
Polymeric substances and biofilms as biomarkers in terrestrial materials: Implications for...
17
JOURNAL OF GEOPHYSICAL RESEARCH, VOL. 105,NO. El0, PAGES 24,511-24,527, OCTOBER 25, 2000 Polymeric substances and biofilms as biomarkers in terrestrial materials' Implications for extraterrestrial samples Frances Westall, •,2 Andrew Steele, • Jan Toporski, 3 Maud Walsh, 4Carlton Allen, sSean Guidry, 6 David McKay, • Everett Gibson, • and Henry Chafetz 6 Abstract. Organic polymeric substances are a fundamental component of microbial biofilms. Microorganisms, especially bacteria, secrete extracellular polymeric substances (EPS) to form slime layers in whichtheyreproduce. In the sedimentary environment, biofilmscommonly contain theproducts of degraded bacteria aswell asallochthonous andautochthonous mineral components. They arecomplex structures whichserve asprotection for thecolonies of microorganisms living in themandalsoact asnutrient traps. Biofilms are almost ubiquitous wherever there is an interface andmoisture (liquid/liquid, liquid/solid, liquid/gas, solid/gas).In sedimentary rocks they are commonly recognized asstromatolites. We alsodiscuss the distinction between bacterial biofilms andprebiotic films. The EPS andcell components of themicrobial biofilmscontain manycation chelation sites which are implicated in the mineralization of the films. EPS,biofilms,andtheirrelated components thus havestrong preservation potential in the rockrecord. Fossilized microbial polymeric substances (FPS) andbiofilms appear to retain the same morphological characteristics asthe unfossilized material andhavebeen recognized in rock formations dating backto the Early Archaean (3.5 b.y.). We describe FPS andbiofilmsfrom hot spring, deep-sea, volcanic lake,and shallow marine/littoral environments ranging up to 3.5 b.y. in age. FPSandbiofilms aremorecommonly observed than fossil bacteria themselves, especially in the older part of the terrestrial record. The widespread distribution of microbial biofilmsandtheir great survival potential makes theirfossilized remains a useful biomarker asa proxyfor life with obvious application to thesearch for life in extraterrestrial materials. 1. Introduction Microbialbiofilmsare almost ubiquitous in nature wherethere is moisture andan interface of anykind (liquid-liquid, such asoil andwater;liquid-gas, such aswater surface; solid-liquid, such as immersed surfaces; andsolid-gas, such assurfaces exposed to the air [Neu, 1994]). The widespread distributionof biofilms, especially in stressed environments, and the specificmicrobial phenotypesforming them suggest that microbial biofilm formation maybe a veryearlycharacteristic of life which perhaps aroseas a natural consequence of the (for modem organisms) harsh conditions existing at that time [Pierson et al., 1993; Costerton et al., 1995]. This is reinforced when the advantages of biofilm formation to an organism are taken into account: protection from dessication, UV radiation, heavy metals, biocides, and other antimicrobial agents [Geesey and Jang, 1989a, 1989b]. The early histories of three of the terrestrial planets, Venus, Mars, andEarth, weresimilar [dakosky, 1998], thus prompting the consideration that microbial biofilms could •NASA Johnson Space Center, Houston, Texas. 2Now atLunar and Planetary Institute, Houston, Texas. 3School of Earth andEnvironmental Sciences, Astrobiology Group, University of Portsmouth, Portsmouth, England, United Kingdom. 4Institute for Environmental Sciences, Louisiana StateUniversity, Baton Rouge. 5Lockheed Martin Space Operations, Houston, Texas. 6Department of Geosciences, University of Houston, Houston, Texas. Copyright 2000 by the American Geophysical Union. Paper number 2000JE001250. 0148-0227/00/2000JE001250509.00 have been common features of potentialearly life on all these planets. Microbial biofilms, including, by definition, stromatolites, appear to be a persistent biomarkerthroughout the history of lifeon Earth and could therefore be a potentially powerful tool in the search for extraterrestrial life. The discussion concerning the evidence of possible life in Martian meteorite ALH84001 [McKay et al., 1996] has stimulated research into all aspects relating to life and its formation and characteristics in the extraterrestial environment and has ledto a recognition of theimportance of interdisciplinary collaboration, as in NASA's virtual Astrobiology Institute. One aspect of this research relates to biomarker identification and characterization. Biomarkers arethemorphological (e.g.,cellular or body fossils),physical (e.g., different crystallography of biogenic minerals), chemical (e.g., traceelements or molecular signatures), or isotopic (e.g., •13C) remains oflife. In the course of investigating the terrestrial recordof fossil bacteria, we became awareof the very widespread distribution of fossilized polymeric substances (FPS) of microbiological origin andfossilized microbial biofilms (definitions given below), both as morphological and, preliminary data suggests, as chemical signatures for life. In fact, FPS seems to be far morecommon than the fossil bacteria themselves, especially in the older parts of the rock record [Walsh, 1992; Westall et al., 2000a]. We therefore consider it appropriate to address polymeric substances and biofilms per se as potentially useful biomarkers for life in both terrestrial and extraterrestrial materials. The followingdefinitions are outlined to clarify the use of acronyms usedin this paper:(1) EPS, extracellular polymeric substances secreted by bacteria through the cell envelope (including capsular material which is closely associated with the cell as well as exopolysaccharides which are more loosely 24,511
-
Upload
carnegiescience -
Category
Documents
-
view
1 -
download
0
Transcript of Polymeric substances and biofilms as biomarkers in terrestrial materials: Implications for...

JOURNAL OF GEOPHYSICAL RESEARCH, VOL. 105, NO. El0, PAGES 24,511-24,527, OCTOBER 25, 2000
Polymeric substances and biofilms as biomarkers in terrestrial materials' Implications for extraterrestrial samples
Frances Westall, •,2 Andrew Steele, • Jan Toporski, 3 Maud Walsh, 4 Carlton Allen, s Sean Guidry, 6 David McKay, • Everett Gibson, • and Henry Chafetz 6
Abstract. Organic polymeric substances are a fundamental component of microbial biofilms. Microorganisms, especially bacteria, secrete extracellular polymeric substances (EPS) to form slime layers in which they reproduce. In the sedimentary environment, biofilms commonly contain the products of degraded bacteria as well as allochthonous and autochthonous mineral components. They are complex structures which serve as protection for the colonies of microorganisms living in them and also act as nutrient traps. Biofilms are almost ubiquitous wherever there is an interface and moisture (liquid/liquid, liquid/solid, liquid/gas, solid/gas). In sedimentary rocks they are commonly recognized as stromatolites. We also discuss the distinction between bacterial biofilms and prebiotic films. The EPS and cell components of the microbial biofilms contain many cation chelation sites which are implicated in the mineralization of the films. EPS, biofilms, and their related components thus have strong preservation potential in the rock record. Fossilized microbial polymeric substances (FPS) and biofilms appear to retain the same morphological characteristics as the unfossilized material and have been recognized in rock formations dating back to the Early Archaean (3.5 b.y.). We describe FPS and biofilms from hot spring, deep-sea, volcanic lake, and shallow marine/littoral environments ranging up to 3.5 b.y. in age. FPS and biofilms are more commonly observed than fossil bacteria themselves, especially in the older part of the terrestrial record. The widespread distribution of microbial biofilms and their great survival potential makes their fossilized remains a useful biomarker as a proxy for life with obvious application to the search for life in extraterrestrial materials.
1. Introduction
Microbial biofilms are almost ubiquitous in nature where there is moisture and an interface of any kind (liquid-liquid, such as oil and water; liquid-gas, such as water surface; solid-liquid, such as immersed surfaces; and solid-gas, such as surfaces exposed to the air [Neu, 1994]). The widespread distribution of biofilms, especially in stressed environments, and the specific microbial phenotypes forming them suggest that microbial biofilm formation may be a very early characteristic of life which perhaps arose as a natural consequence of the (for modem organisms) harsh conditions existing at that time [Pierson et al., 1993; Costerton et al., 1995]. This is reinforced when the advantages of biofilm formation to an organism are taken into account: protection from dessication, UV radiation, heavy metals, biocides, and other antimicrobial agents [Geesey and Jang, 1989a, 1989b]. The early histories of three of the terrestrial planets, Venus, Mars, and Earth, were similar [dakosky, 1998], thus prompting the consideration that microbial biofilms could
•NASA Johnson Space Center, Houston, Texas. 2Now at Lunar and Planetary Institute, Houston, Texas. 3School of Earth and Environmental Sciences, Astrobiology Group,
University of Portsmouth, Portsmouth, England, United Kingdom. 4Institute for Environmental Sciences, Louisiana State University,
Baton Rouge. 5Lockheed Martin Space Operations, Houston, Texas. 6Department of Geosciences, University of Houston, Houston, Texas.
Copyright 2000 by the American Geophysical Union.
Paper number 2000JE001250. 0148-0227/00/2000JE001250509.00
have been common features of potential early life on all these planets. Microbial biofilms, including, by definition, stromatolites, appear to be a persistent biomarkerthroughout the history of life on Earth and could therefore be a potentially powerful tool in the search for extraterrestrial life.
The discussion concerning the evidence of possible life in Martian meteorite ALH84001 [McKay et al., 1996] has stimulated research into all aspects relating to life and its formation and characteristics in the extraterrestial environment
and has led to a recognition of the importance of interdisciplinary collaboration, as in NASA's virtual Astrobiology Institute. One aspect of this research relates to biomarker identification and characterization. Biomarkers are the morphological (e.g., cellular or body fossils), physical (e.g., different crystallography of biogenic minerals), chemical (e.g., trace elements or molecular signatures), or isotopic (e.g., •13C) remains of life.
In the course of investigating the terrestrial record of fossil bacteria, we became aware of the very widespread distribution of fossilized polymeric substances (FPS) of microbiological origin and fossilized microbial biofilms (definitions given below), both as morphological and, preliminary data suggests, as chemical signatures for life. In fact, FPS seems to be far more common than the fossil bacteria themselves, especially in the older parts of the rock record [Walsh, 1992; Westall et al., 2000a]. We therefore consider it appropriate to address polymeric substances and biofilms per se as potentially useful biomarkers for life in both terrestrial and extraterrestrial materials.
The following definitions are outlined to clarify the use of acronyms used in this paper: (1) EPS, extracellular polymeric substances secreted by bacteria through the cell envelope (including capsular material which is closely associated with the cell as well as exopolysaccharides which are more loosely
24,511
24,512 WESTALL ET AL.: EPS AS A FOSSIL BIOMARKER
associated with the cell). This material is polysaccharide rich and extremely hydrated. (2) PS, polymeric substances, which is a generic term for any complex organic molecule within the bacterial biofilm and covers both EPS and cell envelope components to account for the possible presence of degraded (diagenetically altered) biomolecules within the fossilized structures that this paper serves to highlight. (3) FPS, fossilized polymeric substances, which covers both partially mineralized PS that may still contain organic biomolecules and PS that served as a template for mineralization and has since been completely degraded.
Biofilms have been widely studied in the environmental microbiology field [Cohen and Rosenberg, 1989; $tal and Caumette, 1994], and their involvement with biocorrosion and groundwater contaminants [Characklis and Marshall, 1990; Steele et al, 1995] as well as with sediment stability [Krumbein et al., 1994] is the subject of numerous investigations. In the sedimentary record, microbial biofilms are also well known as macroscopic microbial buildups, such as stromatolites [Walter, 1976; Monty, 1981; Krumbein, 1983; Bernier et al., 1991]. However, the indisputable relationship between bacteria and PS within biofilms and the appearance of FPS and fossilized biofilms as reliable biomarkers in the fossil record have yet to be addressed. Our hypothesis in the following is this: Polymeric substances, biofilms, and their fossilized remains can be regarded as proxy evidence for the presence of bacteria.
This paper is part review and part presentation of new data. We review present understanding of the relationships between modem bacteria, PS, and microbial biofilms and present fossil evidence of PS and biofilms ranging in age from the Recent to 3.5 b.y.
2. The Nature of Biofilms
Microbial cells, including bacteria, fungi, and cyanobacteria, attach to surfaces and form continuous sheets in which
extracellular polymeric substances form the matrix which holds the colonies of microbial cells firmly to a surface (structural details given below) [Geesey, 1982; Costerton et al., 1978, 1987; Characklis and Cooksey, 1990; Characklis and Marshall, 1990]. Within this framework, microbial cells metabolize and reproduce, thus extending the matrix of polymeric substances. This process leads to the buildup of a slime sheet known as biofilm. In certain conditions, the secretion of EPS by cells in the planktonic phase can cause the formation of floating microbial aggregates or floccules [Turley and Lochte, 1990]. EPS and cell envelope components are "sticky" and attract allochthonous particles which adhere to them. Biogenically mediated precipitation of minerals occurs within biofilms, resulting in a mucoidal structure which is both complex and heterogeneous [Krumbein and $tal, 1991]. Biofilms will also contain the degradation products of bacteria, such as cell envelope remnants and other cellular structures (e.g., flagella or fibrils), which all contribute to the overall nature of the remnant PS. Detailed definitions and descriptions of microbial biofilms are given by Charaklis and Marshall [1990], Krumbein and $tal [ 1991 ], and Costerton et al. [ 1995].
