
Review of the main morphological characters of the palaearctic notodontid larvae (Lepidoptera,...
18
Lambillionea, CIII. 4. Decembre 2003 607 REVIEW OF THE MAIN MORPHOLOGICAL CHARACTERS OF THE PALAEARCTIC NOTODONTID LARVAE (Lepidoptera, Notodontidae) COMMUNICATION 1. Irina V. DOLINSKAYA & Igor G. PLJUSHCH * * Schmalhausen Institute of Zoology, National Academy of Sciences of Ukraine, B. Chmielnicky str., 15, UA 01030 Kiev, Ukraine Key words: Lepidoptera. Notodontidae, larva. first instar, morphology. chaetotaxv. Palaearctic. Abstract: This is the first communication about palaearctic notodontid larvae. 70 species belonging to 40 genera were examined. Morphological characters of larvae of the first instar have been described and illustrated the comparative aspect. Zusammenfassung: Die erstc Mitteilung iiber palacarctische Raupe (Fam. Notodontidae) ist vorgelegl. Insgesammt 70 Arten aus 40 Gattungen sind untersucht. Die morphologische Merkmale (nebst Abbildungen) von Raupen der 1. Lebenalten sind bctracht. Resume: Premiere comunication concernant les chenilles de Notodontidae palearctiques. Les chenilles de 70 especes appartenant а 40 genres ont ete examinees. Les caracteres morphologiques des chenilles du ler age sont illustres et analyses de maniere comparative. Introduction This paper concerns comparative study of the palaearctic notodontid moths based on the main larval morphological characters of the first instar of 70 species from 40 genera. The reason is that the morphological peculiarities in the early instar are of taxonomic importance and often use to clear up the relationships in this family, whereas middle instar and mature larvae have many similar characters because of their high adaptability to the environment. Moreover, the morphology of the mature larvae of the Notodontidae has been studied in a series of the works The best recent works are these of GODFREY & APPLEBY (1987) about caterpillars of NORTH American specics Nolodontidae and MILLER (1991),which contains cladistics and classification of the Notodontidae based on the detailed morphological investigations larvae and imago. ln 1996 J. MILLER published the work concerning phylogeny of the Neotropical Notodontidae (Dioptinae: Josiini) with detailed descriptions of imago, larvae and pupae of the mentioned Lepidoptera. Several good illustrated atlases of Japanese Notodontidae with brief descriptions of the larvae (HATTORI, 1969; NAKATOMI, 1987) were also published. Bcsides these generalized checklists there are some papers with mоrе or less detailed descriptions of the several species of Notodontidae from different regions of the world. (GARDNER, 1943, 1946; NAKATOMI, 1977; DOLINSKAYA et al„ 1983; SUGI et al., 1988; GARCIA CONDE et al., 1994 and others). ln comparison with mature larvae morphology, tbe first instar larva remain poorly studied. PACKARD'S (1895) monograph is one of the first well-known for us publications with the descriptions and figures of the first instar larvae of Notodontidae. FENILI (1956) gave the detailed description of the first instar iarvae of Cerura vinula (L.). WELLER'S (1987) paper contains descriptions of the first instar larvae of the North American notodonlid Litodonta hydromeli Harvey. Joshi's et al. (1989) described the first instar larva of the Indian notodontid Clostera fulgurita Wlk. and C. restitura Wlk.
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Review of the main morphological characters of the palaearctic notodontid larvae (Lepidoptera,...

Lambillionea, CIII. 4. Decembre 2003 607
REVIEW OF THE MAIN MORPHOLOGICAL CHARACTERS OF THE PALAEARCTIC NOTODONTID LARVAE
(Lepidoptera, Notodontidae)
COMMUNICATION 1.
Irina V. DOLINSKAYA & I g o r G. PLJUSHCH *
* Schmalhausen Institute of Zoology, National Academy of Sciences of Ukraine, B. Chmielnicky str., 15, UA 01030 Kiev, Ukraine
Key words : Lepidoptera. Notodontidae, larva. first instar, morphology. chaetotaxv. Palaearctic.
Abs t rac t : This is the first communication about palaearctic notodontid larvae. 70 species belonging to 40 genera were examined. Morphological characters of larvae of the first instar have been described and illustrated the comparative aspect.
Z u s a m m e n f a s s u n g : Die erstc Mitteilung iiber palacarctische Raupe (Fam. Notodontidae) ist vorgelegl. Insgesammt 70 Arten aus 40 Gattungen sind untersucht. Die morphologische Merkmale (nebst Abbildungen) von Raupen der 1. Lebenalten sind bctracht.
Resume: Premiere c o m u n i c a t i o n concernant les chenilles de Notodontidae palearctiques. Les chenilles de 70 especes appartenant а 40 genres ont ete examinees. Les caracteres morphologiques des chenilles du ler age sont illustres et analyses de maniere comparative.
Introduction
This paper concerns comparative study of the palaearctic notodontid moths based on the main larval morphological characters of the first instar of 70 species from 40 genera. The reason is that the morphological peculiarities in the early instar are of taxonomic importance and often use to clear up the relationships in this family, whereas middle instar and mature larvae have many similar characters because of their high adaptability to the environment. Moreover, the morphology of the mature larvae of the Notodontidae has been studied in a series of the works The best recent w o r k s are t he se of GODFREY & APPLEBY ( 1 9 8 7 ) abou t ca te rp i l l a r s of NORTH A m e r i c a n specics Nolodontidae and MILLER (1991),which contains cladistics and classification of the Notodontidae based on the detailed morphological investigations larvae and imago. ln 1996 J. MILLER published the work concerning phylogeny of the Neotropical Notodontidae (Dioptinae: Josiini) with detailed descriptions of imago, larvae and pupae of the mentioned Lepidoptera. Several good illustrated atlases of Japanese Notodontidae with brief descriptions of the larvae (HATTORI, 1969; NAKATOMI, 1987) were also published. Bcsides these generalized checklists there are some papers with mоrе or less detailed descriptions of the several species of Notodontidae from different regions of the world. (GARDNER, 1943, 1946; NAKATOMI, 1977; DOLINSKAYA et a l „ 1983; SUGI et al. , 1988; GARCIA CONDE et al . , 1994 and others). ln comparison with mature larvae morphology, tbe first instar larva remain poorly studied. PACKARD'S (1895) monograph is one of the first well-known for us publications with the descriptions and figures of the first instar larvae of Notodontidae. FENILI (1956) gave the detailed description of the first instar iarvae of Cerura vinula (L.). WELLER'S (1987) paper contains descriptions of the first instar larvae of the North American notodonlid Litodonta hydromeli Harvey. Joshi's et al. (1989) described the first instar larva of the Indian notodontid Clostera fulgurita Wlk. and C. restitura Wlk.

