
Parallel evolution and phenotypic divergence in lichenized fungi: A case study in the lichen-forming...
19
Parallel evolution and phenotypic divergence in lichenized fungi: A case study in the lichen-forming fungal family Graphidaceae (Ascomycota: Lecanoromycetes: Ostropales) Eimy Rivas Plata a,b,⇑ , H. Thorsten Lumbsch b a Biological Sciences Department, University of Illinois at Chicago, 845 W. Taylor St., Chicago, IL 60607, USA b Department of Botany, The Field Museum, 1400 S. Lake Shore Drive, Chicago, IL 60605, USA article info Article history: Received 12 October 2010 Revised 14 March 2011 Accepted 30 April 2011 Available online 14 May 2011 Keywords: Ancestral character state reconstruction Character evolution Convergent evolution Homoplasy Thelotremataceae abstract A molecular phylogeny of combined mtSSU, nuLSU, and RPB2 data revealed previously unrecognized lev- els of parallel evolution and phenotypic divergence in the lichen family Graphidaceae. Five clades were supported within the family: the Fissurina, Ocellularia, Graphis, Topeliopsis, and Thelotrema clades, contain- ing 33 of the 42 currently accepted genera within the family. The results for the first time provide a fully resolved phylogeny of this family and confirm the synonymy of Graphidaceae and Thelotremataceae. Ancestral character state reconstruction using likelihood, Bayesian, and parsimony approaches indicate that lirellate ascomata evolved independently in each of the five clades. Carbonized ascomata evolved independently in at least four of the five clades. An unexpected result was the independent evolution of columella structures in the Fissurina and Ocellularia clades. Besides these more general findings, we document several cases in which evolution of several traits in parallel resulted in striking look-alikes within unrelated lineages, such as Topeliopsis muscigena and Chapsa meridensis in the Topeliopsis and Thelotrema clades, Leptotrema wightii, Myriotrema laeviusculum, and Leucodecton phaeosporum in the Ocel- lularia and Thelotrema clades, Ocellularia stylothecia and Melanotrema meiosporum in the Fissurina and Ocellularia clades, and Myriotrema pycnoporellum, Myriotrema clandestinum and Wirthiotrema glaucopal- lens in the Fissurina, Ocellularia, and Topeliopsis clades. Pagel’s test of independent character evolution suggested that at least for some of the traits involved in these cases, ecological constraints may have caused their evolution in parallel. The most intriguing find is the correlation between gall-forming thalli and vertical columns of calcium oxalate crystals, suggesting that these crystals do not function as light distributors, as previously assumed, but instead stabilize the thalli which are usually hollow beneath, similar to a dome-shaped structure. Ancestral character state reconstruction together with an approach to visualize the phenotype of putative ancestral lineages suggested the alpha-Graphidaceae to resemble some of the extant species currently classified in Myriotrema s.lat., with pore-like ascomata, and non- amyloid ascospores with lens-shaped lumina. Ó 2011 Elsevier Inc. All rights reserved. 1. Introduction One of the most intriguing evolutionary phenomena is the inde- pendent evolution of similar or identical traits from a distant ancestor that did not have the traits in question. This phenomenon is variously termed ‘parallel’ or ‘convergent’ evolution, or ‘homo- plasy’, but these terms address very specific and conceptually dif- ferent evolutionary phenomena (Moore and Willmer, 1997; Zhang and Kumar, 1997; Olson and Hall, 2003; Futuyma, 2005; Stearns and Hoekstra, 2005; Barton et al., 2007). Convergent evolution de- notes the independent evolution of functionally similar traits based on different structural elements in unrelated or distantly re- lated lineages. These traits involve different structural elements and their underlying body plan shows anatomical and ontogenetic differences; such traits are called ‘analogous’. A classical example for convergent evolution are the wings of bats and pterodactyls: being phylogenetically unrelated, they represent a similar solution to biological ‘engineering’, but use different parts of the vertebrate forelimb (Stearns and Hoekstra, 2005). The opposite of convergent would be divergent evolution, in which related lineages acquire very different traits due to radiation, but based on homologous body parts. Divergent evolution is common in organisms with an open body plan, such as plants and fungi, in which closely related lineages often exhibit phenotypic divergence (Blanco et al., 2004a,b; Tehler and Irestedt, 2007; Mugambi and Huhndorf, 2009; Lumbsch et al., 2010; Parnmen et al., 2010). A prominent 1055-7903/$ - see front matter Ó 2011 Elsevier Inc. All rights reserved. doi:10.1016/j.ympev.2011.04.025 ⇑ Corresponding author at: Department of Botany, The Field Museum, 1400 S. Lake Shore Drive, Chicago, IL 60605, USA. E-mail address: erivasplata@fieldmuseum.org (E. Rivas Plata). Molecular Phylogenetics and Evolution 61 (2011) 45–63 Contents lists available at ScienceDirect Molecular Phylogenetics and Evolution journal homepage: www.elsevier.com/locate/ympev
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Parallel evolution and phenotypic divergence in lichenized fungi: A case study in the lichen-forming...

Parallel evolution and phenotypic divergence in lichenized fungi: A case studyin the lichen-forming fungal family Graphidaceae (Ascomycota: Lecanoromycetes:Ostropales)
Eimy Rivas Plata a,b,⇑, H. Thorsten Lumbsch b
aBiological Sciences Department, University of Illinois at Chicago, 845 W. Taylor St., Chicago, IL 60607, USAbDepartment of Botany, The Field Museum, 1400 S. Lake Shore Drive, Chicago, IL 60605, USA
a r t i c l e i n f o
Article history:Received 12 October 2010Revised 14 March 2011Accepted 30 April 2011Available online 14 May 2011
Keywords:Ancestral character state reconstructionCharacter evolutionConvergent evolutionHomoplasyThelotremataceae
a b s t r a c t
A molecular phylogeny of combined mtSSU, nuLSU, and RPB2 data revealed previously unrecognized lev-els of parallel evolution and phenotypic divergence in the lichen family Graphidaceae. Five clades weresupported within the family: the Fissurina, Ocellularia, Graphis, Topeliopsis, and Thelotrema clades, contain-ing 33 of the 42 currently accepted genera within the family. The results for the first time provide a fullyresolved phylogeny of this family and confirm the synonymy of Graphidaceae and Thelotremataceae.Ancestral character state reconstruction using likelihood, Bayesian, and parsimony approaches indicatethat lirellate ascomata evolved independently in each of the five clades. Carbonized ascomata evolvedindependently in at least four of the five clades. An unexpected result was the independent evolutionof columella structures in the Fissurina and Ocellularia clades. Besides these more general findings, wedocument several cases in which evolution of several traits in parallel resulted in striking look-alikeswithin unrelated lineages, such as Topeliopsis muscigena and Chapsa meridensis in the Topeliopsis andThelotrema clades, Leptotrema wightii, Myriotrema laeviusculum, and Leucodecton phaeosporum in the Ocel-lularia and Thelotrema clades, Ocellularia stylothecia and Melanotrema meiosporum in the Fissurina andOcellularia clades, and Myriotrema pycnoporellum, Myriotrema clandestinum and Wirthiotrema glaucopal-lens in the Fissurina, Ocellularia, and Topeliopsis clades. Pagel’s test of independent character evolutionsuggested that at least for some of the traits involved in these cases, ecological constraints may havecaused their evolution in parallel. The most intriguing find is the correlation between gall-forming thalliand vertical columns of calcium oxalate crystals, suggesting that these crystals do not function as lightdistributors, as previously assumed, but instead stabilize the thalli which are usually hollow beneath,similar to a dome-shaped structure. Ancestral character state reconstruction together with an approachto visualize the phenotype of putative ancestral lineages suggested the alpha-Graphidaceae to resemblesome of the extant species currently classified in Myriotrema s.lat., with pore-like ascomata, and non-amyloid ascospores with lens-shaped lumina.
� 2011 Elsevier Inc. All rights reserved.
1. Introduction
One of the most intriguing evolutionary phenomena is the inde-pendent evolution of similar or identical traits from a distantancestor that did not have the traits in question. This phenomenonis variously termed ‘parallel’ or ‘convergent’ evolution, or ‘homo-plasy’, but these terms address very specific and conceptually dif-ferent evolutionary phenomena (Moore and Willmer, 1997; Zhangand Kumar, 1997; Olson and Hall, 2003; Futuyma, 2005; Stearnsand Hoekstra, 2005; Barton et al., 2007). Convergent evolution de-notes the independent evolution of functionally similar traits
based on different structural elements in unrelated or distantly re-lated lineages. These traits involve different structural elementsand their underlying body plan shows anatomical and ontogeneticdifferences; such traits are called ‘analogous’. A classical examplefor convergent evolution are the wings of bats and pterodactyls:being phylogenetically unrelated, they represent a similar solutionto biological ‘engineering’, but use different parts of the vertebrateforelimb (Stearns and Hoekstra, 2005). The opposite of convergentwould be divergent evolution, in which related lineages acquirevery different traits due to radiation, but based on homologousbody parts. Divergent evolution is common in organisms with anopen body plan, such as plants and fungi, in which closely relatedlineages often exhibit phenotypic divergence (Blanco et al.,2004a,b; Tehler and Irestedt, 2007; Mugambi and Huhndorf,2009; Lumbsch et al., 2010; Parnmen et al., 2010). A prominent
1055-7903/$ - see front matter � 2011 Elsevier Inc. All rights reserved.doi:10.1016/j.ympev.2011.04.025
⇑ Corresponding author at: Department of Botany, The Field Museum, 1400 S.Lake Shore Drive, Chicago, IL 60605, USA.
E-mail address: [email protected] (E. Rivas Plata).
Molecular Phylogenetics and Evolution 61 (2011) 45–63
Contents lists available at ScienceDirect
Molecular Phylogenetics and Evolution
journal homepage: www.elsevier .com/ locate /ympev

example of convergent and divergent evolution in lichenized fungiare the hyphophore anamorphs of the family Gomphillaceae(Lücking et al., 2008). These exhibit remarkably divergent evolu-tion in different lineages whereas some forms convergentlyevolved to resemble the campylidia anamorphs of the Pilocarpa-ceae (Lücking et al., 2008).
Parallel evolution occurs when related lineages evolve similaror identical traits independently but based on the same ancestraltrait or body parts. A spectacular example is the evolution of sim-ilar life forms in the two branches of mammals, placentals andmarsupials, such as the extinct European sabre-tooth tiger (Smil-odon) and the South American marsupial sabre-tooth (Thylacosmi-lus). Other well-known examples are the neotropical poison dartfrogs (Dendrobatidae) versus the Malagasy poison frogs (Mantelli-dae) (Schaefer et al., 2002). In general, the underlying structuresthat evolved in parallel are homologous, but their evolutionarychange occurs independently in each lineage. In fungi, an interest-ing case of parallel evolution is found in termite-egg mimicry inunrelated lineages (Matsuura and Yashiro, 2010). Parallel evolutionoften occurs repeatedly within a lineage, then referred to as evolu-tionary trend. Disentangling phenomena such as divergent, con-vergent and parallel evolution is crucial for our understanding ofevolutionary processes.
Fungi, including lichens, are a prime example of how the lack ofunderstanding of divergent, convergent, and parallel evolutioncaused instable classification schemes over time. Traditional char-acters used to define major lineages within Ascomycota and Basid-iomycota, such as fruiting body type and development, have beenshown to have evolved multiple times even among closely relatedlineages (Berbee and Taylor, 1992; Gargas and Taylor, 1995;Berbee, 1996; Hibbett et al., 2007; Larsson and Jeppson, 2008;Mugambi and Huhndorf, 2009). Striking examples among liche-nized fungi are the order Ostropales in the Ascomycota and thefamily Hygrophoraceae (Agaricales) in the Basidiomycota, whichnow include lineages that previously had been placed in differentclasses due to their different fruiting body types: Porinaceae withperithecia in the otherwise apothecioid Ostropales and Dictyonemawith resupinate basidiocarps in the otherwise mushroom-formingHygrophoraceae (Grube et al., 2004; Lawrey et al., 2009; Schmittet al., 2009; Baloch et al., 2010).
While divergent and parallel evolution of fruiting body typesin fungi has been repeatedly documented in molecular studies(Lutzoni et al., 2004; Hibbett et al., 2007; Schoch et al., 2009),emerging phylogenies of selected clades reveal that divergentand parallel evolution occurred at unexpected levels. Phyloge-netic studies in the lichen family Graphidaceae, the largest fam-ily of tropical lichens with possibly close to 2000 species,suggested that monophyletic genera shared thallus and fruitingbody morphotypes (Staiger, 2002; Frisch et al., 2006), contraryto previous classifications which defined genera based on singu-lar taxonomic characters such as ascospores (Staiger, 2002;Frisch et al., 2006; Lücking, 2009; Rivas Plata et al., 2010). How-ever, a larger taxon sampling showed that these revised genericconcepts can be equally misleading and that identical morpho-types representing an entire suite of phenotypic traits evolvedseveral times independently within the family (Rivas Plataet al., 2011).
In the present paper, we document and analyze specific cases ofparallel evolution and evolutionary divergence of fruiting body andthallus types in the lichen family Graphidaceae. We used a molec-ular phylogeny based on sequences of ribosomal nuclear and mito-chondrial genes and the protein-coding RPB2 gene to reconstructthe phylogeny and better understand the evolution of particularphenotypes in this group of fungi. To that end, we also employedancestral character state reconstruction and tested evolutionarymodels of independent character evolution.
2. Material and methods
2.1. Taxon selection and DNA extraction
Ingroup taxa were selected based on variation of phenotypecharacters, including thallus, ascoma morphology, anatomy, andsecondary chemistry. The molecular dataset included 33 of the42 currently recognized genera. The data matrix from which thephenotype characters were extracted contained a total of 130 char-acters in multistate coding and 215 characters in binary coding. Forour analyses, we used a subset of 128 binary-coded characters(Appendix A). As outgroup taxa we used representatives of threelineages shown to be close to Graphidaceae (Porinaceae,Coenogoniaceae, Gyalectaceae), plus Ramonia valenzueliana(Baloch et al., 2010).
We assembled a molecular dataset of three genes for thisstudy: mitochondrial small subunit ribosomal DNA (mtSSU), nu-clear large subunit ribosomal DNA (nuLSU), and the second larg-est subunit of the nuclear RNA polymerase II (RPB2). Relevantsequences were downloaded from GenBank (Table 1). New se-quences were generated for this study using the SigmaREDExtract-N-Amp Plant PCR Kit (St. Louis, Missouri, SA) forDNA extraction following the manufacturer’s instructions, exceptthat we extracted from small samples of fruiting individualbodies and therefore only 40 ll of extraction buffer and 40 lldilution buffer were used. DNA dilutions (5�) were used in PCRreactions of the genes coding for the nuLSU, mtSSU and RPB2,respectively. Primers for amplification were: (a) for nuLSU:AL2R (Mangold et al., 2008), and nu-LSU-1125-30 (=LR6) (Vilgalysand Hester, 1990), (b) for mtSSU: mr-SSU1 (Zoller et al., 1999)and Mr-SSU3R (Zhou and Stanosz, 2001), and (c) for RPB2:fRPB2-7cF and fRPB2-11aR (Liu et al., 1999). PCR reactions con-tained 5.0 ll R4775 Sigma REDExtract-N-Amp™ PCR ReadyMix,0.5 ll of each primer (10 lM), 2 ll genomic DNA extract and2 ll distilled water for a total of 10 ll. Thermal cycling parame-ters were: (1) for nuLSU: initial denaturation for 5 min at 94 �C,followed by 35 cycles of 30 s at 95 �C, 30 s at 58 �C, 1 min at72 �C, and a final elongation for 10 min at 72 �C; (2) for mtSSU:initial denaturation for 5 min at 95 �C, followed by 35 cycles of45 s at 94 �C, 1 min at 50 �C, 1 min 30 s at 72 �C, and a final elon-gation for 10 min at 72 �C; and (3) for RPB2: initial denaturationfor 3 min at 95 �C, then 1 min at 95 �C, and 37 cycles of 1 minat 57 �C, 1 min at 58 �C, 1 min at 59 �C, 1 min at 60 �C, 1 min at61 �C, 1 min at 62 �C, 1 min at 63 �C, 1 min at 64 �C and 1.5 minat 72 �C, and a final elongation for 10 min at 72 �C. Samples werevisualized on a 1% ethidium bromide-stained agarose gel underUV light and bands were gel extracted, heated at 70 �C for5 min, cooled to 45 �C for 10 min, treated with 1 ll GELase (Epi-centre Biotechnologies, Madison, WI, USA) and incubated at45 �C for at least 24 h. The 10 ll cycle sequencing reactionsconsisted of 1–1.5 ll of Big Dye version 3.1 (Applied Biosystems,Foster City, California, USA), 2.5–3 ll of Big Dye buffer, 6 lMprimer, 0.75–2 ll GELased PCR product and water. Samples weresequenced with PCR primers. The cycle sequencing conditionswere as follows: 96 �C for 1 min, followed by 25 cycles of 96 �Cfor 10 s, 50 �C for 5 s and 60 �C for 4 min. Samples were precipi-tated and sequenced using Applied Biosystems 3730 DNAAnalyzer (Foster City, California, USA), sequences were assem-bled in SeqMan 4.03 (DNASTAR) and submitted to GenBank(Table 1).
2.2. Alignment and phylogenetic analyses
Sequences were arranged into multiple alignments for eachgene using BioEdit 7.09 (Hall, 1999) and automatically prealignedwith CLUSTAL W2 (Larkin et al., 2007) to sort out problematic
46 E. Rivas Plata, H.T. Lumbsch /Molecular Phylogenetics and Evolution 61 (2011) 45–63

