
Taxonomic position and phylogenetic relationships of Colobomycter pholeter, a small reptile from the...
7
Taxonomic position and phylogenetic relationships of Colobomycter pholeter, a small reptile from the Lower Permian of Oklahoma MICHEL LAURIN AND ROBERT R. REISZ Department of Zoology, University of Toronto, Erindale College, 3359 Mississauga Road N, Mississauga, Ont., Canada L5L 1 C6 Received March 25, 1988 Revision accepted July 13, 1988 Redescription of Colobomycterpholeter Vaughn, a small amniote from the Lower Permian of Oklahoma, provides evidence that it is not a mammal-like reptile. Osteological features used to associate Colobomycter with the family Eothyrididae are also present in protorothyridids. Colobomycter has a large sheet-like jugal in the temporal region, precluding the presence of a pelycosaurian lateral temporal opening. Derived osteological features shared with members of the family Protorothyridi- dae suggest that Colobomycter might be a member of that family, but this assignment must remain tentative because of the fragmentary nature of the known specimens. Une redescription de Colobomycter pholeter Vaughn, un petit amniote du Permien infkrieur de I'Oklahoma, dCmontre qu'il n'est pas un reptile mammalien. Les caractkres ostCologiques utilists pour associer Colobomycter avec la famille Eothyrididae sont Cgalement prCsents chez les protorothyridides. Colobomycter a un grand jugal rectangulaire dans la rtgion temporale, excluant la prCsence d'une f6netre temporale lattrale. Des caractkres osttologiques dCrivCs Cgalement prCsents chez les protorothyrididCs suggbrent que Colobomycter peut &treun membre de cette famille, mais cette assignation reste incertaine i cause de 1'Ctat fragmentaire des sptcimens connus. Can. J. Earth Sci. 26, 544-550 (1989) I Introduction One of the most problematic groups of pelycosaurs, the family Eothyrididae, was erected as a provisional group by Romer and Price (1940) for the reception of the small pelyco- saur Eothyris and three much larger forms, Baldwinonus trux, Stereophallodon ciscoensis, and Stereorchachis dominans. The latter three pelycosaurs were included within this family because they are "primitive in most known regards but paralleling the higher sphenacodonts in the development of much enlarged canines and showing a tendency toward elonga- tion of the vertebral column" (Romer and Price 1940). Subsequent studies resulted in the placement of additional new taxa into this family: Bayloria morei (Olson, 1941), represented by a single partial skull; Colobomycter pholeter (Vaughn, 1958), represented by a single partial skull; Delo- rhynchus priscus (Fox, 1962), represented by a few isolated maxillae; and Oedaleops campi (Langston, 1965), represented by two partial skulls and isolated limb elements. Recently, the number of eothyridids has been reduced by the study and removal of many of the above taxa from the family. Reisz and Heaton (1982) demonstrated that Bayloria morei is an immature specimen of Captorhinus aguti. Delorhynchus priscus is now considered as being a reptile of uncertain taxo- nomic position (Reisz 1986). Baldwinonus trux, Stereophallo- don ciscoensis, and Stereorchachis dominans have also been removed from the family and placed within the Ophiacodonti- dae (Reisz 1986: Brinkrnan and Eberth 1986). Three genera now remain in this family: Eothyris parkeyi (the type species), Oedaleops campi, and Colobomycter pho- leter. The subject of this paper, Colobomycter pholeter, is represented by part of a skull (Fig. 1) and an incomplete left maxilla (Fig. 2) from the Lower Permian fissure fills at the Dolese Brothers limestone quarry near Fort Sill, Oklahoma. For the original description, Vaughn (1958) prepared and illustrated the external surface of the skull and compared it with other small Lower Permian reptiles known at that time. Recent preparation of the internal surface of the holotype of C. pholeter permits a reevaluation of the taxonomic position and phylogenetic relationships of this unusual reptile. This study is particularly timely because recent increases in our knowledge of primitive captorhinomorph reptiles permit com- parisons with members of that group as well as with the two remaining eothyridids, Oedaleops campi and Eothyrisparkeyi. Description and comparisons The holotype of Colobomycter pholeter (Field Museum of Natural History, Chicago, Illinois, FMNH 272) consists of the following complete cranial elements from the right side of the skull roof and cheek: nasal, lacrimal, jugal, prefrontal, frontal, and postfrontal. The right maxilla, parietal, and post- orbital bones lacking their posterior margins are also preserved (Fig. 1). The external surface of the nasal, frontal, postfrontal, post- orbital, jugal, and prefrontal are marked by shallow, round pits. Although caseids have this pattern of sculpturing, the protorothyridids Paleothyris acadiana, Protorothyris archeri, and BroufJia orientalis also have similar external pitting on the skull root (Carroll 1969; Carroll and Baird 1972; Clark and Carroll 1973). This is in strong contrast to the condition seen in Oedaleops, where a heavy sculpturing pattern is evident on the skull roof (Langston 1965). The skull is similar in size and shape to those of protoro- thyridids. The distance from the orbit to the naris is 9.0 mrn. The orbit is large (13.5 mm long and 11.5 mm high) and sub- circular in outline. The nasal is larger than in other small reptiles, with an unusually long contact with the maxilla on the external sur- face. It is bordered posterolaterally by the prefrontal. The nasal joins the frontal in a posteriorly concave contact. Anter- iorly, it appears to have contacted the premaxilla (not pre- served) in an interdigitating suture, as seen on the dorsal surface. A posterolateral extension of the nasal, wedged between the maxilla and prefrontal, contacts the lacrimal in a short suture. Printed in Canada 1 Imprim6 au Canada Can. J. Earth Sci. Downloaded from www.nrcresearchpress.com by Renmin University of China on 05/28/13 For personal use only.
