
Relaciones entre el bacterioplancton y fitoplancton en el Amazonas Venezolano.
24
Boletín del Centro de Investigaciones Biológicas ISSN 0375-538X ~ Vol. 47. Nº 3, Septiembre-Diciembre 2013, Pp. 235 - 239 Recibido / Received: 18-07-2013 ~ Aceptado / Accepted: 29-10-2013 Relaciones entre el fitoplancton y el bacterioplancton en el río Cataniapo, estado Amazonas, Venezuela Simón Astiz 1,2 , Haymara Álvarez 1 y Mariana Villaró 1,3 1 Instituto de Recursos Naturales, Universidad Simón Bolívar, Apartado 89000. Caracas 1080-A, Venezuela. [email protected] 2 Doctorado en Ciencia y Tecnologías Ambientales, Universidad Autónoma de Barcelona. Bellaterra, Barcelona 08193, España. 3 Maestría en Ciencias Biológicas, Universidad Simón Bolívar, Apartado 89000. Caracas 1080-A, Venezuela. Resumen Evaluamos las variaciones de las comunidades del fitoplancton y bacterioplancton en un río de aguas claras en la cuenca media del río Orinoco con la finalidad de verificar el acoplamiento entre am- bas comunidades. Los promedios de abundancia de las bacterias fluctuaron entre 0,72 y 4,07 × 10 5 org/mL y sus variaciones espacio- temporales fueron significativas. El fitoplancton osciló entre 1,1 y 22,6 × 10 3 org/L presentando los mayores valores en la cuenca baja del río y durante el período de aguas bajas. Las interrelacio- nes entre la abundancia bacteriana y las variables fisicoquímicas e hidrológicas presentaron correlaciones significativas positivas y negativas con la temperatura y el carbono orgánico disuelto res- pectivamente. El fitoplancton estuvo representado mayormente por las clorofíceas (4,8 × 10 3 org/L), luego las bacillariofíceas (2,1 × 10 3 org/L) y por último las cianobacterias (1,2 × 10 3 org/L). Se regis- traron 49 taxones de organismos fitoplanctónicos (aguas altas), siendo las bacillariofíceas el grupo que presentó la mayor rique- za promedio durante todo el período de estudio. Se presentó una relación negativa no significativa entre el fitoplancton y el bacte- rioplancton y una correlación positiva significativa entre el bacte-
Transcript of Relaciones entre el bacterioplancton y fitoplancton en el Amazonas Venezolano.

Boletín del Centro de Investigaciones Biológicas ISSN 0375-538X ~ Vol. 47. Nº 3, Septiembre-Diciembre 2013, Pp. 235 - 239
Recibido / Received: 18-07-2013 ~ Aceptado / Accepted: 29-10-2013
Relaciones entre el fitoplancton y el bacterioplancton en el río Cataniapo,
estado Amazonas, VenezuelaSimón Astiz1,2, Haymara Álvarez1 y Mariana Villaró1,3
1Instituto de Recursos Naturales, Universidad Simón Bolívar, Apartado 89000. Caracas 1080-A, Venezuela. [email protected] en Ciencia y Tecnologías Ambientales, Universidad Autónoma de Barcelona. Bellaterra, Barcelona 08193, España.3Maestría en Ciencias Biológicas, Universidad Simón Bolívar, Apartado 89000. Caracas 1080-A, Venezuela.
Resumen
Evaluamos las variaciones de las comunidades del fitoplancton y bacterioplancton en un río de aguas claras en la cuenca media del río Orinoco con la finalidad de verificar el acoplamiento entre am-bas comunidades. Los promedios de abundancia de las bacterias fluctuaron entre 0,72 y 4,07 × 105 org/mL y sus variaciones espacio-temporales fueron significativas. El fitoplancton osciló entre 1,1 y 22,6 × 103 org/L presentando los mayores valores en la cuenca baja del río y durante el período de aguas bajas. Las interrelacio-nes entre la abundancia bacteriana y las variables fisicoquímicas e hidrológicas presentaron correlaciones significativas positivas y negativas con la temperatura y el carbono orgánico disuelto res-pectivamente. El fitoplancton estuvo representado mayormente por las clorofíceas (4,8 × 103 org/L), luego las bacillariofíceas (2,1 × 103 org/L) y por último las cianobacterias (1,2 × 103 org/L). Se regis-traron 49 taxones de organismos fitoplanctónicos (aguas altas), siendo las bacillariofíceas el grupo que presentó la mayor rique-za promedio durante todo el período de estudio. Se presentó una relación negativa no significativa entre el fitoplancton y el bacte-rioplancton y una correlación positiva significativa entre el bacte-

Relaciones entre el fitoplancton y el bacterioplancton en el río Cataniapo Astiz et al.
236
rioplancton y el grupo de las bacillariofíceas, por lo que se sugiere un acoplamiento regulado por la composición de la comunidad fitoplanctónica.
Palabras clave: fitoplancton, bacterioplancton, río Cataniapo, es-tado Amazonas, Venezuela.
Relations between Phytoplankton and Bacterioplankton in the Cataniapo River,
State of Amazonas, Venezuela
Abstract
The variations of phytoplankton and bacterioplankton communi-ties in a clearwater river in the middle basin of the Orinoco River were evaluated in order to verify the coupling between the two communities. The average abundance of bacteria fluctuated be-tween 0.72 and 4.07 × 105 org/ml and spatiotemporal variations were significant. Phytoplankton ranged from 1.1 to 22.6 × 103 org/L presenting highest values in the lower river basin during the low-water period. The interrelationships between bacterial abundance and physicochemical and hydrological variables showed significant positive and negative correlations with temperature and organic dissolved carbon, respectively. Phytoplankton was represented principally by Chlorophyceae (4.8 × 103 org/L), then the bacillari-oficeas (2.1 × 103 org/L) and finally, cyanobacteria (1.2 × 103 org/L). 49 taxa of phytoplankton (high water) organisms were recorded; bacillarioficeas were the group with the highest average level dur-ing the entire period studied. There was a non-significant, nega-tive relationship between the phytoplankton and bacterioplank-ton communities and a significant positive correlation between bacterioplankton and the bacillarioficeas group, which suggests a coupling regulated by the composition of the phytoplankton com-munity.
Keywords: phytoplankton, bacterioplankton, Cataniapo River, Amazon State, Venezuela.

Boletín del Centro de Investigaciones Biológicas Vol. 47, Nº 3, septiembre-diciembre 2013, Pp. 235 - 239
237
Introducción
Existen antecedentes que indican que en diversos ecosistemas acuáticos las bacterias y el fitoplancton compiten por los nutrientes inorgánicos, al mismo tiempo, la liberación de materia orgánica disuel-tapor parte del fitoplancton estimula parcialmente el crecimiento del bacterioplancton; este al mismo tiempo libera nutrientes utilizables por la microflora, lo que favorece su desarrollo (Cole et al. 1993, Co-veney y Wetzel 1995, del Giorgio et al. 1997, Havens et al. 2000, Cano-sa y Pinilla 2007, Pinilla et al. 2007). Esta relación depende de factores abióticos que afectan a las dos comunidades como la temperatura, el oxígeno disuelto, la disponibilidad de nutrientes, las variaciones del caudal e influye sobre su productividad, abundancia y composición. Se ha demostrado que la abundancia y la producción del bacterioplancton están relacionadas positivamente con la producción primaria del fito-plancton y que el bacterioplancton utiliza gran parte de las proteínas y aminoácidos provenientes del fitoplancton (Rosenstock y Simon 2001, Van den Meersche et al. 2004, Beckwith 2009, Løvdal et al. 2008). Sin embargo, otros estudios han encontrado que la abundancia y el creci-miento bacterial no están acoplados a la abundancia y producción fito-planctónica (Findlay et al. 1991, Gurung y Urabe 2010).
En las últimas décadas se han realizado diversos estudios en la Orinoquia Venezolana relacionados con la taxonomía y abundancia de las comunidades del fitoplancton y el zooplancton que habitan en algunos ecosistemas acuáticos de esta región (Vásquez 1984a,b, Vás-quez y Sánchez 1984, Zoopi de Roa et al. 1985, Medina y Vásquez 1988, Saunders y Lewis 1988a,b, Martin 1989, Vásquez y Rey 1989,1992, Ha-milton et al. 1990, Weibezahn et al. 1990, Lewis et al. 1990, González 1992, Astiz y Alvarez 1998, Vásquez et al. 1998, Papadakis 2003, Reverol et al. 2008). Sin embargo, se conocen pocos estudios sobre algunos aspectos ecológicos de la comunidad del bacterioplancton en dichos ecosistemas (Lewis et al. 1990, Pérez 1995, Villaró 1997, Montoya 1999, Castillo 2000, Castillo et al. 2003, Bastardo 2006, Bastardo et al. 2007) y muy pocos o ninguno sobre la dinámica conjunta de estas comunida-des. En este estudio se dan a conocer los primeros resultados del estu-dio comparativo y el posible acople entre dos componentes planctóni-cos fundamentales (bacterioplancton y fitoplancton) del río Cataniapo