608 Lambillionea, CIII, 4, Decembre 2003
In the present paper the review of the family based on the characters of the first instar larvae is represented, whereas in the next communication keys to the genera and specics will be given.
Materials and Methods
The work is based on the original materials collected by the authors on the territory of the former USSR (Ukraine, Kazakhstan, northern Kyrghyzstan and Primorski Krai
(Far East. Russia). Females were collected by UV light traps. Emerged larvae were placed into the cold water which then was boiled. After that the caterpillars were fixed in 75% alcohol and then studied with using a binocular microscope MBS-9.Ten specimens of each species have been studied. Terminology for larval morphology, chaetotaxy of the head and ten abdominal segments one follows HLNTON (1946), GERASIMOV (1952) and Stehr (1987). Hinton's (1946) terminology was retained to discuss the body chaetotaxy. The taxonomical arrangement of the Notodontidae follows SCHINTLMEISTER (1989). ln this paper we did not incorporate the descriptions of the mouth parts, not did we
incorporate information on proprioceptor setae because of the minute size of larvae. Such investigations need in using SEM. То this problem will be devoted a separate paper.
Descriptions
General external characters. Body (figs 1-12) mostly cylindrical tapering towards А9-А10; rarely conical, gradually becoming smaller from A2 through A10 (Pygaera Ochs.). Most of the Iarvae are of usual structure, without protrusions or humps; sometimes indistinct tubercle occupies the area of A8 (Spatalia Hbn., Notodonta Ochs. and others). However, in some cases body shape is atypical. For example, larvae of Harpyia Ochs. саn be distinguished by the presence of the well expressed humps in the areas of tergites А1-А6 and A8. Humps of А1-А8 tergites are bifurcated at the apex (fig. 4). Larvae of Stauropus Germar and Cnethodonta Stgr. with well developed double humps on А1-А5 tergites and besides that pleurites on А1-А7 are broadened looking like conical humps. A8 in these genera distincly bulbar and joined lightlv with A9-10;these segments are oval-rounded having a conspicuous shape looking like turtle shell (figs 9, 10). Larvae of Harpyia, Stauropus and Cnelhodonta with large lateral humps in the area of A8-A9.
Cuticle of notodontid larvae mostly shining and impellucid but species of some genera (Gluphisia Bsdv., Semidonta Stgr., Drymonia Hbn. and others) have pellucid cuticle through which pulsating dorsal vessel is visible. Many larvae with body of light ground-colour and with dark, brown or black, strongly pigmented head, prothoracic shield, anal shield, pinacula, setae. Thoracic legs and lateral side of ventral prolegs. Much rarely these structures pigmented by light
colours, less sclerotized becoming almost indistinct on the main ground-colour of the body (Gluphisia, Euhampsonia Dyar. Epodonta Mtsm.). It should be noted that decrcase of sclerotization of the mentioned structures often is correlated with cuticular weakness. Cuticle of notodontid larvae mostly smooth, but species of some genera (Pygaera, Clostera Samouelle, Dicranura Reichenbach and other) have cuticle with numerous small spines. Coloration and pattern. Body coloration mainly monotonous, light. predominantly yellowish-white or greenish white (Epodonta, Gluphisia, Pheosia Hbn. and others), sometimes also lemony-yellow (Ellida Grote), greyish-green (Pygaera) or velvetish-black (Cerura Schrank). Some genera wilh mixed coloration. For example, larvae of Lophontosia Stgr. are pale green with darkish-pink A1, A3 and A7; larvae of Fentonia Butl. with greenish-yellow thoracic and 4-th-5-th abdominal segments while