Transcript of Taxonomic position and phylogenetic relationships of Colobomycter pholeter, a small reptile from the...

Taxonomic position and phylogenetic relationships of Colobomycter pholeter, a small reptile from the Lower Permian of Oklahoma
MICHEL LAURIN AND ROBERT R. REISZ Department of Zoology, University of Toronto, Erindale College, 3359 Mississauga Road N, Mississauga, Ont.,
Canada L5L 1 C6
Received March 25, 1988
Revision accepted July 13, 1988
Redescription of Colobomycterpholeter Vaughn, a small amniote from the Lower Permian of Oklahoma, provides evidence that it is not a mammal-like reptile. Osteological features used to associate Colobomycter with the family Eothyrididae are also present in protorothyridids. Colobomycter has a large sheet-like jugal in the temporal region, precluding the presence of a pelycosaurian lateral temporal opening. Derived osteological features shared with members of the family Protorothyridi- dae suggest that Colobomycter might be a member of that family, but this assignment must remain tentative because of the fragmentary nature of the known specimens.
Une redescription de Colobomycter pholeter Vaughn, un petit amniote du Permien infkrieur de I'Oklahoma, dCmontre qu'il n'est pas un reptile mammalien. Les caractkres ostCologiques utilists pour associer Colobomycter avec la famille Eothyrididae sont Cgalement prCsents chez les protorothyridides. Colobomycter a un grand jugal rectangulaire dans la rtgion temporale, excluant la prCsence d'une f6netre temporale lattrale. Des caractkres osttologiques dCrivCs Cgalement prCsents chez les protorothyrididCs suggbrent que Colobomycter peut &tre un membre de cette famille, mais cette assignation reste incertaine i cause de 1'Ctat fragmentaire des sptcimens connus.
Can. J. Earth Sci. 26, 544-550 (1989) I
Introduction One of the most problematic groups of pelycosaurs, the
family Eothyrididae, was erected as a provisional group by Romer and Price (1940) for the reception of the small pelyco- saur Eothyris and three much larger forms, Baldwinonus trux, Stereophallodon ciscoensis, and Stereorchachis dominans. The latter three pelycosaurs were included within this family because they are "primitive in most known regards but paralleling the higher sphenacodonts in the development of much enlarged canines and showing a tendency toward elonga- tion of the vertebral column" (Romer and Price 1940).
Subsequent studies resulted in the placement of additional new taxa into this family: Bayloria morei (Olson, 1941), represented by a single partial skull; Colobomycter pholeter (Vaughn, 1958), represented by a single partial skull; Delo- rhynchus priscus (Fox, 1962), represented by a few isolated maxillae; and Oedaleops campi (Langston, 1965), represented by two partial skulls and isolated limb elements.
Recently, the number of eothyridids has been reduced by the study and removal of many of the above taxa from the family. Reisz and Heaton (1982) demonstrated that Bayloria morei is an immature specimen of Captorhinus aguti. Delorhynchus priscus is now considered as being a reptile of uncertain taxo- nomic position (Reisz 1986). Baldwinonus trux, Stereophallo- don ciscoensis, and Stereorchachis dominans have also been removed from the family and placed within the Ophiacodonti- dae (Reisz 1986: Brinkrnan and Eberth 1986).
Three genera now remain in this family: Eothyris parkeyi (the type species), Oedaleops campi, and Colobomycter pho- leter. The subject of this paper, Colobomycter pholeter, is represented by part of a skull (Fig. 1) and an incomplete left maxilla (Fig. 2) from the Lower Permian fissure fills at the Dolese Brothers limestone quarry near Fort Sill, Oklahoma. For the original description, Vaughn (1958) prepared and illustrated the external surface of the skull and compared it with other small Lower Permian reptiles known at that time. Recent preparation of the internal surface of the holotype of
C. pholeter permits a reevaluation of the taxonomic position and phylogenetic relationships of this unusual reptile. This study is particularly timely because recent increases in our knowledge of primitive captorhinomorph reptiles permit com- parisons with members of that group as well as with the two remaining eothyridids, Oedaleops campi and Eothyrisparkeyi.
Description and comparisons The holotype of Colobomycter pholeter (Field Museum of
Natural History, Chicago, Illinois, FMNH 272) consists of the following complete cranial elements from the right side of the skull roof and cheek: nasal, lacrimal, jugal, prefrontal, frontal, and postfrontal. The right maxilla, parietal, and post- orbital bones lacking their posterior margins are also preserved (Fig. 1).
The external surface of the nasal, frontal, postfrontal, post- orbital, jugal, and prefrontal are marked by shallow, round pits. Although caseids have this pattern of sculpturing, the protorothyridids Paleothyris acadiana, Protorothyris archeri, and BroufJia orientalis also have similar external pitting on the skull root (Carroll 1969; Carroll and Baird 1972; Clark and Carroll 1973). This is in strong contrast to the condition seen in Oedaleops, where a heavy sculpturing pattern is evident on the skull roof (Langston 1965).
