
Involvement of mitochondrial permeability transition pore opening in alpha-bisabolol induced...
11
Involvement of mitochondrial permeability transition pore opening in a-bisabolol induced apoptosis Elisabetta Cavalieri 1, *, Christian Bergamini 2, *, Sofia Mariotto 1 , Serena Leoni 2 , Luigi Perbellini 3 , Elena Darra 1 , Hisanori Suzuki 1 , Romana Fato 2 and Giorgio Lenaz 2 1 Dipartimento di Scienze Morfologico-Biomediche, Universita ` di Verona, Italy 2 Dipartimento di Biochimica ‘G. Moruzzi’, Universita ` di Bologna, Italy 3 Dipartimento di Medicina e Sanita ` Pubblica, Universita ` di Verona, Italy Introduction a-Bisabolol is a small oily sesquiterpene (m, 222.37 Da), widely present in plants, shrubs and trees. It has been reported to reduce inflammation and arthritis, prevent the development of gastric ulcers and act as an antibacterial and antifungal agent. It also exhibits wound-healing activity and is used in cosmetic products to treat dermatitis [1–5]. More recently, insec- ticidal effects of this oil have also been reported [6]. An antitumor action of a-bisabolol has only recently been discovered [7]. Indeed, we reported that Keywords a-bisabolol; apoptosis; cyclosporine A; mitochondria; mitochondrial permeability transition pore Correspondence H. Suzuki, Dipartimento di Scienze Morfologico-Biomediche, Universita ` di Verona, Strada Le Grazie, 8, 37134 Verona, Italy Fax: +39 045 802 7170 Tel: +39 045 802 7167 E-mail: [email protected] *These authors contributed equally to this work (Received 16 April 2009, revised 15 May 2009, accepted 21 May 2009) doi:10.1111/j.1742-4658.2009.07108.x a-Bisabolol is a natural monocyclic sesquiterpene alcohol. It has been used in cosmetics for hundreds of years because of its perceived skin-healing properties. a-Bisabolol is known to have anti-irritant, anti-inflammatory and antimicrobial properties. In precedent studies, we described how a-bisabolol exerts a selective pro-apoptotic action towards transformed cells [Cavalieri E et al. (2004) Biochem Biophys Res Commun 315, 589–594] and its uptake is mediated by lipid rafts on the plasma membrane [Darra E et al. (2008) Arch Biochem Biophys 476, 113–123]. In this study, we hypothesize that the intracellular target of a-bisabolol may be the mitochondrial perme- ability transition pore (mPTP). To evaluate this hypothesis, we used one transformed cell line (human glioma T67) in comparison with a nontrans- formed one (human fibroblasts). We assessed the effect of a specific mPTP inhibitor (cyclosporine A) on the toxic action of a-bisabolol. Results show that the a-bisabolol-induced decrease in oxygen consumption is abolished by the addition of cyclosporine A in T67 cells, indicating that a-bisabolol may target mPTP. The central role of mitochondria was also demonstrated by using galactose to force cells to a more aerobic metabolism. In this con- dition, we observed higher a-bisabolol toxicity. Furthermore, we studied the effect of a-bisabolol on isolated rat liver mitochondria. This study expands the notion of the specific action of a-bisabolol on transformed cells and sug- gests that it may act by disturbing the structure and function of the mPTP. a-Bisabolol toxicity is clearly related to its cellular uptake, which is higher in transformed cell lines. Abbreviations CsA, cyclosporine A; DCF-DA, dichlorodihydrofluorescein diacetate; DMEM, Dulbecco’s modified Eagle’s medium; FCCP, carbonyl cyanide p-(trifluoromethoxy)-phenylhydrazone; mPTP, mitochondrial permeability transition pore; NAC, N-acetyl-cysteine; PARP, poly(ADP-ribose) polymerase; ROS, reactive oxygen species; TMRM, tetramethylrodamine methyl ester. 3990 FEBS Journal 276 (2009) 3990–4000 ª 2009 The Authors Journal compilation ª 2009 FEBS
Transcript of Involvement of mitochondrial permeability transition pore opening in alpha-bisabolol induced...