2.1. The Formation of EPS and Biofilms
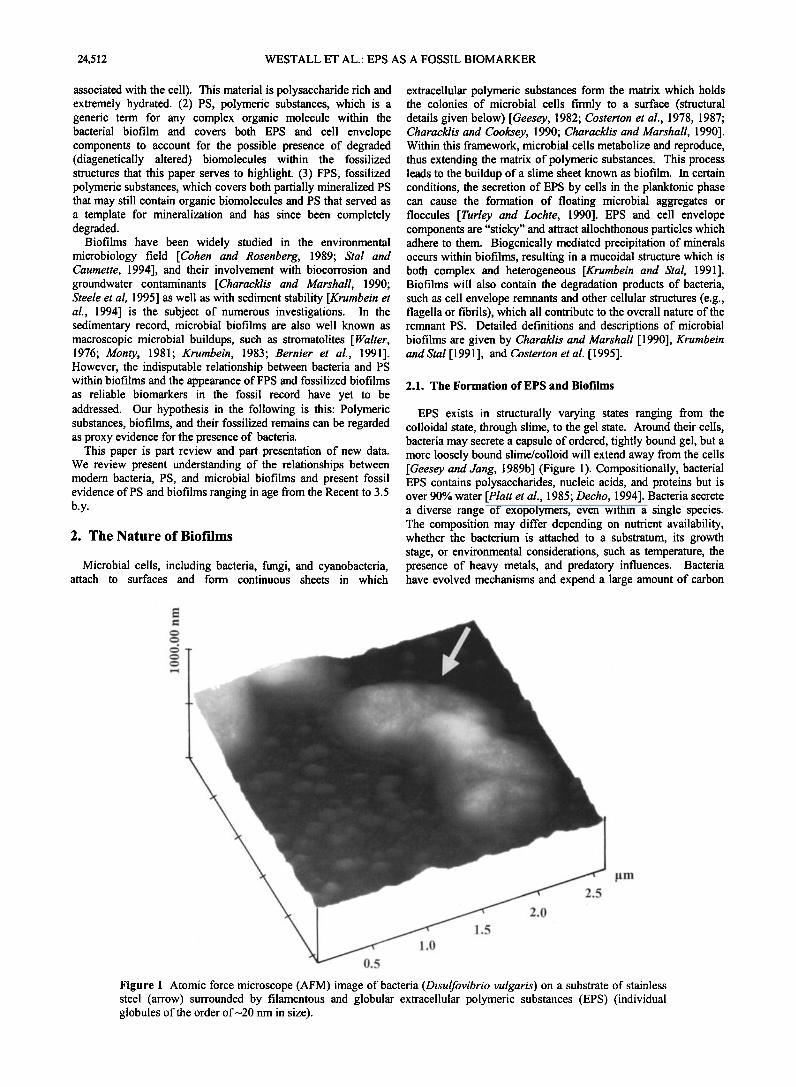
EPS exists in structurally varying states ranging from the colloidal state, through slime, to the gel state. Around their cells, bacteria may secrete a capsule of ordered, tightly bound gel, but a more loosely bound slime/colloid will extend away from the cells [Geesey and Jang, 1989b] (Figure 1). Compositionally, bacterial EPS contains polysaccharides, nucleic acids, and proteins but is over 90% water [Platt et al., 1985; Decho, 1994]. Bacteria secrete a diverse range of exopolymers, even within a single species. The composition may differ depending on nutrient availability, whether the bacterium is attached to a substratum, its growth stage, or environmental considerations, such as temperature, the presence of heavy metals, and predatory influences. Bacteria have evolved mechanisms and expend a large amount of carbon
........ •..•.•.....•7 • ....... •:•m: ½,•7....-...:.•:--- •:--:•.. :.•r•D½• *
:.. -..
1.5
illlB
Figure 1. Atomic force microscope (AFM) image of bacteria (Disulfovibrio vulgaris) on a substrate of stainless steel (arrow) surrounded by filamentous and globular extracellular polymeric substances (EPS) (individual globules of the order of•20 nm in size).

WESTALL ET AL.: EPS AS A FOSSIL BIOMARKER 24,513
dissol'ved organic molecules
attached bacteria multiply feversable
irrever•,•abte att. ac•m•nt
transport to surface
colony formation
in EPS •o• to t
.....
0 = coccoid bacteria
<:::::> = rod-shaped bacteria
C• = authigenic crystal growth i•i = gas bubbles
..-. = allochthonous particles
= EPS
' "• = conditioning layer Illlltlllllilll = substrate
:::::::::::::::::::::::::: :: ::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::: ..... :-.:_. _- ...................... ::.....: .:.:..:..:L...:....• ......
Figure 2. The formation of biofilm: (1) organic molecules nucleate onto a substrate surface (the substrate may be a gas, liquid, or mineral interface) to form a conditioning film; (2) unattached bacteria settle onto the conditioned surface and (3) attach themselves either reversibly or irreversibly; (4) the bacteria multiply and exude EPS to build up a biofilm; (5) allochthonous particles are trapped by the "stickiness" of the PS, authigenic minerals precipitate as a result of bacterial control of the physicochemical conditions within the biofilm, and gas bubbles form owing to bacterial gas production, producing a very complex and heterogeneous biofilm; (6) periodic sloughing of the biofilm occurs.
and energy to secrete these strands through their cell envelope and into the external environment. The advantage of EPS for them is the enormously high water content, which ensures that the cell is protected from desiccation and enables reactions across the cell membrane to continue even in the harshest of conditions.
EPS also provides protection from biocides, antibiotics, predatory organisms, host immune systems, and toxic metal ions and stabilizes the microenvironment around the cell in the case of
sudden changes in the environment [Wimpenny, 1995; Geesey and Jang, 1989a, 1989b]. Finally, EPS is extremely important in helping cells form biofilms by being the adhesive polymer which attaches the cells to surfaces, and therefore EPS is the most common part of a bacterium to interact directly with the substratum.
Changes in the composition of the EPS affect its metal-binding properties and consequently the robustness of its structure [Decho, 1994]. The metal-binding capacity, a critical
characteristic for the fossilization of biofilms, is a useful ecological function for two reasons: in the first place, EPS can trap essential nutrients and trace elements in this way (e.g., Ca, Mg, Na, and Ni to name just a few), and in the second place, the exopolymer matrix protects the cell from toxic metals in the environment (e.g., Pb, Cu, Ni, Cd, Cr, Mo, Hg, U, etc.) by chelating them [Collins and Stotzky, 1989; Allison, 1993; Decho, 1994]. In fact, the production of EPS can increase in response to metal ion concentration [Geesey and Jang, 1989a]. Metal binding to EPS is a passive process, but bacteria can also actively bind (toxic) metal ions within their cells to neutralize them [Brierley et al., 1985, 1989; Beveridge, 1989], hence their application in the bioremediation field.
Biofilm formation on natural and artificial surfaces occurs in a
number of discrete steps (Figure 2) [Costerton et al., 1978; Fletcher and Marshall, 1982; Characklis and Cooksey, 1990; Characklis and Marshall, 1990; Fleming and Geesey, 1991 ].
24,514 WESTALL ET AL.: EPS AS A FOSSIL BIOMARKER
:::::::::::::::::::::::::: ..•.:•i :•:.•::'"'"'•:•i!• ..........
.::. ""'•:•...:..:•. .. .•::)'
•..
e i?:::•?•?•$ii•½):•,,.::•i•..•.%ii?:::•-:--•%.:½!•:•i:•i:½•;:•:'•½!:• •'":'.-• "'"::':":•:•:::::::"•:•' '":'"":•':•'•':'"':':•::'::::::":: '""•:• ß :::• ;;•.;:•:.•:. '•'•::;::•; .• ....... :.:...:•.. ......... •:• ................ •½. •::.>.' .:• ....:•.-- :.....• :;%•
................ • :::.:?•'?•?..•?C.?•:C%•:;•;•5•. .•?.'.• ?.'..•': ,:•?•:•. •::•.:. ß '• :?• ' ' •s•::'.. :' :.5: ...................... :."'•',:•' '"•'?':" ?:?.•'•?.?•...•7 ?:'•"'"•"-•? :5•:5•..•:½•::•:• .-..: '....-..-.:.>•?•- :•.:•.: .:. ",•::.?.' •' , ....... :•' . ,:..
•:•:' •:-,•.:::•.• ...:;;•::•--•..-•... ,...,.• ...• :. ::•)' .-%:?: ...... ,:• •} ?-•. • '::'•. ,.:.•;:;:.::........ %•:. "%.: ..
... ..•:.:•::[:: ::::::::::::::::::::::::::::::::: •-•. • :;•.
..... .... ..... , ,f. ff ..:::. '...:..::.• .... ß .......... :• •.•.:•• ......... .• ............................. •{:•?:.';. ..:•:.-
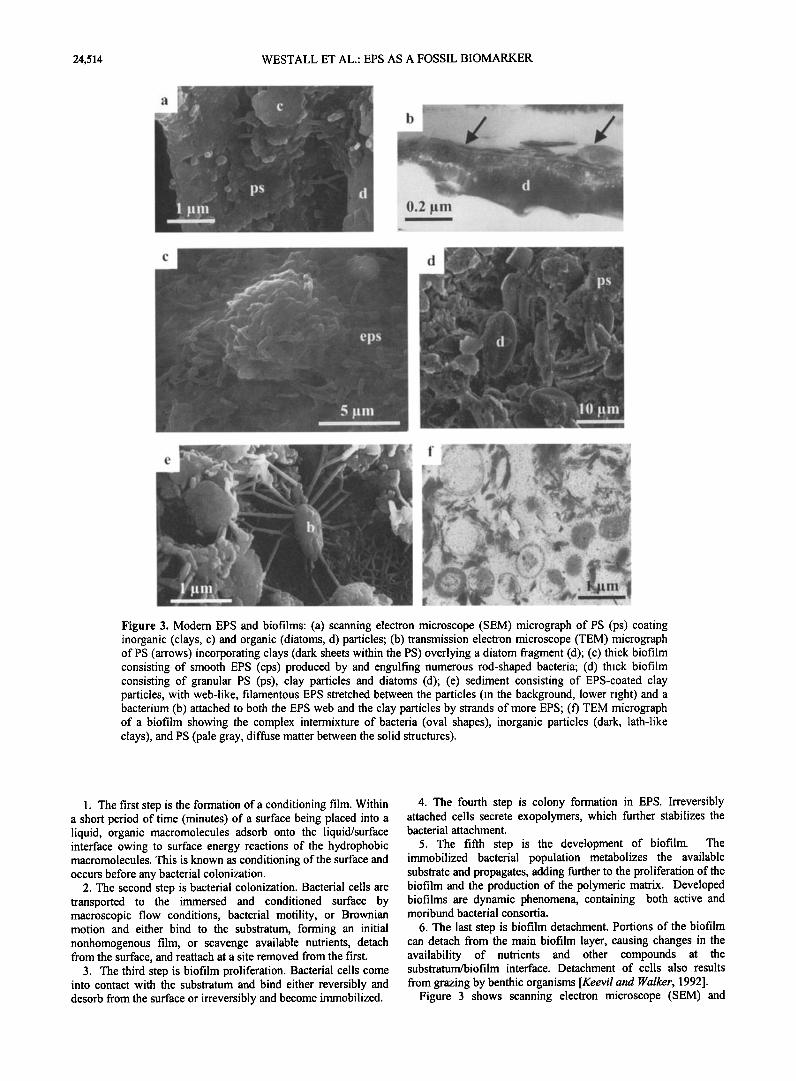
Figure 3. Modem EPS and biofilms: (a) scanning electron microscope (SEM) micrograph of PS (ps) coating inorganic (clays, c) and organic (diatoms, d) particles; (b) transmission electron microscope (TEM) micrograph of PS (arrows) incorporating clays (dark sheets within the PS) overlying a diatom fragment (d); (c) thick biofilm consisting of smooth EPS (eps) produced by and engulfing numerous rod-shaped bacteria; (d) thick biofilm consisting of granular PS (ps), clay particles and diatoms (d); (e) sediment consisting of EPS-coated clay particles, with web-like, filamentous EPS stretched between the particles (in the background, lower right) and a bacterium (b) attached to both the EPS web and the clay particles by strands of more EPS; (f') TEM micrograph of a biofilm showing the complex intermixture of bacteria (oval shapes), inorganic particles (dark, lath-like clays), and PS (pale gray, diffuse matter between the solid structures).
1. The first step is the formation of a conditioning film. Within a short period of time (minutes) of a surface being placed into a liquid, organic macromolecules adsorb onto the liquid/surface interface owing to surface energy reactions of the hydrophobic macromolecules. This is known as conditioning of the surface and occurs before any bacterial colonization.
2. The second step is bacterial colonization. Bacterial cells are transported to the immersed and conditioned surface by macroscopic flow conditions, bacterial motility, or BrownJan motion and either bind to the substratum, forming an initial nonhomogenous film, or scavenge available nutrients, detach from the surface, and teattach at a site removed from the first.
3. The third step is biofilm proliferation. Bacterial cells come into contact with the substratum and bind either reversibly and desorb from the surface or irreversibly and become immobilized.
4. The fourth step is colony formation in EPS. Irreversibly attached cells secrete exopolymers, which further stabilizes the bacterial attachment.
5. The fifth step is the development of biofilm. The immobilized bacterial population metabolizes the available substrate and propagates, adding further to the proliferation of the biofilm and the production of the polymeric matrix. Developed biofilms are dynamic phenomena, containing both active and moribund bacterial consortia.
6. The last step is biofilm detachment. Portions of the biofilm can detach from the main biofilm layer, causing changes in the availability of nutrients and other compounds at the substratum/biofilm interface. Detachment of cells also results
from grazing by benthic organisms [Keevil and •alker, 1992]. Figure 3 shows scanning electron microscope (SEM) and

WESTALL ET AL.: EPS AS A FOSSIL BIOMARKER 24,515
transmission electron microscope (TEM) images of EPS and well-developed microbial biofilms which illustrate the complexity of both cultured and natural biofilms.
One interesting phenomenon related to the formation of microbial biofilms is the fact that some bacterial species forming the biofilms may have a different morphology (phenotype) to that of the same species in the planktonic state. For instance, Charaklis et al. [ 1990] note that most microorganisms in biofilms seem to be filamentous and even bacteria which are not normally filamentous, such as Pseudomonas putida 40, take on a filamentous habit, apparently in response to oxygen stress. Margulls et al. [1980], Krumbein [1983], Neu [1994], and Wachenddirfer et al. [1994], however, note that a variety of small rod-shaped and coccoid-shaped bacteria dominate the subsurface microbial communities in a biofilm. Biofilms are ubiquitous in nature and commonly form in stressed environments such as hypersaline ponds, hot springs, intertidal sediments, high UV environments, and alkaline lakes and streams [Margulls et al., 1976; Cohen and Rosenberg, 1989; Pierson et al., 1993; Castenholz, 1994; Decho, 1994; Costerton et al., 1995].
