
Morphometric study on tadpoles of Bombina variegata (Linnaeus, 1758) (Anura; Bombinatoridae)
Upload
fieldmuseumCategory
view
0download
0
Organization of a Community of Tadpoles in Rain Forest Streams in BorneoAuthor(s): Robert F. Inger, Harold K. Voris and Karl J. FrognerSource: Journal of Tropical Ecology, Vol. 2, No. 3 (Aug., 1986), pp. 193-205Published by: Cambridge University PressStable URL: http://www.jstor.org/stable/2559850 .
Accessed: 01/10/2014 11:10
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Cambridge University Press is collaborating with JSTOR to digitize, preserve and extend access to Journal ofTropical Ecology.
http://www.jstor.org
This content downloaded from 128.135.12.127 on Wed, 1 Oct 2014 11:10:48 AMAll use subject to JSTOR Terms and Conditions

Journal of Tropical Ecology (1986) 2:193-205. With 1 figure
Organization of a community of tadpoles in rain forest streams in Borneo
ROBERT F. INGER, HAROLD K VORIS and KARLJ. FROGNER*
Field Museum of Natural History, Chicago, Illinois, 60605, USA
ABSTRACT. Samples of 29 distinct larval forms of anurans were collected in streams flowing through primary rain forest at Nanga Tekalit, Sarawak, at three periods spaced over 22 years. This assemblage of tadpoles lives in torrents, riffles, shingle areas, open pools, leaf drifts, pro- tected side pools, potholes in rocky banks, bank seepages, and pools of small intermittent streams. Positive associations of species in types of microhabitats and co-occurrence within sites lead to recognition of four taxonomically heterogeneous groups: one concentrated in leaf drifts; one using riffles, shingle areas, and open pools; one concentrated in potholes; and one using side pools and potholes. Most pairs of larval forms having strong negative association are characterized by differences in morphology and feeding behaviour; they show little evi- dence of competitive relations. Between-sites variation in species composition differs among the types of microhabitats, leaf drifts showing the least amount of variation. Species composi- tion within a site shows less change over intervals of 1-5 days than over intervals of 13-18 days. Variation in species composition is greater between sites than within sites over intervals of 9-31 days. Organization in this community seems to be effected by three factors: repro- ductive behaviour of adults and morphological and behavioural adaptations of tadpoles.
KEY WORDS: anuran larvae, Borneo, community organizations, rain forest.
INTRODUCTION
Anurans are probably the principal terrestrial vertebrate insectivores in tropical rain forests and yet the structure and dynamics of this component of a rain forest ecosystem have been dealt with in only a few papers all of which have concerned adult stages (e.g. Crump 1971, Toft & Duellman 1979). Our present study considers tadpoles living in rain forest streams in Borneo and comple- ments a previous analysis of the community organization of adult frogs and toads using the same streams (Inger 1969).
Much of the previous work on population ecology of tadpoles (e.g. Collins & Wilbur 1979, Heyer 1979, Morin 1983, Steinwascher 1979, Travis 1984, Wilbur 1984) has been carried out in the Temperate Zone with its special seasonal effects. The few tropical studies undertaken have involved areas with strong seasons and either pond environments or small (<3 m wide) inter- mittent streams (Heyer 1973, 1976, Heyer et al. 1975). Given habitats with little internal structure and climates providing strong cues, it is not surprising
* Present address: Hawaii Institute of Marine Biology, Kaneohe, Hawaii, 96744, USA.
(193)
This content downloaded from 128.135.12.127 on Wed, 1 Oct 2014 11:10:48 AMAll use subject to JSTOR Terms and Conditions

194 ROBERT F. INGER, HAROLD K. VORIS AND KARL J. FROGNER
that seasonal time is the primary dimension of resource partitioning in these studies (Toft 1985).
Our samples were collected in small to medium-sized streams (1-8 m wide) at one locality in Bornean rain forest over a 22-year period. There is no dry season and, related to that fact, the frogs using these streams for breeding do so throughout the year (Inger 1969, Inger & Bacon 1968). These circumstances provide a different set of abiotic factors and eliminate one of the biotic ele- ments, temporal variation in breeding periods, important in the dynamics of communities from strongly seasonal climates. As we have 29 'species' or dis- tinct larval forms (representing 11 genera), our assemblage is much richer in terms of genera and species than any other previously analysed. Heyer's (1973) sample from northeastern Thailand comes closest with 15 larval forms.
ENVIRONMENT
The location at which our sampling was carried out, Nanga Tekalit, Mengiong River, Seventh Division, Sarawak, is an area of hilly rain forest at 10 38' N/ 1130 35' E. The area has been described in some detail in previous papers (Inger 1979, Inger & Greenberg 1966). Precipitation is heavy throughout the year though the pattern is not uniform. In the only 12 month period (1962/63) for which we have records, 79 days had more than 25 mm of rain and those days were distributed in random fashion (Lloyd et al. 1968). Streams emptying into the Mengiong River vary in width from 3 to 15 m, have bottoms of sand, gravel, and bedrock, and consist of complex, differing mixtures of quiet and turbulent areas. The water is clear except after heavy rain. Most of our tadpole sampling was done in tributaries of the Mengiong 3-8 m wide, beginning about 100 m from the mouth and continuing upstream for 700-900 m. Some work was done in small (ca. 1 m), intermittent streams draining into principal tribu- taries. Water level fluctuated significantly in response to two circumstances: heavy rain in the catchment area of a tributary and back-flooding from the Mengiong rising in response to heavy rain upriver. Intra-basin flooding often raised water level more than 1 m and could convert these normally gentle streams into raging torrents. Back-flooding extended at least 600 m upstream from the mouth, led to temporary deposition of silt, and could allow large fish to enter previously inaccessible portions of streams.
