
Studies on the swimming musculature of the rainbow trout.. II. Muscle metabolism during severe...
9
Reprinted from I. Fish Bioi. (1975) 7, 451-458 Studies on the swimming musculature of the rainbow trout I. Fibre types I. A. JOHNSTON,* P. S. WARDt AND G. GOLDSPINKt * Research Unit for Comparative Animal Respiration, University of Bristol, Woodland Road, Bristol BSB 1 UG, England and t Department ofZoology, University of Hull, Humberside, Yorkshire, England (Accepted 7 August 1974) A histochemical study has been made on the myotomal musculature of the rainbow trout Salmo gairdneri Richardson. Four main types of fibre can be distinguished on the basis of differences in fibre size, lipid content and succinic dehydrogenase and myofibrillar adenosine triphosphatase activity (A TPase activity). Early histological studies have concluded that in addition to a superficial strip of muscle small diameter. red' fibres occur throughout the trout myotome. These small diameter fibres occurring in the bulk of the myotome are shown to differ from the superficial fibres with respect to their oxidative metabolism and myofibrillar A TPase. The relevance of this finding to studies on the energetics and swimmin!t efficiency of this species are discussed. I. INTRODUCnON . The myotome of most teleost fish is composed of distinct red and white muscles. The red muscle forms a thin sheet lying just underneath the lateral line, and runs from behind the head to the caudal fin. Red fibres are known to have high con- centrations of myoglobin, cytochromes and lipids (George, 1962; Matsuura & Hashimoto, 1954), and have high activities of respiratory and citric acid cycle enzymes (Bostrom & Johansson, 1972). The bulk of the remaining muscle is made up of larger white fibres with low lipid contents and a predominantly glycolytic type of metabolism (George, 1962; Hamoir et al., 1972). Barany (1967) has shown that the myofibrillar or myosin A TPase of a number of vertebrate muscles parallels their speed of shortening. Biochemical studies of myofibrillar A TPase in fish have shown that the white and red muscles correspond to' fast' and' slow' muscles respectively (Johnston et al., 1972; Johnston, 1973). In salmonids, small diameter fibres occur throughout the myotome intermingled with the larger fibres, giving the muscle a characteristic mosaic appearance (Boddeke et al., 1959). Early histological studies concluded that this mosaic was comprised of two types of fibre, the smaller of which corresponded to that found in the superficial red muscle. Extensive histological and histochemical investigations of mammalian muscle have indicated that superficial classifications of muscle fibres on the basis of size and colour are misleading (Close, 1972). For example, while many red muscles are' slow' contracting, the bat crichothyroid muscle is an example of a' fast' red muscle (Revel, 1962). In this histochemical study of the trout, evidence is presented to show the red fibres of the mosaic and superficial muscles correspond to different fihre tvnes with resnect to their mvofibrillar A TPase activitv and oxidative metabolism.
Transcript of Studies on the swimming musculature of the rainbow trout.. II. Muscle metabolism during severe...

Reprinted from I. Fish Bioi. (1975) 7, 451-458
Studies on the swimming musculature of the rainbow trout
I. Fibre types
I. A. JOHNSTON,* P. S. WARDt AND G. GOLDSPINKt* Research Unit for Comparative Animal Respiration, University of Bristol,
Woodland Road, Bristol BSB 1 UG, Englandand
t Department ofZoology, University of Hull, Humberside, Yorkshire, England
(Accepted 7 August 1974)
A histochemical study has been made on the myotomal musculature of the rainbow troutSalmo gairdneri Richardson. Four main types of fibre can be distinguished on the basis ofdifferences in fibre size, lipid content and succinic dehydrogenase and myofibrillar adenosinetriphosphatase activity (A TPase activity). Early histological studies have concluded thatin addition to a superficial strip of muscle small diameter. red' fibres occur throughoutthe trout myotome. These small diameter fibres occurring in the bulk of the myotome areshown to differ from the superficial fibres with respect to their oxidative metabolism andmyofibrillar A TPase. The relevance of this finding to studies on the energetics andswimmin!t efficiency of this species are discussed.