Relaciones entre el fitoplancton y el bacterioplancton en el río Cataniapo Astiz et al.
238
en la cuenca media del río Orinoco y su relación con las variaciones am-bientales de este ecosistema.
Área de estudio
La cuenca del río Cataniapo está localizada en el municipio Atu-res, al noroeste del estado Amazonas y al sureste de su capital Puerto Ayacucho (5°25’-5°43’ Norte, 67°05’-67°35’ Oeste), posee la figura admi-nistrativa de área protegida (Plan de ordenamiento y reglamento de uso de la zona protectora de la cuenca hidrográfica del Río Catania-po 2005). Forma parte de la subcuenca del Orinoco medio y tiene un área de 1585 km2. La longitud máxima del cauce principal del río Ca-taniapo es de 108 km, con una descarga promedio en el ámbito de su desembocadura de 82,7 m3/s (Núñez 1993). El clima de esta importante cuenca se puede considerar de tipo tropical monzónico con una corta estación seca comprendida entre los meses de enero y marzo, con una temperatura y precipitación promedio anual de 26,7°C y 2847 mm res-pectivamente. La población de esta área es de 3112 habitantes, siendo el 90% indígena (Graterol y Botto 2003). El río Cataniapo es un río de sexto orden, que recibe las aguas de 116 afluentes, siendo el río Gavilán el mayor de ellos y responsable del 25% del área total de drenaje. Las aguas del río Cataniapo son consideradas como “aguas claras” según la clasificación de Sioli (1984), de poca turbidez y de baja conductividad, ligeramente ácidas y de una coloración verdosa.
Materiales y métodos
Las estaciones para los muestreos de agua y plancton fueron fija-das en la parte central del cauce del río Cataniapo en tres sectores de fácil acceso: el muelle de la comunidad indígena de “Gavilán” corres-pondiente a la estación uno (5°32’ Norte, 67°21’ Oeste) representativa de las condiciones de la sección alta del río Cataniapo; la comunidad indígena de “Las Pavas” correspondiente a la estación dos (5°34’ Norte, 67°30’ Oeste) representativa del tramo “medio” del río y finalmente la estación de aducción del acueducto local o “Toma de la CVG” corres-pondiente a la estación tres (5°36’ Norte, 67°35’ Oeste) representati-va de la sección baja del río, la más antropizada (Infante 1992, Blanco 2005, Astiz 2012). Se realizaron siete muestreos de campo en el período

Boletín del Centro de Investigaciones Biológicas Vol. 47, Nº 3, septiembre-diciembre 2013, Pp. 235 - 239
239
de un año los cuales correspondieron a las hidrofases del río de descen-so de aguas (octubre-diciembre); aguas bajas (abril); ascenso de aguas (mayo-junio) y aguas altas (agosto). Se recolectaron las muestras para la determinación de nutrientes integrando la columna de agua en el punto de mayor velocidad de la corriente con una botella integradora de 3,5 L (Nordin et al. 1983), los recipientes fueron previamente lavados con una solución de HCl al 10% y agua desionizada (APHA 1985). El pH, temperatura, conductividad y oxígeno disuelto se determinó in situ con el uso de sensores de campo, la transparencia con disco de Secchi. Adi-cionalmente se calculó el caudal en cada estación de muestreo.
Las muestras de fitoplancton se recolectaron haciendo un barrido vertical de la columna de agua con la botella integradora. Se extrajeron dos alícuotas de un litro en cada estación y se fijaron en campo, una con forma-lina al 5% para la identificación de las microalgas por grandes grupos; otra con lugol concentrado para el análisis cuantitativo, el cual se realizó en un microscopio invertido según el método de Lund et al. (1958) en cámaras de sedimentación Utermöhl de 10 mL. Las muestras subsuperficiales de bacterioplancton se tomaron directamente del río en frascos estériles y se fijaron con formol al 2%, se mantuvieron refrigeradas y en oscuridad hasta el conteo total de bacterias con la técnica de la epifluorescencia (Kepner y Pratt 1994). Se utilizó como fluorocromo naranja de acridina al 0,2% y un tiempo de contacto de dos minutos; se utilizaron filtros Isopore negros de 0,2 µm de diámetro de poro y un microscopio de epifluorescencia Zeiss, en cada filtro se seleccionaron diez campos al azar y se promedió el conteo de todas las bacterias presentes (Wetzel y Likens 1992).
Se realizó un análisis de varianza (ANOVA) de dos vías para cono-cer posibles diferencias significativas de las variables ambientales (fi-sicoquímicas, hidrológicas y nutrientes) y biológicas (densidad) entre las estaciones y los períodos (hidrofases) estudiadas. Para establecer posibles correlaciones entre las variables ambientales y biológicas se utilizó el índice de correlación de Spearman, ya que algunas variables no pudieron ser corregidas por normalidad luego de su transformación logarítmica. Para los cálculos se utilizó el programa InfoStat versión 2008. Para conocer las relaciones estadísticas entre pares de comuni-dades se utilizaron regresiones simples de sus densidades previa estan-darización de los volúmenes. En todos los casos se consideró un nivel de significancia de P < 0,05.
Relaciones entre el fitoplancton y el bacterioplancton en el río Cataniapo Astiz et al.
240
Resultados
Características fisicoquímicas
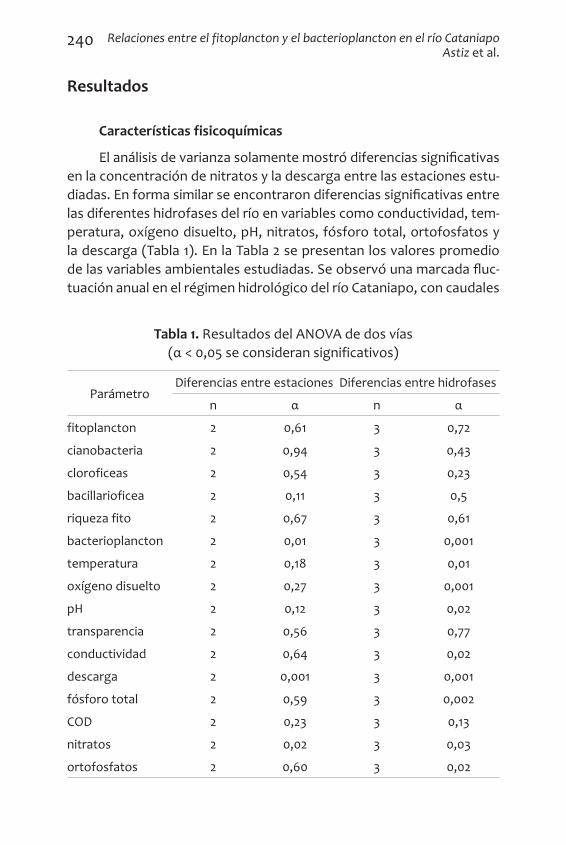
El análisis de varianza solamente mostró diferencias significativas en la concentración de nitratos y la descarga entre las estaciones estu-diadas. En forma similar se encontraron diferencias significativas entre las diferentes hidrofases del río en variables como conductividad, tem-peratura, oxígeno disuelto, pH, nitratos, fósforo total, ortofosfatos y la descarga (Tabla 1). En la Tabla 2 se presentan los valores promedio de las variables ambientales estudiadas. Se observó una marcada fluc-tuación anual en el régimen hidrológico del río Cataniapo, con caudales
Tabla 1. Resultados del ANOVA de dos vías (α < 0,05 se consideran significativos)
ParámetroDiferencias entre estaciones Diferencias entre hidrofases
n α n α
fitoplancton 2 0,61 3 0,72
cianobacteria 2 0,94 3 0,43
cloroficeas 2 0,54 3 0,23
bacillarioficea 2 0,11 3 0,5
riqueza fito 2 0,67 3 0,61
bacterioplancton 2 0,01 3 0,001
temperatura 2 0,18 3 0,01
oxígeno disuelto 2 0,27 3 0,001
pH 2 0,12 3 0,02
transparencia 2 0,56 3 0,77
conductividad 2 0,64 3 0,02
descarga 2 0,001 3 0,001
fósforo total 2 0,59 3 0,002
COD 2 0,23 3 0,13
nitratos 2 0,02 3 0,03
ortofosfatos 2 0,60 3 0,02