We recognize nine types of microhabitats in these streams: (1) torrents, (2) riffles, (3) shingle areas, (4) open pools, (5) leaf drifts, (6) protected side pools, (7) potholes in rocky banks, (8) bank seepages, and (9) pools of intermittent streams. These are described in detail elsewhere (Inger 1985).
We acknowledge variation within each type and some fuzziness at the boun- daries between certain types (e.g. riffles and torrents). Furthermore, every site undergoes change in character in response to flooding. Potholes and side pools may lose their isolation. Intra-basin flooding expands the extent of torrents and increases the strength of current in all microhabitats. Back-flooding reduces current and converts long portions of a stream into one long, deep pool.
This content downloaded from 128.135.12.127 on Wed, 1 Oct 2014 11:10:48 AMAll use subject to JSTOR Terms and Conditions

Tadpole community in rain forest streams 195
MATERIALS AND METHODS
We sampled these streams September 1962-September 1963, April-July 1970, and March 1984. With few exceptions, we used dipnets and strainers of various sizes to collect tadpoles. A few were collected by use of rotenone and minnow seines. In some cases we tried to remove every tadpole from a site but at most sites we made 5-10 sweeps. Riffles and torrents were not sampled as effectively as other microhabitats. We did not attempt to collect in every possible site.
Tadpoles were preserved in formalin immediately after capture. Fewer than 1% were kept alive for rearing. We identified, staged, and measured larvae in our Chicago laboratory. The bases for identification are given elsewhere (Inger 1985). Three larval forms could not be assigned to species of adults. Informa- tion on morphology is taken from Inger (1985) and that on diets from a manu- script in preparation. We note the following taxonomic changes relative to the paper on adults; in each case the name used in Inger (1969) is followed after a dash by the current name: Nesobia mjobergi - Leptobrachella mjobergi, Lepto- brachium hasselti - L. montanum, Megophrys monticola - M. nasuta, Bufo biporcatus - B. divergens, Amolops jerboa - A. phaeomerus and A. poecilus, Rana macrodon - R. ingeri, Rana microdisca - R. palavanensis, Philautus gauni - Rhacophorus gauni, Microhyla annectans - M. petrigena.
DISTRIBUTION IN MICROHABITATS
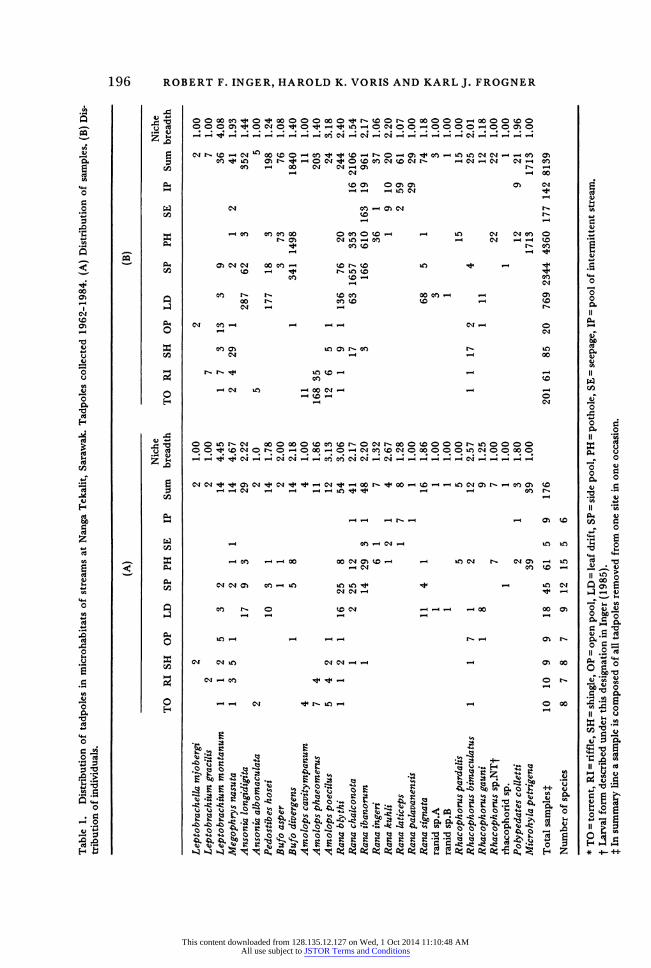
Niche breadth Distributions by number of samples and individuals are given in Table 1.