The skull is similar in size and shape to those of protoro- thyridids. The distance from the orbit to the naris is 9.0 mrn. The orbit is large (13.5 mm long and 11.5 mm high) and sub- circular in outline.
The nasal is larger than in other small reptiles, with an unusually long contact with the maxilla on the external sur- face. It is bordered posterolaterally by the prefrontal. The nasal joins the frontal in a posteriorly concave contact. Anter- iorly, it appears to have contacted the premaxilla (not pre- served) in an interdigitating suture, as seen on the dorsal surface. A posterolateral extension of the nasal, wedged between the maxilla and prefrontal, contacts the lacrimal in a short suture.
Printed in Canada 1 Imprim6 au Canada
Can
. J. E
arth
Sci
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Ren
min
Uni
vers
ity o
f C
hina
on
05/2
8/13
For
pers
onal
use
onl
y.

LAURIN AND REISZ
FIG. 1. Colobomycterpholeter. Holotype (FMNH 272). Partial skull in medial (A) and medioventral (B) views. Abbreviations: bc, braincase; f, frontal; j, jugal; 1, lacrimal; mx, maxilla; n, nasal; p, parietal; pf, postfrontal; prf, prefrontal; ?sph, sphenethmoid (uncertain identification); ?sq, squamosal (uncertain identification). Scale represents 1 cm.
The nasal has short contacts with the maxilla and the pre- frontal medially and meets the frontal and lacrimal in long sutures (Fig. I). Three short grooves on the internal surface end anteroventrally in foramina. These probably continue into the nasal in the form of an intraosseous canal. This canal prob- ably contained the orbitonasal vein as in Eocaptorhinus lati- ceps (Heaton, 1979). In Eocaptorhinus, however, a single large foramen and numerous small foramina communicated with this tube, so the structures are not identical to those of Colobomycter but are at the same location, and both seem to have carried venous blood from the premaxilla.
As in protorothyridids, the maxilla is a narrow, elongate ele- ment that reaches its maximum height above the caniniform tooth. The dorsal expansion is pronounced in lateral view but
is surprisingly small in medial view. In lateral view the dorsal expanse of the maxilla decreases rapidly posteriorly, as in Cephalerpeton aff. C. ventriarmatum (Reisz and Baird, 1983). The suborbital rami of the lacrimal and jugal exclude the maxilla from the orbital margin. The posterior end of the maxilla is missing, but it probably did not extend farther than the posterior border of the jugal, as evidenced by the rate of tapering of this element.
The maxilla bears 12 teeth on the straight alveolar shelf and has alveoli for two more teeth. The posterior margin of the elongated external naris is located slightly anterior to the caniniform tooth. Anteriorly the maxilla has a well-developed premaxillary process that floors the external naris. The dorsal expanse not only forms the posterior border of the external
Can
. J. E
arth
Sci
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Ren
min
Uni
vers
ity o
f C
hina
on
05/2
8/13
For
pers
onal
use
onl
y.
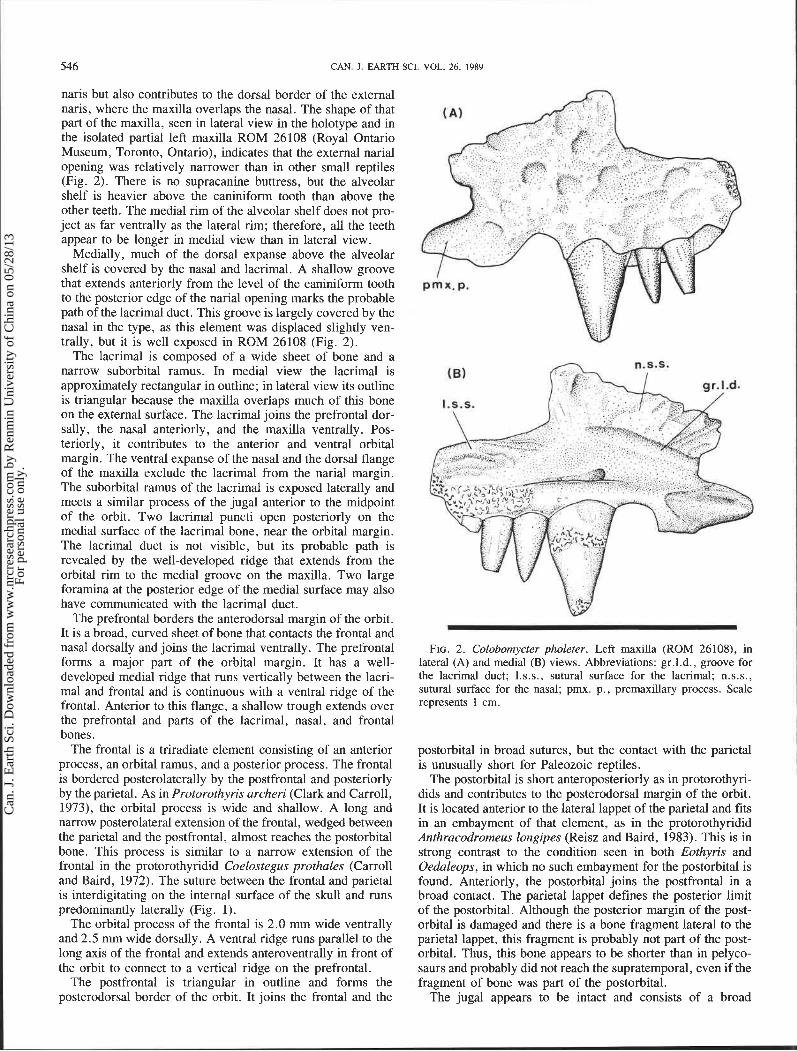
546 CAN. J. EARTH SCI. VOL. 26, 1989 I naris but also contributes to the dorsal border of the external naris, where the maxilla overlaps the nasal. The shape of that part of the maxilla, seen in lateral view in the holotype and in the isolated partial left maxilla ROM 26108 (Royal Ontario Museum, Toronto, Ontario), indicates that the external narial opening was relatively narrower than in other small reptiles (Fig. 2). There is no supracanine buttress, but the alveolar shelf is heavier above the caniniform tooth than above the other teeth. The medial rim of the alveolar shelf does not pro- ject as far ventrally as the lateral rim; therefore, all the teeth appear to be longer in medial view than in lateral view.