Boletín del Centro de Investigaciones Biológicas Vol. 47, Nº 3, septiembre-diciembre 2013, Pp. 235 - 258
241
Tabl
a 2.
Val
ores
de
conc
entr
acio
nes
prom
edio
de
las
varia
bles
fisi
coqu
ímic
as e
hid
roló
gica
s an
aliz
adas
en
las
dife
rent
es h
idro
fase
s y
para
cad
a es
taci
ón
Varia
bles
Hid
rofa
se
Agua
s ba
jas
(Abr
)As
cens
o ag
uas
(May
-Jun
)Ag
uas
Alta
s (A
go)
Des
cens
o ag
uas
(Oct
-Dic
)
Est.
1Es
t. 2
Est.
3Es
t. 1
Est.
2Es
t. 3
Est.
1Es
t. 2
Est.
3Es
t. 1
Est.2
Es
t.3
tem
pera
tura
(°C)
26,2
25
,0
26,0
24
,7
24,3
24
,9
25,4
25
,7
26,9
23
,8
23,2
24
,0
tran
spar
. (m
)1,1
51,4
21,2
0_
1,58
2,60
1,00
1,85
1,55
1,30
1,42
1,30
cond
uct.
(µS/
cm)
-4,
06
4,65
2,
85
2,87
3,
09
3,59
3,
13
3,12
3,
82
4,60
3,
48
pH6,
20
3,60
3,
50
5,25
5,
43
5,50
5,
70
5,90
5,
70
6,00
6,
00
5,50
oxí.
dis.
(mg/
L)9,
60
9,60
9,
80
--
-8,
00
7,60
7,
80
7,40
7,
60
7,40
fósf
oro
tota
l (µg
/L)
8,50
10,5
08,
0018
,50
29,5
039
,00
50,0
052
,50
31,0
010
,00
11,0
021
,00
orto
fosf
atos
(µg/
L) 2
,00
2,00
0,00
3,00
0,00
2,00
10,0
030
,00
0,30
0,00
0,00
0,00
nitr
atos
(µg/
L)14
2,00
156,
9513
5,50
_16
8,95
133,
6014
3,75
151,5
012
6,50
126,
314
0,44
121,1
2
COD
(mg/
L)1,5
01,2
01,0
01,7
01,3
01,9
04,
372,
001,9
02,
001,8
01,8
0
desc
arga
(m³/s
)10
,97
20,0
044
,00
20,0
042
,64
90,0
038
,81
83,6
316
0,90
27,5
630
,00
60,0
0

Relaciones entre el fitoplancton y el bacterioplancton en el río Cataniapo Astiz et al.
242
máximos y mínimos de 160,9 m3/s y 10,97 respectivamente. La tempe-ratura del agua osciló entre los 23,2 y los 26,9°C, los mayores valores se registraron durante la hidrofase de aguas bajas (diciembre). La trans-parencia presentó valores bastante altos y cónsonos con lo reportado para estos ríos de aguas claras, con un promedio de 1,49 m. En relación a la concentración de nutrientes los valores mínimos observados fueron los siguientes: NO3
−: 126,3 µg/L; PO4 ³⁻: 0,0 µg/L; fósforo total: 8,0 µg/L; carbono orgánico disuelto (COD): 1,0 mg/L, los cuales se obtuvieron en la hidrofase de aguas bajas. Se detectaron valores máximos para los NO3
−: 168,95 µg/L; PO4³⁻: 30,0 µg/L; fósforo total: 52,5 µg/L; COD: 4,37 mg/L, la mayoría obtenidos en aguas altas (Tabla 2). El valor promedio del pH osciló alrededor de cinco para las tres secciones del río. Sin em-bargo, en la época de sequía o hidrofase de aguas bajas (diciembre) se observó una disminución del pH hasta las 3,5 unidades en la estación tres (cuenca baja). Los valores de oxígeno disuelto fueron ligeramente superiores en esta misma hidrofase de aguas bajas, alrededor de 9,8 mg/L en comparación con la estación o hidrofase de descenso de aguas (octubre) en donde los valores oscilaron alrededor de los 7,4 mg/L. La conductividad (3,6 µS/cm) y el oxígeno disuelto (8,31 mg/L) presenta-ron valores promedio a lo largo del período de estudio aceptables para el normal desarrollo de la vida acuática y mantuvieron la tendencia ob-servada y reportada en los últimos años (Núñez 1993, Astiz 2012). Los bajos niveles de conductividad observados en el río Cataniapo son el resultado del tipo de suelos que drena la cuenca del río, los cuales son muy antiguos, lixiviados y pobres en carbonatos y sales disueltas.
Las interrelaciones entre las variables fisicoquímicas e hidrológi-cas presentaron seis correlaciones significativas, de las cuales cinco fueron positivas (fósforo total con pH, descarga, COD y los ortofosfa-tos; pH con ortofosfatos) y una negativa (pH con COD) (Tabla 3).
Abundancia bacteriana y fitoplanctónica
Los promedios de abundancia de las bacterias en el río Cataniapo fluctuaron entre 0,72 y 4,07 × 105 org/mL (Fig. 1) y presentaron diferen-cias espaciales y temporales significativas (Tabla 1), con los menores valores en la estación uno representativa de la cuenca alta del río y du-rante el descenso de aguas. Los mayores valores de abundancia bacte-riana se observaron en la estación tres representativa de la cuenca baja

Boletín del Centro de Investigaciones Biológicas Vol. 47, Nº 3, septiembre-diciembre 2013, Pp. 235 - 258
243
y durante la hidrofase de aguas bajas. La forma predominante de la co-munidad bacteriana estuvo representada por los cocos (71%), luego por los bacilos (21%) y en menor proporción los bastones (8%). Las interre-laciones entre la abundancia bacteriana y las variables fisicoquímicas e hidrológicas presentaron solamente dos correlaciones significativas: una positiva con la temperatura y una negativa con el COD (Tabla 3). El mayor número de taxones de organismos fitoplanctónicos registra-dos fue de 49 (aguas altas), siendo las bacillariofíceas el grupo que pre-sentó la mayor riqueza promedio durante todo el período de estudio. La abundancia total del fitoplancton osciló entre 1,1 y 22,6 × 103 org/L, presentando los mayores valores en esta última estación y durante la hidrofase de aguas bajas, sin embargo las fluctuaciones espacio tem-porales en la abundancia no fueron significativas (Tabla 1). Entre los grupos analizados se observó en promedio el predominio de las clo-rofíceas (4,8 × 103 org/L), con mayor presencia de los géneros Stauras-trum y Scenedesmus; luego las bacillariofíceas (2,1 × 103 org/L), con pre-dominio de Pinnularia y Navicula; después las cianobacterias (1,2 × 103
org/L), con mayor participación de Anabaena y Oscillatoria y finalmente los grupos de presencia temporal englobados como otros (euglenofí-ceas, dinofíceas, criptofíceas, pirrofíceas) (0,9 × 103 org/L) (Fig. 1). Las clorofíceas y las bacillariofíceas presentaron mayores abundancias en la hidrofase de aguas bajas (abril) y en las estaciones dos y tres del río, mientras que el grupo de los otros presentaron aumentos importantes en su abundancia durante las aguas altas (agosto) en las mismas esta-ciones. La abundancia de la comunidad fitoplanctónica presentó una alternancia en cuanto al dominio del grupo de las bacillariofíceas con el de las clorofíceas a lo largo del período de estudio (Fig. 1).
Tabla 3. Correlaciones de rangos de Spearman significativas (P < 0,05) entre las variables analizadas en el río Cataniapo
Variables Variables
fósforo total ortofosfato (+), pH (+), descarga (+), COD (+)
pH COD (–), ortofosfato (+)
bacterioplancton temperatura (+), bacillarioficeas (+), COD (-)
fitoplancton cianobacterias (+)
Los signos dentro de los paréntesis indican correlaciones positivas (+) o negativas (–).