2.2. The Structure of Bacterial Biofilms
Understanding of the structure of biofilms has been aided enormously by in situ observation using fluorescent dyes in combination with confocal laser scanning microscopy, atomic force microscopy, and environmental scanning electron microscopy [Costerton et al., 1995; Steele et al., 1994, 1995] as well as the use of microsensors [e.g., DeBeer et al., 1994]. These methods have shown that biofilms are heterogeneous in time and space and that they consist of cell clusters or stacks, i.e., conglomerates of cells embedded within a polymeric substance matrix, which may or may not be in contact with the substratum [Costerton et al., 1995; Lewandowski et al., 1995; Costerton, 1995]. Owing to the spatial arrangement of these stacks within the cellular/EPS matrix, vertical spaces (pores) and horizontal spaces (conduits) are formed and are integral parts of the biofilm total volume. Convective fluid flow through these channels has been demonstrated [Costerton et al., 1995]. However, even this relatively complicated model does not do justice to the complexity ofbiofilm systems [Wimpenny, 1995].
Biofilms are affected by a host of factors, including (1) number and types of species present within the biofilm and their individual and combined metabolisms, (2) nature of the substratum and the bulk medium, (3) fluid flow rates, (4) ability of the organisms to utilize the substratum as an energy source, (5) temperature, (6) pH, and (7) the concentrations of essential nutrients, such as carbon, nitrogen, and phosphorous. A change in any one of these factors would produce a concomitant change in the nature and structure of the biofilm. In fact, the organisms within a biofilm do not exist as colonies comprising individual species but as interactive communities of different species [Haack and McFeters, 1982; Dowling et al., 1991]. These consortia are subject to environmental pressures which influence their speciation and spread [Fera et al., 1989; Fletcher, 1992]. Each species acts cooperatively or aggressively to maintain the balance of nutrient flow, genetic exchange [Costerton et al., 1978], and protection [Pope et al., 1983] within the biofilm.
The complexity of biofilms, the interrelationships between the various components and their interactions have lead some to compare well-developed biofilms with tissues in higher organisms [Wachenddirfer et al., 1994; Costerton et al., 1995, p.713]: "Biofilms resemble the tissues formed by eukaryotic cells, in their physiological cooperativity and in the extent to which they are protected from variations in bulk phase conditions by primitive homeostasis provided by the biofilm matrix".
2.3. The Fossilization of PS
The fossilization of PS within biofilms is a direct consequence of its metal (anionic)-binding capacity. In addition, the metal- binding properties of the cell envelopes of bacteria contribute to this process. The metal-binding sites in EPS and within cell envelopes are the carboxylate, hydroxyl, amine, and phosphate (and rarely thiol) active groups within the polysaccharides of the EPS and the bacterial sheaths, proteins, or glycoproteins in the crystalline proteinaceous S layer present in some bacteria and the lipopolysaccharide units within the cell envelope of Gram- negative bacteria [Geesey and Jang, 1989a, 1989b].
The physicochemical environment also influences the metal- binding capacity of EPS. The effects of temperature and pH on the uptake of metals by polysaccharides were studied by Geddie and Sutherland [1993], who found that high temperatures decrease binding capacity at the ion-binding sites by creating disordered geometry and other associated changes. Furthermore, they noted increased cation uptake at higher pHs probably because at lower pHs, there is competition for the binding sites on the polymer from excess protons [Ferris et al., 1989; Welch et al., 1999].
Bacteria can influence the growth of minerals in biofilms in two ways: (1) by their metabolic control of localized environments in which solutes are concentrated from the bulk
solution, resulting in oversaturation, and (2) by providing active sites for nucleation to take place. These active sites may be local or removed from the bacteria depending on the spread of anionic polymer residues away from the cell [Geesey and Jang, 1989b]. An example of this is carbonate precipitation by cyanobacteria, where precipitation is very much a function of photosynthetic activity and the amount of cells actively respiring. The fixation of CO2 by these bacteria brings about an increase in pH due to bicarbonate production that leads to conditions in the microenvironment around the cells which readily favor carbonate precipitation [Ddfarge et al., 1994]. Biomineralization by the process outlined above is not only confined to carbonate but has been described for a diverse variety of bacteria in the precipitation of sulfide, iron, manganese, phosphate, and siliceous minerals.
In terms of searching for fossil evidence of life, numerous investigations have been undertaken to study the fossilization of bacteria but not EPS or other bacterial PS and therefore not for
biofilms per se. Cyanobacteria, with their robust EPS capsules, were of particular interest in these experiments for a number of reasons: (1) the importance of their role in modem mat-forming communities; (2) their apparent dominance of important microbial structures, such as stromatolites, in the past; and (3) the fact that some of the earliest microbial fossils in the rock record
were interpreted as possible cyanobacteria [e.g., Schopf and Packer, 1987; Schopf, 1993]. The thick EPS capsules of the cyanobacteria lend themselves, particularly, to fossilization owing to the large number of cation-binding sites within them (e.g., experimental fossilizations of Oehler and Schopf [1971], Oehler [ 1976], Walters et al. [ 1977], Francis et al. [1978a, 1978b]). In most cases the death and degradation of the organisms within microbial biofilms enhances their susceptibility to preservation by fossilization by providing further nucleation sites [Ferris et al., 1988] and by the deactivation of mechanisms, such as proton pumps (the proton motive force that bacteria use to synthesize andenosine triphosphate, ATP), within the organisms. However, Chafetz and Buczynski [1992] determined that under their experimental conditions, fossilization of autoclaved mats only took place in the presence of live heterotrophs. At present, we are undertaking experimental mineralization of microbial biofilms in order to better understand the process as related to the
24,516 WESTALL ET AL.' EPS AS A FOSSIL BIOMARKER
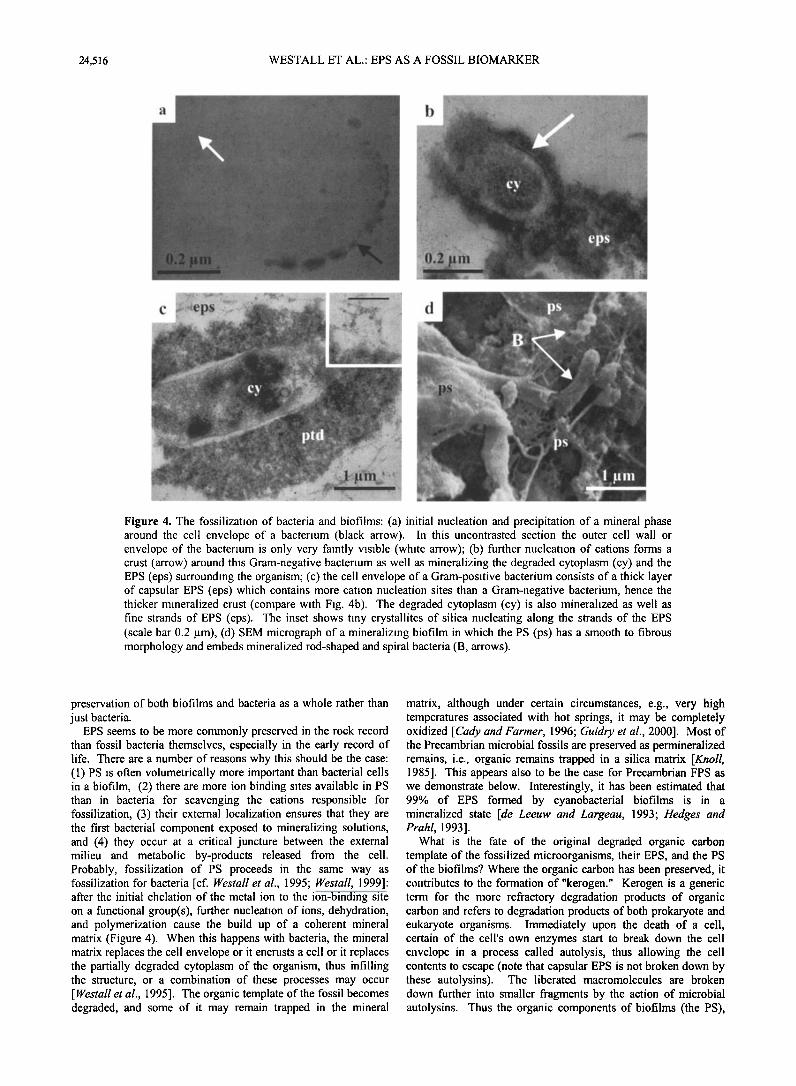
Figure 4. The fossilization of bacteria and biofilms: (a) initial nucleation and precipitation of a mineral phase around the cell envelope of a bacterium (black arrow). In this uncontrasted section the outer cell wall or envelope of the bacterium is only very faintly visible (white arrow'; (b) further nucleation of cations forms a crust (arrow) around this Gram-negative bacterium as well as mineralizing the degraded cytoplasm (cy) and the EPS (eps) surrounding the organism; (c) the cell envelope of a Gram-positive bacterium consists of a thick layer of capsular EPS (eps) which contains more cation nucleation sites than a Gram-negative bacterium, hence the thicker mineralized crust (compare with Fig. 4b). The degraded cytoplasm (cy) is also mineralized as well as fine strands of EPS (eps). The inset shows tiny crystallites of silica nucleating along the strands of the EPS (scale bar 0.2 gm), (d) SEM micrograph of a mineralizing biofilm in which the PS (ps) has a smooth to fibrous morphology and embeds mineralized rod-shaped and spiral bacteria (B, arrows).
preservation of both biofilms and bacteria as a whole rather than just bacteria.
EPS seems to be more commonly preserved in the rock record than fossil bacteria themselves, especially in the early record of life. There are a number of reasons why this should be the case: (1) PS is often volumetrically more important than bacterial cells in a biofilm, (2) there are more ion binding sites available in PS than in bacteria for scavenging the cations responsible for fossilization, (3) their external localization ensures that they are the first bacterial component exposed to mineralizing solutions, and (4) they occur at a critical juncture between the external milieu and metabolic by-products released from the cell. Probably, fossilization of PS proceeds in the same way as fossilization for bacteria [cf. Westall et al., 1995; Westall, 1999]: after the initial chelation of the metal ion to the ion-binding site on a functional group(s), further nucleation of ions, dehydration, and polymerization cause the build up of a coherent mineral matrix (Figure 4). When this happens with bacteria, the mineral matrix replaces the cell envelope or it encrusts a cell or it replaces the partially degraded cytoplasm of the organism, thus infilling the structure, or a combination of these processes may occur [ Westall et al., 1995]. The organic template of the fossil becomes degraded, and some of it may remain trapped in the mineral
matrix, although under certain circumstances, e.g., very high temperatures associated with hot springs, it may be completely oxidized [Cady and Farmer, 1996; Guidry et al., 2000]. Most of the Precambrian microbial fossils are preserved as permineralized remains, i.e., organic remains trapped in a silica matrix [Knoll, 1985]. This appears also to be the case for Precambrian FPS as we demonstrate below. Interestingly, it has been estimated that 99% of EPS formed by cyanobacterial biofilms is in a mineralized state [de Leeuw and Largeau, 1993; Hedges and Prahl, 1993].
What is the fate of the original degraded organic carbon template of the fossilized microorganisms, their EPS, and the PS of the biofilms? Where the organic carbon has been preserved, it contributes to the formation of "kerogen." Kerogen is a generic term for the more refractory degradation products of organic carbon and refers to degradation products of both prokaryote and eukaryote organisms. Immediately upon the death of a cell, certain of the cell's own enzymes start to break down the cell envelope in a process called autolysis, thus allowing the cell contents to escape (note that capsular EPS is not broken down by these autolysins). The liberated macromolecules are broken down further into smaller fragments by the action of microbial autolysins. Thus the organic components of biofilms (the PS),
WESTALL ET AL.: EPS AS A FOSSIL BIOMARKER 24,517
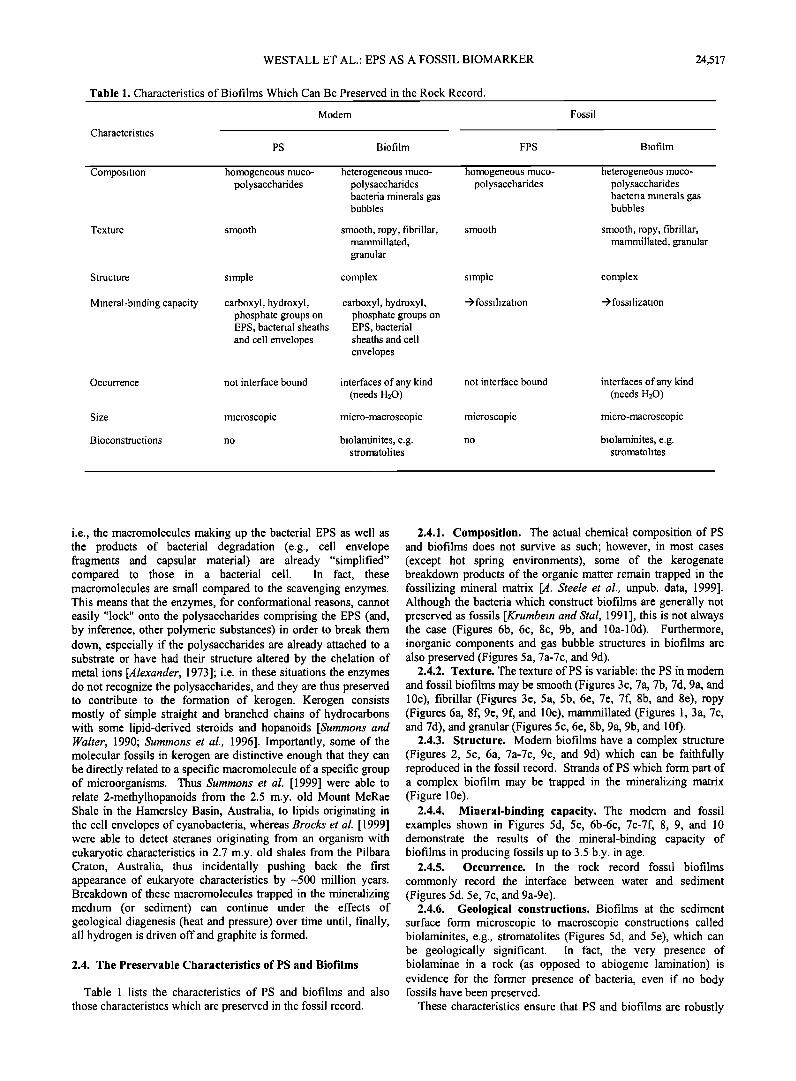
Table 1. Characteristics of Biofilms Which Can Be Preserved in the Rock Record.
