
Revision of the genus Coletinia (Zygentoma: Nicoletiidae) in the Iberian Peninsula, with...
60
Accepted by E. Bernard: 6 Dec 2012; published: 15 Feb. 2013 ZOOTAXA ISSN 1175-5326 (print edition) ISSN 1175-5334 (online edition) Copyright © 2013 Magnolia Press Zootaxa 3615 (1): 001–060 www.mapress.com/ zootaxa/ Monograph http://dx.doi.org/10.11646/zootaxa.3615.1.1 http://zoobank.org/urn:lsid:zoobank.org:pub:D53E1122-AA33-4152-90BE-3D717979C648 ZOOTAXA Revision of the genus Coletinia (Zygentoma: Nicoletiidae) in the Iberian Peninsula, with descriptions of nine new species RAFAEL MOLERO 1 *, CARMEN BACH 2 , ALBERTO SENDRA 3 , SERGIO MONTAGUD 4 , PABLO BARRANCO 5 & MIGUEL GAJU 1 1 Departamento de Zoología, Universidad de Córdoba, Edificio C-1 (Darwin), Campus de Rabanales. 14071 Córdoba, Spain. e-mail: [email protected] 2 Departamento de Biología Animal, Vegetal y Ecología. Facultad de Ciencias. Universidad Autónoma de Barcelona. 08193 Bellaterra (Barcelona), Spain 3 Departamento de Zoología y Antropología Física, Universidad de Alcalá, Alcalá de Henares, Spain. 4 Museu Valencià d’Història Natural (Fundación Entomológica Torres Sala). Paseo de la Pechina 15. 46008 Valencia, Spain 5 Dpto. Biología Aplicada. CITE II-B. Universidad de Almería.04120 Almería *Corresponding author Magnolia Press Auckland, New Zealand 3615 TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Transcript of Revision of the genus Coletinia (Zygentoma: Nicoletiidae) in the Iberian Peninsula, with...

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
ZOOTAXAISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2013 Magnolia Press
Zootaxa 3615 (1): 001–060 www.mapress.com/zootaxa/ Monograph
http://dx.doi.org/10.11646/zootaxa.3615.1.1http://zoobank.org/urn:lsid:zoobank.org:pub:D53E1122-AA33-4152-90BE-3D717979C648
ZOOTAXA
Revision of the genus Coletinia (Zygentoma: Nicoletiidae) in the Iberian Peninsula, with descriptions of nine new species
RAFAEL MOLERO1*, CARMEN BACH2, ALBERTO SENDRA3, SERGIO MONTAGUD4, PABLO BARRANCO5 & MIGUEL GAJU1
1 Departamento de Zoología, Universidad de Córdoba, Edificio C-1 (Darwin), Campus de Rabanales. 14071 Córdoba, Spain. e-mail: [email protected]
2 Departamento de Biología Animal, Vegetal y Ecología. Facultad de Ciencias. Universidad Autónoma de Barcelona. 08193 Bellaterra (Barcelona), Spain
3 Departamento de Zoología y Antropología Física, Universidad de Alcalá, Alcalá de Henares, Spain.4 Museu Valencià d’Història Natural (Fundación Entomológica Torres Sala). Paseo de la Pechina 15. 46008 Valencia, Spain
5 Dpto. Biología Aplicada. CITE II-B. Universidad de Almería.04120 Almería*Corresponding author
Magnolia PressAuckland, New Zealand
3615
Accepted by E. Bernard: 6 Dec 2012; published: 15 Feb. 2013

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
RAFAEL MOLERO, CARMEN BACH, ALBERTO SENDRA, SERGIO MONTAGUD, PABLO BARRANCO & MIGUEL GAJURevision of the genus Coletinia (Zygentoma: Nicoletiidae) in the Iberian Peninsula, with descriptions of nine new species(Zootaxa 3615)
60 pp.; 30 cm.
15 Feb. 2013
ISBN 978-1-77557-106-3 (paperback)
ISBN 978-1-77557-107-0 (Online edition)
FIRST PUBLISHED IN 2013 BY
Magnolia Press
P.O. Box 41-383
Auckland 1346
New Zealand
e-mail: [email protected]
http://www.mapress.com/zootaxa/
© 2013 Magnolia Press
All rights reserved.
No part of this publication may be reproduced, stored, transmitted or disseminated, in any form, or by any
means, without prior written permission from the publisher, to whom all requests to reproduce copyright
material should be directed in writing.
This authorization does not extend to any other kind of copying, by any means, in any form, and for any purpose
other than private research use.
ISSN 1175-5326 (Print edition)
ISSN 1175-5334 (Online edition)
MOLERO ET AL.2 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Table of contents
Abstract . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4Material and methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5Results . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
Anatomic characters with taxonomic relevance in Coletinia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 61. General characters . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6
1a. Body length and shape, antennae and terminal filament length . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 61b. Pigmentation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 61c. Spiralization of macrosetae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 61d. Chaetotaxy of the head. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 81e. Tibial spine size and arrangement . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 81f. Length/width ratio (L/W) of tibiae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 81g. Shape of the posterior border of urotergite X . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 111h. Arrangement of setae on the disc of urotergite X . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 111i. Presence of discal macrosetae on urosternites. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
2. Male characters. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 112a. Apophyses of the pedicellus. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 112b. Shape of the posterior border of urosternite VIII . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 112c. Paramere length . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 112d. Number, position and shape of sensory pegs on urotergite X . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 112e. Presence, number and shape of sensory pegs or spines in terminal filaments . . . . . . . . . . . . . . . . . . . . . . 12
3. Female characters . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 123a. Shape of the subgenital plate . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 123b. Length and number of divisions of the ovipositor . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12
Preliminary phylogenetic groupings . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 121. Group “asymetrica” . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 122. Group “mendesi” . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 123. Group “capolongoi” . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 144. Group “maggii” . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
Descriptions of new species . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15Coletinia herculea n. sp. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15Coletinia vergitana n. sp. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19Coletinia calaforrai n. sp. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24Coletinia intermedia n. sp. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27Coletinia diania n. sp. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29Coletinia longitibia n. sp. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32Coletinia tessella n. sp. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35Coletinia redetecta n. sp. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 38Coletinia hernandoi n. sp. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
Previously known species . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43Coletinia asymetrica Bach de Roca, Mendes & Gaju Ricart . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43Coletinia mendesi Wygodzinsky . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43Coletinia capolongoi Wygodzinsky . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47Coletinia tinauti Molero-Baltanás, Gaju-Ricart & Bach de Roca . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49Coletinia maggii (Grassi) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 52Coletinia sp. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 52
Discussion and general conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 541. Species of the “asymetrica group” . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 542. Species of the “mendesi group” . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 543. Species of the “capolongoi group” . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 554. Species of the “maggii group” . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 55
Identification key for Iberian Coletinia species. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 57Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58About the authors. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 60
Zootaxa 3615 (1) © 2013 Magnolia Press · 3REVISION OF IBERIAN COLETINIA

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Abstract
The discovery of several members of the genus Coletinia Wygodzinsky, 1980, from subterranean habitats (endogean and troglobiont), prompted the review of this genus in the Iberian Peninsula. Most of the samples came from caves of the Mediterranean basin of Spain, from Cádiz to the Tarragona province. As a result of this revision, nine new species have been established: C. herculea n. sp., an endogean from Cádiz; C. vergitana n. sp. from the Gádor calcareous mountains in Almería; C. calaforrai n. sp. from the gypsum karst in Almería; C. intermedia n. sp. from caves in Murcia and Alicante; C. diania n. sp., found in the north of the province of Alicante; C. longitibia n. sp. and C. tessella n. sp., both troglobites from Valencia; C. redetecta n. sp. from Castellón caves and finally C. hernandoi n. sp., an endogean from Tarragona. Moreover, Coletinia maggii (Grassi, 1887) is reported for the first time in the Iberian Peninsula, and new data are presented regarding C. mendesi, C. tinautiand C. capolongoi that widen their geographic distribution and enhance the information about their anatomic characteristics and biology. These results increase the number of known species of this genus to 14 in the region and to 21 in the world. The new species are described and compared with the most closely related previously known species of the genus. Characters with the most taxonomic relevance are discussed using optical and scanning microscope studies. A key for the identification of the Iberian Coletinia species and a distribution map including all of them are also provided.
Key words: Coletiniinae, Thysanura, Spain, taxonomy, identification key, endogean fauna, troglobitic fauna
Resumen El hallazgo de numerosas muestras del género Coletinia Wygodzinsky, 1980, integrado por especies subterráneas (endogeas y troglobias), permite la revisión de dicho género en la Península Ibérica. La mayoría de estas muestras proceden de cuevas situadas en provincias mediterráneas, desde Cádiz a Tarragona. Como resultado de esta revisión, se describen nueve nuevas especies: C. herculea n. sp., endogea de Cádiz; C. vergitana n. sp. procedente de la sierra de Gádor en Almería; C. calaforrai n. sp. del karst en yesos de Almería; C. intermedia n. sp., de cuevas de Murcia y Alicante; C. diania n. sp., encontrada en el norte de la provincia de Alicante; C. longitibia n. sp. y C. tessella n. sp., ambas troglobias de Valencia; C. redetecta n. sp., de cuevas de Castellón, y finalmente C. hernandoi n. sp., endogea de Tarragona. Además, C. maggii (Grassi, 1887) se cita por primera vez en la Península Ibérica, y también se aportan nuevos datos sobre C. mendesi, C. tinauti y C. capolongoi que amplían su distribución geográfica y la información disponible sobre sus caracteres anatómicos y biología. Estos resultados incrementan el número de especies conocidas de este género hasta 14 en el área ibérica y hasta 21 a nivel mundial. Las nuevas especies se describen y comparan con las previamente conocidas y más estrechamente relacionadas del género. Se discuten los caracteres con mayor interés taxonómico, utilizando estudios tanto de microscopía óptica como de microscopio electrónico de barrido. Se proporciona también una clave para la identificación de las Coletinia ibéricas y un mapa de distribución de las mismas.
Introduction
The genus Coletinia Wygodzinsky, 1980 belongs to the family Nicoletiidae (order Zygentoma = Thysanura s. str.) and includes species from subterranean environments collected in the Southwest Palaearctic region; one species has also been described from Brazil (Mendes & Ferreira, 2002). Prior to this study, 12 species belonging to this genus were known, 4 of which were endemic to the Iberian Peninsula (continental Portugal and Spain): C. mendesiWygodzinsky, 1980, C. capolongoi Wygodzinsky, 1980, C. asymetrica Mendes et al. 1985 and C. tinauti Molero et al., 1997. Additional data about the previous knowledge of these 12 species are presented in Table 1.
Difficulties in canvassing the habitats (endogean medium and caves) where these insects occur have resulted in a scarcity of knowledge about this genus. Fortunately, samples recently provided by Spanish biospeleologic teams have increased the number of specimens of Nicoletiidae available for taxonomic and biologic research on these poorly known thysanurans. As a result of the study of these specimens, nine new species and new faunistic data are presented in this work.
MOLERO ET AL.4 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
TABLE 1. Previously known species of Coletinia, including geographic distribution and remarks. Species marked with * are only known from original description. The references column includes only those that are additional to their original description and to the revision of the genus reported by Wygodzinsky (1980).
Material and methods
Specimens were collected directly with an entomological aspirator or indirectly using traps. These insects were most commonly found in caves or in the soil, often under large stones. The sublithic specimens sometimes shared their habitat with social insects such as ants or termites. Once captured, the insects were immediately fixed with 70º alcohol and labelled in vials. Most of the specimens were dissected for microscopic studies and then, either all the entire exoskeletons were mounted in Tendeiro medium for optical microscope (OM) study, or the specimens were prepared for scanning electron microscope (SEM) studies. Tendeiro medium was prepared following the protocol described in Molero et al. (2000). For SEM studies, selected pieces of the insects were dehydrated with absolute alcohol and finally with hexamethyldisilazane (following Ubero-Pascal et al. 2005); afterwards they were coated with gold.
Length-to-width ratios are indicated as L/W.The studied material has been deposited in the following institutions: Museo Nacional de Ciencias Naturales, Madrid, Spain (MNCN); Museu Valencià d’Història Natural (Fundación Entomológica Torres Sala), Valencia, Spain (MVHN); Departamento de Zoología, Universidad de Córdoba, Córdoba, Spain (UCO); Centro de Zoología, IICT, Lisbon, Portugal. (CZ)
Species Distribution Remarks References
C. maggii (Grassi, 1887) Central and southern Europe
Described from Sicily, widespread in Europe
Parona, 1888; Grassi, 1887; Mendes, 1981; Mendes, 1992; Mendes, 1996; Paclt & Christian, 1996
C. subterranea (Silvestri, 1902) Continental Italy, Sicily, Sardinia
Escherich, 1905; Silvestri, 1908; Mendes, 1992
C. corsica (Chopard, 1924)* Corsica Insufficient original description; known from the female only
C. jeanneli (Silvestri, 1938)* Southern France (Var Department)
C. bulgarica (Kozaroff, 1939)* Bulgaria
C. capolongoi Wygodzinsky, 1980*
Eastern Spain (Valencia Province)
C. mendesi Wygodzinsky, 1980 Southern Portugal, southwestern Spain
Bach et al., 1985; Mendes, 1985; Mendes, 1996
C. setosula Wygodzinsky, 1980* Sicily
C. asymetrica Bach de Roca, Mendes & Gaju Ricart, 1985*
Southern Spain (Córdoba Province)
C. tinauti Molero-Baltanás, Gaju-Ricart & Bach de Roca, 1997
Southern Spain (Jaén and Córdoba Provinces)
Troglobic and edaphic
C. longissima Mendes, 1988 Turkey Mendes, 1992
C. brasiliensis Mendes & Ferreira, 2002*
Brazil (!)
Zootaxa 3615 (1) © 2013 Magnolia Press · 5REVISION OF IBERIAN COLETINIA

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Results
Our results can be divided into five sections: a) remarks about the anatomic characteristics considered for the taxonomic studies within the genus Coletinia, concluding that at least four groups of species can be distinguished; b) descriptions of nine new species; c) remarks about the remaining Spanish species, more extensive for those where new material is available and new faunistic data are provided; d) a discussion justifying the establishment of these new species, which ends with some general conclusions and a distribution map that summarizes the updated faunistic knowledge of the genus in the Iberian Peninsula; and e) a key for the identification of the Iberian species.
Anatomic characters with taxonomic relevance in Coletinia
The structures that deserve the most taxonomic interest in Coletinia were determined to be the following:
1. General characters. This section includes those characteristics shared by both sexes.
1a. Body length and shape, antennae and terminal filament length. The maximum body length in both sexes is an interesting characteristic because some small species are less than 10 mm and some large ones are greater than 15 mm. As Nicoletiidae are ametabolous insects, it could be argued that the small specimens correspond to immature specimens and could grow to reach a larger size. To solve this problem, it must be determined whether a given specimen is an adult. Adult Coletinia males can be recognized by the well-developed sensory pegs of the urotergite X and terminal filaments; the antennal apophyses and their glandular cone must also be well differentiated. Adult Coletinia females show a well-developed ovipositor.
Although all Coletinia species have long, subcylindrical bodies, strict troglobitic forms manifest narrower bodies and longer antennae and terminal filaments, which are usually longer than the body length, whilst other species found more frequently as edaphic forms are relatively wide and have shorter appendages (Fig. 1). Therefore, it is interesting to know the thorax width and the maximum length preserved of the antennae, cerci and paracercus. Unfortunately, these appendages are very often broken.
1b. Pigmentation. The integument of these insects contains whitish or yellowish pigments. Apart from the tonality, the pigment concentration varies; thus, several species are relatively transparent while others have a more intense tone. The darker colorations observed in some species (the abdomen is usually darker than the head and thorax) are due to gut contents.
1c. Spiralization of macrosetae. This is a new taxonomic character that has not been reported previously in Coletinia species. In many species, most macrosetae, especially those inserted in the head and appendages (such as coxae, antennal pedicels, palps) show a strong tendency to twist over their own longitudinal axis. This twisting is evident in the spiral curving of the striated surface and appears most clearly in specimens mounted in Tendeiro medium (Fig. 2). Herein, this twisting is called spiralization. The species with the most pronounced spiralization is Coletinia redetecta n. sp., which bears macrosetae with 8 or more turns (Fig. 2A). Additional species, such as C. tessella n. sp., C. longitibia n. sp. and C. capolongoi (Fig. 2B) exhibit a moderate degree of macrosetae spiralization, and some others, such as C. tinauti, C. intermedia n. sp. and C. calaforrai have macrosetae with a straighter striation, with at most 1–2 turns (Fig. 153).
It can be argued that this spiralization is caused by the mounting medium (the curving of the striated surface of the setae is evident in SEM photographies, but less intense) and can be interpreted as an artifact of the protocol. Nevertheless, the degree of twisting of macrosetae is independent from the batch of Tendeiro liquid, but is variable depending on the species (it is similar in different specimens of the same species). Unfortunately, there are not enough specimens of those species with a greater degree of spiralization to enable SEM studies. In any event, we think that the differences in the spiralization correspond to different taxa and that including this character in the descriptions of species is interesting when Tendeiro medium is used (and prepared as indicated in the section of methods, the more usual protocol among Zygentoma specialists). If any entomologist in the future uses a different technique of preservation, we do not know if this character could be taken into account. Thus, the use of the spiralization in the identification keys of the last section of this work is very limited (only included as auxiliary for distinguishing some species whose females are very similar), but is indicated in the description of all the species.
MOLERO ET AL.6 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURE 1. Photographs of living specimens of Iberian Coletinia. A. C. mendesi (photograph by R. Molero). B. C. tinauti(photograph by M. Gaju). C. C. redetecta n. sp. (photograph by S. Montagud). Scales: 2 mm.
Zootaxa 3615 (1) © 2013 Magnolia Press · 7REVISION OF IBERIAN COLETINIA

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURE 2. OM photographs of coxal spiralized macroseta showing the different degrees of twisting of the tegument. A. Coletinia redetecta n. sp. B. C. capolongoi. Scales: 50 µm.
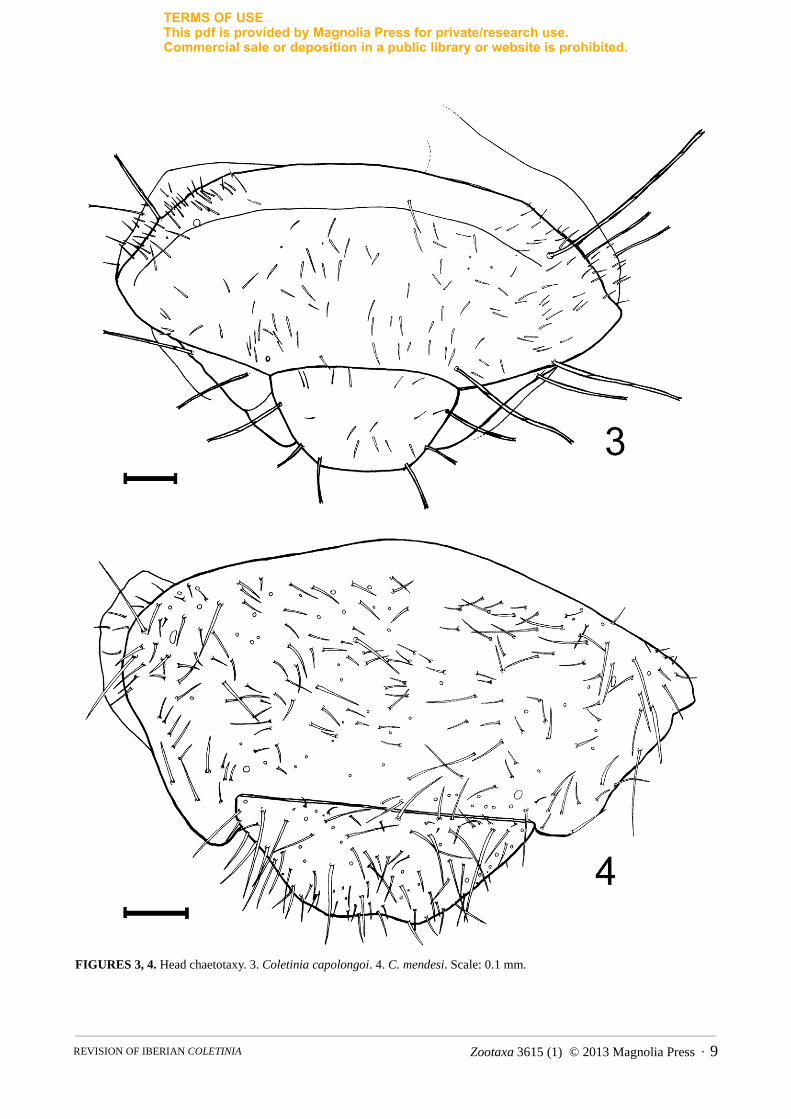
1d. Chaetotaxy of the head. In the genus Coletinia two different types of cephalic setation have been reported. Most species have scattered, short hairs and a few regularly arranged macrosetae with bifid apex (in the frons, these macrosetae are inserted mainly along the borders, as illustrated in Fig. 3). However, in a small group of species that includes C. mendesi, the head bears numerous subequal strong setae and some small and dispersed setae, but bifid macrosetae are absent (Fig. 4).
1e. Tibial spine size and arrangement. Coletinia species usually show acute spines inserted on the tibiae. These spines are normally slender and shorter than or as long as the diameter of the article. Some of them are inserted in dorsal position of the tibia (usually 1–3, in subbasal, medial or subdistal positions). A higher number (usually four bifid spines) are found in ventral position: two are inserted mediobasally very close to each other, and two are inserted subdistally (also very close). In some species, additional basal spines are present on the ventral side. A lateral short spine is also inserted in the basal half of the tibia (Fig. 5). Although some intraspecific variability could be detected, the size, number and arrangement of these spines appear to be species-specific characteristics. The most evident differences were observed in C. mendesi, whose tibial spines are robust and long (in the mesotibia, their length is more than the diameter of the article) and which lacks lateral spines (Fig. 6). The high number of ventral bifid spines in C. redetecta n. sp. (Figs. 105–107) is also characteristic.
1f. Length/width ratio (L/W) of tibiae. This characteristic is very useful for distinguishing between many species, especially the parameter of the length/width ratio of the metatibiae. Although there is some intraspecific variability, the differences between related species can be very clear. Juvenile specimens manifest shorter tibiae, so these parameters must be measured in adults. Larger species usually have longer tibiae, and it seems clear that, within the same species, the L/W ratio of the tibiae is higher in longer specimens. However, in the Iberian fauna there are large species with relatively short tibiae, such as C. capolongoi, and small species with relatively narrow tibiae, such as C. calaforrai n. sp.
MOLERO ET AL.8 · Zootaxa 3615 (1) © 2013 Magnolia Press
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 3, 4. Head chaetotaxy. 3. Coletinia capolongoi. 4. C. mendesi. Scale: 0.1 mm.
Zootaxa 3615 (1) © 2013 Magnolia Press · 9REVISION OF IBERIAN COLETINIA