Table 1Species and specimens included in the analysis with GenBank numbers indicated. All ingroup specimens were critically revised by the authors and collection information oningroup voucher specimens is given. Species names with an asterisk indicate corrected taxonomic identifications compared to GenBank data.
Species mtSSU nuLSU RPB2 Voucher
OutgroupRamonia valenzueliana AY300921 AY300871 – –Belonia russula AY648888 FJ941887 – –Coenogonium luteum AY584699 AF279387 AY641038 –Coenogonium pineti AY300884 AY300834 – –Gyalecta hypoleuca – AF465453 AY641060 –Gyalecta jenensis AF431956 AF465450 AY641043 –Gyalecta ulmi AY300888 AF465463 AY641044 –Porina aenea DQ168411 – – –Porina epiphylla DQ168374 – – –Porina limbulata DQ168403 – – –Porina nitidula DQ168392 – – –Porina nucula DQ168388 – – –
IngroupAcanthothecis peplophora HQ639593 HQ639623 – Florida, Lücking & Rivas Plata 26501 (F)Ampliotrema amplius JF828958 JF828973 – Peru, Rivas Plata 8-9 (F)Carbacanthographis chionophora JF828959 JF828974 – Peru, Rivas Plata 8-9-A (F)Carbacanthographis chionophora – – JF828939 Brazil, Caceres 6040 (F)Chapsa astroidea EU075566 EU075614 – Australia, Lumbsch 19166nChapsa meridensis EU075610 EU075655 JF828940 Costa Rica, Lücking 17770 (F)Chapsa pulchra EU075571 EU075619 JF828941 Australia, Lumbsch 19129 f (F)Chroodiscus defectus FJ708497 FJ708490 – Thailand, Papong 5118Ocellularia thailandica JF828960 JF828975 – Thailand, Lumbsch 19955d (F)Diorygma poitaei HQ639596 HQ639627 JF828942 Nicaragua, Lücking 28538 (F)Dyplolabia afzelii HQ639594 HQ639628 – Florida, Lücking & Rivas Plata 26509 (F)Fissurina astroisidiata DQ431954 DQ431923 – Mexico, Lücking RLD051Fissurina nigromarginata JF828961 JF828976 JF828943 Philippines, Rivas Plata 1198B (F)Glyphis cicatricosa HQ639610 HQ639630 – El Salvador, Lücking 28047 (F)Glyphis scyphulifera DQ431956 – – Dominican Republic, Kalb 33140Glyphis substriatula DQ431982 DNA3191 DNA3191 El Salvador, Lücking 28009Graphis illinata HQ639614 HQ639634 – Mexico, Lumbsch 19639 (F)Graphis librata HQ639621 HQ639636 JF828945 El Salvador, Lücking 28001 (F)Gyrotrema wirthii HQ639591 – – Costa Rica, Lücking 16301 (F)Halegrapha chimera JF505933 – – Philippines, Rivas Plata 1022C (F)Hemithecium chlorocarpum HQ639595 HQ639650 JF828946 Guatemala, Lücking 25522d (F)Hemithecium implicatum DQ431978 HQ639654 JF828947 El Salvador, Lücking 28039 (F)Leptotrema wightii EU075574 JF828977 JF828948 El Salvador, Lücking 28095 (F)Leucodecton compunctellum DQ384904 – – Tanzania, Frisch 99/Tz66Leucodecton compunctellum – JF828978 – Australia, Lumbsch 19092q (F)Leucodecton occultum HQ639611 HQ639657 JF828949 El Salvador, Lücking 28098 (F)Leucodecton phaeosporum JF828962 – – Australia, Mangold 22zl (F)Melanotrema columellatum DQ384894 – – Australia, Kalb 34014Melanotrema endomelaenum FAB1414 – – Australia, Lumbsch 19136l (F)Melanotrema meiospermum DQ384890 – – Tanzania, Frisch 99/Tz470aMyriotrema clandestinum JF828963 – – Australia, Lumbsch 19132 v.A (F)Myriotrema laeviusculum – AY605070 – Costa Rica, Sipman 47896Myriotrema microporum EU075578 EU075626 – Australia, Lumbsch 19092oMyriotrema peninsulae HQ639616 – JF828950 Florida, Lücking & Rivas Plata 26542 (F)Myriotrema pycnoporellum HQ639584 HQ639658 – Florida, Lücking & Rivas Plata 26545 (F)Nadvornikia hawaiiensis EU075581 HQ639659 – Australia, Mangold 36w (F)Ocellularia bahiana – AY605067 – Costa Rica, Nelsen 2032aOcellularia diacida EU075583 EU075630 – Australia, Lumbsch 19120jBOcellularia obturascens JF828965 – – Australia, Lumbsch 19120jD (F)Ocellularia obturascens JF828967 JF828979 – Florida, Lücking & Rivas Plata 26553a (F)Ocellularia papillata EU075586 HQ639661 – Mexico, Lücking RLD063 (F)Ocellularia profunda EU075590 EU075636 – Australia, Lumbsch 19123kOcellularia stylothecium HQ639597 HQ639662 – Nicaragua, Lücking 28636 (F)Ocellularia terebrata JF828964 – – Australia, Lumbsch 19132zd (F)Pallidogramme chlorocarpoides JF828968 – – Philippines, Rivas Plata 1020E (F)Pallidogramme chlorocarpoides – DQ431932 – Australia, Kalb 33926Pallidogramme chlorocarpoides – – JF828951 Philippines, Rivas Plata 2008 (F)Phaeographis dendritica HQ639592 – – Costa Rica, Luecking 21030 (F)Phaeographis intricans DQ431960 – – Tanzania, Frisch 99/Tz1863Phaeographis intricans – DQ431927 – Australia, Kalb 33934Platythecium sphaerosporellum – AY605071 – Costa Rica, Lücking 15070aRedingeria glaucoglyphica HQ639601 – – Peru, Rivas Plata 8-2-1 (F)Reimnitzia santensis HQ639622 HQ639664 JF828952 El Salvador, Lücking 2801 (F)5Sarcographa labyrinthica JF828969 – JF828953 Florida, Lücking & Rivas Plata 26559 (F)Schistophoron tenue EU544933 EU544932 – UnknownStegobolus subemersus DQ384887 – – Brazil, Kalb 31665Thalloloma hypoleptum HQ639609 – – Costa Rica, Lücking & Rivas Plata 17573b (F)Thalloloma hypoleptum – HQ639665 JF828954 Florida, Lücking & Rivas Plata 26564 (F)Thecaria quassiicola JF828971 – – Philippines, Rivas Plata 2011 (F)
(continued on next page)
E. Rivas Plata, H.T. Lumbsch /Molecular Phylogenetics and Evolution 61 (2011) 45–63 47

sequences and finally aligned with MAFFT using the auto option(Katoh and Toh, 2005). The unaligned sequences were also submit-ted to the GUIDANCE web server at http://guidance.tau.ac.il (Pennet al., 2010a,b) to assess alignment uncertainty and delimit ambig-uously aligned regions (mostly for mtSSU) which were then ex-cluded from further analysis. This resulted in alignments of 834bases for mtSSU, 944 for nuLSU, and 963 for RPB2. The individualgene trees were tested for topological conflicts (Mason-Gamerand Kellogg, 1997; Miadlikowska and Lutzoni, 2000; Kauff andLutzoni, 2002), and since no supported conflict was found, com-bined into a single multiple alignment of 2741 bases.
The alignment was subjected to maximum likelihood searchusing RAxML 7.2.6 (Stamatakis and Ludwig, 2005; Stamatakis,2006), with parametric bootstrapping using 500 replicates. Weused the program jMODELTEST (Posada, 2008) to detect the mostappropriate evolutionary model for each gene partition and ap-plied the GTR model for all three gene partitions. Monophyly con-straints were tested using the Shimodaira–Hasegawa (SH) test(Shimodaira and Hasegawa, 1999) as implemented in RAxML(Rambaut and Drummond, 2007). To test for congruence betweentree searching methods, we also employed Bayesian analysis inMrBAYES 3.1.1 (Huelsenbeck and Ronquist, 2001) and maximumparsimony in PAUP� (Swofford, 2003). MrBayes was run for 5 mil-lion generations with two chains and trees were sampled every500 generations, resulting in 10,000 trees per chain. The first2500 trees of each chain were discarded as the ‘burn in’. We usedAWTY (Nylander et al., 2008) to compare splits frequencies be-tween runs and to plot cumulative split frequencies to ensure thatstationarity was reached, and obtained a sample of 13,892 treesfrom both chains. PAUP was run using the following parameters:gaps treated as missing data, 100 addition sequence replicates,TBR as branch swapping algorithm, steepest descent option notin effect, ‘MulTrees’ option in effect, bootstrapping with 50%resampling and 500 replicates.
2.3. Phenotype traits and ancestral character state reconstruction
Ancestral character state reconstruction was done in MESQUITE2.73 (Maddison and Maddison, 2010) based on the single mostlikely tree found in the ML analysis, the set of 162 equally parsimo-nious trees found in a MP analysis, and the set of 13,892 trees re-tained by the Bayesian analysis, using the Trace Character Historyfunction. For this purpose, the phenotype character matrix of 128binary-coded characters was mapped onto the molecular trees.For each character, the node position of likely gains and losses oftraits occurring over the tree(s) was determined. We also recon-structed the ancestral character states for the most basal ingroupnode and the basal nodes of the major clades and subsequentlycomputed a linear correlation between the node character statesand those of all currently accepted species within the family, usingour database, to visualize the most likely phenotypes of putative
ancestral lineages. For this purpose, character states at each nodewere coded as either 0 or 1 if the likelihood for that state was atleast 75% and as equivocal if the likelihood was between 25% and75%. ADOBE Photoshop™ CS2 was used to create phenotype visual-izations based on the most similar extant species that agreed in allmorphological characters with the reconstructed ancestral nodes.
To address possible evolutionary correlations between pheno-type traits, we used Pagel’s (1994) test for association among dis-crete variables, as implemented in MESQUITE 2.73. This methodestimates transition rates from the data and uses a probabilisticmodel to infer the likelihood of joint changes between two charac-ters versus a model of independent evolution. The test was done onthe ultrametrized combined ML tree (Leclere et al., 2009). The P va-lue for each pair of characters was estimated from 100 or 1000 sim-ulations. Usually 1000 simulations for each case is recommended,but it was found to be prohibitive in terms of computing time. In-stead, we analyzed character pairs of interest and ran a higher num-ber of simulations only in cases where the P value resulted close toeither 0.05 or 0.001. The number of analyzed pairs amounted to 30.
We computed distance matrices from both molecular (mtSSU)and phenotype data (divided into morphology and ascoma anat-omy) using PAUP� (Swofford, 2003). A Jukes-Cantor model wasused for the molecular data and total character difference for thephenotype data. For the molecular data we used the mtSSU parti-tion since this was the most complete set and this gene is suffi-ciently variable to allow for quantification of genetic differencesbetween closely related taxa. We compared the molecular andmorphological distances for selected pairs of taxa to quantity thelevel of parallel evolution in distantly related and phenotype diver-gence in closely related taxa.
3. Results
3.1. Phylogenetic analyses
Maximum likelihood analysis of combined mtSSU, nuLSU, andRPB2 data resulted in a single most likely tree with a log-likelihoodof �37048.225 (Fig. 1). Rate parameters were estimated as follows:A–C = 0.851, A–G = 3.626, A–T = 1.694, C–G = 0.819, C–T = 5.400,and G–T = 1.000. Base frequencies were estimated at pi(A) =0.285, pi(C) = 0.197, pi(G) = 0.259, and pi(T) = 0.260. Maximumparsimony analysis resulted in 162 equally parsimonious treeswith a length of 7656 steps each. Bayesian analysis recovered13,892 credible trees. Maximum likelihood (ML), maximum parsi-mony (MP), and Bayesian analyses recovered five major clades asmonophyletic with support and identical backbone topology(Fig. 1). The outgroup was paraphyletic with Porinaceae sister tothe remaining taxa, Coenogoniaceae sister to Gyalectaceae plusGraphidaceae, and Gyalectaceae sister to Graphidaceae. Outgrouptopology was identical in both analyses and partially supportedin a Bayesian framework and under parsimony.
Table 1 (continued)
Species mtSSU nuLSU RPB2 Voucher
OutgroupThecaria quassiicola HQ639617 HQ639667 – Florida, Lücking & Rivas Plata 26567 (F)Thelotrema bicinctulum EU075598 EU075642 JF828955 Australia, Mangold 34f (F)Thelotrema diplotrema EU075599 EU075643 – Australia, Lumbsch 19127vThelotrema expallescens – AY605072 – Costa Rica, Lücking 15069Thelotrema lepadinum AY300916 AY300866 – Unknown, Printzen 5745Thelotrema porinaceum EU675291 – JF828956 Australia, Lumbsch 19156d (F)Thelotrema porinaceum – AY605084 JF828956 Australia, Lumbsch 19108d (F)Topeliopsis muscigena EU075611 EU075656 JF828957 Australia, Mangold 5d2 (F)Wirthiotrema glaucopallens JF828972 – – Australia, Lumbsch 19167c (F)Wirthiotrema glaucopallens – AY605069 – Costa Rica, Lücking 15620
48 E. Rivas Plata, H.T. Lumbsch /Molecular Phylogenetics and Evolution 61 (2011) 45–63

Graphidaceae was strongly supported as monophyletic in allanalyses. The family formed two major clades with strong support:the Fissurina clade including Fissurina and Dyplolabia as well as thefollowing species: Myriotrema pycnoporellum, Ocellularia aff. cruen-tata, and Ocellularia stylothecium, and a clade including the remain-ing taxa. The latter contains four supported clades: the Ocellulariaclade, with the genus Ocellularia s.lat. and related genera (Amplio-trema, Gyrotrema, Leptotrema,Melanotrema,Myriotrema, Redingeria,Reimnitzia, Stegobolus); the Graphis clade, with the bulk of the lirel-late Graphidaceae (Graphis, Diorygma, Glyphis, Halegrapha, Hemi-thecium, Pallidogramme, Phaeographis, Platythecium, Sarcographa,Schistophoron, Thecographa, Thalloloma); the Topeliopsis clade,which includes the genera Acanthothecis, Carbacanthographis,Nadvornikia, Topeliopsis, and Wirthiotrema, as well as some or-phaned species currently placed in Myriotrema and Thelotrema;and the Thelotrema clade, which includes Thelotrema and the re-lated genera Chapsa, Chroodiscus, and Leucodecton.
Of the currently accepted genera within Graphidaceae, thefollowing appeared polyphyletic in this analysis: Fissurina, with
M. pycnoporellum nested within, suggesting that at least two fis-surinoid lineages with either non-carbonized or carbonized excip-ulum are involved; Ocellularia s.lat., with at least six separatelineages in the Ocellularia clade, which differ in thallus anatomyand ascospore types; Myriotrema, with four lineages in additionto the type lineage (M. microporum which is closely related tothe type M. olivaceum), two of which appear outside of the Ocellu-laria clade clustering with Fissurina and Nadvornikia and species ofThelotrema s.lat., respectively; Graphis and Hemithecium, with twoseparate lineages in the Graphis clade; Chapsa, with two separatelineages in the Thelotrema clade, and Thelotrema s.lat., with twospecies clustering with Nadvornikia in the Topeliopsis clade. Theclade formed by the mazediate Nadvornikia and the species Myrio-trema peninsulae, Thelotrema bicinctulum, and T. expallescens, isstrongly supported. The polyphyly of the aforementioned generais currently under study and we maintained current generic assig-nations pending formal changes. In all cases, monophyly of thesegenera was rejected at P = 0.001 by the Shimodaira–Hasegawa(SH) test.
Fig. 1. Most likely tree found using maximum likelihood search in RAxML 7.2.6. Values above and below or left of branches indicate bootstrap support values from maximumlikelihood search (500 replicates) and maximum parsimony search (500 replicates) are indicated above branches and posterior probabilities from Bayesian analysis. Grey dotsdenote notes for which ancestral character states were reconstructed.
E. Rivas Plata, H.T. Lumbsch /Molecular Phylogenetics and Evolution 61 (2011) 45–63 49

