
Converting the cigarette butts into valuable products using the ...

335
International Journal of Medicinal Mushrooms, 11(4):335–350 (2009)
1521-9437/09/$35.00© 2009 by Begell House, Inc.
FOR AUTHOR U
SE ONLY
Mycelial Submerged Culture of New Medicinal Mushroom, Humphreya coffeata (Berk.) Stey. (Aphyllophoromycetideae) for the Production of Valuable Bioactive Metabolites with Cytotoxicity, Genotoxicity, and Antioxidant Activity
Sandra M. Porras-Arboleda,1,2 Norma A. Valdez-Cruz,3 Benjamín Rojano,1 Cecilia Aguilar,4 Leticia Rocha-Zavaleta,4 & Mauricio A. Trujillo-Roldán1,5,*1Universidad Nacional de Colombia, Sede Medellín, Calle 59A No 63 – 20, Medellín, Colombia; 2Universidad EAFIT, Carrera 49 #7 sur 50, A.A. 3300, Medellín, Colombia; 3Departamento de Medicina Molecular y Bioprocesos, Instituto de Biotecnología, Universidad Nacional Autónoma de México, AP. 510-3, Cuernavaca, CP. 62250 Morelos, México; 4Departamento de Biología Molecular y Biotecnología, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México, AP. 70228, México, D.F., CP. 04510, México; 5Unidad de Bioprocesos, Departamento de Inmunología, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México, AP. 70228, México, D.F., CP. 04510, México
* Address all correspondence to Mauricio A. Trujillo-Roldán, Unidad de Bioprocesos, Departamento de Inmunología, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México, AP. 70228, México, D.F., CP. 04510, México; Tel.: +52 55 56229192; Fax: +52 5 6223369; [email protected]; [email protected]
ABSTRACT: The mycelial submerged culture of the Colombian mushroom Humphreya coffeata, the production of exopolysaccharides (EPS), and the evaluation of the supernatant components in terms of bioactivity were investigated. Three carbon sources, three concentrations of carbon and nitrogen sources, and three pH values were evaluated. The optimal source was lactose (over sucrose and glucose), and the maximal biomass and EPS concentrations (15.5 g/L and 6.9 g/L, respectively) were obtained using lactose at 50 g/L, yeast extract 10 g/L, and an initial pH of 4.5. The cytotoxic, proliferative, genotoxic, and antioxidant activities were evaluated in lyophilized culture fi ltrates. Cytotoxicity was analyzed on human nontumorigenic keratinocytes (HaCaT), two human epithelial cell lines of cervical cancer (HeLa and InBl), and one lymphoma cell line (Jurkat). None of the fi ltrate concentrations that showed a cytotoxic effect on Jurkat cells (250–2500 μg/mL) were toxic for control HaCat cells and cervical cancer (HeLa and InBl). Furthermore, DNA damage was observed at a high extract concentration (2500 μg/mL), and genoprotection was observed using the lowest concentration tested (250 μg/mL). In addition, antioxidant activity was observed via the superoxide anion and the DPPH scavenging. This suggests that H. coffeata extracts can repress initiation and/or break the chain of oxidation reactions.
KEY WORDS: medicinal mushrooms, Humphreya coffeata, submerged culture, exopolysaccharides, genotoxicity, cytotoxicity, antioxidant activity
I. INTRODUCTION
An overview estimation of the number of different strains of mushrooms existing on Earth is approxi-
mately 150,000, and probably only less than 10% of them have been described.1,2 Molecular evidence has provided insight into several basidiomycete speciation rates in conjunction with biogeography

336 International Journal of Medicinal Mushrooms
S. M. PORRAS-ARBOLEDA ET AL.
FOR AUTHOR U
SE ONLYpatterns3,4 related to their complex growing environ-
ments and genetic background. A large portion of the ancient knowledge about the use of mushrooms as a medical resource is used nowadays in Eastern Europe and Asia. Mushrooms such as Ganoderma lucidum (Reishi), Lentinus edodes (Shiitake), Inonotus obliquus (Chaga), and many others have been in use continuously. In the Southern Hemi-sphere, particularly in Colombia, the fruit body of the medicinal mushroom species Humphreya cof-feata (Berk.) Stey. (Ganodermataceae) is used as a medical resource in small villages in the North of the country and is collected by elderly women who frequently pick the mushrooms together with their children.
For these reasons, we decided to analyze cul-tured extracts on two human epithelial cell lines of cervical cancer (HeLa and InBl), one lymphoma cell line (Jurkat), and a human nontumorigenic keratinocyte (HaCaT) as a positive control. Nowa-days, the accepted scientifi c name is H. coffeata5 (=Ganoderma coffeatum (Berk.) Furt.).
Many higher Basidiomycetes have several different biologically active compounds, such as intra- and extracellular polysaccharides, which have an effect on many types of cancer cells.6,7 More-over, these fungi also produce many oxygenated triterpenes (especially ganoderic acid), which affect cell cycle regulation, signal transduction cascades, apoptosis, angiogenesis, and metastasis.8–11 Those compounds are found in fruit bodies, cultured mycelium, and cultured broth.8,11,12 However, cultivating the fruit body of a mushroom usually takes several months. In natural conditions, the fruit body growth of H. coffeata takes between 3 and 5 months. For this reason, submerged culture
techniques have received a great deal of attention as a new, rapid, and reproducible alternative for the effi cient production of these metabolites.13
Submerged cultures of mushrooms as a higher productivity alternative was developed during the 1950s.14 However, compared to other fi lamentous models (e.g., Streptomyces), a literature survey of mushrooms will show few reports on the de-velopment of higher fungi bioprocesses.15 Yields and productivity of mycelium-submerged cultures depend on the mushroom itself, the substrate, and the culture conditions. To our knowledge, there are no reports in the literature about the submerged culture of H. coffeata, the production of its me-tabolites, or its bioactivity characterization. In this work, the effects of the initial pH of the medium and the concentration and type of the carbon and nitrogen sources were studied regarding the biomass growth and exopolysaccharide (EPS) production. Selection of the initial culture conditions and nutrients concentration are considered important for the development of industrial-scale cultures of valuable fungi.16 Moreover, cytotoxic, genotoxic, and antioxidant activities of lyophilized culture fi ltrates were evaluated in order to determine the feasibility of H. coffeata as a promising medicinal mushroom. Because this fungi H. coffeata (known popularly as an herbal medicine) evolved in a dif-ferent biogeographically environment from other Asiatic fungi described (G. lucidum,17–19 Lentinus edodes,20 Inonotus obliquus,21,22 and other me-dicinal mushroom species2,13,23–25 ), it has opened up the possibility to discover novel activities and components. This study presents a bioprocess to produce bioactive compounds from a promising natural tool, in submerged cultures. To the best of
ABBREVIATIONS
ABTS: 2,2’-azino-bis(3-ethylbenzothiazoline-6-sulphonic acid); BHT: butylated hydroxytoluene; DMEM: Dulbecco’s Modified Eagle Medium; DMSO: dimethyl sulfoxide; DPPH: 1,1-diphenyl-2-picrylhydrazyl; EC50: effective con-centration at which the antioxidant activity was 50%; EPS: exopolysaccharides; FBS: fetal bovine serum; HaCaT: human nontumorigenic keratinocytes; HeLa and InBl: two human epithelial cell lines of cervical cancer; Jurkat: immortalized line of T lymphocytes or a lymphoma cell line; NBT: nitroblue tetrazolium; NADH: β-nicotinamide adenine dinucleotide; PDA: potato dextrose agar; PMS/NADH: phenazine methosulfate system-β-nicotinamide adenine dinucleotide; RNS: reactive nitrogen species; ROS: reactive oxygen species; RPMI-1640: Roswell Park Memorial Institute animal cells medium.

Volume 11, Issue 4, 2009 337
MYCELIAL SUBMERGED CULTURE OF HUMPHREYA COFFEATA FOR THE PRODUCTION OF VALUABLE METABOLITES
FOR AUTHOR U
SE ONLY
the authors’ knowledge, this is the fi rst detailed production of bioactive components in submerged cultures of H. coffeata.
II. MATERIALS AND METHODS
A. Mushroom Strain
Humphreya coffeata was acquired from the cul-ture collection of the Universidad de Antioquia (Medellín, Colombia), which had been originally isolated from the North Colombian town of Tierra Alta, (Cordoba, Colombia). H. coffeata was co-cul-tured and stored in the Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México (Mexico City). The stock culture was maintained on potato dextrose agar (PDA) slants. The slants were incubated at 30ºC for 7 days, and then stored at 4ºC and maintained ready for cultur-ing for 4–5 weeks. Preservation of stock cultures was done by freezing at –80ºC in 1.5 mL cryotubes with 40% glycerol.