This table shows a wide variety of patterns. We compared the array for each species represented in seven or more samples against the distribution of all samples (bottom line of Table 1). All but one of the 17 tested, Bufo diver- gens, had distributions that differed significantly (P<0.01, chi-square tests) from expectation.
The species clearly differ in breadth of distribution. Again we limit con- sideration to forms appearing in at least seven samples. Based solely on number of microhabitats occupied, seven forms have broad niches, occurring in four or more microhabitats. But 90% of the samples of two of these, Rana chalconota and R. ibanorum, came from just two microhabitats. The other five, Lepto- brachium montanum, Megophrys nasuta, Amolops poecilus, Rana blythi, and Rhacophorus bimaculatus, were evenly distributed Applying the Simpson index to calculate niche breadth and using number of samples (Table 1A) give similar results: the last five species have the largest niche breadths in the assemblage (2.57-4.67) and the first two smaller ones (2.17-2.20). Calculations using number of individuals (Table 1B) yield similar results except that Rana ibanorum has a higher niche breadth (2.20) than Megophrys nasuta (1.90). At the other end of the scale, two forms were found in only one type of micro- habitat (Microhyla petrigena and Rhacophorus sp.NT) and four species in two (Rana ingeri, R. laticeps, Amolops phaeomerus, and Rhacophorus gauni).
This content downloaded from 128.135.12.127 on Wed, 1 Oct 2014 11:10:48 AMAll use subject to JSTOR Terms and Conditions
196 ROBERT F. INGER, HAROLD K. VORIS AND KARL J. FROGNER
a 4 oC oo oo+bQobo ooooo _ ooo C +o+_ _oC oo_ o_o
m ~~Z _ ____ccici i___ci_ _
Q
bQNe _e++ o_o+ eNN_
A D ee or+_o"+o-e"X"s _N_N Ne00 c g V) Ch _ oo Ct Ct _ o t- _~~~~~~~~~~~~~~~~~0
O~~~~~~~~~~~~~~~~~~~~~c 0 cn _oO o +
O 0
O-taNl
.U = _ e e e X O e O Q _ _ e N N e O ~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~- * 01 _c 0 T*
u~~~~~~~~~~~~~~0 fo cs to _ o
00 0
Y Q Y o o + Q Ct ~~~~o t- t o_o _ 0C too tooo
Ev cn _ _- cf _ ++_ _e ,
0 | : | Cn _ _ t0 N _ _ _ _ 00 _ | OO CA
in. 0 sd O - _ _ _ _ t- _ o t- 11 ?~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~- .QZ = N N e N N _ _ _ o X O ar~~~~~~~~~~~~~~~~~~~~~~~~~ S
V l S I M - e + + _ ? b I %'V oX~~~~~~~~~~~
sd~~ ~ ~ ~~~~ !O . to 0 r- 0 - 0 00 -4 0 to 0 00 0 0 0 in 0 0 00 0C<<228t o
H b 9 9 9 ~~~~~~t -l M -t ,e W t- "4 00 to t- f~ M to v
This content downloaded from 128.135.12.127 on Wed, 1 Oct 2014 11:10:48 AMAll use subject to JSTOR Terms and Conditions

Tadpole community in rain foeest streams 197
An important feature of both narrow- and broad-niched groups is that each is composite in terms of microhabitats exploited by its members and in terms of general morphology and habits. For example, the species with broad niches live in strong (Amolops poecilus), moderate (Leptobrachium montanum), or weak current (Rana blythi). This set includes one funnel-mouth, surface film feeder (Megophrys nasuta), one that feeds by scraping rocks while attached to substrate by an abdominal sucker (Amolops poecilus), and several less specialized morphologically and behaviourally. The group with narrow niches exploits strong current (Amolops phaeomerus), slowly flowing water (Rana laticeps), and standing water (Microhyla petrigena). The last is very small, lacks beaks and keratinized denticles, and is a midwater feeder. Amolops phaeomerus is a large tadpole and has the same morphological and behavioural specializations as A. poecilus. Rana laticeps closely resembles R. blythi. Rhaco- phorus sp.NT has a specialized oral disk unlike those of other forms living in these streams (Inger 1985).
Niche overlap There are some obvious differences between certain species in microhabitats
used (e.g. Microhyla petrigena and species of Amolops). Because the statistical significance of the difference between some pairs of species is not obvious, we carried out G tests for all pairs of the 17 relatively common species (number of samples >6). We used number of samples rather than individuals because large collections were not equally probable for all species. To reduce the num- ber of empty cells, we have grouped several microhabitats - torrent with riffle, shingle with open pool, and bank seepage with intermittent stream - combining microhabitats that are relatively close in bottom type and current.
One hundred and thirty six pair-wise tests were possible; the difference are statistically significant (P<0.05) in 118 tests. The species that do not differ form four clusters:
1. A group of five centered on leaf drifts: Ansonia longidigita, Pedostibes hosei, Rana blythi, R. signata, and Rhacophorus gauni.
2. Three using riffles, shingle areas, and open pools: Leptobrachium monta- num, Megophrys nasuta, and Rhacophorus bimaculatus. Amolops poecilus does not differ from M. nasuta, but does from the other two. Leptobrachium gracilis and Leptobrachella mjobergi would probably have been included here if we had had more samples.