3.2. Phenotype traits
All five clades were found to be phenotypically diverse and in-clude species with rounded (previously separated in the familyThelotremataceae) and lirellate ascomata (Figs. 2 and 3). Anatom-ical features previously regarded as characterizing natural groupsappear dispersed over the tree. One example is the formation ofa columella, a sterile tissue within the hymenium, which is foundin species of the Fissurina clade (Ocellularia stylothecia) and theOcellularia clade (Fig. 2(8 and 12 and 3(21–23)). A further trait isa carbonized excipulum which appears to alternate withuncarbonized excipula in pairs of closely related species, such asFissurina, several instances within the Ocellularia clade (e.g.,Ocellularia papillata versus Myriotrema clandestinum), the two Gra-phis/Hemithecium subclades, and the Carbacanthographis/Acantho-thecis subclade.
Myriotremoid ascomata consist of immersed pores with a non-carbonized excipulum lacking periphysoids (lateral paraphyseswith horizontal orientation), and are often found in combination
with a shiny thallus featuring a prosoplectenchymatous, splittingcortex; this combination occurs in several lineages throughoutthe tree, such as in M. pycnoporellum (Fig. 2(5)) in the Fissurinaclade,M. clandestinum (Fig 3(1–3)) in the Ocellularia clade, andWir-thiotrema (Fig 3(4 and 5) in the Topeliopsis clade. Taxa with lepadi-noid or immersed-lepadinoid ascomata, in which the apotheciaform a characteristic double margin and the thallus lacks a com-pact cortex (Fig. 2(28, 29, 31, 11 and 15), occur in the Ocellulariaclade (Fibrillithecis halei, Myriotrema microporum, Melanotremaendomelaenum), the Thelotrema clade (Thelotrema diplotrema,Thelotrema lepadinum, Leucodecton occultum), and the Topeliopsisclade (M. peninsulae, T. bicinctulum, Thelotrema expallescens).Chroodiscoid ascomata in which the disc is exposed by recurvinglobules (Fig. 2(2, 9) and 3(16–19) evolved in all clades and subc-lades, such as Ocellularia aff. cruentata in the Fissurina clade, Gyro-trema and Reimnitzia in the Ocellularia clade, Phaeographis lobata inthe Graphis clade (not included), and Chapsa and Chroodiscus in theThelotrema clade. In many of these cases, the disc is brightly pig-mented (Fig. 2(2 and 9).
Fig. 2. (1–8) Species of the Fissurina clade. (1) Fissurina astroisidiata. (2 and 4) Ocellularia aff. cruentata (in four non-amyloid ascospores with diamond-shaped lumina). (3)Fissurina nigrolabiata. (5) Myriotrema pycnoporellum. (6 and 8) Ocellularia stylothecia (in eight section through apothecium showing broad-stump-shaped, carbonizedcolumella). (7) Dyplolabia afzelii. (9–12) Species of the Ocellularia clade resembling those in the Fissurina clade. (9–10) Gyrotrema wirthii (in 10 amyloid ascospore with lens-shaped lumina). (11–12) Melanotrema meiospermum (in 12 section through apothecium showing broad-stump-shaped, carbonized columella). (13–21) Species of theOcellularia and Thelotrema clades with gall-forming thalli. (13–15) Leptotrema wightii (in 14 section through thallus showing vertical columns of crystals, in 15 thallus andapothecia enlarged). (16–18) Myriotrema laeviusculum (in 17 dark brown ascospores, in 18 thallus and apothecia enlarged). (19–21) Leucodecton phaeosporum (in 20 sectionthrough thallus showing vertical columns of crystals, in 21 dark brown ascospores). (22–27) Phenotype divergence in the Glyphis and Sarcographa/Thecaria subclade of theGraphis clade. (22–24) Glyphis scyphulifera, G. substriatula, and G. cicatricosa with rounded, lirellate, and stromatic ascomata. (25–26) Thecaria quassiicola with rounded andwith lirellate ascomata (often ocurring on the same thallus). (27) Sarcographa labyrinthica with stromatic ascomata. (28–33) Species of the Nadvornikia subclade in theTopeliopsis clade and the genus Thelotrema in the Thelotrema clade. (28–31) Lepadinoid apothecia in Thelotrema lepadinum, Myriotrema peninsulae (in 30 ascospores), andThelotrema expallescens. (32–33) Nadvornikia hawaiiensiswith mazaediate ascomata (in 33 ascospores). (34–35) Topeliopsis muscigena (Topeliopsis clade) and Chapsa meridensis(Thelotrema clade) with topeliopsidoid ascomata growing over bryophytes.
50 E. Rivas Plata, H.T. Lumbsch /Molecular Phylogenetics and Evolution 61 (2011) 45–63

Another frequently evolved type are graphidoid ascomata,which are lirellate and produce thick, carbonized excipula(Fig. 2(3 and 7) and 3(24–27); such ascomata are found in Fissurinaand Dyplolabia in the Fissurina clade, Graphis s.lat., Halegrapha, andGlyphis in the Graphis clade, and Carbacanthographis in the Topeli-opsis clade. Hemithecioid ascomata are similar but lack carboniza-tion (Fig. 3(28–31); such ascomata evolved frequently in theFissurina clade, the Graphis clade (Hemithecium s.lat., Pallido-gramme), and the Topeliopsis clade (Acanthothecis). Mazediate asco-mata are found in two unrelated lineages, Schistophoron in theGraphis clade and Nadvornikia in the Topeliopsis clade (Fig. 2(32)and 3(20)).
3.3. Ancestral character state reconstruction
Ancestral character state reconstruction (both likelihood andparsimony) suggested that round ascomata are ancestral withinthe family and lirellate ascomata are derived (Table 2). Roundascomata were also reconstructed as ancestral for the Fissurina,Ocellularia, Topeliopsis, and Thelotrema clades and lirellate
ascomata for the Graphis clade. Under likelihood, the ancestralexcipular type in the family was indicated as uncarbonized, pros-oplectenchymatous, and lacking lateral paraphyses, but as uncar-bonized to dark brown under parsimony. Full carbonization, i.e.black excipula, evolved several times independently in theFissurina and Topeliopsis clades. The ancestral state for theOcellularia and Graphis clades was reconstructed as carbonized,which suggests that reversal to uncarbonized excipula occurredseveral times within these clades.
A paraplectenchymatous excipulum and/or lateral paraphysesare synapomorphies for most lineages within the Topeliopsis andThelotrema sister clades, but ancestral state reconstruction wasequivocal for the presence of lateral paraphyses (Table 2).Pigmented ascospores evolved multiple times within Graphida-ceae. The ancestral ascospore type for the family was recon-structed as hyaline and non-amyloid with thickened walls andlens-shaped to rounded lumina, a type which in extant taxa isonly found in certain species of Fissurina, Myriotrema protoalbum,Carbacanthographis and in some species of Chapsa andThelotrema.
Fig. 3. (1–5) Species of the Ocellularia and Topeliopsis clade with myriotremoid apothecia. (1–3) Myriotrema clandestinum (in 2, 3 ascospores with lens-shaped lumina beforeand after staining with Iodine solution). (4–5) Wirthiotrema glaucopallens (in E non-amyloid ascospore with angular lumina). (6–7) Species of the Ocellularia clade withampliotremoid apothecia. (6) Ampliotrema amplius. (7) Ocellularia bahiana. (8–10) Species of the Ocellularia and Thelotrema clades with porinoid apothecia. (8) Ocellulariaprofunda. (9) Thelotrema porinaceum. (10) Leucodecton compunctellum. (11–15) Species of the Ocellularia, Topeliopsis, and Thelotrema clades with immersed-lepadinoidapothecia. (11)Myriotrema microporum. (12)Melanotrema endomelaenum. (13) Thelotrema bicinctulum. (14) Thelotrema diplotrema. (15) Leucodecton occultum. (16–19). Speciesof the Ocellularia and Thelotrema clades with chroodiscoid apothecia. (16) Reimnitzia santensis. (17) Chroodiscus defectus. (18) Chapsa pulchra. (19) Chapsa astroidea. (20)Schistophoron tenue with mazaediate lirellae. (21–23) Species of the Ocellularia clade with stegoboloid apothecia. (21) Melanotrema columellatum. (22) Stegobolus subemersus.(23) Ocellularia obturascens. (24–27) Species of the Graphis and Topeliopsis clades with graphidoid lirellae. (24) Graphis illinata. (25) Carbacanthographis chionophora. (26)Graphis librata. (27) Halegrapha chimera. (28–31) Species of the Graphis and Topeliopsis clades with hemithecioid lirellae. (28) Hemithecium chlorocarpum. (29) Hemitheciumimplicatum. (30) Pallidogramme chrysenteron. (31) Acanthothecis peplophora. (32–33) Species of the Ocellularia and Graphis clades with diorygmoid lirellae. (32) Redingeriaglaucoglyphica. (33) Diorygma poitaei. (34–35) Species of the Graphis clade with thallolomoid lirellae. (34) Thalloloma hypoleptum. (35) Kalbographa lueckingii.
E. Rivas Plata, H.T. Lumbsch /Molecular Phylogenetics and Evolution 61 (2011) 45–63 51

Table
2Anc
estral
characterstatereco
nstruc
tion
sforGraph
idacea
eingrou
pno
des(m
arke
dby
grey
dots
inFig.
1).R
ange
ofch
aracterstates
aregive
nacco
rdingto
likelihoo
dof
node
reco
nstruc
tion
;if75
%or
high
er,o
nlyon
estateis
indicated,
ifless
than
75%,the
ntherang
eof
possible
states
isindicated.
Differentialch
aracters
betw
eenclad
esarehigh
lighted
initalics.
Baseof
Graph
idacea
eFissurinaclad
eOcellu
lariaclad
eGraph
isclad
eTo
pelio
psis
clad
eTh
elotremaclad
e
Character
Like
lihoo
dBay
esian
Parsim
ony
Like
lihoo
dLike
lihoo
dLike
lihoo
dLike
lihoo
dLike
liho
odSu
bstrate
Cortico
lous
Cortico
lous
Cortico
lous
Cortico
lous
Cortico
lous
Cortico
lous
Cortico
lous
Cortico
lous
Surface
Smoo
thSm
ooth
Smoo
thSm
ooth
Smoo
thSm
ooth
Smoo
thSm
ooth
Gallform
ations
––
––
––
––
Veg
etativeprop
agules
––
––
––
––
Oxa
late
crystals
Not
columnar
Not
columnar
Not
columnar
Not
columnar
Not
columnar
Not
columnar
Not
columnar
Not
columnar
Cortex
Loos
eto
compa
ctLo
oseto
compa
ctLo
oseto
compa
ctPros
oplect
Pros
oplect
Pros
oplect
Pros
oplect
Loos
eto
compa
ctAscom
aem
erge
nce
Erum/prom
Erum/prom
Erumpe
nt
Erum/prom
Erum/prom
Erum/prom
Erum/prom
Erum/prom
Ascom
ash
ape
Rou
nd-irregu
lar
Rou
nd-irregu
lar
Irregu
lar
Irregu
lar
Rou
nd-irregu
lar
Lirella
teIrregu
lar
Rou
nd-irregu
lar
Discex
posu
reCov
ered
/pore
Cov
ered
/pore
Cov
ered
Cov
ered
/pore
Cov
ered
/pore
Covered/slit
Cov
ered
/pore
Cov
ered
/pore
Ascom
amargin
Entire/fissured
Entire/fissured
Fissured
Entire/fissured
Entire/fissured
Entire/fissured
Entire/fissured
Entire/lob
ulate
Dou
blemargin
––
––
––
––
Columella
––
––
––
––
Columella
shap
e–
––
–Fing
er-like
––
–Columella
pigm
entation
––
––
Carbon
ized
––
–Ex
cipu
lum
pigm
entation
Hya
lineto
pale
Hya
lineto
pale
Hya
lineto
dark
Dark/carbon
ized
Carbon
ized
Dark/carbon
ized
Hya
line/da
rkHya
line
/dark
Excipu
lum
stru
cture
Pros
oplect
Pros
oplect
Pros
oplect
Prosop
lect
Prosop
lect
Prosop
lect
Paraplect
Paraplect
Periph
ysoids
––
––
––
–/Present
–/Present
Paraph
yses
capitate
––
––
––
––
Paraph
yses
warts
––
––
––
––
Insp
ersion
––
––
––
––
Maz
aedium
––
––
––
––
Ascos
pore
numbe
r8/Ascus
8/Ascus
8/Ascus
8/Ascus
8/Ascus
8/Ascus
8/Ascus
8/Ascus
Ascos
pore
size
10–5
0�
5–10
lm
10–5
0�
5–10
lm
10–2
5�
5–10
lm
10–2
5�
5–10
lm
10–5
0�
5–10
lm
10–5
0�
5–10
lm
10–5
0�
5–20
lm
10–5
0�
5–20
lm
Ascos
pore
lumina
Lens-sh
aped
Lens-sh
aped
Lens-sh
aped
Lens-sh
aped
Lens-sh
aped
Lens-sh
aped
Subrectangu
lar
Subrectangu
lar
Ascos
pore
ontoge
ny
Macroceph
alic
Macroceph
alic
Macroceph
alic
Macroceph
alic
Macroceph
alic
Macroceph
alic
Macroceph
alic
Macroceph
alic
Ascos
pore
outerwall
Thin
Thin
Thin
Thin
Thin
Thin
Thin
Thin
Ascos
pore
color
Hya
line
Hya
line
Hya
line
Hya
line
Hya
line
Hya
line
Hya
line
Hya
line
Ascos
pore
ornam
entation
––
––
––
––
Ascos
pore
amyloidity
––
––/W
eak
Presen
tPresen
t–
–Tran
sverse
septa
3–9
3–9
3–9
3–9
3–9
3–9
3–9
3–9(–1
9)Lo
ngitudinal
septa
0–2
0–2
1–3
0–2
0–2
0–2
0–2
0–2
Seco
nda
rysu
bstances
––
––
––
––
Med
ullarypigm
ents
––
––
––
––
Apo
thecialpigm
ents
––
––
––
––
52 E. Rivas Plata, H.T. Lumbsch /Molecular Phylogenetics and Evolution 61 (2011) 45–63