Relaciones entre el fitoplancton y el bacterioplancton en el río Cataniapo Astiz et al.
244
Figura 1. Variación espacio-temporal en la abundancia de las comunidades planctónicas en relación a la descarga en el río
Cataniapo durante el período de estudio.

Boletín del Centro de Investigaciones Biológicas Vol. 47, Nº 3, septiembre-diciembre 2013, Pp. 235 - 258
245
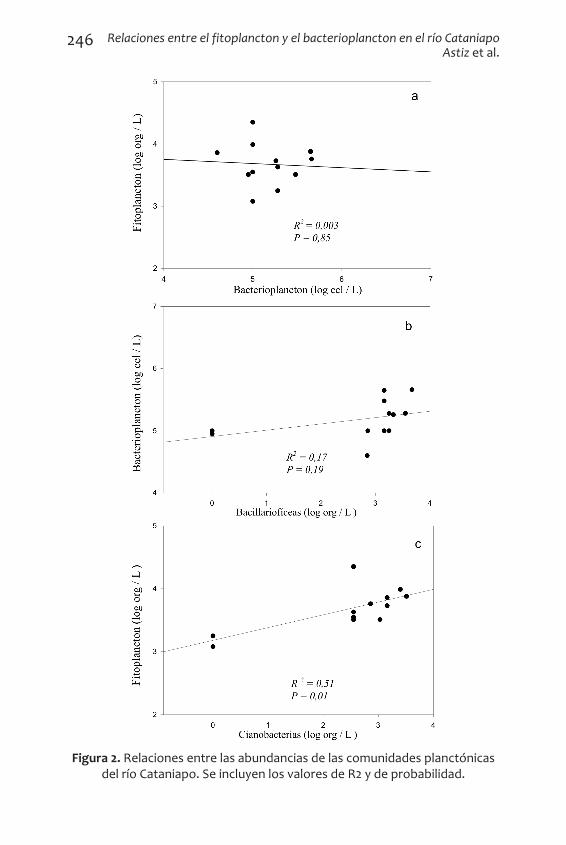
Relaciones entre las comunidades
Las abundancias de las comunidades fitoplanctónicas y del bacte-rioplancton presentaron una muy leve relación negativa no significati-va entre (R2 = 0,003) (Fig. 2a). Por otro lado, se observó una correlación positiva significativa entre el bacterioplancton y el grupo de las baci-llariofíceas (Tabla 3 y Fig. 2b), por lo que el bacterioplancton prospera cuando lo hace también las bacillariofíceas. La regresión lineal entre el fitoplancton y el grupo de las cianobacterias reflejó una relación positiva significativa entre ambas comunidades (R2 = 0,01) (Tabla 3 y Fig. 2c). Es importante recordar que las regresiones lineales sólo ex-plican parcialmente las relaciones entre los organismos planctónicos, ya que existen muchos fenómenos ecológicos que las abundancias no consideran; no obstante, constituyen un reflejo de las tendencias de cómo los cambios en una comunidad pueden estar afectados por las variaciones en las otras.
Discusión
Las variables fisicoquímicas e hidrológicas mostraron que cada hi-drofase del río Cataniapo tiene características limnológicas distintas, lo que aparentemente influyó en la presencia y abundancia del bacte-rioplancton pero no en la densidad del fitoplancton. El análisis de las variaciones espaciales y temporales de la gran mayoría de las variables ambientales estudiadas en la primera parte de este estudio se reportan en el trabajo de Astiz (2012). En general estas variaciones responden a las fluctuaciones del régimen hidrológico del río, el cual a su vez está afectado por la estacionalidad de las precipitaciones en la Orinoquia venezolana determinada por el movimiento latitudinal de la zona de convergencia intertropical, originándose una época seca de escasas precipitaciones que se extiende regularmente desde el mes de Ene-ro hasta Marzo y una intensa época lluviosa el resto del año (Vásquez 1989, Astiz y Alvarez 1998, Lewis et al. 2000, Montoya et al. 2011).
Las concentraciones de nitratos, fósforo total y ortofosfatos ob-servados presentaron diferencias significativas entre las estaciones estudiadas y entre las diferentes hidrofases, con poca variación en comparación con estudios anteriores realizados en esta cuenca y en donde se señala un aumento de estos nutrientes durante la hidrofase
Relaciones entre el fitoplancton y el bacterioplancton en el río Cataniapo Astiz et al.
246
Figura 2. Relaciones entre las abundancias de las comunidades planctónicas del río Cataniapo. Se incluyen los valores de R2 y de probabilidad.

Boletín del Centro de Investigaciones Biológicas Vol. 47, Nº 3, septiembre-diciembre 2013, Pp. 235 - 258
247
de ascenso de aguas (inicio de la época lluviosa), debido a que estos elementos son fácilmente lavados de los suelos por la escorrentía su-perficial. Los valores moderados de nitratos observados en el río Ca-taniapo podrían ser el resultado de que la cuenca es ineficiente en la retención del nitrógeno inorgánico o que el sistema libera el exceso en función de las entradas por precipitación o por un balance positivo para el primer término en la tasa de nitrificación-desnitrificación, prin-cipalmente bajo la acción de la vegetación. Las bajas concentraciones obtenidas para el fósforo disuelto son el resultado de una menor dispo-nibilidad del elemento en la cuenca y por una mayor eficiencia biológica de retención en los sistemas dominados por bosques húmedos tropi-cales en equilibrio (Weibezahn 1990, Núñez 1985,1993, Lucchetti 1996, Lewis et al. 2000). El río Cataniapo, de aguas claras, presentó concen-traciones de carbono orgánico disuelto (COD) que oscilaron entre 1,00 y 4,37 mg/L. Weibezahn (1990) registró concentraciones de COD para los ríos Ventuari y Vichada, ambos de aguas claras y ubicados también en la cuenca media del río Orinoco, de 2,9 y 2,3 mg/L respectivamente y muy similares a las obtenidas en este estudio. Otros reportes de la cuenca baja del río Orinoco han determinado concentraciones que os-cilaron entre 3,04 y 6,2 mg/L (Paolini 1990, Lewis y Saunders 1990). En relación a las variaciones estacionales en las concentraciones de COD, si bien no fueron significativas, los valores máximos observados en el mes de agosto son consecuencia del período de lluvia, ya que la mate-ria orgánica particulada y disuelta originada en la cuenca es lavada por efecto de las precipitaciones y depositada en el cauce del río, en don-de parte de ella es procesada y parte exportada fuera de la cuenca. El origen autóctono del COD es prácticamente despreciable debido a las bajas densidades de organismos fitoplanctónicos del río Cataniapo y de la marcada influencia de la descarga sobre la producción bacteriana, estas evidencias fortalecen la hipótesis de que la principal fuente de materia orgánica del río es de origen alóctono (Villaró 1997, Castillo et al. 2004).
Los promedios de abundancia de las bacterias en el río Cataniapo fluctuaron entre 0,4 y 4,6 × 105 org/mL. Estas abundancias resultaron ser similares a las reportadas por Castillo et al. (2004) para este mismo río (6,5 × 105 org/mL) y por Schmidt (1970) para los ríos Negro y Solimôes, las cuales fueron de 2,5 y 5,0 × 105 org/mL respectivamente. En general