B. Inoculum Preparation, Media, and Experimental Design
One complete colony of H. coffeata grown on PDA medium was transferred to seed submerged cultures in 250-mL fl asks containing 50 mL of culture medium at 120 rpm, 30°C, initial pH of 5.5 for 7 days in darkness. The culture medium previously reported for G. lucidum16,26 containing (in g/L) glucose, 35.0; peptone, 5.0; yeast extract, 2.5; KH2PO4⋅H2O, 1.0; MgSO4⋅7H2O, 0.5; and vitamin B1, 0.05 was used. All experiments were performed in 250-mL Erlenmeyer fl asks containing 45 mL of culture media and 5 mL of seed culture, and all were performed at least in triplicate (with the exception of the experiments shown in Fig. 1, which were done in duplicate).
Three levels of experiments were carried out. First, three carbon sources were evaluated (glu cose, lactose, and sucrose) at 35 g/L, using yeast extract and peptone at 5.0 g/L, and an initial pH of 5.5. After selecting the best carbon source, three levels of carbon source (20, 35, and 50 g/L) and yeast
FIGURE 1. Biomass growth (•), EPS production (∇), and reducing sugar concentration (■) of cultures of Humphreya coffeata carried out in conventional shake fl asks at 30°C, 120 rpm, and darkness, using optimized culture medium (pH 4.5, lactose 50 g/L, and yeast extract 10 g/L).

338 International Journal of Medicinal Mushrooms
S. M. PORRAS-ARBOLEDA ET AL.
FOR AUTHOR U
SE ONLY
extract (2.5, 5.0, and 10.0 g/L), and three initial pH values (4.5, 5.5, and 6.5), were evaluated in order to optimize biomass and EPS production. Finally, the optimized conditions were used to determine biomass growth, reducing sugars consumption, and EPS production kinetics. All cultures were carried out for 14 to 16 days at 30°C, 120 rpm, and under darkness.
C. Analytical Methods
1. Cell Concentration
To obtain representative data, the contents of all the fl asks were used as a sample. The dry cell weight was measured by fi ltering the sample through a Whatman #2 fi lter paper. The fi ltrate was set aside to determine reducing sugars and extracel-lular polysaccharide concentration. The cell pellet obtained was washed twice with distilled water and dried overnight to a constant weight at 50ºC for 24 hours. The cell concentration is expressed as the gram dry cell weight per liter.
2. Reducing Sugar and Extracellular Polysaccharide Concentration
A major part of the fi ltrate through a Whatman #2 fi lter paper was lyophilized on a Lyph-Lock 4.5 (Labconco, Kansas City, MO, USA). For measuring reducing sugars and to obtain a small fraction of exopolysaccharides (EPS), 1.0 mL of fi ltrate taken prior to lyophilization was incubated with 4.0 mL of 95% (v/v) cold ethanol and centrifugated at 5000 × g for 15 minutes. The precipitate containing all EPS was redissolved in NaOH 1M at 60°C for 1 hour, and the EPS were determined by the modifi ed phenol–sulfuric acid method or Dubois method for total sugars at 492 nm.27,28 Moreover, supernatant was used for the determination of reducing sugars by the dinitrosalicylic acid (DNS) method.29 On the other hand, when necessary, lyophilized culture fi ltrates were dissolved in cancer cell culture medi-ums (DMEM supplemented with 10% fetal bovine serum or RPMI-1640) to the desired concentrations, and stored at 4ºC for later use.
D. Cytotoxicity and Proliferation Assays
Cytotoxicity analysis was determined using three human epithelial cell lines: two cervical cancer cell lines, HeLa and InBl,30 and HaCaT (a nontumorigenic keratinocyte). Cells were maintained in DMEM medium supplemented with 10% fetal bovine se-rum and penicillin-streptomycin (2%). Lyophilized H. coffeata fi ltrates were diluted in supplemented cell culture medium. To evaluate cytotoxicity, 5 × 104 cells were cultured in the presence of different concentrations of H. coffeata fi ltrates (30, 60, 120, 250, 500, 1000, 3000, 6000, 12000, 25000, and 50000 μg/mL) at 37°C for 24 hours. For the cell proliferation experiments, 5 × 103 cells were cultured in the presence of 10, 25, 50, 100, and 250 μg/mL of H. coffeata fi ltrates at 37°C for 24, 48, 72, and 96 hours. The number of cells was determined by the MTT assay.31,32 Briefl y, 50 μL MTT (0.5 mg/mL) were added to each well and incubated at 37°C for 2 hours. MTT-containing medium was discarded, and 100 μL/well of isopropanol was added. Formazan precipitate was dissolved by pipetting each well until no precipitate was visible. The plates were read by an ELx800 (BIO-TEK) plate reader at 570 nm. Normalization of the proliferation shown in Figure 2 was done based on the cell line number grown at each time point in the absence of H. coffeata extracts. In addition, a human leukemia cell line (Jurkat) was used, cultured in RPMI-1640 medium (Sigma Chemicals Corp, St. Louis, MO, USA), supplemented with SBF 10% and penicillin–streptomycin (2%). The cytotoxicity of the lyophilized culture fi ltrates was determined by colorimetric MTT assay. In this case, triplicate dilutions of the lyophilized culture fi ltrates (250, 500, 1000, 1500, and 2500 μg/mL) were added to 1.5 × 104 cells/mL on 96-well fl at-bottomed plates and incubated at 37ºC under 5% CO2 and humid atmosphere for 48 and 96 hours.
On the other hand, to observe if the osmotic equilibrium is altered by the addition of lyophilized culture fi ltrates, we measured the conductivity in a Radiometer Copenhagen Conductivity Meter CDM2f (Copenhagen, Denmark). It is noteworthy that there can be no simple relationship between conductivity and osmotic pressure. However, both quantities increase with concentration in a rather linear manner, so it is possible to “calibrate” con-

Volume 11, Issue 4, 2009 339
MYCELIAL SUBMERGED CULTURE OF HUMPHREYA COFFEATA FOR THE PRODUCTION OF VALUABLE METABOLITES
FOR AUTHOR U
SE ONLY
FIGURE 2. Normalized proliferation, at different Humphreya coffeata extract concentrations, of nontumorigenic keratino-cytes HaCaT (•) and two human cervical cancer cell lines, HeLa (▼) and InBl (○), at 24 (A), 48 (B), and 96 hours (C).
ductivity measurements with osmotic pressures by the diffi culty of the last to be measured.
E. Genotoxicity Assays
Genotoxicity was determined using a human leu-kemia cell line (Jurkat), cultured in RPMI-1640 medium (Sigma), supplemented with SBF 10% and penicillin–streptomycin (2%). Genotoxicity
was measured using the Comet Assay; this is a single-cell-based technique allowing the detection and quantifi cation of DNA damage.33–36 In dark-ness, 1.0 × 106 cells/mL were exposed to 50 μL of H. coffeata culture fi ltrates (dissolved in deionized water to 250, 1000, and 2500 µg/mL). As a positive control (C+), 50 μL of H2O2 (100 μM) was used, and 50 µL of DMSO was used as negative control (C–). After incubation (4ºC for 1 hour), 10 µL of cell suspension were mixed with low melting point

340 International Journal of Medicinal Mushrooms
S. M. PORRAS-ARBOLEDA ET AL.
FOR AUTHOR U
SE ONLY
agarose (0.5% w/v in PBS, Ca++, Mg++ free), using a pretreated slide with 100 μL of normal melting point agarose (0.5% w/v in PBS, Ca++, Mg++ free), and incubated at 4ºC for 5 minutes. After that, slides were submerged in lysis solution (2.5 M NaCl, 0.1 M EDTA, 10 mM Tris-HCl 10% w/v DMSO, 1% Triton X-100, pH 10) for 1 hour at 4ºC. Finally, two washes in cold PBS were performed, followed by incubation in darkness at 4ºC for 0.5 hours. Electrophoresis was done at 25 V for 20 minutes. Subsequently, slides were covered with neutralizing buffer (Tris-HCl 0.4 M, pH 7.5) for 15 minutes and washed with cold methanol. Last, each slide was colored with ethidium bromide (2 µg/mL) and observed at 20× under a microscope (Carl Zeiss SMT Corp., Peabody, MA, USA) with fl uorescence. Twenty-fi ve photos were taken of each sample (SONY DSC-S85) and analyzed using Comet Score software (TriTek-Corp., www. autocomet.com). Tail moment is defi ned as the product of the tail length and the fraction of total DNA in the tail. Tail mo-ment incorporates a measure of both the smallest detectable size of migrating DNA (refl ected in the comet tail length) and the number of relaxed/broken pieces (represented by the intensity of DNA in the tail). Genoprotective activity was measured using the same concentrations as above, in a similar technique (Comet Assay), but 50 μL of H2O2 (100 μM) was used in the incubation at 4ºC for 1 hour as negative control (C–), and 50 μL of DMSO was used as positive control (C+).