Characteristics
Modem
PS Biofilm FPS
Fossil
Biofilm
Composition homogeneous muco- polysaccharides
Texture smooth
heterogeneous muco- polysaccharides bacteria minerals gas bubbles
smooth, ropy, fibrillar, mammillated, granular
homogeneous muco- polysaccharides
heterogeneous muco- polysaccharides bacteria minerals gas bubbles
smooth smooth, ropy, fibrillar, mammillated, granular
Structure simple complex simple complex
Mineral-binding capacity carboxyl, hydroxyl, phosphate groups on EPS, bacterial sheaths and cell envelopes
carboxyl, hydroxyl, phosphate groups on EPS, bacterial sheaths and cell
envelopes
-> fossilization -> fo ssilization
not interface bound interfaces of any kind (needs H20)
micro s copi c mi cro-macro s copi c
no biolaminites, e.g. stromatolites
Occurrence not interface bound interfaces of any kind (needs H20)
Size microscopic micro-macroscopic
Bioconstructions no biolaminites, e.g. stromatolites
i.e., the macromolecules making up the bacterial EPS as well as the products of bacterial degradation (e.g., cell envelope fragments and capsular material) are already "simplified" compared to those in a bacterial cell. In fact, these macromolecules are small compared to the scavenging enzymes. This means that the enzymes, for conformational reasons, cannot easily "lock" onto the polysaccharides comprising the EPS (and, by inference, other polymeric substances) in order to break them down, especially if the polysaccharides are already attached to a substrate or have had their structure altered by the chelation of metal ions [Alexander, 1973]; i.e. in these situations the enzymes do not recognize the polysaccharides, and they are thus preserved to contribute to the formation of kerogen. Kerogen consists mostly of simple straight and branched chains of hydrocarbons with some lipid-derived steroids and hopanoids [Summons and Walter, 1990; Summons et al., 1996]. Importantly, some of the molecular fossils in kerogen are distinctive enough that they can be directly related to a specific macromolecule of a specific group of microorganisms. Thus Summons et al. [1999] were able to relate 2-methylhopanoids from the 2.5 m.y. old Mount McRae Shale in the Hamersley Basin, Australia, to lipids originating in the cell envelopes of cyanobacteria, whereas Brocks et al. [1999] were able to detect steranes originating from an organism with eukaryotic characteristics in 2.7 m.y. old shales from the Pilbara Craton, Australia, thus incidentally pushing back the first appearance of eukaryote characteristics by-500 million years. Breakdown of these macromolecules trapped in the mineralizing medium (or sediment) can continue under the effects of geological diagenesis (heat and pressure) over time until, finally, all hydrogen is driven off and graphite is formed.
2.4. The Preservable Characteristics of PS and Biofilms
Table 1 lists the characteristics of PS and biofilms and also
those characteristics which are preserved in the fossil record.
2.4.1. Composition. The actual chemical composition of PS and biofilms does not survive as such; however, in most cases (except hot spring environments), some of the kerogenate breakdown products of the organic matter remain trapped in the fossilizing mineral matrix [A. Steele et al., unpub. data, 1999]. Although the bacteria which construct biofilms are generally not preserved as fossils [Krumbein and Stal, 1991 ], this is not always the case (Figures 6b, 6c, 8c, 9b, and 10a-10d). Furthermore, inorganic components and gas bubble structures in biofilms are also preserved (Figures 5a, 7a-7c, and 9d).
2.4.2. Texture. The texture of PS is variable: the PS in modem
and fossil biofilms may be smooth (Figures 3c, 7a, 7b, 7d, 9a, and 10c), fibrillar (Figures 3e, 5a, 5b, 6e, 7e, 7f, 8b, and 8e), ropy (Figures 6a, 8f, 9e, 9f, and 10e), mammillated (Figures 1, 3a, 7c, and 7d), and granular (Figures 5c, 6e, 8b, 9a, 9b, and 1 Of).
2.4.3. Structure. Modem biofilms have a complex structure (Figures 2, 5c, 6a, 7a-7c, 9c, and 9d) which can be faithfully reproduced in the fossil record. Strands of PS which form part of a complex biofilm may be trapped in the mineralizing matrix (Figure 10e).
2.4.4. Mineral-binding capacity. The modem and fossil examples shown in Figures 5d, 5e, 6b-6e, 7c-7f, 8, 9, and 10 demonstrate the results of the mineral-binding capacity of biofilms in producing fossils up to 3.5 b.y. in age.
2.4.5. Occurrence. In the rock record fossil biofilms
commonly record the interface between water and sediment (Figures 5d. 5e, 7c, and 9a-9e).
2.4.6. Geological constructions. Biofilms at the sediment surface form microscopic to macroscopic constructions called biolaminites, e.g., stromatolites (Figures 5d, and 5e), which can be geologically significant. In fact, the very presence of biolaminae in a rock (as opposed to abiogenic lamination) is evidence for the former presence of bacteria, even if no body fossils have been preserved.
These characteristics ensure that PS and biofilms are robustly
24,518 WESTALL ET AL.' EPS AS A FOSSIL BIOMARKER
"::•":i: ..;:7;.:. :?.':::".." ...
.':';:'*".'-'; .'.' .7: ..... - :':-';!.Z".;•. ' .
:;..:.:"" ;.'".;..... •:-'{i.i": •/::'•:• '•.'""½" .,•.......,..•r :.......
"½ :: i':':':.!:.'.. ': i,"t,Lm
.... ..::.. ....... . .....
.. ,
ß t. .........
..... .....
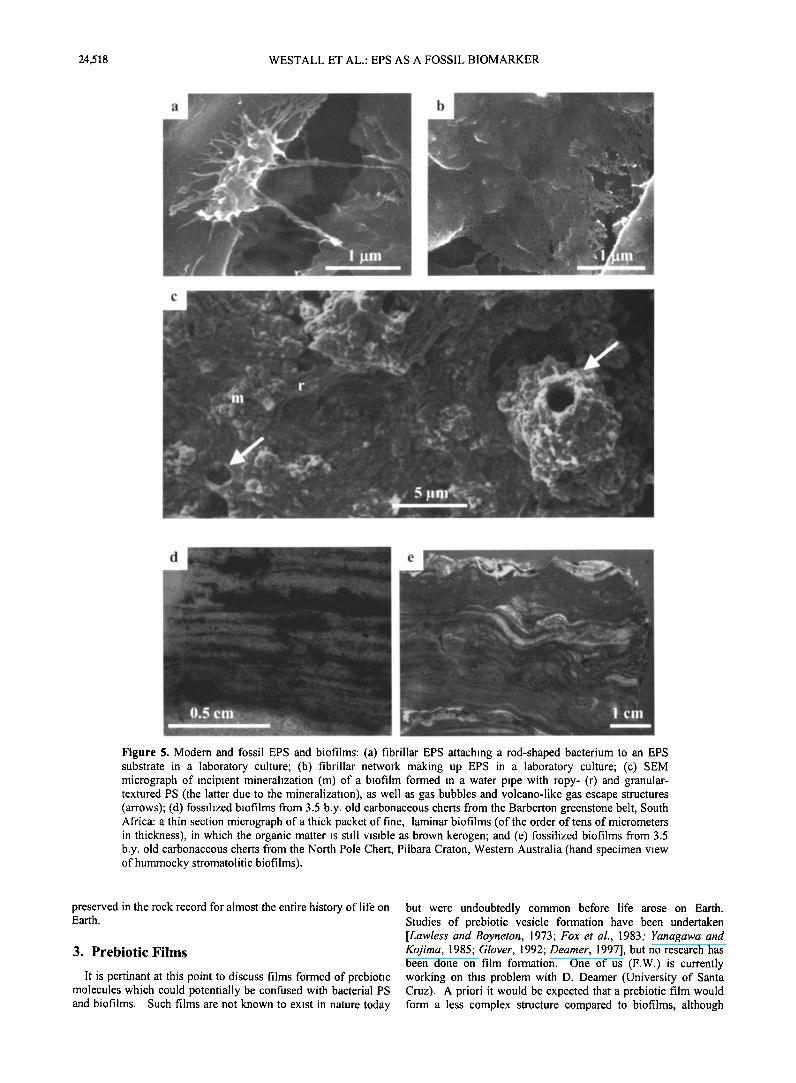
Figure 5. Modern and fossil EPS and biofilms: (a) fibrillar EPS attaching a rod-shaped bacterium to an EPS substrate in a laboratory culture; (b) fibrillar network making up EPS in a laboratory culture; (c) SEM micrograph of incipient mineralization (m) of a biofilm formed in a water pipe with ropy- (r) and granular- textured PS (the latter due to the mineralization), as well as gas bubbles and volcano-like gas escape structures (arrows); (d) fossilized biofilms from 3.5 b.y. old carbonaceous cherts from the Barberton greenstone belt, South Africa: a thin section micrograph of a thick packet of fine, laminar biofilms (of the order of tens of micrometers in thickness), in which the organic matter is still visible as brown kerogen; and (e) fossilized biofilms from 3.5 b.y. old carbonaceous cherts from the North Pole Chert, Pilbara Craton, Western Australia (hand specimen view of hummocky stromatolitic biofilms).
preserved in the rock record for almost the entire history of life on Earth.
3. Prebiotic Films
It is pertinant at this point to discuss films formed of prebiotic molecules which could potentially be confused with bacterial PS and biofilms. Such films are not known to exist in nature today
but were undoubtedly common before life arose on Earth. Studies of prebiotic vesicle formation have been undertaken [Lawless and Boyneton, 1973; Fox et al., 1983; Yanagawa and Kojima, 1985; Glover, 1992; Deamer, 1997], but no research has been done on film formation. One of us (F.W.) is currently working on this problem with D. Deamer (University of Santa Cruz). A priori it would be expected that a prebiotic film would form a less complex structure compared to biofilms, although
WESTALL ET AL.' EPS AS A FOSSIL BIOMARKER 24,519
m
I0 pm
b ..
.-::•':•,•....-.'i::•i•.:,.:-."*•=-"•";•?';• •:'!.. ' >;=.>..--:.• =? ........ ..:,-.•. "•:;;::•'-:..:.-... .... :-='= ß """" =•:-:-.----- ' '%:.•3=!! ....... '<:.:•.::'•;:' ....... "•-.=-' ..,.?:•,' ...... :"a•':•::' .........
.... :.. ..... .;.- ....:...?...
ß . ........ =;.>..=-:.•.%•:(...": ....... '""1%?...... ."-: <--
ß : '..:-=.. .'.'-•:•-.,:..= ... :-. •'--'--., :..:;i: •,/:•'"•. ,: ,.. ' '"'" "•:= - '• .'!'-. - '" •....2 5½ ' •"... ' ;:,;..:, -. :.. % •'-
ß .=/• .... ,,:-.(:..•=.!:.'3.::) "?'.=-';½'..;:.•.;.:.;'• •-.•.:.:,.; ":•7.•?.-=-.•::'- .
' :!<.•? ',.';;:!: - ............. •,, .== .•=• ........ 5 pm ...... ..*...,.'-. ,.:;5¾ . = '•::'"'• ' 'J":'•;=-" '•"',• •' ................ " " .. ' .:?-,-::..:-'%¾: •---:..:=-,•... - .............