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 5, 6. SEM photographs of Coletinia tibiae. 5. Tibia III of C. capolongoi, showing the typical position of the tibial spines: d: dorsal; l: lateral; v: ventral. 6. Tibia II of C. mendesi, without lateral spines and with long and stronger ventral spines. Scales: 0.2 mm.
MOLERO ET AL.10 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
For allometric growth studies of specimens preserved in alcohol, it is preferable to use the length of the metatibia as indicative of the size of the insect, because intersegmental membranes can extend in these specimens.
1g. Shape of the posterior border of urotergite X. This urotergite shows sexual dimorphism. In males, two posterolateral lobes are developed in all species, but the shape of the posterior border between these lobes can be more or less straight (Fig. 147) or curved (concave), as in Fig. 46. In females, the posterior border can be slightly concave or straight depending on the species (Figs. 111 and 167, respectively). Because the integument in the median area of this urotergite is very fragile, the posterior border sometimes appears broken in microscopic slides, and this shape is not well visible.
1h. Arrangement of setae on the disc of urotergite X. Setae covering the surface of this urotergite can be sparse or dense and uniformly or irregularly distributed over the disc depending on the species. When there are few setae, these are often concentrated in the posterolateral areas of the urotergite (Fig. 159). Moreover, these discal setae can be more or less strong.
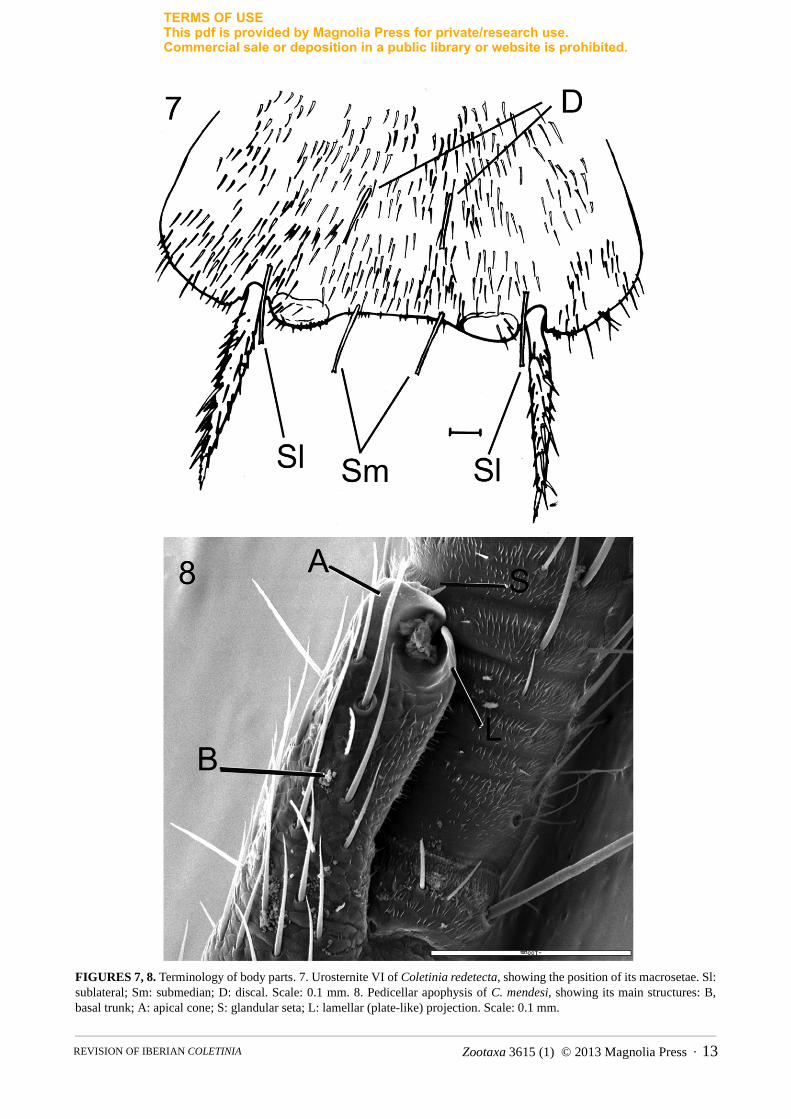
1i. Presence of discal macrosetae on urosternites. All Coletinia species studied bear 2+2 macrosetae inserted in the hind border of the urosternites; one pair is inserted internally to the insertion of the stylets, named here the sublateral macrosetae (“posterosublateral” in Wygodzinsky 1980), and one pair is inserted near the exsertile vesicles, designated the submedian macrosetae in this work (“posterosubmedian” in Wygodzinsky’s nomenclature). Moreover, many species have 1+1 macrosetae inserted into the median position of the disc, termed here the discal macrosetae and called “anteromedian” by Wygodzinsky (Fig. 7); the presence of discal macrosetae is usual, but they are absent in some species such as C. mendesi.
2. Male characters2a. Apophyses of the pedicellus. This is one of the best characteristics for distinguishing between species.
Males bear one apophysis whose shape and particularities are exclusive to each species, which is usually in dorsointernal position on basal trunk of the pedicellus of each antenna. In a typical apophysis, we can distinguish a glandular cone, recognizable because it bears a glandular seta on top (Figs. 8, 143). The simplest apophysis, such as that of C. maggii, has a cylindrical shape with the aforementioned glandular cone in the apex (Fig. 164), and several (usually 5) long, bifid macrosetae inserted basally, similar to the 5 macrosetae that females usually bear in the pedicellus. However, in different species some modifications of this basic pattern can be detected, such as the presence of lateral projections with different shape and development (plate-like, hook-shaped, etc.), specialized sclerotized zones of the tegument or additional setae (Figs. 11–12, 28–29, 128). Moreover, some species have symmetrical antennae, while in others, the apophysis of the left pedicellus is more developed and contains different modifications than the right (curiously, all species known are “left-handed”!). The three-dimensional structure of these apophyses must be observed carefully in microscopic slides because similar antennae placed in different positions could be interpreted as different shapes, but upon rotating the antennae, similar structures can be recognized. This problem can be solved (when enough material is available) with the scanning electron microscope (SEM), where an antenna can be observed from different points of view (see Figs. 41A–D, 134–136, 141–143 or 155–157). Finally, the size of the apophysis can be used as a taxonomic character. In this work the level of the flagellum that the top of the apophysis reaches has been indicated in the descriptions, but conclusions on this character should be taken with caution since probably the size of the apophysis grows over successive moults.
2b. Shape of the posterior border of urosternite VIII. In males, the posterior border of this urosternite usually protrudes between stylets VIII, but its shape can range between straight and truncated (Fig. 97 or Fig. 137) and convex and rounded (Fig. 47). In some species, this shape is constant, and in others, it appears to be variable.
2c. Paramere length. Two parameters can be taken into account: the length/width ratio of the parameres (short in C. diania n. sp., C. mendesi or C. tinauti as shown in Figs. 72, 138 and 161, and long in C. capolongoias shown in Fig. 150) and the “paramere length/stylets IX length” ratio.
2d. Number, position and shape of sensory pegs on urotergite X. The setation of the tenth urotergite of males is modified: some setae, usually some inserted in the lateral margins, are transformed into sensory pegs with blunt apexes. The shape of these pegs seems to be variable; for example, upon studying C. capolongoi by optical microscopy, Wygodzinsky (1980) described the pegs as bulb-shaped. Nevertheless, SEM studies reveal
Zootaxa 3615 (1) © 2013 Magnolia Press · 11REVISION OF IBERIAN COLETINIA

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
a relatively homogeneous structure in all of the species observed (see Fig. 9). All of the pegs have a striated tegument that curves in the apex; all are cylindrical in shape and have an apical furrow where the tegument grooves finish. Interspecific differences in length cannot be considered important, because intraspecific (and even individual) variability in this parameter has been detected. However, the number and position of these pegs seems to be species-specific (although intraspecific variability has also been detected).
2e. Presence, number and shape of sensory pegs or spines in terminal filaments. Coletinia species usually have sensory pegs (absent in C. brasiliensis) with striated tegument (Fig. 10) in the base of the cerci and paracercus. In some species, these pegs are absent in the paracercus, as found in some specimens belonging to C. tinauti or C. redetecta n. sp. Some species have few pegs (1–3) in the cerci, while some others bear a higher number (4 or more). The shape of these pegs is also variable; they can be short and blunt, similar to those of the tenth urotergite, or longer and more acute in other species, as shown in Fig. 10.
3. Female characters3a. Shape of the subgenital plate. The median urosternite of the eighth segment in females has a variable
shape; it can be more or less semicircular, semielliptical or subtriangular. The main characteristics useful for comparing species are the length/width at the base ratio and the shape of the posterior border of the sclerite, which can range from straight (truncate, as in Fig. 162) to convex (rounded, as in Fig. 87). In some species, such as C. calaforrai n. sp. and C. intermedia n. sp., intraspecific variability has been detected (see below).
3b. Length and number of divisions of the ovipositor. Some Coletinia species bear a long and thin ovipositor with many divisions (more than 20), while others have shorter and stouter gonapophyses with fewer divisions. Apart from this characteristic, the most useful parameter is the ratio between the length of the portion of the ovipositor that surpasses the tip of the ninth stylets and the length of these stylets.
Preliminary phylogenetic groupings
Analysis of all of these characteristics has led to the discrimination of four groups within the genus Coletinia for practical purposes. A subgeneric division with a phylogenetic sense of the genus requires a deep redescription of the species from other countries where no well-preserved material is available. Furthermore, molecular studies and cladistic analysis of the morphologically variable features are also needed. Nevertheless, a hypothesis about the phylogenetic relationships of the following groups is proposed:
1. Group “asymetrica”. This group is typified by asymmetric antennae in males; in all species observed, with the left antenna exhibiting a more developed apophysis than the right. The posterior border of the eighth urosternite in Iberian species is convex and protrudes to varying extents; it is straight only in C. subterranea(Silvestri, 1902) from Italy. The “asymetrica” group includes three previously known species: the aforementioned Italian species, C. asymetrica from Spain and C. brasiliensis from Brazil. These three species appear to be phylogenetically less interrelated than the species in the other groups. Two new species belonging to this group are described in this work, C. vergitana n. sp. and C. herculea n. sp.; both are likely more closely related to C. asymetrica than to the other species of the group, which probably had independent origins from a symmetrical ancestor similar to C. maggii. Nevertheless, all of the species coincide in that the reduced apophysis occurs on the right side.
2. Group “mendesi”. This group is characterized by the setation of the head, with numerous subequal strong setae and without bifid strong macrosetae (Fig. 4). The posterior border of the urosternite VIII is truncated and straight in males (Fig. 137). Antennae of the male are symmetrical and the apophyses have subapical plate-like structures. This group includes only two species: C. mendesi from Portugal and Spain and C. setosulaWygodzinsky (1980) from Sicily (Italy). These species likely share other characteristics, such as the absence of discal macrosetae in the urosternites and lateral spines in the tibiae, but these features were not provided in the original description of C. setosula and the type of this species has not been re-examined. Many characteristics exclusive to both species are likely to be apomorphic, and there is most likely a close relationship between this group and the Iberian species of the subgroup “capolongoi” (see below), because of the similarities in the configuration of the antennal apophyses in males.
MOLERO ET AL.12 · Zootaxa 3615 (1) © 2013 Magnolia Press
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 7, 8. Terminology of body parts. 7. Urosternite VI of Coletinia redetecta, showing the position of its macrosetae. Sl: sublateral; Sm: submedian; D: discal. Scale: 0.1 mm. 8. Pedicellar apophysis of C. mendesi, showing its main structures: B, basal trunk; A: apical cone; S: glandular seta; L: lamellar (plate-like) projection. Scale: 0.1 mm.
Zootaxa 3615 (1) © 2013 Magnolia Press · 13REVISION OF IBERIAN COLETINIA

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 9, 10. SEM photographs of sensory pegs in Coletinia. 9. Pegs of urotergite X from several species: A, C. capolongoi; B, C. tinauti; C: C. intermedia n. sp. Scales: 5, 10 and 9 µm, respectively. 10. Sensory peg of the cercus of C. capolongoi. Scale: 10 µm.
3. Group “capolongoi”. This group includes species with a cephalic chaetotaxy similar to the “maggii” and “asymetrica” groups (Fig. 3). Antennal apophyses of males have subapical platelike structures in addition to the apical cone, as in C. capolongoi (see typical structure in Fig. 8). The tenth urotergite of females usually has a slightly concave posterior border. The eighth urosternite of males of most species possesses a convex posterior border. Within this group, there appears to be a tendency in the north of its distribution area toward increased body size and increased degree of macroseta spiralization. This is the most diversified subgroup of the Iberian fauna. Eight species can be included in this group, 2 previously known (C. capolongoi and C. tinauti) and 6 newly described species: C. calaforrai n. sp., C. intermedia n. sp., C. diania n. sp., C. longitibia n. sp., C. tessella n. sp. and C. redetecta n. sp. These 8 species show a relatively homogeneous configuration of the antennal apophysis, indicating a very recent phylogenetic relationship. Some other Mediterranean species, such as C. jeanneli (Silvestri, 1938) from southern France, C. bulgarica (Kozaroff, 1939) from Bulgaria and C. longissima Mendes, 1988, from Turkey, are most likely related to this group but their different antennal processes (especially in the Turkish species) suggest an independent origin from an ancestor with a simple apophysis, such as C. maggii.
MOLERO ET AL.14 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
4. Group “maggii”. Males of these species bear symmetrical and simple subcylindrical apophyses without lamellar projections. The head has two very different types of setae: few macrosetae and a variable number (usually scarce) of short, scattered bristles (as in Fig. 3). In Spain this group is represented by C. maggii (reported here for the first time in the Iberian fauna) and C. hernandoi n. sp. Coletinia maggii is most likely the most plesiomorphic species within the genus.
Descriptions of new species
Coletinia herculea Molero, Bach & Gaju, new speciesFigs. 11–26
Studied material: Cádiz: Algeciras, Mirador del Estrecho, 12 January 2007, A. Tinaut leg., male holotype deposited in MNCN, type number 2236, and 1 subadult male paratype (deposited in UCO, ref. Z2170).
Description. Body length of the holotype 8.5 mm, subadult, 6.2 mm. Antennae broken, maximum preserved length 6.5 mm. Terminal filaments broken, a cercus with only 1.5 mm preserved. General body shape campodeiform, with yellowish pigment uniformly distributed, slightly more intense in the posterior part. Head with scattered short hairs and few macrosetae.
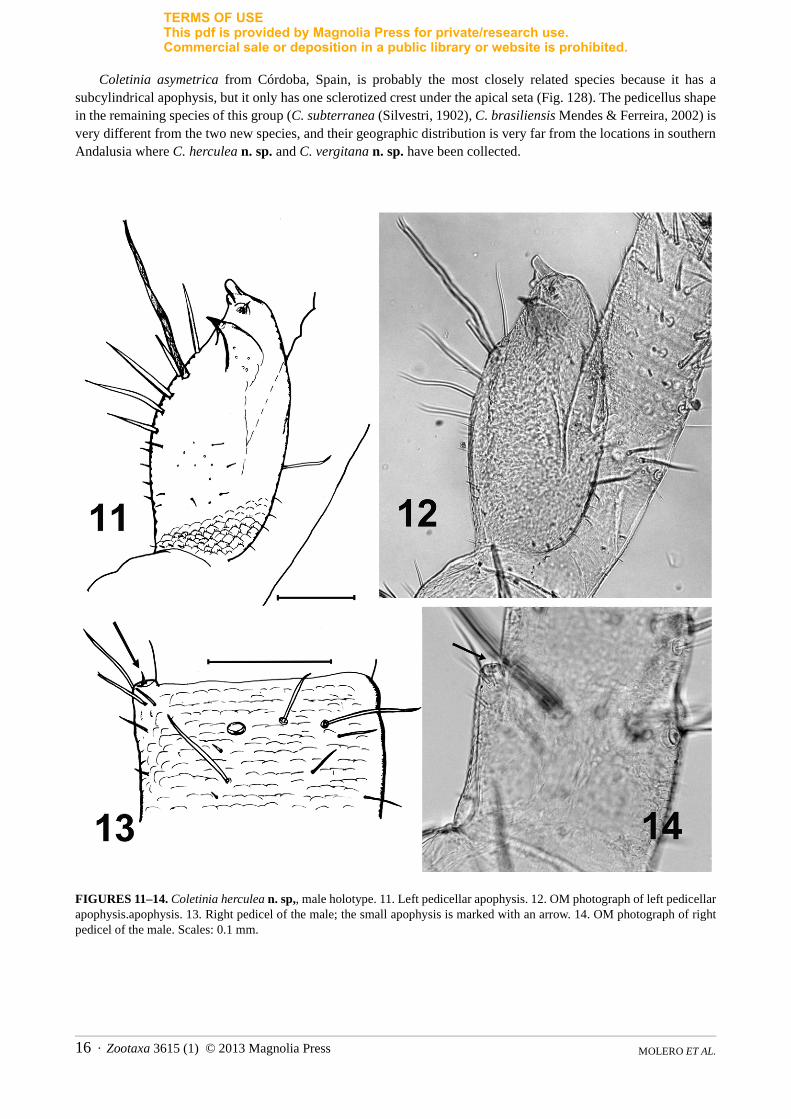
Antennae asymmetric in the holotype: left pedicellus with well-developed apophysis (Figs. 11, 12); right pedicellus with minute apophysis (Figs. 13, 14), very difficult to distinguish due to its width less than one-eighth the diameter of the pedicellus, its top not surpassing distal limit of this antennal segment; its setation consisting only of an apical glandular seta and four thin setae inserted in the trunk of the apophysis.
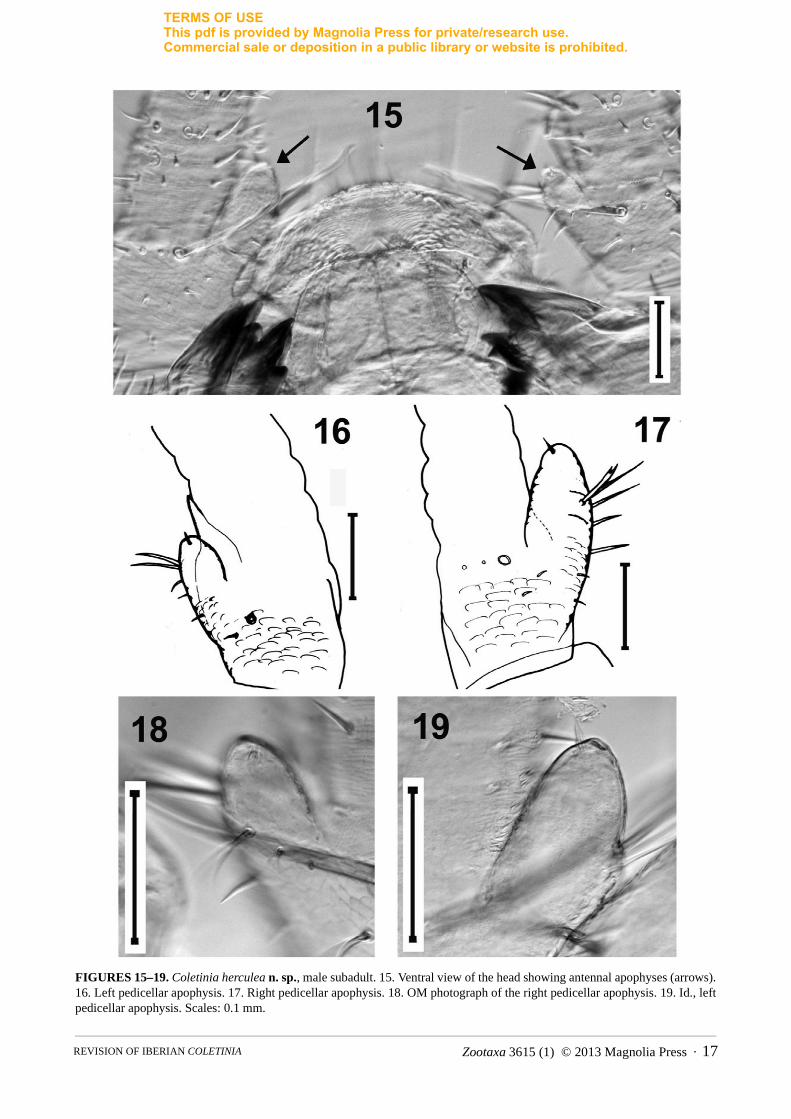
Left apophysis wider than diameter of pedicellus, apical region reaching at least third division of flagellum; shape subovoid to subcylindrical with two lamellae or processes, one of which surrounds the apical glandular seta and the other with an acute sclerotized point (hook-shaped) under the aforementioned seta. These projections oriented to the inner side of the antenna and on this side below the lamellae, trunk of the apophysis with a strong bifid macroseta and some thinner and shorter setae. In the subadult specimen, asymmetric condition of the antennae less evident: both antennae with similar cylindrical apophysis (more simple than in adult) with apical glandular seta (Figs. 15–19).
Mouthparts typical for the genus, the maxillae and the labial palp as illustrated in Figs. 20 and 21. Distal article of maxillary palp 6.5 times longer than wide and 1.7 times longer than penultimate article.
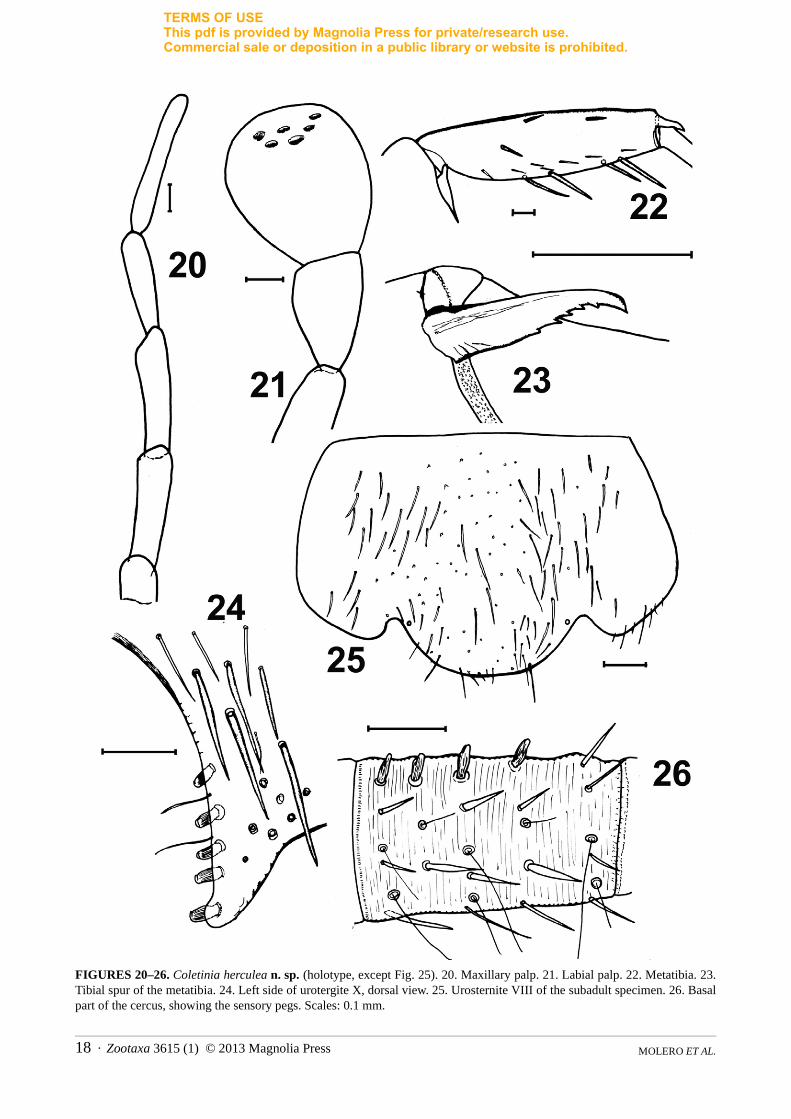
Thoracic nota as typical in Coletinia spp. Tibiae with two dorsal, one lateral and four ventral spiniform macrosetae; these setae shorter than the diameter of the tibiae (Fig. 22). Tibial spur denticulated in its hind margin (Fig. 23). L/W ratio of tibiae: protibiae = 2.95; mesotibiae = 3.2; metatibiae = 3.7.
Urotergites as typical for the genus. Tenth urotergite with posterior border slightly concave; posterolateral processes with 5+5 sensory pegs (Fig. 24). Pegs subcylindrical in shape, with slight apical emargination (similar to those in C. tinauti). Disc of urotergite X covered with robust setae.
Urosternites I-VI as typical in the genus; posterior urosternites and genitalia not well preserved, but bearing 1+1 submedian, 1+1 sublateral and 1+1 distal bifid macrosetae. Urosternites VII, VIII and parameres not well preserved in holotype. In subadult, urosternite VIII with convex posterior border, protruding between the stylets (Fig. 25); parameres subcylindrical, 3.4 times longer than wide and slightly wider in the distal half than in the basal half; nearly as long as the ninth urosternite stylets.
Cerci with 4 sensory pegs with blunt apex (Fig. 26), paracercus lost (broken in basal region).Etymology. The species is named after Hercules, the god who, according to Roman and Greek mythology,
separated the two sides of the Gibraltar Straits, on the northern side of which this new species was found.Remarks. Coletinia herculea n. sp. is a member of the “asymetrica” group. The shape of the left apophysis is unique in C. herculea n. sp. because it bears two lamellae with different
shapes and positions, one apical and above the glandular seta and the other situated under this seta and with a sclerotized point. Coletinia vergitana n. sp. also has a nearly subcylindrical apophysis, but the shape of the expansions and sclerotizations, as well as the setation of the basal trunk of the apophysis, are very distinctive. Uniquely, the bifid apex has an expansion next to the glandular cone and field of setae that extends over the external margin of the apophysis.
Zootaxa 3615 (1) © 2013 Magnolia Press · 15REVISION OF IBERIAN COLETINIA
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Coletinia asymetrica from Córdoba, Spain, is probably the most closely related species because it has a subcylindrical apophysis, but it only has one sclerotized crest under the apical seta (Fig. 128). The pedicellus shape in the remaining species of this group (C. subterranea (Silvestri, 1902), C. brasiliensis Mendes & Ferreira, 2002) is very different from the two new species, and their geographic distribution is very far from the locations in southern Andalusia where C. herculea n. sp. and C. vergitana n. sp. have been collected.
FIGURES 11–14. Coletinia herculea n. sp,, male holotype. 11. Left pedicellar apophysis. 12. OM photograph of left pedicellar apophysis.apophysis. 13. Right pedicel of the male; the small apophysis is marked with an arrow. 14. OM photograph of right pedicel of the male. Scales: 0.1 mm.
MOLERO ET AL.16 · Zootaxa 3615 (1) © 2013 Magnolia Press
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 15–19. Coletinia herculea n. sp., male subadult. 15. Ventral view of the head showing antennal apophyses (arrows). 16. Left pedicellar apophysis. 17. Right pedicellar apophysis. 18. OM photograph of the right pedicellar apophysis. 19. Id., left pedicellar apophysis. Scales: 0.1 mm.
Zootaxa 3615 (1) © 2013 Magnolia Press · 17REVISION OF IBERIAN COLETINIA
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 20–26. Coletinia herculea n. sp. (holotype, except Fig. 25). 20. Maxillary palp. 21. Labial palp. 22. Metatibia. 23. Tibial spur of the metatibia. 24. Left side of urotergite X, dorsal view. 25. Urosternite VIII of the subadult specimen. 26. Basal part of the cercus, showing the sensory pegs. Scales: 0.1 mm.
MOLERO ET AL.18 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Coletinia vergitana Molero, Barranco, Bach & Gaju new speciesFigs. 27–40
Studied material. Almería: Berja, Sierra de Gádor, Cueva del Águila, 22 March 2004, P. Barranco and J. G. Mayoral col., male holotype, 1 female paratype and 1 juvenile paratype, deposited in MNCN, type number 2237.
Description. Holotype body length: approximately 13 mm. Antennae broken, maximum length preserved 6.5 mm. Subadult length 10.5 mm. Body pale yellowish, with the pigment slightly more intense in the posterior segments.
Head with sparse short setae and regularly arranged macrosetae (Fig. 27). Lateral region of head where maxillae are inserted (condyles) very conspicuous due to intense sclerotization and dark pigmentation.
Antennae with left pedicellus showing a very developed apophysis, 1.2 times wider than pedicellus and nearly 2 times longer than wide, and reaching seventh joint of flagellum (Figs. 28–30). Basal portion of left apophysis subcylindrical and bearing on the outer side a longitudinal field of thin setae whose length increases distally. Apical portion of apophysis bifid, with two processes, the usual cone with a glandular seta and a lateroexternal expansion with a rounded shape and two bifid macrosetae on the external side; these macrosetae similar to the three remaining macrosetae inserted in the pedicellus, near the base of the apophysis. On the external side of the lateroexternal expansion, a thin seta next to the glandular cone and surpassing the top of the cone. The tegument of this expansion pilose, covered with small hairs; external side of the trunk of the pedicellus also pilose and covered with small hairs in the region of the longitudinal field of thin setae. Glandular cone with truncated apex and relatively long glandular seta; cone tegument smooth, but on the external side next to the lateroexternal expansion, tegument with a conspicuous hook-shaped sclerotized prolongation with a basal zone bearing some teeth in the margin resembling the molar area of a mandible (Fig. 30). Right pedicellus with small, rounded apophysis, in its apical internal angle bearing six thin setae; length/width ratio 0.5, width of apophysis/width of pedicellus approximately 0.3. Pedicellus with four long bifid macrosetae (Figs. 31, 32).
Apophyses of subadult specimen similar, about 2 times longer than wide, but less developed than in the adult and narrower than the pedicellus (width of the apophysis/width of the pedicellus ratio approximately 0.4) and their tips reaching only to fourth subdivision of the flagellum; special structures lacking except for the apical glandular seta. A few thin setae inserted in the apophysis. Four long, bifid macrosetae inserted in the pedicellus, absent from apophysis (Fig. 33).
Basal part of maxilla with two pigmented areas (condyles). Distal article of maxillary palp in male about 7 times longer than wide (5.25× in the female and 5× in subadult) and 1.3–1.5 times longer than the penultimate article. Labial palp without remarkable features.
Thoracic nota typical of the genus. Length/width ratio of tibiae as follows: protibiae 3.5–4, mesotibiae 4–4.5, metatibiae 5.2–5.4. Pro—and mesotibiae with 2 dorsal (one medial, one distal) and 4 ventral (2 subdistal, 2 proximal) spines, metatibiae with 5 spines (1 inserted basally, 2 medially, 2 subdistally). All tibiae with few additional small basal and medial spines (Fig. 34). Lateral spine usually present.
Urotergites typical for Coletinia. Tenth urotergite of male with concave posterior border and 6+7 sensory pegs inserted in lateral margin of the posterolateral lobes and 1+1 sensory pegs inserted in disc of urotergite, at the level of the apex of the posterolateral lobes. All pegs short, subcylindrical, truncated at the apex. Lateral area of disc covered with very robust, acute setae (Fig. 35).
Tenth urotergite of female with 1+1 strong bifid macrosetae inserted in posterolateral angles, and around them, 1+1 fields of strong acute setae (Fig. 36). Posterior border not clearly observed, but straight or slightly concave.
Urosternite I divided, urosternites II–VII with 1+1 submedian, 1+1 sublateral and 1+1 discal bifid macrosetae. Eighth urosternite of male with its hind border slightly convex, not much protruding, with subtrapezoidal shape, resembling a blunt obtusangle triangle (Fig. 37). This margin is bent in the subadult specimen, but it seems to have a concave median region.
Subgenital plate of the female semielliptical, wider than long (length/width ratio about 0.85), its posterior border rounded, but slightly truncated in the median region (Fig. 38).
Parameres 4.3 times longer than wide, very similar in the subadult specimen.Ovipositor long, with 30 divisions, extending beyond the apex of the ninth stylets more than three times their
length (Fig. 39).Cerci and paracercus broken, the cerci of the male with 2–3 subacute basal pegs, paracercus with 5 acute
pigmented spines in its basal divisions (Fig. 40).
Zootaxa 3615 (1) © 2013 Magnolia Press · 19REVISION OF IBERIAN COLETINIA