F. Antioxidant Activity of Lyophilized Culture Filtrates
Five aliquots taken from the H. coffeata culture fi ltrates at different phases of growth (early: day 4; mid: day 8; ending exponential: day 12; and station-ary phase: days 16 and 21) were evaluated by various methods searching for antioxidant activity.
1. Scavenging Effect on 1,1-diphenyl-2-picrylhydrazyl (DPPH) Radicals
The effect of lyophilized culture fi ltrates on the DPPH radical was estimated according the method
reported by Hatano et al.37 Culture fi ltrates (10 μL) were incubated with 990 μL of DPPH (20 mg/L) in triplicate. The mixture was shaken vigorously and allowed to stand for 0.5 hour in darkness, and measured at 517 nm (Jenway 6405, England). The scavenging ability was calculated by scavenging ability (%) = [(∆A517 control – ∆A517 sample)/∆A517 control] × 100. The blank used was methanol (1.0 mL) and sample solution (10 μL). The control was DPPH solution (1.0 mL) and DMSO (10 μL).37,38
2. ABTS Radical Scavenging Assay by Spectrometry
This method is based on the ability different compounds have to scavenge the radical cation ABTS+ (2,2’-azino-bis[3-ethylbenzothiazoline-6-sulphonic acid]) compared to the standard antioxi-dant Trolox (Sigma).39 ABTS+ was produced by reacting 3.5 mmol/L ABTS stock solution with 1.5 mmol/L potassium persulfate overnight, in dark-ness and at room temperature. The ABTS+ solu-tion was diluted with PBS to reach 0.70 (± 0.05) at 734 nm. Then, 10 μL of sample or butylated hydroxytoluene ([BHT], Sigma) were added to 990 μL of diluted ABTS+ solution. After 0.5 hour in darkness, the change in absorbance is proportional to the quenched ABTS+. The culture fi ltrates were assayed in triplicate, and the calibration was car-ried out using Trolox as the standard (between 0.8 and 16 μM). The EC50 (mg/mL) is the effective concentration at which the antioxidant activity was 50% and was obtained by interpolation of the linear regression analysis.
3. Superoxide Anion Scavenging Activity
Superoxide radicals are generated in a phenazine methosulfate system (PMS)-β-nicotinamide adenine dinucleotide (NADH), by the oxidation of NADH, and assayed by the reduction of nitroblue tetrazo-lium (NBT). Measurement of superoxide anion scavenging activity was done based on a method described, with some modifi cations.40 The reaction solution was prepared using 0.25 mL of NBT (156 μM in 100 mM phosphate buffer, pH 7.4), 0.25
Volume 11, Issue 4, 2009 341
MYCELIAL SUBMERGED CULTURE OF HUMPHREYA COFFEATA FOR THE PRODUCTION OF VALUABLE METABOLITES
FOR AUTHOR U
SE ONLY
mL of NADH (468 μM in 100 mM phosphate buffer, pH 7.4), and 1 mL of the culture fi ltrates in methanol. The reaction started by adding 1 mL of PMS (60 μM in 100 mM phosphate buffer, pH 7.4) to the mixture. The reaction was incubated at 25ºC and measured at 560 nm for 5 minutes. (+) Catechin was used as a standard. The inhibition ratio (%) was calculated using the following formula: inhibition percentage = 100 × (slope of sample – slope of control)/slope of control. The IC50 values were calculated by exponential regression plots, where the abscissa represented the concentration
of samples, and the ordinate represented the inhibi-tion ratio percent.
G. Statistical Analysis
Data were expressed as the average ± standard devia-tion of at least three independent experiments (Figs. 3 and 4), or just as the average of two independent ex-periments (Fig. 1). The cytotoxicity and genotoxicity were analyzed by ANOVA and Fisher average analysis using Statgraphics Plus software, version 4.0.
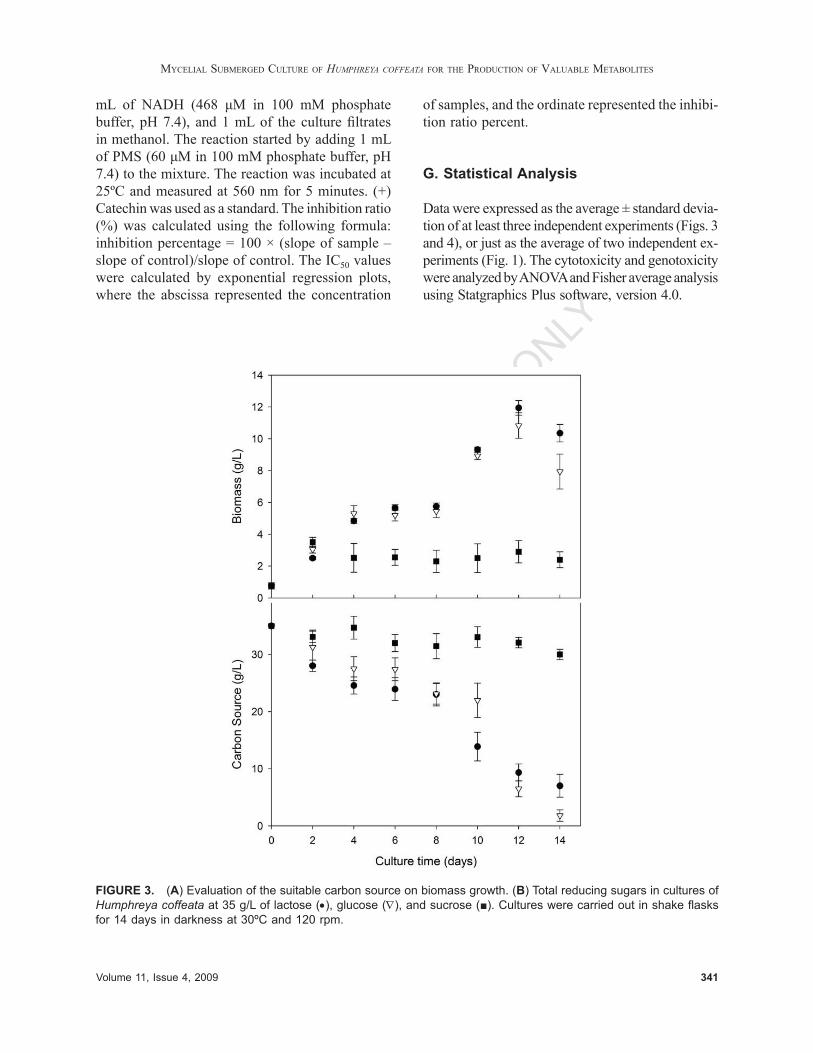
FIGURE 3. (A) Evaluation of the suitable carbon source on biomass growth. (B) Total reducing sugars in cultures of Humphreya coffeata at 35 g/L of lactose (•), glucose (∇), and sucrose (■). Cultures were carried out in shake fl asks for 14 days in darkness at 30ºC and 120 rpm.