3. A pothole group of three forms: Rana ingeri, Rhacophorus sp.NT, and Microhyla petrigena. Larval Rhacophorus pardalis probably belongs here (Table 1) though we had too few samples for testing.
4. Two species using side pools and potholes: Bufo divergens and Rana ibanorum. The pattern of R. ingeri does not differ statistically from these, though we suspect that it would differ from B. divergens if more samples had been available. The distribution of R. chalconota differs from that of R. ibanorum but not from that of B. divergens.
This content downloaded from 128.135.12.127 on Wed, 1 Oct 2014 11:10:48 AMAll use subject to JSTOR Terms and Conditions

198 ROBERT F. INGER, HAROLD K. VORIS AND KARLJ. FROGNER
Co-occurrence The fact that two species use the same type of microhabitat does not neces-
sarily mean that they occur in the same sites. Larvae of Rhacophorus sp.NT and Microhyla petrigena were found only in potholes, yet only 3 of the 7 samples of the former also included the latter. To evaluate co-occurrence of every pair of species we applied Fisher's exact probability test for the reasons given by Pielou (1969), using only the 17 species occurring in at least 7 samples. Of the 136 tests, there were 15 instances of significant positive association and 19 significant negative ones (P<0.05).
The tests of co-occurrence produce just two strong clusters. The first consists of the five species in the leaf drift group mentioned earlier and accounts for 10 of the 15 positive associations. Co-occurrences of these species are not limited to leaf drifts as each form also occurred in at least one additional type of microhabitat with the others. Rana chalconota has a significant (P<0.01) positive association with R. blythi but no other member of this group.
The other cluster of strongly associated species are three forms of the riffle-shingle group (see above). Rhacophorus bimaculatus, Megophrys nasuta, and Amolops poecilus not only have similar distributions; they also occur in the same sites more often than expected (P<0.02). But Leptobrachium mon- tanum, though using the same types of microhabitats, is not strongly associated with them (P>0.10). Amolops poecilus also co-occurs with A. phaeomerus in torrents and riffles (P = 0.01), but the latter's association with the other riffle- shingle forms is not significant (P >0.3 0).
Strong negative associations may reflect differences in adaptations of tad- poles to the physical circumstances of particular habitats as well as competitive interactions. Fifteen of the 19 statistically significant negative associations involve clear differences in adaptations, either in mode of feeding or morpho- logy. For example, Microhyla petrigena, which participates in nine of the negative associations, is the only midwater filter feeder in this fauna. Four additional cases of negative association involve larvae of Amolops with those of three species of Rana. These two groups differ greatly in morphology and feeding behaviour. Two more cases involve larval Megophrys nasuta, a surface film feeder, with larval Rana blythi and R. chalconota, which are bottom feeders. There is little evidence that competition accounts for negative associa- tions.
The frequency of statistically significant negative associations is depressed by the small number of samples for many species, in which cases the expected value is too close to zero. Consequently, strict adherence to the usual levels of statistical significance may result in an unacceptable number of Type II errors and the ignoring of biologically significant relations. We believe a more realistic assessment of the number of negative associations in this data set is achieved by including those pairs of species that show no instances of co-occurrence and whose distribution patterns are significantly different (P<0.01, G tests). This procedure adds 45 instances of negative associations.
This content downloaded from 128.135.12.127 on Wed, 1 Oct 2014 11:10:48 AMAll use subject to JSTOR Terms and Conditions

Tadpole community in rain forest streams 199
Twenty-four of these cases are probably accounted for by morphological and behavioural specializations and involve larval Amolops, Megaphrys nasuta, and Microhyla petrigena with other, more generalized tadpoles. Three more cases involve larval Leptobrachium montanum, which unlike the others is a leaf snip- per (Inger 1983). Four cases consist of pairs of larval Rana and three of larval Rhacophorus. Competition may be at issue in these seven congeneric pairs, but stronger evidence is needed. The remaining 11 cases are interfamilial pairs.
VARIATION AMONG MICROHABITATS IN SPECIES COMPOSITION
Differences within and between types of microhabitats Not every site of a given microhabitat type yielded the same species or even
the same number of species. Torrents had 1-4 species, riffles 1-4, shingle areas 1-4, open pools 1-3, leaf drifts 2-7, side pools 1-6, and potholes 1-6. Dif- ferences among types of microhabitats in number of species per site are statis- tically significant (P<0.001, Kruskal-Wallis ANOVA), with leaf drifts having the largest number and open pools the smallest.
Microhabitats differed in terms of between-sites variation in species composi- tion. We measured between-sites variation by calculating the Jaccard similarity coefficient (Sneath & Sokal 1973) for every pair of sites within each type of microhabitat. Leaf drifts had the highest median coefficient (S = 0.50), and the frequency distribution of these coefficients differed significantly (P<0.001, Kolmogorov-Smirnov test) from those of all other types of microhabitats. At the other extreme, the median value of coefficients for both riffles and open pools was S = 0. The frequency distribution of coefficients for these two types differed significantly (P<0.05) from those of all other microhabitats except torrents. Potholes and shingle areas had higher median coefficients (S = 0.25) than side pools and torrents (S = 0.20), but only potholes and side pools dif- fered significantly (P<0.001). Thus the ranking of microhabitats from least to most variable in species composition is: leaf drifts<potholes<side pools< riffles and open pools. Shingle areas and torrents seem to lie at about the level of potholes and side pools.