Secondary chemistry is highly variable even among closely re-lated species, although taxa with psoromic and protocetraric acidsand related substances are most commonly found in the Ocellulariaclade, whereas species in the Graphis, Topeliopsis, and Thelotremaclades frequently produce stictic and norstictic acids and relatedsubstances or lack secondary compounds. For all basal ingroupnodes, the ancestral state was reconstructed as lacking secondarysubstances, which is in line with the fact that the closest outgrouptaxa lack secondary substances typical of Graphidaceae and sug-gests that secondary chemistry evolved multiple times within thefamily.
Visualization of ancestral lineages (Fig. 4) suggests a myrio-tremoid lineage as basal for the Graphidaceae and for the Fissurinaclade, an ocellularioid lineage as basal for the Ocellularia clade, anda graphioid as basal for the Graphis clade.
Ancestral character state reconstruction has to be done withcare since it is affected by taxon sampling. In the present case,
we tried to achieve a representative balance between differentthallus and fruiting body types occurring in the Graphidaceae.The total number of taxa sampled is not critical as long as the taxonrepresenting a given lineage likely corresponds to the ancestralstates of that lineage. For example, assuming that the genusAmpliotrema is monophyletic and species of the genus agree in rel-evant charactes used for ancestral character state analysis, theanalysis will not be affected by the number of species of Amplio-trema included in the analysis. The same applies to sister claderelationships: if two sister clades differ in a particular characterbut the species within each clade are consistent regarding thischaracter, the number of species sampled in each clade is irrele-vant. We can therefore assume that the phylogeny used in thisanalysis, even if only including a fraction of all species acceptedin the family, is representative of a larger taxon sampling andthe main conclusions are reliable, in particular with regard toancestral ascoma morphology, ascospore type, and chemistry. Anexample is the reconstruction of the excipulum in the Graphis cladeas carbonized, even if the proportion of carbonized versus uncar-bonized taxa (about 1:1) in our sampling was lower than for theentire clade if all known species were included (about 2:1).
3.4. Examples of parallel evolution involving suites of traits
Pagel’s test (1994) was used to determine the correlation of se-lected traits in three pairs of species (Table 3). The most conspicu-ous example of parallel evolution in the family is O. stylothecia inthe Fissurina clade versus Melanotrema meiospermum in the Ocellu-laria clade. Both lineages include species with ecorticate or looselycorticate thalli often producing stictic acid or lacking secondarysubstances, erumpent ascomata that rupture to form small excipu-lar lobules and expose a broad-stump-shaped columella, and heavycarbonization of excipulum and columella (Fig. 2(6, 8, 11 and 12).O. stylothecia and M. meiospermum are morphologically very simi-lar and can only be reliably distinguished by the shape of theirascospore lumina: diamond-shaped in O. stylothecia and lens-shaped in M. meiospermum.
Another striking case is found in Leptotrema wightii and Myrio-trema laeviusculum in the Ocellularia clade and Leucodecton phaeo-sporum in the Thelotrema clade. These species produce thalli thatresemble gall-like structures overgrowing loose bark and bryo-phytes, with the cavities beneath the thallus often inhabited byants. All three have a loose cortex and form characteristic verticalwalls and columns of calcium oxalate crystals. All three have small,pore-like apothecia and small, submuriform, dark brown ascosp-ores, two of the three species lack secondary substances(Fig. 2(13–21). The three species are commonly confused with eachother. Pagel’s tests revealed a strong correlation between a gall-forming thallus, columnar clusters of crystals, and dark brownascospores (Table 3), suggesting evolutionary dependency be-tween these traits.
A third case of parallel evolution involving multiple charactersis the couple Topeliopsis muscigena in the Topeliopsis clade andChapsa meridensis in the Thelotrema clade. Topeliopsis species typi-cally overgrow bryophytes on soil and form sessile, round asco-mata with a denticulate margin and partly covered disc, lateralparaphyses, large muriform ascospores in most species, and a sim-ple chemistry or lack of secondary substances. C. meridensis agreeswith Topeliopsis s.str. in all these features, but molecular evidenceshow that this species is unrelated to Topeliopsis. Pagel’s test re-vealed significant correlation between a muscicolous habit andsessile ascomata with fissured margins, lateral paraphyses, andlarge muriform ascospores (Table 3). Further cases of parallel evo-lution include M. clandestinum in the Ocellularia clade versusWirthiotrema glaucopallens in the Topeliopsis clade (Fig. 3(1–5)),the couple Ampliotrema amplius and Ocellularia bahiana in the
Fig. 4. Visualization of most likely phenotypes of ancestral lineages. (1) Basalancestral lineage of Graphidaceae and Fissurina clade (myriotremoid). (2) Ancestrallineage of Ocellularia clade (resembling Ocellularia zamboangensis). (3) Ancestrallineage of Graphis clade (resembling Graphis scripta).
E. Rivas Plata, H.T. Lumbsch /Molecular Phylogenetics and Evolution 61 (2011) 45–63 53
Ocellularia clade (Fig. 3(6–7)), and the species Ocellularia profunda,Thelotrema porinaceum, and Leucodecton compunctellum in theOcellularia and Thelotrema clades (Fig. 3(8–10)). Stegoboloidapothecia, that is apothecia with complex columella structures(Fig. 3(21–23)), evolved independently in several clades and subc-lades. The same applies for the lirellate ascomata of the Diorygma(Fig. 3(32–33)) and Thalloloma type (Fig. 3(34 and 35)).
3.5. Phenotypic divergence in closely related taxa
Multiple cases of parallel evolution go along with remarkablephenotypic divergence within certain clades. In the Fissurina clade,O. stylothecia, Ocellularia aff. cruentata, and Dyplolabia afzelii arefound on a single clade but exhibit highly disparate morphological,anatomical, and chemical characters. The only shared trait is theascospore type with diamond-shaped lumina (Fig 2(4)). Ascomamorphology is also variable within the genus Glyphis, and closelyrelated species exhibit different ascoma types which in earlier clas-sifications were used to distinguish families: round in G. scyphylif-era, lirellate in G. substriatula, and stromatic in G. cicatricosa(Fig. 2(22–24)). These species had previously been assigned to dif-ferent genera (Gyrostomum, Graphina, Glyphis). Strikingly similardivergence is found in the closely related taxa Sarcographa andThecaria (Fig. 2(25–27). A third example for phenotypic divergenceis found in the lineage including the mazediate Nadvornikia and thethree orphaned species of Myriotrema and Thelotrema in theTopeliopsis clade (Fig. 2(28–33)). The mazediate ascomata of
Nadvornikia are so different from other Graphidaceae that thegenus had previously been placed within the unrelated family, Cal-iciaceae (Lecanorales). There is no morphological transition be-tween Nadvornikia and the other species supported within thisclade.
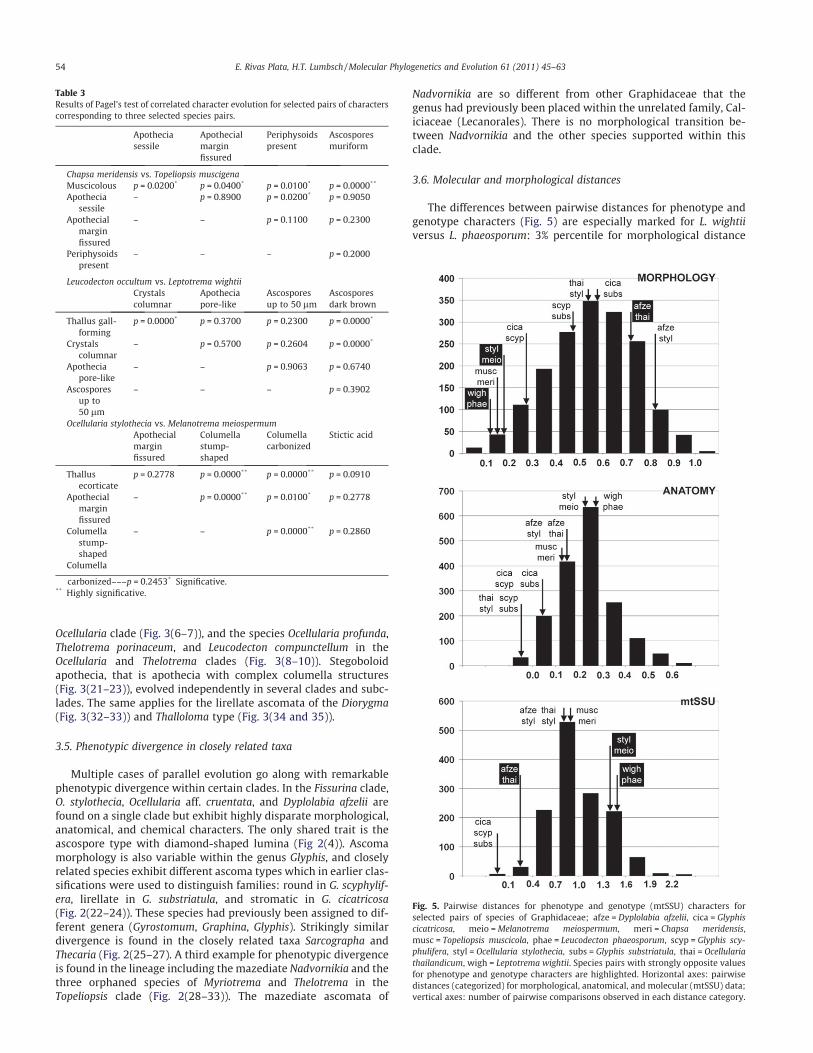
3.6. Molecular and morphological distances
The differences between pairwise distances for phenotype andgenotype characters (Fig. 5) are especially marked for L. wightiiversus L. phaeosporum: 3% percentile for morphological distance
Fig. 5. Pairwise distances for phenotype and genotype (mtSSU) characters forselected pairs of species of Graphidaceae; afze = Dyplolabia afzelii, cica = Glyphiscicatricosa, meio =Melanotrema meiospermum, meri = Chapsa meridensis,musc = Topeliopsis muscicola, phae = Leucodecton phaeosporum, scyp = Glyphis scy-phulifera, styl = Ocellularia stylothecia, subs = Glyphis substriatula, thai = Ocellulariathailandicum, wigh = Leptotrema wightii. Species pairs with strongly opposite valuesfor phenotype and genotype characters are highlighted. Horizontal axes: pairwisedistances (categorized) for morphological, anatomical, and molecular (mtSSU) data;vertical axes: number of pairwise comparisons observed in each distance category.
Table 3Results of Pagel’s test of correlated character evolution for selected pairs of characterscorresponding to three selected species pairs.
Apotheciasessile
Apothecialmarginfissured
Periphysoidspresent
Ascosporesmuriform
Chapsa meridensis vs. Topeliopsis muscigenaMuscicolous p = 0.0200* p = 0.0400* p = 0.0100* p = 0.0000**
Apotheciasessile
– p = 0.8900 p = 0.0200* p = 0.9050
Apothecialmarginfissured
– – p = 0.1100 p = 0.2300
Periphysoidspresent
– – – p = 0.2000
Leucodecton occultum vs. Leptotrema wightiiCrystalscolumnar
Apotheciapore-like
Ascosporesup to 50 lm
Ascosporesdark brown
Thallus gall-forming
p = 0.0000* p = 0.3700 p = 0.2300 p = 0.0000*
Crystalscolumnar
– p = 0.5700 p = 0.2604 p = 0.0000*
Apotheciapore-like
– – p = 0.9063 p = 0.6740
Ascosporesup to50 lm
– – – p = 0.3902
Ocellularia stylothecia vs. Melanotrema meiospermumApothecialmarginfissured
Columellastump-shaped
Columellacarbonized
Stictic acid
Thallusecorticate
p = 0.2778 p = 0.0000** p = 0.0000** p = 0.0910
Apothecialmarginfissured
– p = 0.0000** p = 0.0100* p = 0.2778
Columellastump-shaped
– – p = 0.0000** p = 0.2860
Columella
carbonized–––p = 0.2453* Significative.** Highly significative.
54 E. Rivas Plata, H.T. Lumbsch /Molecular Phylogenetics and Evolution 61 (2011) 45–63

but in the 94% percentile for mtSSU distance; an identical patternis seen for O. stylothecia versus M. meiospermum, meaning thatthese species are morphologically extremely similar but geneti-cally very distant when compared to the overal distribution of dis-tances for all possible species pairs. Less marked but stillsubstantial deviations between morphological and mtSSU dis-tances are found for the pair T. muscigena versus C. meridensis.On the other hand, the pair D. afzelii versus Ocellularia thailandicashows the opposite trend: 91% percentile for morphologicaldistances compared to 3% percentile for mtSSU distances, meaningthat these two species are genetically very close but morphologi-cally very disparate. A less marked trend is found in Glyphis cicatri-cosa versus G. substriatula, G. scyphulifera versus G. substriatula, andD. afzelii versus O. stylothecia. Anatomical distances between thesepairs are intermediate but usually closer to the mtSSU distances,suggesting that anatomical characters, such as hamathecium,ascus, and ascospore type, are better predictors of geneticrelatedness.
4. Discussion
4.1. Parallel evolution in lichenized fungi and Graphidaceae
Parallel evolution in distantly related or unrelated and pheno-typic divergence in closely related lineages have been repeatedlydocumented in Fungi, including lichenized forms (Grube andKantvilas, 2006; Hibbett et al., 2007; Larsson and Jeppson, 2008;Lawrey et al., 2009; Mugambi and Huhndorf, 2009; Baloch et al.,2010). Both phenomena are the rule rather than the exception inthis kingdom and, as a consequence, the classification of Fungihas undergone substantial changes (Hibbett et al., 2007; Kirket al., 2008). At the family level, however, molecular phylogeniesseemed to indicate that monophyletic lineages comprising naturaltaxa are characterized by a rather uniform ascoma and thallusmorphology and anatomy and largely vary in ascospore charactersand chemistry or details in thallus morphology (Staiger, 2002;Miller, 2003; Miller and Huhndorf, 2005; Frisch et al., 2006; Staigeret al., 2006; Mangold et al., 2008). It thus appeared that genera canbe recognized morphologically but require anatomical and chemi-cal details to separate species. This concept was used to revise thegeneric delimitation in Graphidaceae and Thelotremataceae(Staiger, 2002; Frisch et al., 2006), the latter now included inGraphidaceae (Mangold et al., 2008). However, with more molecu-lar data available, it became obvious that similar morphotypeshave evolved in parallel in distantly related lineages withinfamilies of lichenized fungi. In Parmeliaceae, the brown speciesofMelanelia s.lat. evolved both in the parmelioid and in the cetrari-oid clade (Thell et al., 2002; Blanco et al., 2004a; Crespo et al.,2010). Another case are the genera Bulbothrix versus Relicina,believed to be closely related and only differing by cortical chem-istry, whereas molecular data suggest that these genera representdistant, unrelated lineages (Crespo et al., 2010). For most families,detailed molecular phylogenies are not yet available to elucidatethese patterns, and the present study is the first one at family levelin which parallel evolution and phenotypic divergence are amplydocumented using ancestral character reconstruction of a widearray of phenotypic characters.
Our study demonstrates that parallel evolution occurred fre-quently in Graphidaceae and at previously unrecognized levels.Prominent examples are O. stylothecia versus M. meiospermum inthe Fissurina and Ocellularia clades, L. wightii, M. laeviusculum,and L. phaeosporum in the Ocellularia and Thelotrema clades, andT. muscigena versus C. meridensis in the Topeliopsis and Thelotremaclades, but many more have been documented in this study. Thishas consequences for classification, since it suggests that morpho-logical characters, even if used in combination, are not reliable as
predictors of phylogenetic relationships. The evolutionary mecha-nisms and constraints leading to such a high level of parallel evo-lution and phenotypic divergence are unknown and leave room forspeculation, considering that thallus morphology and anatomyprovide the interface with the environment regarding nutrient up-take and photosynthesis and thus competitive strength, ascomamorphology and anatomy guarantee sexual reproduction, asco-spore type relates to successful dispersal and establishment, andsecondary chemistry has diverse functions such as protectionagainst UV radiation and herbi- and fungivores.
4.2. Parallel evolution of thallus morphology in Graphidaceae
Graphidaceae that grow in the shaded rainforest understory,such as most species of Acanthotrema, Fissurina, Myriotrema s.lat.,Ocellularia s.lat., and Wirthiotrema (Rivas Plata et al., 2008), havea thin but distinct, very compact upper cortex with a continuousphotobiont layer below, giving them a shiny, more or less greenappearance when fresh. The cortex is hydrophobic which isprobably an adaptation to prevent the thallus from becomingpermanently and fully hydrated under a constantly humid envi-ronment, since full hydration would impede rapid diffusion ofCO2 within the thallus and specifically the photobiont layer(Lakatos et al., 2006). Yet, the cortex is sufficiently thin to permitentering of sufficient light under the comparatively low levels ofthe rainforest understory to maintain a positive photosyntheticbalance and permit thallus growth. Thus, the independent evolu-tion of similar thallus types can be interpreted as the result of eco-logical constraints.
Few lichens are known in which the photobiont layer is orga-nized in vertical rather than horizontal layers, such as Labyrintha(Malcolm, 1995). This anatomy has been compared with that ofthe ‘window plants’ (Fenestraria) of the Namib desert, in whichvertical crystal columns distribute the light to the photosynthetictissue of the pebble-like, succulent plants that are semi-immersedin the soil (Vogel, 1955; Ihlenfeldt, 1994; Attenborough, 1995).L. wightii, L. phaeosporum, and M. laeviusculum show such a photo-biont arrangement: the photobiont layer is divided into verticalcolumns by very large, columnar clusters of calcium oxalate crys-tals that form a network of vertical ‘walls’ when seen from above.One would assume that this anatomy serves to improve the distri-bution of light within the thallus and thus enhance photosynthesis.However, gall-forming species do not appear to have preferencesfor specific light conditions (Rivas Plata et al., 2008). Instead, thecorrelation between gall-forming thalli and columnar crystals sup-ports an alternative hypothesis, namely that the vertical arrange-ment of the crystal columns is an adaptation to stabilize thesethalli, which are hollow underneath and thus more susceptible tomechanical stress than other thallus types. Such thalli appear torepresent a biological analogy to the architectural solutions of sta-bilizing dome structures (Buckminster Fuller, 1961; Escrig et al.,1997). Thus, the phenotypic similarity of these species can be ex-plained by a combination of ecological and structural constraintson thallus morphology plus the retaining of a plesiomorphic, myr-iotremoid ascoma type.
A third, common type is white, corticate thalli which feature acontinuous layer of calcium oxalate crystals above the photobiontlayer, as found in species of the genus Graphis (Lücking, 2009). Thewhite color is due to reflection of incoming light by the crystallayer, which most probably prevents damage of the photobiontlayer under high light levels, specifically UV. Species with suchthalli are frequently found in exposed habitats and microhabitats,such as the forest canopy (Lücking et al., 2008; Rivas Plata et al.,2008). The observation that species of different lineages withinthe family that coexist in the same microhabitats have similar thal-lus types, including the examples studied in this paper but also
E. Rivas Plata, H.T. Lumbsch /Molecular Phylogenetics and Evolution 61 (2011) 45–63 55