Medially, much of the dorsal expanse above the alveolar shelf is covered by the nasal and lacrimal. A shallow groove that extends anteriorly from the level of the caniniform tooth to the posterior edge of the narial opening marks the probable path of the lacrimal duct. This groove is largely covered by the nasal in the type, as this element was displaced slightly ven- trally, but it is well exposed in ROM 26108 (Fig. 2).
The lacrimal is composed of a wide sheet of bone and a narrow suborbital ramus. In medial view the lacrimal is approximately rectangular in outline; in lateral view its outline is triangular because the maxilla overlaps much of this bone on the external surface. The lacrimal joins the prefrontal dor- sally, the nasal anteriorly, and the maxilla ventrally. Pos- teriorly, it contributes to the anterior and ventral orbital margin. The ventral expanse of the nasal and the dorsal flange of the maxilla exclude the lacrimal from the narial margin. The suborbital ramus of the lacrimal is exposed laterally and meets a similar process of the jugal anterior to the midpoint of the orbit. Two lacrimal puncti open posteriorly on the medial surface of the lacrimal bone, near the orbital margin. The lacrimal duct is not visible, but its probable path is revealed by the well-developed ridge that extends from the orbital rim to the medial groove on the maxilla. Two large foramina at the posterior edge of the medial surface may also have communicated with the lacrimal duct.
The prefrontal borders the anterodorsal margin of the orbit. It is a broad, curved sheet of bone that contacts the frontal and nasal dorsally and joins the lacrimal ventrally. The prefrontal forms a major part of the orbital margin. It has a well- developed medial ridge that runs vertically between the lacri- mal and frontal and is continuous with a ventral ridge of the frontal. Anterior to this flange, a shallow trough extends over the prefrontal and parts of the lacrimal, nasal, and frontal bones.
The frontal is a triradiate element consisting of an anterior process, an orbital ramus, and a posterior process. The frontal is bordered posterolaterally by the postfrontal and posteriorly by the parietal. As in Protorothyris archeri (Clark and Carroll, 1973), the orbital process is wide and shallow. A long and narrow posterolateral extension of the frontal, wedged between the parietal and the postfrontal, almost reaches the postorbital bone. This process is similar to a narrow extension of the frontal in the protorothyridid Coelostegus prothales (Carroll and Baird, 1972). The suture between the frontal and parietal is interdigitating on the internal surface of the skull and runs predominantly laterally (Fig. 1).
The orbital process of the frontal is 2.0 mm wide ventrally and 2.5 mm wide dorsally. A ventral ridge runs parallel to the long axis of the frontal and extends anteroventrally in front of the orbit to connect to a vertical ridge on the prefrontal.
The postfrontal is triangular in outline and forms the posterodorsal border of the orbit. It joins the frontal and the
FIG. 2 . Colobomycter pholeter. Left maxilla (ROM 26108), in lateral (A) and medial (B) views. Abbreviations: gr.l.d., groove for the lacrimal duct; I.s.s., sutural surface for the lacrimal; n.s.s., sutural surface for the nasal; pmx. p., premaxillary process. Scale represents 1 cm.
postorbital in broad sutures, but the contact with the parietal is unusually short for Paleozoic reptiles.
The postorbital is short anteroposteriorly as in protorothyri- dids and contributes to the posterodorsal margin of the orbit. It is located anterior to the lateral lappet of the parietal and fits in an embayment of that element, as in the protorothyridid Anthracodromeus longipes (Reisz and Baird, 1983). This is in strong contrast to the condition seen in both Eothyris and Oedaleops, in which no such embayment for the postorbital is found. Anteriorly, the postorbital joins the postfrontal in a broad contact. The parietal lappet defines the posterior limit of the postorbital. Although the posterior margin of the post- orbital is damaged and there is a bone fragment lateral to the parietal lappet, this fragment is probably not part of the post- orbital. Thus, this bone appears to be shorter than in pelyco- saurs and probably did not reach the supratemporal, even if the fragment of bone was part of the postorbital.
The jugal appears to be intact and consists of a broad
Can
. J. E
arth
Sci
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Ren
min
Uni
vers
ity o
f C
hina
on
05/2
8/13
For
pers
onal
use
onl
y.