342 International Journal of Medicinal Mushrooms
S. M. PORRAS-ARBOLEDA ET AL.
FOR AUTHOR U
SE ONLY
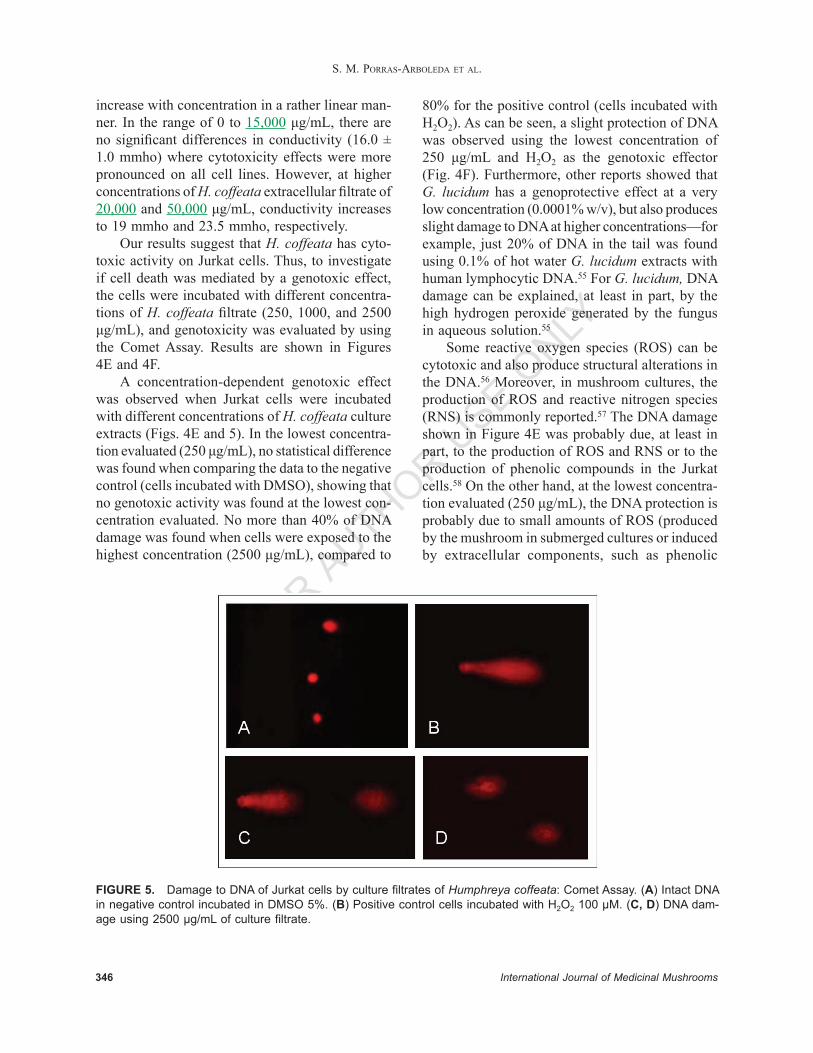
FIGURE 4. (A) Cytotoxicity of culture fi ltrate extracts of Humphreya coffeata in two human cervical cancer cell lines—HeLa (•) and InBl (○); and (B) nontumorigenic keratinocytes (HaCaT) as a normal cell control. Cytotoxicity of culture fi ltrate extracts of H. coffeata in Jurkat cells after 48 hours (C) and 96 hours (D) of incubation. C– (DMSO 5%) and C+ (H2O2 100 µM). (E) Genotoxicity of culture fi ltrate extracts of H. coffeata in Jurkat cells. C– (DMSO 5%) and C+ (H2O2 100 µM), evaluated by Comet Assay. (F) Genoprotection of cellular DNA of Jurkat cells in the presence of aqueous culture fi ltrate extracts and H2O2 100 µM. C+ (DMSO 5%) and C– (H2O2 100 µM) evaluated by Comet Assay. Data represent an average of Olive tail moment with standard deviation.

Volume 11, Issue 4, 2009 343
MYCELIAL SUBMERGED CULTURE OF HUMPHREYA COFFEATA FOR THE PRODUCTION OF VALUABLE METABOLITES
FOR AUTHOR U
SE ONLY
III. RESULTS AND DISCUSSION
A. Effect of the Carbon Source
To fi nd out the suitable carbon source that maxi-mized biomass mycelial growth of H. coffeata, and to evaluate the average duration of H. coffeata cultures, three carbon sources (glucose, lactose, and sucrose) were evaluated and provided at 35 g/L. Among the carbon sources evaluated, lactose showed the highest biomass concentration (11.9 ± 0.5 g/L, at day 12, Fig. 3), and no signifi cant dif-ferences were found with cultures carried out with glucose (10.8 ± 0.8 g/L, also at day 12). However, by using sucrose, just 2.9 ± 0.7 g/L were obtained. It is possible that sucrose has an effect on the catabolic repression of the cellular secondary metabolism, as previously reported for other mushrooms.41 The biomass yield and the productivity of the culture using lactose were 0.46 gbiom/glac and 0.99 gbiom/L day, respectively. Alternatively, when using glucose, the yields were 0.38 gbiom/gglu and 0.90 gbiom/L day. Therefore, lactose was selected as the carbon source in the following experiments, which was additionally supported by data previously reported in fed batch cultures for G. lucidum,28 which state that lactose is the best carbon source for biomass growth compared to maltose and glucose, and there is practically no biomass mycelial growth when cul-tured using sucrose.28 In cultures of G. lucidum, the β-galactosidase activity was measured and observed in exponential and stationary phases of growth, sug-gesting the existence of a lactose permease system in the cells.42 Similar behavior needs to be analyzed in H. coffeata cultures in future work.
Two growth phases were observed in H. cof-feata kinetics using lactose and glucose as carbon sources (Fig. 3). For the fi rst 4 days of culture, H. coffeata grew at a specifi c rate of 0.46, 0.48, and 0.30 day–1 using lactose, glucose, and sucrose as car-bon sources. From days 4 to 8, H. coffeata stopped growing, and from days 8 to 12, a second growth phase was observed at specifi c growth rates of 0.18 and 0.17 day–1 for lactose and glucose, respectively; however, no growth was observed for sucrose. A similar case of a diauxic growth phenomenon was demonstrated for G. lucidum43 and elucidated when the cultures were complemented using complex
sources of carbon and nitrogen, signifying a catabo-lite repression. Moreover, authors proposed that the production of some metabolites, as ganoderic acids, was inhibited by the high concentrations of glucose and peptone, which is consistent with the production of other secondary metabolites.43
B. Effect of the Initial Carbon and Nitrogen Concentration and pH
To investigate the suitable culture conditions for EPS and biomass production, three levels of carbon source (20, 35, and 50 g/L of lactose), nitrogen source (2.5, 5.0, and 10.0 g/L of yeast extract, supplemented with 5.0 g/L of peptone), and an initial pH (4.5; 5.5; 6.5) were evaluated (Table 1). The use of organic complex nitrogen sources (peptone and yeast extract) was based on the fact that most Basidiomycetes prefer these complex sources for a favorable culture because certain essential amino acid(s) can scarcely be synthesized from inorganic nitrogen sources in the cultivation of higher fungi.44
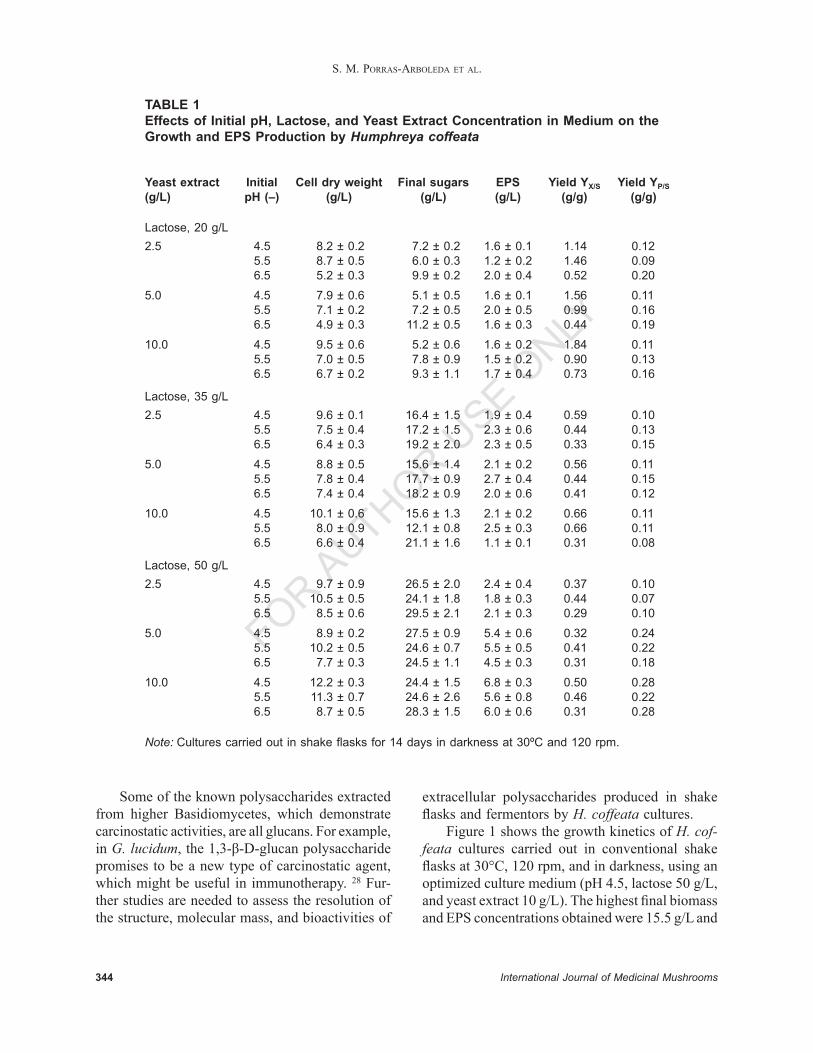
Table 1 shows data for the average and standard deviation of at least three shake fl asks cultured for each condition. As can be seen, the lowest pH evaluated (4.5), with 50 g/L of lactose and 10 g/L of yeast extract, resulted in the higher fi nal biomass concentration (12.2 ± 0.3 g/L). The same triplet of conditions resulted in the highest EPS production (6.8 ± 0.3 g/L), consuming 25.6 ± 1.5 g/L of lactose. The highest biomass yield (YX/S of 1.84 gbiom/glac) was found using the lowest carbon source concen-tration (20 g/L), a three times higher yield than that reported for G. lucidum (0.577 g/g at 20 g/L of glucose) in similar conditions.15 Moreover, Table 1 shows that the highest EPS yield (YP/S) observed was 0.28 g/g (with a maximum EPS production of 6.8 ± 0.3 g/L) using the highest evaluated lactose concentration. A similar pattern was reported for G. lucidum, with a maximum yield of 0.028 g/g (with a maximum EPS concentration of 1.08 g/L) at the highest carbon source concentration.15 These comparisons present H. coffeata as a potential new strain able to produce high levels of EPS and new culture strategies, such as fermentors and high cell density cultures, which are desirable in order to increase biomass and EPS production.