others such as Graphis and Allographa (Rivas Plata et al., 2011),underlines the notion that ecological constraints may cause paral-lel evolution in thallus morphology.
4.3. Parallel evolution of ascoma morphology in Graphidaceae
Very little is known about the detailed mechanisms of thereproductive biology of lichenized fungi (Awasthi, 1983; Hale,1983; Nash, 2008) and much less about tropical families, such asGraphidaceae. Thus, although the family exhibits the highest levelof variation of ascoma types known among all Ascomycota, closelymatched only by the Roccellaceae (Grube, 1998; Tehler andIrestedt, 2007; Nelsen et al., 2009), nothing is known about theadaptive value of different ascoma types. Pagel’s test (Pagel,1994) suggested that the sessile apothecia shared by T. muscigenaand C. meridensis are the result of ecomorphological constraintsrelated to the growth over bryophytes, which requires the apothe-cia to expose themselves vertically to ensure successful ascosporedispersal. It is usually assumed that ascospores are dispersed byabiotic vectors, such as wind and rain, but this would not explainthe morphological variation of ascoma types observed in thefamily. Dispersal via inspects, specifically ants (Lorentsson andMattsson, 1999; Lücking and Bernecker-Lücking, 2000), is apossible explanation for the morphological appearance of certainascoma and ascospore types, but this has not been studied. Else,one would assume that evolution of ascoma morphology and anat-omy is neutral, allowing for shapes and colors to evolve freely, butsince sexual reproduction via ascospores is essential to theselichenized fungi, this seems unlikely.
Another explanation for anatomical features of the ascomatacould be the protection of the hymenial layer, asci, and ascosporesduring development, as it has been observed that the hymenium,for example, is specifically targeted by fungivorous insects suchas psocids (Seaward, 1988; Lücking and Bernecker-Lücking,2000). Many Graphidaceae feature heavily carbonized ascomataor a tendency to completely conceal the hymenium until and evenduring maturity (Staiger, 2002; Frisch et al., 2006), thus potentiallypreventing fungivores from feeding on hymenial structures. Colu-mella structures reduce the hymenium to a narrow, completelyconcealed and protected ring in the periphery of the ascomata. Thiscould explain their independent evolution in the Fissurina andOcellularia clades, although their absence in the remaining cladesmay suggests some level of evolutionary constraint. Species withexposed hymenia, such as in chroodiscoid ascomata, often producea layer of crystal granules and dead hyphae (‘pruina’) on the sur-face of the disc, which might also help to prevent attack byfungivores.
One of the most prominent features of Graphidaceae is thedevelopment of elongate, lirellate ascomata or ascomata groupedin (pseudo-)stromata. Our analysis suggests that lirellate ascomataevolved in each of five clades independently: besides the predom-inantly lirellate Graphis clade, lirellae are found in Dyplolabia andFissurina (Fissurina clade), species of Redingeria and Stegobolus(Ocellularia clade), Acanthothecis and Carbacanthographis (Topeliop-sis clade), and species of Chroodiscus and Chapsa (Thelotrema clade).The immediate advantage of lirellate over round ascomata is notnecessarily evident, but the obvious difference is one of growthpattern: lirellate ascomata can grow relatively unlimited in twodirections, and hence produce large quantities of ascospores, with-out giving up protection of the hymenium by covering it throughthe lateral excipulum. In round ascomata this is not possible, sincecontinuous growth will expose the hymenium. The observationthat species with lirellate ascomata are much more abundant inthe tropics could be explained by the higher abundance and diver-sity of herbivorous and fungivorous invertebrates in tropical eco-
systems (Price et al., 1995; Lücking and Bernecker-Lücking, 2000;Novotny et al., 2006).
4.4. Parallel evolution of ascospore morphology in Graphidaceae
Morphology and anatomy of ascospores are expected to be re-lated to functional aspects of dispersal and establishment. Twotendencies in ascospore morphology are obvious in Graphidaceaeand shared by other families of lichenized tropical fungi, such asPyrenulaceae and Trypetheliaceae (Harris, 1989; Aptroot et al.,2008): pigmented and/or muriform ascospores. The immediateadvantage of pigmented ascospores is unknown. If pigments pro-vide protection against UV radiation after ascospore discharge, itwould be expected that they are more common in species occur-ring in exposed habitats. Data on ecological preferences of Graph-idaceae (Lücking et al., 2008; Rivas Plata et al., 2008) do notsupport this hypothesis: species of Graphiswith hyaline ascosporesare frequently found in exposed habitats, whereas species of Phae-ographis with pigmented ascospores are more common in partiallyshaded microhabitats. A lack of correlation between microhabitatand ascospore pigmentation is also found in the genus Stegobolus,a characteristic element of the shady rainforest understory butwith species having either hyaline or pigmented ascospores (RivasPlata et al., 2008).
In most fungi, the tendency to produce large ascospores goesalong with either the formation of a thickened outer wall or thecompartimentation of the ascospores by three-dimensional celldivision, resulting in ‘muriform’ ascospores. Muriform cell divisionstabilizes the ascospores against both internal (turgescence) andexternal mechanical stress and thus maintains their shape, evenif the septa are thin with little investment of wall material. Thiscorresponds to the concept of ‘tensegrity’ as a means to stabilizestructures by minimal structural investment, such as in geodesicdomes (Buckminster Fuller, 1961; Pugh, 1976; Ingber, 1998;Gómez Jáuregui and Valentín, 2010). The reason for the tendencyto produce such large ascospores could be the need for an in-creased period of survival in order to reassociate with an appropri-ate photobiont, and large ascospores can store more nutrients.Invertebrates that feed on ascospores potentially serve as dispers-ers, particularly psocids and ants (McCarthy and Healy, 1978;Seaward, 1988; Lorentsson and Mattsson, 1999; Lücking andBernecker-Lücking, 2000). Many ants in tropical rainforest live insymbioses with plants, which provide shelter and food in formof specialized, minute structures formed on the leaf petioles, suchas the so-called Beltian and Müllerian bodies in Acacia and Cecro-pia plants (Rickson, 1969, 1976; Eubanks et al., 1997; Herrera andOlle, 2002; Meehan et al., 2009). Large ascospores resemble thesebodies in appearance and size, as they can reach sizes of up to0.7 � 0.07 mm, e.g. in Ocellularia subpraestans, and hence mightappear attractive to ants. Unfortunately, the role of symbiotic antsin ascospore dispersal of tropical lichens has not been studied sofar.
4.5. Parallel evolution of secondary chemistry in Graphidaceae
The presence of secondary substances in lichenized fungi is verywell-documented with over 1000 different substances identified(Huneck and Yoshimura, 1996; Lumbsch, 2002; Nash, 2008). Eco-physiological studies suggest that they may serve as sun screens,protection against fungivores and herbivores, and to control CO2
diffusion in the lichen thallus due to their often hydrophobic nat-ure (Lawrey, 1983a,b; Golojuch and Lawrey, 1988; Rikkinen,1995; Fernández et al., 1996; Proksch and Hesbacher, 1997;Solhaug et al., 2003; Gauslaa and McEnvoy, 2005; Asplund et al.,2009; Boehm et al., 2009). Flower colors in vascular plants andespecially their patterns in UV light are correlated to attraction
56 E. Rivas Plata, H.T. Lumbsch /Molecular Phylogenetics and Evolution 61 (2011) 45–63

by pollinators. For lichens and fungi, no such evidence is available,although many species produce pigmented ascomata. Apart fromsun lichens in which pigments serve as sunscreens, such as Telos-chistaceae and the genera Haematomma and Ramboldia, Graphida-ceae contains the highest number of shade species that producemedullary and/or apothecial pigments in any known group of lich-enized Ascomycota. This is intriguing but at present, any possiblecorrelation with potential ascospore dispersal by biotic vectorscan only be speculative.
4.6. Parallel evolution versus plesiomorphy
When assessing potential cases of parallel evolution, it isimportant to distinguish evidence for plesiomorphy. For example,crustose species in the collective genera Buellia and Rinodina(Physciaceae) or Verrucaria (Verrucariaceae) have been shownto be highly polyphyletic (Grube and Arup, 2001; Wedin et al.,2002; Helms et al., 2003; Cubero et al., 2004; Lohtander et al.,2008; Gueidan et al., 2009; Nadyeina et al., 2010). In these cases,it appears that plesiomorphic morphotypes have been retained inmany lineages. In the Graphidaceae, the basal plesiomorphic typeappears to be myriotremoid apothecia and thalli, explaining theoccurrence of such types in at least four of the five major cladeswithin the family. Within the Ocellularia clade, small ocellularioidascomata with columella and carbonization appear plesiomor-phic, explaining the occurrence of such forms dispersed overthe clade whereas other morphotypes appear derived and oftenmonophyletic. Cases of convergent evolution of thallus or ascomatypes within Graphidaceae are not evident at first glance, sincethe underlying structures are homologous in most cases, butone example can be mentioned: whereas in Gyrotrema (Ocellular-ia clade) the red-pigmented disc represents the hymenial surface,in Ocellularia aff. cruentata and Acanthothecis sanguinoloba (notincluded in this study), the red pigment is formed on the
exposed outer layer of the excipulum that covers the(unpigmented) disc.
Documenting cases of parallel evolution and phenotypicdivergence helps to identify model cases that can be used in ad-vanced studies focused on the evolutionary mechanisms andconstraints that might lead to such cases. With the enormousdiversity of Fungi, it is important that many more taxa bestudied in detail, to unravel the diversity of evolutionarymechanisms at work in this kingdom. Such studies are also nec-essary to evaluate current classifications, as they shed new lighton the taxonomic importance of characters and charactercomplexes.
Acknowledgments
Material from which new sequences were obtained was col-lected in the frame of the NSF-funded projects Ticolichen (DEB0206125 to The Field Museum; PI R. Lücking), Phylogeny and Tax-onomy of Ostropalean Fungi, with Emphasis on the Lichen-formingThelotremataceae (DEB 0516116 to The Field Museum; PI H.T.Lumbsch; Co-PI R. Lücking), and Neotropical Epiphytic Microli-chens – An Innovative Inventory of a Highly Diverse yet LittleKnown Group of Symbiotic Organisms (DEB 715660 to The FieldMuseum; PI R. Lücking). We would like to thank K. Papong (Thai-land) and M. Cáceres (Brazil) for arranging fieldwork and allow ac-cess to personal collections. We are indebted to R. Lücking and M.Nelsen for their discussion and suggestions. All the lab work wasperformed in the Pritzker Laboratory of Molecular Systematics atthe Field Museum (USA).
Appendix A
See Table A1.
Table A1Binary morphological data matrix used in the analysis (interleaved).
Species 0 0 0 0 0 0 0 0 0 1 1 1 1 1 1 1 1 1 1 2 2 2 2 2 2 2 2 2 2 3 3 31 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2
Ramonia valenzueliana 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 1Belonia russula 0 0 0 1 0 0 0 0 0 0 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 1 1 0 0 0 0Coenogonium luteum 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0Coenogonium pineti 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0Gyalecta hypoleuca 0 0 0 1 0 0 0 0 0 0 1 0 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1Gyalecta jenensis 0 0 0 1 0 0 0 0 0 0 1 0 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1Gyalecta ulmi 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1Porina aenea 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 1 1 0 0 0 0 0 0 0 0 1 1 1 0 0 0 0Porina epiphylla 0 0 1 0 0 0 0 1 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 0 1 1 1 0 0 0 0Porina limbulata 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 1 1 0 0 0 0Porina nitidula 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 1 1 1 0 0 0 0 0 0 0 1 1 1 0 0 0 0Porina nucula 1 0 0 0 1 0 0 1 0 0 1 0 0 0 0 1 1 1 0 0 0 0 0 0 0 1 1 1 0 0 0 0Acanthothecis peplophora 1 0 0 0 0 0 0 1 0 0 0 0 0 1 0 1 0 0 1 1 1 1 0 0 0 1 1 1 0 0 0 0Ampliotrema amplius 1 0 0 0 0 0 0 1 0 0 0 0 1 0 0 1 1 1 0 0 0 0 0 0 0 1 0 0 1 0 0 0Carbacanthographis chionophora 1 0 0 0 0 0 0 1 0 0 0 1 0 0 0 1 1 1 1 1 1 1 0 0 0 1 1 1 0 0 0 0Chapsa astroidea 1 0 0 0 0 0 0 1 0 0 0 0 0 1 0 1 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0Chapsa meridensis 0 1 0 0 0 0 0 1 0 0 0 0 0 1 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 1Chapsa pulchra 1 0 0 0 0 0 0 1 0 0 1 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 1 0 1 0Chroodiscus defectus 0 0 1 0 0 0 0 1 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Clandestinotrema stylothecium 1 0 0 0 0 0 0 1 0 0 1 0 0 0 0 1 0 0 1 0 0 0 0 0 0 1 0 0 1 0 0 1Cruentotrema thailandicum 1 0 0 0 0 0 0 0 0 0 0 0 0 1 1 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0Diorygma poitaei 1 0 0 0 0 0 0 1 0 0 1 0 0 0 0 1 0 0 1 1 1 1 0 0 0 1 1 0 1 0 0 0Dyplolabia afzelii 1 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 1 1 1 1 1 0 0 0 0 1 1 1 1 0 0 0Fissurina astroisidiata 1 0 0 0 0 0 1 1 0 0 0 0 0 1 1 1 0 0 1 1 1 1 1 0 0 1 1 0 0 0 0 1Fissurina nigromarginata 1 0 0 0 0 0 0 1 0 0 0 0 0 1 1 1 0 0 1 1 1 1 0 0 0 1 1 1 0 0 0 0Glyphis cicatricosa 1 0 0 0 0 0 0 1 0 0 0 0 0 1 0 0 0 0 1 1 0 0 0 1 0 0 0 0 1 1 0 0Glyphis scyphulifera 1 0 0 0 0 0 0 1 0 0 0 0 0 1 0 1 1 1 0 0 0 0 0 0 0 0 0 0 1 1 0 0Glyphis substriatula 1 0 0 0 0 0 0 1 0 0 0 0 0 1 0 1 1 1 1 1 1 1 0 0 0 1 1 1 1 1 0 0
(continued on next page)
E. Rivas Plata, H.T. Lumbsch /Molecular Phylogenetics and Evolution 61 (2011) 45–63 57