Variation among types of microhabitats in species richness was modest, with five having 7-9 species, only two with more, and two with fewer (Table 1). As is evident in the table, overlap between microhabitats in species composition varied widely and similarity between microhabits, measured by Jaccard's coefficient, varied from 0 to 0.66. The four types of microhabitats having moderate to strong current - torrents, riffles, shingle areas, open pools - share five species and constitute a set in which each pair has relatively high similarity (S = 0.45-0.66). The high similarities among these four are probably related to similarities in substrate and the proximity of shingle areas and open pools to riffles and torrents. Both factors increase the likelihood of a larva crossing the boundary between microhabitat types. Potholes and side pools have relatively
This content downloaded from 128.135.12.127 on Wed, 1 Oct 2014 11:10:48 AMAll use subject to JSTOR Terms and Conditions

200 ROBERT F. INGER, HAROLD K. VORIS AND KARL J. FROGNER
high S (0.50) as do side pools and shingle areas (0.43). The similarity of pot- hole and side pool assemblages seems to depend on two things: the presence of species restricted to areas of weak or no current (e.g. Bufo divergens, Rana ibanorum) and the presence of species having wide distributions (e.g. Rana blythi, Mecophrys nasuta). For all remaining pairs of microhabitats, S does not exceed 0.40. The physical circumstances of leaf drifts may account for the low similarity of this assemblage with others. Larvae, such as those of Amolops, that feed by scraping algae from rocks would not find appropriate food under leaf drifts, and larvae that apparently can survive only in the weakest of currents (e.g. Bufo divergens, Microhyla petrigena) would probably not succeed in gaining entrance to leaf drifts.
Variation over time within sites A wild assemblage of tadpoles is affected by predation, rate of development
of the constituent species, breeding activity of adults, competition, and fluctua- tion in physical conditions (Collins & Wilbur 1979, Heyer 1973, 1979, Wilbur 1984). In the case of the community we studied, in contrast to those investiga- ted by Heyer (north temperate zone and strongly seasonal tropics) and Collins and Wilbur (north temperate zone), the critical aspect of adult behaviour is aseasonal (Inger 1969, Inger & Bacon 1968) and the most important abiotic elements are aperiodic changes in current and water level (Inger 1969). The combined effect of these aperiodic factors on the assemblage of tadpoles living in one site should be gradual change in species composition and in relative abundances of species. If that conclusion is correct, the assemblage at a given site should remain relatively constant over a very short period (say, 1-5 days) but show signs of change over a slightly longer period (say, 10-30 days). How- ever, we do not expect the relationship between sampling interval and degree of change to be linear or to continue beyond a certain limit. Over long periods (say, 1-2 years), we expect continuity within limits, assuming the site is not severely damaged, because species do not alter breeding habits rapidly, because populations of adults apparently do not oscillate drastically over intervals of this length in Bornean forests, and because most larval forms are restricted to a limited number of types of microhabitats.
Our sampling procedures do not permit examination of changes in relative abundances, but are adequate for studies of species composition. Five sites - one pothole, two side pools, and two leaf drifts - were sampled after intervals of 1-5 and 13-18 days. For each site the similarity coefficient after the shorter interval was larger than that after the longer. The probability of this result is 0.03. The difference between coefficients is not related to the number of species involved; in four of the five pairs of coefficients, the number of species in the shorter interval equalled or exceeded by one the number of species involved in the longer. There is, therefore, more change in species composition over the longer period.
We have only two observations relevant to the issue of stability over the
This content downloaded from 128.135.12.127 on Wed, 1 Oct 2014 11:10:48 AMAll use subject to JSTOR Terms and Conditions

Tadpole community in rain forest streams 201
longest interval. Adult Bufo divergens were seen breeding in the same pothole on one stream in 1962, 1970, and 1984; in 1970 and 1984 tadpoles of that species were collected from that pothole. Larvae of Microhyla petrigena were collected from an adjacent pothole in 1970 and 1984; in 1962 adults of that species were caught on the rocks beside that pool. While these observations do not enable us to compare long- and short-term similarity in species composition, they demonstrate that at least some species use particular breeding sites for years, which places limits on variation.
Another approach to assessing stability within sites is by comparison of within- and between-sites variation. The behaviour of adults on these streams suggests the likelihood of individuals breeding repeatedly in the same small area. Adults of the four largest species marked and recaptured on these streams (Bufo asper, Rana blythi, R. ibanorum, R. ingeri) had small home ranges; 3 1% (of 452) had home ranges smaller than 10 m of bank length (Inger 1979). Bufo divergens was observed calling from a single large pothole on 15 nights spaced over a 12 month period. Ten males of this species were recaptured at this breeding site after intervals of 14 or more days, but none was recaptured at other sites on the same stream.