LAURIN AND REISZ 547
posterior sheet of bone and a narrow anterior suborbital ramus; it contributes to the posterior and ventral margins of the orbit. As in all captorhinomorph reptiles, the jugal of Colobomycter is not divided behind the orbit into a posterior or subtemporal ramus and a narrow dorsal postorbital ramus but is a large, broad sheet. In contrast, the jugal of all known pelycosaurs is a triradiate element with discrete narrow sub- orbital, dorsal, and posterior or subtemporal rami, with the latter two forming part of the border of the temporal fenestra. The posterior margin of the jugal of Colobomycter is almost vertical, seems to be intact, and shows no sign of the presence of a lateral temporal fenestra. The jugal contacts the post- orbital in a deeply concave suture that runs anteroventrally and swings anterodorsally shortly before reaching the orbit. In Eothyris and Oedaleops, the jugal-postorbital suture runs posterodorsally, as in most pelycosaurs.
The parietal is a broad, flat sheet of bone with a lateral lappet. The relatively large pineal foramen is located midway along the medial edge of the parietal. The posterolateral wing of the parietal is not preserved. This part of the bone is impor- tant because it usually carries a groove for the supratemporal. We are thus able to determine the shape of the groove and hence an indication of the size of the missing supratemporal.
The internal sutures between the postorbital, postfrontal, parietal, and jugal are hidden by two flat pieces of bone and fragments of the braincase. One of these pieces lies on the postorbital and surrounding bones. It is incomplete, approxi- mately circular in outline, and tentatively identified as the sphenethmoid. On the posteroventral part of the orbit lies the other, triangular piece of bone. A medial fold along its ventral margin indicates that it might be a piece of the squamosal, but this identification is also tentative.
Discussion In the original description of Colobomycter, Vaughn (1958)
assigned this small reptile to the Eothyrididae, on the basis of the following perceived similarities between Colobomycter and Eothyris :
(1) Both Eothyris and Colobomycter have a short face. Vaughn used the ratio between the interorbital distance and the distance between the orbit and the naris to measure the relative length of the face. This method seems inappropriate because the above ratio depends on the width of the skull, the degree to which the skull roof is emarginated above the orbit, the size of the orbit, the length of the face, and the location, size, and shape of the external naris. This ratio is affected by the fact that Colobomycter has a larger orbit than Eothyris and that the region of the naris has been greatly modified in Colobomycter. In order to evaluate the length of the face, it is preferable to compare the estimated antorbital length with the total length of the skull. This comparison indicates that Colobomycter, Cephalerpeton ventriarmatum, and Oedaleops have similar ratios of antorbital length / total length of the skull. This morphological feature therefore appears to be shared by Colobomycter, eothyridids, and some protorothyridids.
(2) According to Vaughn, the size of the pineal foramen of Colobomycter is similar to that of Eothyris. Measurements indicate, however, that the pineal foramen of Colobomycter (2.5 mm in diameter) is smaller than that of Eothyris (4.5 mm), but it falls within the size range of pineal foramina of captorhinomorphs of similar size (1.5 - 3.0 mm in diam- eter). As the skulls of the genera listed above are of similar
size, direct comparisons of the diameter of the pineal foramen are meaningful.
(3) According to Vaughn, the dentition of Colobomycter and that of Eothyris are similar. The maxillary dentition is reduced in both to 14 teeth, and the first and second maxillary teeth are said to be enlarged. Among primitive reptiles, the protoro- thyridid Cephalerpeton and the eothyridid Oedaleops have also reduced their dentition to 16 maxillary teeth. Further- more, Oedaleops has an enlarged precaniniform tooth, in con- trast to the condition seen in Eothyris. The first maxillary tooth of Colobomycter is small, but Vaughn suggested that it would have grown larger and become a caniniform. The evidence provided by Vaughn for this is the apparent large size of the socket. As seen in medial view the alveolar shelf of the holo- type is too thin above this tooth to support a caniniform tooth. In addition, the position of the socket is too far forward for a caniniform tooth, as it is located below the external naris. No known pelycosaur, protorothyridid, or captorhinid has a caniniform tooth located below the nostril. Since the gap behind the second maxillary tooth is small, there is only one caniniform tooth in Colobomycter, in contrast to the condition seen in Oedaleops and Eothyris, where two caniniform teeth are found.
(4) The size of the naris is similar in Colobomycter and Eothyris. The size of the naris of these reptiles may be similar, but their shape and morphology appear to be different: the naris is a narrow, elongate slit in Colobomycter, whereas in both Oedaleops and Eothyris it appears to be a large, oval opening. In the latter two genera, the lacrimal borders the naris posteriorly, the maxilla defines the floor of the nostril, and the nasal forms a shelf above the naris. In Colobomycter the lacrimal is excluded from the narial margin by the maxilla and nasal. Thus, the maxilla contributes to the posterior and ventral margins of this opening. Furthermore, there is no shelf above the naris on the nasal of Colobomycter. Because of these osteological differences, a feature such as size of narial opening becomes largely meaningless in evaluating rela- tionships.
(5) According to Vaughn, both Colobomycter and Eothyris have a broad postorbital bar. No direct comparison is possible between Colobomycter and Eothyris in this region of the skull because there is no evidence that there was a lateral temporal fenestra in Colobomycter or a pelycosaurian postorbital bar. In all pelycosaurs, including Eothyris and Oedaleops, the jugal and postorbital form a postorbital bar that separates the orbit from the temporal fenestra. There is no evidence that the jugal of Colobomycter has a reduced postorbital region that would form a postorbital bar. In Eothyris the width of the jugal is equal to one quarter the length of the orbit, whereas the width of the jugal is equal to one third the length of the orbit in Colobomycter.