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Etymology. The specific name of this new taxon refers to Vergis, the Roman name of Berja, a town in the province of Almería (SE Spain). The cave where C. vergitana n. sp. was found is within the municipal boundaries of this town.
Remarks. Coletinia vergitana n. sp. is a member of the “asymetrica” group. Distinguishing characters for this new species are given in Remarks under C. herculea n. sp., above.
FIGURES 27–30. Coletinia vergitana n. sp., holotype male. 27. Setation of the head. 28. OM photograph of the left pedicellar apophysis. 29. Left apophysis, entire. 30. Left apophysis, detail of the apical region. Scales: 0.1 mm.
MOLERO ET AL.20 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 31–36. Coletinia vergitana n. sp. 31. OM photograph of the right pedicellar apophysis of the holotype male. 32. Right pedicellar apophysis, entire. 33. Pedicellar apophysis of the subadult paratype. 34. Metatibia of the holotype, showing spines and additional small spines. 35. Urotergite X of the male holotype, posterolateral lobe with conules and setae of the disc. 36. Urotergite X of the female paratype, right side. Scales: 0.1 mm.
Zootaxa 3615 (1) © 2013 Magnolia Press · 21REVISION OF IBERIAN COLETINIA

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 37–40. Coletinia vergitana n. sp. 37. Urosternite VIII of the male holotype. 38. Subgenital plate of the female paratype. 39. Ovipositor (paratype). 40. Basal part of the terminal filaments of the male holotype. Scales: 0.1 mm, except Fig. 39, 0.5 mm.
MOLERO ET AL.22 · Zootaxa 3615 (1) © 2013 Magnolia Press
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
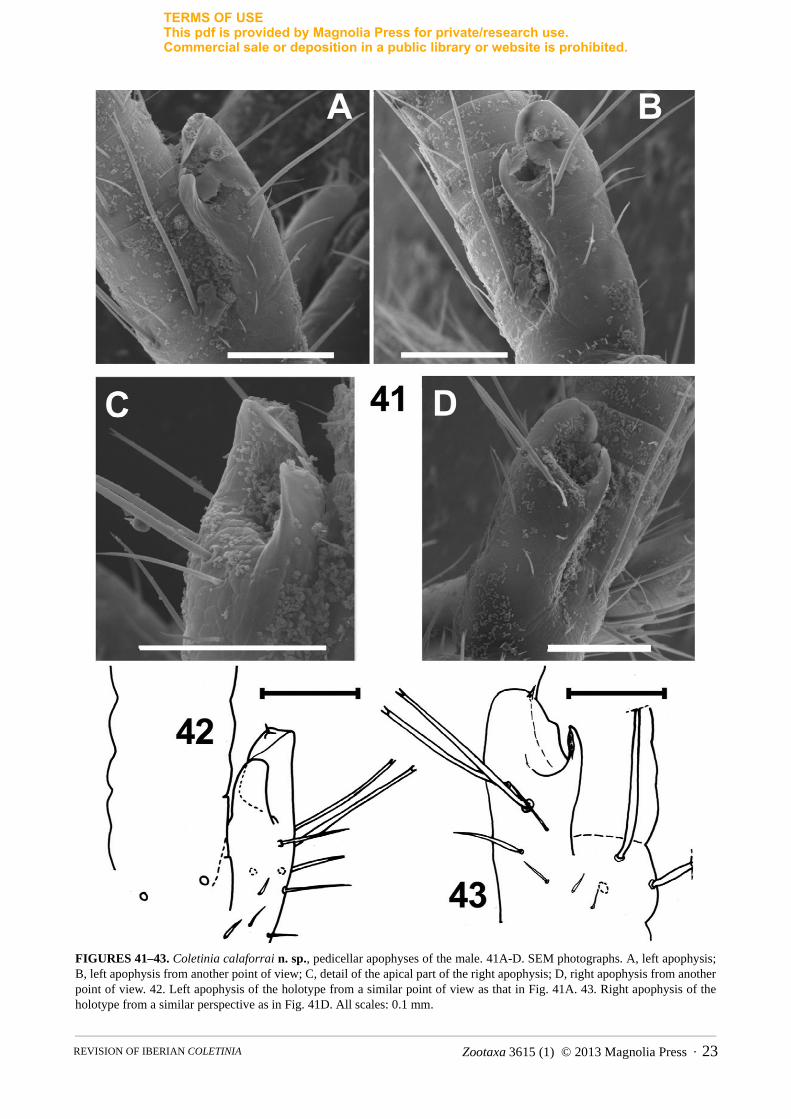
FIGURES 41–43. Coletinia calaforrai n. sp., pedicellar apophyses of the male. 41A-D. SEM photographs. A, left apophysis; B, left apophysis from another point of view; C, detail of the apical part of the right apophysis; D, right apophysis from another point of view. 42. Left apophysis of the holotype from a similar point of view as that in Fig. 41A. 43. Right apophysis of the holotype from a similar perspective as in Fig. 41D. All scales: 0.1 mm.
Zootaxa 3615 (1) © 2013 Magnolia Press · 23REVISION OF IBERIAN COLETINIA