344 International Journal of Medicinal Mushrooms
S. M. PORRAS-ARBOLEDA ET AL.
FOR AUTHOR U
SE ONLY
TABLE 1Effects of Initial pH, Lactose, and Yeast Extract Concentration in Medium on the Growth and EPS Production by Humphreya coffeata
Yeast extract Initial Cell dry weight Final sugars EPS Yield YX/S Yield YP/S(g/L) pH (–) (g/L) (g/L) (g/L) (g/g) (g/g)
Lactose, 20 g/L2.5 4.5 8.2 ± 0.2 7.2 ± 0.2 1.6 ± 0.1 1.14 0.12 5.5 8.7 ± 0.5 6.0 ± 0.3 1.2 ± 0.2 1.46 0.09 6.5 5.2 ± 0.3 9.9 ± 0.2 2.0 ± 0.4 0.52 0.20
5.0 4.5 7.9 ± 0.6 5.1 ± 0.5 1.6 ± 0.1 1.56 0.11 5.5 7.1 ± 0.2 7.2 ± 0.5 2.0 ± 0.5 0.99 0.16 6.5 4.9 ± 0.3 11.2 ± 0.5 1.6 ± 0.3 0.44 0.19
10.0 4.5 9.5 ± 0.6 5.2 ± 0.6 1.6 ± 0.2 1.84 0.11 5.5 7.0 ± 0.5 7.8 ± 0.9 1.5 ± 0.2 0.90 0.13 6.5 6.7 ± 0.2 9.3 ± 1.1 1.7 ± 0.4 0.73 0.16
Lactose, 35 g/L2.5 4.5 9.6 ± 0.1 16.4 ± 1.5 1.9 ± 0.4 0.59 0.10 5.5 7.5 ± 0.4 17.2 ± 1.5 2.3 ± 0.6 0.44 0.13 6.5 6.4 ± 0.3 19.2 ± 2.0 2.3 ± 0.5 0.33 0.15
5.0 4.5 8.8 ± 0.5 15.6 ± 1.4 2.1 ± 0.2 0.56 0.11 5.5 7.8 ± 0.4 17.7 ± 0.9 2.7 ± 0.4 0.44 0.15 6.5 7.4 ± 0.4 18.2 ± 0.9 2.0 ± 0.6 0.41 0.12
10.0 4.5 10.1 ± 0.6 15.6 ± 1.3 2.1 ± 0.2 0.66 0.11 5.5 8.0 ± 0.9 12.1 ± 0.8 2.5 ± 0.3 0.66 0.11 6.5 6.6 ± 0.4 21.1 ± 1.6 1.1 ± 0.1 0.31 0.08
Lactose, 50 g/L 2.5 4.5 9.7 ± 0.9 26.5 ± 2.0 2.4 ± 0.4 0.37 0.10 5.5 10.5 ± 0.5 24.1 ± 1.8 1.8 ± 0.3 0.44 0.07 6.5 8.5 ± 0.6 29.5 ± 2.1 2.1 ± 0.3 0.29 0.10
5.0 4.5 8.9 ± 0.2 27.5 ± 0.9 5.4 ± 0.6 0.32 0.24 5.5 10.2 ± 0.5 24.6 ± 0.7 5.5 ± 0.5 0.41 0.22 6.5 7.7 ± 0.3 24.5 ± 1.1 4.5 ± 0.3 0.31 0.18
10.0 4.5 12.2 ± 0.3 24.4 ± 1.5 6.8 ± 0.3 0.50 0.28 5.5 11.3 ± 0.7 24.6 ± 2.6 5.6 ± 0.8 0.46 0.22 6.5 8.7 ± 0.5 28.3 ± 1.5 6.0 ± 0.6 0.31 0.28
Note: Cultures carried out in shake fl asks for 14 days in darkness at 30ºC and 120 rpm.
Some of the known polysaccharides extracted from higher Basidiomycetes, which demonstrate carcinostatic activities, are all glucans. For exam ple, in G. lucidum, the 1,3-β-D-glucan polysaccharide promises to be a new type of carcinostatic agent, which might be useful in immunotherapy. 28 Fur-ther studies are needed to assess the resolution of the structure, molecular mass, and bioactivities of
extracellular polysaccharides produced in shake fl asks and fermentors by H. coffeata cultures.
Figure 1 shows the growth kinetics of H. cof-feata cultures carried out in conventional shake fl asks at 30°C, 120 rpm, and in darkness, using an optimized culture medium (pH 4.5, lactose 50 g/L, and yeast extract 10 g/L). The highest fi nal biomass and EPS concentrations obtained were 15.5 g/L and

Volume 11, Issue 4, 2009 345
MYCELIAL SUBMERGED CULTURE OF HUMPHREYA COFFEATA FOR THE PRODUCTION OF VALUABLE METABOLITES
FOR AUTHOR U
SE ONLY
6.8 g/L, respectively, consuming almost 30 g/L of lactose. In Table 1, an EPS production of 6.8 ± 0.3 g/L at day 14 of culture is reported. However, in Figure 1, the EPS production on day 14 was 6.1 g/L; and on days 15, 16, and 17 the EPS production was 6.8, 6.7, and 6.9 g/L, respectively, showing a production[[AU: “PRODUCTIVE”?]] stationary phase by the end of the culture. These results show that it is likely that in the fi rst trial of experiments (shown in Table 1), this stationary phase was reached one day earlier than those phases presented in Figure 1.
In a 5.0-L stirred tank bioreactor and an opti-mized medium, Kim et al.45 reported a maximum EPS concentration of up to 3.0 g/L and a maximum mycelial Agrocybe cylindracea growth of up to 11.0 g/L. Moreover, in G. lucidum shake fl ask cultures using feeding strategies and optimized culture media, the maximum biomass and EPS concentrations were up to 16.0 g/L and 6.0 g/L, respectively.46 The data obtained from our experi-ments, as shown in Figures 1 and 3 and Table 1, are within the same scale of magnitude as the data reported in the literature,15,16,28,45,46 showing H. cof-feata as an excellent candidate for the production of bioactive metabolites.
C. Cytotoxic, Proliferative and Genotoxic Effects of Humphreya coffeata on Human Cells [[AU: SHOULD COMMAS BE REMOVED
FROM HIGHLIGHTED/UNDERLINED TEXT, AS YOU REQUEST ON
PAGE 338?]]