I. INTRODUCnON .
The myotome of most teleost fish is composed of distinct red and white muscles.The red muscle forms a thin sheet lying just underneath the lateral line, and runsfrom behind the head to the caudal fin. Red fibres are known to have high con-centrations of myoglobin, cytochromes and lipids (George, 1962; Matsuura &Hashimoto, 1954), and have high activities of respiratory and citric acid cycle enzymes(Bostrom & Johansson, 1972). The bulk of the remaining muscle is made up oflarger white fibres with low lipid contents and a predominantly glycolytic type ofmetabolism (George, 1962; Hamoir et al., 1972). Barany (1967) has shown that themyofibrillar or myosin A TPase of a number of vertebrate muscles parallels theirspeed of shortening. Biochemical studies of myofibrillar A TPase in fish have shownthat the white and red muscles correspond to' fast' and' slow' muscles respectively(Johnston et al., 1972; Johnston, 1973).
In salmonids, small diameter fibres occur throughout the myotome intermingledwith the larger fibres, giving the muscle a characteristic mosaic appearance (Boddekeet al., 1959). Early histological studies concluded that this mosaic was comprised oftwo types of fibre, the smaller of which corresponded to that found in the superficialred muscle. Extensive histological and histochemical investigations of mammalianmuscle have indicated that superficial classifications of muscle fibres on the basis ofsize and colour are misleading (Close, 1972). For example, while many red musclesare' slow' contracting, the bat crichothyroid muscle is an example of a' fast' redmuscle (Revel, 1962). In this histochemical study of the trout, evidence is presentedto show the red fibres of the mosaic and superficial muscles correspond to differentfihre tvnes with resnect to their mvofibrillar A TPase activitv and oxidative metabolism.

452 I. A. JOHN-"TON ET AL
In the light of recent criticisms of the procedures used for demonstrating myofibrillarATPase (Guth, 1973), a critical examination has been also made of the specificity ofthis techniaue.
n. MATERIALS AND METHODS
Rainbow trout Salmo gairdneri Richardson about 10-15 cm long were maintained intanks of circulating tap water at 9.0::1: 1.0°C. The trout were stunned by a blow to the headand killed by decapitation. Muscle samples were taken from the region underneath theadipose fin. A region of muscle was dissected which included not only superficial red musclebut substantial amounts of mosaic muscle [Plate I(a)]. A mosaic muscle sample was takenfrom the deep ~paxial musculature adjacent to the vertebral column. Blocks of muscle werequickly dissected from the fish and mounted between mouse liver. The tissue was frozen byimmersion in dichlorodifluoromethane (Arcton 12, I.C.I.), cooled to its melting point( -158°C) with liquid nitrogen. Blocks were placed in a refrigerated cabinet at -25°C for1 h and then cryostat sections were cut at 8-12 ~ thickness. and mounted on coverslips.
HISTOCHEMICAL METHODS
LipidLipid was demonstrated by the method of Pearse (1960) using Sudan Black B.
Succinic dehydrogenase (SDH)
Sections were stained for succinic dehydrogenase activity using the nitro-blue tetrazoliummethod of Nachlas et at. (1957). Controls were incubated with either malonate replacingsuccinate in the incubation medium or with the nitro-blue tetrazolium salt bein2 omitted.
Myofibrillar adenosine triphosphatase (ATPase)MyofibriJlar A TPase activity was demonstrated using the method of Guth & SamaM
(1970) which is a modification of the technique of Padykula & Herman (1955), Somesections were prefixed in 13% sucrose, 100 mM sodium cacodylate pH 7'4, 1% paraformal-dehyde at 2°C, Fixation caused substantial inactivation of all the fibres in the trout asreported for other fish species (Johnston et al" 1974a, b), Sections were preincubatedwithout prior fixation for periods ranging from 0'5-10 min in a soJution of 18 mM CaCI2,100 mM 2-amino-2-methyl-l-propanol buffer at a series of alkaline pH's (10'0, 10-1, 10-2,10-3,10-4), Incubation for myofibrillar ATPase was for 20min at 21°C in the medium ofGuth & Samaha (1969), To inhibit Ca2+ activated mitochondrial ATPases, some sectionswere incubated with 3'5 mM sodium azide in the incubation medium. In addition somesections were treated with a 1% solution of the anionic detergent Triton X-I00 for 1 hat4°C to soJubilize membranous ATPase (Schiaffino & Bormioli, 1973), Sections treated inthis manner were repeatedly washed with 18 mM CaCIs, 100 mM tris-HCI pH 7'8, beforebeing subjected to incubation for myofibrillar A TPase activity as described above-
Serial sections were used in order to compare the results for the different treatments andfor staining for Jipid, myofibrillar A TPase and succinic dehydrogenase activities.