Lambillionea, СШ, 4, Decembre 2003 609
the rest one are reddish-brown; larvae of Harpyia are dark claret with lemony-yellow spots on tergites of A2, A5-A6. At the end of the first instar a slight pattern contour of the mature larvae appears. In the genus Stauropus mediodorsal and subdorsal lines both slightly visible; larvae of Hexafrenum Mtsm. have dorsal brown line, larvae of Phalera Hbn. with distinct subdorsal, stigmatal and basal lines. Head of orthognatic type, usually rounded wiih slightly expressed vertexal tops. Head position is free one, except of genera Cerura and Furcula Lamarck, with posterior part of the head being retracted into prothorax. Parietal suture of the most Notodontids somewhat exceeds (by 1/3-1/4) frontal length (figs 13, 15). However, sometimes it can be on the contrary (Dicranura, Shaka Mtsm., Nerice Walker and others) (fig, 19). Head cuticle usually impellucid except of Gluphisia and Dicranura which epicranial and occipital parademes are distincly visible through cuticle. Most of the notodontid larvae have unicolour head, dark brown or black (Pheosia, Spatalia Hbn, Nerice and others), sometimes of paler tints fusing with the main ground-colour of the body, or whitish-green (Drymonia, Gonoclostera Butl.) or yellowish-white (Gluphisia, Epodonta). In the other cases there is a head pattern. For instance, larvae of Pterostoma griseum Br. have whitish head with slight brownish infumation and with light brownish dots. Larvae of Lophocosma Stgr. can be recognized by light green head marked with slight brown tint and by brown marginal and two parallel middle stripes (figs 17, 18); Iarvae of Semidonta with brown head edges while the rest spacc is yellowish-brown. Stemmata as in a majority of lepidopterous insects arranged in an arched row, where either 6 stemmata located in the row (figs 16, 18) or 4 stemmata in the row and 2 apart (figs 20, 21). In this area there is often a dark smear (fig. 18). Mouthparts. Some works are devoted to the morphology of larval mouthparts (GODFREY, 1 9 8 4 ; 1990 ; GODFREY e t al . , 1 9 8 9 ; MILLLER, 1 9 9 1 ) In t h e s e w o r k s o n l y mature larvae were studied. Mouthparts of the first instar larvae of some Notodontid species have been been investigated by WELLER, 1987 and DOCKTER, 1993. Prothoracic shield (figs 22-29) usually dark brown or black, strongly sclerotized (Notodonta, Pheosia, Pterostoma Germar and others).Sometimes it is slightly scierotizes fusing with ground-colour of body, then its shape can be recognized only by located there setae (Gonoclostera, Lophocosma, Epodonta). Prothoracic shield in the most species elongated, narrow (about 2.5-5.0 times as long as broad), usually with smooth-rounded margins and its hind margin (sometimes anterior one also) with small groove. Anterior margin of prothoracic shield mostly longer than posterior one and sometimes elevated (Spatalia, Lophocosma., Epodonta and others). Some genera differ by largc lateral projections directed upwards and forwards. For example, Harpyia larvae have anterior margin of prothoracic shield with long, slender, spinulated projcctions (fig.27). In Cerura and Furcula genera such projections are large, wide, with many conical setaceous tubercles (fig.28), whereas in Hemifentonia Kiriakoff, Fentonia and Wilemanus Nagano genera they are accordingly smaller and without spinulae and tubercles (fig. 29). Anal shield (figs 30-37) usually of the same colour and degree of sclerotization as prothoracic one or paler and less distinct. In the most cases il is targe, occupying almost completely tergal surface. Posterior margin of anal shield more or less smooth-rounded, elongated; anterior one broadened, trapeziform, often with a small, rounded or acute-angled medial cut. Sometimes anal shield universely trapeziform (Micromelalopha Nagano); in Cerura and Furcula genera it is extremely elongated along its longitudinal axis (fig.32). In the other cases (Allodonta Stgr., Phalera and other) there are enlarged seta. locatcd on large pinacula which imitate an undulating surface of proximal margin (figs 30, 31). Preanal lobes (paraproctes) raether well developed, representing more or less largc papilliform projections which in apical part are brown, each with a stout seta. The bcst expressed paraproctes in Cerura and Furcula larvae being farge, sabreform.

610 Lambillionea, CIII. 4. Decembre 2003 610
protruding outside of the anal shield (fig.32). In genera Harpyia paraproctes are abscnt. Setae of notodoniid Iarvae uniform, hairlike, mainly small, brown, more or less distinct usually 1.5-4.0 times (Pheosia, Lophontosia, Semidonta and other) (fig. 1 )or even 4.0-6.0 times (Dicranura, Wilemanus, Stauropus) (fig. 9) shorter than A1 along dorso-ventral axis. Sometimes setae as long as A1 height, or its length 1/3-1/4 exceeds the last one (Micromelalopha,Ptilodon ladislai Oberth). The longest setae are noted in Hexafrenum and Allodonta genera (fig. 3) being 2.0-3.0 times as long as A1 (along dorso-ventral axis). Setae disposed on larval body are of various size, coloration and strength. For
example. Fentonia species have the most long and stout sctae arranged on the anal prolegs (fig. 6), whereas Phalera species, besides that. have similar setae both on prothoracic and anal shields (fig. 7), Notodontid larvae often have the longest and stoul D1, D2 and SD1 of thoracic and abdominal segments while the rest of them
mоrе small and slender. For instance, Euhampsonia species have more stout, dark D1 and D2 of thoracic and abdominal segments, also SD1 of thoracic and 8-th-9-th abdominal segments, also X-seta of 9-th abdominal segment. The other setae are whitish and thin being indistinct on the ground-colour of body. Pinacula of this most notodontid species dislinct, large. strongly sclerotized. pigmented (black or brown). Sometimes they without dark pigmentation, slightly sclerotizes fusing with the body ground-colour (Euhampsonia, Epodonta and others). In some cases pinacula very small and scarcely visiblc (Stauropus, Harpyia, Cerura) Pinacula mostly of different size. Usually pinacula of D1 and SD1 of thoracic and abdominal segments are the largest ones (Micrometalopha, most of species Plilodon and other) (Fig. 2). Only D1 of thoracical and abdominal segments of Phalera. Spatalia and Uropyia Stgr.have the largest pinacula (figs 7, 8). D1, D2 and L1 of A8 Stauropus species have the largest pinacula. Sometimes only D1of A8 (Pheosia, Ellida and others) with the largest pinacula (fig. 1. For the majority species of Notodontidae is of character pinacula with small, slightly cxprcssed elevation in the middle (Fentonia, Pterostoma and other). More rarely this elevation distinct and larger (Spatalia, Uropyia, Allodonta) (figs 3. 8). In_Pygaera (fig. 5) pinacula are espccially distinct, of large size with large elevations in the centre and some of them have two setae, Sometimes such elevations appear independently without pinacula (Ptilodon cucullina Schiff.) (fig. 12). lt must be noted that during the development of the mentioned structure there is а gradual transition from slightly expressed pinacula with indistinct elevation in the middle to the well expressed pinacula like in the genus Pygaera. It is very interesting to note that in the 2-nd instar Pygaera specics have instead of pinacula only verrucae covered with numerous setae. Probably pinacula in the first instar of Pygaera may be treated as prototype of verrucae. In the most of Notodontid species pinacula since the 2-nd instar became much smaller and slightly developed,
Chaetotaxy. Chaetotaxy of Notodontidae in general is similar to that of Noctuidae (Merzheevskaya, 1967). Some information concerning chaetotaxy of Notodontid larvae of the first inslar is given in the above mentioned works and also noted by Hinton (1946) and Gerasimov (1952). In particular Hinton (1946) characterized chaetotaxy of the first instar larva of Cerura vinula L.; including descriptions of tactile and proprioceptor setae. Head chaetotaxy is common. Sometimes additional setae are present. For example, there are two addilional stout setae on the middle part of the head of Uropyia (flgs 15, 16). Head of Harpyia larvae covered with numerous small secondary setae while the primary one invisible among such great number of setae. Primary setae mostly of moderate length, distinct. stout. brown (Ellida, Pheosiopsis Bryk, Drymonia), sometimes elongated (figs 13, 14. 20, 21). Rather often setae are small, slender.