Recently reported data suggest that a variety of compounds derived from higher Basidiomycetes, such as Sarcodon scabrosus,47 G. lucidum,48 and Leucopaxillus giganteus,49 have antiproliferative and cytotoxic activities on human cervical cancer cells. Thus, to determine the effect of H. coffeata on cervical tumor cells, two cervical cancer-derived cells lines, HeLa and InBl, along with a nontumori-genic human cell line (HaCat), were incubated with different concentrations of the lyophilized H. cof-feata extracellular fi ltrate (30, 60, 120, 250, 500, 1000, 3000, 6000, 12,000, 25,000, and 50,000 μg/mL). As shown in Figure 4A, a signifi cant cyto toxic effect (more than 50% cytotoxicity) was observed by incubating cervical tumor cells with high concentra-tions of H. coffeata extracellular fi ltrate (≥ 25,000
μg/mL). A similar cytotoxic effect was observed in the control, nontumorigenic HaCat cells, when they were incubated with high concentrations of the fi ltrate (12,000–50,000 μg/mL) (Fig. 4B). In contrast, incubation with low concentrations of H. coffeata extracellular fi ltrate (up to 250 μg/mL) seemed to increase the proliferation of tumor cells. To study this effect in more detail, a time-course experiment was carried out by incubating the cells with low concentrations of H. coffeata extracellular fi ltrate (10–250 μg/mL) for 96 hours (Fig. 2). After incubation for 48 hours, the proliferation level of tumor cells was signifi cantly higher than that ob-served in the control cells in the presence of 250 μg/mL of H. coffeata extracellular fi ltrate (Fig. 2). Interestingly, after 96 hours, the H. coffeata fi ltrate induced an increase up to 40% in the proliferation of tumor cells but not of nontumorigenic control cells at different concentrations (50–250 μg/mL). Moreover, it is reported that alcohol extracts and acid hydrolysates of fi ve higher Basidiomycetes species (Coprinus comatus, Agaricus campestris, A. bisporus, Lentinus edodes, and Tricholoma matsutake) also positively effected the proliferation of HeLa cells.50
There are also a number of reports showing that extracts of some Basidiomycetes have activity against hematologic malignancies, such as leukemia, lymphoma, and multiple myeloma.17,51–54 Therefore, to explore the effect of H. coffeata on human leu-kemia, different concentrations (250, 500, 1000, 1500, and 2500 μg/mL) of H. coffeata extracellular fi ltrate were incubated with Jurkat cells. As shown in Figure 4C, a dose-dependent cytotoxic effect was observed after a 48-hour culture. After 96 hours, 80% cytotoxicity was observed for all the H. coffeata fi ltrate concentrations assayed (Fig. 4D). Interest-ingly, none of the fi ltrate concentrations that showed a cytotoxic effect on Jurkat cells (250–2500 μg/mL) were toxic for control HaCat cells, suggesting that H. coffeata antitumoral activity might be specifi c for haematologic malignancies, as previously reported for other higher Basidiomycetes.51
With the aim to verify that osmotic equilibrium is not altered by the addition of lyophilized culture fi ltrates, we measured the conductivity of differ-ent concentrations of the lyophilized H. coffeata extra cellular fi ltrate solubilized on culture medium. That is because conductivity and osmotic pressure
346 International Journal of Medicinal Mushrooms
S. M. PORRAS-ARBOLEDA ET AL.
FOR AUTHOR U
SE ONLY
increase with concentration in a rather linear man-ner. In the range of 0 to 15,000 μg/mL, there are no signifi cant differences in conductivity (16.0 ± 1.0 mmho) where cytotoxicity effects were more pronounced on all cell lines. However, at higher concentrations of H. coffeata extracellular fi ltrate of 20,000 and 50,000 μg/mL, conductivity increases to 19 mmho and 23.5 mmho, respectively.
Our results suggest that H. coffeata has cyto-toxic activity on Jurkat cells. Thus, to investigate if cell death was mediated by a genotoxic effect, the cells were incubated with different concentra-tions of H. coffeata fi ltrate (250, 1000, and 2500 μg/mL), and genotoxicity was evaluated by using the Comet Assay. Results are shown in Figures 4E and 4F.
A concentration-dependent genotoxic effect was observed when Jurkat cells were incubated with different concentrations of H. coffeata culture extracts (Figs. 4E and 5). In the lowest concentra-tion evaluated (250 μg/mL), no statistical difference was found when comparing the data to the negative control (cells incubated with DMSO), showing that no genotoxic activity was found at the lowest con-centration evaluated. No more than 40% of DNA damage was found when cells were exposed to the highest concentration (2500 μg/mL), compared to
80% for the positive control (cells incubated with H2O2). As can be seen, a slight protection of DNA was observed using the lowest concentration of 250 μg/mL and H2O2 as the genotoxic effector (Fig. 4F). Furthermore, other reports showed that G. lucidum has a genoprotective effect at a very low concentration (0.0001% w/v), but also produces slight damage to DNA at higher concentrations—for example, just 20% of DNA in the tail was found using 0.1% of hot water G. lucidum extracts with human lymphocytic DNA.55 For G. lucidum, DNA damage can be explained, at least in part, by the high hydrogen peroxide generated by the fungus in aqueous solution.55
Some reactive oxygen species (ROS) can be cytotoxic and also produce structural alterations in the DNA.56 Moreover, in mushroom cultures, the production of ROS and reactive nitrogen species (RNS) is commonly reported.57 The DNA damage shown in Figure 4E was probably due, at least in part, to the production of ROS and RNS or to the production of phenolic compounds in the Jurkat cells.58 On the other hand, at the lowest concentra-tion evaluated (250 μg/mL), the DNA protection is probably due to small amounts of ROS (produced by the mushroom in submerged cultures or induced by extracellular components, such as phenolic
FIGURE 5. Damage to DNA of Jurkat cells by culture fi ltrates of Humphreya coffeata: Comet Assay. (A) Intact DNA in negative control incubated in DMSO 5%. (B) Positive control cells incubated with H2O2 100 µM. (C, D) DNA dam-age using 2500 µg/mL of culture fi ltrate.

Volume 11, Issue 4, 2009 347
MYCELIAL SUBMERGED CULTURE OF HUMPHREYA COFFEATA FOR THE PRODUCTION OF VALUABLE METABOLITES
FOR AUTHOR U
SE ONLY
compounds), inducing a cellular adaptation that will benefi t Jurkat cells in terms of resistance to this challenge, or inducing proliferation, as is the case with cervical cancer cells. A previously reported concept of habituation to small levels of ROS has been called “hormesis.”55–59
D. Antioxidant Activity of Humphreya coffeata Extracts
The antioxidant activity of culture fi ltrates from days 4, 8, 12, 16, and 21 are shown (Table 2) by means of various antioxidant analytical methods. As is shown in Table 2, the percentage of DPPH scavenging increases almost 4.5 times by day 12 of culture, and later on this value seems to remain constant. Similar values of % DPPH scavenging were reported for four wild edible mushrooms, at a concentration of 0.5 mg/mL—Russula cyanoxan-tha, approximately 10%; Amanita rubescens, 60%; Suillus granulates, just 40%; and Boletus edulis, 60%.60 In hot water-extracted polysaccharides from Ganoderma tsugae culture fi ltrates, the scavenging abilities on DPPH radicals can reach 100% at 20 mg/mL.38 Moreover, at 5 mg/mL, the scavenging abilities from hot water extracts of the same mush-room were in the range of 50% to 70%.61
In terms of EC50 of ABTS radical scaveng-ing activity, there are no signifi cant differences between the EC50 of culture fi ltrates at different culture ages. However, to the best of our know-
ledge, there are no data in the literature reporting the EC50 for ABTS, and, therefore, no comparison can be made. In vivo, the fi rst step of defense is based on the preventive antioxidants, by suppressing free radicals formation, which can be measured by the ABTS assay.62 And as observed in this work, H. coffeata culture supernatant does not appear to demonstrate an increase during submerged culture. However, the data for DPPH demonstrate that the exo-components of H. coffeata at the end of the culture can be used for scavenging free radicals, repressing chain initiation and/or breaking the chain of propagation reactions, as the second step in the line of defense of antioxidants.
A decrease in the concentration of extract to IC50(PMS)/NADH can be observed in Table 2, showing important antioxidant activity in the cap-ture of superoxide radicals. This activity is related to the stationary cell growth phase, during which H. coffeata probably produces certain enzymes during its secondary metabolism, which in nature are involved in ligninolytic activities capable of breaking the lignin and comprise reactions catalyzed by oxidases and peroxidases,63 thereby producing the whitening of wood. Although in this work H. coffeata is not attached to wood, it is assumed that it presents a similar responsive performance in submerged culture growing conditions, as is reported for some other fungi of the Basidiomyco-tine division.64,65
Data suggest that the antioxidant activity of taking up superoxide radicals is due to enzyme
TABLE 2Antioxidant Activity of Five Humphreya coffeata Culture Filtrates Measured by DPPH Scavenging, ABTS, and NADH
Scavenging DPPH EC50 ABTS EC50 NADHDay of culture (%) (mg/mL) (mg/mL)
4 6.23 29.21 82.12 8 N.D. 23.89 73.94 12 28.85 32.43 53.44 16 22.66 36.60 38.00 21 32.13 35.36 24.17
Note: Cultures carried out in shake fl asks for 21 days in darkness at 30ºC and 120 rpm. N.D. = not determined.