Fibre size measurements
Fibre diameters of the three types of fibre distinguished on the basis of differences inmyofibrillar ATPase activity were measured in each of five rainbow trout. A total of 100fibres were measured for each of five fish. The areas of sampling for superficial fibres,intermediate' pink' fibres and mosaic fibres are shown in Fig. 1.
m. RESUL TSLIPID
Only the superficial red fibres of region A (Fig. I) stained heavily for lipid. Theintermediately positioned fibres of high myofibrillar ATPase (see below) situatedbetween the superficial and mosaic muscle regions also had a slight staining for lipid.

453TROUT MUSCLE FIBRES
In contrast, the small and large diameter fibres of the mosaic muscle were completely
unstained [Plate I(a)].
SUCCINIC DEHYDROGENASE (SDH) ACTIVITYThe superficial red fibres in the region of the lateral line were heavily stained for
SDH activity. The pink fibres also showed some staining for this enzyme although
~ Pink muscle
~ Mosaic muscle
FIn
considerably less than the superficial red fibres. The fibres of the mosaic muscle wereunstained as were all the fibres of control sections. These results are illustrated in
Plate I(b ).
MYOFIBRILLAR ATPASE ACnVITYThe pattern of cobalt sulphide staining obtained on incubating tissue sections for
mvofibrillar A TPase was found to be dependent both on the time and pH of the
-.A diagram to show the distribution of muscle fibre types in the trunk musculature of the
rainbow trout Salmo f!airdneri.
454 I. A. JOHNSTON RT AT-
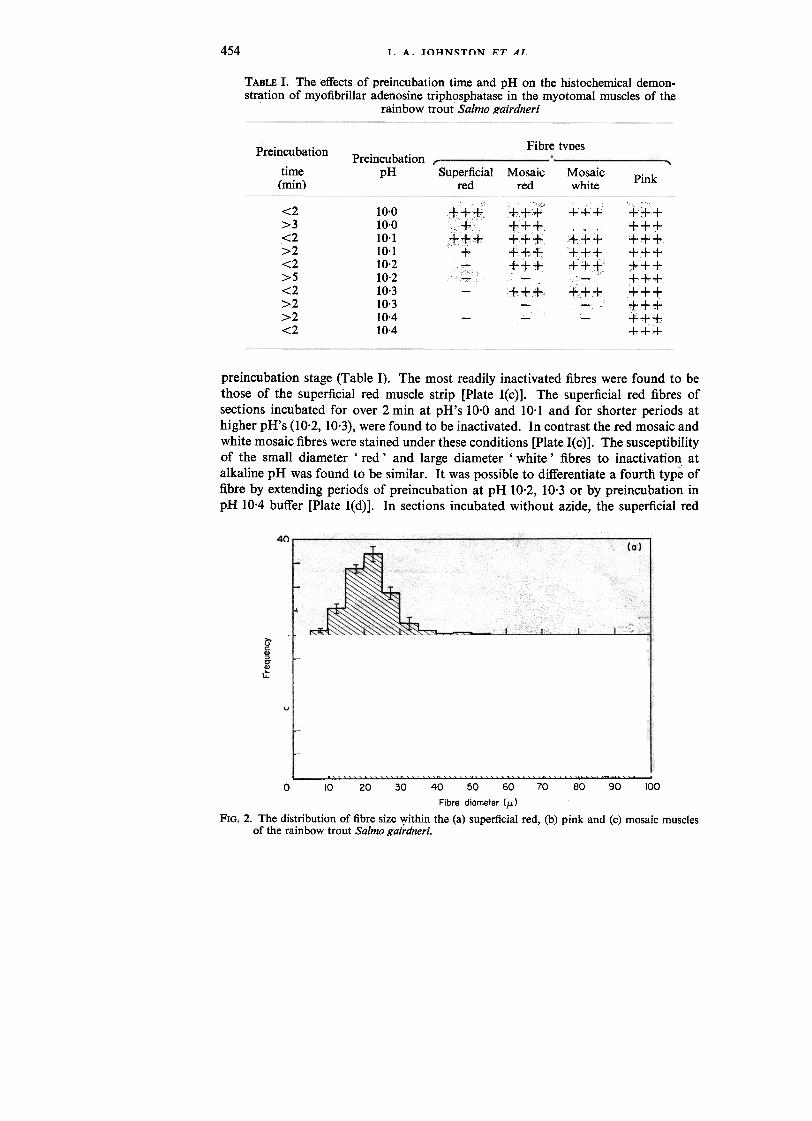
TABLE I. The effects of preincubation time and pH on the histochemical demon-stration of myofibrillar adenosine triphosphatase in the myotomal muscles of the
rainbow trout Salmo gairdneri
Fibre typesPreincubation
time(min)
PreincubationpH Superficial
redMosaic
redMosaicwhite
Pink
<2>3.<2>2<2>5<2>2>2<2
10.010.010.110.110.210.210.310.310.410.4
;"+++," "" c,,"+"
"'++c+c,
+
lT;:1;.~{C
+
+
+
+
+++i ++++f+++++
+
tcf
-