Relaciones entre el fitoplancton y el bacterioplancton en el río Cataniapo Astiz et al.
248
las abundancias del bacterioplancton obtenidas en este estudio fueron ligeramente inferiores a las reportadas en otros ecosistemas acuáticos de las cuencas de los ríos Orinoco y Amazonas. Existe suficiente evi-dencia de que las abundancias del bacterioplancton en la mayoría de los ecosistemas acuáticos epicontinentales fluctúan entre uno o dos órdenes de magnitud, lo que podría indicar que no existen variaciones extremas en la abundancia del bacterioplancton, independientemente de las características ecológicas de estos ambientes (Lewis et al. 1990, Benner et al. 1995, Pérez 1995, Villaró 1997, Montoya 1999, Carvalho et al. 2003, Castillo et al. 2004, Pinilla et al. 2007, Canosa y Pinilla 2007, Teixeira et al. 2011, Portillo et al. 2012).
Las mayores densidades bacterianas observadas en la estación tres representativa de las condiciones de la cuenca baja del río Cata-niapo estarían reflejando la respuesta positiva de esta comunidad a las mayores entradas de material orgánico e inorgánico alóctono origina-do en esta zona de la cuenca con mayor grado de antropización, el cual ha sido producto de la mayor presencia de asentamientos indígenas, campesinos y turísticos que estarían modificando la cobertura vegetal, las condiciones del suelo y generando residuos sólidos y líquidos que son arrastrados al curso fluvial, estimulando el desarrollo de la comu-nidad del bacteriaplancton entre otras (Villaró 1997, Astiz 2012). El pa-trón de variación temporal de la abundancia del bacterioplancton en el río Cataniapo presentó un comportamiento similar al encontrado por Lewis et al. (1990), Leff et al. (1998), Carvalho et al. (2003) y Palijan et al. (2007) donde los máximos en las abundancias se observaron en la hidrofase de ascenso de aguas y, según estos autores, estuvo relacio-nado con las concentraciones más altas de COD para ese período a dife-rencia de lo encontrado en este estudio, en donde el bacterioplancton y el pH estuvieron correlacionados negativamente con el COD. Mon-toya (1999) y Castillo et al. (2004) reportaron en forma similar la falta de correlación entre la producción y abundancia bacteriana y la con-centración de COD en los ríos Caura y Cataniapo (ligeramente ácidos) respectivamente. Esta falta de vínculos entre las variables menciona-das se podrían originar por el aumento de la acidez del agua, ya que al disminuir los valores de pH en el agua los compuestos húmicos pre-sentes se vuelven más hidrofóbicos y por consiguiente menos lábiles a la degradación bacteriana, lo que explicaría la ausencia de respuesta

Boletín del Centro de Investigaciones Biológicas Vol. 47, Nº 3, septiembre-diciembre 2013, Pp. 235 - 258
249
en la abundancia del bacterioplancton al aumentar las concentraciones de COD. Actualmente existen evidencias que factores como una baja disponibilidad de nutrientes, pH o condiciones físicas extremas pueden controlar la proporción de bacterias activas en un sistema acuático (Pu-lido et al. 2003, Harsha et al. 2007).
A diferencia del bacterioplancton, la abundancia del fitoplancton fue mayor en la hidrofase de aguas bajas, con un marcado predominio de las algas clorofíceas. Los valores del río Cataniapo oscilaron entre 1,1 y 22,6 × 103 org/L, muy por debajo a los de otros ríos de la región. Para el río Orinoco se han registrado abundancias entre 5,3 × 104 y 1,1 × 106
org/L (Vásquez y Sánchez 1984, Alvarez et al. 1992), para el río Ventuari (afluente del alto Orinoco) 1,7 × 104 org/L (Carvajal, 1994), para el río Caura (bajo Orinoco) 1,02 × 105 org/L (Ibáñez y Lara 1997). Para otros ríos en Suramérica como el Paranapanema en Brasil se han registrado abundancias promedio de 2,0 ×105 org/L (Nogueira et al. 2010) y en el río Paraná en Argentina se han encontrado hasta 2,5 × 103 org/L (Zalocar et al. 2007). Los “picos” máximos de abundancia en la comunidad fito-planctónica durante las aguas bajas frecuentemente superan por un factor de diez a los valores mínimos observados durante las aguas altas (Sánchez y Vásquez 1986, Lewis et al. 1990). Estos máximos durante las aguas bajas son interesantes porque indicarían el origen autóctono de la comunidad fitoplanctónica, ya que no habría ningún aporte adi-cional de los organismos planctónicos que proliferan en las lagunas de inundación que se encuentran aisladas del río. Debido a que los ríos no están en contacto con estas lagunas durante la hidrofase de aguas ba-jas, el fitoplancton prospera en pequeños hábitats de escasa corriente y mayor transparencia formados dentro del curso fluvial, ya sea en las cabeceras, remansos o en las riberas con depresiones protegidas de la corriente que eventualmente serían incorporados al canal principal del río (Lewis et al. 1990, Wetzel 2001).
Por otro lado es importante destacar el aumento significativo en la abundancia del grupo fitoplanctónico denominado en este estudio como otros (euglenofíceas, dinofíceas, criptofíceas) durante la hidro-fase de las aguas altas en las estaciones estudiadas del río Cataniapo, período en el cual se presentan los mayores valores de conductividad y de concentración de sólidos suspendidos totales (Astiz 2012), este florecimiento temporal sería favorecido por la mayor concentración

Relaciones entre el fitoplancton y el bacterioplancton en el río Cataniapo Astiz et al.
250
de nutrientes y de bacterioplancton en este período de máxima des-carga del río. Existen reportes que estos grupos prosperan en aguas con abundante materia orgánica y carga bacteriana en detrimento de otros grupos como las clorofíceas y bacillariofíceas (Mirande y Tracan-na 2005, Nabout et al. 2006).
Las bacillariofíceas fue el grupo de mayor riqueza promedio en el río Cataniapo. Resultados similares se han reportado para el río Orino-co (Lewis y Weibezahn 1976, Weibezahn et al. 1990, Álvarez et al. 1992) y podría ser explicado por la mayor adaptabilidad que presenta este grupo fitoplanctónico a las condiciones de inestabilidad que presenta el ecosistema lótico (Wetzel 2001, Pérez 2002). Las cianobacterias a pesar de ser poco abundantes y con menor cantidad de formas fue el grupo que presentó la mayor relación proporcional significativa con la abundancia del fitoplancton total, por lo que podría pensarse en una sincronización entre las variaciones de ambas abundancias a lo largo del período de estudio. En general, las abundancias fitoplanctónicas observadas en el río Cataniapo corresponden a ambientes oligotrófi-cos, en especial durante la hidrofase de aguas altas o de máxima inun-dación, característica común en la mayoría de ecosistemas acuáticos de la selva lluviosa tropical (Núñez 1985, Allan y Castillo 2007).
La regresión entre las abundancias del fitoplancton y el bacterio-plancton total no fue significativa, sin embargo indicó una tendencia de que el fitoplancton disminuye cuando el bacterioplancton aumenta, lo que podría mostrar algún tipo de competencia entre las dos comu-nidades, posiblemente por fósforo (Del Giorgio et al. 1997, Bastardo 2006, Pinilla et al. 2007). Por otro lado, se obtuvo una relación directa-mente proporcional aunque no significativa entre el bacterioplancton y el grupo de las bacillariofitas, indicando algún tipo de simbiosis entre ambas comunidades, siendo este grupo fitoplanctónico más oportunis-ta y adaptable a condiciones ambientales muy variables (Wetzel 2001, Pérez 2002) podría verse favorecido por los productos de excreción (nutrientes inorgánicos) del bacterioplancton.
El acoplamiento entre el bacterioplancton y el fitoplancton no siempre se presenta en los ecosistemas acuáticos, y cuando sucede al-gún grado de dependencia es muy variable y específico (Wetzel 2001, Costa y Miyashiro 2005, Petrucio et al. 2006, Canosa y Pinilla 2007, Gao et al. 2007, Pinilla et al. 2007, Faithfull et al. 2011). Al parecer el aco-