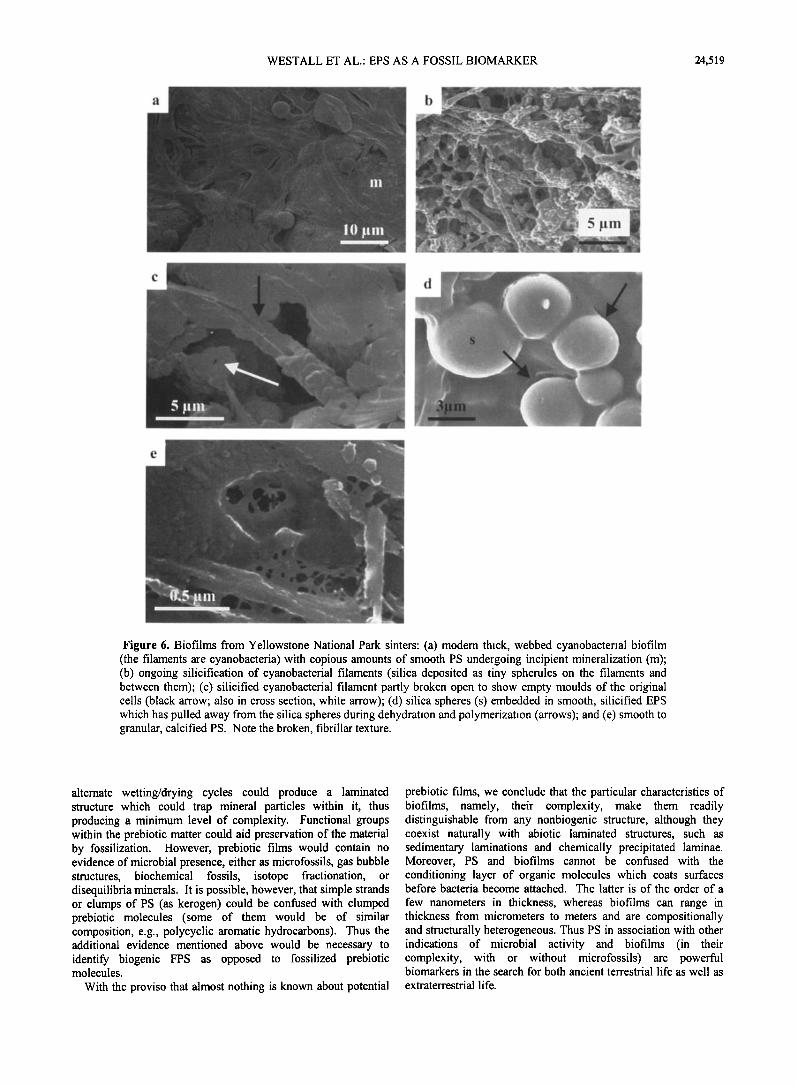
Figure 6. Biofilms from Yellowstone National Park sinters: (a) modem thick, webbed cyanobacterial biofilm (the filaments are cyanobacteria) with copious amounts of smooth PS undergoing incipient mineralization (m); (b) ongoing silicification of cyanobacterial filaments (silica deposited as tiny spherules on the filaments and between them); (c) silicified cyanobacterial filament partly broken open to show empty moulds of the original cells (black arrow; also in cross section, white arrow); (d) silica spheres (s) embedded in smooth, silicified EPS which has pulled away from the silica spheres during dehydration and polymerization (arrows); and (e) smooth to granular, calcified PS. Note the broken, fibrillar texture.
alternate wetting/drying cycles could produce a laminated structure which could trap mineral particles within it, thus producing a minimum level of complexity. Functional groups within the prebiotic matter could aid preservation of the material by fossilization. However, prebiotic films would contain no evidence of microbial presence, either as microfossils, gas bubble structures, biochemical fossils, isotope fractionation, or disequilibria minerals. It is possible, however, that simple strands or clumps of PS (as kerogen) could be confused with clumped prebiotic molecules (some of them would be of similar composition, e.g., polycyclic aromatic hydrocarbons). Thus the additional evidence mentioned above would be necessary to identify biogenic FPS as opposed to fossilized prebiotic molecules.
With the proviso that almost nothing is known about potential
prebiotic films, we conclude that the particular characteristics of biofilms, namely, their complexity, make them readily distinguishable from any nonbiogenic structure, although they coexist naturally with abiotic laminated structures, such as sedimentary laminations and chemically precipitated laminae. Moreover, PS and biofilms cannot be confused with the conditioning layer of organic molecules which coats surfaces before bacteria become attached. The latter is of the order of a
few nanometers in thickness, whereas biofilms can range in thickness from micrometers to meters and are compositionally and structurally heterogeneous. Thus PS in association with other indications of microbial activity and biofilms (in their complexity, with or without microfossils) are powerful biomarkers in the search for both ancient terrestrial life as well as extraterrestrial life.
24,520 WESTALL ET AL.: EPS AS A FOSSIL BIOMARKER
i:.•i .... -,-'.::•,½'":..?:' "%1:i:,::•:.•:::• ..... i'•:::... ..
?&::. -> .... .... '%:?.
;::•
.
7' ::•?•'..:':
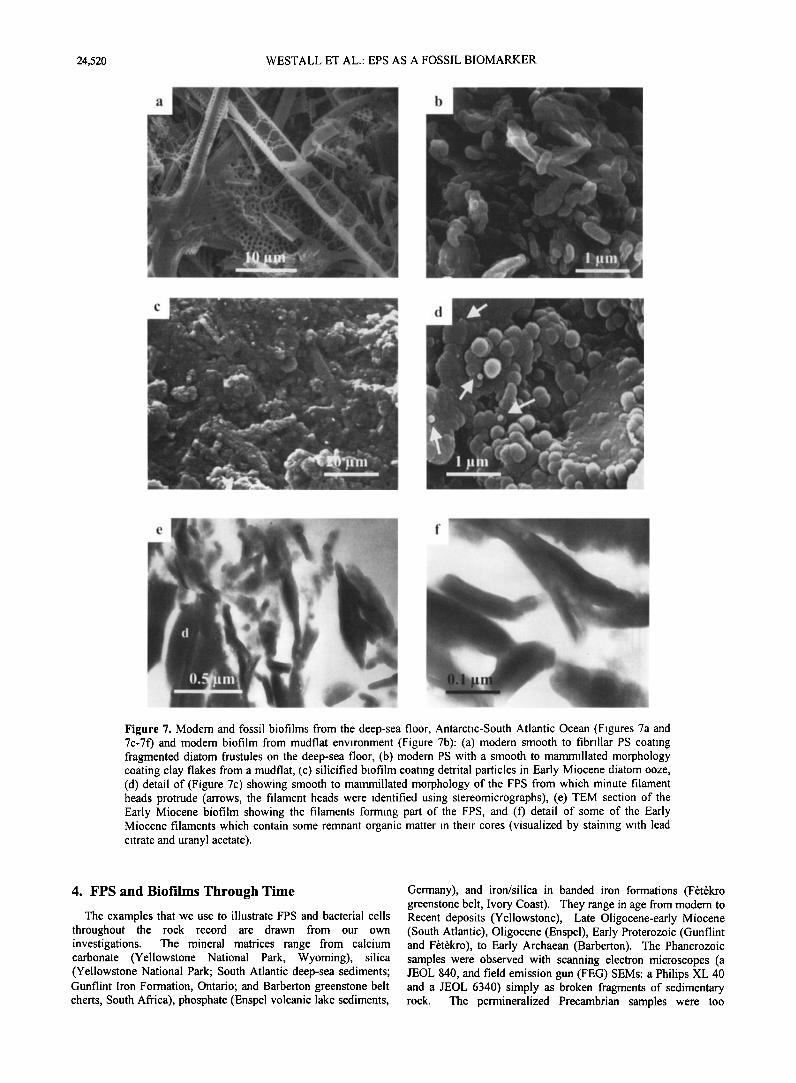
Figure 7. Modem and fossil biofilms from the deep-sea floor, Antarctic-South Atlantic Ocean (Figures 7a and 7c-7f) and modem biofilm from mudflat environment (Figure 7b): (a) modem smooth to fibriliar PS coating fragmented diatom frustules on the deep-sea floor, (b) modem PS with a smooth to mammillated morphology coating clay flakes from a mudflat, (c) silicified biofilm coating detrital particles in Early Miocene diatom ooze, (d) detail of (Figure 7c) showing smooth to mammillated morphology of the FPS from which minute filament heads protrude (arrows, the filament heads were identified using stereomicrographs), (e) TEM section of the Early Miocene biofilm showing the filaments forming part of the FPS, and (f) detail of some of the Early Miocene filaments which contain some remnant organic matter in their cores (visualized by staining with lead citrate and uranyl acetate).
4. FPS and Biofilms Through Time
The examples that we use to illustrate FPS and bacterial cells throughout the rock record are drawn from our own investigations. The mineral matrices range from calcium carbonate (Yellowstone National Park, Wyoming), silica (Yellowstone National Park; South Atlantic deep-sea sediments; Gunflint Iron Formation, Ontario; and Barberton greenstone belt cherts, South Africa), phosphate (Enspel volcanic lake sediments,
Germany), and iron/silica in banded iron formations (F•t•kro greenstone belt, Ivory Coast). They range in age from modem to Recent deposits (Yellowstone), Late Oligocene-early Miocene (South Atlantic), Oligocene (Enspel), Early Proterozoic (Gunflint and F•t•kro), to Early Archaean (Barberton). The Phanerozoic samples were observed with scanning electron microscopes (a JEOL 840, and field emission gun (FEG) SEMs: a Philips XL 40 and a JEOL 6340) simply as broken fragments of sedimentary rock. The permineralized Precambrian samples were too
WESTALL ET AL.: EPS AS A FOSSIL BIOMARKER 24,521
a "'
ß 5 ß 0,'5 .pro
..
:..
...............
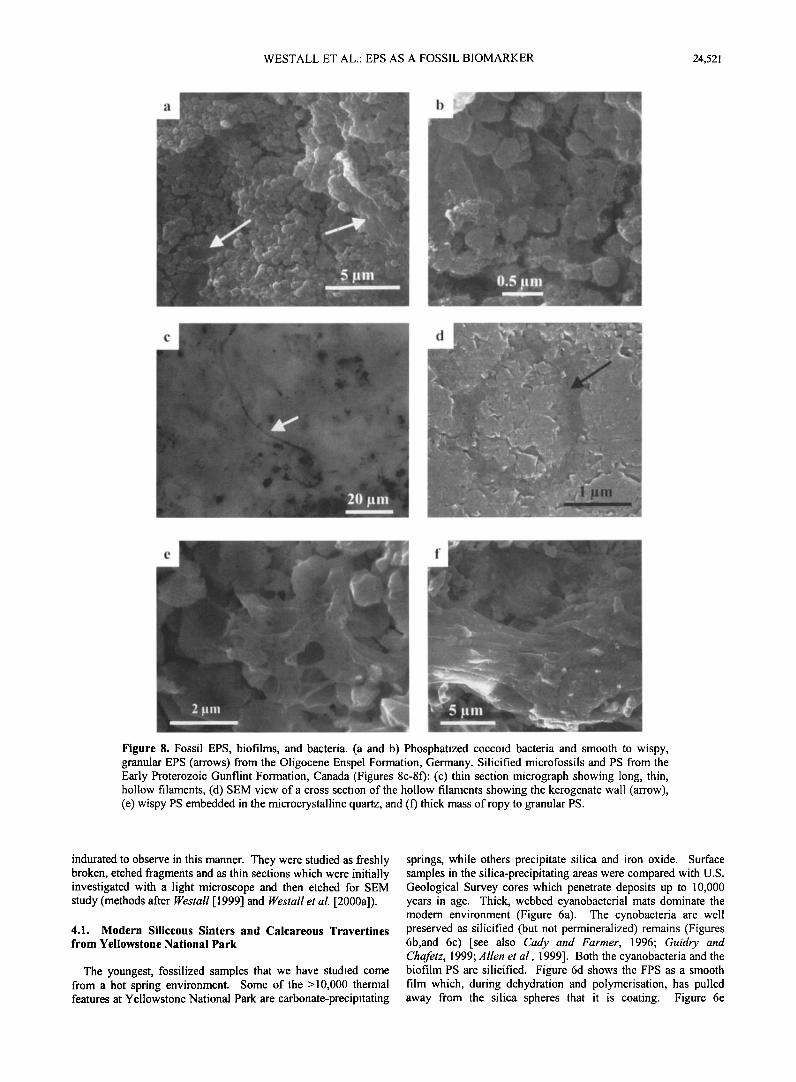
Figure 8. Fossil EPS, biofilms, and bacteria. (a and b) Phosphatized coccoid bacteria and smooth to wispy, granular EPS (arrows) from the Oligocene Enspel Formation, Germany. Silicified microfossils and PS from the Early Proterozoic Gunflint Formation, Canada (Figures 8c-8f): (c) thin section micrograph showing long, thin, hollow filaments, (d) SEM view of a cross section of the hollow filaments showing the kerogenate wall (arrow), (e) wispy PS embedded in the microcrystalline quartz, and (f) thick mass of ropy to granular PS.
indurated to observe in this manner. They were studied as fleshly broken, etched fragments and as thin sections which were initially investigated with a light microscope and then etched for SEM study (methods after Westall [1999] and Westall et al. [2000a]).