preincubation stage (Table I). The most readily inactivated fibres were found to bethose of the superficial red muscle strip [Plate I(c)]. The superficial red fibres ofsections incubated for over 2 min at pH's 10.0 and 10.1 and for shorter periods athigher pH's (10.2, 10.3), were found to be inactivated. In contrast the red mosaic andwhite mosaic fibres were stained under these conditions [Plate I(c)]. The susceptibilityof the small diameter' red' and large diameter' white' fibres to inactivation atalkaline pH was found to be similar. It was possible to differentiate a fourth type offibre by extending periods of preincubation at pH 10.2, 10.3 or by preincubation inpH 10.4 buffer [Plate I(d)]. In sections incubated without azide, the superficial red
30
20
10
~ c
c0)5- 200)
~10
(b)
-~~~~~ ~ I(c)
20
10
.-~~~~.~ I0 10 20 30 40 50 60 70 80 90 100
Fibre diameter (fJ.)
FIG. 2. The distribution of fibre size Vjithin the (a) superficial red, (b) pink and (c) mosaic musclesof the rainbow trout Salmo gairdneri.
+++++
+
+'++
~+
+
""+"+++++"++++
++++++++++
.
+
+
+
"...,
+
-
++f
+

.
.~
,.,_.,,"~,--,$$$ --~---PLATE I. (a) A section of rainbow trout myotome cut in the region of the lateral line which has
been stained for lipid. Note the heavily stained triangle of superficial red fibres (R). Fibreslabelled (P) show slight lipid staining while the fibres of the mosaic muscle are unstained.x 14. (b) Muscle stained for succinic dehydrogenase activity. Superficial red fibres areintensely stained but fibres of the mosaic muscle are unstained. x 20. (c) Myotomal musclepreincubated at pH 10.2 for 2 min before incubation for myofibrillar ATPase. Superficial redmuscle cells (R) have been inactivated, while small and large diameter fibres of the mosaicmuscle (W) have stained intensely. x 40. (d) Muscle incubated for myofibrillar ATPase inthe presence of 3.5 mM sodium azide. x36. (e) Muscle preincubated at pH 10.4 for 3 minprior to incubation for myofibrillar ATPase. Only pink fibres (P) have stained under theseconditions. x 57.

455TROIJT MIJ~rLR FIRRE~
fibres had a slight brown colouration [Plate I(c)] presumably due to staining attri-butable to Ca2+-activated mitochondrial ATPases. This effect was abolished insections incubated in the presence of 3.5 mM sodium azide [Plate I(e)]. The generalpattern of staining, however, remained the same, with the mosaic and intermediatelypositioned pink fibres stained heavily and superficial fibres completely unstained[Plate I(e)]. Treatment with Triton-X-loo only removed part of the brown colourationof the superficial red fibres after incubation at pH 9.4.