Involvement of mitochondrial permeability transition poreopening in a-bisabolol induced apoptosisElisabetta Cavalieri1,*, Christian Bergamini2,*, Sofia Mariotto1, Serena Leoni2, Luigi Perbellini3,Elena Darra1, Hisanori Suzuki1, Romana Fato2 and Giorgio Lenaz2
1 Dipartimento di Scienze Morfologico-Biomediche, Universita di Verona, Italy
2 Dipartimento di Biochimica ‘G. Moruzzi’, Universita di Bologna, Italy
3 Dipartimento di Medicina e Sanita Pubblica, Universita di Verona, Italy
Introduction
a-Bisabolol is a small oily sesquiterpene (m, 222.37 Da),
widely present in plants, shrubs and trees. It has been
reported to reduce inflammation and arthritis, prevent
the development of gastric ulcers and act as an
antibacterial and antifungal agent. It also exhibits
wound-healing activity and is used in cosmetic
products to treat dermatitis [1–5]. More recently, insec-
ticidal effects of this oil have also been reported
[6]. An antitumor action of a-bisabolol has only
recently been discovered [7]. Indeed, we reported that
Keywords
a-bisabolol; apoptosis; cyclosporine A;
mitochondria; mitochondrial permeability
transition pore
Correspondence
H. Suzuki, Dipartimento di Scienze
Morfologico-Biomediche, Universita di
Verona, Strada Le Grazie, 8, 37134 Verona,
Italy
Fax: +39 045 802 7170
Tel: +39 045 802 7167
E-mail: [email protected]
*These authors contributed equally to this
work
(Received 16 April 2009, revised 15 May
2009, accepted 21 May 2009)
doi:10.1111/j.1742-4658.2009.07108.x
a-Bisabolol is a natural monocyclic sesquiterpene alcohol. It has been used
in cosmetics for hundreds of years because of its perceived skin-healing
properties. a-Bisabolol is known to have anti-irritant, anti-inflammatory
and antimicrobial properties. In precedent studies, we described how
a-bisabolol exerts a selective pro-apoptotic action towards transformed cells
[Cavalieri E et al. (2004) Biochem Biophys Res Commun 315, 589–594] and
its uptake is mediated by lipid rafts on the plasma membrane [Darra E et al.
(2008) Arch Biochem Biophys 476, 113–123]. In this study, we hypothesize
that the intracellular target of a-bisabolol may be the mitochondrial perme-
ability transition pore (mPTP). To evaluate this hypothesis, we used one
transformed cell line (human glioma T67) in comparison with a nontrans-
formed one (human fibroblasts). We assessed the effect of a specific mPTP
inhibitor (cyclosporine A) on the toxic action of a-bisabolol. Results show
that the a-bisabolol-induced decrease in oxygen consumption is abolished
by the addition of cyclosporine A in T67 cells, indicating that a-bisabololmay target mPTP. The central role of mitochondria was also demonstrated
by using galactose to force cells to a more aerobic metabolism. In this con-
dition, we observed higher a-bisabolol toxicity. Furthermore, we studied the
effect of a-bisabolol on isolated rat liver mitochondria. This study expands
the notion of the specific action of a-bisabolol on transformed cells and sug-
gests that it may act by disturbing the structure and function of the mPTP.
a-Bisabolol toxicity is clearly related to its cellular uptake, which is higher
in transformed cell lines.
Abbreviations
CsA, cyclosporine A; DCF-DA, dichlorodihydrofluorescein diacetate; DMEM, Dulbecco’s modified Eagle’s medium; FCCP, carbonyl cyanide
p-(trifluoromethoxy)-phenylhydrazone; mPTP, mitochondrial permeability transition pore; NAC, N-acetyl-cysteine; PARP, poly(ADP-ribose)
polymerase; ROS, reactive oxygen species; TMRM, tetramethylrodamine methyl ester.