(6) Eothyris and Colobomycter are said to have large supra- temporals. The size of this bone is unknown in Colobomycter because neither the supratemporal nor the embayment of the parietal into which it fits is preserved. Vaughn estimated the size of the supratemporal from the length of the posterolateral edge of the parietal. This method is not convincing, because there is no simple correlation between the part of the parietal that is preserved in Colobomycter and the size of the supra- temporal. The reason why no such correlation can be estab- lished is that the shape of the posterolateral wing of the parietal varies considerably among primitive reptiles. It is wide and short anteroposteriorly in Anthracodromeus longipes (Reisz
Can
. J. E
arth
Sci
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Ren
min
Uni
vers
ity o
f C
hina
on
05/2
8/13
For
pers
onal
use
onl
y.

548 CAN. J. EARTH SCI. VOL. 26, 1989
and Baird, 1983), whereas it is narrow and long anteropos- teriorly in Coelostegus prothales (Carroll and Baird, 1972). The posterior half of the parietal may widen posteriorly, as in Protorothyris archeri (Clark and Carroll, 1973), or narrow posteriorly, as in Anthracodromeus longipes. Furthermore, the supratemporal may be narrower than the posterolateral wing of the parietal, as in Protorothyris archeri, or larger than this part of the parietal, as in Coleostegus prothales or Oedaleops. Thus, the size of the supratemporal of Colobo- mycter cannot be determined from the available information.
The similarities of the snout mentioned by Vaughn (1958), such as a short face, a reduced dentition, anteriorly located caniniform teeth, and a large naris, seem to have evolved inde- pendently in Colobomycter and Eothyris, since the pattern of bones in this area is very different in these two genera. In Eothyris the long lacrimal reach the external naris, the nasal forms a shelf above the naris, and the maxilla is long, slender, and probably thin, as in Oedaleops. Colobomycter has a ven- trally enlarged nasal and a dorsally enlarged maxilla that exclude the lacrimal from the narial margin. Furthermore, the dorsal expanse of the maxilla is thick and high above the caniniform tooth, and there is no shelf above the narrow naris.
Thus, Colobomycter shares few similarities with Eothyris. Important differences between Colobomycter and Eothyris or Oedaleops include the following:
(1) The skull of Colobomycter appears to be proportionately higher and narrower than the skull of either Eothyris or Oedaleops.
(2) The jugal of Colobomycter is not a triradiate structure, whereas Eothyris and all known pelycosaurs have a triradiate jugal. In pelycosaurs, the jugal expands posteriorly in a sub- temporal ramus, which contributes to the ventral margin of the lateral temporal fenestra. In Colobomycter, this element con- sists of a broad sheet of bone and a narrow suborbital ramus. The jugal has no subtemporal ramus. The suture between the jugal and postorbital runs anteroventrally and swings antero- dorsally shortly before reaching the orbit. The contact between this bone and the postorbital runs anteroventrally in Eothyris and Oedaleops.
( 3 ) The lacrimal of Colobomycter is excluded from the external narial margin by a dorsal expansion of the maxilla and a lateral lappet of the nasal bone, whereas in both Oedaleops and Eothyris the lacrimal reaches the margin of the narial opening.
(4) The postorbital is anterior to the lateral lappet of the parietal in Colobomycter, whereas it is situated laterally to this lappet in both Oedaleops and Eothyris. The postorbital there- fore fits into an embayment of the parietal in Colobomycter, but no such embayment is found in either Oedaleops or Eothyris. In Colobomycter, the postorbital is short antero- posteriorly and probably does not reach the supratemporal, a derived condition found only among protorothyridids, capto- rhinids, and diapsids (Heaton and Reisz 1986). Eothyris and all pelycosaurs have an anteroposteriorly long postorbital that reaches the supratemporal.
(5) The dorsal expansion of the maxilla reaches its greatest height above the caniniform tooth in Colobomycter and decreases rapidly posteriorly. In Eothyris and Oedaleops, the dorsal expansion of this bone does not show a marked increase in height about the caniniform teeth. The maxilla of Colobo- mycter is therefore very different from that of Eothyris or Oedaleops .
(6) The lateral process of the frontal entering the orbit is
wide in Colobomycter, whereas it is narrow in Oedaleops and absent in Eothyris.
(7) The sculpturing of the skull roof of Colobomycter con- sists of shallow, round pits. This is different from the heavy pattern of corrugations and grooves of Oedaleops (the sculpturing pattern of Eothyris is unknown because its surface appears to have been damaged during preparation).
The aforementioned differences between Colobomycter and Eothyris indicate that they do not belong to the same family. Vaughn (1958) assigned Colobomycter to the Pelycosauria on the basis of the following features:
(1) The exclusion of the lacrimal from the narial margin. This character may be shared with Cephalerpeton aff. C. ven- triarmatum, in which the dorsally enlarged maxilla may have excluded the lacrimal from the narial margin, at least laterally (Reisz and Baird 1983). Among pelycosaurs, this feature is found only in Mycterosaurus (Berman and Reisz 1982) and advanced sphenacodontids. Thus, this character of the lacrimal cannot be used to include Colobomycter in the Pelycosauria, because this is not a diagnostic character of pelycosaurs.