MEASUREMENTS OF FIBRE SIZEThe distribution of the fibre size within the superficial red, pink and mosaic muscle
regions is illustrated in Fig. 2. The superficial red fibres ranged in diameter from10-50 !l with a peak frequency around 20-25 !l [Fig. 2(a)]. The pink muscle showed awider range of fibre sizes than the red muscle (10-75 !l) with the majority of fibreshaving a diameter of 25-45 !l [Fig. 2(b)]. The distribution of fibres in the mosaicmuscle was larger, ranging from 15-95 !l [Fig. 2(c)]. There was an almost continuousdistribution of fibre size in this muscle type, with the majority of fibres havingdiameters of 35-55 !l. There were, however, a significant number of larger fibres
(50-75 J.l) [Fig. 2(c)].
IV. DISCUSSION
Biochemical determinations of myofibrillar ATPase activities in a number ofteleosts have shown that the white fibres have a specific activity 2-4 times higher thanthat of the superficial red fibres (Johnston et al., 1972; Johnston, 1973). A recentstudy of actomyosin ATPase activities (Nag, 1972) has given similar results for thered and white muscles of the rainbow trout Salmo gairdneri. The myofibrillar A TPaseof a muscle is of particular interest since it has been shown to parallel the speed ofcontraction of different muscles (Barany, 1967). Histochemical methods for demon-strating myofibrillar or myosin ATPase based on those ofPadykula & Herman (1955)have been shown to give results which correlate with biochemical determinations ofthese enzymes (Guth & Samaha, 1969, 1970). These methods involve selectivelyinactivating specific groups of fibres by preincubating at either acid or alkaline pH(Guth & Samaha, 1969, 1970). Mammalian fibres of low ATPase activity have beenshown to be acid stable and alkaline labile (Guth & Samaha, 1969; Close, 1972).
Fish muscle fibres are much more susceptible to inactivation at low and high pH'sthan mammalian fibres (Johnston et al., 1974a, b). Indeed the instability of the myosinand myofibrillar ATPases ofteleost white muscle i~ well documented (Johnston et al.,1973; Syrovy et al., 1970). In a previous study of a number of teleost species it wasfound that selective inactivation of the slow red fibres could be obtained by pre-incubating sections for short periods at alkaline pH (Johnston et al., 1974b). Themodification of the mammalian technique for demonstrating myofibrillar ATPaseactivity gives results for fish muscle which are correlated with biochemical data forthe same species (Johnston, 1973). The technique of Guth & Samaha (1969) fordemonstrating myofibrillar A TPase has recently been criticized on the grounds thatit also reflects mitochondrial ATPases and is therefore non-specific (Guth, 1973).
To eliminate the effect of mitochondrial A TPases, sections were' incubated in thepresence of sodium azide at a concentration known to inhibit all mitochondrialATPase activity (Brierly et al., 1964). Sonie sections were also treated with thedetergent Triton X-100 in order to solubilize sarcoplasmic ATPases (Schiaffino &

456 I. A. JOHNSTON ET AL
Bormiolli, 1973). Since essentially the same pattern of staining was obtained with allthese treatments, it appears that interference by non-myofibrillar ATPases is not verysignificant at pH 9.4 in fish muscle [Plate I(c), (e)]. In common with other teleosts(Johnston et al., 1974b), the superficial red fibres of the rainbow trout were foundto be readily inactivated by a brief exposure to alkaline pH. However, the mosaicfibres stained intensely following a similar period of preincubation. The histo-chemical results obtained for the trout agree with the biochemical studies of actomyosinATPase in the red and mosaic muscles of the same species (Nag, 1972).
Many species of fish also have a layer of fibres of intermediate size situated betweenthe red and white muscle regions (Bone, 1966; Brotchi, 1968; Walker, 1970). Thesefibres have an intermediate level of staining for oxidative enzymes and lipid betweenthe red and white fibres (George & Bokdawala, 1964; Ogata & Mori, 1964). Similarresults have been obtained for the citric acid cycle enzyme succinic dehydrogenaseand lipid in the present study. The pink fibres of the trout also correspond to theintermediately positioned fibres of other species with respect to their staining formyofibrillar ATPase (Johnston, 1973; Johnston et al., 1974b). Pink fibres havebeen found to stain similarly to the white fibres following short periods of pre-incubation at alkaline pH (Johnston et al., 1974b). These pink fibres are the typemost resistant to inactivation during exposure to alkaline pH (Table I). The pinkfibre region of the rainbow trout is relatively restricted compared to other speciessuch as the carp Carassius carassius and the grey mullet Mugil cephalus (Johnston,1973; Johnston et al., 1974b). In the trout these fibres occupy a thin band 3-4 cellsdeep situated between the superficial red and mosaic muscle regions (Fig. 1). Thepink fibres do not constitute a discrete region but gradually merge into fibres of the
..mosaic muscle [Plate I(c)].