348 International Journal of Medicinal Mushrooms
S. M. PORRAS-ARBOLEDA ET AL.
FOR AUTHOR U
SE ONLY
systems such as superoxide dismutase, laccases, and peroxidases that might be present during the stationary phase. The elucidation of the specifi c mechanisms of this fungus becomes a challenge for further studies.
IV. CONCLUSIONS
In this study, a culture medium was designed and optimized for mycelial submerged culture of a new strain of H. coffeata from the Southern Hemisphere, so as to obtain an effi cient production of bioactive compounds, mainly EPS and those with cytotoxic, genotoxic, and antioxidant activities. The effect of the carbon source and the initial lactose concen-tration on the cell growth and the EPS production was studied, showing comparable results to that of other higher Basiomycetes. The optimal carbon, nitrogen, and pH conditions for the cell growth and the EPS accumulation were identifi ed, and the culture fi ltrates were evaluated for bioactivity. We demonstrated that H. coffeata culture fi ltrates have a dramatic cytotoxic effect on lymphoma cells, but no effect on nontumorigenic keratinocytes and cervical cancer cell lines, at similar concentrations, were found. Moreover, proliferative effects on cervical cancer cell lines at very low concentrations were presented. Data show correspondence to the partial genoprotective effect at very low concentrations on lymphoma cells. However, a careful study of the composition of the culture fi ltrates and other different extracts (hot water, ethyl ether, or metha-nolic extracts) is required to determine the com-ponents with important activities, their structural characterizations, and operating mechanisms. We are currently studying the effect of some culture parameters on productivity and on certain biologi-cal activities. We are also evaluating the structure, molecular mass, and bioactivities of extracellular polysaccharides produced in shake fl asks and fer-mentors by H. coffeata cultures.
ACKNOWLEDGMENTS
The authors thank Andrés Pareja-López (Univer-sidad Nacional de Colombia, Medellin Campus,
Colombia) for his technical assistance in genotox-icity and cytotoxicity assays of Jurkat cells, and Dr. Ana Esperanza Franco-Molano (Universidad de Antioquia, Medellín, Colombia) for her identifi ca-tion and classifi cation of H. coffeata. We also thank Ana Delgado for reviewing the English version of the manuscript.
REFERENCES
1. Zaidman BZ, Wasser SP, Nevo E, Mahajna J. Androgen receptor-dependent and -independent mechanisms medi-ate Ganoderma lucidum activities in LNCaP prostate cancer cells. Int J Oncol. 2007;31:959–67.
2. Wasser SP. Medicinal mushrooms as a source of anti-tumor and immunomodulating polysaccharides. Appl Microbiol Biotechnol. 2002;60:258–74.
3. Isikhuemhen OS, Moncalvo JM, Nerud F, Vilgalys R. Mating compatibility and phylogeography in Pleurotus tuberregium. Mycol Res. 2000;104:732–7.
4. Moncalvo JM, Buchanan PK. Molecular evidence for long distance dispersal across the Southern Hemisphere in the Ganoderma applanatum-australe species complex (Basidiomycota). Mycol Res. 2008;112:425–36.
5. Steyaert RL. Species of Ganoderma and related genera mainly of the Bagor and Leiden Herbaria. Persoonia. 1972;7:55–118.
6. Gao Y, Tang W, Dai X, Gao H, Chen G, Ye J, Chan E, Koh HL, Li X, Zhou S. Effects of water-soluble Ganoderma lucidum polysaccharides on the immune functions of patients with advanced lung cancer. J Med Food. 2005;8:159–68.
7. Cao QZ, Lin ZB. Ganoderma lucidum polysaccharides peptide inhibits the growth of vascular endothelial cell and the induction of VEGF in human lung cancer cell. Life Sci. 2006;78:1457–63.
8. Li CJ, Li YM, Sun HH. New ganoderic acids, bioactive triterpenoid metabolites from the mushroom Ganoderma lucidum. Nat Prod Res. 2006;20:985–91.
9. Liu J, Shimizu K, Kondo R. Ganoderic acid TR, a new lanostanoid with 5 alpha-reductase inhibitory activity from the fruiting body of Ganoderma lucidum. Nat Prod Comm. 2006;1:345–50.
10. Tang W, Liu HW, Zhao WM, Wei DZ, Zhong JJ. Ganoderic acid T from Ganoderma lucidum mycelia induces mitochondria mediated apoptosis in lung cancer cells. Life Sci. 2006;80:205–11.
11. Yang HL. Ganoderic acid produced from submerged culture of Ganoderma lucidum induces cell cycle arrest and cytotoxicity in human hepatoma cell line BEL7402. Biotechnol Lett. 2005;27:835–8.
12. Wang HX, Ng TB. Ganodermin, an antifungal pro-tein from fruiting bodies of the medicinal mushroom Ganoderma lucidum. Peptides. 2006;27:27–30.

Volume 11, Issue 4, 2009 349
MYCELIAL SUBMERGED CULTURE OF HUMPHREYA COFFEATA FOR THE PRODUCTION OF VALUABLE METABOLITES
FOR AUTHOR U
SE ONLY
13. Yamac M, Bilgili F. Antimicrobial activities of fruit bodies and/or mycelial cultures of some mushroom isolates. Pharma Biol. 2006;44:660–7.
14. Yang FC, Liau CB. Effects of cultivating conditions on the mycelial growth of Ganoderma lucidum in sub merged fl ask cultures. Bioprocess Eng. 1998;19:233–6.
15. Fang QH, Zhong JJ. Submerged fermentation of higher fungus Ganoderma lucidum for production of valuable bioactive metabolites—ganoderic acid and polysaccha-ride. Biochem Eng J. 2002;10:61–5.
16. Fang QH, Zhong JJ. Effect of initial pH on production of ganoderic acid and polysaccharide by submerged fermentation of Ganoderma lucidum. Process Biochem. 2002;37:769–74.
17. Yuen JWM, Gohel MDI. Anticancer effects of Gano-derma lucidum: a review of scientifi c evidence. Nutr Cancer. 2005;53:11–7.
18. Sliva D. Ganoderma lucidum (Reishi) in cancer treat-ment. Integr Cancer Ther. 2003;2:358–64.
19. Sliva D. Cellular and physiological effects of Gano-derma lucidum (Reishi). Mini Rev Med Chem. 2004;4:873–9.
20. Jong SC, Birmingham JM. Medicinal and therapeutic value of the shiitake mushroom. Adv Appl Microbiol. 1993;39:153–84.
21. Rhee SJ, Cho SY, Kim KM, Cha DS, Park HJ. A com-parative study of analytical methods for alkali-soluble beta-glucan in medicinal mushroom, Chaga (Inonotus obliquus). Lwt-Food Sci Technol. 2008;41:545–9.
22. Nakajima Y, Sato Y, Konishi T. Antioxidant small phe-nolic ingredients in Inonotus obliquus (Persoon) Pilat (Chaga). Chem Pharm Bull. 2007;55:1222–6.
23. Zou X. Optimization of nutritional factors for exopoly-saccharide production by submerged cultivation of the medicinal mushroom Oudemansiella radicata. World J Microbiol Biotechnol. 2005;21:1267–71.
24. Zaidman BZ, Yassin M, Mahajna J, Wasser SP. Medicinal mushroom modulators of molecular targets as can-cer therapeutics. Appl Microbiol Biotechnol. 2005;67:453–68.
25. Zhong JJ, Tang YJ. Submerged cultivation of medicinal mushrooms for production of valuable bioactive me-tabolites. Biomanufacturing. 2004;87:25–59.
26. Fang QH, Tang YJ, Zhong JJ. Signifi cance of inocula-tion density control in production of polysaccharide and ganoderic acid by submerged culture of Ganoderma lucidum. Proc Biochem. 2002;37:1375–9.
27. Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F. Colorimetric method for determination of sugars and related substances. Anal Chem. 1956;28:350–6.
28. Tang YJ, Zhong JJ. Fed-batch fermentation of Gano-derma lucidum for hyperproduction of polysaccharide and ganoderic acid. Enzyme Microb Technol. 2002;31:20–8.