3990 FEBS Journal 276 (2009) 3990–4000 ª 2009 The Authors Journal compilation ª 2009 FEBS

a-bisabolol is able to kill by apoptosis human and
mouse glioma cell lines such as U87, T67 and C6,
which are highly resistant to common antitumor treat-
ments. a-Bisabolol, at a concentration of 2 lm, which
is estimated to be the EC50 value, induces rapid apop-
tosis in these cells. At higher concentrations (10 lm),
a-bisabolol requires only a few hours to completely kill
these cell lines. a-Bisabolol quickly induces caspase-3
activation, poly(ADP-ribose) polymerase (PARP)
cleavage and DNA ladder formation, which are associ-
ated with an increase in the hypo-G1 fraction of the
cell population [7].
Because neither the antagonist anti-Fas IgG nor the
caspase-8 inhibitor block a-bisabolol-induced cell
death, a-bisabolol has been presumed to trigger apop-
tosis in a Fas-receptor-independent manner, through
the intrinsic (i.e. mitochondrially related) pathway [7].
Furthermore, treatment of cells with a-bisabolol rap-
idly induces loss of mitochondrial inner transmem-
brane potential (DWm) and the release of cytochrome c
from mitochondria.
Another striking feature of a-bisabolol is that it is
less toxic to normal cells, such as astroglial cells [7],
than to transformed cells, in line with its nontoxicity
in adult mice and rats (reported LD50 values are
14–15 gÆkg)1) [8].
Apoptosis is a physiological, energy-requiring pro-
cess that is genetically programmed [9]. Tumor cells
are believed to be more resistant to apoptosis than
normal cells [10,11]. Unfortunately, a majority of the
antitumor compounds reported to date induce apopto-
sis indiscriminately in normal and tumor cells [12]. A
preferential apoptotic action of a-bisabolol towards
tumor cells, therefore, should represent a remarkable
advantage for therapeutics.
The higher toxicity of a-bisabolol towards tumor cell
lines may be related to a different uptake efficiency
between normal and tumor cells that might be medi-
ated by plasma membrane lipid composition. Because
a-bisabolol is a quite hydrophobic compound, it is rea-
sonable to hypothesize that the lipid composition of
the plasma membrane may play a crucial role in its
uptake. In a recent study, Darra et al. [13] described
how a-bisabolol can be rapidly incorporated into lipid
rafts, highly dynamic membrane domains enriched
with sterol and sphingolipid that play an important
role in intracellular protein transport and membrane
fusion [14]. In this regard, it has been reported that
the plasma membranes of tumor cells contain many
more lipid rafts than the plasma membranes of normal
cells. Moreover, Darra et al. [13] have demonstrated
that a-bisabolol induces the transfer of Bid from cyto-
plasm to lipid rafts and interacts with Bid in a highly
specific way, suggesting that Bid may be directly
involved in a-bisabolol transport from lipid rafts to
the mitochondrial membrane.
In this study, we confirmed the high a-bisabololuptake in a T67 glioma cell line in comparison with
normal fibroblasts and we investigated the molecular
mechanism by which a-bisabolol efficiently and rapidly
induces apoptosis in T67 cells. Based on the previous
observation that a-bisabolol induces apoptosis throughthe mitochondrial intrinsic pathway, rather than the
receptor-mediated pathway, we analyzed the relation
between mitochondrial damage induced by a-bisabololand the viability of the human glioblastoma cell line
and human fibroblasts.
Because cytochrome c release is an early event
induced by treatment with a-bisabolol and could be
mediated by mitochondrial permeability transition
pore (mPTP) opening or by members of the Bcl-2 fam-
ily, we analyzed both the effect of cyclosporine A
(CsA), a well-characterized inhibitor of mPTP opening,
and the amount of Bax and Bcl-2 on a-bisabololaction in glioblastoma cells and fibroblasts.