Earlier histological studies concluded that the superficial and mosaic red fibres ofsalmonids were identical fibre types mainly from considerations of colour, lipidcontent and fibre size (Boddeke et al., 1959; Greene & Greene, 1913). Webb (1970)considered that these fibres were identical in structure and function. The presentstudy shows that mosaic red fibres differ from superficial red fibres in respect of theirsuccinic dehydrogenase and myofibrillar A TPase activities and correspond to differentfibre types. There is also physiological evidence that the two types of red fibre havedifferent thresholds of activation and different functions in the division of labourbetween the swimming muscles. It is known from electromyographical recordingsthat only the superficial red muscle of the trout is active during low speed swimming(Hudson, 1973). The mosaic muscle in contrast only becomes active at intermediatecruising speeds and during bursts of activity (Webb, 1970; Hudson, 1973).
Studies on the energetics of mammalian muscles with different speeds of con-traction have shown that they have different thermodynamic efficiencies with respectto the amount of work done per unit of high energy phosphate utilized (Awan et al.,1972). Since the superficial and mosaic red fibres of the trout differ in myofibrillarA TPase and hence, speed of shortening, it is likely that they will also have verydifferent energetic properties. This finding is of particular relevance to studies on theenergetics and efficiency of swimming in this species, since previous studies (Webb,1971) have assumed that the two types of red myotomal fibre are equivalent.
I. A. Johnston is grateful to the Natural Environmental Research Council for a post-doctoral fellowship and to Professor G. M. Hughes for his encouragement.

457TROUT MUSCLE FIBRES
References
Awan, M. Z., Frearson, N., Goldspink, G. & Waterson, S. W. (1972). Biochemical efficiencyof smooth muscle and different fibre types of striated muscle. J. Mechanochem. CellMotility 1, 225-232.
Barany, M. (1967). ATPase activity of myosin correlated with speed of muscle shortening.J. gen. Physiol. 50, 197-219.
Boddeke, R., Slijper, E. J. & van der Stelt, A. (1959). Histological characteristics of the bodymusculature of fishes in connection with their mode of life. Koninklijke ned. Akad.Wetenscappen Ser. C 62, 576-588.
Bone, Q. (1966). On the function of the two types ofmyotomal muscle fibre in elasmobranchfish. J. mar. bioI. Ass. U.K. 46, 321-349.
Bostrom, S. L. & Johansson, .R. G. (1972). Enzyme activity patterns in white and redmuscles of the eel (Anguilla anguilla) at different developmental stages. Comp.Biochem. Physiol. 42B, 533-542.
Brierley, G. P., Murer, E. & Bachman, E. (1964). Studies on ion transport III. The accumu-lation of calcium and inorganic phosphate by heart mitochondria. Archs Biochem.Biophys. 105, 89-102.
Brotchi, J. (1968). Identification histoenzymologique des fibres lentes et rapides dans lesmuscles squellettiques des vertebretes. Archs int. Physiol. Biochim. 76, 299-310.
Close, R. I. (1972). Dynamic properties of mammalian skeletal muscles. Physiol. Rev. 52,129-310.
George, J. C. (1962). A histophysiological study of the red and white muscles of the mackerel.Am. MidI. Nat. 68, 487-494.
George, J. C. & Bokadawala, F. D. (1964). Cellular organization and fat utilization in fishmuscle. J. Anim. Morph. Physiol. 11, 124-132.
Greene, F. C. & Greene, C. H. (1913). The skeletal musculature of the king salmon. Bull.Bur. Fish., Wash. 33, 25-59.
