19* 11 29 7 11 0 12 11 8 9 15 8 8* 20 13

156
7 8* 4* 8* 7 12 13 6 12
11 3* 17*
5
Grooming SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
0 3 2 4 4 6 10 3 1 3 3 4 0 4 3 3 1 2 6 3 5 10 3 3 2 4 5 6 2 6 3 2 0 2 3 4 2 11 11 4 1 1 3 3 1 7 8 2 4 1 3 1 2 6 3 1 2 3 1 3 1 0 2 2 1 2 2 0 5 1 2 1 1 3 1 2 7 3 3 2 2 1 5 1 5 3 4 2 5 3 2 4 3 0 1 2 2 4 4 1 2 2 2 2 0 2 6 5 1 1 0 1 2 5 8 2 1 4 5 4 2 2 4 4 5 5 6
9* 3 5 4 7 2

157
3 3
TESTE DE IMERSÃO DE CAUDA - FÊMEAS Imersão de Cauda
SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP 364 602 502 410 364 482 434 2,37* 370 486 420 416 386 486 400 2,75* 408 592 484 448 404 386 380 2,6* 372 520 360 420 402 454 440 362 440 455 392 460 396 444 438 282 364 812 400 452 414 486 440 304 340 452 364 404 402 424 444 312
1212 410 566 462 440 362 280 440 442 512 398 438 360 320 438 612* 460 436 372 260 440 462 482 366 354 364 444 384 414 408 350 382 674 404 440 482 422 1084 444 524 280
TESTE DE IMERSÃO DE CAUDA - MACHOS Imersão de Cauda
SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP 326 514 422 362 362 4,69* 1* 1212* 332 612 492 400 422 1,33* 1,07* 422 364 634 428 368 368 1,34* 1,41* 464

158
400 552 414 446 364 0,72* 1,37* 392 844* 564 410 346 348 1,25* 282 402 330 652 540 428 410 1,41* 374 1288* 340 530 520 304 408 1,78* 368 380 364 540 345 448 370 316 410 484 352 552 348 458 412 376 382 542 342 546 470 1212* 404 326 372 660* 414 496 560 422 424 416 360 420 440 596 494 314 402 528 322 830* 430 482 310 382 666* 416 424 362 544 364 400 680* 374 382 316 460 360 408 308 400 384 284 330 432
344 316 476 408 310 532 320 358 396 350 394
TESTE DE NATAÇÃO - FÊMEAS Natação
SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP 4 4 4 4 4 3 3 3 4 3 4 4 4 3 4 3 4 3 4 4 4 3 3 4 4 3 4 4 4 3 4 3 4 3 4 4 4 3 3 3

159
4 4 4 4 4 4 3 3 4 3 4 4 4 4 3 3 4 3 4 3 4 4 4 3 3 4 4 4 4 4 4 3 3 3 3 4 4 4 3 3 4 4 4 4 3 4 4 4 4 3 4 3 3 3 4 3 3
TESTE DE NATAÇÃO - MACHOS Natação
SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP 4 4 4 4 4 3 3 4 4 3 4 4 4 4 4 4 3 4 4 4 3 4 3 4 3 4 3 4 3 4 3 3 3 3 4 4 3 4 3 4 3 4 4 4 3 4 3 4 3 4 4 4 4 3 4 4 3 4 4 4 4 4 4 4 2 4 4 3 3 3 4 4 3 4 4 4 2 4 3 4 4 4 4 3 3 2 4 4 3 4

160
4 3 3 2 3 4 4 3 3 4 4 3 3 4 2
TESTE ROTAROD - FÊMEAS Tempo 1° Queda
SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP 140 21 174 102 34 54 118 40 89 49 56 47 67 41 145 10
108 117 145 124 30 68 31 46 153 16 120 42 35 117 57 152 39 44 107 60 38 27 28 139
187 50 108 44 51 138 30 43 37 114 116 161 103 87 32 43
149 12 88 64 66 54 103 133 57 87 158 40 84 128 143 139 28 177 126 67 172 54 60 163 30 115 114 42 46 172 105