4.1. Modern Siliceous Sinters and Calcareous Travertines from Yellowstone National Park
The youngest, fossilized samples that we have studied come from a hot spring environment. Some of the >10,000 thermal features at Yellowstone National Park are carbonate-precipitating
springs, while others precipitate silica and iron oxide. Surface samples in the silica-precipitating areas were compared with U.S. Geological Survey cores which penetrate deposits up to 10,000 years in age. Thick, webbed cyanobacterial mats dominate the modem environment (Figure 6a). The cynobacteria are well preserved as silicified (but not permineralized) remains (Figures 6b,and 6c) [see also Cady and Farmer, 1996; Guidry and Chafetz, 1999; Allen et al., 1999]. Both the cyanobacteria and the biofilm PS are silicified. Figure 6d shows the FPS as a smooth film which, during dehydration and polymerisation, has pulled away from the silica spheres that it is coating. Figure 6e
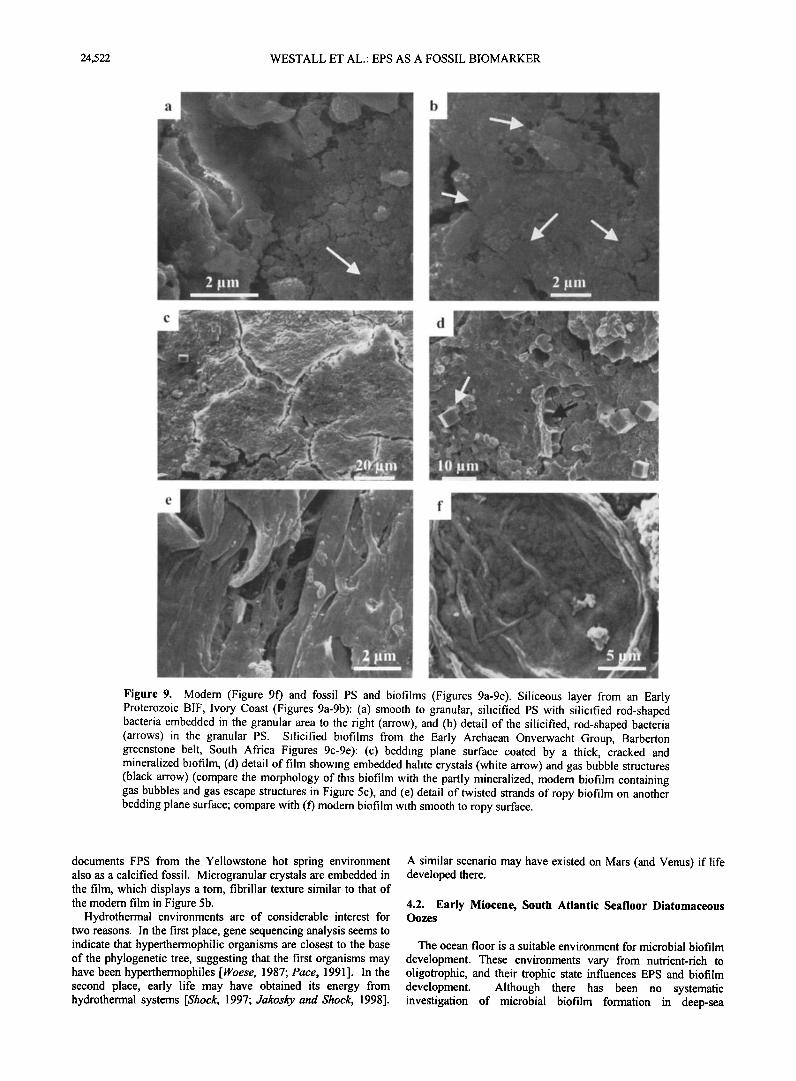
24,522 WESTALL ET AL.' EPS AS A FOSSIL BIOMARKER
Figure 9. Modem (Figure 9f) and fossil PS and biofilms (Figures 9a-9e). Siliceous layer from an Early Proterozoic BIF, Ivory Coast (Figures 9a-9b): (a) smooth to granular, silicified PS with silicified rod-shaped bacteria embedded in the granular area to the right (arrow), and (b) detail of the silicified, rod-shaped bacteria (arrows) in the granular PS. Silicified biofilms from the Early Archaean Onverwacht Group, Barberton greenstone belt, South Africa Figures 9c-9e): (c) bedding plane surface coated by a thick, cracked and mineralized biofilm, (d) detail of film showing embedded halite crystals (white arrow) and gas bubble structures (black arrow) (compare the morphology of this biofilm with the partly mineralized, modem biofilm containing gas bubbles and gas escape structures in Figure 5c), and (e) detail of twisted strands of ropy biofilm on another bedding plane surface; compare with (f) modem biofilm with smooth to ropy surface.
documents FPS from the Yellowstone hot spring environment also as a calcified fossil. Microgranular crystals are embedded in the film, which displays a tom, fibrillar texture similar to that of the modem film in Figure 5b.
Hydrothermal environments are of considerable interest for two reasons. In the first place, gene sequencing analysis seems to indicate that hyperthermophilic organisms are closest to the base of the phylogenetic tree, suggesting that the first organisms may have been hyperthermophiles [Woese, 1987; Pace, 1991]. In the second place, early life may have obtained its energy from hydrothermal systems [Shock, 1997; Jakosky and Shock, 1998].
A similar scenario may have existed on Mars (and Venus) if life developed there.
4.2. Early Miocene, South Atlantic Seafloor Diatomaceous Oozes
The ocean floor is a suitable environment for microbial biofilm development. These environments vary from nutrient-rich to oligotrophic, and their trophic state influences EPS and biofilm development. Although there has been no systematic investigation of microbial biofilm formation in deep-sea
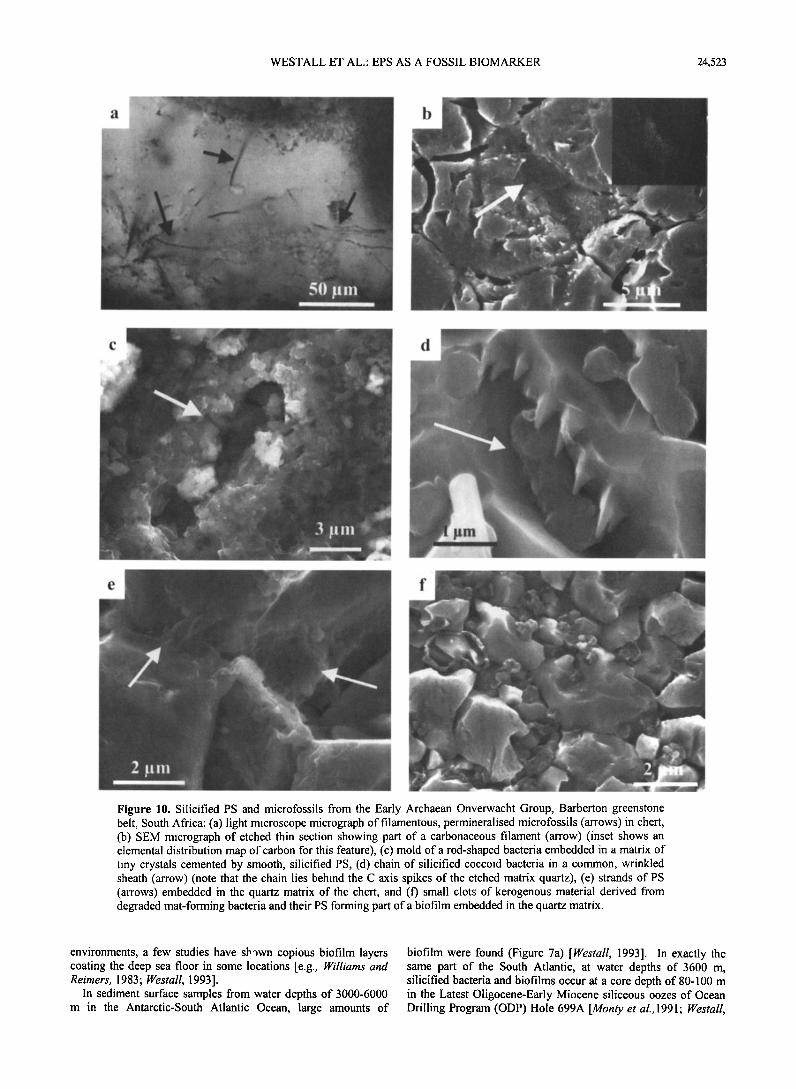
WESTALL ET AL.' EPS AS A FOSSIL BIOMARKER 24,523
5'0
Figure 10. Silicified PS and microfossils from the Early Archaean Onverwacht Group, Barberton greenstone belt, South Africa: (a) light microscope micrograph of filamentous, permineralised microfossils (arrows) in chert, (b) SEM micrograph of etched thin section showing part of a carbonaceous filament (arrow) (inset shows an elemental distribution map of carbon for this femure), (c) mold of a rod-shaped bacteria embedded in a matrix of tiny crystals cemented by smooth, silicified PS, (d) chain of silicified coccoid bacteria in a common, wrinkled shemh (arrow) (note that the chain lies behind the C axis spikes of the etched matrix quartz), (e) strands of PS (arrows) embedded in the quartz matrix of the chert, and (f) small clots of kerogenous mmerial derived from degraded mat-forming bacteria and their PS forming part of a biofilm embedded in the quartz matrix.
environments, a few studies have sb'•wn copious biofilm layers coming the deep sea floor in some locmions [e.g., Williams and Reimers, 1983; Westall, 1993].
In sediment surface samples from water depths of 3000-6000 m in the Antarctic-South Atlantic Ocean, large amounts of
biofilm were found (Figure 7a) [Westall, 1993]. In exactly tke same part of the South Atlantic, at water depths of 3600 m, silicified bacteria and biofilms occur at a core depth of 80-100 m in the Latest Oligocene-Early Miocene siliceous oozes of Ocean Drilling Program (ODP) Hole 699A [Monty et a1.,1991; Westall,

24,524 WESTALL ET AL.: EPS AS A FOSSIL BIOMARKER
1994]. The Latest Oligocene-Early Miocene biofilms are strongly similar in morphology to modem biofilms (Figure 7b) [Westall, 1994; Westall and Rinc•, 1994]. The silicified biofilms coat all particle surfaces and are characterized by a mainly globular or mammillated texture with some smooth areas (Figures 7c, and 7d). In addition, some of the fibrillar nature of the biofilm has also been retained (Figures 7d, 7e, and 7f). The degree of preservation is so good that some structural detail within the fibrils may be observed.
In terms of environmental significance, there are indications that a seafloor environment may have existed on early Mars. The recent orbital altimetry data from Mars (Mars Orbiter Laser Altimetry, MOLA) corroborate a previous hypothesis that there were water-filled basins in the Northern Plains [Parker et al., 1989, 1993; Head et al., 1998, 1999]. Head et al. [1999] estimate average water depths for the early ocean at -620 rn with a maximum of 2000 m.
4.3. Oligocene Volcanic Lacustrine Environments, Germany
The Enspel fossil site in central Germany is a volcanic, lacustrine environment. This location is well known for its
superb fossil preservation, including the "Hauptschatten" or skin shadows of the vertebrates. The 25 m.y. old Enspel site is similar in environmental setting and fossil preservation to the nearby, older (Eocene) Messel site [Wuttke, 1983; Liebig et al., 1996]. The preservation of the soft tissues of the organisms in the anaerobic volcanic muds was due to the fossilization
(phosphatization) of the microbial biofilms which coated the larger, dead and decaying organisms. FPS is not always present in these samples even though phosphatized bacteria are ubiquitous. This may be explained in the following manner: Phosphate is usually a limiting nutrient, and it's presence in excess to such an extent that it becomes a mineralizing medium indicates that the environment was probably nutrient rich, in which case, capsular EPS was probably not formed in the first place. Figures 8a and 8b show colonies of phosphatized, oval- shaped coccoid bacteria coating the preserved soft tissues of a fossil fish. Associated with the bacteria are wispy, granular strands of fossilized PS as well as smooth FPS coating cells and stretched between cells. Part of the smooth FPS film is puckered and tom, probably due to dehydration during the fossilization process, and resembles sirnilar features in modem PS (Figure 5b). In contrast to the soft tissues, the surrounding sediment is devoid of fossil bacteria and FPS.
Similar volcanic, lacustrine environmental settings existed on Mars (and Venus). Geomorphological data indicate that there were craters containing standing bodies of water [e.g., de Hon, 1992; Ori and Baliva, 1998], and such locations are being considered in the search for life on Mars.
4.4. Early Proterozoic Gunflint Formation, Canada
Stromatolites are perhaps the most characteristic biogenic structures of the Precambrian. Most of those identified to date
appear to be of a shallow, marine origin. Their gradual decrease in abundance throughout the Phanerozoic has been much discussed and is thought to be due to environmental changes (PCO2) as well as competition from and grazing by eukaryotes [Riding, 1994].
The 2.0 b.y. old Gunflint Iron Formation in Ontario, Canada, was the first formation in which Precambrian microfossils were
recognized [Barghoorn and Tyler, 1965] and is perhaps the most famous of the Precambrian fossiliferous sites. The Gunflint is an
iron-rich stromatolitic deposit which formed in shallow, silica- rich waters [Awramik, 1976]. The microfossils occur in the
cherty horizons. The permineralized filamentous and coccoid fossils are readily visible in thin section and with an SEM (Figures 8c and 8d). Silicified PS has the same ropy, smooth to granular appearance as in younger samples (e.g., Figures 8e and 80.
Environments similar to those in which the Precambrian
stromatolites thrived were likely to have existed on Mars, for example, around the edges of the water-filled basins of the Northern Plains or around the edges of any other body of standing water. In contrast to the terrestrial situation, if life developed on Mars, it is unlikely that eukaryotes would have evolved because of the early deterioration in the surface environment (loss of atmosphere, surface water and cooling,-4.0-3.8 b.y.) [Jakosky and Shock, 1998; Westall, 1999]; therefore organisms which could have constructed stromatolites would not have been
affected by competition or grazing by eukaryotes.
4.5. Kouroudia Banded Iron Formation, Ivory Coast
Given the recent finding of several large deposits of iron minerals on Mars [Christensen et al., 2000], a consideration of terrestrial iron deposits of biological relevance is pertinent. Banded iron formations (BIFs) have come under scrutiny in this respect because of the discussions about possible biological influences in their formation [e.g., Robbins et al., 1987; Krumbein and Stal, 1991 ].
The BIF located in the F•t•kro greenstone belt of the Ivory Coast is 2.1 b.y. old [Leake, 1992]. In contrast to the 2.0 b.y. old silicified Gunflint stomatolites, the Early Proterozoic BIF from the Ivory Coast appears to be less diagenetically altered. From within the cherty layers, Figure 9a shows remnant FPS with a partly ropy to smooth, and partly granular surface texture. Clearly embedded within the granular FPS film are fossil bacteria having bacillus and coccoid morphologies (Figure 9b). Both the PS and bacteria have been permineralized. The slight cracking of the film is probably due to dehydration during permineralization.
4.6. Early Archaean Barberton Greenstone Belt, South Africa
Rocks from the earliest period of Earth's history are rare, most deposits having been destroyed by plate tectonics or severely altered by metamorphism. The Barberton greenstone belt, South Africa, is one of the regions with the best preserved Early Archaean sequences. This series of volcanic strata, volcaniclastics, and siliciclastics has an age range of 3.2-3.5 b.y., close in age to the period when there was relatively abundant liquid water on the surface of Mars and when life could have developed on that planet [Cart, 1996]. A number of samples of carbonaceous cherts from the 3.3-3.5 b.y. old Onverwacht Group within the Swaziland Supergroup yield well-preserved biofilms, FPS, and microfossils [Walsh and Lowe, 1985, 1999; Walsh, 1992; Westall and Gemeke, 1998; Westall, 1999; Westall et al., 2000a]. The great age of these examples demonstrates the preservation potential of FPS and biofilms through time. All the samples are of shallow water origin, and some show clear evidence of subaerial exposure in an evaporitic environment.