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Coletinia calaforrai Molero, Barranco, Bach & Gaju new speciesFigs. 41–53
Studied material. Almería: Paraje Natural del Karst en Yeso de Sorbas, Covadura, C. Ruiz-Portero, coll., 11 May 2000, male holotype deposited in MNCN, type number 2238. Twenty-two paratypes, same locality and collector, other data as follows: 1 female (UCO Ref. Z2182), no date; 1 female (MNCN, type number 2238), Complejo GEP,21 August 2000; 1 female (UCO Ref. Z2183), 21 September 2000; 1 female, (UCO Ref. Z2184), Cueva del Agua, 9 November 2000; 1 female, 1 juvenile (UCO Ref. Z2185), GEP, 27 February 2001; 1 female (UCO Ref. Z2186),C-3, 30 July 2001; 1 female (deposited in CZ), GEP, 3 August 2001; 1 female, 1 incomplete juvenile (UCO Ref. Z2187), C3-R1, 19 October 2001; 2 males, 4 females, 3 juveniles, 2 incomplete specimens in alcohol, GEP, 24 October 2001, one mounted for scanning (UCO Ref. Z2177); 1 male (deposited in CZ), PB-4, 9 November 2003,.
FIGURES 44–49. Coletinia calaforrai n. sp. 44. SEM photograph of the metatibia. 45. Metatibia of the holotype, showing spines. 46. Urotergite X of the male. 47. Urotergite X of the male, detail showing sensory pegs. 48. Urotergite X of the female. 49. Hind border of urosternite VIII of the male. Scales: 0.1 mm.
MOLERO ET AL.24 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 50–53. Coletinia calaforrai n. sp. 50. Subgenital plate of female, with subconvex hind margin. 51. Subgenital plate of another specimen, with subtruncate hind margin. 52. Ovipositor. 53. Sensory pegs of the cercus. Scales: 0.1 mm, except Fig. 52, 0.5 mm.
Description. Body length of holotype 10.0 mm, maximum female length 11.2 mm. Antennae broken on all specimens, holotype with longest remnant, 5.5 mm long. Terminal filaments broken on all specimens, holotype with only a basal portion of 1–2 mm length preserved; longest preserved section in a paratype 5.5 mm (female paracercus).
Body pale yellowish, in some specimens appearing slightly darker in abdominal region due to gut contents.Macrosetae slightly spiralized. Head with few scattered short setae and few regularly arranged macrosetae (see Fig. 3).
Antennae symmetrical, pedicellus with subcylindrical apophysis at the apical part with a typical glandular cone with smooth tegument on its inner side, and a curved lateral lamella shorter than the glandular cone. Two bifid macrosetae and few short setae n basal trunk of apophysis near the cavity between the lamella and the cone (Figs. 41-43). Three additional long, bifid macrosetae inserted in pedicellus near base of apophysis. Apical part of apophysis reaching fourth division of the flagellum.
Length/width ratio of the distal article of the maxillary palp 5 in holotype, 4.2–5.5 in the remaining specimens; distal article 1.1–1.5 times longer than the antedistal article. Labial palp typical.
Tibiae slender. The L/W ratio in holotype 4.2 for protibiae, 4.4 for mesotibiae and 5.7 for metatibiae (see Table 2). Pro—and mesotibiae with 2 dorsal spines, metatibia with 1 dorsal spine; all tibiae wuth 1 lateral spine and 4 ventral spines (2 mediobasal and 2 subdistal), usually with 1 or 2 thin, short additional basal spines (Figs. 44, 45).
Urotergites without remarkable characters. Tenth urotergite of male with concave posterior margin (Fig. 46) and 4–5 + 4–5 sensory pegs arranged in a row on the external margin of the posterolateral lobes of the urotergite (Fig. 47); sensory pegs short and typical, with furrow in the dorsoapical region. Disc completely covered with thin setae. In female posterior margin of urotergite X slightly concave, disc covered with very few and sparse setae (Fig. 48).
Urosternites I-VIII with 1+1 submedian, 1+1 sublateral and 1+1 discal bifid macrosetae. Eighth urosternite of male protruding moderately, posterior margin slightly convex and subtruncate (Fig. 49).
Subgenital plate of female semielliptical, shorter than wide at its base (length/width ratio about 0.8–0.9), its posterior margin nearly straight, clearly truncate (Fig. 51) to slightly convex (Fig. 50).
Zootaxa 3615 (1) © 2013 Magnolia Press · 25REVISION OF IBERIAN COLETINIA
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Parameres 5.5 times longer than wide, as long as the ninth-segment stylets.Ovipositor relatively short and stout, with 14–16 divisions, surpassing the tip of tenth-segment stylets
approximately 1–1.7 times their length (Fig. 52). Basal division of cerci usually without sensory pegs (one specimen with one small peg), second division with 3
acute pegs (Fig. 53). Broken paracercus in holotype, with acute spine on basal division.Etymology. This new species is dedicated to José María Calaforra, a specialist in the gypsum karst of Almería
(including the caves where C. calaforrai n. sp. has been collected), who promoted the study of the troglobitic fauna of this special habitat.
Remarks. Coletinia calaforrai n. sp. is a member of the “capolongoi” group and differentiation of this species can be found in the Discussion section below.
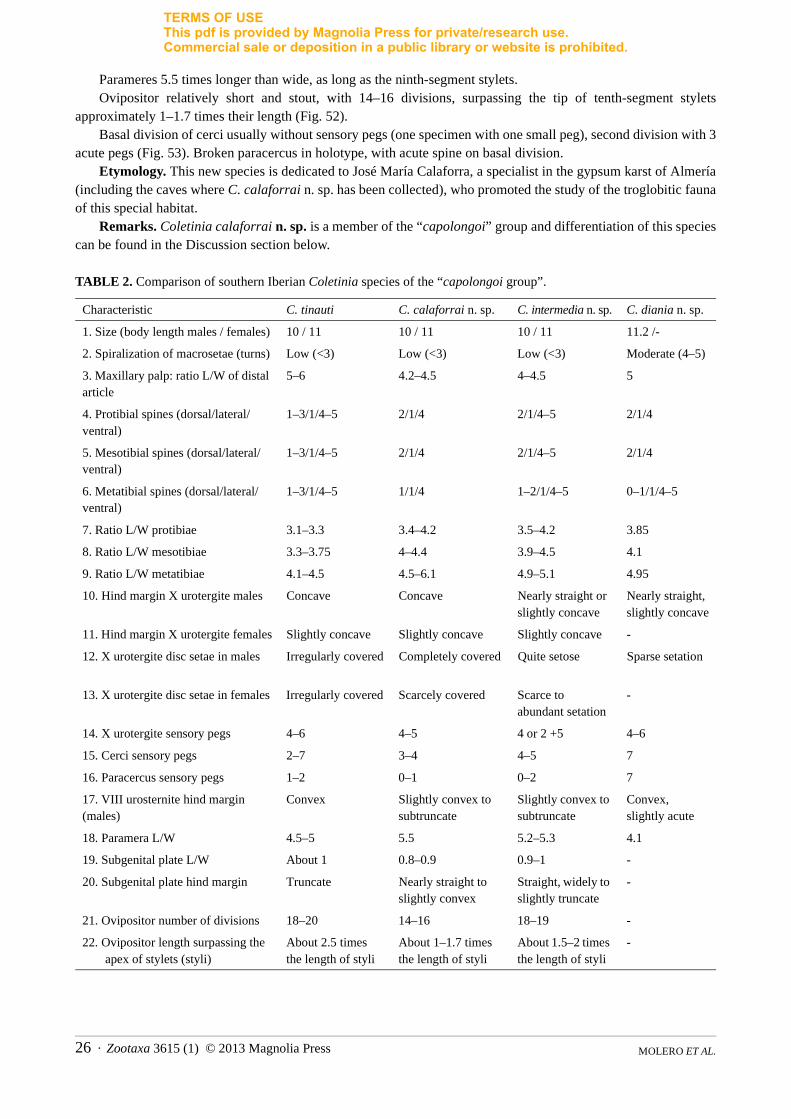
TABLE 2. Comparison of southern Iberian Coletinia species of the “capolongoi group”.
Characteristic C. tinauti C. calaforrai n. sp. C. intermedia n. sp. C. diania n. sp.
1. Size (body length males / females) 10 / 11 10 / 11 10 / 11 11.2 /-
2. Spiralization of macrosetae (turns) Low (<3) Low (<3) Low (<3) Moderate (4–5)
3. Maxillary palp: ratio L/W of distal article
5–6 4.2–4.5 4–4.5 5
4. Protibial spines (dorsal/lateral/ventral)
1–3/1/4–5 2/1/4 2/1/4–5 2/1/4
5. Mesotibial spines (dorsal/lateral/ventral)
1–3/1/4–5 2/1/4 2/1/4–5 2/1/4
6. Metatibial spines (dorsal/lateral/ventral)
1–3/1/4–5 1/1/4 1–2/1/4–5 0–1/1/4–5
7. Ratio L/W protibiae 3.1–3.3 3.4–4.2 3.5–4.2 3.85
8. Ratio L/W mesotibiae 3.3–3.75 4–4.4 3.9–4.5 4.1
9. Ratio L/W metatibiae 4.1–4.5 4.5–6.1 4.9–5.1 4.95
10. Hind margin X urotergite males Concave Concave Nearly straight or slightly concave
Nearly straight, slightly concave
11. Hind margin X urotergite females Slightly concave Slightly concave Slightly concave -
12. X urotergite disc setae in males Irregularly covered Completely covered Quite setose Sparse setation
13. X urotergite disc setae in females Irregularly covered Scarcely covered Scarce to abundant setation
-
14. X urotergite sensory pegs 4–6 4–5 4 or 2 +5 4–6
15. Cerci sensory pegs 2–7 3–4 4–5 7
16. Paracercus sensory pegs 1–2 0–1 0–2 7
17. VIII urosternite hind margin (males)
Convex Slightly convex to subtruncate
Slightly convex to subtruncate
Convex, slightly acute
18. Paramera L/W 4.5–5 5.5 5.2–5.3 4.1
19. Subgenital plate L/W About 1 0.8–0.9 0.9–1 -
20. Subgenital plate hind margin Truncate Nearly straight to slightly convex
Straight, widely to slightly truncate
-
21. Ovipositor number of divisions 18–20 14–16 18–19 -
22. Ovipositor length surpassing the apex of stylets (styli)
About 2.5 times the length of styli
About 1–1.7 times the length of styli
About 1.5–2 times the length of styli
-
MOLERO ET AL.26 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Coletinia intermedia Molero, Bach & Gaju new speciesFigs. 9C, 54–64
Studied material. Alicante, Alfas del Pi, Cova del Far, UTM 30SYH5672, 8-IV-2006, male holotype mounted on a slide, deposited in MVHN, Cod.: 080406SG2. Four paratypes: 1 male in 70% alcohol (partially dissected for SEM study, deposited in UCO, Ref. Z2188) and 3 females (1 on a slide, deposited in UCO, Ref. Z2188 and 2 in alcohol, deposited in MVHN, Cod.: 080406SG2) same collection data as holotype.
Additional specimens: Murcia, Calasparra, Cueva del Puerto, Sala de los Corales, 5 July to August 2003, 1 male collected in a beer trap (MVHN, Cod.: 201206RV57) and 3 females and 2 juveniles, not well preserved, collected in a similar trap (MVHN, Cod.: 201206RV59).
Description. Body length of holotype 10 mm, up to 11 mm in the remaining specimens. Subcylindrical shape, with very narrow thorax, maximum width 1.5 mm. Body light yellowish to nearly transparent, gut contents often visible. Antennae as long as the body or slightly longer, broken in the holotype (maximum length preserved: 11.5 mm). Macrosetae with a low degree of spiralization. Head with typical setation, with few short and thin setae and very few bifid macrosetae.
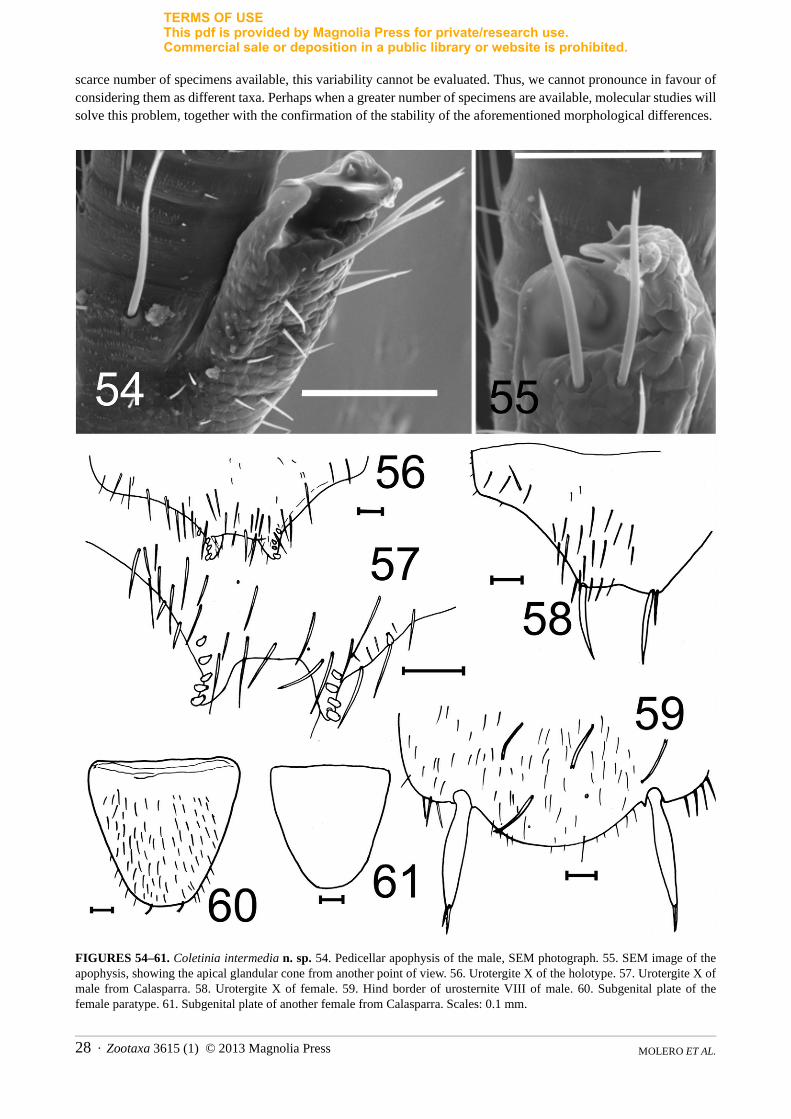
Antennae symmetrical, in male with a pedicellar apophysis that resembles that of C. tinauti or C. calaforrai n. sp., reaching fourth joint of flagellum. Apical cone joined internally on side of flagellum to the lamellar expansion, which is shorter than the cone. At the apical part of the external side of the trunk, two bifid macrosetae inserted near the cavity formed by the apical cone and the expansion; tegument of this cavity smooth, tegument of trunk rugose and scale-like (Figs. 54, 55). Three additional long bifid macrosetae inserted in the pedicellus of the antenna.
Distal article of maxillary palp relatively short and wide, approximately 3.8–4.5 times longer than wide and about 1.3 times longer than penultimate article. Labial palp with typical configuration of the genus, with 6 basiconic sensilla along the outer side and 4–5 along the inner side.
L/W ratios of tibiae given in Table 2. Pro—and mesotibiae with 2 dorsal, 1 lateral and 4–5 ventral macrosetae, metatibiae usually with one dorsal and five ventral macrosetae. Tibial spurs slightly denticulate.
Urotergites I-IX without special features. Tenth urotergite of male with posterior border nearly straight or slightly concave (Fig. 56); posterolateral lobes with 4+4 sensory pegs appearing bilobulate apically when visualized by OM (corresponding to example photographed in Fig. 9C). In addition to 4+4 conules, one male from Murcia with 1+1 additional anterior pegs and 2 spiniform pegs on one side (Fig. 57). Long setae spread over extension of the disc of the urotergite in both sexes, particularly in the centre and hind margin of the disc, scarcer in the lateral region. In females, posterior border slightly concave and flanked by 1+1 long spines (Fig. 58); tenth urotergite of a female from Murcia with less setose disc.
Urosternites I-VIII with 1+1 submedian, 1+1 sublateral and 1+1 discal bifid macrosetae. Eighth urosternite of male convex but slightly truncated at posterior border (Fig. 59).Length and basal width of female subgenital plate equal (ratio = 0.9–1), with straight posterior border that is
widely truncated in the female paratype (Fig. 60), less widely so in female from Murcia (Fig. 61). Parameres 5.2–5.3 times longer than wide, as long as ninth stylets. Ovipositor long, surpassing level of ninth stylets by nearly 2 times their length (Fig. 62), with 18–19 divisions.
Paracercus without sensory pegs in both sexes, at most with two acute spines not clearly equivalent to pegs. Second division of cerci bearing 4+4 sensory pegs with blunt apex (Fig. 63); in a male from Murcia these pegs more acute and 1+1 pegs on the first cercal segment (Fig. 64).
Etymology. This new species is called intermedia because of the presence of intermediate characteristics within the “capolongoi” group (such as the L/W ratio of tibiae, shape of eighth urosternite of the male, number of divisions of the ovipositor) and because of the central geographical position within the distribution of the species belonging to this group (Fig. 173).
Remarks. Coletinia intermedia n. sp. is a member of the “capolongoi” group and differentiation of this species can be found in the Discussion section below.
Some variability has been observed between the specimens of the type locality in Alfas del Pi (Alicante) and those from the cave of Calasparra (Murcia) assigned to the same new species. In spite of the differences observed (different number of sensory pegs on urotergite X of the male, a different number of setae in the disc of this urotergite in females, different shape in the posterior border of the subgenital plate and one additional sensory peg in the first segment of the cerci in males from Murcia), the insects from both caves are very similar and because of the
Zootaxa 3615 (1) © 2013 Magnolia Press · 27REVISION OF IBERIAN COLETINIA
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
scarce number of specimens available, this variability cannot be evaluated. Thus, we cannot pronounce in favour of considering them as different taxa. Perhaps when a greater number of specimens are available, molecular studies will solve this problem, together with the confirmation of the stability of the aforementioned morphological differences.
FIGURES 54–61. Coletinia intermedia n. sp. 54. Pedicellar apophysis of the male, SEM photograph. 55. SEM image of the apophysis, showing the apical glandular cone from another point of view. 56. Urotergite X of the holotype. 57. Urotergite X of male from Calasparra. 58. Urotergite X of female. 59. Hind border of urosternite VIII of male. 60. Subgenital plate of the female paratype. 61. Subgenital plate of another female from Calasparra. Scales: 0.1 mm.
MOLERO ET AL.28 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 62–64. Coletinia intermedia n. sp. 62. Ovipositor and stylet IX. 63. Basal portion of the cercus of the holotype, showing the positions of the sensory pegs. 64. Basal portion of the cercus of a male from Calasparra. Scales: 0.5 mm for Fig. 62, 0.1 mm for Figs. 63 and 64.
Coletinia diania Molero, Bach & Gaju new speciesFigs. 65–75
Studied material. Alicante, Denia, Cova de la Punta de Benimaquia. UTM 31SBD451011. 31 October 2004, male holotype. A. Sendra et al., coll., deposited in MVHN, Cod.: 201206RV85.
Description. Body length 11.2 mm. Antennae broken, only 1 mm preserved. Terminal filaments broken, maximum length of cercus 6 mm. Body uniformly yellowish. Macrosetae clearly spiralized, particularly evident in the head and coxae, each containing 4–5 turns of the spiral (Fig. 65).
Head with relatively abundant short, thin setae and few bifid macrosetae (Fig. 66). Antennal apophysis reaching fourth joint of flagellum, structure similar to those of C. capolongoi or C. calaforrai n. sp., with a lateral lamella shorter than the apical cone (Figs. 67, 68). Partially sclerotized region of the tegument visible in the internal region of the cavity between the lamella and the cone, next to the flagellum (Fig. 67). Pedicellus with 5 long, bifid, spiralized macrosetae, 2 of which are in the distal region of the basal trunk of the apophysis, below the apical cone. Basal trunk with very setose tegument and 4 or 5 long, thin setae as long as the diameter of the apophysis.
Distal article of maxillary palp about 5 times longer than wide and 1.25 times longer than the penultimate article. Labial palp without remarkable features, each distal article with two basal groups of 4–5 basiconic sensillae, one on the inner side, one on the outer side.
L/W ratios of tibiae given in Table 2. Pro—and mesotibiae with 2 dorsal, 1 lateral and 4 ventral spines; metatibia, with 0–1 dorsal and 5 ventral spines.
Nota and urotergites I-IX typical. Posterior margin of tenth urotergite nearly straight, slightly concave (Fig. 69), with 7+10 sensory pegs extending from posterolateral lobes to the medial region of the lateral margin of the urotergite (Fig. 70); disc covered dorsally with sparse, thin, long setae.
Urosternites I-VIII with 1+1 discal, 1+1 submedian and 1+1 sublateral bifid macrosetae. Posterior margin of the eighth urosternite protruding, convex, rounded and slightly acute at its apex (Fig. 71). Parameres relatively short, with length-width ratio about 4.1, shorter than the ninth stylets (Fig. 72).
Cerci with 7+7 sensory pegs, 2 in each basal segment and 1+(1–2)+(1–2)+1 in the second division (Figs. 73, 74). Cerci in medial portion with very long bothriotricha, more than five times longer than width of the appendage. Paracercus with 5 acute pegs medially and 2 additional pegs laterally (Figs. 73, 75).
Female unknown.Etymology. This species is called “diania” after the Latin name "Dianium" of the municipality of Denia (likely
derived from the name of the Roman goddess Diana), in which the cave where this new taxon was found is located.
Zootaxa 3615 (1) © 2013 Magnolia Press · 29REVISION OF IBERIAN COLETINIA

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 65–71. Coletinia diania n. sp., holotype male. 65. Spiralized macroseta. 66. Head chaetotaxy. 67. Pedicellar apophysis. 68. Detail of the pedicellar apophysis. 69. Urotergite X. 70. Sensory pegs of urotergite X. 71. Urosternite VIII. Scales: 0.1 mm.
MOLERO ET AL.30 · Zootaxa 3615 (1) © 2013 Magnolia Press
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
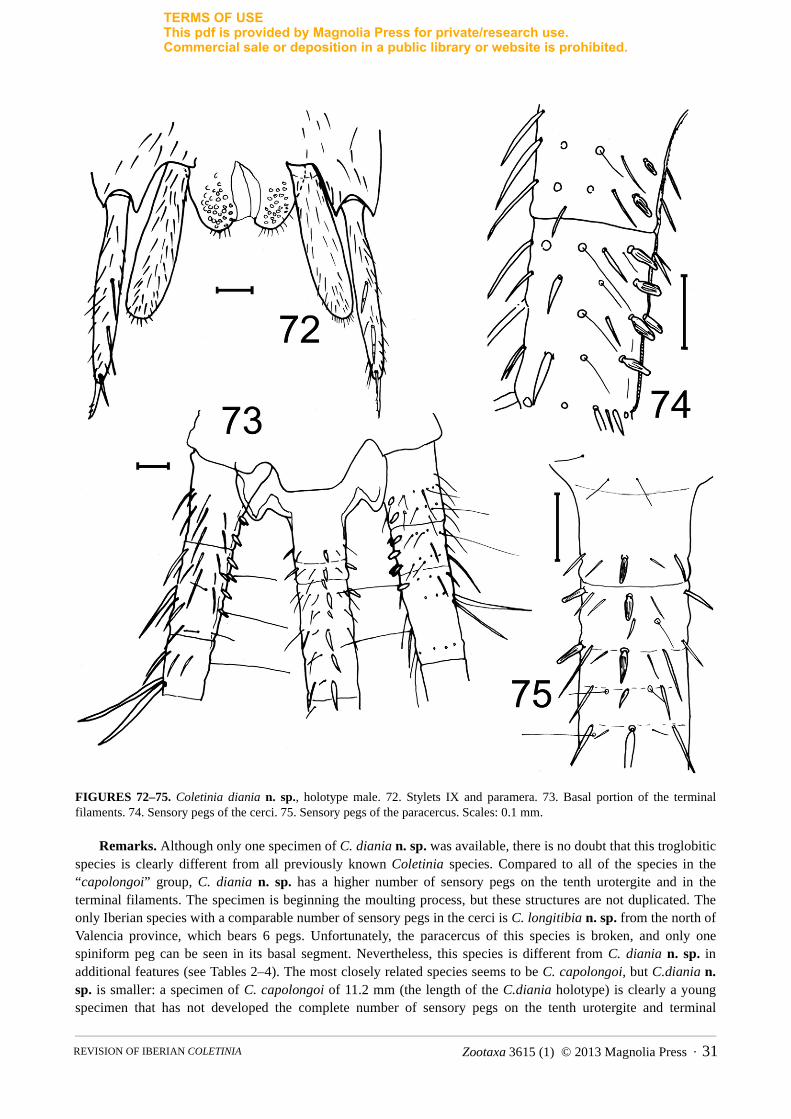
FIGURES 72–75. Coletinia diania n. sp., holotype male. 72. Stylets IX and paramera. 73. Basal portion of the terminal filaments. 74. Sensory pegs of the cerci. 75. Sensory pegs of the paracercus. Scales: 0.1 mm.
Remarks. Although only one specimen of C. diania n. sp. was available, there is no doubt that this troglobitic species is clearly different from all previously known Coletinia species. Compared to all of the species in the “capolongoi” group, C. diania n. sp. has a higher number of sensory pegs on the tenth urotergite and in the terminal filaments. The specimen is beginning the moulting process, but these structures are not duplicated. The only Iberian species with a comparable number of sensory pegs in the cerci is C. longitibia n. sp. from the north of Valencia province, which bears 6 pegs. Unfortunately, the paracercus of this species is broken, and only one spiniform peg can be seen in its basal segment. Nevertheless, this species is different from C. diania n. sp. in additional features (see Tables 2–4). The most closely related species seems to be C. capolongoi, but C.diania n. sp. is smaller: a specimen of C. capolongoi of 11.2 mm (the length of the C.diania holotype) is clearly a young specimen that has not developed the complete number of sensory pegs on the tenth urotergite and terminal
Zootaxa 3615 (1) © 2013 Magnolia Press · 31REVISION OF IBERIAN COLETINIA