Lambillionea, CIII. 4. Decembre 2003 5
whitish and almost indistinct (Dicranura. Micromelalopha, Lophocosma and other) (figs 17-19). Prothoracic segment. Like in the most of Lepidoptera D1, D2, XD1, XD2 located on the large prothoracic shield: D1 and D2 occupy its posterior part. XD1 and XD2 -
anterior one (figs 23, 25, 26). Divergence of setae disposition insignificant, that is when D2 located anleriorly (fig.24) or when XD1 and XD2 are placed closely on verrucae together with additional seta (fig. 22). Modification of chaetotaxy correlated also with the development of the projections of prothoracic shield, with removing selac XD1 and XD2 which located there (figs 27-29). Sometimes (Harpyia) XD1 transposed to thc middle ot this shield (fig. 27). Setae of prothoracic shield usually are rnore or less long. XD1 and XD2 1.5-3.0 limcs as long as shield breadth (Notodonta, Euhampsonia, Pterostoma and others), or 4.0-9.0 times (figs 22, 23), equal to its width or their length 1/3-1/4 exceeds shield breadth (figs 24-26). Sometimes 1.5-2.0 times shorter than shield breadth (fig. 27). In the other cases there are numerous secondary setae which are of the same length and toughness that primary one (fig. 5). D1 and D2, as in the most of Lepidoptera, distinctly more slender and shorter than XD2 and XD1. SD1, SD2, L1, L2, SV1 and SV2 mostly arranged in pairs located on common pinaculum (figs. 5, 8). Sometimes these sctae are separated from each other and located on isolated pinacula (figs 4, 6). Rarely SD1, SD2 and also L1, L2 are locatcd оn trisetose verrucae (fig. 7). As usual, notodontid larvae have SV1 and SV2. Sometimes the number of setae varies from 2 till 4 (fig. 7). HINTON (1946) at Cerura vinula L. discovered 3 proprioceptor setae- MXD1, MV2 and MV3.
Meso - and metathoracic segments. D1, D2, located either in pairs or these ones are separatcd from each other. Thus in Pheosia, Pygaera and others D1 and D2 located on large common pinaculum (figs 1, 2, 5). In Hexafrenum, Fentonia, Clostera and otber these setae are separated from each other and located separately (figs 3, 6). Usually these setae characterized by vertical position that is typical for Lepidoptera. However, D2 of Stauropus larvae which located on T2 directed somewhat forwards, whereas on T3 it is directed backwards (fig. 9). In the most cases setae SD1 and SD2 are separated from each other. Sometimes SD1 and SD2 iocated in pairs on common pinaculum (Phalera bucephala).Usually these sctae characterized by vertical posilion, however in Harpyia SD2 directed somewhat forwards (fig. 4). L group is rcpresented only by a single L1. As usual, notodontid larvae have only SV1 (figs 1, 2. 12). Sometimes SV2 also is present (figs 4, 6. 11). Sometimes on those segments, as well as on prothoracic ones, several verrucaes are present. Thus in the area of D1 (Phalera) appear three setose verrucae. D2 and L -bisetose ones. HINTON (1946) found out the following proprioceptor setae on the meso-and metathorax in Cerura vinula - MD1, MD2, (the latter on the metathorax is absent), MSD1, MSD2, MV1, MV2 and MV3.
HINTON (1946) noticed that MD-group on the metathorax which consists of two sctae is of great interest being an argument of relationship of the many families. Unfortunalely, he invesligatcd Cerura vinula taking it in account as example for the family, and therefore based his conclusions concerning the relationships of Notodontidae and the other families using the characters of this species. We consider that Cerura species are not good examples for such discussion as the mentioned genus is very specialized, and that is why cannot be used for the total discussion of the family. 1-9-th Abdominal segments. Setae of these segments are rather well developed, mostly of common position. rarely slight disposition is observed. For example, larvae