(2) A short face. As described earlier in this article, some protorothyridids also share this character with Colobomycter. Therefore, this feature cannot be used to include Colobo- mycter in the Pelycosauria. Because protorothyridids have been described in detail only in more recent work (Carrol 1964, 1969; Carroll and Baird 1972), Vaughn could not com- pare Colobomycter with these small reptiles.
(3) The location of the largest maxillary tooth far forward. This feature seems to depend on the diet and mode of life of the animal and probably evolved independently in Colobo- mycter and in Eothyris or Oedaleops. Increase in size of the caniniform tooth and decrease in number of precaniniform maxillary dentition occur at least twice among pelycosaurs and is also noted in other primitive reptiles. Among protorothyri- dids, Cephalerpeton aff. C. ventriarmatum (Reisz and Baird, 1983) shows a marked increase in the size of the caniniform tooth over the geologically older holotype of Cephalerpeton ventriarmatum (Carroll and Baird, 1972).
Similarities to protorothyridids include the following: The lateral process of the frontal entering the orbital margin is wide, as in Protorothyris archeri. As in Cephulerpeton aff. C. ventriarmatum (Reisz and Baird, 1983) and Cephalerpeton ventriarmatum (Carroll and Baird, 1972), much of the orbital margin of the lacrimal is covered medially by a ventral process of the prefrontal. The distinctive posterior margin of the jugal of Colobomycter resembles that of Protorothyris archeri and Anthracodromeus longipes; it is almost vertical in these three genera. The lacrimal appears similar to that of Cephalerpeton aff. C. ventriarmatum and is probably excluded from the narial opening by a dorsal extension of the maxilla. Colobo- mycter and Cephalerpeton aff. C. ventriarmatum share the derived condition of a maxilla with a significant dorsal expan- sion above the caniniform tooth. This is different from the maxillae of Eothyris and Oedaleops, which do not show a dis- tinct increase in height above the caniniform teeth.
Conclusion
The protorothyridids were poorly known when Vaughn (1958) described Colobomycter. It was, therefore, natural for him to compare this interesting specimen with the better known primitive synapsid Eothyris. Restudy of Colobomycter
Can
. J. E
arth
Sci
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Ren
min
Uni
vers
ity o
f C
hina
on
05/2
8/13
For
pers
onal
use
onl
y.

LAURIN A ,ND REISZ 549
indicates that it is not a primitive pelycosaur. The shape of the jugal, indicating the lack of lateral temporal fenestrae, is the most important primitive osteological feature of Colobomycter that appears in derived form in all pelycosaurs. Colobomycter shows an advanced condition over eothyridids in the following features: short postorbital that probably did not reach the supratemporal; narrow and deep snout; short lacrimal excluded from the naris by the maxilla and the nasal; and wide orbital process of the frontal. The removal of Colobomycter from the Eothyrididae leaves only two very similar genera, Eothyris and Oedaleops, in that family.
Similarities between the skull of Colobomycter and protoro- thyridid captorhinomorphs indicate that this small reptile from Lower Permian deposits near Fort Sill, Oklahoma, may be a member of that family. Derived characters of Colobomycter shared with protorothyridids include short postorbital that does not reach the supratemporal; wide orbital process of the frontal and narrow and deep skull. Although these characters are also found in other groups, they suggest that Colobomycter is a protorothyridid. Assignment of Colobomycter to the family Protorothyrididae must remain tentative because the holotype and only known specimen of C. pholeter is incom- plete. In addition, the Protorothyrididae group is problematic because it is defined on the basis of shared ~rimitive characters and shared derived characters also found in other groups; it may be an artificial assemblage of reptiles representing several lineages (Carroll 1982; Heaton and Reisz 1986).
Colobomycter seems to be closely related to Cephalerpeton aff. C. ventriarmatum, a small protorothyridid from the Late Pennsylvanian of Linton, Ohio; they both share the following derived characters: a reduced number of maxillary teeth, enlarged caniniform tooth, well-developed ventral flange of the prefrontal, dorsally enlarged maxilla, and short lacrimal. They may represent a lineage of protorothyridids that includes the geologically older Cephalerpeton ventriamtum (from the Middle Pennsylvanian of Mazon Creek). In C. ventriamtum, the lacrimal is elongate, with a long groove on the medial surface where the lacrimal duct becbmes subosseous. In Cephalelpeton aff. C. ventriamtum, the maxilla is enlarged dorsally, the lacrimal is shorter than in Cephalelpeton ven- triarmatum, and the groove for the lacrimal duct is located at the anterior end of the lacrimal. With continued expansion of the maxilla, the groove for the lacrimal duct may have extended onto the medial surface on the maxilla in Colobo- mycter. Thus, Cephalelpeton ventriarmatum, cephalelpeton aff. C. ventriarmatum, and Colobomycter may represent a phylogenetic line within the protorothyridids, with evidence of a progressive reduction of the length of the lacrimals. Mem- bers of this line are characterized by a reduction of the dental formula, enlargement of the caniniform tooth, and enlarge- ment of the maxilla.