Microbial biofilms and associated filamentous and smaller, non filamentous bacteria occur in finely laminated, carbonaceous cherts (Figures 9c-9f, and 10). SEM study of rock chips documented well-developed, permineralized biofilms on a few bedding plane surfaces (Figures 9c-9e). These bedding plane biofilms are highly complex structures which bear remarkable resemblance to modem biofilms (Figure 9f). The PS of the biofilms consists of thick, cracked crusts of twisted and interwoven strands which in some places has a granular texture

WESTALL ET AL.: EPS AS A FOSSIL BIOMARKER 24,525
and in others is smooth. Embedded within these biofilms are
small crystals of calcite, halite, possibly gypsum (or anhydrite?), and tourmaline. The latter are common in these strata because the
whole depositional environment was affected by hydrothermal activity. The association of the other minerals together with the cracked appearance of the biofilm suggests that these particular films were subaerially exposed in an evaporitic setting. One of the biofilms contains bubble-like structures consisting of subrounded hollows surrounded by raised rims (Figure 9d). These structures are similar to gas bubbles within modem biofilms (Figure 5c): gas production by the bacteria inhabiting a biofilm is a common phenomenon which creates bubbles within films as well as volcano-like gas escape structures. Isolated FPS strands, globs of FPS and more granular, wispy FPS are common in the carbonaceous cherts (Figures 10e and 1 Of).
$. Conclusions
Bacteria produce exopolymeric substances (EPS) and colonies of bacteria secrete EPS which, on an interface, forms slime layers called biofilms. Biofilms are compositionally complex, containing a variety of microbes and their degradation products as well as allochthonous and autochthonous particles. They are also structurally complex. This complexity is a characteristic of microbial biofilms, which would not be found in potential prebiotic organic films. Furthermore, because of their high metal-binding capacity, biofilms are readily fossilizable. We have documented fossil biofilms ranging in age from the Recent to the Early Archaean (3.5 b.y.) occurring in deposits whose environmental settings correspond well with environments which would have existed on Mars and early Venus (hot springs, lacustrine volcanic, littoral, shallow marine, and deeper marine). The biofilms were preserved by permineralization in silica, phosphate, and calcite as well as by mineral replacement (silica).
Biofilms in their complexity therefore constitute reliable and robust fossil evidence of life. The discovery of complex biofilms in any state of preservation on pristine, uncontaminated extraterrestrial material would constitute very strong evidence for life in that material and its parent body. However, it is recognized that additional criteria, such as in situ biochemical or isotope analyses, would be useful to unequivocally identify biofilms in both returned samples and remotely on a planetary robotic mission.
6. Future Work
Although the morphological characteristics of, especially, fossil biofilms constitute strong evidence for biogenicity, further studies to promote understanding of biofilms would contribute to their reliability as a robust biomarker: (1) Comparison between biofilms and experimentally produced films of prebiotic molecules in order to obtain a "base" control. Such a study is underway by F. Westall and D. Deamer (Santa Cruz). (2) Most of the FPS and fossil biofilms contain remnant organic molecules which could retain traces of their biological ancestry. In fact, preliminary studies by A. Steele et al. (see Westall et al., 2000b) have tentatively identified a specific biomolecule or fragment thereof in all the examples we have cited here (except for the carbon-less Yellowstone hot spring sample).
Acknowledgments. F.W. acknowledges funding from the National Research Council, J. H6gbom, and M.J. de Wit; J.T. acknowledges funding by the DAAD. A.S. would like to acknowledge Recep Avci for ToFSIMS analysis. M. de Wit and J. Dann are thanked for supplying some of the samples. We are very grateful to Gordon Southam and an unnamed reviewer for their helpful critiques. F.W. thanks D. Deamer for fruitful discussion concerning the prebiotic world.
References
Alexander, J., Non biodegradable and other recalcitrant molecules, Biotechnol. Bioeng. 15, 611-647, 1973.
Allen, C.C., F.G. Albert, H.S. Chafetz, J. Crombie, C.R. Graham, T.L. Kieft, S.J. Kivett, D.S. McKay, A. Steele, A.E. Taunton, M.R. Taylor, K.L. Thomas-Keprta, and F. Westall, Physical biomarkers in carbonate thermal springs: Implications for Mars, Icarus, in press, 2000.
Allison, D.G., Exopolysaccharide synthesis in bacterial biofilms, in The Life and Death of Biofilm, edited by J. Wimpenny et al., pp. 25-30, Bioline, Cardiff, Wales, 1993.
Awramik, S.M., Gunflint stromatolites: Microfossil distribution in relation to stromatolite morphology, in Stromatolites, Dev. in Sedimentol., vol. 20, edited by M.R. Walter, pp. 311-320, Elsevier Sci., New York, 1976.
Barghoom, E.S., and S.A. Tyler, Micro-organisms from the Gunflint Chert, Science, 147, 563-577, 1965.
Bemier, P., C. Gaillard, J.C. Gall, G. Barale, J.P. Bourseau, E. Buffetaut, and S. Wenz, Morphogenetic impact of microbial mats on surface structures of Kimmeridgian micritic limestones (Cerin, France), Sedimentology, 38, 127-136, 1991.
Beveridge, T.J., Metal ions and bacteria, in Metal Ions and Bacteria, edited by T.J. Beveridge and D.J. Doyle, pp. 1-30, John Wiley, New York, 1989.
Brierley, C.L., D.P. Kelly, K.J. Seal, and D.J. Best, Materials and biotechnology, in Biotechnology: Principles and Applications, edited by I.J. Higgins, D.J. Best, and J. Jopnes, pp. 163, Blackwell, Malden, Mass., 1985.
Brierley, C.L., J.A. Briefly, and M.S. Davidson, Applied microbial processes for metals recovery and removal from wastewater, in Metal Ions and Bacteria, edited by T.J. Beveridge and D.J. Doyle, pp. 359- 383, John Wiley, New York, 1989.
Brocks, J.J., G.A. Logan, R. Buick, and R.E. Summons, Archaean molecular fossils and the early rise of eukaryotes, Science, 285, 1033- 1036, 1999.
Cady, S.L., and J.D. Farmer, Fossilization processes in siliceous thermal springs: trends in preservation along the thermal gradient, in Evolution of Hydrothermal Ecosystems on Earth (and Mars D, CIBA Found. Syrup., 202 vol., edited by G.R. Bock and J.A. Goode, pp. 150-169, John Wiley, New York, 1996.
Carr, M.H., Water on Mars, 229 pp., Oxford Univ. Press, New York, 1996.
Castenholz, R.W., Microbial mat research: The recent, past and new perspectives, in Microbial Mats: Structure, Development and Environmental Significance, NATO ASI Ser. G. vol 35, edited by L.J. Stal and P. Caumette, pp. 3-18, Springer-verlag, New York, 1994.
Chafetz, H.S., and C. Buczynski, Bacterially induced lithification of microbial mats, Palaios, 7, 277-293, 1992.
Charaklis, W.G., and K.E. Cooksey, Biofilms and microbial fouling, Adv. Appl. Microbiol., 29, 93-138, 1990.
Charaklis, W.G., and K.C. Marshall, Biofilms: A basis for interdisciplinary research, in Biofilms, edited by W.G. Charaklis and K.C. Marshall, pp. 3-16, John Wiley, New York, 1990.
Charaklis, W.G., G.A. McFetters, and K.C. Marshall, Physiological ecology in biofilm systems, in Biofilms, edited by W.G. Charaklis and K.C. Marshall, pp. 341-394, 1990.
Christensen, P.R., et al., Detection of crystalline hematite mineralization on Mars by the thermal emission spectrometer: Evidence for near- surface water, d. Geophys. Res.,105, 9623-9642, 2000.
Cohen, Y., and E. Rosenberg (Eds.), Microbial Mats, Am. Soc. for Microbiol., Washington, D.C., 494 pp., 1989.
Collins, Y.E., and G. Stozky, Factors affecting the toxicity of heavy metals to microbes, in Metal Ions and Bacteria, edited by T.J. Beveridge and D.J. Doyle, pp. 31-90, John Wiley, New York, 1989.
Costerton, J.W., Transport through porous media, in The Life and Death of Biofilm, edited by J. Wimpenny, P. Handley, P. Gilbert, and H. Lappin-Scott, pp. 127-128, Bioline, Cardiff, Wales, 1995.
Costerton, J.W.J.C. Nickel, and T.I. Ladd, How bacteria stick, $ci. Am., 238 (1), 86-95, 1978.
Costerton, J., Z. Lewandowski, D.E. Caldwell, D.R. Korber, and H.M. Lappin-Scott, Microbial biofilms, Annu. Rev. Microbiol., 49, 711-745, 1995.
Deamer, D.W., The first living systems: A bioenergetic perspective, Microbiol. Mol. Biol. Rev., 61,239-261, 1997.
DeBeer, D, P. Stoodley, F.L. Roe, and Z. Lewandowski, Effects of

24,526 WESTALL ET AL.: EPS AS A FOSSIL BIOMARKER
biofilm structures on oxygen distribution and mass transport, Biotech. Bioeng., 43, 1131-1138, 1994.
Decho, A.W., Molecular-scale events influencing the macro-scale cohesiveness of exopolymers, in Biostabilization of Sediments, edited by W.E. Krumbein, D.M. Paterson, and L.J. Stal, pp. 135-149, BIS, Oldenburg, Germany, 1994.
D6farge, C., J. Trichet, A. Maurin, and M. Hucher, Kopara in Polynesian atolls, early formation of calcareous stromatolites, Sediment. Geol., 89, 9-23, 1994.
de Hon, R.A., Martian lake basins and lacustrine plains, Earth Moon Planets, 56, 95-122, 1992.
de Leeuw, J.W., and C. Largeau, A review of macromolecular organic compounds that compare living organisms and their role in kerogen, coal and petroleum formation, in Organic Geochemistry.' Principles and Applications, edited by M. Engel and S.A. Macko, pp. 56-57, Plenum Press, NewYork, 1993.
Dowling, N.J.E., M.W. Mittlman, and D.C. White, The role of consortia in microbially influenced corrosion, in Mixed Cultures in Biotechnology, edited by J.G. Zeikus, pp. 341-372, McGraw-Hill, New York, 1991.
Fera, R., M.A. Siebel, W.G. Charaklis, and D. Prieur, Seasonal variations in bacterial colonization of stainless steel, aluminium and polycarbonate surfaces in a seawater flow system, Biofouling, 1,251- 261, 1989.
Ferris, F.G., W.S. Fyfe, and T.J. Beveridge, Metallic ion binding by Bacillus subtills: Implications for the fossilization of microorganisms, Geology, 16, 149-152, 1988.
Ferris, F.G., S. Schultz, T.C. Witten, W.S. Fyfe, and T.J. Beveridge, Metal interactions with microbial biofilms in acidic and neutral pH environments, Appl. Environ. Microbiol., 55, 1249-1257, 1989.
Fleming, H.C., and G.G. Geesey (Eds.), Biofouling and Biocorrosion in Industrial Water Systems, 162 pp., Springer-Verlag, New York, 1991.
Fletcher, M., The measurement of bacterial attachment to surfaces in static systems, in Biofilms-Science and Technology, edited by L.F. Melo, pp. 105-110, Kluwer Acad., Norwell, Mass., 1992.
Fletcher, M., and K.C. Marshall, Bubble contact angle method for evaluating substratum interfacial characteristics and its relevance to bacterial attachment, Appl. Environ. Microbiol, 44, 184-192, 1982.
Fox, S.W., R.M. Syren, M. Ingram, B.J. Price, and J. Costello, Ancient microspheres: Abiogenic, protobiogenic, or biogenic?, Precambrian Res., 23, 1-8, 1983.
Francis, S., L. Margulis, and E.S. Barghoom, On the experimental silicification of microorganisms, II, On the time of appearance of eukaryotic organisms in the fossil record, Precambrian Res.,6, 65-100, 1978a.
Francis, S., E.S. Barghoom, and L. Margulis, On the experimental silicification of microorganisms, III, Implications of the preservation of the green prokaryotic alga Prochloron and other coccoids for the interpretation of the fossil record, Precambrian Res, 7, 377-383, 1978b.
Geddie, J.L., and I.W. Sutherland, Uptake of metals and polysaccharides, d. Appl. Bacteriol., 74, 467-472, 1993.
Geesey, G.G., Microbial exopolymers: Ecological and economic considerations, ASM News, 48, 9-14, 1982.
Geesey, G.G., and L. Jang, Interactions between metal ions and capsular polymers, in Metal Ions and Bacteria, edited by T.J. Beveridge and D.J. Doyle, pp. 325-358, John Wiley, New York, 1989a.
Geesey, G.G., and L. Jang, Extracellular polymers for metal binding, in Metal Ions and Bacteria, edited by T.J. Beveridge and D.J. Doyle, pp. 223-247, John Wiley, New York, 1989b.
Glover, J.E., Sediments of the Early Archaear, coastal plains: A possible environment for the origin of life, Precambrian Res., 56, 159-166, 1992.
Guidry, S.A., and H.S. Chafetz, Preservation of microbes in geyserite and siliceous sinter: Yellowstone National Park, Wyoming, Lunar Planet. $ci. [CD-ROM], XXX, Abstract 1152, 1999.
Guidry, S.A., H.S. Chafetz, A. Steele, and J.K.W. Toporski, A preliminary ToF-SIMS assessment of preservation potential of organic biomarkers in modem siliceous sinter and core, Yellowstone National Park, Wyoming. Lunar Planet. Sci. [CD-ROM], XXXI, Abstract 1100, 2000.
Haack, T.K., and G.A. McFeters, Nutritional relationships among microorganisms in an epilithic biofilm community, Microbiol. Ecol., 8, 115-126, 1982.
Head, J.W., D. Smith, M. Zuber, and the MOLA Team, Mars: Assessing
evidence for an ancient Northern Ocean with MOLA data, Meteorit. Planet. $ci., 33, A66, 1998.
Head, J.W., M. Kreslavsky, H. Hiesinger, and S. Pratt, Northern seas and oceans in the past history of Mars: new evidence from Mars Orbiter Laser Altimeter (MOLA) data, Lunar Planet. Sci. [CD-ROM], XXX,, Abstract 1352, 1999.