612 Lambillionea, CIII, 4, Decembre 2003
of Harpyia, Stauropus, Cnethodonta which have tubercles on the abdominal segments are characterized by transposition of D1 and D2 on the mentioned tubercles, and the lattcr scta displaced ventrally (figs 4, 9). SD1 of Leucodonta larvae on A7-A8 dispaced ventrally, SD2 was invisible in the all species studied by us. In Harpyia genus on А1-А8 in the area of SD2 disposition we found out 1-3 additional setae (one of which larger and identical to the primary setae) (fig. 4). Hinton (1946) recorded this large seta on 1-8 abdominal segments of Cerura vinula. Miller(199I) studied SD2 with SEM. Basing on his investigations he concluded that "this seta is morphologically identical to abdominal MD proprioceptor. Following by interpretation of Kitching (1984), Singh and Goel (1987) he named SD2 as MSD2. ' S D 1 , L1 and L2 on A7-A8 of Dicranura separated from each other by the rather big space (comparing wiith the rest examined species). As it is known (Hinton, 1946) L3 is subprimary seta, and that is why it is absent among all studied species. L group on A9 is unisetose. HlNTON (1946) designated this seta as L1 for Notodontidae, Sphingidae and other. STEHR (1987) noted that on A9 the L group is often unisetose or bisetose; L1 is always present.GERASIMOV (1952) have noted that often it is difficult to elucidate which seta of L-group is present. Among studied by us caterpillars of the first instar this seta by its structure and disposition has intennediate position between L1 and L2.However in the 2-nd instar above mentioned characters are close to those of L2. Evidently, studying of musculature can elucidate this problem. Number of SV setae usually varies. There can be on A1 only SVl (Pheosia, Notodonta, Shaka and other) or SV1 and SV2 (Cnethodonta, Allodonta, Lophontosia and other).On the A2 present mostly SV1 and SV2 (Peridea, Hagapteryx and other). Sometimes in the areas of subventral group therc are additional secondary very small setae (Phalera, Spatalia and other). For example, А1-А2 of Spatalia with 3-8 setae, whereas Cerura erminea accordingly with 4-5 setae. Additional secondary setae in the areas of subventral group of setae are present on the lateral tubercle of A8-A9 (Harpyia, Slauropus) where their number varies from 6 to 13 (figs 4, 9). Additional setae appear also in the area of the other setae, In Pygaera timon and Phalera bucephala L. D1 is bisetose on segment A1 through A9 (flgs 5, 7) HlNTON (1946) noticed a presence of three proprioceptor setae on A1 - MDl, MD2 and MV3 and A2-A9 - MD1 , MV3 at Cerura vinula. 10-th Abdominai segnient1. Chaetotaxy of anal shield (figs 30-37) are uniform for the most of notodontid larvae. D1 occupies medial part of shield somewhat above its middle; sometimes both of D1 located on two together fused tubercles (Notodonta jankowski Oberth.). D2, SD1 and SD2 mostly arranged proportionally along posterior margin of shield. Often SD1 replaced towards D2 (Ptilodon ladislai. Dicranura ulmi Schiff. and other); such replacement in the best way expressed in Stauropus species (fig. 35); sometimes setae are concentrated in the posterior part of shield (Harpyia) (fig. 37). In the other cases (Phalera, Clostera, Pygaera) there is additional seta. located near D2 (figs 30-33). Stehr (1987) called it "EXD3" On the anal shield of many species setae as long as body length; in the most of genera the longest is D2, ils length 1/3-1/4 exceeds shield breadth (Leucodonta,
Micromelalopha, Fentonia). Larvae of Clostera albosigma curtuloides Erschoff have the longest SD1 and SD2 which 4.5-5.0 times as long as breadth of anal shield and 3.0-4.0 times as long as thc rest setac. Intersegmental X seta located on the antero-lateral corner of the anal shield (figs 1-13). F.STEHR (1987) and then J. MlLLER (1991) used term E-area for designation а disposilion of this seta. Lateral setae group (Ll, L2, L3) located in the middle part of the leg; on the lateral part- SV2, SV4 and SV1 (figs 38, 39, 43, 44, 45), SV3 and VI located on the inner
' Chaetotaxy of (this segment has distinct diagnostic characters for genera and species, which in more details will be discussed in our next communication.