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
filaments. Even when these pegs are well developed (in a male 14–15 mm long), the number of pegs in C. capolongoi is lower (4–6 on the tenth urotergite, 4–6 on the cerci and 0–5 on the paracercus).
TABLE 3. Comparison of northern Iberian Coletinia species of the “capolongoi group”.
Coletinia longitibia Molero, Bach & Gaju new speciesFigs. 76–90
Studied material. Valencia, Serra, Sima Pla dels Llomes. 3 May 2006, male holotype and 2 female paratypes, collected with bait traps. Deposited in MVHN, Cod.: 030606PL1.
Description. Male length 14.5 mm, females up to 17 mm. Colour uniformly pale yellowish. Macrosetae with a high degree of spiralization, with 4–5 or more turns (Fig. 76). Head with scarce long macrosetae and a few sparse thin setae.
Characteristic C. capolongoi C. longitibia n. sp. C. tessella n. sp. C. redetecta n. sp.
1. Length (males/females) (mm) 15 / 19.5 14.5 / 17 12 / 13.5 11.5 / 15.5
2. Spiralization of macrosetae (turns) Moderate (3–5) Moderate to high (4–5) Moderate (3–5) High (8–10)
3. Maxillary palp L/W ratio of distal article
5–5.2 4.5–5 5.75 5.7
4. Protibial spurs (dorsal/lateral/ventral)
2/1/4–5 2–4/1/4–6 2/1/4 2–3/1–2/6–8
5. Mesotibial spurs (dorsal/lateral/ventral)
2/1/4–5 2/1/4–6 2/1/4–6 2–3/1–2/6–8
6. Metatibial spurs (dorsal/lateral/ventral)
2/1/4–6 2/1/4–6 1–2/1/4–6 1–3/1–2/6–8
7. Ratio L/W protibiae 3.5–3.75 4.25–4.9 4.1–4.4 3.6–4.1
8. Ratio L/W mesotibiae 3.7–3.9 5 4.5–4.95 3.7–4.5
9. Ratio L/W metatibiae 5–5.3 (7?) 6.8–7 5.5–5.95 4.4–5.6
10. Hind margin male urotergite X Straight Not well visible Concave Slightly concave
11. Hind margin female urotergite X Slightly concave Slightly concave Nearly straight Concave
12. Urotergite X disc setae, males Extensely covered Sparsely covered Irregularly covered
Extensely covered
13. Urotergite X disc setae, females Irregularly covered Scarce, more abundant in posterolateral areas
Irregularly covered
Extensely covered
14. Urotergite X sensory pegs 7–10 8 6–8 4–8
15. Cerci sensory pegs 4–6 6 5 0–3
16. Paracercus sensory pegs 0–5 1+? 1–3 0
17. Urosternite VIII hind margin, males
Very convex Convex but slightly subtruncate
Straight, clearly truncate
Convex but nearly straight medially
18. Paramera L/W ratio 5.9 5 5–8 5–6
19. Subgenital plate L/W ratio 0.85 0.87 0.85 1–1.1
20. Subgenital plate hind margin Convex, rounded Convex, rounded Straight, truncate
Convex, rounded
21. Ovipositor divisions 15–16 18 15–18 19–20
22. Ovipositor length surpassing apex of stylets (styli)
About 1.5–2 times the length of styli
About 1.1–1.5 times the length of styli
About 0.6–1 times the length of styli
About 1.5–2 times length of styli
MOLERO ET AL.32 · Zootaxa 3615 (1) © 2013 Magnolia Press
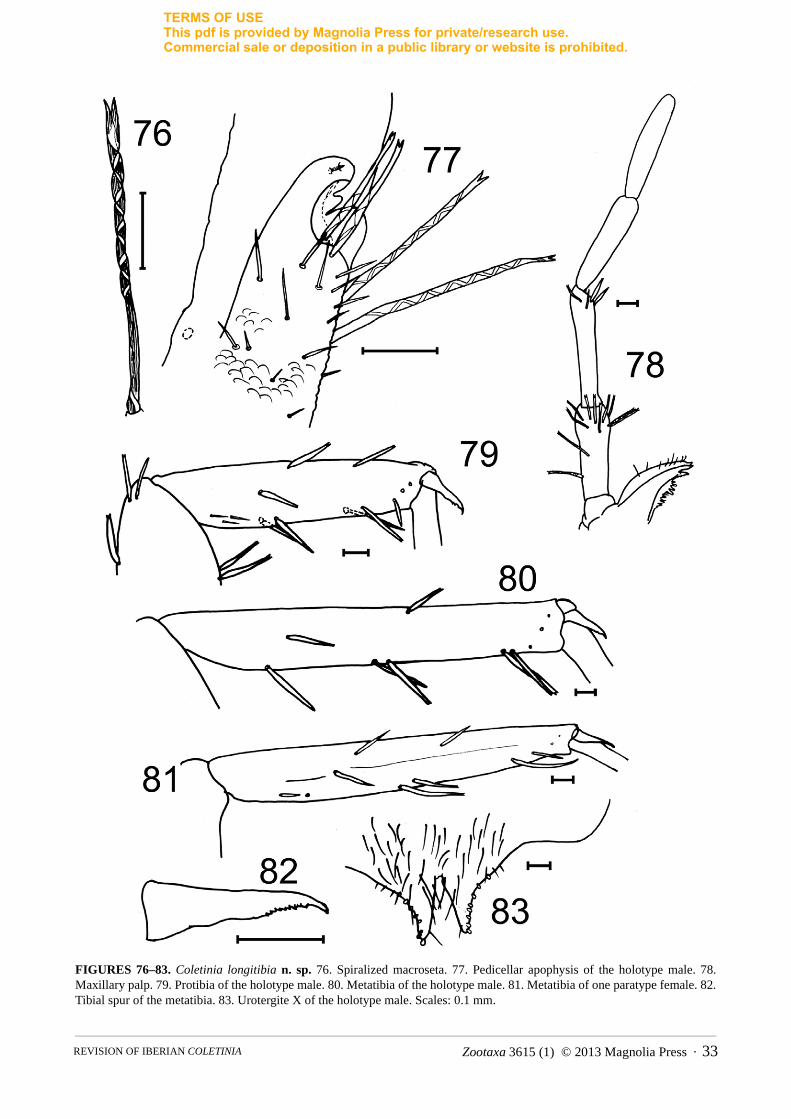
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 76–83. Coletinia longitibia n. sp. 76. Spiralized macroseta. 77. Pedicellar apophysis of the holotype male. 78. Maxillary palp. 79. Protibia of the holotype male. 80. Metatibia of the holotype male. 81. Metatibia of one paratype female. 82. Tibial spur of the metatibia. 83. Urotergite X of the holotype male. Scales: 0.1 mm.
Zootaxa 3615 (1) © 2013 Magnolia Press · 33REVISION OF IBERIAN COLETINIA
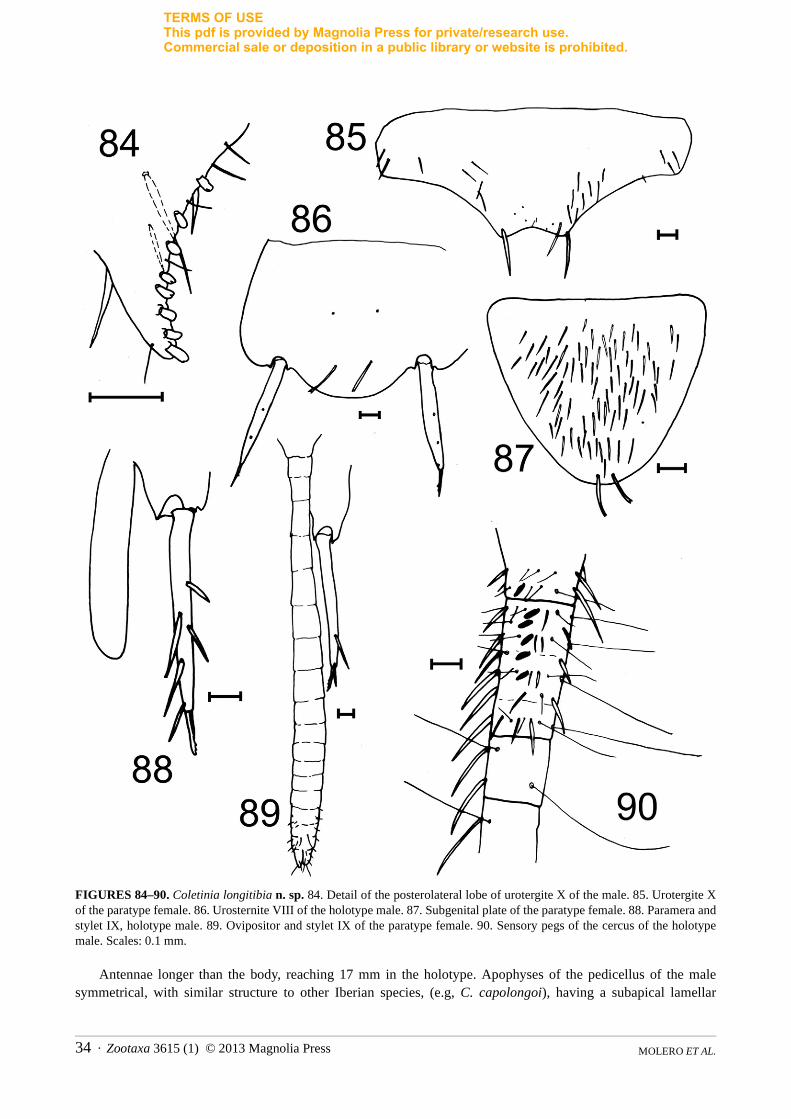
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 84–90. Coletinia longitibia n. sp. 84. Detail of the posterolateral lobe of urotergite X of the male. 85. Urotergite X of the paratype female. 86. Urosternite VIII of the holotype male. 87. Subgenital plate of the paratype female. 88. Paramera and stylet IX, holotype male. 89. Ovipositor and stylet IX of the paratype female. 90. Sensory pegs of the cercus of the holotype male. Scales: 0.1 mm.
Antennae longer than the body, reaching 17 mm in the holotype. Apophyses of the pedicellus of the male symmetrical, with similar structure to other Iberian species, (e.g, C. capolongoi), having a subapical lamellar
MOLERO ET AL.34 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
projection connected by an indented and somewhat sclerotized region to the glandular cone (Fig. 77), delimiting a subcircular smooth cavity. Basal trunk of pedicellus with few thin acute setae and five bifid macrosetae: one strong macroseta in basal region of the pedicellus opposite the base of the apophysis, two long macrosetae inserted in the median region of the basal trunk and two subequal, shorter macrosetae near the subapical process. Apical part of apophysis reaching the suture between the fourth and fifth joints of the flagellum.
Distal article of maxillary palp about 4.5–5 times longer than wide and slightly longer than penultimate article (ratio of distal to penultimate less than 1.1) (Fig. 78). Labial palp without remarkable features.
Tibiae very long, L/W ratio given in Table 2. Metatibiae 6.8–7 times longer than wide (Figs. 79–81). Tibial spines shorter than diameter of tibia; usual number of tibial spines as 2 dorsal (1 medial and 1 distal), 1 lateral and 4–6 ventral; dorsal side of protibia sometimes bearing 3 or 4 spines. Tibial spur clearly denticulated (Fig. 82).
Tenth urotergites of the male and female specimens folded on slides, posterior borders not seen clearly. Male posterolateral lobes bearing 8+8 sensory pegs (Fig. 83), disc likely densely covered with setae (Fig. 84). Posterior border of female probably slightly concave and the disc with few setae, which are more concentrated in the posterolateral regions (Fig. 85).
Urosternites typical, with 1+1 submedian, 1+1 sublateral and 1+1 discal bifid macrosetae. Posterior border of urosternite VIII of male protruding and convex but nearly straight or truncated near the midline (Fig. 86). Subgenital plate of female semielliptical, with posterior border strongly convex, arched (Fig. 87), L/W ratio of this sclerite approximately 0.87.
Parameres about 5 times longer than wide and as long as the ninth stylets (Fig. 88). Ovipositor long, with 18 divisions, surpassing paramere apex by 1.1 times their length, or 1.5 times if apical spine of stylet is not taken into account (see Fig. 89).
Terminal filaments broken. Cerci up to 16 mm long, in the male with 6 acute pegs with a blunt apex (Fig. 90). Paracercus broken at its base in the holotype, with only 1 medioventral spiniform peg visible; paracercus up to 14 mm long in females.
Etymology. The specific name for C. longitibia n. sp. refers to the extreme relative length of the tibia, greater in this species than in any other European species.
Remarks. C. longitibia n. sp. differs from all Iberian Coletinia in the high L/W ratio of the tibiae. Only one other Coletinia species, C. longissima from Turkey (Mendes, 1988), has longer tibiae (length/width ratio about 9), but the Turkish species differs in the shape of the male pedicellar apophysis, which is longer and bears a thin, curved, long expansion (nothing similar within Spanish Coletinia), and in other features such as the higher number of sensory pegs on the tenth urotergite and the shorter ovipositor. Some large specimens belonging to C. capolongoi can reach a similar L/W ratio of the tibiae, as the illustrations in Wygodzinsky (1980) suggest, but these specimens are nearly 20 mm long, and specimens of C. longitibia n. sp. with an L/W ratio of the metatibiae of approximately 7 are smaller.
Coletinia longitibia n. sp. belongs to the group “capolongoi” and is particularly related to those species found in the northern Spanish Mediterranean (C. capolongoi, C. redetecta n. sp.), having larger size and a convex hind border of the eighth urosternite in males and subgenital plate in females.
Coletinia tessella Molero, Bach & Gaju new species Figs. 91–101 Studied material. Valencia, Vilamarxant, Cueva de los Murciélagos (30SYJ0479),. 12 March 2006, male holotype, 1 female and 1 juvenile paratypes, A. Sendra et al., coll. Deposited in MVHN, Cod.: 120306MV5; Additional specimens: Valencia, Lliria, Cova del Cavall (30SYJ0688), 23 December2006, 1 male, 1 female. A. Sendra et al., coll. Deposited in UCO, Ref. Z2189; Valencia, Serra. Cova de l’Alt del Pí, 20 December 2004, 1 female, A. Sendra et al., coll. Deposited in MVHN, Cod.: 201206RV43.
Description. Body length up to 12 mm in males (holotype 12 mm), up to 13.5 mm in females. Body yellowish, abdomens of some specimens darkener yellowish brown due gut contents. Macrosetae with moderate degree of spiralization, those inserted in the antennal pedicellus the most twisted. Head with scattered short hairs and few macrosetae arranged as in Fig. 91.
Zootaxa 3615 (1) © 2013 Magnolia Press · 35REVISION OF IBERIAN COLETINIA
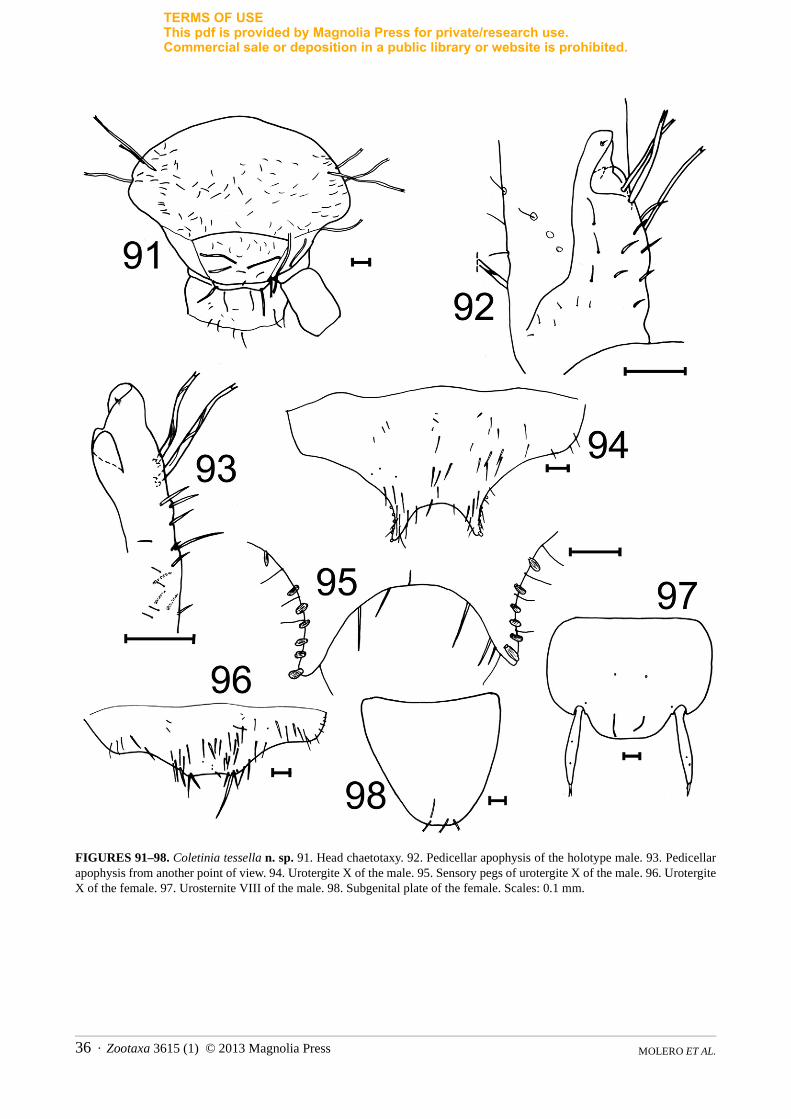
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 91–98. Coletinia tessella n. sp. 91. Head chaetotaxy. 92. Pedicellar apophysis of the holotype male. 93. Pedicellar apophysis from another point of view. 94. Urotergite X of the male. 95. Sensory pegs of urotergite X of the male. 96. Urotergite X of the female. 97. Urosternite VIII of the male. 98. Subgenital plate of the female. Scales: 0.1 mm.
MOLERO ET AL.36 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 99–101. Coletinia tessella n. sp. 99. Ovipositor and stylet IX. 100. Basal portion of the cercus, showing the sensory pegs. 101. Basal portion of the paracercus. Scales: 0.1 mm.
Antennae symmetrical, the apophysis of the pedicellus with a plate-like subapical process similar to those described for the related species C. capolongoi or C. longitibia n. sp. (Figs. 92, 93); subapical process clearly shorter than glandular cone and connected to the cone by a somewhat sclerotized region defining a cavity, and with an indented superior margin resembling a small second process; apical part of apophysis reaching fourth joint of the flagellum.
Mouthparts typical, the distal article of the maxillary palp about 5.75 times longer than wide and 1.1–1.2 times longer than the penultimate article.
Thoracic nota as usual in Coletinia species. Tibiae long, L/W ratios given in Table 2. Tibiae usually with 2 dorsal, 1 lateral and 4 ventral spines, all of them shorter than the diameter of the article. Sometimes the mesotibiae and metatibiae with 5 or 6 ventral spines metatibiae of some specimens with only one dorsal spine.
Urotergites I-IX typical. Tenth urotergite with its posterior border concave (Figs. 94, 95); posterolateral processes with 6–8 sensory pegs each, disc not uniformly covered with setae. Setae more abundant in the lateral margins, in the median region of the disc and near the posterolateral lobes, and sparser in the centrolateral region (Fig. 95). In the female, tenth urotergite with a nearly straight posterior border and 1+1 long spines inserted in the posterolateral angles; disc irregularly covered with short thin setae (Fig. 96).
Urosternites I-VII typical, with 1+1 submedian, 1+1 sublateral and 1+1 discal bifid macrosetae. Urosternite VIII of the male with protruding, straight and widely truncate posterior border (Fig. 97). Parameres more than 5 times longer than wide and as long as the ninth stylets. Subgenital plate of the female about 0.85 times longer than wide at its base, semielliptical to subtriangular, with a straight and truncate posterior border (Fig. 98). Ovipositor relatively short and stout, with 15–18 divisions, surpassing the level of the apex of ninth stylets by 0.6–1 times their length (Fig. 99).
Male cerci each with 4 or 5 acute sensory pegs (Fig. 100). Paracercus apparently lacking pegs in the holotype (at least with 1 short spine visible), but in this specimen, the terminal filaments are not in good condition; three acute pegs visible in the other examined male (Fig. 101).
Etymology. The many different species of Coletinia discovered in the Spanish Mediterranean slope constitutes a complex mosaic, and within it, C. tessella n. sp. represents a new piece, thus receiving the name because “tessella” is the Roman word for a piece of a mosaic (tile).
Remarks. C. tessella n. sp. belongs to the “capolongoi” group because its cephalic setation and symmetric condition of the antennae of the male. In this group, only C. calaforrai n. sp. and C. intermedia n. sp. share with C.
Zootaxa 3615 (1) © 2013 Magnolia Press · 37REVISION OF IBERIAN COLETINIA