Bax and Bcl-2 have opposite effects on apoptotic
cell death: the first induces cell death and the second
inhibits it. Bax is a cytosolic, monomeric protein
that, following a death stimulus, translocates from
the cytoplasm to the mitochondria, where it becomes
an integral membrane protein. Bcl-2 is an integral
membrane protein, heavily localized in the mito-
chondria [15].
Results
a-Bisabolol uptake is higher in human glioma cell
line T67 than in fibroblasts
Our previous results [7] showed that 5 lm a-bisabololis able to induce apoptotic cell death in all examined
malignant cell lines and is far less effective in killing
human normal cells. Table 1 shows the effect of 5 lm
a-bisabolol on cell viability, DNA ladder and PARP
cleavage in different cell lines. The data clearly show
that a-bisabolol toxicity is higher in malignant than
nontransformed cell lines; this behavior may be related
to differences in cellular uptake. In fact, intracellular
concentration analysis shows that in the T67 cell line,
a-bisabolol is rapidly distributed in the membrane,
cytosolic and nuclear fractions, whereas in fibroblasts
its intracellular distribution is significative lower
(Fig. 1). These results are in line with previous
data obtained in other cell lines [13] and suggest that
a-bisabolol, once absorbed into plasma membrane
lipid rafts, is transported inside cells.
E. Cavalieri et al. a-Bisabolol, an efficient apoptosis inducer
FEBS Journal 276 (2009) 3990–4000 ª 2009 The Authors Journal compilation ª 2009 FEBS 3991

a-Bisabolol-induced apoptosis is independent of
Bax and Bcl-2 and of reactive oxygen species
Cytochrome c release and loss of mitochondrial mem-
brane potential are early events in the apoptotic cell
death induced by a-bisabolol treatment [7]; both events
may be related to a change in the amounts of pro- ⁄antiapoptotic proteins of the Bcl-2 family or to a direct
involvement of the mPTP.
To investigate the molecular mechanism involved in
a-bisabolol toxicity, we examined the change in the
amounts of Bcl-2 family proteins and the role of
mPTP in a-bisabolol-induced apoptosis. Expression of
these two proteins in T67 cells was examined by wes-
tern blotting after a-bisabolol treatment over periods
of 15–60 min, corresponding to the period in which we
observed cytochrome c release [7]. As shown in Fig. 2,
there was no change in the amounts of cytosolic and
mitochondrial Bax, or in total Bcl-2 proteins at any
time during treatment.
However, cell treatment with CsA, a specific inhibi-
tor of mPTP, induced an increase in cell survival in
the presence of a-bisabolol, as confirmed by the retain-
ing of membrane potential (Fig. 7A,B). Because mPTP
opening can be induced by reactive oxygen species
(ROS), we studied their contribution to a-bisabolol-induced apoptosis. For this purpose, we measured the
ROS level in both control and a-bisabolol-treated T67
and fibroblast cell lines. Treatment with 5 lm a-bisab-olol increases the ROS level in T67 cells. This effect
was counteracted by adding 10 mm N-acetyl-cysteine
(NAC). Moreover, we did not observe any increase in
ROS levels in treated fibroblasts (Fig. 3A). Neverthe-
less, in T67 cells, NAC did not prevent PARP cleavage
or DNA ladder formation after 3 or 5 h of a-bisabololtreatment (Fig. 3B,C). These data indicate that oxida-
tive damage, although present, is not involved in the
apoptotic process induced by a-bisabolol.
a-Bisabolol affects oxygen consumption in intact
cells and rat liver mitochonria
Mitochondrial involvement in a-bisabolol-induced cell
death was confirmed by measuring oxygen consump-
tion by intact cells. Because this (Table 1) and previous
studies [7] have shown that the toxic effect is higher in
cancer cells than in normal cells, we measured oxygen
consumption in two different cell types: T67 and fibro-
blasts. We measured the endogenous respiration rate
in the presence of glucose. The maximum respiration
rate was achieved by adding carbonyl cyanide p-(triflu-
oromethoxy)-phenylhydrazone (FCCP) as an electron
transport uncoupler.