Boletín del Centro de Investigaciones Biológicas Vol. 47, Nº 3, septiembre-diciembre 2013, Pp. 235 - 258
251
plamiento entre fitoplancton y bacterioplancton depende no solo de la disponibilidad de COD sino de otros factores como limitaciones de nitrógeno o fósforo, intensidad lumínica, predación y herbivoría y para-sitismo por virus (Wetzel 2001, Farnell y Ward 2003, Zöllner et al. 2003). Existe evidencia de que el acoplamiento entre bacterioplancton y fito-plancton sería más a nivel metabólico (producción) que entre las abun-dancias o las biomasas y que esta interacción sería más intensa en lagos oligotróficos, donde la fuente principal de carbono orgánico disuelto (COD) es el fitoplancton. Mariazzi et al. (1998) estudiando un embalse del río Negro en Argentina demostraron que estas dos producciones estuvieron significativamente correlacionadas, aunque la producción bacteriana no mostró correlación con la abundancia de bacterias. En este estudio realizado en un río oligotrófico esta interacción no sería tan marcada debido al origen alóctono de casi la mayoría del COD pre-sente en el río Cataniapo (Villaró 1997, Castillo et al. 2004). De forma similar a lo encontrado en este estudio Biddanda et al. (2001), observa-ron una mayor participación del bacterioplancton en la biomasa planc-tónica total de aguas oligotróficas en comparación con las eutróficas, lo que estaría indicando una carencia de acoplamiento entre este y el fitoplancton. Adicionalmente, la correlación positiva observada entre el bacterioplancton y la temperatura indicaría la importancia de esta variable para el bacterioplancton, lo que hace pensar que bacterias y fitoplancton responden independientemente a los cambios en la tem-peratura del agua (Abarzúa et al. 1995, Wetzel 2001, Niu et al. 2011).
Este estudio presentó datos preliminares e inéditos sobre la abun-dancia, riqueza y variación de la comunidad fitoplanctónica y su rela-ción con el bacterioplancton en un río de aguas claras, el río Cataniapo, en la cuenca media del río Orinoco. Sería fundamental establecer los mecanismos de control de ambas comunidades. Los resultados indica-rían que no sólo los factores abióticos como la descarga, temperatura, pH o la disponibilidad de nutrientes afectarían su desarrollo sino fac-tores bióticos como la herbivoría, depredación o parasitismo podrían explicar la falta de acoplamiento entre ambas comunidades. Nuevos estudios que involucren muestreos más intensivos y un mayor grupo de microorganismos participantes en las cadenas tróficas acuáticas ayudarían a entender mejor las interacciones entre el bacterioplancton y el fitoplancton en el ecosistema estudiado.

Relaciones entre el fitoplancton y el bacterioplancton en el río Cataniapo Astiz et al.
252
Agradecimientos
Este estudio fue realizado gracias al financiamiento del Fondo Na-cional de Ciencia, Tecnología e Innovación (FONACIT) antiguo CONICIT, a través del Proyecto “Balance Biogeoquímico de las Cuencas de los Ríos Autana y Cataniapo. Edo. Amazonas. Venezuela”, código MPS-RP-VIII 250060. Esta investigación es otra contribución del Proyecto Ecosistema Orinoco (PECOR) del Instituto de Recursos Naturales de la Universidad Simón Bolívar. A Maciel Melguero y a Humberto Carvajal-Chitty por su apoyo en el análisis de las muestras fitoplanctónicas.
Literatura citada
Abarzúa, M., S. Basualto y H. Urrutia. 1995. Relación entre la abundancia y biomasa de fitoplancton y bacterioplancton heterotrófico en aguas su-perficiales del golfo de Arauco, Chile. Invest. Mar. Valparaiso 23: 67-74.
Allan, D. and M. Castillo. 2007. Stream Ecology. Springer, The Netherlands. 436 pp.
Alvarez, H., H. Carvajal y A. Martínez. 1992. Monitoreo de parámetros fisico-químicos y del plancton en el alto y medio Orinoco. Instituto de Recursos Naturales. Universidad Simón Bolívar. Caracas. 98 pp.
APHA. 1985. Standard methods for examination of water and wastewater. American Public Health Association, Nueva York. 2275 pp.
Astiz, S. y H. Álvarez. 1998. El zooplancton en el alto y medio río Orinoco, Venezuela. Acta Científica Venezolana 49: 5-18.
Astiz, S. 2012. Deterioro del recurso agua en el río Cataniapo, Amazonas, Vene-zuela. Tecnología y Ciencias del Agua III (3): 5-20.
Bastardo, A. 2006. Dinámica y funcionalidad espacio-temporal de las bacte-rias heterótrofas del agua en el corredor ribereño del bajo río Orinoco. Tesis de Maestría en Ciencias Ambientales. Universidad Experimental de Guayana. Ciudad Guayana. 118 pp.
Bastardo, A., H. Bastardo y J. Rosales. 2007. Diversidad funcional de las bac-terias heterótrofas del bajo río Orinoco, Venezuela. Ecotropicos 20(1): 15-23.
Beckwith, M. 2009. Coupling of autotrophic and heterotrophic plankton food web components in the tidal-freshwater James river, USA. M.Sc. thesis, Virginia Commonwealth University. 46 pp.
Benner, R., S. Opsahl, G. Chin-Leo, J. Richey y B. Forsberg. 1995. Bacterial carbon metabolism in the Amazon river system. Limnol. Oceanogr. 40: 1262-1270.

Boletín del Centro de Investigaciones Biológicas Vol. 47, Nº 3, septiembre-diciembre 2013, Pp. 235 - 258
253
Biddanda, B., M. Ogdahl y J. Cotner. 2001. Dominance of bacterial metabo-lism in oligotrophic relative to eutrophic waters. Limnol. Oceanogr. 46: 730-739.
Blanco, H. 2005. The tension between developmental land uses and the con-servation area designation: a case study of the creoles in Cataniapo river basin, Venezuela. M.Sc. thesis. Athens. University of Ohio. 100 pp.
Canosa, A., y G. Pinilla. 2007. Relaciones entre las abundancias del bacterio-plancton y del fitoplancton en tres ecosistemas lenticos de los Andes Colombianos. Rev. Biol. Trop. 55(1): 135-146.
Carvajal, H. 1994. Phytoplankton in the upper and middle Orinoco river. Verh. Internat. Verein. Limnol. 25: 1856-1861.
Carvalho, P., S. Thomaz y L. Bini. 2003. Effects of water level, abiotic and biotic factors on bacterioplankton abundance in lagoons of a tropical floodplain (Paraná river, Brazil). Hydrobiologia 510: 67-74.
Castillo, M. 2000. Influence of hydrological seasonality on bacterioplankton in two neotropical floodplain lakes. Hydrobiologia 437: 57-65.
Castillo, M., G. Kling y J.-D. Allan. 2003. Bottom-up controls on bacterial pro-duction in tropical lowland rivers. Limnology and Oceanography 48(4): 1466-1475.
Castillo, M., J.D. Allan, R. Sinsabaugh y G. Kling. 2004. Seasonal and inter-annual variation of bacterial production in lowland rivers of the Orinoco basin. Freshwat. Biol. 49: 1400-1414.
Cole, J., M. Pace, N. Caraco y G. Steinhart. 1993. Bacterial biomass and cell size distributions in lakes: More and larger cells in anoxic waters. Limnol. Oceanogr. 38: 1627-1632.
Costa, C. y A. Miyashiro. 2005. Relationships between rotifers, phytoplank-ton and bacterioplankton in the Corumbá reservoir, Goiás state, Brazil. Hydrobiologia 546: 415-421.
Coveney, M. y R. Wetzel. 1995. Biomass, production, and specific growth rate of bacterioplancton and coupling to phytoplankton in an oligotrophic lake. Limnol Oceanogr. 40: 1187-1200.
Del Giorgio, P., J. Cole y A. Cimbleris. 1997. Respiration rates in bacteria ex-ceed phytoplankton production in unproductive aquatic systems. Na-ture 385: 148-151.
Faithfull, C., M. Huss, T. Vrede y A. Bergström. 2011. Bottom-up carbon sub-sidies and top-down predation pressure interact to affect aquatic food web structure. Oikos 120: 311-320.
Farnell, E. y K. Ward. 2003. Sasonal patterns of viruses, bacteria and dis-solved organic carbon in a riverine wetland. Freshwat. Biol. 48: 841-851.