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
tessella n. sp. a truncated posterior border of the eighth urosternite in males, and also a truncated straight posterior border of the subgenital plate in females. Nevertheless, the eighth urosternites of both species are not as straight posteriorly as in C. tessella n. sp. These three species are related, but C. tessella n. sp. is longer, has a higher number of sensory pegs on urotergite X and a higher tibial L/W ratio than the other two species.
Compared with C. calaforrai n. sp., C. tessella n. sp. shows a more irregular covering of setae on the tenth urotergite and a higher ratio L/W of the distal article of the maxillary palp.
Compared with C. intermedia n. sp., C. tessella n. sp. shows also a more concave posterior margin of the Xth urotergite in males and more straight in females, a slightly shorter subgenital plate and shorter ovipositor.
Coletinia redetecta Molero, Bach & Gaju new speciesFigs. 2A, 7, 102–116
= Coletinia sp. II in Wygodzinsky (1980).
Studied material. Castellón, Sant Mateu, Cova dels Encenalls, 21 March 2003, male holotype and 1 female paratype, S. Montagud et al. coll., captured directly, deposited in MVHN, Cod.: 231108BM32; Castellón, Cabanes, Avenc d’En Serenge, 2 February 2003, 1 male paratype, S. Montagud et al. coll., deposited in MVHN, Cod.: 231108BM33; Castellón, Coves de Vinromá, Cova del Mas d’Abat, 17 June 2005, 1 female paratype, F. Fadrique leg., deposited in UCO, Ref. Z2171.
Description. Body length 11–11.5 mm in males, up to 15.5 mm in females. Body uniformly pale yellowish to pale brown, or yellowish with a brownish abdomen.
Macrosetae with a high degree of spiralization; on the head, pedicellus and coxae the number of turns reaching 8–10, these highly twisted macrosetae also inserted on the maxillae and femora (Fig. 2A).
Head with scattered, thin and short setae and a few bifid macrosetae (Fig. 102). Antennae of the male symmetrical; pedicellar apophysis with its structure resembling C. capolongoi, bearing
an apical glandular cone (in males, internal glands behind the setae clearly visible by O.M.) and a subapical platelike process with rounded upper margin connecting to the base of the glandular cone through a region of smooth and more sclerotized tegument with an indentation in the upper margin and defining a subapical cavity (Fig. 103). In both observed males, one bifid macroseta inserted near this cavity and an insertion of another macroseta; pedicellus also bearing three long, highly spiralized macrosetae. Tegument of the scapus, pedicellus and basal trunk of the apophysis squamose; basal trunk bearing few thin and relatively long setae. The apical part of the apophysis reaching its limit between the fourth and fifth joints of the flagellum.
Distal article of maxillary palp 5.7 times longer than wide and approximately 1.2 longer than the penultimate article (Fig. 104).
Nota as is usual in the genus. Tibiae relatively long; their L/W ratios given in Table 2. Most tibial spines equal to or shorter than diameter of the tibiae, subdistal ventral spines slightly longer than width of the tibiae. All tibiae with 2 or 3 dorsal, 1 or 2 lateral and usually more than 6 ventral spines; metatibia sometimes with only one dorsal spine. Ventral spines of 2 subdistal, 2 mediobasal and 2 or more basal spimes; additional thin and short spines present mediobasally or basally on the ventral side of tibiae (Figs. 105–107).
Urotergites I-IX typical. Tenth urotergite of male with a concave rear edge between the posterolateral lobes (Fig. 108); margin with obtuse angle in the holotype, more rounded in the other male observed (Fig. 110). Urotergite of holotype appearing somewhat folded in its midline. Posterolateral lobes with 4–8 sensory pegs each; in holotype only 4+4 pegs visible (Fig. 109), but number possibly as posterior segments not well preserved; the other male bearing 7+8 pegs (Fig. 110). In the female, tenth urotergite has with concave posterior border, and its disc extensively covered with thin setae (Fig. 111).
Urosternites I-VII without remarkable characters, with 1+1 discal, 1+1 submedian and 1+1 sublateral bifid macrosetae. Eighth urosternite of the male protruding, convex, nearly flattened and straight near the midline (Fig. 112).
Parameres as long as ninth stylets, somewhat collapsed in the holotype, about 5–6 times longer than wide (Fig. 113).Subgenital plate of the female with strongly convex hind border and slightly longer than wide at its base (Fig.
114). Ovipositor long with 19–20 divisions, surpassing the level of the ninthth stylets by 1.5–2 times their length (Fig. 115).
MOLERO ET AL.38 · Zootaxa 3615 (1) © 2013 Magnolia Press
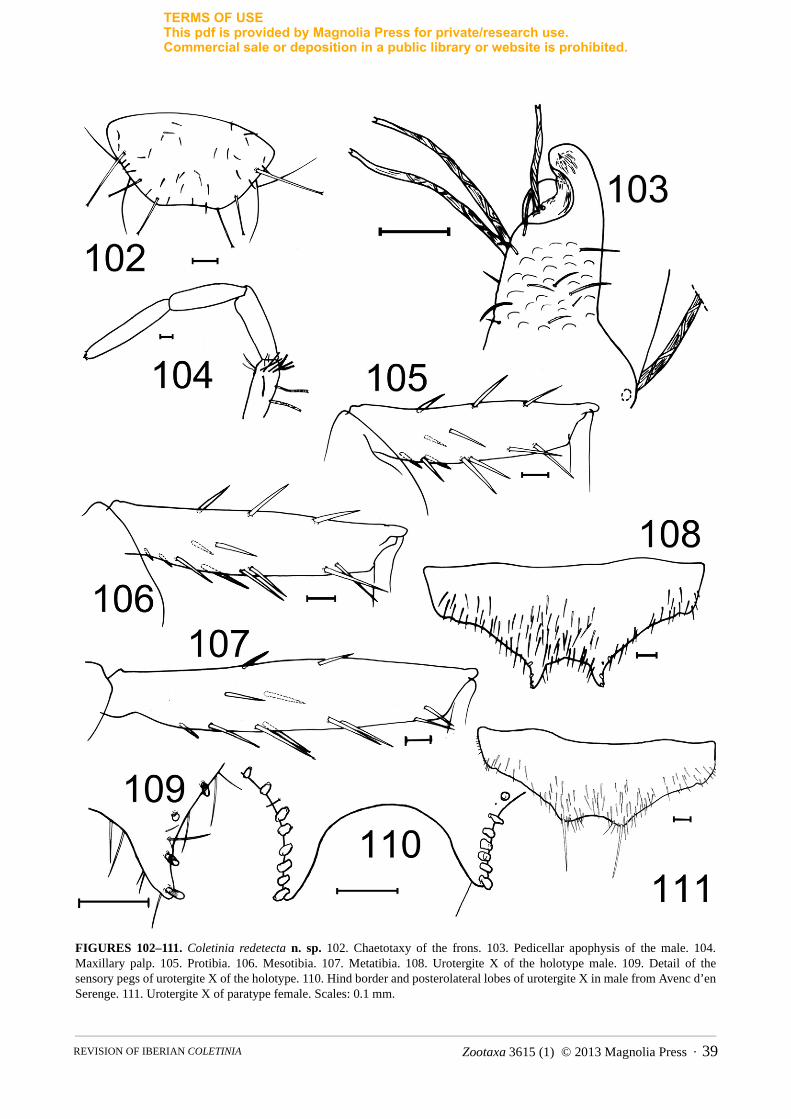
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 102–111. Coletinia redetecta n. sp. 102. Chaetotaxy of the frons. 103. Pedicellar apophysis of the male. 104. Maxillary palp. 105. Protibia. 106. Mesotibia. 107. Metatibia. 108. Urotergite X of the holotype male. 109. Detail of the sensory pegs of urotergite X of the holotype. 110. Hind border and posterolateral lobes of urotergite X in male from Avenc d’en Serenge. 111. Urotergite X of paratype female. Scales: 0.1 mm.
Zootaxa 3615 (1) © 2013 Magnolia Press · 39REVISION OF IBERIAN COLETINIA
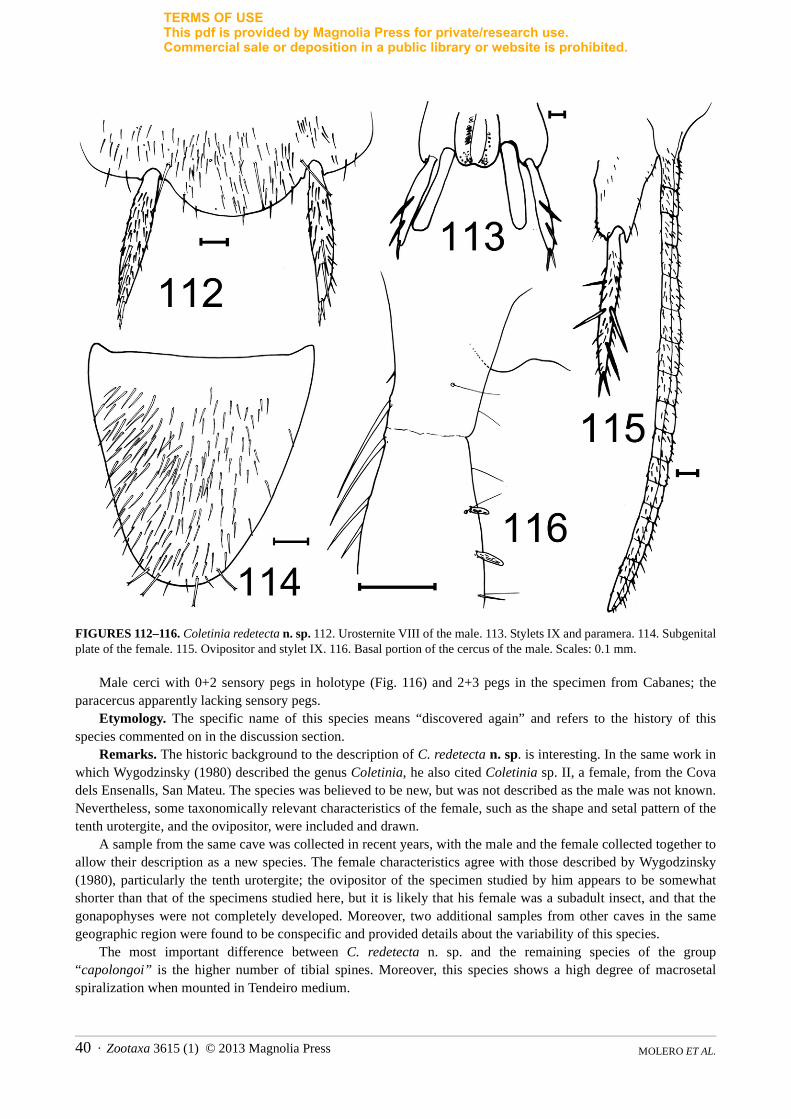
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 112–116. Coletinia redetecta n. sp. 112. Urosternite VIII of the male. 113. Stylets IX and paramera. 114. Subgenital plate of the female. 115. Ovipositor and stylet IX. 116. Basal portion of the cercus of the male. Scales: 0.1 mm.
Male cerci with 0+2 sensory pegs in holotype (Fig. 116) and 2+3 pegs in the specimen from Cabanes; the paracercus apparently lacking sensory pegs.
Etymology. The specific name of this species means “discovered again” and refers to the history of this species commented on in the discussion section.
Remarks. The historic background to the description of C. redetecta n. sp. is interesting. In the same work in which Wygodzinsky (1980) described the genus Coletinia, he also cited Coletinia sp. II, a female, from the Cova dels Ensenalls, San Mateu. The species was believed to be new, but was not described as the male was not known. Nevertheless, some taxonomically relevant characteristics of the female, such as the shape and setal pattern of the tenth urotergite, and the ovipositor, were included and drawn.
A sample from the same cave was collected in recent years, with the male and the female collected together to allow their description as a new species. The female characteristics agree with those described by Wygodzinsky (1980), particularly the tenth urotergite; the ovipositor of the specimen studied by him appears to be somewhat shorter than that of the specimens studied here, but it is likely that his female was a subadult insect, and that the gonapophyses were not completely developed. Moreover, two additional samples from other caves in the same geographic region were found to be conspecific and provided details about the variability of this species.
The most important difference between C. redetecta n. sp. and the remaining species of the group “capolongoi” is the higher number of tibial spines. Moreover, this species shows a high degree of macrosetal spiralization when mounted in Tendeiro medium.
MOLERO ET AL.40 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Coletinia hernandoi Molero, Bach & Gaju new speciesFigs. 117–127
Studied material. Tarragona, Sierra de Llabería, 4 May 2009, from soil samples collected by C. Hernando, male holotype and 1 female paratype, deposited in MNCN, type number 2239.
Description. Body length of male 10 mm, female 11 mm. Body yellowish, dorsally more intensely pigmented. Macrosetae with a moderate degree of spiralization; those inserted in the coxae with 4–6 turns (Fig. 117). Head with few bifid macrosetae and dispersed thin and short setae, which are more abundant in the occipital region (Fig. 118).
Antennae of the male symmetrical; pedicellar apophysis subcylindrical as in C. maggii but more narrow, approximately 5 times longer than the basal width and reaching the level of the fourth joint of the flagellum. Width of apophysis about half that of basal part of flagellum. Apical seta not well developed, almost completely sunken in the tegument (Fig. 119).
Distal article of the maxillary palp 3.4–3.8 times longer than wide and about 1.5 times longer than the penultimate article (Fig. 120). Labial palp typical of the genus.
Nota without special characters. Tibiae short (L/W ratio of protibia, mesotibia and metatibia: 3.15–3.35, 3.35–3.5 and 4.20–4.35, respectively), with 1 or 2 dorsal and 4 ventral spines; lateral spine usually present, lacking on some protibiae and mesotibiae.
Urotergites I–IX typical of the genus. Tenth urotergite of male with concave hind border between the posterolateral lobes, the concavity about as wide as long (Fig. 121). Four sensory pegs on each posterolateral lobe and an additional peg present in a more basal position (Fig. 122). Female tenth urotergite with concave posterior border, its disc densely covered with thin setae (Fig. 123).
Urosternites I–VII as usual, bearing 1+1 discal macrosetae. Eighth urosternite of male with a very protruding and convex hind margin (Fig. 124).
Parameres as long as stylets IX, and almost 5 times longer than wide. Subgenital plate of the female nearly as wide as long (L/W ratio about 0.9), with its apex acutely convex (Fig. 125). Ovipositor long, with 23 divisions, surpassing the level of the ninth stylets by more than 3 times their length (Fig. 126).
Male cerci with 6 or 7 elongated, acute pegs, with two pairs inserted at the same level (Fig. 127). Paracercus with a basal series of 4 long, striated acute spines.
Etymology. The specific name is dedicated to Carlos Hernando from Barcelona, who collected the soil samples where these insects were found.
Remarks. Coletinia hernandoi n. sp. is closely related to C. maggii because of the subcylindrical shape of the pedicellar apophyses; the remaining known species of Iberian Coletinia have more complex antennal apophyses. Some additional features are within the normal variation of C. maggii, such as the shape and number of sensory pegs of the tenth urotergite in the male, L/W ratio and setation of the tibiae, and length and number of divisions of the ovipositor. However, these specimens cannot be included as C. maggii because of the different development of the male antennal apophyses, which are very narrow and long compared with those of C. maggii. In Iberian C. maggii, these apophyses are about 3 times longer than wide, and their width is only slightly less than the width of the basal divisions of the flagellum. Central European C. maggii have apophyses with a similar L/W ratio and the basal flagellum is about 2 times wider than the apophysis (Wygodzinsky, 1980, also see Figs. 3B, 3C, 4A, 4B and 5E of current paper). In C. hernandoi n. sp., the male antennal apophyses are approximately 5 times longer than wide and very narrow compared to the width of the flagellum (Fig. 119). It is not reasonable to consider this difference to fall within the range of variability of C. maggii.
The shape and setation of the tenth urotergite of females of C. maggii and C. hernandoi n. sp. are clearly different. Females of C. maggii show a straight hind margin of this urotergite, but the specimen from Tarragona shows a clearly concave hind margin. In C. maggii the disc of the tenth urotergite of females is devoid of setae, with at most a few small setae inserted near the lateral margins, but in the new Iberian species, the female bears dense setation on the disc of this urotergite (Fig. 123). Some other features appear to be different in the new species, such as the shape of the eighth urosternite of the male, whose hind margin is not as truncated as in C. maggii (compare Figs. 124 and 168 in the current paper and Figs. 3L and 5A in Wygodzinsky, 1980), but more robust conclusions cannot be made unless more specimens are available.
Zootaxa 3615 (1) © 2013 Magnolia Press · 41REVISION OF IBERIAN COLETINIA
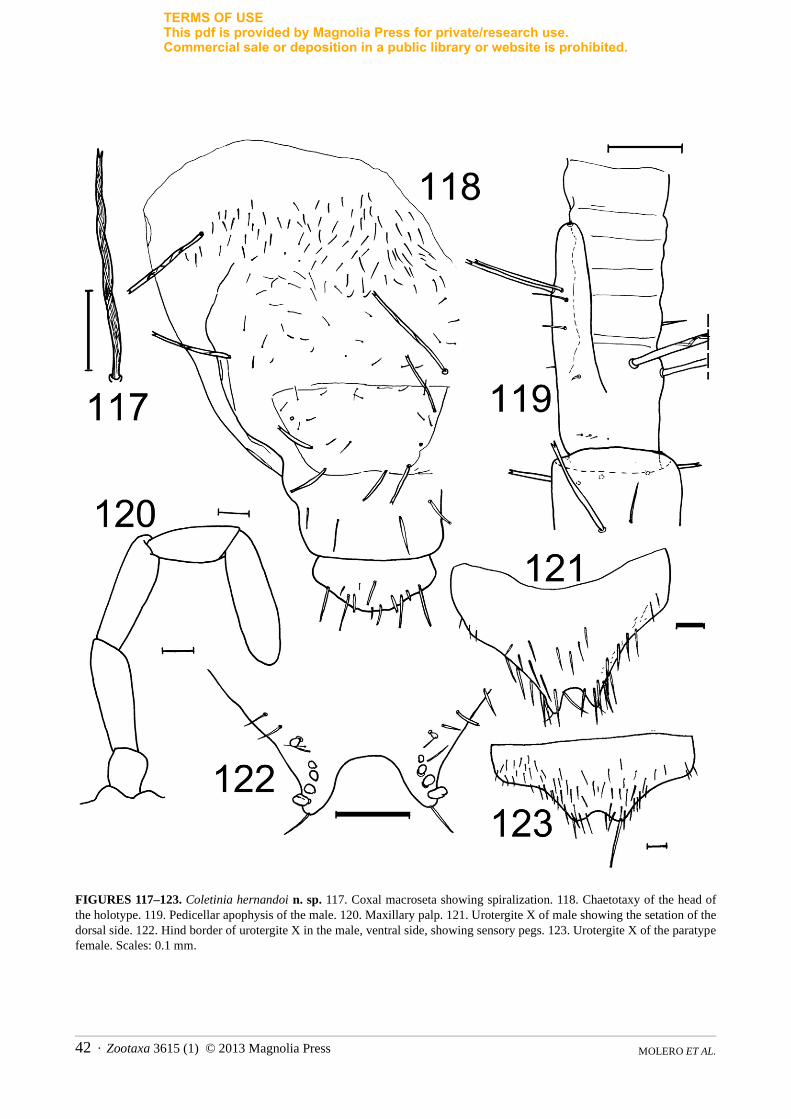
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 117–123. Coletinia hernandoi n. sp. 117. Coxal macroseta showing spiralization. 118. Chaetotaxy of the head of the holotype. 119. Pedicellar apophysis of the male. 120. Maxillary palp. 121. Urotergite X of male showing the setation of the dorsal side. 122. Hind border of urotergite X in the male, ventral side, showing sensory pegs. 123. Urotergite X of the paratype female. Scales: 0.1 mm.
MOLERO ET AL.42 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 124–127. Coletinia hernandoi n. sp. 124. Urosternite VIII of the male. 125. Subgenital plate of the paratype female. 126. Ovipositor and stylet IX. 127. Basal portion of the terminal filaments of the male, showing spiniform sensory pegs. Scales: 0.1 mm.
Previously known species
Coletinia asymetrica Bach de Roca, Mendes & Gaju Ricart, 1985Figs. 128–133
Studied material. No additional material for this species has been found since its original description, but for this revision some specimens belonging to the type series have been re-examined.
Descriptive remarks. This species is unique because of the configuration of the asymmetrical pedicellar apophyses in the male (Figs. 128, 129); subadult specimens also exhibit this asymmetry, which distinguishes this species from C. herculea n. sp. and C. vergitana n. sp. Each tibiae has 2 dorsal, 1 lateral (sometimes absent) and 4 ventral spines. The posterior margin of the tenth urotergite of males is variable, straight in adult specimens (Fig. 130) or concave in juveniles, and each posterolateral lobe of the urotergite possesses 4–6 sensory pegs. As in C. maggii, the posterior border of the tenth urotergite in the female is straight and the disc is devoid of setae, with only sparse setae near the margin of the urotergite (Fig. 131). The eighth urosternite of males is protruding, convex and rounded posteriorly as in other asymmetric species (Fig. 132). The subgenital plate of the female isas long as wide at its base (Fig. 133) and has a rounded posterior border. The ovipositor long and the gonapophyses have a high number of divisions (33–35). More details about this species can be found in the original description (Bach et al.1985).
Coletinia mendesi Wygodzinsky, 1980Figs. 1A, 4, 6, 8, 134–140
Studied material. Cádiz, San Roque, 31 December 1979, 1 female, under stones; published as Coletinia sp. V in Molero-Baltanás et al. (1997); deposited in UCO, Ref. Z0303. Córdoba, Palenciana, 28 June 2001, captured in Sentri-Sol traps for termites, 3 males (partially dissected for SEM study) and 2 juveniles, Ref. Z2172. Córdoba, edaphic in decomposed wood, August 2001, 1 juvenile male, UCO, Ref. Z2173.
Part of the material from Portugal reported by Mendes (1985, 1996) has also been examined.
Zootaxa 3615 (1) © 2013 Magnolia Press · 43REVISION OF IBERIAN COLETINIA
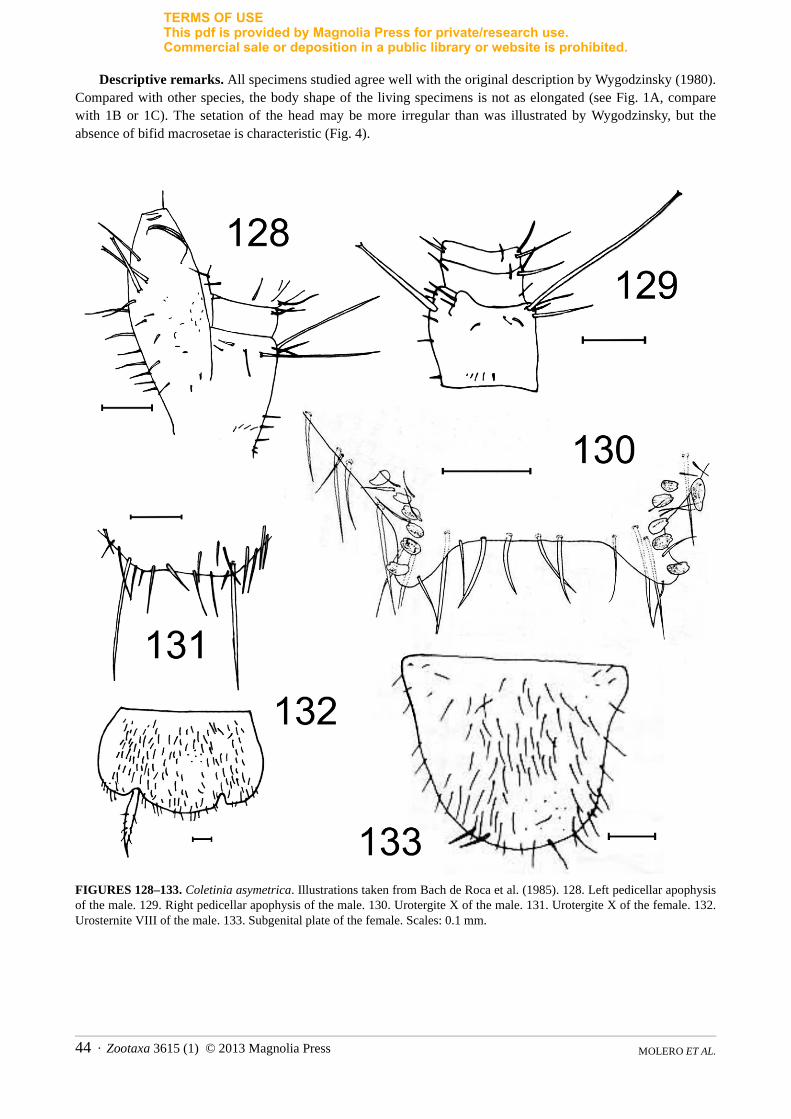
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Descriptive remarks. All specimens studied agree well with the original description by Wygodzinsky (1980). Compared with other species, the body shape of the living specimens is not as elongated (see Fig. 1A, compare with 1B or 1C). The setation of the head may be more irregular than was illustrated by Wygodzinsky, but the absence of bifid macrosetae is characteristic (Fig. 4).
FIGURES 128–133. Coletinia asymetrica. Illustrations taken from Bach de Roca et al. (1985). 128. Left pedicellar apophysis of the male. 129. Right pedicellar apophysis of the male. 130. Urotergite X of the male. 131. Urotergite X of the female. 132. Urosternite VIII of the male. 133. Subgenital plate of the female. Scales: 0.1 mm.
MOLERO ET AL.44 · Zootaxa 3615 (1) © 2013 Magnolia Press
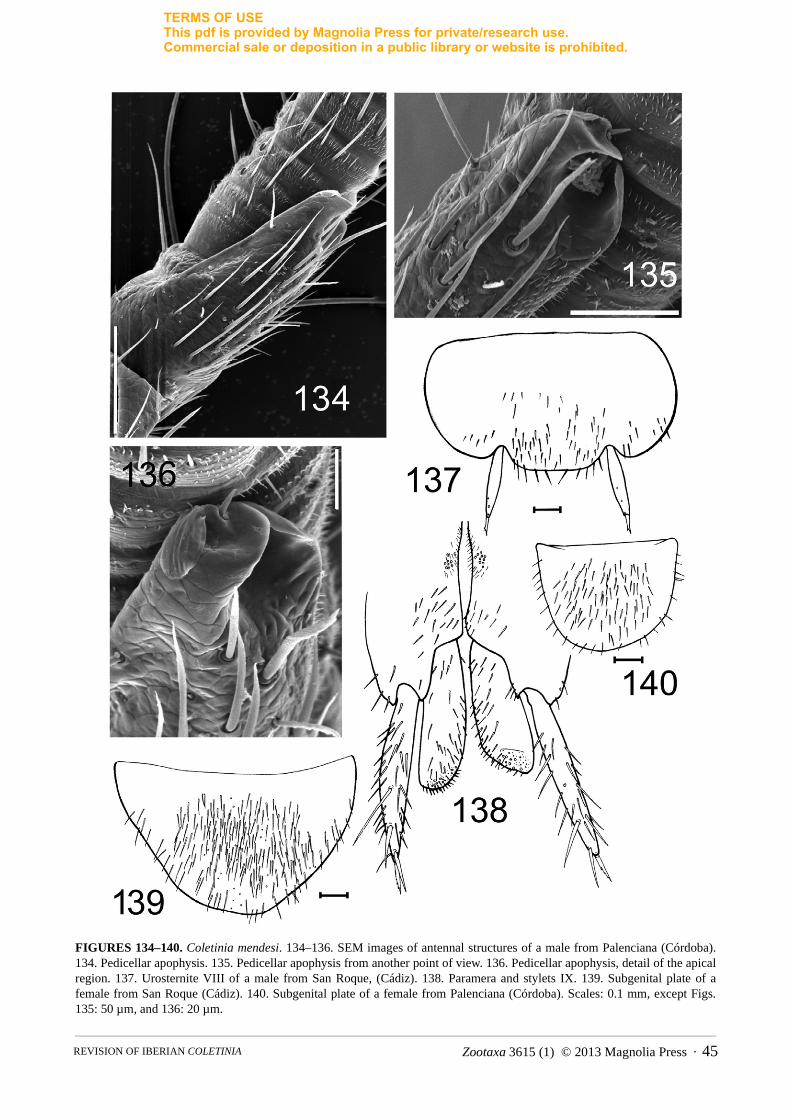
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 134–140. Coletinia mendesi. 134–136. SEM images of antennal structures of a male from Palenciana (Córdoba). 134. Pedicellar apophysis. 135. Pedicellar apophysis from another point of view. 136. Pedicellar apophysis, detail of the apical region. 137. Urosternite VIII of a male from San Roque, (Cádiz). 138. Paramera and stylets IX. 139. Subgenital plate of a female from San Roque (Cádiz). 140. Subgenital plate of a female from Palenciana (Córdoba). Scales: 0.1 mm, except Figs. 135: 50 µm, and 136: 20 µm.
Zootaxa 3615 (1) © 2013 Magnolia Press · 45REVISION OF IBERIAN COLETINIA

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 141–144. Coletinia capolongoi, pedicellar apophysis of males from Cueva del Barranco Hondo (Cheste, Valencia). 141. SEM photograph of the left apophysis. 142. SEM photograph of the left apophysis from another point of view. 143. Detail of the apical part of the apophysis showing the glandular setae (S) and the distal portion of the lamellar expansion with an indentation (I). 144. OM photograph of the apophysis of another male from the same location. Scales: 0.1 mm, except for Fig. 143: 20 µm.
MOLERO ET AL.46 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
The availability of many specimens from Córdoba has allowed us to study one of them with SEM, especially the secondary sexual characters of the male. The antennal apophysis was visualized and photographed from different angles to obtain a tri-dimensional image of this feature (Figs. 8, 134–136) and to clarify Wygodzinsky’s illustrations made with OM techniques. These photographs reveal the configuration of the subapical plate-like process; it has an internal position and is curved towards the glandular cone. The cone is also curved apically to the subapical process, delimiting a nearly circular cavity with a smooth tegument (Figs. 135, 136). The tegument is scale-like externally and basally in the basal trunk of the apophysis, and hairy (with many short setae) in the internal side, adjacent to the basal annuli of the flagellum. The setae on the external side of the trunk are longer, robust and relatively numerous (Figs. 134, 135), which results in the most distinctive appearance of the apophysis in this species. In some other aspects, the antennal apophysis of male C. mendesi is not significantly different from those species belonging to the “capolongoi” subgroup; this similarity suggests that C. mendesi (and likely, the very closely related C. setosula) are apomorphic and derive from the “capolongoi” group of species, as the diagnostic features of the “mendesi” group of species coincide with their apomorphic exclusive characters. The apical part of the apophysis attains the level of the fourth division of the flagellum, or even the demarcation between the fourth and fifth joints.
In this species, the lateral spines of the tibiae are absent in all examined specimens (Fig. 6); the discal bifid macrosetae of the urosternites also are absent, but are present in the remaining species of Coletinia studied (see Fig. 7). Another species-specific characteristic is found in the ventral spines of the mesotibiae, which are strong and longer than the diameter of the tibiae (Fig. 6). Other diagnostic features of this species are the straight hind border of the eighth urosternite in males (Fig. 137), the short paramera (approximately 3–3.5 times longer than their maximum width and slightly shorter than the ninth stylets, see Fig. 138) and the semicircular subgenital plate in females (shorter than wide at the base and with convex hind margin, but with evident intraspecific variation as shown in Figs. 139 and 140). The original description by Wygodzinsky is sufficient to distinguish this species; the examination of more specimens allows us to widen the variability of some characteristics, such as the aforementioned shape of the subgenital plate or the proportions of the maxillary palp, in which the distal article can be 4.8–6 times longer than wide and 1.3–1.5 times longer than the penultimate article.
Coletinia capolongoi Wygodzinsky, 1980Figs. 1, 2B, 3, 5, 9A, 10, 145–152
Studied material. Valencia, Cheste, Cueva del Barranco Hondo, 8 March 2003, samples BH-4 and BH-10, captured with beer + chloral hydrate traps, 2 males, 1 female and many incomplete specimens deposited in MVHN, Cod. 130208DS15: 1 male (partially dissected for SEM study) and 1 female deposited in UCO, Ref. Z2190.
Descriptive remarks. This species was known previously only from one male and one female from the Cueva de las Maravillas in Llombai (Valencia) Wygodzinsky (1980). Many specimens (some of them incomplete or in poor condition) were collected from a nearby location. Despite minor differences in some of the characteristics from those presented in the original description, these specimens are assigned to C. capolongoi because the differences are minor; some are most likely due to intraspecific variability and others to the interpretation of some features with OM and now visualized with SEM.
The size of the available specimens is slightly smaller than was given in the original description: the male length reaches 15 mm and the female length is up to 17 mm (up to 19.5 mm in the type material). Cephalic setation is shown in Fig. 1. Macrosetae are moderately spiralized; the more twisted are those inserted in the coxae and pedicels, with 4 or 5 turns (Fig. 2B). The antennal apophyses of the male were illustrated in the original description, indicating that there are “two subapical projections closely adpressed to the glandular cone.” The SEM images reveal that the structure of the pedicellar apophysis is similar to related species (Figs. 141–143); the lateral plate-like projection has an indentation that resembles a division between two parts (Fig. 143) and is not adpressed to the glandular cone as it appears from some points of view (Figs. 141 or 144), as it defines a median cavity (Fig. 142). The aspect of the antenna seen in OM (Fig. 144) is similar to Wygodzinsky’s illustrations, providing support that the examined specimens conspecific with C. capolongoi. In the available males, the apical part of the apophysis reaches the fifth joint of the flagellum.
Zootaxa 3615 (1) © 2013 Magnolia Press · 47REVISION OF IBERIAN COLETINIA