Figure 4A shows that the oxygen consumption in T67
cells exposed to 5 lm a-bisabolol for 20 min was � 50%
Table 1. a-Bisabolol (5 lM) induced apoptotic cell death in a
number of transformed and normal human cells. PARP,
poly(ADP-ribose) polymerase.
Cells
% Cell
deatha
DNA
ladderb
PARP
cleavagec
T67 glioblastoma 90 + +
U87 glioblastoma 60 + +
MDA-MB231 breast cancer 86 + +
LNCap prostate cancer 73 + +
PC3 prostate cancer 90 + +
HL-60 leukemia 67 + +
Lymphocytes T normal cell 2 ) )Neutrophiles normal cell 1 ) )Fibroblasts normal cell 20 ) )
a Cell viability was examined after 5 h. b DNA ladder formation was
examined after 5 h. c PARP cleavage was analyzed after 3 h.
Fig. 1. Measurement of a-bisabolol concentration in T67 cells and
fibroblasts. Concentration of a-bisabolol in human fibroblasts and
T67 cell lines after 30 min of treatment. a-Bisabolol measured in
cytosolic, membrane and nuclear fractions using the gas chroma-
tography method. Data are presented as ± SD of six independent
measurements.
Fig. 2. Mitochondrial and cytosolic levels of Bax and Bcl-2 are unaf-
fected by a-bisabolol treatment. T67 cells were treated with 5 lM
a-bisabolol or with vehicle solvent over a period of 15–60 min. The
amounts of cytosolic and mitochondrial Bax and total Bcl-2 were
visualized by western blot using anti-Bax and anti-Bcl-2 IgG, respec-
tively. The results are indicative of three independents blots.
a-Bisabolol, an efficient apoptosis inducer E. Cavalieri et al.
3992 FEBS Journal 276 (2009) 3990–4000 ª 2009 The Authors Journal compilation ª 2009 FEBS
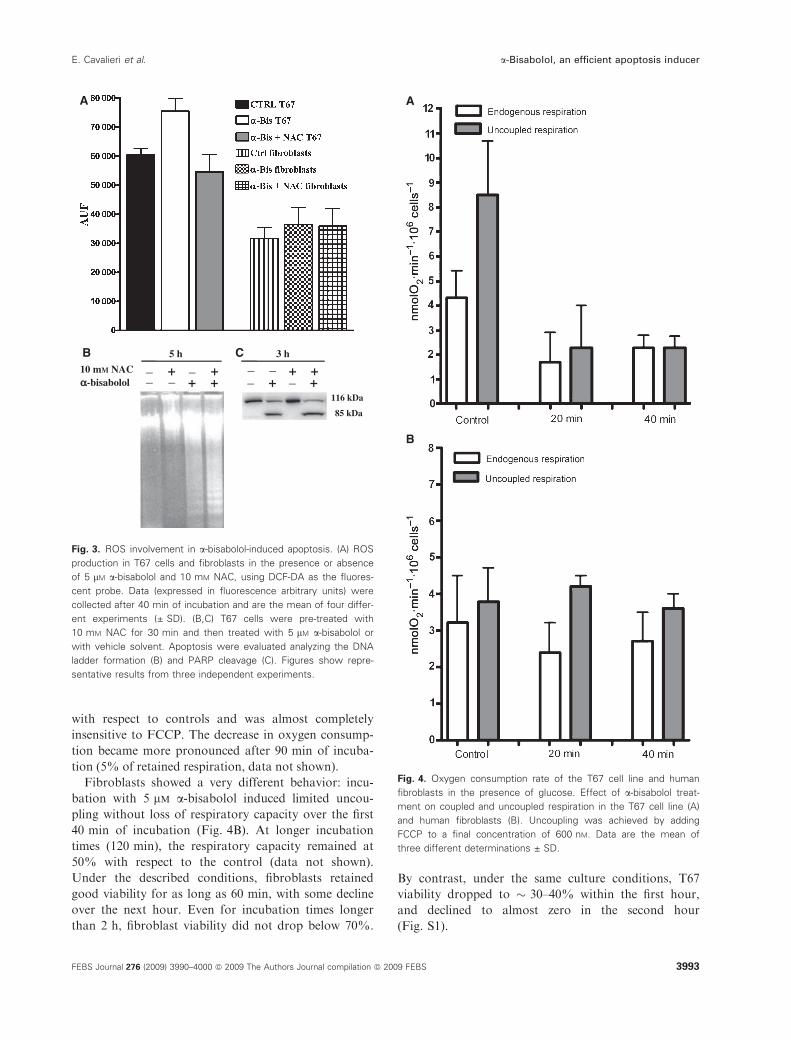
with respect to controls and was almost completely
insensitive to FCCP. The decrease in oxygen consump-
tion became more pronounced after 90 min of incuba-
tion (5% of retained respiration, data not shown).
Fibroblasts showed a very different behavior: incu-
bation with 5 lm a-bisabolol induced limited uncou-
pling without loss of respiratory capacity over the first
40 min of incubation (Fig. 4B). At longer incubation
times (120 min), the respiratory capacity remained at
50% with respect to the control (data not shown).
Under the described conditions, fibroblasts retained