161
76 32 58 48 37 86 223 47 150 70 102 26 154 40
56 106 47 46 86 54 52 43
RPM 1° Queda SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
21 7 25 16 8 11 13 9 15 10 11 10 12 9 17 6 17 18 21 19 8 13 8 10 22 6 19 9 9 19 11 22 9 9 17 12 9 7 7 21
27 10 17 9 10 21 8 9 8 18 18 24 16 15 8 9
22 6 15 12 12 11 16 20 11 15 23 9 14 19 21 21 7 25 19 12 25 11 11 24 8 18 18 9 10 25 17 13 8 11 10 8 14 31 10 22 12 16 7 22 9
11 17 10 10 14 11 10 9

162
Tempo Final SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
167 108 185 150 100 180 247 110 119* 80 220 160 107 180 324 96 174 133 164 155 97 117 206 89 202 117 182 136 105 172 263 177 158 163 156 150 89 197 248 158 220 103 156 104 75 168 256 103 218 125 240 214 162 266 262 114 215 35 266 191 264 241 180 180 263* 224 221 144 260 276 188 199 248 234 173 92 257 298 172 212 256* 227 256 110 190 289 249 234 262* 247 239 123 222 267 111 243 114 228 87 241 118
160 245 129 97 120 154 116 122
RPM Final SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
24 17 26 22 16 27 24 17 18 14 30 23 17 27 28 16 25 20 24 23 16 19 19 15 28 18 26 20 17 25 36 25 23 24 23 22 15 28 34 23 30 16 23 17 13 24 35 16 30 19 33 30 24 36 36 18

163
30 8* 36 27 36 33 26 26 36 31 31 21 35 37 27 27 34 32 25 16 35 40 25 30 35 31 35 17 27 39 34 32 36 34* 33 19 31 35 17 33 18 31 15 33 18
23 33 19 16 19 22 18 19
Bolos fecais SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
6 0 1 0 0 0 2 0 6 3 3 0 0 6 0 0 6 4 11 0 0 0 0 0 4 0 0 2 4 0 6 5 5 1 6 0 0 0 4 6 3 2 4 0 3 10 5 3 3 4 4 0 3 3 3 4 3 2 3 2 0 7 1 3 6 2 2 3 4 0 0 0 4 5 0 1 7 5 0 0 5 4 0 2 5 3 0 1 3 0 6 1 0 4 1 4 0 0 2 0 3 4 0 1 4 0 3

164
0 0
TESTE ROTAROD - MACHOS Tempo 1° Queda
SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP 12* 93* 70 30 55 265* 101 77 9* 72 59 38 52 74 92 25* 56 45 77 30 65 62 38 150
103 65 67 47 54 150* 73 52 49 36 88 44 52 40 34 46*
10* 120* 58 50 117 104 65 163 39 80 8* 9* 34 40 29 45*
35* 93* 9* 10* 84 26 10* 158 45 86 8* 10* 112 29 45 28
24* 40 15 76 49 27 23* 10 54 150* 8 34 29* 27 90 96 56 96* 12 49 153 77 100 101
158 40 115 64 20* 99 110 114 86 20 98 35 27 17* 44
123 32 57 42 152 61 74 40 86 60 195 48 90 16 82 141 53
96* 47 65 175 50 65 46 36 43 43

165
RPM 1° Queda
SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP 5* 9 13 8 11 15 8 13 5* 8 11 9 10 7 7 7* 11 7 13 9 12 6 5* 22 17 7 12 10 10 10 7 10 10 6 15 10 10 9 9 10 5* 12 11 10 18 16 12 24 8 14 5 6 8 9 7 9 8 15 5 6 14 6 5* 23
10 14 5 6 17 9 9 7* 7 9 6 13 10 7 7 5*
10 22* 5 8 7 7 15 16 11 16* 6 10 23 13 16 16 23 9 17 12 7 16 17 18 14 7 16 8 7 6* 9 19 8 11 9 23 11 13 9 14 11 27 10 15 6 14 21 10
16* 10 12 25 10 12 10 8 9 9
Tempo Final SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
45 225 113 90 146 300 300 105