Systematic paleontology CLASS Reptilia Linnaeus, 1758
SUBCLASS Eureptilia Olson, 1947 ORDER Captorhina Olson, 1947
FAMILY Protorothyrididae Price, 1937 GENUS Colobornycter Vaughn, 1958
Colobomycter pholeter Vaughn, 1958
Holotype-FMNH 272. Part of the right cheek and the skull table.
Referred specimen-ROM 26108. A broken piece of a left maxilla.
Horizon and locality-Fissure deposit in Arbuckle limestone at the Dolose Brothers limestone quarry, north of Fort Sill, in sec. 3 1, tp. 4N, rge. 11 W, Commanche County, Oklahoma. These sediments are of Early Permian age, possibly equivalent to the Arroyo Formation, lower Clear Fork Group of Texas (Heaton 1979).
Revised diagnosis-Average-sized protorothyridid capto- rhinomorph distinguished from all other known protorothyri- dids by its reduced dentition. Characterized by a short face, elongated external naris, simple, conical teeth, and large caniniform tooth. Lacrimal is excluded from the narial open- ing by the dorsally enlarged maxilla and a ventrolateral exten- sion of the nasal bone.
Acknowledgments
We are indebted to Ms. Heather Wilson and Ms. Diane Scott, who critically read the manuscript. Ms. Diane Scott also drew the illustration of the holotype of C. pholeter. We thank Dr. Pierre Couillard, whose support allowed one of us (Michel Laurin) to receive a scholarship for this research. Thanks are also due to Dr. John Bolt, who lent us the holotype of Colobomycter. This research was supported by the Natural Sciences and Engineering Research Council of Canada.
BERMAN, D. S., and REISZ, R. R. 1982. Restudy of Mycterosaurus longiceps (Reptilia, Pelycosauria) from the Lower Permian of Texas. Annals of the Carnegie Museum, 51: 423-453.
BRINKMAN: D., and EBERTH, D. A. 1986. The anatomy and relation- ships of Stereophallodon and Baldwinonus (Reptilia, Pelyco- sauria) . Breviora, 485: 1 - 34.
CARROLL, R. L. 1964. The earliest reptiles. Journal of the Linnean Society, Zoology, 45: 61 - 83.
1969. A middle Pennsylvanian captorhinomorph, and the interrelationships of primitive reptiles. Journal of Paleontology, 43: 151 -170.
1982. Early evolution of reptiles. Annual Review of Ecology and Systematics 13: 87- 109.
CARROLL, R. L., and BAIRD, D. 1972. Carboniferous stem reptiles of the family Romeriidae. Bulletin of the Museum of Comparative Zoology at Harvard University, 143: 32 1 - 364.
CLARK, J., and CARROLL, R. L. 1973. Romeriid reptiles from the Lower Permian. Bulletin of the Museum of Comparative Zoology at Harvard University, 144: 353 -407.
Fox, R. C. 1962. Two new pelycosaurs from the Lower Permian of Oklahoma. University of Kansas Publications of the Museum of Natural History, 12: 297 - 307.
HEATON, M. J. 1979. Cranial anatomy of primitive captorhinid rep- tiles from the Late Pennsylvanian and Early Permian Oklahoma and Texas. Oklahoma Geological Survey, Norman, OK, Bulletin 127, pp. 1-84.
HEATON, It$. J., and REISZ, R. R. 1986. Phylogenetic relationships of captorhinomorph reptiles. Canadian Journal of Earth Sciences, 23: 4 0 2 ~ 4 1 8 .
LANGSTON, W., JR. 1965. Oedaleops campi (Reptilia; Pelycosauria). New genus and species from the Lower Permian of New Mexico and the family Eothyrididae. Texas Memorial Museum Bulletin, No. 9, pp. 1-48.
OLSON, E:, C. 1941. New specimens of Permian vertebrates in Walker Museum. Journal of Geology (Chicago), 49: 753-763.
REISZ, R. R. 1980. A protorothyridid captorhinomorph reptile from the Lower Permian of Oklahoma. Royal Ontario Museum Life Sciences Contributions, No. 121, pp. 1 - 16.
Can
. J. E
arth
Sci
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Ren
min
Uni
vers
ity o
f C
hina
on
05/2
8/13
For
pers
onal
use
onl
y.

550 CAN. 1. EARTH SCI. VOL. 26, 1989
1986. Encyclopedia of palaeoherpetology . Part 17A. Pelyco- hinus aguti (Cope, 1882). Canadian Journal of Earth Sciences, 19: sauria. Gustav Fischer Verlag, Stuttgart, pp. 1 - 102. 1232-1234.
REISZ, R. R., and BAIRD, D. 1983. Captorhinomorph "stem" reptiles ROMER, A. S., and PRICE, L. I. 1940. Review of the Pelycosauria. from the Pennsylvanian coal-swamp deposit of Linton, Ohio. Geological Society of America, Special Paper 28, pp. 1-588. Annals of the Carnegie Museum, 52: 393 -41 1. VAUGHN, P. P. 1958. On a new pelycosaur from the Lower Permian
REISZ, R. R., and HEATON, M. J. 1982. Bayloria morei Olson 1941 of Oklahoma, and on the origin of the family Caseidae. Journal of identified as an immature specimen of the Permian reptile Captor- Paleontology, 32: 981 -991.
Can
. J. E
arth
Sci
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Ren
min
Uni
vers
ity o
f C
hina
on
05/2
8/13
For
pers
onal
use
onl
y.




