Hedges, J.I.. and F.G. Prahl, Early diagenesis: Consequences for application of molecular biomarkers, in Organic Geochemistry, edited by M. Engel and S.A. Macko, pp. 237-254, Plenum, New York, 1993.
Jakosky, B.M., The Search for Life on Other Planets, 326 pp., Cambridge Univ. Press., New York, 1998.
Jakosky, B.M., and E.L. Shock, The biological potential of Mars, the early Earth, and Europa, o r. Geophys. Res., 103, 19,359-19,364, 1998.
Keevil, C.W., and J.T. Walker, Normarski DIC microscopy and image analysis, Binary, 4, 93-95, 1992.
Knoll, A.H., Exceptional preservation of photsynthetic organisms in silicified carbonates and silicified peats, Philos. Trans. R. $oc. London $er. B, 311, 111-122, 1985.
Krumbein, W.E., Stromatolites-the challenge of a term in space and time, Precambrian Res., 20, 493-531, 1983.
Krumbein, W.E. and L.-J. Stal, The geophysiology of marine cyanobacterial mats and biofilms, Kiel. Meeresforsch. Sonderh., 8, 137-145, 1991.
Krumbein, W.E., D.M. Paterson, and L.J. Stal (Eds.), Biostabilization of Sediments, 526 pp, BIS, Oldenburg, Germany, 1994.
Krumbein, W.E., D.M. Paterson, and L.J. Lucas (Eds.), Biostabilization of sediments, 526 pp., BIS, Oldenburg, 1994.
Lawless, J.G., and C.D. Boyneton, Thermal synthesis of amino acids from a simulated primitive atmosphere, Nature, 243,405-407, 1973.
Leake, M.H., The petrogenesis and structural evolution of the Early Proterozoic F•t•kro greenstone belt, Dabakala Region, NE C6te d'Ivoire, Ph.D. thesis, Univ. of Portsmouth, Portsmouth, England, 1992.
Lewandowski, Z., P. Stoodley, and S. Altobelli, Experimental and conceptual studies on mass transport in biofilms, Water $ci. Technol., 31, 153-162, 1995.
Liebig, K., F. Westall, and M. Schmitz, A study of fossil microstructures from the Eocene Messel Formation using transmision electron microscopy, Neues. orahrb. Geol. Palaeontol. Monatsh., 4, 218-231, 1996.
Margulis, L., J.C.G. Walker, and M. Rambler, Reassessment of roles of oxygen and ultraviolet light in Precambrian evolution, Nature, 264, 620-624, 1976.
Margulis, L., E.S. Barghoom, D. Ashendorf, S. Banerjee, D. Chase, S. Francis, S. Giovannoni, and J. Stolz, The microbial community in the layered sediments at Laguna Figueroa, Baja California, Mexico: Does it have Precambrian analogues?, Precambrian Res., 11, 93-12, 1980.
McKay, D.S., E.K. Gibson, K.L. Thomas-Keprta, H. Vali, C.S. Romanek, S.J. Clemett, X.D.F. Chillier, S.R. Maedling, and R.N. Zare, Search for past life on Mars: Possible relic biogenic activity in Martian meteorite ALH84001, Science, 273,487-489, 1996.
Monty, C.L.V., Phanerozoic $tromatolites, Case Histories, 249 pp., Springer-Verlag, New York, 1981.
Monty, C.L.V., F. Westall, and S. van der Gaast, Diagenesis of siliceous particles in sub-Antarctic sediments, ODP Leg 114, Hole 699A: Possible microbial mediation, Proc. Ocean Drill. Program Sci. Results, 114, pp. 685-710, 1991.
Neu, T.R., Biofilms and microbial mats, in Biostabilization of Sediments, edited by 'vvr. E. Krumbein, D.M. Paterson, and L.J. Stal, pp. 9-16, BIS, Oldenburg, Germany, 1994.
Oehler, J.H., Experimental studies in Precambrian paleontology: Structural and chemical changes in blue-green algae during simulated fossilization in synthetic chert, Bull. Geol. $oc. Am., 87, 117-129, 1976.
Oehler, J.H. and J.W. Schopf, Artificial microfossils: Experimental studies of permineralization of blue-green algae in silica, Science, 174, 1229-1231, 1971.
Ori, G.G. and A. Baliva, Lacustrine and fluvial sedimentary environments in Ismenius Lacus and Memnonia, Mars, Lunar Planet. $ci. [CD- ROM], XXIX, Abstract 1601, 1998.
Pace, N.R., Origin of life- Facing up to the physical setting, Cell, 65, 531-533, 1991.
Parker, T.J., R.S. Saunders, and D.M. Schneeberger, Transitional morphology in the west Deuteronilus Mensae region of Mars: Implications for modification of the lowland/upland boundary, Icarus, 82, 111-145, 1989.

WESTALL ET AL.: EPS AS A FOSSIL BIOMARKER 24,527
Parker, T.J., D.S. Gorsline, R.S. Saunders, D.C. Pieri, and D.M. Schneeberger, Coastal geomorphology of the Martian Northern Plains, J. Geophys. Res., 98, 11,061-11,078, 1993.
Pierson, B.K., H.K. Mitchell, and A.L. Ruff-Roberts, Chlorofiexus aurantiacus and ultraviolet radiation: Implications for Archean shallow water stromatolites, Origin Life Evol. Biosphere, 23, 243-260, 1993.
Platt, R.M., G.G. Geesey, J.D. Davies, and D.C. White, Isolation and partial chemical analysis of firmly bound exoploysaccharide from adherent cells of freshwater sediment bacteria, Can. J. Microbiol., 31, 675-680, 1985.
Pope, D.H., D.J. Duquette, A.H. Johannes, and P.C. Wayner, MIC of industrial alloys, in Corrosion 83, p. 247, Natl. Assoc. Corrosion Engineers, Houston, Tex., 1983.
Riding, R., Stromatolite survival and change: The significance of Shark Bay and Lee Stocking Island subtidal columns, in Biostabilization of Sediments, edited by W.E. Krumbein, D.M. Paterson, and J.L. Lucas, pp. 183-202, BIS, Oldenburg, Germany, 1994.
Robbins, E.I., G.L. La Berge, and R.G. Schmidt, A model for the biological precipitation of Precambrian formations. B: Morphological evidence and modem analogs, in Precambrian Iron-Formations, edited by P.W.U. Appel and G.L. La Berge, pp. 97-139, Theophrastes, Athens, 1987.
Schopf, J.W., Microfossils of the Early Archaean Apex Chert: New evidence of the antiquity of life, Science, 260, 640-646, 1993.
Schopf, J.W., and B.M. Packer, Early Archaean (3.3-billion to 3.5-billion- year-old) microfossils from the Warrawoona Group, Australia, Science, 237, 70-73, 1987.
Shock, E.L., High-temperature life without photosynthesis as a model for Mars, J. Geophys. Res., 102, 23,687-23,694, 1997.
Stal, L.J. and P. Caumette (Eds.), Microbial Mats, NATO ASI $er. G., vol. 35, 463 pp., Springer-Verlag, New York, 1994.
Steele, A., D.T. Goddard and I.B. Beech, An atomic force microscopy study of the biodeterioration of stainless steel in the presence of bacterial biofilms, Int. Biodeterioration Biodegradation, 33(5), 35-46, 1994.
Steele, A., D.T. Goddard, and I.B. Beech, A quantitative study of the corrosion damage of stainless steel in the presence of bacterial biofilms using Atomic Force Microscopy, in Corrosion 95, paper 73, Nat. Assoc. of Corrosion Eng. Houston, Tex., 1995.
Summons, R.E., and M.R. Walter, Molecular fossils and microfossils of prokaryotes and protists from Proterozoic sediments, Am. J. $ci., 290- A, 212-244, 1990.
Summons, R.E., L.L. Jahnke, and B.R.T. Simoneit, Lipid biomarkers for bacterial ecosystems: Studies of cultured organisms, hydrothermal environments and ancient sediments, in Evolution of Hydrothermal Ecosystems on Earth (and Mars?), CIBA Found. Symp., 202 vol., edited by G.R. Bock and J.A. Goode, pp. 174-192, John Wiley, New York, 1996.
Summons, R.E., L.L. Jahnke, J.M. Hope, and G.A. Logan, 2- Methylehopanoids as biomarkers for cyanobacterial oxygenic photosynthesis, Nature, 400, 554-557, 1999.
Turley, C.M., and K. Lochte, Microbial response to the input of fresh detritus to the deep-sea bed, Palaeogeogr. Palaeoclimatol. Palaeoecol., 89, 3-23, 1990.
Wachend6rfer, V., W.E. Krumbein, and H.J. Schellnhuber, Bacteriogenic porosity of marine sediments-a case for biomorphogenesis of sedimentary rocks, in Biostabilization of Sediments, edited by W.E. Krumbein, D.M. Paterson, and L.J. Stal, pp. 203-220, BIS, Oldenburg, Germany, 1994.
Walsh, M.M., Microfossils and possible microfossils from the Early Archaean Onverwacht Group, Barberton Mountain Land, South Africa, Precambrian Res.,$4, 271-293, 1992.
Walsh, M.M., and D.R. Lowe, Filamentous microfossils from the 3,500- Myr-old Onverwacht Group, Barberton Mountain Land, South Africa, Nature, 314, 530-532, 1985.
Walsh, M.M., and D.R. Lowe, Modes of accumulation of carbonaceous matter in the Early Archaean: A petrographic and geochemical study of the carbonaceous cherts of the Swaziland Supergroup, in Geologic Evolution of the Barberton Greenstone Belt, South Africa, edited by
D.R. Lowe and G.R. Byerly, Spec. Pap. Geol. $oc. Am., 329, 115-132, 1999.
Walter, M.R. (Ed.), $tromatolites, Dev. $edimentol., vol. 20, 790 pp., Elsevier Sci., New York, 1976.
Walters, C.C., L. Margulis, and E.S. Barghoom, On the experimental silicification of microorganisms. I. Microbial growth on organosilicon compounds, Precambrian Res., 5, 241-248, 1977.
Welch, S.A., W.W. Barker, and J.F. Banfield, Microbial extracellular polysaccharides and plagioclase dissolution, Geochim. Cosmochim. Acta, 63, 1405-1419, 1999.
Westall, F., An experimental investigation into the influence of bacteria on the dissolution and precipitation of silica (early diagensis) and the fossilization of micro-organisms in deep-sea siliceous sediments under low temperature/high pressure conditions, Report to the Commission of European Communities, November 1993, 80 pp, Comm. of Eur. Commun., 1993.
Westall, F., Silicified bacteria and associated biofilm from the deep-sea sedimentary environment, Kaupia Darmstd. Beitr. Naturwissensch., 4, 29-43, 1994.
Westall, F., The nature of fossil bacteria: A guide to the search for extraterrestrial life, J. Geophys. Res., 104, 16,437-16,451, 1999.
Westall, F., and D. Gemeke, Electron microscope methods in the search for the earliest life forms on earth (in 3.5-3.3. Ga cherts from the Barberton greenstone belt, South Africa): Applications for extraterrestrial life studies, Proc. $PIE Int. $oc. Opt. Eng., 3441, 158- 169, 1998.
Westall, F., and Y. Rinc6, Bacteria-particle interactions in a littoral diatomaceous sediment, Sedimentology, 44, 147-162, 1994.
Westall, F., L. Boni, M.E. Guerzoni, The experimental silicification of microorganisms, Palaeontology, 38, 495-528, 1995.
Westall, F., M.J. de Wit, J. Dann, S. van der Gaast, C.E.J. de Ronde, and D. Gemeke, Early Archaean fossil bacteria and biofilms in hydrothermally-influenced, shallow water sediments, Barberton greenstone belt, South Africa, Precambrian Res., in press, 2000a.
Westall, F. A. Steele, J. Toporski, M.M. Walsh, C.C. Allen, S. Guidry, D.S. McKay, E.K. Gibson, and H. Chafetz, A 3.8 b.y. history of bacterial biofilms and their significance in the search for extraterrestrial life, Lunar Planet. $ci. [CD-ROM,] X_XXI, Abstract 1707, 2000b.
Williams, L.A., and C. Reimers, Role of bacterial mats in oxygen deficient marine basins and coastal upwelling regimes: Preliminary report, Geology, I 1,267-269, 1983.
Wirepenny, J.W.T., The spatial organisation of biofilms, in The Life and Death of Biofilm, edited by J. Wirepenny et al., pp. 1-6, Bioline, Cardiff, Wales, 1995.
Woese, C.R., Bacterial evolution, Microbiol. Rev., 51, 221-271, 1987. Wuttke, M., "Weichteilerhaltung" durch lithifizierte Mikroorganismen bei
mittel-eozlinen Vertebraten aus den Olschiefem der "Grube Messel" bei Darmstadt, $enckenberg. Lethaiea, 65, 509-527, 1983.
Yanagawa, H., and K. Kojima, Thermophilic microspheres of peptide-like polymers and silicates formed at 250 degrees C, J. Biochem., 97, 1521- 1524, 1985.
C. Allen, Lockheed Martin Space Operations, Houston, Texas, 77058. H. Chafetz and S. Guidry, Department of Geosciences, University of
Houston, Texas 77204.
E. Gibson, D. McKay, and S. Steele, NASA Johnson Space Center, NASA Road 1, Houston, Texas 77058.
J. Toporski, School of Earth and Environmental Sciences, Astrobiology Group, University of Portsmouth, Portsmouth P01 3QL, England, United Kingdom.
M. Walsh, Institute for Environmental Sciences, Louisiana State University, Baton Rouge, LA 70803.
F. Westall, Lunar and Planetary Institute, 3600 Bay Area Boulevard, Houston, Texas 77058. (Westall•lpi.usra.edu)
(Received February 16, 2000; revised June 5, 2000; accepted June 28, 2000)




