Lambillionea, C I I I . 4. Decembre 2003 613
part of the leg (fig. 40). On the preanal Lobe placed preanal seta РР1 (figs 38, 39. 43-45, 47). Disposition of lateral setae-group mostly constant -L1 in the upper leg part, L3 is located somewhat inferiorly and laterally, L2 placed more inferiorly than previous setae at the middle of the leg (flg. 45). L1 and L3 can be located almost on the s a e level (fig.38, 39). SV4 and SV2 located on the proximal and SVl - on the distal edge of the leg. It should be noted that larvae with modificated anal legs have primary chaetotaxy of the leg invariable. (fig. 43). Phalera bucephala has two additional setae on the external side of leg as well as on the inner one (fig. 46). Uropyia has primary chaetotaxy slightly changed. SV4 and V1 are located on the inner part of the leg. SV4 are located on the proximal part of the leg and V1 -on the distal one (flg. 47). The rest of the setae are transfered on the lateral part of the leg. Moreover SV2 and SV1 locatcd on sides of the leg and parallel to each other. Harpyia has 9 setae on the anal leg, 4 of which located on the external part of the leg and 5 setae - on the inner one (figs 41, 42). Thoracic legs usually of constant shape and with typical arrangement of setae.
Hoowever, in some genera those characters are more or less modified. For example, Stauropus and Cnethodonta larvae have meso-and metathoracic legs considerably elongated, 4.0-6.0 times as long as А1 Mesothoracic legs of these genera about 1/5 as long as metathoracic ones. Along all of the leg surface there are numerous small setae
placed on tubercles (fig. 9). Legs of the most of notodontid larvae are of the same coloration as body, with distinct large, black or brown pinacula. Some of the representatives of Epodonta, Euhampsonia, Phesiopsis genera with pinacula of light colour, thus fusing with general body ground colour. Ventral prolegs. Meso-ventral prolegs well developed; any cases of their reductions are known. l-st-4-th pairs of ventral prolegs with more or less distinct lateral pinacula which mainly are large, occupy the most part of proleg surface, often with rounded groove on the anterior margin (Semidonta, Shaka, Hagapteryx and other). Rarely pinacula are indistinct because of the same with proleg ground-colour (Ptilodon ladislai, Euhampsonia, Gonoclostera). Sometimes pinacula are visible only on certain prolegs. For instance, Fentonia and Hemifentonia genera have distinct, large brown pinacula only on 1-st and 4-th pairs of proleg, while on 2-nd and 3-rd pairs they are fused with ground-colour. In the area of lateral pinacula there are numerous secondary setae among which setae of SV-group are indistinct. Sometimes the latter being long and stout are well visible (Allodonta, Shaka). Total number of setae located on lateral pinacula varies from 6 to 17. Rather often secondary setae arranged in a straight line along inferior margin of pinacula (Leucodonta, Lophontosia, Lophocosma). Anal prolegs characterized by their different structure. For example, in some genera prolegs transformed into appendices which do not take part in the body rcmoving. Above mentioned modifications can be of various kinds. For instance, anal prolegs become more slender and clongated (Fentonia, Hemifentonia, Wilemanus) (fig. 6); considerably decreascd in their size and disposed parallel to body axis (figs 4, 41), in the same time books on the foot and primary chaetotaxy are preserved. Then they transform into small preanal furca (Phalera, Uropyia), where hooks are absent, but primary chaetotaxy is present yet (figs 7, 8, 46, 47): after that preanal furca becomes extremely elongated (fig. 11) exceeding body length and accompanied by development of glandular filaments (Cerura, Furcula). In the genera Stauropus and Cnethodonta these structures transform into projections tbat resemble palpae (figs 9. 10) and where hooks of the foot are absent but the numerous secondary setae appear. Such some transformed structures recorded by Gerasimov (1952) and Holloway (1983). The latter consider that this character is enough good to separate the subfamilies. Such modification of anal proleg structure can be explained by the fact of lacking their main ability to push on the body. Instead of this above mentioned larvae hold upwards these appendices of prolegs as well as the last abdominal segments.

614 Lambillionea, CIII, 4, Decembre 2003
The proleg base of the most of notodontid larvae is a subspherical planta (figs 48-50), while at Cerura the planta is redused outside. The planta base has clearly expressed muscle present as dark spot (fig. 51), Crochets of notodontid larvae disposed on thc planta inside, in uniordinal mesal penellipse (figs 48, 49. 51). Heteroideous mesoseries is typical for Dicranura, Clostera, Pygaera species, where hooks are minute in the medial part of range, being more large along the edges (fig. 50). The number of crochets disposed in mesal penellipse varies from 10 to 30 (usually to 20). For example, in Lophocosma and Phalera species this number is 10-17, while in Pheosia, Pterostoma and the rest genera accordingly is 15-30. Crochets disposed in heteroideous mesoseries have accordingly 10-16. The number of crochets which occupy an anal pair of proleg is the less one as usual. Phesiopsis larvae with 16-21 crochets on l-st-4-th pairs of ventral prolegs and with 13-17 on the anal pair, Hagapteryx larvae have accordingly 14-17 and 9-12 crochets. Besidcs that, the number of crochets somewhat varies on the different prolegs. For instance, in Phesiopsis larvae this correlation consequently is following: 23-29; 24-30; 24-31; 25-30 and 18-23.
Acknowledgements
We wish to express our thanks to Dr. M. OMELKO (Gorno-Tayozhnaya Stantsiya, Primorski Krai. Russia) for his kind assistance during our field work in Primorski Krai and granting us their stationary facilities. We are obliged to Mr. 1. K O S T Y U K (Zoological Museum of the Shevchcnko National University in Kiev), to Dr. A. SCHINTLMEISTER (Germany), to Dr. G. L. GODFREY (Center for Biodiversity , Illinois Naturai History Survey) and to Dr. J. S. MILLER (American Museum of Natural History) who kindly have sent us the reprints of their works.