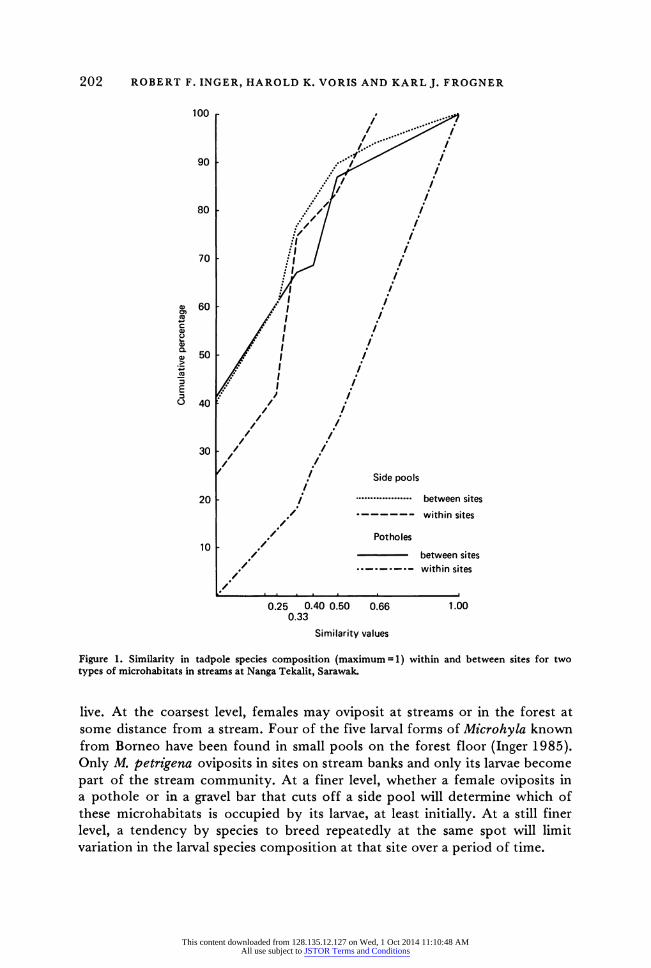
We had repeated larval samples after intervals of 9-31 days from 9 potholes having 1-5 species and from 10 side pools having 2-5 species. Within-sites similarity coefficients of potholes are significantly higher than those between sites (P<0.02, Kolmogorov-Smirnov test; see Figure 1). Although within-sites coefficients of side pools tend to be larger than those between sites (medians 0.33 and 0.20), the frequency distributions are not significantly different (P>0.10). The difference between these microhabitats in this regard is pro- bably related to their physical circumstances. Side pools are at stream bed level. A slight rise in water level results in renewed or stronger flow through side pools and, perhaps more important, deepens connections with the main flow allowing fish to enter. Potholes, on the other hand, are raised 1-3 m above the stream bed and are less subject to flooding and invasions by fish large enough to prey on tadpoles. During a 30-day period in 1984, these streams rose 0.6 m on two occasions, changing the size of side pools and deepening and enlarging their connections with the main flow, but not affecting potholes.
DISCUSSION AND CONCLUSION
This community of stream tadpoles has two major persistent features: groups of co-occurring larval forms and differences in the assemblages of species occurring in various types of microhabitats. There are also indications of sta- bility in species composition within sites over very short (<6 days) and rather longer periods (> 1 year). These regularities in organization appear to be mediated by three factors:
(i) Reproductive behaviour of adults. Sites of oviposition determine the range of possible microhabitats in which tadpoles of a particular species may
This content downloaded from 128.135.12.127 on Wed, 1 Oct 2014 11:10:48 AMAll use subject to JSTOR Terms and Conditions
202 ROBERT F. INGER, HAROLD K. VORIS AND KARLJ. FROGNER
100
so i<~ ~ ~~~~~/ !....
/lS / 90 '
80 .
70 ' / / /
a) 60 / I ,/
8'0 .. /
E / j /~~~/
E/ ,/
70 i
30 -
/ ~~~Side pools
20 - ...........-------- between sites , ~ ~~~ -- -- - -within sites
, ~~~~~Potholes 10 .
, ~~~~~~~~~between sites , * - - - -..... with in sites
C
0.25 0.40 0.50 0.66 1.00 0.33
Similarity values
Figure 1. Similarity in tadpole species composition (maximum =l) within and between sites for two types of microhabitats in streams at Nanga Tekalit, Sarawak.
live. At the coarsest level, females may oviposit at streams or in the forest at some distance from a stream. Four of the five larval forms of Microhyla known from Borneo have been found in small pools on the forest floor (Inger 1985). Only M. petrigena oviposits in sites on stream banks and only its larvae become part of the stream community. At a finer level, whether a female oviposits in a pothole or in a gravel bar that cuts off a side pool will determine which of these microhabitats is occupied by its larvae, at least initially. At a still finer level, a tendency by species to breed repeatedly at the same spot will limit variation in the larval species composition at that site over a period of time.