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 145–152. Coletinia capolongoi, specimens from Cueva del Barranco Hondo, Cheste (Valencia). 145. Metatibia of a male. 146. Urotergite X of a male. 147. Urotergite X of a male, detail of the hind border and posterolateral lobes with sensory pegs as visualized by OM. 148. SEM photograph of a posterolateral lobe of urotergite X showing some sensory pegs. Scale: 50 µm. 149. SEM photograph of a sensory peg. Scale: 7 µm. 150. SEM photograph of the paramera and stylets IX. Scale: 0.2 mm. 151. Urosternite VIII of a male. 152. Subgenital plate of a female. Scales: 0.1 mm except as noted.
MOLERO ET AL.48 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
The L/W ratios of the tibiae of available specimens are presented in Table 2. Each tibia has 2 dorsal, 1 lateral and 4 ventral spines; in some specimens, the metatibiae have some additional small basal spines (Fig. 145). The L/W ratio of the metatibiae drawn by Wygodzinsky appears to be approximately 7, but in the recently found specimens, the tibiae are somewhat shorter (L/W about 5). This discrepancy can be explained by different assumptions: a) a high variability in this feature in C. capolongoi, with larger specimens like those studied by Wygodzinsky having longer tibiae; b) Wygodzinsky’s drawing of the tibia is not very precise; c) the specimens now available are not conspecific with C. capolongoi. The latter hypothesis is rejected by the large number of anatomic similarities between the specimens available and those in the original description and the geographic proximity of their localities. Coletinia longitibia n. sp. has an L/W ratio very similar to Wygodzinsky’s drawings, but is different because of the different shape of the eighth urosternite in males and the higher number of sensory pegs in the tenth urotergite.
The posterior margin of the tenth urotergite of the male is straight, as originally described (Fig. 146). The number of sensory pegs in each lobe of the tenth urotergite is 4–5 in the original description and 4–6 in the available specimens. These sensory pegs were originally described as bulb-shaped and this shape as observed by OM is confirmed (Fig. 147). In SEM of one specimen, the pegs are subcylindrical, with striated tegument; these striations are subparallel and converge in a ventroapical furrow (Figs. 9A, 148, 149). This shape and structure are very similar to those of other species that we have also observed with SEM (see Figs. 9B and 9C for C. tinauti and C. intermedia n. sp., respectively).
The tenth urotergite of the female fits the original description, slightly concave at the posterior border and with the disc covered irregularly with setae. The hind margin of the eighth urosternite of the male is protruding and convex (Fig. 151).
Paramera are long, approximately 6 times longer than wide, reaching the apical fourth of the tenth stylet (Fig. 150). The subgenital plate of the female is semielliptical, rounded posteriorly (Fig. 152), wider at the base than long (L/W about 0.85). The ovipositor is as originally described; the gonapophyses have 15–16 divisions. Males have 4–6 sensory pegs in the basal part of each cercus; the shape of these pegs is almost conical, more acute apically than those of the tenth urotergite (Fig. 10) but show the same striated tegument. The paracercus possesses a row of acute sensory spines.
Coletinia tinauti Molero-Baltanás, Gaju-Ricart & Bach de Roca, 1997Figs. 1B, 6B, 9B, 153–163
Studied material. Córdoba, Los Villares, 11 July 2007, 1 male and 1 juvenile (UCO, Ref. Z2174); Córdoba, Hornachuelos, M. Baena leg., 19 October 1991, 2 males (UCO, Ref. Z2075); Córdoba; Palenciana, 5 December 2002, 2 juveniles (UCO, Ref. Z2176);
Córdoba, Alcolea, 10 September 2002, 2 males (1 dissected for SEM studies) and 2 juveniles (UCO, Ref. Z2175). The type material from Peal de Becerro (Jaén) also was re-examined.
Descriptive remarks. The availability of additional specimens of this species (some edaphic and others from caves) has enabled us to widen our knowledge of its intraspecific variability and geographic distribution. One specimen was used for SEM studies in order to make the description of C. tinauti more precise and to compare it with the remaining Coletinia spp. Characteristics sufficiently described in the original description are not commented on here, except those that require correction or redescription to be compared with related new species or for inclusion in the identification key presented below.
The body shape (Fig. 1B) is subcylindrical (length up to 11 mm) and more slender than C. mendesi (compare with Fig. 1A). The macrosetae of C. tinauti show a low degree of spiralization (Fig. 153). The antennal apophysis of the male pedicel is typical of the Mediterranean species of the group “capolongoi.” Its apical part reaches the third or fourth joint of the flagellum. The subapical plate-like expansion can be observed clearly with SEM; in the original description, it was interpreted as folds with an internal bulb, which is how it appears with OM (Fig. 154). The lateral projection and the glandular cone define a cavity with smooth tegument and the projection is folded forming a tectum (roof) that covers the cavity (Figs. 155–157). The tegument of the trunk of the apophysis appears to be more pilose than in other related species, and its scale-like rugosities are not as marked. However, detailed examination with OM of additional specimens revealed that this characteristics appears to be somewhat variable within this species. The female pedicel drawn in Molero et al. (1997, Fig. 4.1) does not exhibit the typical shape because it is collapsed, but does include the usual five macrosetae inserted into this antennal division.
Zootaxa 3615 (1) © 2013 Magnolia Press · 49REVISION OF IBERIAN COLETINIA
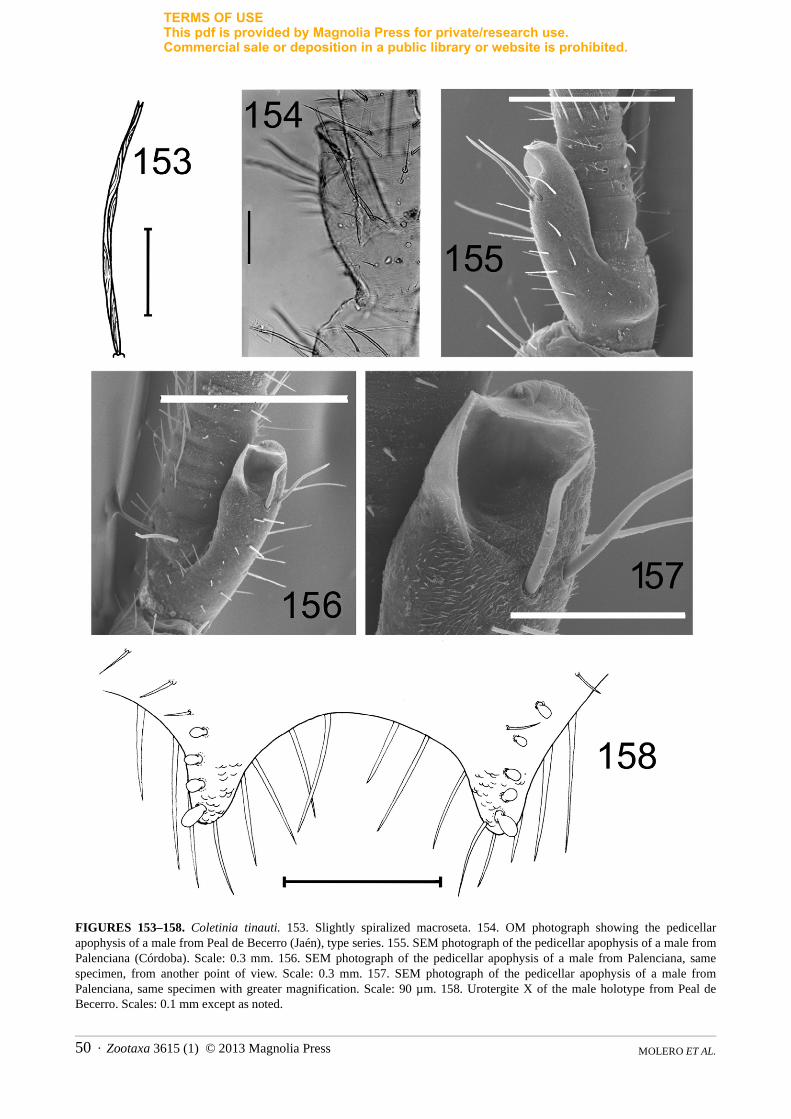
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 153–158. Coletinia tinauti. 153. Slightly spiralized macroseta. 154. OM photograph showing the pedicellar apophysis of a male from Peal de Becerro (Jaén), type series. 155. SEM photograph of the pedicellar apophysis of a male from Palenciana (Córdoba). Scale: 0.3 mm. 156. SEM photograph of the pedicellar apophysis of a male from Palenciana, same specimen, from another point of view. Scale: 0.3 mm. 157. SEM photograph of the pedicellar apophysis of a male from Palenciana, same specimen with greater magnification. Scale: 90 µm. 158. Urotergite X of the male holotype from Peal de Becerro. Scales: 0.1 mm except as noted.
MOLERO ET AL.50 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 159–163. Coletinia tinauti. 159. Urotergite X of a female paratype from Peal de Becerro (Jaén). 160. Hind border of urosternite VIII of a male from Peal de Becerro. 161. SEM photograph of the paramera of a male from Palenciana (Córdoba). 162. Subgenital plate of a female from Peal de Becerro. 163. Sensory pegs of the terminal filaments. Scales: 0.1 mm, except Fig. 161: 0.5 mm.
Zootaxa 3615 (1) © 2013 Magnolia Press · 51REVISION OF IBERIAN COLETINIA

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
The distal article of the maxillary palp is relatively short, about 5–5.75 times longer than wide and 1.15–1.45 times longer than the penultimate article.
L/W ratios of the tibiae are presented in Table 2; C. tinauti has shorter tibiae than related species, in which the metatibiae are approximately 4.5 times longer than wide. All of the tibiae have 1–3 dorsal, 1 lateral and 4–5 ventral spines, and sometimes 1–2 additional short ventrobasal spines (see Fig. 2.3 of the original description).
The tenth urotergite has a concave posterior border (Fig. 158) and 4–6 sensory pegs. These pegs are similar to those of C. capolongoi, subcylindrical, with striated tegument and a ventroapical furrow where the striations converge (Fig. 9B). Setae irregularly cover the disc of the urotergite in both sexes. In females, the posterior border is slightly concave (Fig. 159).
The eighth urosternite of the male protrudes, with its posterior border convex and rounded (Fig. 160). Paramera are relatively short, about 4-5,5 times longer than wide (Fig. 161). The subgenital plate of the female is approximately as long as wide at the base or slightly longer than wide, with a truncated and almost straight posterior border (Fig. 162). The long ovipositor long has 18–20 divisions and extends beyond the tip of the ninth stylets by 2.4–2.7 times their length. Cerci of the male have 2–6 acute sensory pegs; the paracercus lacks true pegs but possesses 1 or 2 short spines (Fig. 163).
Coletinia maggii (Grassi, 1887)Figs. 164–172
Studied material. Burgos, Cueva Grande, Atapuerca. July 2003. J. Zaragoza, V. Ortuño and A. Sendra leg., 1 male, deposited in MVHN, Cod.: 201206RV53; Burgos, Cueva de Atapuerca, 30 May 1975, A. Serra leg., 1 female, published in Molero-Baltanás et al. (1997) as Coletinia sp. VI, deposited in UCO, Ref. Z1959; Burgos, Barbadillo de Herreros, road to Monterrubio de la Demanda, 25 August 1992, inside a nest of Messor capitatus(Formicidae), 1 female, published in Molero-Baltanás et al. (1997) as Coletinia sp. VI, deposited in UCO, Ref. Z1870. A male specimen from northern Italy, lent by L. F. Mendes, also was examined.
Descriptive remarks. The specimens from Burgos agree well with the redescription by Wygodzinsky (1980). The body length of the male is 9 mm, and both females are approximately 10 mm long. Macrosetae arew moderately twisted, those of the coxa with 3–5 turns. Apophyses of the antennal pedicellus of the male are simple, subcylindrical and apically rounded (Fig. 164, also see Wygodzinsky 1980, Figs. 4A, 4B). In the male from Burgos, the apical part of the apophysis reaches the fourth joint of the flagellum. L/W ratio of tibiae are as follows: protibiae 3.1–3.8, mesotibiae 3.5–4.1, metatibiae 4–4.5. Each tibia with 1 dorsal spine inserted in the distal half, 1 lateral spine inserted basally, and 4 ventral spines. Lateral spines on the protibiae and mesotibiae are inserted more dorsally than those on the metatibiae. The tenth urotergite of the male has a nearly semicircular emargination in the posterior border between the posterolateral lobes (Fig. 165); the shape in our specimens fall within the range of variability given in Wygodzinsky (1980). The disc of this urotergite is irregularly covered with thin setae (male) and devoid of setae in females, where the hind margin is typically straight across (Fig. 166); posterolateral lobes in the male specimen have 8+8 sensory pegs arranged in two irregular rows (Fig. 167). Urosternite VIII of the male protrudes posteriorly and has a straight hind margin (Fig. 168). Paramera slightly shorter than the ninth stylets and about 4–4.5 times longer than wide (Fig. 169). Subgenital plate semielliptical, wider at the base than long (Fig. 170) and with a rounded posterior border. The ovipositor has 20 and 21 divisions in the two females available, and extends beyond the ninth stylets by 2–3 times their length (Fig. 171). The cerci of the Iberian male bear 3–4 acute sensory pegs and the paracercus has 2 spiniform basal pegs (Fig. 172).
Coletinia sp.
Studied material. Castellón, Eslida, Cova Ferrera, 11 January 2004, 1 male and 1 juvenile (both in poor condition), deposited in UCO, Ref. Z2191; Córdoba. Montoro, 12 September 1991, 1 subadult male in poor condition (UCO Ref. Z2073); Jaén, Villacarrillo, Cueva de la Morciguilla, 24 March 2002, GEV leg., 1 female (UCO Ref. Z2178); Málaga, Antequera, Cueva del Yeso III, 28 September 2008, GEV leg., 1 female (UCO Ref. Z2179); same locality, 4 May 2009, GEV leg., 1 incomplete male (UCO, Ref. Z2180).
MOLERO ET AL.52 · Zootaxa 3615 (1) © 2013 Magnolia Press
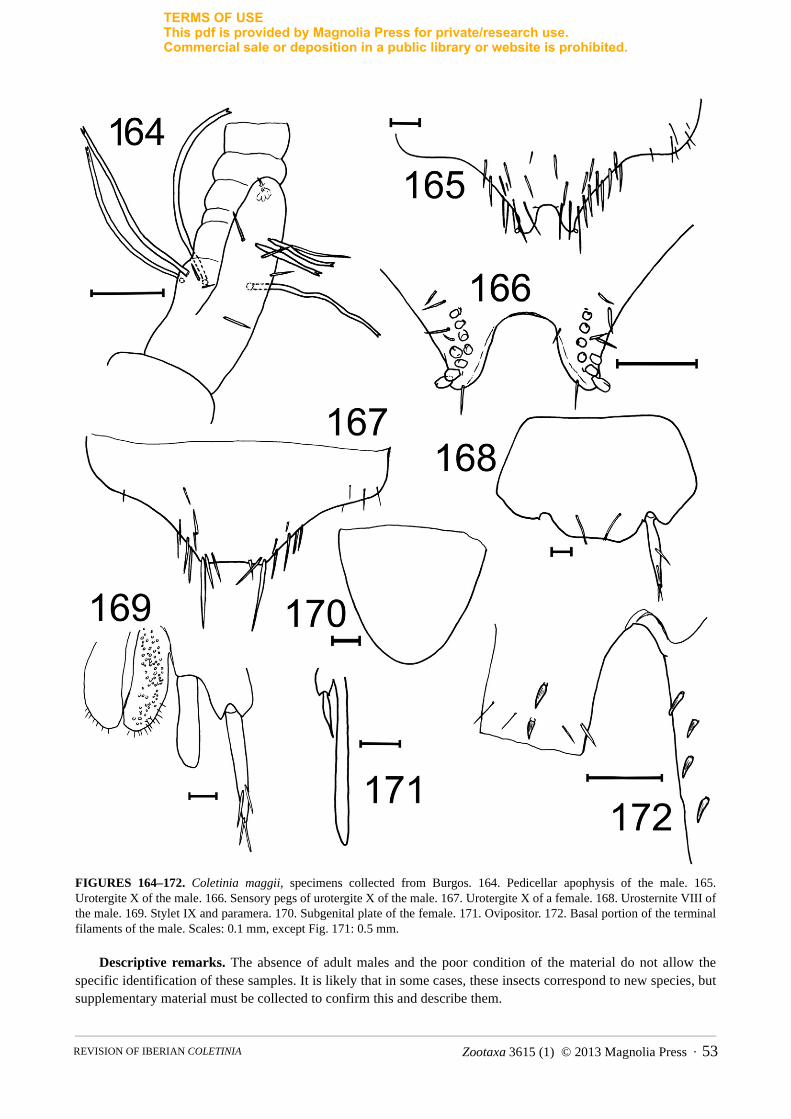
TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURES 164–172. Coletinia maggii, specimens collected from Burgos. 164. Pedicellar apophysis of the male. 165. Urotergite X of the male. 166. Sensory pegs of urotergite X of the male. 167. Urotergite X of a female. 168. Urosternite VIII of the male. 169. Stylet IX and paramera. 170. Subgenital plate of the female. 171. Ovipositor. 172. Basal portion of the terminal filaments of the male. Scales: 0.1 mm, except Fig. 171: 0.5 mm.
Descriptive remarks. The absence of adult males and the poor condition of the material do not allow the specific identification of these samples. It is likely that in some cases, these insects correspond to new species, but supplementary material must be collected to confirm this and describe them.
Zootaxa 3615 (1) © 2013 Magnolia Press · 53REVISION OF IBERIAN COLETINIA

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Discussion and general conclusions
1. Species of the “asymetrica group”. Two new species are described belonging to this group: Coletinia herculea n. sp. and C. vergitana n. sp. In addition to the presence of asymmetric apophyses in the pedicel of males (the left apophysis is much more developed than the right one), all of the species in this group also share other features, such as the cephalic setation, a long and thin ovipositor with a high number of divisions (20 or more) and, at least in the Iberian species, a convex and more or less protruding posterior border of the VIIIth urosternite in males.
The most surprising feature of both new species is the different condition of the antennae in the adult and subadult specimens. While the juvenile has nearly symmetrical apophyses, adult males have a greatly reduced right apophysis and a strongly developed left one. The following two hypotheses can be proposed:
1) The subadult specimens belong to a different symmetrical species (they are not adults because they show immature characteristics, such as the absence of sensory pegs in the tenth urotergite and terminal filaments);
2) The adult and subadult specimens are conspecific, which implies that the asymmetrical condition is a character that appears very late in the postembryonic development of C. herculea and C. vergitana. Juveniles first develop symmetrical antennae, and when they reach the sexual maturity, the left pedicellar apophysis grows and differentiates lamellae and other processes or sclerotized projections, and the right atrophies.
In this work the second hypothesis is assumed, which is supported by the following reasons:
a) The adult and subadult specimens were collected together in the same sample (locality and date);b) The adult and subadult specimens have many similar non-sexual features, such as habitus, pigmentation
and cephalic and tibial setation;c) This postembryonic change from symmetrical to asymmetrical condition of the antennal apophyses likely
occurs in other related Coletinia species, such as C. vergitana n. sp. (in this case, three specimens have been collected in the same cave, and the juvenile is symmetrical, while the adult male shows asymmetrical antennae).
However, in another related species, C. asymetrica, a subadult male specimen (length 6.2 mm) collected together with the adult holotype (8.4 mm) bears asymmetrical antennae very similar to those of the adult male (Bach et al.1985).
The finding of more specimens of these insects will clarify the doubts concerning the identity of the subadult specimen and the postembryonic development of asymmetrical antennae.
The shape of the eighth urosternite in males of C. vergitana n. sp. differs from previously known species. Another unique characteristic of the male of this species is the presence of 1+1 discal sensory pegs on the tenth urotergite. Moreover, this species from Almería shows a characteristic intense pigmentation of the condylic region of the maxillae and head.
Comparing females, Coletinia vergitana n. sp. is different from other species because it has a high number of divisions in its long ovipositor (30); only C. asymetrica bears 33–35 divisions. Females of both species have similar subgenital plates and tibial setation, but the new species has more robust setation on the tenth urotergite (Figs. 35, 36).
Unfortunately, no females of C. herculea n. sp. have yet been found, so nothing can be said about the differences between this new species and other related ones in terms of female characters.
2. Species of the “mendesi group”. Only one species of this group, Coletinia mendesi, occurs in Spain. The description of this species is improved and its geographic distribution is extended.
3. Species of the “capolongoi group”. All of the Iberian species of this group share features such as symmetrical antennae in males, cephalic setation and configuration of the male pedicellar apophysis, with lamellar expansions beside the glandular cone as photographed in Fig. 8. This type of apophysis appears only in the Iberian species of this group and in the two known species of the group “mendesi” (including C. mendesi and C. setosula). Viewed with OM, the configuration of the pedicellar apophysis in all the species of this group seems to be very similar, and
MOLERO ET AL.54 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
the apparent differences may be due to different mounting angles. The relevant details can only be discerned with SEM. These observations have been made on four species; three of these seem to be very closely interrelated, in agreement with their more meridional geographic distribution: C. tinauti, C. calaforrai n. sp. and C. intermedia n. sp. Compared with the remaining species of this group, these three species have a combination of shorter tibiae, smaller sizes and a lower degree of macrosetal spiralization. The fourth species visualized with SEM is C. capolongoi. The low number of specimens of the four remaining new species has prevented any SEM studies of these taxa.
Our comparison of apophyses of C. calaforrai n. sp. (Figs. 41–43), C. intermedia n. sp. (Figs. 54, 55), C. capolongoi (Figs. 141–144) and C. tinauti (Figs. 154–157) shows that their structure is very similar in these four species, especially when they are observed from the same point of. Only the apophysis of C. tinauti appears to be slightly more distinctive: its basal trunk is more setose, and the lateral lamella is as long as the apical cone and joins with to form a roof (tectum); this plate-like projection is shorter in C. calaforrai n. sp. and C. capolongoi. In C. intermedia the roof is not complete.
Although the structure of the pedicellar apophysis in the males does not clarify the differences among the species of this group, other characteristics allow separation of the six new species. These characters (22 in total) are summarized in Table 2 and are explained in the first section of the results of this work. The main features that are useful for each distinction are included in Tables 3 and 4. This is also complemented by the identification key that follows the discussion.
TABLE 4. Cross-wise comparison of species of the Coletinia "capolongoi" group. The numbers in each box refer to the characteristics detailed in table 2 that are most relevant to distinguish between each pair of species.
4. Species of the “maggii group”. Coletinia hernandoi n. sp. and C. maggii are the known members of this group, characterized by the subcylindrical shape of the pedicellar apophyses. Differences between the two species are discussed under the description of C. hernandoi n. sp.
This is the first report of C. maggii in the Iberian Peninsula. Few differences were observed between Iberian specimens and the central European specimens on which Wygodzinsky (1980) based his redescription of the species. Males from central Europe have a slightly different arrangement of the sensory pegs (1 row on
C. tinauti C. calaforrai n. sp.
C. intermedia n. sp.
C. diania n. sp.
C. capolongoi C. longitibia n. sp.
C. tessella n. sp.
C. redetecta n. sp.
1, 2, 4, 5, 6, 7, 8, 9, 16, 20
1, 2, 3, 4, 5, 6, 19, 20, 21, 22
1, 2, 3, 4, 5, 6, 19, 20
4, 5, 6, 12, 15, 16, 18
1, 4, 5, 6, 10, 13, 19, 21
1, 2, 4, 5, 6, 7, 8, 9, 12, 15, 19, 22
4, 5, 6, 11, 15, 16, 17, 19, 20, 21, 22
C. tessella n. sp.
1, 2, 7, 8, 9, 17, 19, 21, 22
1, 2, 3, 8, 11, 12, 14, 15, 16, 17, 20, 22
1, 2, 3, 9, 10, 11, 12, 14, 17, 19, 22
7, 8, 9, 10, 15, 16, 17, 18
1, 7-9?, 10, 17, 20
1, 3, 9, 17, 20
C. longitibia n. sp.
1, 2, 7, 8, 9, 14, 19, 20, 22
1, 2, 7, 8, 9, 14, 15, 20, 21
1, 2, 8, 9, 12, 14, 15, 19, 20
1, 7, 8, 9, 14, 16, 17, 18
7-9?, 17, 18, 21
C. capolongoi 1, 2, 9, 10, 14, 19, 20, 21
1, 2, 10, 14, 15, 17, 20
1, 2, 14, 17, 19, 20, 21
1, 8, 9, 15, 16, 18
C. diania n. sp.
1, 2, 7, 8, 9, 10, 16, 18
1, 2, 3, 10, 12, 15, 16, 17, 18
2, 3, 12, 15, 16, 17, 18
C. intermedia n. sp.
3, 7, 8, 9, 10, 12, 17, 18, 22
10, 13, 19, 21, 22
C. calaforrai n. sp.
3, 7, 8, 9, 17, 19, 21, 22
Zootaxa 3615 (1) © 2013 Magnolia Press · 55REVISION OF IBERIAN COLETINIA