Guth, L. (1973). Fact and artifact in the histochemical procedure for myofibrillar ATPase.Exptl Neurol. 41, 440-450.
Guth, L. & Samaha, F. J. (1969). Qualitative differences between actomyosin ATPase dfslow and fast mammalian muscle. Exptl Neurol. 25, 138-152.
Guth, L. & Samaha, F. J. (1970). Procedure for the histochemical demonstration of acto-myosin ATPase. Exptl Neurol. 28,365-367.
Hamoir, G., Focant, B. & Disteche, M. (1972). Proteinic criteria of differentiation of white,cardiac and various red muscles in the carp. Comp. Biochem. Physiol. 41B, 665-674.
Hudson, R. C. L. (1973). On the function of the white muscles in teleosts at intermediateswimming speeds. J. exp. BioI. 58, 509-522.
Johnston, I. A. (1973). Biochemical and physiological studies on the swimming muscles ofteleosts. Ph.D. Thesis, University of Hull, England.
Johnston, I. A., Frearson, N. & Goldspink, G. (1972). Myofibrillar ATPase activities of thewhite and red muscles of marine fish. Experentia 28, 713-714.
Johnston, I. A., Frearson, N. & Goldspink, G. (1974a). The effects of environmentaltemperature on the properties of myofibrillar adenosine triphosphatases from variousspecies of fish. Biochem. J. 133, 735-738.
Johnston, I. A., Patterson, S., Ward, P. S. & Goldspink, G. (1974b). The histochemicaldemonstration of myofibrillar adenosine triphosphatase activity in fish muscle. Can.J. Zool. 52, 871-877.
Matsuura, F. & Hashimoto, K. (1954). Chemical studies on the red muscle (' chai ') ofgoldfishes. II. Determinations of the contents of haemoglobin, myoglobin, andcytochrome c in the muscles of fishes. Bull. Jap. Soc. scient. Fish. 20, 308-312.
Nachlas, M. M., Tsou, K. C., De Sonza, E., Cheng, C. S. & Seligman, A. M. (1957). Cyto-chemical demonstration of succinic dehydrogenase by the use of anew p-nitrophenylsubstituted ditetrazale. J. Histochem. Cytochem. 5, 420-436.
Nag, A. C. (1972). Ultrastructure and adenosine triphosphatase activity of red and whitemuscle fibres of the caudal region of a fish, Salmo gairdneri. J. Cell BioI. 55, 42-57.
Ogata, T. & Mori, M. (1964). Histochemical study of oxidative enzymes in vertebratemll~rl,,~ -' H;"tnrhpm rvtnrhpm 12- 17.1-1R2

458 I. A. JOHNSTON ET AL.
Padykula, H. A. & Herman, E. (1955). The specificity of the histochemical method foradenosine triphosphatase. I. Histochem. Cytochem.3, 110-183.
Pearse, A. G. E. (1960). In Histochemistry, 2nd edn. Edinburgh & London: ChurchiIlLivingstone.
Revel, J. P. (1962). The sarcoplasmic reticulum of the bat cricothyroid muscle. I. Cell Bioi.12, 571-588.
Schiaffino, S. & BormioIli, S. P. (1973). Histochemical characterisation of adenosine tri-phosphatases in skeletal muscle fibres by selective extraction procedures. I. Histochem.Cytochem. 21, 140-145.
Syrovy, I., Gaspar-Godfroid, A. & Hamoir, G. (1970). Comparative study of the myosinsfrom red and white muscles of the carp. Archs Int. Physiol. Biochim. 78,919-934.
Walker, M. G. (1970). Growth and development of the skeletal muscle fibres of the cod(Gadus morhua L:). I. Cons. perm. int. Explor. Mer 33, 228-244.
Webb, P. W. (1970). Some aspects of the energetics of swimming offish with special referenceto the cruising performance of rainbow trout Salmo gairdneri Richardson. Ph.D.Thesis, University of Bristol.
Webb, P. W. (1971). The swimming energetics of trout II. Oxygen consumption andswimming efficiency. I. exp. Bioi. 55, 521-540.
Printed in Great Britain by Henry Ling Ltd.. at the Dcrset Press. Dorchester. Dorset




