Table A1 (continued)
Species 0 0 0 0 0 0 0 0 0 1 1 1 1 1 1 1 1 1 1 2 2 2 2 2 2 2 2 2 2 3 3 31 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2
Graphis illinata 1 0 0 0 0 0 0 1 0 0 0 0 0 1 0 1 1 0 1 1 1 1 0 0 0 1 1 1 0 0 0 0Graphis librata 1 0 0 0 0 0 0 1 0 0 0 0 0 1 0 1 0 0 1 1 1 1 0 0 0 1 1 1 0 0 0 0Gyrotrema wirthii 1 0 0 0 0 0 0 1 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Halegrapha chimera 1 0 0 0 0 0 0 1 0 0 0 0 0 1 0 1 1 0 1 1 1 1 0 0 0 1 1 1 0 0 0 0Hemithecium chlorocarpum 1 0 0 0 0 0 0 1 0 0 0 0 0 1 0 1 1 0 1 1 1 1 0 0 0 1 1 1 0 0 0 0Hemithecium implicatum 1 0 0 0 0 0 0 1 0 0 0 0 0 1 0 1 1 0 1 1 1 1 0 0 0 1 1 1 0 0 0 0Leptotrema wightii 1 0 0 0 1 1 0 1 1 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0Leucodecton compunctellum 1 0 0 0 1 0 0 1 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 1 0 0 0 0Leucodecton occultum 1 0 0 0 0 0 0 1 0 0 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0Leucodecton phaeosporum 1 0 0 0 1 1 0 1 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 1 0 0 0 0Melanotrema columellatum 1 0 0 0 0 0 0 1 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 1Melanotrema endomelaenum 1 0 0 0 0 0 0 1 0 0 1 0 0 0 0 1 0 0 1 0 0 0 0 0 0 1 0 0 1 0 0 1Melanotrema meiospermum 1 0 0 0 0 0 0 1 0 0 1 0 0 0 0 1 0 0 1 0 0 0 0 0 0 1 0 0 1 0 0 1Myriotrema clandestinum 1 0 0 0 0 0 0 1 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 1 1 0 1 0 0 0Myriotrema laeviusculum 1 0 0 0 1 1 0 1 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 1 1 0 1 0 0 0Myriotrema microporum 1 0 0 0 0 0 0 1 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 1 1 0 1 0 0 1Myriotrema peninsulae 1 0 0 0 0 0 0 1 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 1Myriotrema pycnoporellum 1 0 0 0 0 0 0 1 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 1 1 0 1 0 0 0Nadvornikia hawaiiensis 1 0 0 0 0 0 0 1 0 0 1 0 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1Ocellularia bahiana 1 0 0 0 1 0 0 1 1 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0Ocellularia diacida 1 0 0 0 0 0 0 1 0 0 0 0 1 0 0 1 0 0 1 0 0 0 0 0 0 1 1 0 1 0 0 0Ocellularia obturascens 1 0 0 0 1 0 0 1 1 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0Ocellularia papillata 1 0 0 0 0 0 0 1 0 0 0 0 0 1 1 1 0 0 1 0 0 0 0 0 0 1 1 0 1 0 0 0Ocellularia profunda 1 0 0 0 1 0 0 1 1 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 1 1 0 1 0 0 0Ocellularia terebrata 1 0 0 0 0 0 0 1 0 0 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 1 1 0 1 0 0 0Pallidogramme chlorocarpoides 1 0 0 0 0 0 0 1 0 0 0 0 0 1 0 1 1 0 1 1 1 1 0 0 0 1 1 1 0 0 0 0Phaeographis dendritica 1 0 0 0 0 0 0 1 0 0 1 0 0 0 0 1 0 0 1 1 1 1 0 0 0 1 0 0 1 1 0 1Phaeographis intricans 1 0 0 0 0 0 0 1 0 0 0 0 0 1 0 0 0 0 1 1 1 1 1 1 0 0 0 0 1 0 0 0Platythecium sphaerosporellum 1 0 0 0 0 0 0 1 0 0 0 0 0 1 1 1 0 0 1 1 1 1 0 0 0 1 1 0 0 0 0 1Redingeria glaucoglyphica 1 0 0 0 0 0 0 1 0 0 1 0 0 0 0 1 0 0 1 1 0 0 0 0 0 1 1 0 1 0 0 1Reimnitzia santensis 1 0 0 0 1 1 1 1 1 0 1 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0Sarcographa labyrinthica 1 0 0 0 0 0 0 1 0 0 0 0 0 1 0 0 0 0 1 1 0 1 1 1 1 0 0 0 1 0 0 0Schistophoron tenue 1 0 0 0 0 0 0 1 0 0 1 0 0 0 0 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0Stegobolus subemersus 1 0 0 0 0 0 0 1 0 0 0 0 0 1 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 1Thalloloma hypoleptum 1 0 0 0 0 0 0 1 0 0 1 0 0 0 0 1 0 0 1 1 1 1 0 0 0 1 0 0 1 1 0 1Thecaria quassiicola 1 0 0 0 0 0 0 1 0 0 0 0 0 1 0 1 1 1 1 1 1 1 0 0 0 1 0 0 1 0 0 0Thelotrema bicinctulum 1 0 0 0 0 0 0 1 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 1Thelotrema diplotrema 1 0 0 0 0 0 0 1 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 1Thelotrema expallescens 1 0 0 0 0 0 0 1 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0Thelotrema lepadinum 1 0 0 0 0 0 0 1 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 0 1 0 0 1 0 0 1Thelotrema porinaceum 1 0 0 0 0 0 0 1 0 0 1 0 0 0 0 1 0 0 1 0 0 0 0 0 0 1 1 0 0 0 0 0Topeliopsis muscigena 0 1 0 0 0 0 0 1 0 0 0 0 0 1 0 1 1 1 1 0 0 0 0 0 0 1 0 0 1 0 0 1Wirthiotrema glaucopallens 1 0 0 0 0 0 0 1 0 0 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 1 1 0 1 0 0 0
Species 3 3 3 3 3 3 3 4 4 4 4 4 4 4 4 4 4 5 5 5 5 5 5 5 5 5 5 6 6 6 6 63 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2 3 4
Ramonia valenzueliana 0 0 0 1 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 1 1 0 1 1 0 0 0 1 0 0 0 0Belonia russula 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0Coenogonium luteum 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0Coenogonium pineti 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 1 0Gyalecta hypoleuca 0 0 0 1 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 1 0Gyalecta jenensis 0 0 0 1 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 1 0Gyalecta ulmi 0 0 0 1 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 1 0Porina aenea 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 1 0 0 0 0 0 0 1 0Porina epiphylla 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 1 0Porina limbulata 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 1 0Porina nitidula 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 1 0 0 0 0 0 0 1 0Porina nucula 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 1 0Acanthothecis peplophora 0 1 0 1 1 0 1 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 1 0Ampliotrema amplius 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 0 0 1Carbacanthographis chionophora 0 0 0 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 1 0 1 1 0 0 0 0 0 0 0Chapsa astroidea 1 0 0 1 0 0 1 1 0 0 1 1 0 0 0 0 0 0 0 1 0 0 1 1 0 0 0 0 0 1 0 0Chapsa meridensis 0 0 0 1 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 1 1 0 1 1 0 0 0 0 0 0 0 0Chapsa pulchra 1 0 0 1 0 0 1 1 0 0 1 1 0 0 0 0 0 0 0 1 0 0 1 1 0 0 0 0 0 1 0 0Chroodiscus defectus 1 0 0 1 0 0 1 1 0 0 1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 1 0Clandestinotrema stylothecium 0 0 0 0 0 0 1 0 0 0 1 1 1 1 1 1 1 1 0 0 1 1 0 0 0 0 0 0 0 0 0 0Cruentotrema thailandicum 1 0 0 1 0 0 1 1 0 0 1 1 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0Diorygma poitaei 0 0 0 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 1Dyplolabia afzelii 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 1Fissurina astroisidiata 0 0 0 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Fissurina nigromarginata 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0Glyphis cicatricosa 0 0 1 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 1 0 0 0 1 1 0 0 0 0 1Glyphis scyphulifera 0 0 0 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 1 0 0 0 1 1 0 0 0 0 1Glyphis substriatula 0 1 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 1 1 0 0 0 0 1Graphis illinata 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 1 0 1 0
58 E. Rivas Plata, H.T. Lumbsch /Molecular Phylogenetics and Evolution 61 (2011) 45–63

Table A1 (continued)
Species 0 0 0 0 0 0 0 0 0 1 1 1 1 1 1 1 1 1 1 2 2 2 2 2 2 2 2 2 2 3 3 31 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2
Graphis librata 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0Gyrotrema wirthii 1 0 0 1 0 0 1 1 0 0 1 1 0 0 0 0 0 0 1 0 1 1 0 0 0 0 0 0 0 0 0 0Halegrapha chimera 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0Hemithecium chlorocarpum 0 1 0 1 1 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 1 0Hemithecium implicatum 0 1 0 1 1 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Leptotrema wightii 0 0 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 1Leucodecton compunctellum 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0Leucodecton occultum 0 0 1 1 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 1 1 0 1 0 0 0 0 0 0 0 0 0Leucodecton phaeosporum 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0Melanotrema columellatum 0 0 1 0 0 0 1 0 0 0 1 1 1 1 1 1 1 1 0 0 1 1 0 0 0 0 0 0 0 0 0 0Melanotrema endomelaenum 0 0 1 0 0 0 1 0 0 0 1 1 1 1 1 1 1 1 0 0 1 1 0 0 0 0 0 0 0 0 0 0Melanotrema meiospermum 0 0 0 0 0 0 1 0 0 0 1 1 1 1 1 1 1 1 0 0 1 1 0 0 0 0 0 0 0 0 0 0Myriotrema clandestinum 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Myriotrema laeviusculum 0 0 0 0 0 0 0 0 0 0 1 1 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Myriotrema microporum 0 0 1 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0 0Myriotrema peninsulae 0 0 1 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Myriotrema pycnoporellum 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0Nadvornikia hawaiiensis 0 0 0 1 1 0 1 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Ocellularia bahiana 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0 0Ocellularia diacida 0 0 0 0 0 0 0 0 1 1 1 1 1 1 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0Ocellularia obturascens 0 0 0 0 0 0 0 0 0 0 1 1 1 0 1 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0 0Ocellularia papillata 0 0 0 0 0 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0Ocellularia profunda 0 0 0 0 0 0 0 0 0 0 1 1 1 1 0 0 1 1 0 0 1 1 0 0 0 0 0 0 0 0 0 0Ocellularia terebrata 0 0 0 0 0 0 0 0 0 0 1 1 1 1 1 0 1 1 0 0 1 1 0 0 0 0 0 0 0 0 0 0Pallidogramme chlorocarpoides 0 1 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Phaeographis dendritica 0 0 0 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 1 0 0 0 0 0 0Phaeographis intricans 0 0 0 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0Platythecium sphaerosporellum 0 0 0 1 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Redingeria glaucoglyphica 0 0 0 1 0 0 1 0 0 0 1 1 1 1 1 0 1 1 0 0 1 1 0 0 0 0 0 0 0 0 0 0Reimnitzia santensis 1 0 0 1 0 0 1 1 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 1 1Sarcographa labyrinthica 0 0 0 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 1 0 0 0 1 0 0 0 0 0 0Schistophoron tenue 0 0 0 1 1 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0Stegobolus subemersus 0 0 0 0 0 0 1 0 0 0 1 1 1 1 1 1 1 1 0 0 1 1 0 0 0 0 0 0 0 0 0 0Thalloloma hypoleptum 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0 0Thecaria quassiicola 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 1 1 0 0 0 1 0 0 0 0 0 0Thelotrema bicinctulum 0 0 1 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 1 0 0 1 1 0 0 0 0 0 0 0 0Thelotrema diplotrema 0 0 1 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 1 0 0 1 1 0 0 0 0 0 0 0 0Thelotrema expallescens 0 0 1 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 1 0 0 1 1 0 0 0 0 0 0 0 0Thelotrema lepadinum 0 0 1 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 1 0 0 1 1 0 0 0 0 0 0 0 0Thelotrema porinaceum 0 0 1 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 1 0 0 1 1 0 0 0 0 0 0 0 0Topeliopsis muscigena 0 0 0 0 0 0 1 0 0 0 1 0 0 0 0 0 0 0 0 1 0 0 1 1 0 0 0 0 0 0 0 0Wirthiotrema glaucopallens 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 1 1 0 1 0 0 0 0 0 0 0 0 0
Species 6 6 6 6 6 7 7 7 7 7 7 7 7 7 7 8 8 8 8 8 8 8 8 8 8 9 9 9 9 9 9 95 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6
Ramonia valenzueliana 0 0 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 1 0Belonia russula 0 0 0 0 1 0 0 0 0 1 0 0 0 0 0 1 1 1 1 0 0 1 1 1 1 1 0 0 0 0 1 0Coenogonium luteum 0 0 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0Coenogonium pineti 0 0 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0Gyalecta hypoleuca 0 0 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 1 0Gyalecta jenensis 0 0 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 1 0Gyalecta ulmi 0 0 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 1 0Porina aenea 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0Porina epiphylla 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 1 0Porina limbulata 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0Porina nitidula 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 1 0Porina nucula 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 1 1 0 1 0 0 1 1 0 0 0 0 1 0Acanthothecis peplophora 0 0 1 1 0 1 1 1 1 0 0 0 0 0 0 1 1 0 1 1 1 1 0 0 0 0 0 1 0 0 0 0Ampliotrema amplius 0 0 0 0 1 1 1 1 1 0 0 0 0 0 0 1 0 0 1 0 0 1 0 0 0 0 0 1 1 0 0 0Carbacanthographis chionophora 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 1 1 0 0 0Chapsa astroidea 0 0 1 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 1 0 0 0 0Chapsa meridensis 0 0 0 0 1 1 1 1 1 0 0 0 1 1 1 1 1 1 1 1 1 1 0 0 0 0 0 1 0 0 0 0Chapsa pulchra 0 0 1 0 0 1 1 1 1 0 0 0 0 0 0 1 1 0 1 0 0 1 1 0 1 0 0 1 0 0 0 0Chroodiscus defectus 0 0 1 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0Clandestinotrema stylothecium 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 1 1 1 0 0Cruentotrema thailandicum 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 1 1 1 0 0Diorygma poitaei 1 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 1 0 1 1 0 1 0 0 0 0 0 1 0 0 0 0Dyplolabia afzelii 1 1 0 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 1 1 1 0 0Fissurina astroisidiata 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 1 1 0 0 0Fissurina nigromarginata 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 1 1 0 0 0Glyphis cicatricosa 1 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 1 0 1 0 0 1 1 0 0 0 1 1 1 0 0 0Glyphis scyphulifera 1 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 0 0 1 1 0 1 0 0 0 0 0 1 1 0 0 0Glyphis substriatula 1 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 0 0 1 1 0 1 0 0 0 0 0 1 1 0 0 0Graphis illinata 0 0 0 0 0 1 1 1 1 0 0 0 1 1 1 1 1 1 1 1 1 1 1 0 0 0 0 1 1 0 0 0
(continued on next page)
E. Rivas Plata, H.T. Lumbsch /Molecular Phylogenetics and Evolution 61 (2011) 45–63 59