Relaciones entre el fitoplancton y el bacterioplancton en el río Cataniapo Astiz et al.
254
Findlay, S., M. Pace, D. Lints, J. Cole, N. Caraco y B. Peierls. 1991. Weak cou-pling of bacterial and algal production in a heterotrophic ecosystem: the Hudson River estuary. Limnol. Oceanogr. 36: 268-278.
Gao, G., B. Qin y R. Sommaruga. 2007. The bacterioplankton of lake Taihu, China: abundance, biomass, and production. Hydrobiologia 581: 177-188.
González, S. 1992. Composición, distribución y abundancia de las poblaciones de copépodos en el embalse de Guri. Tesis de Grado, Universidad Simón Bolívar, Caracas. 169 pp.
Graterol, B. y C. Botto. 2003. Los procesos de ocupación del espacio en la cuenca del Cataniapo, estado Amazonas: su impacto sobre el ecosiste-ma y la salud humana. Memorias del V Congreso Venezolano de Ecolo-gía, Puerto Ordaz, Estado Bolívar, Venezuela. 89 pp.
Gurung, T. y J. Urabe. 2010. Trophic and nutrient dynamic aspect of aquatic microbial ecology. Ecoprint 17: 9-16.
Hamilton, S., S. Sippel, W. Lewis Jr y J. Saunders. 1990. Zooplankton abun-dance and evidence for its reduction by macrophyte mats in two Ori-noco floodplain lakes. J. Plank. Res. 12: 345-363.
Harsha, T., S. Yamakanamardi y M. Mahadevaswamy. 2007. Heterotrophic free-living and particle-bound bacterial cell size in the river Cauvery and its downstream tributaries. J. Biosci. 32(2): 363-374.
Havens, K., K. Work y T. East. 2000. Relative efficiencies of carbon transfer from bacteria and algae to zooplankton in a subtropical lake. J. Plankton Res. 22: 1801-1809.
Ibañez, A. y J. Lara. 1997. Algunos aspectos fisicoquímicos y biológicos de las aguas del río Caura (Venezuela), en su parte media. En: J. Rosales y O. Huber (eds), Ecología de la Cuenca del Río Caura, Venezuela. II. Estudios especiales. Scientia Guaianae 6: 29-33.
Infante, J. 1992. Diagnóstico físico conservacionista de la cuenca del río Cata-niapo con fines de ordenamiento. MPPAMB. Puerto Ayacucho, Estado Amazonas, Venezuela. 50 pp.
Kepner, R. y J. Pratt. 1994. Use of fluorochromes for direct enumeration of total bacteria in environmental samples: past and present. Microbiol. Rev. 58: 603-615.
Leff, L., A. Leff y M. Lemke. 1998. Seasonal chamges in planktonic bacterial as-semblages of two Ohio streams. Freshwat. Biol. 39: 129-134.
Lewis Jr, W. y F. Weibezahn. 1976. Chemistry, energy, flow y community struc-ture in some Venezuelan freshwater. Archiv für Hydrobiologie 50: 145-207.
Lewis Jr, W. y J. Saunders III. 1990. Chemistry y elementary transport by the Orinoco main stem y lower tributaries. In: F. Weibezahn, H. Álvarez y W.

Boletín del Centro de Investigaciones Biológicas Vol. 47, Nº 3, septiembre-diciembre 2013, Pp. 235 - 258
255
Lewis Jr. (eds), El río Orinoco como Ecosistema: 211-240. Editorial Galac, Caracas.
Lewis Jr, W., J. Saunders III y R. Dufford. 1990. Suspended organism y bio-logical carbon flux along the lower Orinoco river. In: F. Weibezahn, H. Álvarez y W. Lewis Jr. (eds), El río Orinoco como Ecosistema: 269-300. Editorial Galac, Caracas.
Lewis Jr, W, S. Hamilton, M. Lasi, M. Rodríguez y J. Saunders III. 2000. Eco-logical determinism on the Orinoco floodplain. BioScience 50(8): 681-692.
Lǿvdal, T., C. Eichner, H. Grossart, V. Carbonell, L. Chou, V. Martin y T. Thingstad. 2008. Competition for inorganic y organic forms of nitrogen y phosphorous between phytoplankton y bacteria during an Emiliana huxleyi sprin bloom. Biogeosciences 5: 371-383
Lucchetti, A. 1996. Estudio de las variaciones del carbono, nitrógeno y fósfo-ro en el río Cataniapo, Edo. Amazonas. Tesis de Grado. Coordinación de Biología, Universidad Simón Bolívar, Caracas. 77 pp.
Lund, J., C. Kipling y E. Le Cren. 1958. The inverted microscope method of es-timating algal number y the statistical basis of estimations by counting. Hydrobiologia 11: 143-170.
Mariazzi, A., M. Di Siervi, J. Donadelli y L. Albino. 1998. The annual cycle of bacterial numbers, biovolumes y productivities in the Exequiel Ramos Mexía reservoir (Río Negro, Argentina). Verh. Internat. Verein. Limnol., 26: 1631-1635.
Martin, E. 1989. Variaciones espaciales y temporales de las características físi-co-químicas del agua y de la comunidad zooplanctónica de dos lagunas rebalseras del Orinoco (Matanzas, Edo. Bolívar) y su relación con des-cargas industriales. Tesis de Grado, Universidad Simón Bolívar, Caracas. 234 pp.
Medina, M. y E. Vásquez. 1988. Estudio de los rotíferos de una laguna de inun-dación de aguas negras del bajo Caroní, Venezuela. Mem. Soc. Cienc. Nat. La Salle 130: 105-120.
Mirye, V. y B. Tracanna. 2005. Fitoplancton de un río del noroeste Argentino contaminado por efluentes azucareros y cloacales. Bol. Soc. Argent. Bot. 40(3-4): 169-182.
Montoya, J. 1999. Abundancia y biomasa del bacterioplancton en la Cuenca baja del río Caura, Edo. Bolívar, Venezuela. Tesis de Grado. Universidad Simón Bolívar. Caracas. 120 pp.
Montoya, J., M. Castillo y L. Sánchez. 2011. La importancia de las inundacio-nes periódicas para el funcionamiento y conservación de los ecosiste-mas inundables de gryes ríos tropicales: estudios en la cuenca del Orino-co. Interciencia 36(12): 900-907.