29. Miller GL. Use of dinitrosalicilic acid reagent for de-termination of reducing sugar. Anal Chem. 1959;31(3):426–8.
30. Caceres-Cortes JR, Alvarado-Moreno JA, Waga K, Rangel-Corona R, Monroy-Garcia A, Rocha-Zavaleta L, Urdiales-Ramos J, Weiss-Steider B, Haman A, Hugo P, Brousseau R, Hoang T. Implication of tyrosine ki-nase receptor and steel factor in cell density-dependent growth in cervical cancers and leukemias. Cancer Res. 2001;61:6281–9.
31. Hu HB, Ahn NS, Yang XL, Lee YS, Kang KS. Ganoderma lucidum extract induces cell cycle arrest and apoptosis in MCF-7 human breast cancer cell. Int J Cancer. 2002;102:250–3.
32. Bernas T, Dobrucki J. Mitochondrial and nonmito-chondrial reduction of MTT: interaction of MTT with TMRE, JC-1, and NAO mitochondrial fl uorescent probes. Cytometry. 2002;47:236–42.
33. Ostling O, Johanson KJ. Microelectrophoretic study of radiation-induced DNA damages in individual mam-malian cells. Biochem Biophys Res Commun. 1984;123:291–8.
34. Singh NP, McCoy MT, Tice RR, Schneider EL. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp Cell Res. 1988;175:184–91.
35. Horner WE, Helbling A, McCants ML, Salvaggio JE, Lehrer SB. Allergenic activity of two species of Ganoderma (G). J Allergy Clin Immunol. 1991;87:184.
36. Olive PL, Banath JP, Durand RE. Detection of etoposide resistance by measuring DNA damage in individual Chinese hamster cells. J Natl Cancer Inst. 1990;82:779–83.
37. Hatano T, Kagawa H, Yasuhara T, Okuda T. Two new fl avonoids and other constituents in licorice root: their relative astringency and radical scavenging effects. Chem Pharm Bull. 1988;36:1097.
38. Tseng YH, Yang JH, Mau JL. Antioxidant properties of polysaccharides from Ganoderma tsugae. Food Chem. 2008;107:732–8.
39. Re R, Pellegrini N, Proteggente A, Pannala A, Yang M, Rice-Evans C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic Biol Med. 1999;26:1231–7.
40. Yamaguchi F, Ariga T, Yoshimura Y, Nakazawa H. Antioxidative and anti-glycation activity of garcinol from Garcinia indica fruit rind. J Agric Food Chem. 2000;48:180–5.
41. Mao XB, Eksriwong T, Chauvatcharin S, Zhong JJ. Optimization of carbon source and carbon/nitrogen ratio for cordycepin production by submerged cultiva-tion of medicinal mushroom Cordyceps militaris. Proc Biochem. 2005;40:1667–72.
42. Tang YJ, Zhong JJ. Exopolysaccharide biosynthesis and related enzyme activities of the medicinal fungus, Ganoderma lucidum, grown on lactose in a bioreactor. Biotechnol Lett. 2002;24:1023–6.
43. Xu P, Ding ZY, Qian Z, Zhao CX, Zhang KC. Improved production of mycelial biomass and ganoderic acid by submerged culture of Ganoderma lucidum SB97 using

350 International Journal of Medicinal Mushrooms
S. M. PORRAS-ARBOLEDA ET AL.
FOR AUTHOR U
SE ONLY
complex media. Enzyme Microb Technol. 2008;42:325–31.
44. Shih IL, Pan K, Hsieh CY. Infl uence of nutritional components and oxygen supply on the mycelial growth and bioactive metabolites production in submerged culture of Antrodia cinnamomea. Proc Biochem. 2006;41(5):1129–35.
45. Kim HO, Lim JM, Joo JH, Kim SW, Hwang HJ, Choi JW, Yun JW. Optimization of submerged culture condition for the production of mycelial biomass and exopolysaccharides by Agrocybe cylindracea. Bioresour Technol. 2005;96:1175–82.
46. Wagner R, Mitchell DA, Sassaki GL, de Almeida Amazonas MA. Links between morphology and physi-ology of Ganoderma lucidum in submerged culture for the production of exopolysaccharide. J Biotechnol. 2004;114:153–64.
47. Dong M, Chen SP, Kita K, Ichimura Y, Guo WZ, Lu S, Sugaya S, Hiwasa T, Takiguchi M, Mori N, Kashima A, Morimura K, Hirota M, Suzuki N. Anti-proliferative and apoptosis-inducible activity of Sarcodonin G from Sarcodon scabrosus in HeLa cells. Int J Oncol. 2009;34:201–7.
48. Guan SH, Xia JM, Yang M, Wang XM, Liu X, Guo DA. Cytotoxic lanostanoid triterpenes from Ganoderma lucidum. J Asian Nat Prod Res. 2008;10:705–10.
49. Ren G, Zhao YP, Yang L, Fu CX. Anti-proliferative effect of clitocine from the mushroom Leucopaxillus giganteus on human cervical cancer HeLa cells by inducing apoptosis. Cancer Lett. 2008;262:190–200.
50. Chung KS. The effects of mushroom components on the proliferation of HeLa cell line in vitro. Arch Pharm Res. 1979;2:25–33.
51. Muller CI, Kumagai T, O’Kelly J, Seeram NP, Heber D, Koeffl er HP. Ganoderma lucidum causes apoptosis in leukemia, lymphoma and multiple myeloma cells. Leuk Res. 2006;30:841–8.
52. Muller CI, Kumagai T, O’Kelly J, Seeram N, Heber D, Koeffl er HP. Anticancer activity and induction of multinucleation in hematologic cell lines mediated by Ganoderma lucidum. Blood. 2004;104(11):208B.
53. Xie YZ, Li SZ, Yee A, La Pierre DP, Deng Z, Lee DY, Wu QP, Chen Q, Li C, Zhang Z, Guo J, Jiang Z, Yang BB. Ganoderma lucidum inhibits tumour cell prolifera-tion and induces tumour cell death. Enzyme Microb Technol. 2006;40:177–85.
54. Wu DY, Pae MK, Ren ZH, Guo ZY, Smith D, Meydani
SN. Dietary supplementation with white button mush-room enhances natural killer cell activity in C57BL/6 mice. J Nutr. 2007;137:1472–7.
55. Wachtel-Galor S, Choi SW, Benzie IFF. Effect of Ganoderma lucidum on human DNA is dose depen-dent and mediated by hydrogen peroxide. Redox Rep. 2005;10:145–9.
56. Wiseman H, Halliwell B. Damage to DNA by reac-tive oxygen and nitrogen species: role in infl amma-tory disease and progression to cancer. Biochem J. 1996;313:17–29.
57. Acharya K, Yonzone P, Rai M, Rupa A. Antioxidant and nitric oxide synthase activation properties of Ganoderma applanatum. Indian J Exp Biol. 2005;43:926–9.
58. Wei S, Helsper JPFG, Van Griensven LJLD. Phenolic compounds present in medicinal mushroom extracts generate reactive oxygen species in human cells in vitro. Int J Med Mushr. 2008;10:1–13.
59. Finkel T, Holbrook NJ. Oxidants, oxidative stress and the biology of ageing. Nature. 2000;408:239–47.
60. Ribeiro B, Lopes R, Andrade PB, Seabra RM, Goncalves RF, Baptista P, Quelhas I, Valentao P. Comparative study of phytochemicals and antioxidant potential of wild edible mushroom caps and stipes. Food Chem. 2008;110:47–56.
61. Mau JL, Tsai SY, Tseng YH, Huang SJ. Antioxidant properties of hot water extracts from Ganoderma tsugae Murrill. LWT - Food Sci Technol. 2005;38:589–97.
62. Lakshmi B, Tilak JC, Adhikari S, Devasagayam TPA, Janardhanan KK. Evaluation of antioxidant activity of selected Indian mushrooms. Pharma Biol. 2004;42:179–85.
63. Belinky PA, Goldberg D, Krinfeld B, Burger M, Rothschild N, Cogan U, Dosoretz C. Manganese-containing superoxide dismutase from the white-rot fungus Phanerochaete chrysosporium: its function, expression and gene structure. Enzyme Microb Technol. 2002;31:754–64.
64. Bockle B, Martinez MJ, Guillen F, Martinez AT. Mechanism of peroxidase inactivation in liquid cultures of the ligninolytic fungus Pleurotus pulmonarius. Appl Environ Microbiol. 1999;65:923–8.
65. Rice-Evans CA. Free radicals and antioxidants in nor-mal and pathological processes. In: Rice-Evans CA, Bruckdorfer KR, editors. Oxidative stress, lipoproteins and cardiovascular dysfunction. London: London Portland Press; 2005. p. 1–32.
Copyright © 2022 FDOKUMEN




