Lambillionea, C I I I . 4. Decembre 2003 615
References
DOCKTER D. E. 1993. Developmental changes and wear of larval mandibles in Heterocampa guttivilta and H subrotata (Notodontidae). J. Lep.. Soc. 47 (1): 32-48. DOLINSKAYA I. V., LOMAKIN M. D.. 1983. Morphology of the immature stages of the Notodonta dromedarius L.(Lepidoptera, Notodontidae). Vestnik zooologii 4: 61 -65. FENILI G., 1956, Contributo alla conoscenza morfobiologica della Dicranura vinula L. (Lepidoptera, Notodontidae). Redia. 41: 343-510. GARCIA CONDE M.R. SCHMIDT G. H. 1994. Das Borsten-und Porenmuster der Raupenstadien des Pinienprozessionsspinners Thaumetopoea pityocampa (Den. & Schiff.) (Insecta: Lepidoptera: Thaumetopoeidae) aus Sudgriechenland. Zool. Jb. Syst. 121: 374-406. GARDNER J. С. M.. 1943. lmmature stages of lndian Lepidoplera. Indian. J. Entomol. 5: 89-102. GARDNER J. С. M., 1946. Immature stages of lndian Lepidoptera. Ibid. 7: 139-144. GERASIMOV A. M. 1952. Larvae of Lepidoptera of U.S.S.R. Fauna U.S.S.R.. insects: Lepidoptera, vol. 1. no. 2. Zool. Inst. Acad. Nauk USSR (n. ser.): 338 pp. [in Russian]. GODFREY G. L.. 1984. Notes on the larva of Cargida pyrrha (Notodontidae).J. Lep. Soc. 38 (2): 88-91. GODFREY G L. .1990. Larval mandible of Cargida pyrrha (Notodontidae). J. Lep. Soc. 44 (2): 93-94. GODFREY G. L., APPLEBY J. E.. 1987. Notodontidae (Noctuoidea): the notodontids and prominents. In F. W. STEHR (ed.). Immature insects, pp. 524-533. Dubuque, lowa: Kendall/Hunt. GODFREY G. L., MILLER J. S. and CARTER D. J.. 1989. Two mouthpart modifications in larval Notodontidae (Lepidoptera): their taxonomic distributions and putaiive tunctions. J. New York Entomol. Soc. 97 (4): 455-470. HATTORI I., 1969. Notodontidae. In S. ISSIKI (rev.), Early stages of Japanese moths in colour. vol.ll, Osaka: Hoikusha [in Japanese]. HINTON H. E.. 1946. Оn the homology and nomenclature of the setae of Lepidoplerous larvae with some notes on the phylogeny of Lepidoptera. Trans. R. Ent. Soc. London 97: 1 -37. HOLLOWAY J. D., 1983. The moths of Borneo: family Notodontidae. Malay. Nat. J. 37:1-107.. JOSHl К. C., GURUNG D. & SARMA Р. C. 1989, External morphology of the larvae of Closlera
fulgurita Wlk. and Clostera reslitura Wlk. (Lepidoptera: Notodontidae). Indian Forester I 15 (10): 754-761. KITCHING I. J., 1984. The use of larval chaetotaxy in butterfly systematics, with special reference to the Danaini (Lepidoptera, Nymphalidae). Syst. Entomol.9: 49-61. MERZHEEVSKAYA О. I., 1967. Larvae of Owlet Moths (Noctuidae), Biology and morphology (Key) Minsk: Science and technics, 1967.: 452 pp .[in Russian]. MILLER J. S., 1991. Cladistics and classification of the Notodontidae(Lepidoptera: Noctuoidea) based on larval and adult morphology, Bull. Amer. Mus. Nat. Hist. 204: 1-230. Fig. 541. Tabl. 8. MILLER J. S.. 1996. Phvlogeny of the Neotropical moth tribe Josiini (Notodontidae: Dioptinae): a hidden case of Mullerian mimicry. Zool. J. Linn. Soc. 118: M5. Fig. 23. NAKATOMI K.. 1977. A new subspecies of Dudusa sphingiformis from Korea and Tsushima Island with description of the last instar larva (Lepidoptera, Notodontidae). Ent. Rev. Japan. 30 (1/2): 41-42. NAKATOMI K., 1987. Notodontidae In S. Sugi (ed.), Larvae of Larger Moths in Japan. 453 pp. Kodansha [in Japanese wilh English summary]. PACKARD A.. S., 1895. Monograph of the bombycine moths of America north of Mexico including their transformations and origin of the larval markings and armature., part 1, Notodontidae. Mem. Nat. Acad. Sci.: 291 pp , plate XLIX. SCHINTLMEISTER A., 1989. Zoogeorraphie der palaearktischen Notodontidae (Lepidoptera). Neue Ent.
Nachr. 25:3-117. SINGH G. P. and GOEL S. C„ 1987, Body chaetotaxy of three noctuid caterpillars. Disch. Entomol. Z, 34: 373-381. STEHR F. W„ 1987. Order Lepidoptera. In F. W. STEHR (ed.), Immature insects. pp.288-305. Dubuque, Iowa: Kendall /Hunt.
SUGl S., NAKAMURA M., 1988. Taxonomic and biological notes on Periergos magnus (Matsumura) (Notodontidae), with descriptions of its larva and pupa. Japan. Heteroc. J. 150: 385-389 [in Japanese with English summary]. WELLER S. J., 1987. Litodonta hydromeli Harvey (Notodontidae): description of life stages. J. Lep.Soc.41 (4): 187-194.

616 Lambillionea, CIII, 4, Decembre 2003
Figs 1-9. Habitus of larval Notodontidae in lateral view. 1. Pheosia gnoma. Scale line = 0.25 mm
2. Ptilodon capucina. Scale line = 0.25 mm 3. Hexafrenum leucodera. Scale line = 0.25 mm

Lambillionea, CIIl, 4. Decembre 2003 617
4. Harpvia umbrosa. Scale line = 0.25 mm 5. Pygaera timon. Scale line = 0.25 mm

618 Lambillionea, CIII, 4, Decembre 2003
6. Fentonia ocypete. Scale line = 0.25 mm 7. Phalera bucephala. Scale line =0.5 mm 8. Uropyia meticulodina. Scale line = 0.25 mm

Lambillionea, CIII, 4. Decembre 2003 619
9. Stauropus fagi. Scale line = 0.25 Fig. 10. Habitus of larva Slauropus fagi in dorsal view. Scale line = 0.5 mm Figs 11-12. Habitus of larval Notodontidae in lateral view. 11. Cerura erminea. Scale line — 0.25 mm 12. Ptilodon cucullina. Scaie line = 0.25 mm

620 Lambillionea, CIII, 4, Decembre 2003
Fig 13. Head of Allodonta plebeja in frontal view. Scale line = 0.15 mm. Fig. 14. Head of Allodonta plebeja in lateral view. Scale line = 0.15 mm. Fig. 15. Head of Uropyia meticulodina in frontal view. Scale line = 0.15 mm. Fig. 16. Head of Uropyia meticulodina in lateral view. Scale line = 0.15 mm Fig. 17. Head of Lophocosma atriplaga in frontal view. Scale line = 0.15 mm Fig. 18. Head of Lophocosma atriplaga in lateral view. Scale line = 0.15 mm Fig. 19. Head of Dicranura ulmi in frontal view. Scale line = 0.15 mrn. Fig. 20. Head of Hexafrenum leucodera in lateral view. Scale line = 0.15 mm Fig. 21. Head of Ptilodon ladislai in lateral view. Scale line = 0.15 mm