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
the tenth urotergite, 4 or 5 blunt pegs on the cerci and more than 2 sensory spines in the basal region of the paracercus), and females have a shorter subgenital plate, according to Fig. 5H in Wygodzinsky (1980). In comparison of Fig. 164 with Wygodzinsky’s Figs. 4B and 4C, the antennal process of the Spanish male seems to be somewhat wider than those from central Europe. The number of divisions of the ovipositor in Iberian females (20–21) is less than that described by Wygodzinsky (25). Nevertheless, we believe that these differences are not substantial enough to consider the Iberian specimens as a different species, taking into account the intraspecific variability in many of the characteristics reported in Wygodzinsky’s description. The availability of additional new specimens from different countries and molecular complementary studies will clarify the specific identity of the Iberian insects. Coletinia maggii is widespread in central Europe (Paclt & Christian, 1996; Mendes, 1992) from northwest Italy to Hungary, but there are no reports from southern France, so there is a gap of more than 1000 km between the Iberian localities and the nearest previously known populations. This gap may indicate an important geographic isolation or perhaps only the scarcity of samplings of endogean Nicoletiidae.
The known distributions of Iberian Coletinia spp. are given in Fig. 173. This map is certainly provisional, as new prospecting will likely provide specimens to describe additional new taxa and enlarge the geographic distribution of the previously known species. Also, as commented previously, more material is needed to adequately describe the intraspecific variability of these taxa to ensure they can be adequately differentiated from each other, and especially for studies on the phylogeny of these edaphic or cave-living insects.
FIGURE 173. Distribution map of the known species of Coletinia in the Iberian Peninsula. 1—Coletinia maggii. 2—C. mendesi. 3—C. capolongoi. 4—C. asymetrica. 5—C. tinauti. 6—C. herculea n. sp. 7—C. vergitana n. sp. 8—C. calaforrai n. sp. 9—C. intermedia n. sp. 10—C. diania n. sp. 11—C. tessella n. sp. 12—C. longitibia n. sp. 13—C. redetecta n. sp. 14—C. hernandoi n. sp. The numbering is consistent with the chronological order of their original description. Squares indicate new reports, circles indicate previous reports. Samples identified as Coletinia sp. are not included.
MOLERO ET AL.56 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Identification key for Iberian Coletinia species
1. Head with numerous subequal strong setae, lacking bifid macrosetae (Fig. 4). Ventral macrosetae of tibiae strong and (at least in mesotibiae) clearly longer than the diameter of the tibia (Fig. 6). Lateral spine of tibiae absent. Discal macrosetae of uroster-nites absent . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. mendesi
– Head with dispersed small setae and few regularly arranged bifid macrosetae (Fig. 3). Ventral macrosetae of tibiae slender, usually shorter or as long as the diameter of the tibia. Lateral spine of tibiae usually present. Discal macrosetae of urosternites present . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
2. Males . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3– Females . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 153. Antennae asymmetrical, the apophysis of the left pedicellus in adult specimens more developed than the right . . . . . . . . . . . 4– Antennae symmetrical, the pedicellus of both antennae with similarly developed apophyses . . . . . . . . . . . . . . . . . . . . . . . . . 64. Posterior margin of eighth urosternite not very protruding and slightly convex (Fig. 37). Apophysis of the left pedicellus of
adults as in Figs. 28–30, with a sclerotized region next to the glandular cone and a subapical lateroexternal process very well-developed, showing no sclerotizations but densely covered with small hairs . . . . . . . . . . . . . . . . . . . . . . . . C. vergitana n. sp.
– Posterior margin of eighth urosternite more protruding and convex (Figs. 25, 160). Apophysis of the left pedicellus of adults different; if bearing a lateral process, it is hook-shaped and not densely pilose . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5
5. Apophysis of the left pedicellus of adults with a sclerotized crest in the subapical zone of the basal trunk and two short bifid macrosetae (Fig. 128). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .C. asymetrica
– Apophysis of the left pedicellus of adults with two lamellar processes, one apical without sclerotization, and one subapical with hook-shaped sclerotization and with a long and strong macrochaeta plus a few thin setae (Figs. 11, 12) C. herculea n. sp.
6. Pedicellar apophysis simple, cylindrical, with no plate-like structures (Fig. 119) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7– Pedicellar apophysis complex, with plate-like (lamellar) subapical expansions and/or sclerotizations next to the glandular cone
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87. Pedicellar apophyses relatively wide and short (Fig. 164); their L/W ratio less than 2.5; base of the flagellum less than twice
the width of the apophysis. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. maggii– Pedicellar apophyses more narrow and long (Fig. 119) their L/W ratio more than 2.5; base of the flagellum more than 2 times
the width of the apophysis. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. hernandoi n. sp.8. Posterior border of urosternite VIII protruding, convex and rounded (Fig. 112); if somewhat straight or truncated in the median
region, then fewer than 6+6 sensory pegs on the tenth urotergite . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9– Posterior border of urosternite VIII protruding but straight (rounded truncate) in its median region (Fig. 97). Sensory pegs of
the tenth urotergite 6+6 or more (Fig. 95) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. tessella n. sp.9. Metatibia with at least 6 ventral bifid spines (Fig. 107). No more than 3 sensory pegs on terminal filaments C. redetecta n. sp.– Metatibia with 4–5 ventral bifid spines; if 6 or more, sensory pegs of terminal filaments very numerous. Cerci with at least 4–5
pegs each . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1010. Larger species, body length of adults surpassing 13 mm. Metatibiae usually long, L/W ratio greater than 6.5 except in some
specimens of C. capolongoi . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14– Smaller species. Metatibiae short, L/W ratio clearly less than 6.5. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1111. Hind border of tenth urotergite straight between the posterolateral lobes or slightly concave (Figs. 70, 147) . . . . . . . . . . . . 12– Hind border of the tenth urotergite clearly concave and rounded between the posterolateral lobes (Figs. 47, 95, 158) . . . . . 1312. At least 5 sensory pegs in the paracercus; cerci with more than 6 pegs. Paramera short, no more than 4.5 times longer than wide
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. diania n. sp.– Paracercus with few sensory pegs (0–2), cerci with no more than 6 pegs, often fewer. Paramera longer, usually 5 or more times
longer than wide . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. intermedia n. sp.13. Tibiae relatively short, L/W ratio of mesotibiae less than 4 of metatibiae less than 4.5. Eighth urosternite with very convex hind
margin (Fig. 160) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. tinauti– Tibiae relatively long or slender, L/W ratio of mesotibiae 4 or more, of metatibiae 4.5 or more. Eighth urosternite variable,
sometimes with rounded hind margin (Fig. 49) but usually less convex, more straight and truncated posteriorly . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. calaforrai n. sp.
14. L/W ratio of metatibiae greater than 6. Paramera about 5 times longer than wide. Posterolateral lobes of urotergite X with 8 sensorial pegs each . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. longitibia n. sp.
– L/W ratio of metatibiae variable, greater than 6 in larger specimens (18 mm or more). Paramera about 6 times longer than wide. Posterolateral lobes of urotergite X with at most 6 sensorial pegs each. . . . . . . . . . . . . . . . . . . . . . . . . . . . C. capolongoi
15. Ovipositor long, with more than 25 divisions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16– Ovipositor with fewer than 25 divisions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1716. Ovipositor with about 30 divisions. Robust and dense setae covering disc of tenth urotergite. Subgenital plate wider at the base
than long (Fig. 38) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. vergitana n. sp.– Ovipositor with 33–35 divisions. Setae of the disc of the tenth urotergite scarce and thin (Fig. 131). Subgenital plate as long as
wide at the base . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. asymetrica17. Posterior border of the tenth urotergite straight, the disc lacking setae (Fig. 167) . . . . . . . . . . . . . . . . . . . . . . . . . . . C. maggii– Posterior border of the tenth urotergite slightly concave, the disc with more or less abundant setae . . . . . . . . . . . . . . . . . . . 1818. Posterior border of the subgenital plate rounded-truncate, nearly near the midline (Fig. 162) . . . . . . . . . . . . . . . . . . . . . . . . 19– Posterior border of the subgenital plate arched, convex, semielliptical (Fig. 87) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2219. Ovipositor short, surpassing the apex of the ninth stylets by less than their length (Fig. 99). Macrosetae mounted in Tendeiro
Zootaxa 3615 (1) © 2013 Magnolia Press · 57REVISION OF IBERIAN COLETINIA

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
medium with a high degree of spiralization . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. tessella n. sp.– Ovipositor longer, usually surpassing the apex of the ninth stylets by more than their length; some specimens of C. calaforrai
with shorter ovipositor but macrosetae mounted in Tendeiro medium with a low degree of spiralization . . . . . . . . . . . . . . . 2120. Ovipositor with a low number of divisions (less than 17). Subgenital plate relatively short, L/W ratio no more than 0.9 . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. calaforrai n. sp.– Ovipositor with more than 17 divisions and surpassing the apex of the ninth stylets by at least 1.5 times their length. L/W ratio
of the subgenital plate 0.9 or more . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2121. Metatibiae not conspicuously long, L/W ratio about 4.5 or lower. Very scarce and dispersed setae covering disc of tenth
urotergite (Fig. 159). Ovipositor long, surpassing the apex of the ninth stylets by at 2 or more times their length . . . C. tinauti– Metatibiae with L/W ratio clearly more than 4.5. Setae of the tenth urotergite scattered irregularly all over the disc (Fig. 58).
Ovipositor shorter, surpassing the apex of the ninth stylets by 1.5–2 times their length . . . . . . . . . . . . . . . C. intermedia n. sp. 22. Tibiae very long, L/W ratio of the metatibiae greater than 6.5. Ovipositor with about 18 divisions. Body length up to 17 mm .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. longitibia n. sp.– Tibiae shorter, L/W ratio of the metatibiae clearly less than 6; if longer (only in large specimens more than 18 mm long) bear-
ing ovipositor with 15–16 divisions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2323. Ovipositor with more than 22 divisions. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. hernandoi n. sp.– Ovipositor with a low number of divisions (20 or less). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2424. Body length of adults usually more than 12 mm. Macrosetae mounted in Tendeiro medium with high spiralization, usually
with 4–5 or more turns . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26– Body length less than 12 mm. Macrosetae mounted in Tendeiro medium slightly spiralized, no more than 2–3 turns . . . . . 2525. Ovipositor with 17 or more divisions. Disc of the tenth urotergite covered rather densely with setae (Fig. 58) . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. intermedia n. sp.– Ovipositor with 16 or fewer divisions. Disc of the tenth urotergite with very scarce setae (Fig. 48) . . . . . . C. calaforrai n. sp.26. Ovipositor with more than 18 divisions. Tibiae usually with 6 macrosetae and additional short spines (Fig. 107). Subgenital
plate longer than wide at the base or as long as wide (Fig. 114) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. redetecta n. sp.– Ovipositor with fewer than 17 divisions. Tibiae usually with 4 macrosetae, apart from additional short spines (Fig. 145). Sub-
genital plate wider at the base than long (Fig. 152) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . C. capolongoi
Acknowledgements
We thank Dr. Luis F. Mendes from IICT (Lisbon) for the loan of the material of C. mendesi and C. asymetrica, and for scientific advice. We are grateful to Dr. Alberto Tinaut for transfer of the type material of C. herculea n. sp. and to Carlos Hernando for providing the type material of C. hernandoi n. sp. We also express our gratitude to the biospeleologic group GEV from Villacarrillo (Jaén) that has provided us with some of the material from caves and to Manuel Baena, who provided us with some samples and allowed us to be in contact with GEV and Carlos Hernando. We also thank Manuel Urbano for the design of some of the figures describing C. tinauti, C. redetecta, C. capolongoi and C. mendesi.
Samplings of material collected by A. Sendra and S. Montagud were financed by the Consellería de Territori i Habitatge, Servei de Conservació i Gestió de la Biodiversitat, Generalitat Valenciana, within the Project “I+D a la conservación de los Invertebrados cavernícolas de la Comunidad Valenciana”. Some samplings and research works were financed with the support of Spanish MCYT “Fauna Ibérica VII Project” (REN2001-1956-C17-05).
Finally, we wish to thank the Spanish Education and Science Ministry for the funding of the project CICYT 1FD97-1577 to sample the caves of the Paraje Natural del Karst en Yeso de Sorbas where C. calaforrai n. sp. was found. We also thank Carmen Ruiz Portero and Ángel Fernández Cortés, who took the samples.
References
Bach de Roca, C., Mendes, L.F. & Gaju Ricart, M. (1985) Sur une nouvelle espèce et une nouvelle citation de Nicoletiinae de Sierra Morena (Cordoue, Espagne) (Nicoletiidae Zygentoma). Bollettino della Società Entomologica Italiana, Genova, 117 (8–10), 132–140.
Chopard, L. (1924) Description d'un Nicoletia de Corse (Thys. Lepismidae). Bulletin de la Société Entomologique de France, 16, 174–176.
Escherich, K. (1905) Das System der Lepismatiden. Zoologica (Stuttgart) 43, 1–164.Grassi, B. (1887) I progenitore dei Miriapodi e degli Insetti. Altre recerche sui Tisanuri. Nota preliminare. Bullettino della Società
Entomologica Italiana, 19, 52–74.Kozaroff, G. (1939) Eine neue Nicoletiaart in Bulgarien. Bull. Soc. Ent. Bulgarie, Izvestiia na Bulgarskoto Entomologichno Dru-
zhestvo, 10, 45–50.
MOLERO ET AL.58 · Zootaxa 3615 (1) © 2013 Magnolia Press

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Mendes, L.F. (1981) Nova nota sobre os tisanuros (Apterygota, Microcoryphia e Zygentoma) da Europa e da bacia mediterranica. Boletim da Sociedade Portuguesa de Entomologia, 18, 1–9.
Mendes, L.F. (1985) Nota preliminar sobre os tisanuros (Microcoryphia e Zygentoma) do Algarve (Portugal). Boletim da Sociedade Portuguesa de Entomologia, 1 (suppl. 1), 239–262.
Mendes, L.F. (1988) Sur deux nouvelles Nicoletiidae (Zygentoma) cavernicoles de Grèce et de Turquie et remarques sur la systémat-ique de la famille. Revue Suisse de Zoologie, 95 (3), 751–772.
Mendes, L.F. (1992) Some new data on the Nicoletiidae (Insecta: Zygentoma) from Europe and Asia Minor. Revue Suisse de Zoologie, 99 (4), 821–834.
Mendes, L.F. (1996) Further data on the Nicoletiidae (Zygentoma) with description of a new species from Mauritius. Revue Suisse de Zoologie, 103 (3), 749–756.
Mendes, L.F. & Ferreira, R.L. ( 2002) On a new cave-dwelling Nicoletiidae (Zygentoma : Insecta) from Brazil. Garcia de Orta, Série de Zoologia, Lisboa, 24, 101–106.
Molero-Baltanás, R., Gaju-Ricart, M. & Bach de Roca, C. (1997) Coletinia tinauti n. sp., nueva especie de Nicoletiidae de Andalucía (Insecta, Apterygota, Zygentoma). Zoologica Baetica, 8, 95–110.
Molero-Baltanás, R., Gaju-Ricart, M. & Bach de Roca, C. (2000) On the taxonomic use of the distribution pattern of the antennal asteriform sensilla in Neoasterolepisma and Tricholepisma (Insecta, Zygentoma, Lepismatidae). Pedobiologia, 44, 248–256. http://dx.doi.org/10.1078/S0031-4056(04)70045-1
Paclt, J. & Christian, E. (1996) Die Gattung Coletinia in Mitteleuropa (Thysanura: Nicoletiidae). Deutsche Entomologische Zeitschrift, 43, 275–279. http://dx.doi.org/10.1002/mmnd.19960430211
Parona, C. (1888) Res Ligusticae. VI. Collembole e Tisanuri finora riscontrante in Liguria. Annali del Museo Civico di Storia Naturale di Genova, Ser. 2a, 6, 133–154.
Silvestri, F. (1902) Materiali por lo studio dei Tisanuri. III. Nuove specie di Nicoletia. Bollettino della Società Entomologica Italiana,33, 223–227.
Silvestri, F. (1908) Materiali per lo studio dei Tisanuri. X. Su alcuni Tisanuri di Corfu. Bollettino del Laboratorio di Entomologia Agraria, Portici, 2, 381–393.
Silvestri, F. (1938) Descrizione di una nuova specie di Nicoletia vivente in una grotta della Francia (Insecta Thysanura). Revue Française d'Entomologie, 5, 188–193.
Ubero-Pascal, N., Fortuño, J.M. & Puig, M.A. (2005) New application for air-drying techniques for studying Ephemeroptera and Ple-coptera eggs by Scanning Electron Microcopy. Microscopy Research and Technique, 68, 264–271. http://dx.doi.org/10.1002/jemt.20248
Wygodzinsky, P. (1980) A survey of the Nicoletiidae of Europe (Nicoletiidae, Thysanura, Insecta). American Museum Novitates, 2695, 1–24.
Zootaxa 3615 (1) © 2013 Magnolia Press · 59REVISION OF IBERIAN COLETINIA

TERMS OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
About the authors
Doctor Rafael Molero Baltanás is currently working as Associated Professor of Zoology at the University of Córdoba (Spain) and has been investigating the biology and taxonomy of the order Zygentoma since 1987. His Ph. D. Thesis dealt with the Zygentoma fauna of Spain, and since then, working together with Carmen Bach and Miquel Gaju, he has described more than twenty new species of insects belonging to the orders Zygentoma and Microcoryphia, and clarified the status of several previously known species of Lepismatidae with very ancient and obsolete descriptions. He has also concentrated his work on the biology of those Zygentoma species living with ants or in human dwellings.
Prof. Dr. Carmen Bach de Roca is an Entomologist. She studied Biology at the University of Barcelona. From 1966 till 1975 she worked at the University of Salamanca where she got her PhD (1971). From 1975 till 1979 she worked at the Autonomous University of Barcelona (UAB); after her habilitation as Professor (1979) she moved to the University of Cordoba where created the Department of Zoology and was head of it. In 1992 she moved again to the UAB where she worked till 2011 and now is in contact with this University. Her principal area of professional research in all Universities is the systematic, biology and biogeography of the orders Microcoryphia and Zygentoma on which subject she has written over 150 papers. Later she focused her work also in control of termites (Isoptera)
Doctor Alberto Sendra got his Ph.D. in Biology in 1988 at the University of Valencia (Spain). His main research interests are the taxonomy, ecology and biogeography of subterranean fauna, with main expertise in a peculiar and barely unknown arthropod group called Diplura. He has published more than fifty papers, edited six books and described more than thirty new taxa, including new species and genus for science, from different regions of the planet. Among his scientific contributions is included the discovery of the world’s deepest subterranean invertebrate community in world's deepest cave (Krubera-Voronja) and the description of the remarkable bizarre troglobiont Gollumjapyx smeagol.
Sergio Montagud studied Biology at the University of Valencia. He is currently working on his PhD project in collaboration with the Institute of Evolutionary Biology (CSIC-UPF) in Barcelona. Now he is associate professor of Zoology at the University of Valencia and curator of its Natural History collections. Previously he has been keeper and researcher at the Natural History Museum of Valencia. Its working groups are Lepidoptera and Coleoptera as taxonomy, and subterranean wildlife and endangered species as ecology. He has written about 20 articles, three books and several chapters on these topics.
Doctor Pablo Barranco obtained his Ph.D. in Biology in 1997 at the University of Granada (Spain) working on Dociostaurus maroccanus locust plague. Actually he teaches Applied Entomology and Pest Management at the University of Almería (Spain). His fields of study are pest control, insect physiology, subterranean fauna and taxonomy. His main groups in taxonomy are orthoptera, trombidoidea acari and palpigradi. He is author or co-author of more than 140 journal articles, books and book chapters. He has described twenty six new taxa, including new species and genus for Science, mainly Iberian Fauna but also from different regions of the planet as Neotropical region and Australia.
Doctor Miquel Gaju Ricart got the Ph. D. in Biology in 1985 at the University of Córdoba (Spain). His research has two lines of interest: taxonomy, biology and biogeography of Microcoryphia and Zygentoma and biology and control of termites (Isoptera). He has published more than fifty papers and described more than twenty new taxa (genera and species). At the present time he is assistant at the Zoology Department in the Córdoba University, teaching on Zoology and Insects Biology.
MOLERO ET AL.60 · Zootaxa 3615 (1) © 2013 Magnolia Press




