good viability for as long as 60 min, with some decline
over the next hour. Even for incubation times longer
than 2 h, fibroblast viability did not drop below 70%.
By contrast, under the same culture conditions, T67
viability dropped to � 30–40% within the first hour,
and declined to almost zero in the second hour
(Fig. S1).
B
A
C
++ __10 mM NAC
5 h 3 h _ +_ +
116 kDa
85 kDa
+ + + +__αα-bisabolol _ _
Fig. 3. ROS involvement in a-bisabolol-induced apoptosis. (A) ROS
production in T67 cells and fibroblasts in the presence or absence
of 5 lM a-bisabolol and 10 mM NAC, using DCF-DA as the fluores-
cent probe. Data (expressed in fluorescence arbitrary units) were
collected after 40 min of incubation and are the mean of four differ-
ent experiments (± SD). (B,C) T67 cells were pre-treated with
10 mM NAC for 30 min and then treated with 5 lM a-bisabolol or
with vehicle solvent. Apoptosis were evaluated analyzing the DNA
ladder formation (B) and PARP cleavage (C). Figures show repre-
sentative results from three independent experiments.
A
B
Fig. 4. Oxygen consumption rate of the T67 cell line and human
fibroblasts in the presence of glucose. Effect of a-bisabolol treat-
ment on coupled and uncoupled respiration in the T67 cell line (A)
and human fibroblasts (B). Uncoupling was achieved by adding
FCCP to a final concentration of 600 nM. Data are the mean of
three different determinations ± SD.
E. Cavalieri et al. a-Bisabolol, an efficient apoptosis inducer
FEBS Journal 276 (2009) 3990–4000 ª 2009 The Authors Journal compilation ª 2009 FEBS 3993

In experiments performed on isolated rat liver mito-
chondria, we observed that the decrease in oxygen con-
sumption induced by a-bisabolol treatment was not
related to a direct inhibitory effect on the mitochon-
drial respiratory enzymes. In fact, the respiration rate
observed in the uncoupled condition was insensitive to
a-bisabolol treatment (Table 2). The decrease in state 3
respiration in the presence of a-bisabolol is partially
counteracted by adding more ADP. This behavior sug-
gests adenine nucleotide translocator or ATP syntase
impairment. Preliminary results show that a-bisabololcan affect adenine nucleotide translocator activity
(data not shown).