Table A1 (continued)
Species 0 0 0 0 0 0 0 0 0 1 1 1 1 1 1 1 1 1 1 2 2 2 2 2 2 2 2 2 2 3 3 31 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2
Graphis librata 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 0 0 1 0 0 1 1 0 1 1 0 1 1 0 0 0Gyrotrema wirthii 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 0 0 1 0 0 1 0 0 0 0 0 1 1 0 0 0Halegrapha chimera 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 0 0 1 0 0 1 0 0 0 0 0 1 1 0 0 0Hemithecium chlorocarpum 0 0 0 0 0 1 1 1 1 0 0 0 1 1 1 1 1 1 1 1 1 1 0 0 0 0 0 1 1 0 0 0Hemithecium implicatum 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 0 0 1 0 0 1 1 0 0 0 0 1 1 0 0 0Leptotrema wightii 0 1 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0 0 1 1 0 1 0 0 0 0 0 1 1 0 0 0Leucodecton compunctellum 0 0 0 0 0 1 1 1 1 0 0 0 1 0 0 1 1 1 1 1 1 1 0 0 0 0 0 1 0 0 0 0Leucodecton occultum 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 0 0 1 1 0 1 0 0 0 0 0 1 1 0 0 0Leucodecton phaeosporum 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 0 0 1 1 0 0 0 0 0 0 0 1 1 0 0 0Melanotrema columellatum 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 1 1 0 0 0Melanotrema endomelaenum 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 0 0 1 0 0 1 0 0 1 1 1 1 1 0 0 0Melanotrema meiospermum 0 0 1 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 1 1 0 0 0Myriotrema clandestinum 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 1 1 0 0 0Myriotrema laeviusculum 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 1 1 0 0 0Myriotrema microporum 0 0 1 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 1 1 0 0 0Myriotrema peninsulae 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 1 0 1 1 1 0 0 0Myriotrema pycnoporellum 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 1 1 0 0 0Nadvornikia hawaiiensis 0 0 0 0 0 1 1 1 1 0 1 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 1 0 0 0Ocellularia bahiana 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 0 0 1 1 0 1 0 0 0 0 0 1 1 0 0 0Ocellularia diacida 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 0 0 1 0 0 1 1 0 1 1 0 1 1 0 0 0Ocellularia obturascens 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 1 1 0 0 0Ocellularia papillata 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 0 0 1 0 0 1 0 0 0 0 0 1 1 0 0 0Ocellularia profunda 0 0 0 0 0 1 1 1 1 0 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1 0 0 1 0 0 0 0Ocellularia terebrata 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 0 0 1 1 0 1 0 0 0 0 0 1 1 0 0 0Pallidogramme chlorocarpoides 0 0 0 0 1 1 1 1 1 0 0 0 1 0 0 1 1 0 1 1 1 1 0 0 0 0 0 1 1 0 0 0Phaeographis dendritica 0 0 0 0 1 1 1 1 1 0 0 0 0 0 0 1 0 0 1 0 0 1 1 0 0 0 1 1 1 0 0 0Phaeographis intricans 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 1 1 0 0 0Platythecium sphaerosporellum 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 1 0 0 0Redingeria glaucoglyphica 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 1 1 0 0 0Reimnitzia santensis 0 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 1 1 0 0 0Sarcographa labyrinthica 0 0 0 0 1 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 1 1 0 0 0Schistophoron tenue 0 0 0 0 0 1 1 1 1 0 1 1 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 1 0 0 0 0Stegobolus subemersus 0 0 1 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 1 1 0 0 0Thalloloma hypoleptum 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 0 0 1 0 0 1 0 0 0 0 0 1 1 0 0 0Thecaria quassiicola 0 0 0 0 1 1 1 1 1 0 0 0 0 0 0 1 1 0 1 1 1 1 1 0 0 0 0 1 1 0 0 0Thelotrema bicinctulum 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 0 0 1 0 0 1 1 0 0 0 1 1 1 0 0 0Thelotrema diplotrema 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 1 0 1 1 0 1 1 0 1 1 1 1 1 0 0 1Thelotrema expallescens 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 1 0 0 0 0Thelotrema lepadinum 0 0 0 0 0 1 1 1 1 0 0 0 1 0 0 1 1 1 1 1 1 1 1 0 0 0 0 1 1 0 0 1Thelotrema porinaceum 0 0 0 0 0 1 1 1 1 0 0 0 1 1 0 1 1 1 1 1 1 1 1 0 1 0 0 1 0 0 0 0Topeliopsis muscigena 0 0 0 0 0 1 1 1 1 0 0 0 1 1 1 1 1 1 1 1 1 1 0 0 0 0 0 1 0 0 0 0Wirthiotrema glaucopallens 0 0 0 0 0 1 1 1 1 0 0 0 0 0 0 1 0 0 1 1 0 1 0 0 0 0 0 1 0 0 0 0
Species 0 0 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 19 9 9 0 0 0 0 0 0 0 0 0 0 1 1 1 1 1 1 1 1 1 1 2 2 2 2 2 2 2 2 27 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8
Ramonia valenzueliana 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Belonia russula 0 0 0 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Coenogonium luteum 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Coenogonium pineti 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Gyalecta hypoleuca 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Gyalecta jenensis 0 0 0 0 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Gyalecta ulmi 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Porina aenea 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Porina epiphylla 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Porina limbulata 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Porina nitidula 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Porina nucula 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Acanthothecis peplophora 0 0 0 0 0 0 1 1 1 1 1 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Ampliotrema amplius 0 0 0 1 1 0 1 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0Carbacanthographis chionophora 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 1 0 0 0 0Chapsa astroidea 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Chapsa meridensis 0 0 0 0 0 0 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Chapsa pulchra 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Chroodiscus defectus 0 0 0 0 0 0 1 0 0 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Clandestinotrema stylothecium 0 0 0 0 0 0 1 0 0 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Cruentotrema thailandicum 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1Diorygma poitaei 0 0 0 1 1 0 1 1 1 1 1 1 0 0 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0Dyplolabia afzelii 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Fissurina astroisidiata 0 0 0 1 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Fissurina nigromarginata 0 0 0 1 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Glyphis cicatricosa 0 0 0 1 1 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Glyphis scyphulifera 0 0 0 1 1 0 1 1 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Glyphis substriatula 0 0 0 1 1 0 1 1 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Graphis illinata 0 0 0 1 1 0 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
60 E. Rivas Plata, H.T. Lumbsch /Molecular Phylogenetics and Evolution 61 (2011) 45–63

References
Aptroot, A., Lücking, R., Sipman, H.J.M., Umaña, L., Chaves, J.L., 2008. Pyrenocarpouslichens with bitunicate asci: a first assessment of the lichen biodiversityinventory in Costa Rica. Biblioth. Lichenol. 97, 1–162.
Asplund, J., Solhaug, K.A., Gauslaa, Y., 2009. Fungal depsidones – an inducible orconstitutive defence against herbivores in the lichen Lobaria pulmonaria? BasicAppl. Ecol. 10, 273–278.
Attenborough, D., 1995. The Private Life of Plants: A Natural History of PlantBehavior. BBC Books, London.
Awasthi, D.D., 1983. Reproduction in lichens. Phytomorphology 33, 26–30.Baloch, E., Lücking, R., Lumbsch, H.T., Wedin, M., 2010. Major clades and
phylogenetic relationships between lichenized and non-lichenized lineages inOstropales (Ascomycota: Lecanoromycetes). Taxon 59, 1483–1494.
Barton, N.H., Briggs, D.E.G., Eisen, J.A., Goldstein, D.B., Patel, N.H., 2007. Evolution.Harbor Laboratory Press, Cold Spring.
Berbee, M.L., 1996. Loculoascomycete origins and evolution of filamentousascomycete morphology based on 18S rRNA gene sequence data. Mol. Biol.Evol. 13, 462–470.
Berbee, M.L., Taylor, J.W., 1992. Convergence in ascospore discharge mechanismamong pyrenomycete fungi based on 18S ribosomal RNA gene sequence. Mol.Phylogenet. Evol. 1, 59–71.
Blanco, O., Crespo, A., Divakar, P.K., Esslinger, T.L., Hawksworth, D.L., Lumbsch, H.T.,2004a. Melanelixia and Melanohalea, two new genera segregated from Melanelia(Parmeliaceae) based on molecular and morphological data. Mycol. Res. 108,873–884.
Blanco, O., Crespo, A., Elix, J.A., Hawksworth, D.L., Lumbsch, H.T., 2004b. A molecularphylogeny and a new classification of parmelioid lichens containingXanthoparmelia-type lichenan (Ascomycota: Lecanorales). Taxon 53, 959–975.
Boehm, F., Clarke, K., Edge, R., Fernández, E., Navaratnam, S., Quilhot, W., Rancan, F.,Truscott, T.G., 2009. Lichens-photophysical studies of potential new sunscreens.J. Photochem. Photobiol. B: Biol. 9, 40–45.
Buckminster Fuller, R., 1961. Tensegrity. Portfolio Art News Ann. 4, 112–127, 144,148.
Crespo, A., Ferencova, Z., Perez-Ortega, S., Elix, J.A., Divakar, P.D., 2010.Austroparmelina, a new Australasian lineage in parmelioid lichens(Parmeliaceae, Ascomycota). Syst. Biodivers., 209–221.
Cubero, O.F., Crespo, A., Esslinger, T.L., Lumbsch, H.T., 2004. Molecular phylogeny ofthe genus Physconia (Ascomycota, Lecanorales) inferred from a Bayesiananalysis of nuclear ITS rDNA sequences. Mycol. Res. 108, 498–505.
Escrig, F., Cobreros, A., Valcarcel, J.P., 1997. The elliptical dome. A survey ofconstructive techniques to stabilize a sopisticated structure. Adv. Architect. Ser.3, 355–364.
Eubanks, M.D., Kimberly, A., Nesci Mette, K., Petersen, Z.L., Horacio Bonfil, Sanchez,1997. The exploitation of an ant-defended host plant by a shelter-buildingherbivore. Oecologia 109, 454–460.
Fernández, E., Quilhot, W., González, I., Hidalgo, M.E., Molina, X., Meneses, I., 1996.Lichen metabolites as UVB filters. Cosm. Toil. 111, 69–74.
Frisch, A., Kalb, K., Grube, M., 2006. Contributions towards a new systematics of thelichen family Thelotremataceae III. Molecular phylogeny of theThelotremataceae. Bibl. Lichenol. 92, 517–539.
Futuyma, D.J., 2005. Evolution. Sinauer Associates, Sunderland.Gargas, A., Taylor, J.W., 1995. Phylogeny of Discomycetes and early radiations of the
the apothecial Ascomycotina inferred from SSU rDNA sequence data. Exper.Mycol. 19, 7–15.
Gauslaa, Y., McEvoy, M., 2005. Seasonal changes in solar radiation drive acclimationof the sun-screening compound parietin in the lichen Xanthoria parietina. BasicAppl. Ecol. 6, 75–82.
Golojuch, S.T., Lawrey, J.D., 1988. Quantitative variation in vulpinic and pinastricacids produced by Tuckermannopsis pinastri (lichen-forming Ascomycotina,Parmeliaceae). Am. J. Bot. 75, 1871–1875.
Gómez Jáuregui, V., 2010. Tensegrity Structures and their Application toArchitecture. Servicio de Publicaciones de la Universidad de Cantabria,Santander.
Table A1 (continued)
Species 0 0 0 0 0 0 0 0 0 1 1 1 1 1 1 1 1 1 1 2 2 2 2 2 2 2 2 2 2 3 3 31 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2 3 4 5 6 7 8 9 0 1 2
Graphis librata 0 0 0 1 1 0 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Gyrotrema wirthii 0 0 0 1 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0Halegrapha chimera 1 0 0 1 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Hemithecium chlorocarpum 0 0 0 1 1 0 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Hemithecium implicatum 0 0 0 1 1 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Leptotrema wightii 1 1 1 1 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1Leucodecton compunctellum 1 1 0 1 0 0 1 1 1 1 1 1 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0Leucodecton occultum 1 1 0 1 0 0 1 0 0 1 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Leucodecton phaeosporum 1 1 1 1 0 0 1 0 0 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Melanotrema columellatum 1 1 0 1 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Melanotrema endomelaenum 0 0 0 1 1 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0Melanotrema meiospermum 1 1 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Myriotrema clandestinum 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0Myriotrema laeviusculum 1 1 0 1 0 0 1 0 0 1 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0Myriotrema microporum 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0Myriotrema peninsulae 0 0 0 0 0 0 1 0 0 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Myriotrema pycnoporellum 0 0 0 1 1 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Nadvornikia hawaiiensis 1 1 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0Ocellularia bahiana 1 1 0 1 0 0 1 0 0 1 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0Ocellularia diacida 0 0 0 1 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 1 0 0 1Ocellularia obturascens 1 1 0 1 0 0 1 0 0 1 1 0 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0Ocellularia papillata 0 0 0 1 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Ocellularia profunda 0 0 0 1 1 0 1 1 1 1 1 1 0 0 0 0 0 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0Ocellularia terebrata 0 0 0 1 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0Pallidogramme chlorocarpoides 0 0 0 1 1 0 1 1 1 1 1 1 0 0 1 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0Phaeographis dendritica 1 0 0 1 0 1 1 1 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Phaeographis intricans 1 0 0 1 0 1 1 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Platythecium sphaerosporellum 0 0 0 1 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 0 0Redingeria glaucoglyphica 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Reimnitzia santensis 1 1 1 1 0 0 1 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Sarcographa labyrinthica 1 0 0 1 0 1 1 0 0 0 0 0 0 0 1 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0Schistophoron tenue 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Stegobolus subemersus 0 0 0 1 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 1 0 0 0 0 0 0Thalloloma hypoleptum 0 0 0 1 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Thecaria quassiicola 1 0 0 1 0 1 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Thelotrema bicinctulum 0 0 0 1 1 0 1 0 0 0 0 0 0 0 1 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0Thelotrema diplotrema 0 0 0 1 1 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Thelotrema expallescens 0 0 0 0 0 0 1 0 0 1 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Thelotrema lepadinum 0 0 0 1 1 0 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Thelotrema porinaceum 0 0 0 0 0 0 1 1 1 1 1 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Topeliopsis muscigena 0 0 0 1 1 0 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0Wirthiotrema glaucopallens 0 0 0 0 0 0 1 0 0 1 1 0 0 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
E. Rivas Plata, H.T. Lumbsch /Molecular Phylogenetics and Evolution 61 (2011) 45–63 61