This content downloaded from 128.135.12.127 on Wed, 1 Oct 2014 11:10:48 AMAll use subject to JSTOR Terms and Conditions

Tadpole community in rain forest streams 203
Breeding throughout the year appears to be characteristic of the species using these streams (Inger 1969, Inger & Bacon 1968, and unpublished data), and this phenomenon also restricts variation in species composition at a site over a period of months.
(ia) Responses of morphotypes to physical conditions. Certain shapes and other morphological modifications are clearly associated with particular en- vironmental conditions. There are several clear examples, one of them being larval Ansonia albomaculata, which have streamlined bodies and expanded suctorial lips (Inger 1985). Larval Amolops have abdominal suckers (Inger 1985) enabling them to cling to rocks and maintain position in strong currents. Probably none of these suctorial devices function well, if at all, on silty sub- strates, limiting occurrence of larval Ansonia albomaculata and Amolops. The slender body of Leptobrachella larvae allows them to wriggle into interstices and avoid the force of current in riffles and torrents (Inger 1983). This beha- viour would not be appropriate over large areas of bedrock, which can be utilized by larval A. albomaculata and Amolops. Larvae having spheroidal bodies and deep tail' fins (e.g. Bufo divergens and Rhacophorus pardalis; see Inger 1985) are confined to areas of weak or no current. To the extent that morphology restricts distribution, variation among types of microhabitats in average current conditions and in bottom characteristics operates as a filter and leads to association of some larval forms and to ecological isolation of others.
(iii) Methods of feeding. On the basis of both direct observation of behaviour and indirect evidence of gut contents and oral and buccal morphology, we recognize five modes of feeding (see Satel & Wassersug 1981): obligate benthic, creation of bottom suspensions ('generalist' of Satel & Wassersug), macro- phagous, midwater suspension feeding, and particulate surface film feeding. We observed larval Amolops poecilus and Ansonia albomaculata using the first mode, Bufo divergens and Rana chalconota the second, Leptobrachium mon- tanum the third (Inger 1983), and Microhyla petrigena the fourth. Larval Mego- phrys have long been known to use the fifth method (Pope 1931, Smith 1926).
Each of these functions best within a limited range of microhabitats. Scraping/grazing on rocks harvests epilithic protists that cannot develop on bottoms covered by silt or dead leaves. Consequently, leaf drifts and silty side pools are not suitable for tadpoles, such as Ansonia albomaculata and Amolops.
Most Bornean larval forms feed by creating a suspension just above bottom debris and filtering the particles produced. We observed larval Bufo divergens, Rana chalconota, Rana ibanorum, and Rhacophorus pardalis moving slowly over the bottom debris in shallow side pools and potholes and presumed they were feeding in this manner. In moderate or strong currents such suspensions may be swept quickly beyond the reach of the tadpole creating them, and thus this method of feeding seems functional mainly in leaf drifts, side pools, pot- holes, and bank seepages.
Midwater filtering probably requires standing water, and Microhyla petri- gena, the only tadpole in our series using this mode of feeding, is restricted to
This content downloaded from 128.135.12.127 on Wed, 1 Oct 2014 11:10:48 AMAll use subject to JSTOR Terms and Conditions

204 ROBERT F. INGER, HAROLD K. VORIS AND KARL J. FROGNER
potholes. Surface film feeding should be possible wherever water is not turbu- lent; we found larval Megophrys nasuta, which uses this method, in a wide range of microhabitats (Table 1).
Dead leaves and their fragments accumulate in most stream microhabitats; they may even occur in crevices in the bottom of riffles and torrents. Tadpoles, such as those of Leptobrachium montanum (Inger 1983), that use leaf snipping can find food almost anywhere in these streams and live in a variety of micro- habitats (Table 1).
Considering this rain forest community in the light of Toft's (1985) analysis of resource partitioning sharpens the contrast between it and tadpole assemblages living in ponds in strongly seasonal climates. For the latter group, structural diversity of the habitat is low and climatic environmental cues strong. The community we studied lives in perennial streams, all of which are structurally heterogeneous, and in a climate lacking a dry season and having only about 10 minutes distinguishing the longest from the shortest day. For this Bornean community the opportunity to partition habitat is maximized while the oppor- tunity to partition seasonal time is minimized. The clusters of larval forms associated with particular types of microhabitats reflect the opportunity to partition space. The fact that these same clusters produce most of the cases of co-occurrence within samples shows that seasonal time is not important. On the basis of our restricted analysis of diet, this tadpole community seems to partition food, although the method of feeding and, therefore, types of food eaten are intertwined with distribution in space. Both of these dimensions of resource partitioning appear to have large historical or phylogenetic compo- nents and no detectable relation to competition.
ACKNOWLEDGEMENTS
We wish to express our thanks first to men of the Iban longhouse, Rumah Jimbong. Without their able assistance, none of the work reported here would have taken place. We also wish to acknowledge Messrs. Lucas Chin, Director, and Charles Leh, Zoologist, both of the Sarawak Museum, who arranged for government permits and eased many logistical problems. Four professional colleages, J. P. Bacon, W. Hosmer, F. W. King, and P. Walker, helped us collect tadpoles at various times. We received valued technical assistance from R. Arrigo, P. Gritis, M. Ozaki, A. Resetar, and K. Salvino at the Field Museum. Field and laboratory work were partially supported by the National Science Foundation (grants G20867, GB7845X, DEB82-04072) and the Allen-Heath Memorial Foundation.