Relaciones entre el fitoplancton y el bacterioplancton en el río Cataniapo Astiz et al.
256
Nabaout, J., I. Nogueira y L. Oliveira. 2006. Phytoplankton community of floodplain lakes of the Araguaia river, Brazil, in the rainy y dry seasons. J. Plank. Res. 28(2): 181-193.
Niu, Y., H. Shen, J. Chen, P. Xie, X. Yang, M. Tao, Z. Ma y M. Qi. 2011. Phyto-plankton community succession shaping bacterioplankton community composition in lake Taihu, China. Wat. Res. 45: 4169-4182.
Nogueira, M., M. Ferrareze, M. Moreira y R. Gouvêa. 2010. Phytoplankton assemblages in a reservoir cascade of a large tropical-subtropical river (SE, Brazil). Braz. J. boil. 70(3): 781-793.
Nordin, C., C. Cranston y A. Mejia. 1983. New technology for measurement water y suspended sediment discharge of large rivers. Proc. 2nd. Int. Symp. On River Sedimentation. Water Resources y Electric Power Press. Beijing. 1145 pp.
Núñez, M. 1985. Flujo de fósforo a través de la Cuenca del Río Cataniapo, T.F.A., y su influencia sobre la química de ese elemento en las aguas de escorrentía de cauce. Tesis de Grado, Universidad Simón Bolívar, Cara-cas. 161pp.
Núñez, M. 1993. Diagnóstico del estado actual de la cuenca del Cataniapo, con énfasis en calidad de agua. Tesis de Maestría. Universidad Simón Bolívar. Caracas. 141 pp.
Palijan, G., I. Bogut y J. Vidakovic. 2007. Effects of high water levels on bac-terioplankton abundance in the Danuba river floodplain (Kopački Rit, Croatia). Polish. J. of Environ. Stud. 16(1): 113-121.
Paolini, J. 1990. Carbono orgánico disuelto y particulado en gryes ríos de la América del Sur. Interciencia 15(6): 358-366.
Papadakis, J. 2003. Relaciones espaciales y temporales del zooplancton en el río Orinoco en su cuenca media. Tesis de Grado, Universidad Simón Bolí-var, Caracas. 138 pp.
Pérez, J. 1995. Estudio del bacterioplancton en el embalse de Guri y su relación con parámetros limnológicos. Tesis de Grado. Universidad Simón Bolí-var. Caracas. 151 pp.
Pérez, M. 2002. Fitoplancton del río Negro, Uruguay. Limnetica 21(1-2): 81-92.Petrucio, M., F. Barbosa y A. Furtado. 2006. Bacterioplankton y phytoplank-
ton production in seven lakes in the middle río Doce basin, south-east Brazil. Limnologica 36: 192-203.
Pinilla, G., A. Canosa, A. Vargas, M. Gavilán y L. López. 2007. Acoplamiento entre las comunidades planctónicas de un lago amazónico de aguas cla-ras (lago Boa, Colombia). Limnetica 26(1): 53-65.
Plan de Ordenamiento y Reglamento de Uso de la Zona Protectora de la Cuenca Hidrográfica del Río Cataniapo. 2005. Consulta pública de la

Boletín del Centro de Investigaciones Biológicas Vol. 47, Nº 3, septiembre-diciembre 2013, Pp. 235 - 258
257
propuesta del Ministerio del Ambiente, 79 pp. Puerto Ayacucho, Vene-zuela.
Portillo, M., S. Yerson y N. Fierer. 2012. Temporal variability in the diversity y composition of stream bacterioplankton communities. Environ. Micro-biol.
Pulido, E., E. Ortega, R. Morales y I. Reche. 2003. El papel de la escala en los patrones de variación del bacterioplancton en lagunas de alta montaña. Limnetica 22(1-2): 183-193.
Reverol, Y., J. Delgado, C. López y L. Sánchez. 2008. Zooplankton commu-nity composition in floodplain lakes of Caura river, Venezuela. Boletín del Centro de Investigaciones Biológicas 42(1): 53-72.
Rosenstock, B. y M. Simon. 2001. Sources y sinks of dissolved free amino acids y protein in a large y deep mesotrophic lake. Limnol. Oceanogr. 46:644-654.
Sánchez, L. y E. Vásquez. 1986. Estudio estacional y longitudinal de la hidroquí-mica y fitoplancton en una sección del bajo Orinoco, (Venezuela). Mem. Soc. Cienc. Nat. La Salle 125-126.
Saunders, J. y W. Lewis Jr. 1988a. Zooplankton abundance y transport in a tropical white-water river. Hydrobiologia 162: 147-155.
Saunders, J. y W. Lewis Jr. 1988b. Dynamics y control mechanism in a tropical zooplankton community (Lake Valencia, Venezuela). Ecological Mono-graphs 58: 337-353.
Sioli, H. 1984. The Amazon limnology y lyscape ecology of a mighty tropical river y its basin. Dr. W. Junk Publishers, Boston. 763 pp.
Schmidt, G. 1970. Numbers of bacteria y algae y their interrelations in some Amazonian waters. Amazoniana. 2(4): 393-400.
Teixeira, M., N. Santana, J. Rodrigues y T. Pagioro. 2011. Bacterioplankton features y its relations with DOC characteristics y other limnological vari-ables in Paraná river floodplain environments (PR/MS-Brazil). Braz. J. Mi-crobiol. 42: 897-908.
Van Den Meersche, K., J. Middelburg, K. Soetaert, P. Van Rijswijk, H. Bosch-ker, y C. Heip. 2004. Carbon-nitrogen coupling y algal-bacterial interac-tions during an experimental bloom: Modeling a 13C tracer experiment. Limnol. Oceanogr. 49(3): 865-878.
Vásquez, E. 1984a. Estudio de las comunidades de rotíferos del Orinoco Me-dio, Bajo Caroní y algunas lagunas de inundación. Mem. Soc. Cienc. Nat. La Salle 44: 95-108.
Vásquez, E. 1984b. El zooplancton de la sección baja de un río de aguas negras (río Caroní) y de un embalse hidroeléctrico (Macagua), Venezuela. Mem. Soc. Cienc. Nat. La Salle 44: 109-129.

Relaciones entre el fitoplancton y el bacterioplancton en el río Cataniapo Astiz et al.
258
Vásquez, E. y L. Sánchez. 1984. Variación estacional del plancton en dos sec-tores del río Orinoco y una laguna de inundación adyacente. Mem. Soc. Cienc. Nat. La Salle 121: 11-34.
Vásquez, E. y J. Rey. 1989. A longitudinal study of zooplankton along the lower Orinoco river y its Delta (Venezuela). Annals of Limnology. 25:107-120.
Vásquez, E. 1989. The Orinoco river: a review of hydrobiological research. Reg-ulated Rivers 3: 381-392.
Vásquez, E. y J. Rey. 1992. Composition, abundance y biomass of zooplankton in Orinoco floodplain lakes, Venezuela. Annals of Limnology. 28:3-18.
Vásquez, E., M. Pardo, E. Zoppi De Roa y C. López. 1998. Rotifer fauna from Venezuela. Amazoniana 15: 11-24.
Villaró, M. 1997. Variaciones del bacterioplancton y carbono orgánico disuel-to en dos ríos del Edo. Amazonas. Tesis de Maestría. Universidad Simón Bolívar. Caracas. 206 pp.
Weibezahn, F. 1990. Hidroquímica y sólidos suspendidos en el alto y medio Ori-noco. En: F. Weibezahn, H. Álvarez y W. Lewis (eds.). El río Orinoco como ecosistema: 151-210. Editorial Galac. Caracas, Venezuela.
Weibezahn, F., H. Álvarez, H. Carvajal y V. Park. 1990. Investigaciones sobre el plancton del río Orinoco en sus sectores alto y medio. Instituto de Re-cursos Naturales, Universidad Simón Bolívar, Caracas. 142 pp.
Wetzel, R. y G. Likens. 1992. Limnological Analysis. 2nd edition. Springer-Verlag. New York. 391 pp.
Wetzel, R. 2001. Limnology-Lake y River Ecosystems, 3th ed. Academic Press, San Diego. 1006 pp.
Zalocar, Y., A. Poi y S. Casco. 2007. Abundance y diversity of phytoplankton in the Paraná river (Argentina) 220 km downstream of the Yaciretá res-ervoir. Bras. J. Biol. 67(1): 53-63.
Zöllner, E., B. Santer, M. Boersma, H. Hoppe y K. Jürgens. 2003. Cascading predation effects of Daphnia y copepods on microbial food web compo-nents. Freshwat. Biol. 48: 2174-2193.
Zoppi De Roa, E., F. Michelangelli y L. Segavia. 1985. Cladocera (Crustacea, Branchipoda) de sabanas inundables de Mantecal, Estado Apure, Vene-zuela. Acta Biológica Venezuelica 12: 43-55.




