Grube, M., 1998. Classification and phylogeny in the Arthoniales (lichenizedAscomycetes). Bryologist 101, 377–391.
Grube, M., Arup, U., 2001. Molecular and morphological evolution in thePhysciaceae (Lecanorales, lichenized Ascomycotina), with special emphasis onthe genus Rinodina. Lichenologist 33, 63–72.
Grube, M., Kantvilas, G., 2006. Siphula represents a remarkable case ofmorphological convergence in sterile lichens. Lichenologist 38, 241–249.
Grube, M., Baloch, E., Lumbsch, H.T., 2004. The phylogeny of Porinaceae(Ostropomycetidae) suggests a neotenic origin of perithecia inLecanoromycetes. Mycol. Res. 108, 1111–1118.
Gueidan, C., Savic, S., Thus, H., Roux, C., Keller, C., Tibell, L., Prieto, M., Heiðmarsson,S., Breuss, O., Orange, A., Froberg, L., Amtoft Wynns, A., Navarro-Rosine’s, P.,Krzewicka, B., Pykala, J., Grube, M., Lutzoni, F., 2009. Generic classification of theVerrucariaceae (Ascomycota) based on molecular and morphological evidence.Recent progress and remaining challenges. Taxon 58, 184–208.
Hale, M.E.Jr., 1983. The Biology of Lichens, third ed. Edward Arnold, Baltimore.Hall, T.A., 1999. BioEdit: a user-friendly biological sequence alignment editor
and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 41, 95–98.
Harris, R.C., 1989. A sketch of the family Pyrenulaceae (Melanommatales) in easternNorth America. Memoirs N.Y. Bot. Garden 49, 74–107.
Helms, G., Friedl, T., Rambold, G., 2003. Phylogenetic relationships of thePhysciaceae inferred from rDNA sequence data and selected phenotypiccharacters. Mycologia 95, 1078–1099.
Herrera, C.M., Olle, P., 2002. Plant–Animal Interactions: An Evolutionary Approach.Blackwell Publishing, Oxford.
Hibbett, D.S., Binder, M., Bischoff, J.F., Blackwell, M., Cannon, P.F., Eriksson, O.E.,Huhndorf, S., James, T., Kirk, P.M., Lucking, R., Lumbsch, H.T., Lutzoni, F.,Matheny, P.B., McLaughlin, D.J., Powell, M.J., Redhead, S., Schoch, C.L., Spatafora,J.W., Stalpers, J.A., Vilgalys, R., Aime, M.C., Aptroot, A., Bauer, R., Begerow, D.,Benny, G.L., Castlebury, L.A., Crous, P.W., Dai, Y.C., Gams, W., Geiser, D.M.,Griffith, G.W., Gueidan, C., Hawksworth, D.L., Hestmark, G., Hosaka, K., Humber,R.A., Hyde, K.D., Ironside, J.E., Koljalg, U., Kurtzman, C.P., Larsson, K.H.,Lichtwardt, R., Longcore, J., Miadlikowska, J., Miller, A., Moncalvo, J.M.,Mozley-Standridge, S., Oberwinkler, F., Parmasto, E., Reeb, V., Rogers, J.D.,Roux, C., Ryvarden, L., Sampaio, J.P., Schussler, A., Sugiyama, J., Thorn, R.G.,Tibell, L., Untereiner, W.A., Walker, C., Wang, Z., Weir, A., Weiss, M., White,M.M., Winka, K., Yao, Y.J., Zhang, N., 2007. A higher-level phylogeneticclassification of the Fungi. Mycol. Res. 111, 509–547.
Huelsenbeck, J.P., Ronquist, F., 2001. MRBAYES: Bayesian inference of phylogeny.Bioinformatics 17, 754–755.
Huneck, S., Yoshimura, I., 1996. Identification of Lichen Substances. Springer, Berlin.Ihlenfeldt, H.D., 1994. Diversification in an arid world: the Mesembryanthemaceae.
Ann. Rev. Ecol. Syst. 25, 521–546.Ingber, D.E., 1998. The architecture of life. Sci. Am. Mag. 278, 48–57.Katoh, K., Toh, M., 2005. MAFFT version 5: improvement in accuracy of multiple
sequence alignment. Nucl. Acids Res. 33, 511–518.Kauff, F., Lutzoni, F., 2002. Phylogeny of Gyalectales and Ostropales (Ascomycota,
Fungi): among and within order relationships based on nuclear ribosomal RNAsmall and large subunits. Mol. Phyl. Evol. 25, 138–156.
Kirk, P.M., Cannon, P.F., Minter, D.W., Stalpers, J.A., 2008. Dictionary of the Fungi,10th ed. CABI, Wallingford.
Lakatos, M., Rascher, U., Büdel, B., 2006. Functional characteristics of corticolouslichens in the understory of a tropical lowland rain forest. New Phytol. 172,679–695.
Larkin, M.A., Blackshields, G., Brown, N.P., Chenna, R., McGettigan, P.A., McWilliam,H., Valentin, F., 2007. ClustalW and ClustalX version 2. Bioinformatics 23, 2947–2948.
Larsson, E., Jeppson, M., 2008. Phylogenetic relationships among species and generaof Lycoperdaceae based on ITS and LSU sequence data from north Europeantaxa. Mycol. Res. 112, 4–22.
Lawrey, J.D., 1983a. Lichen herbivore preference. A test of two hypotheses. Am. J.Bot. 70, 1188–1194.
Lawrey, J.D., 1983b. Vulpinic and pinastric acids as lichen antiherbivorecompounds: contrary evidence. Bryologist 86, 365–369.
Lawrey, J.D., Lücking, R., Sipman, H.J.M., Chaves, J.L., Redhead, S.A., Bungartz, F.,Sikaroodi, M., Gillevet, P.M., 2009. High concentration of basidiolichens in asingle family of agaricoid mushrooms (Basidiomycota: Agaricales:Hygrophoraceae). Mycol. Res. 113, 1154–1171.
Leclere, L., Schuchert, P., Cruaud, C., Couloux, A., Manuel, M., 2009. Molecularphylogenetics of thecata (Hydrozoa, Cnidaria) reveals long-term maintenanceof life history traits despite high frequency of recent character changes. Syst.Biol. 58, 509–526.
Liu, Y.J., Whelen, S., Hall, B.D., 1999. Phylogenetic relationships amongascomycetes: evidence from the RNA polymerase II subunit. Mol. Biol. Evol.16, 1799–1808.
Lohtander, K., Ahti, T., Stenroos, S., Urbanavichus, G., 2008. Is Anaptychiamonophyletic? A phylogenetic study based on nuclear and mitochondrialgenes. Ann. Bot. Fennici 45, 55–60.
Lorentsson, S., Mattsson, J.-E., 1999. New reports of soredia dispersed by ants,Formica cunicularia. Lichenologist 31, 204–207.
Lücking, R., 2009. The taxonomy of the genus Graphis sensu Staiger (Ascomycota:Ostropales: Graphidaceae). Lichenologist 41, 319–362.
Lücking, R., Bernecker-Lücking, A., 2000. Lichen feeders and lichenicolous fungi: dothey affect dispersal and diversity in tropical foliicolous lichen communities?Ecotropica 6, 23–41.
Lücking, R., Chaves, J.L., Sipman, H.J.M., Umaña, L., Aptroot, A., 2008. A firstassessment of the Ticolichen Biodiversity Inventory in Costa Rica: the genusGraphis, with notes on the genus Hemithecium (Ascomycota: Ostropales:Graphidaceae). Fieldiana Bot., New Ser. 46, 1–126.
Lumbsch, H.T., 2002. Analysis of phenolic products in lichens for identification andtaxonomyc. In: Kranner, I., Beckett, R.P., Varma, A.K. (Eds.), Protocols inLichenology. Culturing, Biochemistry, Ecophysiology and Use inBiomonitoring. Springer, Berlin, Germany, pp. 281–295.
Lumbsch, H.T., Parnmen, S., Rangsiruji, A., Elix, J.A., 2010. Phenotypic disparity andadaptive radiation in the genus Cladia (Lecanorales, Ascomycota). Austral. Syst.Bot. 23, 239–247.
Lutzoni, F., Kauff, F., Cox, C., McLaughlin, D., Celio, G., Dentinger, B., Padamsee, M.,Hibbett, D., James, T.Y., Baloch, E., Grube, M., Reeb, V., Hofstetter, V., Schoch, C.,Arnold, A.E., Miadlikowska, J., Spatafora, J., Johnson, D., Hambleton, S., Crockett,M., Shoemaker, R., Sung, G.-H., Lücking, R., Lumbsch, H.T., O’Donnell, K., Binder,M., Diederich, P., Ertz, D., Gueidan, C., Hansen, K., Harris, R.C., Hosaka, K., Lim, Y.-W., Matheny, B., Nishida, H., Pfister, D., Rogers, J., Rossman, A., Schmitt, I.,Sipman, H., Stone, J., Sugiyama, J., Yahr, R., Vilgalys, R., 2004. Assembling thefungal tree of life: progress, classification, and evolution of subcellular traits.Am. J. Bot. 91, 1446–1480.
Maddison, W.P., Maddison, D.R., 2010. Mesquite: A Modular System forEvolutionary Analysis. Version 2.73. <http://mesquiteproject.org>.
Malcolm, W.M., 1995. Light transmission inside the thallus of Labyrintha implexa(Porpidiaceae, lichenized Ascomycetes). Bibl. Lichenol. 58, 275–280.
Mangold, A., Martin, M.P., Lücking, R., Lumbsch, H.T., 2008. Molecular phylogenysuggests synonymy of Thelotremataceae within Graphidaceae (Ascomycota:Ostropales). Taxon 57, 476–486.
Mason-Gamer, R.J., Kellogg, E.A., 1997. Testing for phylogenetic conflict amongmolecular data sets in the tribe Triticeae (Gramineae). Syst. Biol. 45, 524–545.
Matsuura, K., Yashiro, T., 2010. Parallel evolution of termite-egg mimicry bysclerotium-forming fungi in distant termite groups. Biol. J. Linn. Soc. 100, 531–537.
McCarthy, P.M., Healy, J.A., 1978. Dispersal of lichen propagules by slugs.Lichenologist 10, 131–132.
Meehan, C.J., Olson, E.J., Reudink, M.W., Kurt, T., Curry, R.L., 2009. Herbivory in aspider through exploitation of an ant–plant mutualism. Curr. Biol. 19, 1591–1682.
Miadlikowska, J., Lutzoni, F., 2000. Phylogenetic revision of the genus Peltigera(lichen-forming Ascomycota) based on morphological, chemical and largesubunit nuclear ribosomal DNA data. Int. J. Plant Sci. 161, 925–968.
Miller, A.N., 2003. A reinterpretation of the pseudo-bombardioid ascomal wall intaxa in the Lasiosphaeriaceae. Sydowia 55, 267–273.
Miller, A.N., Huhndorf, S.M., 2005. Multi-gene phylogenies indicate ascomal wallmorphology is a better predictor of phylogenetic relationships than ascosporemorphology in the Sordariales (Ascomycota). Mol. Phyl. Evol. 35, 60–75.
Moore, J., Willmer, P., 1997. Convergent evolution in invertebrates. Biol. Rev. 72, 1–60.
Mugambi, G.K., Huhndorf, S.M., 2009. Parallel evolution of hysterothecial ascomatain ascolocularous fungi (Ascomycota, Fungi). Syst. Biodivers. 7, 453–464.
Nadyeina, O., Grube, M., Mayrhofer, H., 2010. A contribution to the taxonomy of thegenus Rinodina (Physciaceae, lichenized Ascomycotina) using combined ITS andmtSSU rDNA data. Lichenologist 42, 521–531.
Nash, T.H., III., 2008. Lichen Biology, second ed. Cambridge University Press,Cambridge.
Nelsen, M.P., Lücking, R., Grube, M., Mbatchou, J.S., Muggia, L., Rivas Plata, E.,Lumbsch, H.T., 2009. Unravelling the phylogenetic relationships of lichenisedfungi in Dothideomyceta. Stud. Mycol. 64, 135–144.
Novotny, V., Drozd, P., Miller, S.E., Kulfan, M., Janda, M., Basset, Y., Weiblen, G.D.,2006. Why are there so many species of herbivorous insects in tropicalrainforests? Science 313, 1115–1118.
Nylander, J.A.A., Wilgenbush, J.C., Warren, D.L., Swofford, D.L., 2008. AWTY (are wethere yet?): a system for graphical exploration of MCMC convergence inBayesian phylogenetics. Bioinformatics 24, 581–583.
Olson, W., Hall, B.K., 2003. Keywords and Concepts in Evolutionary DevelopmentalBiology. Harvard University Press, Cambridge.
Pagel, M., 1994. Detecting correlated evolution on phylogenies: a general methodfor the comparative analysis of discrete characters. Proc. Roy. Soc. Lond. B 255,37–45.
Parnmen, S., Rangsiruji, A., Mongulsuk, P., Boonpragob, K., Elix, J.A., Lumbsch, H.T.,2010. Morphological disparity in Cladoniaceae: the foliose genus Heterodeaevolved from fruticose Cladia species (Lecanorales, Ascomycota). Taxon 59,841–849.
Penn, O., Privman, E., Landan, G., Graur, D., Pupko, T., 2010a. An alignmentconfidence score capturing robustness to guide-tree uncertainty. Mol. Biol. Evol.27, 1759–1767.
Penn, O., Privman, E., Ashkenazy, H., Landan, G., Graur, D., Pupko, T., 2010b.GUIDANCE: a web server for assessing alignment confidence scores. Nucl. AcidsRes. 38, W23–W28.
Posada, D., 2008. JModelTest: phylogenetic model averaging. Mol. Biol. Evol. 25,1253–1256.
Price, P.W., Diniz, I.R., Morais, H.C., Marques, E.S.A., 1995. The abundance of insectherbivore species in the tropics: the high local richness of rare species.Biotropica 27, 468–478.
Proksch, P., Hesbacher, S., 1997. Lichen products in lichen–herbivore interactions.Bibl. Lichenol. 67, 91–102.
62 E. Rivas Plata, H.T. Lumbsch /Molecular Phylogenetics and Evolution 61 (2011) 45–63

Pugh, A., 1976. An Introduction to Tensegrity. University of California Press,Berkeley and Los Angeles.
Rambaut, A., Drummond, A.J., 2007. Tracer v1.4. <http://beast.bio.ed.ac.uk/Tracer>.Rickson, F.R., 1969. Developmental aspects of the shoot apex, leaf and Beltian bodies
of Acacia cornigera L. Am. J. Bot. 56, 196–200.Rickson, F.R., 1976. Anatomical development of the leaf trichilium and Müllerian
bodies of Cecropia peltata L. Am. J. Bot. 63, 1266–1271.Rikkinen, J., 1995. What’s behind the pretty colours? A study on the photobiology of
lichens. Bryobrothera 4, 1–239.Rivas Plata, E., Lücking, R., Lumbsch, H.T., 2008. When family matters: an analysis of
Thelotremataceae (Lichenized Ascomycota: Ostropales) as bioindicators ofecological continuity in tropical rainforests. Biodiv. Conserv. 17, 1319–1351.
Rivas Plata, E., Lücking, R., Sipman, H.J.M., Mangold, A., Kalb, K., Lumbsch, H.T., 2010.A world-wide key to the thelotremoid Graphidaceae, excluding the Ocellularia–Myriotrema–Stegobolus clade. Lichenologist 42, 139–185.
Rivas Plata, E., Hernández, J.E., Lücking, R., Staiger, B., Kalb, K., Cáceres, M.E.S., 2011.Graphis is two genera – a remarkable case of parallel evolution in lichenizedAscomycota. Taxon 60, 99–107.
Schaefer, H.-C., Vences, M., Veith, M., 2002. Molecular phylogeny of Malagasypoison frogs, genus Mantella (Anura: Mantellidae): homoplastic evolution ofcolour pattern in aposematic amphibians. Org. Divers. Evol. 2, 97–105.
Schmitt, I., Del Prado, R., Grube, M., Lumbsch, H.T., 2009. Repeated evolution ofclosed fruiting bodies is linked to ascoma development in the largest groupof lichenized fungi (Lecanoromycetes, Ascomycota). Mol. Phyl. Evol. 52, 34–44.
Schoch, C.L., Sung, G.H., Lopez-Giraldez, F., Townsend, J.P., Miadlikowska, J.,Hofstetter, V., Robbertse, B., Matheny, P.B., Kauff, F., Wang, Z., Gueidan, C.,Andrie, R.M., Trippe, K., Ciufetti, L.M., Wynns, A., Fraker, E., Hodkinson, B.P.,Bonito, G., Groenewald, J.Z., Arzanlou, M., de Hoog, G.S., Crous, P.W., Hewitt, D.,Pfister, D.H., Peterson, K., Gryzenhout, M., Wingfield, M.J., Aptroot, A., Suh, S.O.,Blackwell, M., Hillis, D.M., Griffith, G.W., Castlebury, L.A., Rossman, A.Y.,Lumbsch, H.T., Lucking, R., Budel, B., Rauhut, A., Diederich, P., Ertz, D., Geiser,D.M., Hosaka, K., Inderbitzin, P., Kohlmeyer, J., Volkmann-Kohlmeyer, B.,Mostert, L., O’Donnell, K., Sipman, H., Rogers, J.D., Shoemaker, R.A., Sugiyama,J., Summerbell, R.C., Untereiner, W., Johnston, P.R., Stenroos, S., Zuccaro, A.,Dyer, P.S., Crittenden, P.D., Cole, M.S., Hansen, K., Trappe, J.M., Yahr, R., Lutzoni,F., Spatafora, J.W., 2009. The Ascomycota tree of life: a phylum-wide phylogenyclarifies the origin and evolution of fundamental reproductive and ecologicaltraits. Syst. Biol. 58, 224–239.
Seaward, M.R.D., 1988. Contribution of lichens to ecosystems. In: Galun, M. (Ed.),CRC Handbook of Lichenology, vol. II. CRC Press Inc., Boca Raton, pp. 107–129.
Shimodaira, H., Hasegawa, M., 1999. Multiple comparisons of log-likelihoods withapplications to phylogenetic inference. Mol. Biol. Evol. 16, 1114–1116.
Solhaug, K.A., Gauslaa, Y., Nybakken, L., Bilger, W., 2003. UV-induction of sun-screening pigments in lichens. New Phytol. 158, 91–100.
Staiger, B., 2002. Die flechtenfamilie Graphidaceae. Studien in Richtung einernatürlicheren Gliederung. Bibl. Lichenol. 85, 1–526.
Staiger, B., Kalb, K., Grube, M., 2006. Phylogeny and phenotypic variation in thelichen family Graphidaceae (Ostropomycetidae, Ascomycota). Mycol. Res. 110,765–772.
Stamatakis, A., 2006. RAxML-VI-HPC: maximum-likelihood-based phylogeneticanalyses with thousands of taxa and mixed models. Bioinformatics 22, 2688–2690.
Stamatakis, A., Ludwig, T., Meier, H., 2005. RAxML-III: a fast program for maximumlikelihood-based inference of large phylogenetic trees. Bioinformatics 21, 456–463.
Stearns, S., Hoekstra, R., 2005. Evolution: An Introduction. Oxford University Press,Oxford.
Swofford, D.L., 2003. PAUP�. Phylogenetic Analysis Using Parsimony (�and OtherMethods). Version 4. Sinauer Associates, Sunderland, Massachusetts.
Tehler, A., Irestedt, M., 2007. Parallel evolution of lichen growth forms in the familyRoccellaceae (Arthoniales, Ascomycota). Cladistics 23, 432–454.
Thell, A., Stenroos, S., Feuerer, T., Kärnefelt, I., Myllys, L., Hyvönen, J., 2002.Phylogeny of cetrarioid lichens (Parmeliaceae) inferred from ITS and beta-tubulin sequences, morphology, anatomy and secondary chemistry. Mycol.Progr. 1, 335–354.
Vilgalys, R., Hester, M., 1990. Rapid genetic identification and mapping ofenzymatically amplified ribosomal DNA from several Cryptococcus species. J.Bacteriol. 172, 4238–4246.
Vogel, S., 1955. Niedere ‘‘Fensterpflanzen’’ in der südafrikanischen Wüste. Eineökologische Schilderung. Beitr. Biol. Pfl. 31, 45–135.
Wedin, M., Baloch, E., Grube, M., 2002. Parsimony analyses of mtSSU and nITS rDNAsequences reveal the natural relationships of the lichen families Physciaceaeand Caliciaceae. Taxon 51, 655–660.
Zhang, J., Kumar, S., 1997. Detection of convergent and parallel evolution at theamino acid sequence level. Mol. Biol. Evol. 14, 527–536.
Zhou, S., Stanosz, G.R., 2001. Primers for amplification of mtSSU rDNA, and aphylogenetic study of Botryosphaeria and associated anamorphic fungi. Mycol.Res. 105, 1033–1044.
Zoller, S., Scheidegger, C., Sperisen, C., 1999. PCR primers for the amplification ofmitochondrial small subunit ribosomal DNA of lichen-forming ascomycetes.Lichenologist 31, 511–516.
E. Rivas Plata, H.T. Lumbsch /Molecular Phylogenetics and Evolution 61 (2011) 45–63 63




