Lambillionea, CIII. 4. Decembre 2003 621
Figs 22-29. Prothoracic shield in dorsal view. fig. 22. Phalera bucephala. Scale line = 0.25 mm
fig. 23. Allodonta plebeja. Scale line =0.15 mm fig. 24. Stauropus fagi. Scale line = 0.15 mm fig. 25. Hagapteryx admirabilis. Scale line = 0.15mm fig. 26. Pheosiopsis cinerea. Scale line = 0.15 mm fig. 27. Harpyia umbrosa. Scale line = 0.15 mm fig. 28. Cerua erminea. Scale line = 0.15 mm fig. 29 Hemifentonia mandschurica. Scale line = 0.15 mm
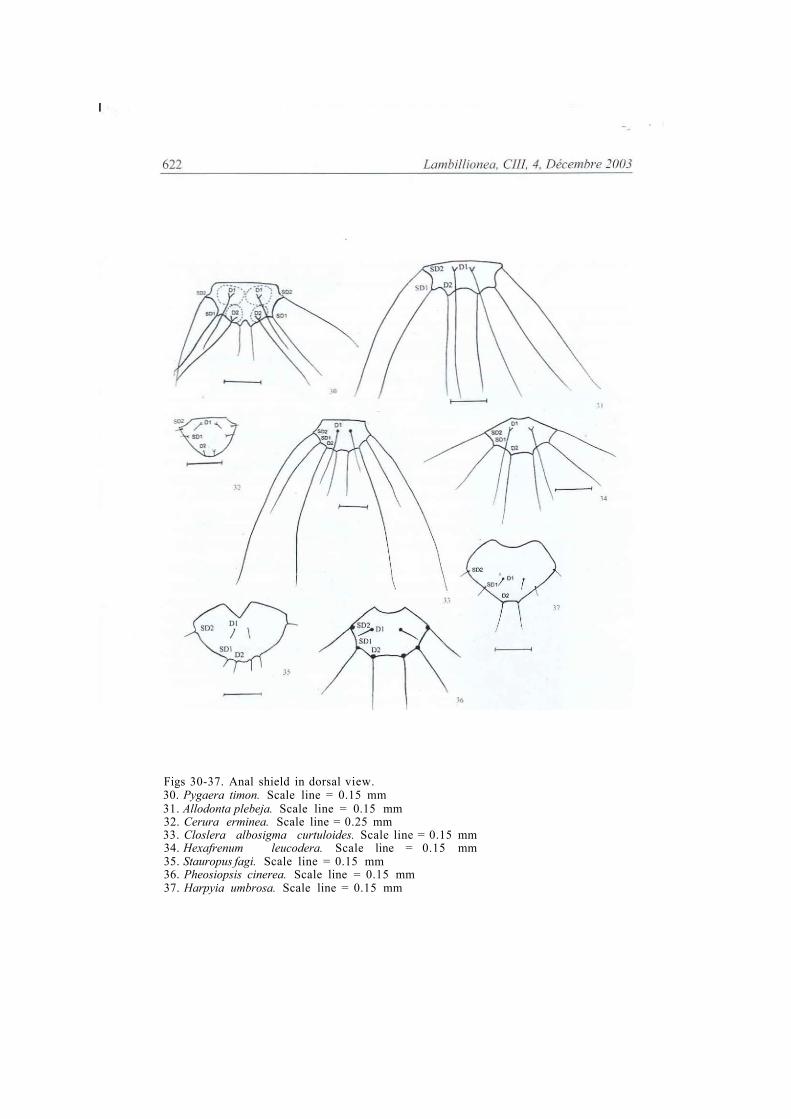
Figs 30-37. Anal shield in dorsal view. 30. Pygaera timon. Scale line = 0.15 mm 31. Allodonta plebeja. Scale line = 0.15 mm 32. Cerura erminea. Scale line = 0.25 mm 33. Closlera albosigma curtuloides. Scale line = 0.15 mm 34. Hexafrenum leucodera. Scale line = 0.15 mm 35. Stauropus fagi. Scale line = 0.15 mm 36. Pheosiopsis cinerea. Scale line = 0.15 mm 37. Harpyia umbrosa. Scale line = 0.15 mm

Lambillionea, CIII, 4. Decembre 2003 623
Figs 38-39. Abdominal segment 10 from the outside. 38 Pheosiopsis cinerea. Scale line = 0.15 mm 39. Lophocosma atriplaga, Scale line= 0.15 mm Fig. 40. Anal proleg of Lophocosma atriplaga from the inside. Scale line = 0.15 mm Fig. 41. Abdominal segment 10 ot Harpyia umbrosa from the outside. Scale lin = 0.15mm Fig.42. Abdominal segment 10 of Harpyia umbrosa from the inside. Scale line = 0.15 mm Fig 43. Abdominal segment 10 of Dicranura ulmi in dorsal view. Scale line = 0.15 mm. Figs 44-45. Anal proleg from the outside. 44. Lophontosia cuculus, Scale line - 0.15 mm 45. Shaka atrovitlata. Scale linc = 0.15 mm 46. Phalera bucephala. Scale line = 0.15 mm 47. Uropyia meticulodina. Scale line = 0.15 mm

624 Lambillionea, CIII, 4, Decembre 2003
LEFT VENTRAL PROLEGS
-CEPHALAD CAUDAD lateral margin
49
48
Figs 48-51. Planta and the crochet arrangements 48. Ellida branickii. Scale line = 0.15 mm 49. Harpyia milhauseri. Scale line = 0.15 mm 50. Clostera albosigma curtuloides. Scale line = 0.15 mm 5 I. Cerura erminea. Scale line - 0.1 5 mm




