166
43 241 113 90 143 277 300 70* 90 229 112 97 126 300 300 290
115 230 103 108 100 300 300 100 95 242 121 88 95 90 155 80 44 230 76 108 155 119 133 180
125 140 37 63 132 65 156 71* 97 147 38 35 146 97 211 180 93 142 40 47 188 97 135 95
100 136 40 108 170 88 202 142 89 180 54 127 82 90 316 146
186 135 97 127 174 214 137 180 259 120 189 171 50 165 155 249 101 140 191 227 132 188 220 115 148 190 47* 103 268 164 188 256 201 95 220 168 234 230 110
135 256 122 239 116 242 102 278 91 90 90
RPM Final SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP
10 19 18 15 22 15 15 17 9 28 18 15 21 14 15 13
15 26 18 16 19 15 15 27* 18 27 16 17 16 15 15 16 15 28 19 15 16 15 23 14

167
9 27 13 17 23 18 20 26* 20 9 12 20 12 18 13 22 9 8 22 16 20 26* 20 9 10 25 16 15 15 19 9 17 25 15 19 21 26 10 19 14 15 28 22 20 16 19 25 30 21 26*
19 19 27 25 10 24 23 16 16 21 27 31 20 27* 15 18 22 27 10 16 16 24 27 35 28 16 15 24 32 32 17 26 20 35 19 35 33 18 34 33 16 31 37 16 36 15 31 15
Bolos fecais
SHAM AVP Mix 1 Mix 2 Mix 3 MIX1 + AVP MIX2 + AVP MIX3 + AVP 0 7 7 0 0 3 0 0 4 4 0 0 0 4 6 0 2 3 6 1 0 4 0 8 2 6 2 0 0 1 0 0 6 8 6 0 0 0 6 0 1 6 4 1 0 1 0 0 2 3 0 0 3 2 8 4 3 4 0 0 3 0 6 5 0 2 3 0 4 0 6 6

168
1 3 2 6 5 3 7 4 3 1 1 6 5 0 5 5 5 2 0 3 5 6 6 8 1 4 5 8* 3 4 2 3 2 0 9* 5 3 4 1 4 4 7 2 0 4 7 1 2 3 1 8 2 5 2 2 5 7 0 3 0 6 0 5 0 0 0

169
APÊNDICE B - VALORES BRUTOS REFERENTES AOS RESULTADOS DA
ANÁLISE PCR quantitativa (RT-qPCR)
Valores de Cq por grupo
Cq Dnmt3a Chd2
Média Desvio padrão Média Desvio padrão SHAM 1 2,21 0,25 2,06 0,96 SHAM 2 2,44 0,34 2,11 1,23 SHAM 3 0,25 0,04 0,39 0,01 SHAM 4 0,33 0,01 0,41 0,06 SHAM 5 0,63 0,06 0,51 0,08 SHAM 6 0,22 0,16 0,38 0,04 SHAM 7 0,34 0,06 0,55 0,08
Cq Dnmt3a Chd2
Média Desvio padrão Média Desvio padrão AV 1 1,00 0,06 1,23 0,71 AV 2 0,93 0,02 1,34 0,90 AV 3 0,27 0,06 0,29 0,01 AV 4 0,32 0,01 0,46 0,03 AV 5 0,29 0,04 0,37 0,00
Cq Dnmt3a Chd2
Média Desvio padrão Média Desvio padrão MIX 3_1 1,05 0,06 1,39 1,05 MIX 3_2 1,24 0,02 1,55 0,89 MIX 3_3 0,25 0,00 0,43 0,07 MIX 3_4 0,19 0,02 0,31 0,07 MIX 3_5 0,20 0,01 0,16 0,08 MIX 3_6 0,24 0,06 0,26 0,08

170
Cq
Dnmt3a Chd2 Média Desvio padrão Média Desvio padrão
AV+MIX 3_1 1,25 0,47 1,89 1,58 AV+MIX 3_2 0,92 0,03 1,22 0,66 AV+MIX 3_3 0,17 0,01 0,11 0,13 AV+MIX 3_4 0,21 0,00 0,31 0,04 AV+MIX 3_5 0,16 0,01 0,29 0,08 AV+MIX 3_6 0,39 0,49 0,15 0,18
Médias
Cq Dnmt3a Chd2
Média Desvio padrão Média Desvio padrão SHAM 0,92 0,97 0,91 0,80 AV 0,56 0,37 0,74 0,50 MIX 3 0,53 0,48 0,68 0,62 AV+MIX 3 0,52 0,46 0,66 0,73

171
ANEXO A- PARECER DE APROVAÇÃO DO COMITÊ DE ÉTICA




