LITERATURE CITED
COLLINS, J. P. & WILBUR, H. M. 1979. Breeding habits and habitats of the amphibians of the Edwin S. George Reserve, Michigan, with notes on the local distribution of fishes. Occasional Paper No. 686, Museum of Zoology, University of Michigan, Ann Arbor. 34 pp.
This content downloaded from 128.135.12.127 on Wed, 1 Oct 2014 11:10:48 AMAll use subject to JSTOR Terms and Conditions

Tadpole community in rain forest streams 205
CRUMP, M. L. 1971. Qualitative analysis of the ecological distribution of a tropical herpetofauna. Occa- sional Paper No. 3. University of Kansas Museum of Natural History, Lawrence. 62 pp.
HEYER, W. R. 1973. Ecological interactions of frog larvae at a seasonal tropical location in Thailand. Journal of Herpetology 7:337-361.
HEYER, W. R. 1976. Studies in larval amphibian habitat partitioning. Smithsonian Contributions in Zoology No. 242, Smithsonian Institution, Washington, DC. 27 pp.
HEYER, W. R. 1979. Annual variation in larval amphibian populations within a temperate pond. Journal of Washington Academy of Sciences 69:65-74.
HEYER, W. R., MCDIARMID, R. W. & WEIGMANN, D. L. 1975. Tadpoles, predation and pond habitats in the tropics. Biotropica 7:100-111.
INGER, R. F. 1969. Organization of communities of frogs along small rain forest streams in Sarawak. Journal of Animal Ecology 38:123-148.
INGER, R. F. 1979. Abundances of amphibians and reptiles in tropical forests of South-east Asia. Pp. 93-110 in Marshall, A. G. (ed.). Abundance of animals in Malesian rain forest. Transactions of the Sixth Aberdeen-Hull Symposium on Malesian Ecology, Department of Geography Miscellaneous Series No. 22, University of Hull, Hull.
INGER, R. F. 1983. Larvae of Southeast Asian species of Leptobrachium and Leptrobrachella (Anura: Pelobatidae). Pp. 13-32 in Rhodin, A. G. J. & Miyata, K. (eds). Advances in herpetology and evolu- tionary biology. Museum of Comparative Zoology, Cambridge, Mass.
INGER, R. F. 1985. Tadpoles of the forested regions of Borneo. Fieldiana: Zoology (NS) no. 26. 89 pp. INGER, R. F. & BACON, J. P. 1968. Annual reproduction and clutch size in rain forest frogs from
Sarawak. Copeia 1968:602-606. INGER, R. F. & GREENBERG, B. 1966. Ecological and competitive relations among three species of
frogs (genus Rana). Ecology 47:746-759. LLOYD, M., INGER, R. F. & KING, F. W. 1968. On the diversity of reptile and amphibian species in a
Bornean rain forest. American Naturalist 102:497-515. MORIN, P. J. 1983. Predation, competition, and the composition of larval anuran guilds. Ecological
Monographs 53:119-138. PIELOU, E. C. 1969. An introduction to mathematical ecology. Wiley, New York. POPE, C. H. 1931. Notes on amphibians from Fukien, Hainan, and other parts of China. Bulletin of the
American Museum of Natural History 59:397-611. SATEL, S. L & WASSERSUG, R. J. 1981. On the relative sizes of buccal floor depressor and elevator
musculature in tadpoles. Copeia 1981:129-137. SMITH, M. A. 1926. The function of the 'funnel' mouth of the tadpoles of Megophrys, with a note on
M. aceras Boulenger. Proceedings of the Zoological Society of London 1926:983-988. SNEATH, P. H. & SOKAL, R. R. 1973. Numerical taxonomy; the principles and practice of numerical
taxonomy. Freeman, San Francisco. STEINWASCHER, K. 1979. Competitive interactions among tadpoles: responses to resource level. Eco-
logy 60:1172-1183. TOFT, C. A. 1985. Resource partitioning in amphibians and reptiles. Copeia 1985:1-21. TOFT, C. A. & DUELLMAN, W. E. 1979. Anurans of the Lower Rio Llullapichis, Amazonian Peru: a
preliminary analysis of community structure. Herpetologica 35:71-77. TRAVIS, J. 1984. Anuran size at metamorphosis: experimental test of a model based on intraspecific
competition. Ecology 65:1155-1160. WILBUR, H. M. 1984. Complex life cycles and community organization in amphibians. Pp. 195-224
in Price, P. W., Slobodchikoff, C. N. & Gaud, W. S. (eds). A new ecology: novel approaches to inter- active systems. Wiley, New York.
Accepted 15 March 1986
This content downloaded from 128.135.12.127 on Wed, 1 Oct 2014 11:10:48 AMAll use subject to JSTOR Terms and Conditions
Copyright © 2022 FDOKUMEN




