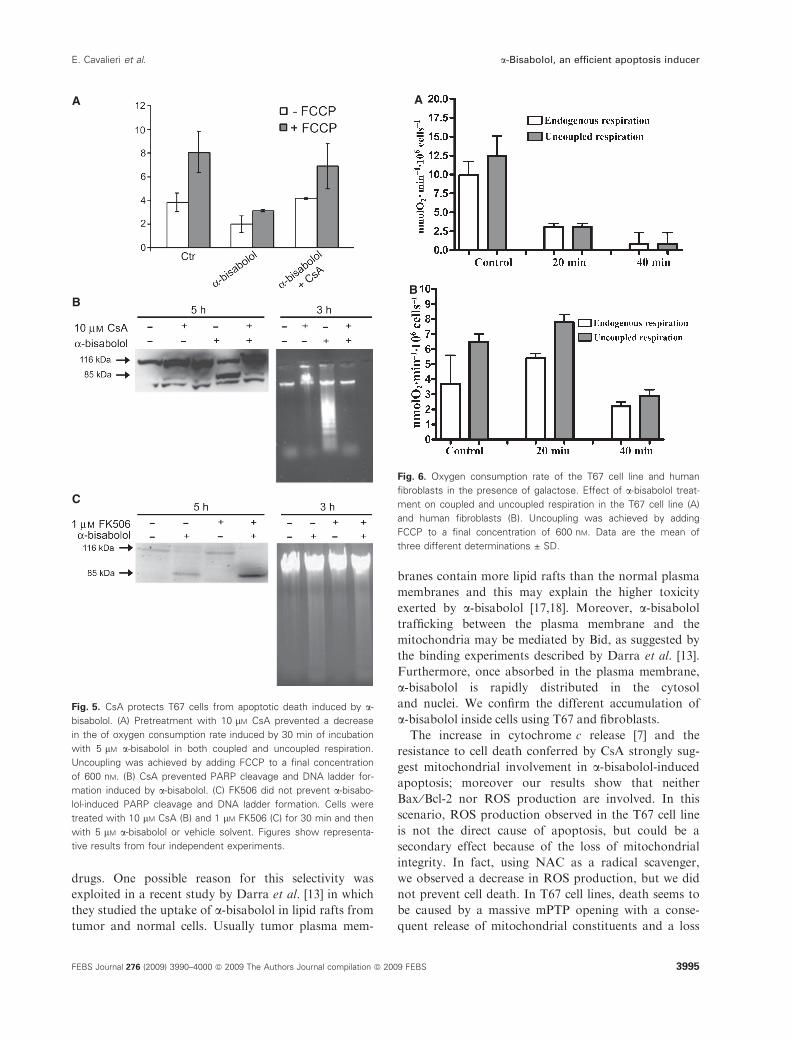
To further test whether the toxic action of a-bisabo-lol affects mPTP, we examined the effect of CsA on
cellular respiration and apoptosis in T67 cells.
Figure 5A shows that the toxic effect of a-bisabolol oncellular respiration was abolished by the presence of
10 lm CsA in the culture medium. Moreover, T67 cells
preincubated with CsA and then exposed to 5 lm a-bi-sabolol, showed no PARP cleavage after 3 h and no
DNA ladder formation after 5 h (Fig. 5B). At this
concentration, CsA has numerous nonspecific effects,
including inhibition of calcineurin, a protein phospha-
tase differently involved in apoptotic process. There-
fore, to exclude that the effect of a-bisabolol could be
mediated by calcineurin, the same experiments were
performed in the presence of the calcineurin inhibitor
FK506. Pretreatment of T67 cells with 1 lm FK506
for 30 min was not able to prevent the apoptotic effect
of a-bisabolol (Fig. 5C).
a-Bisabolol toxicity is higher under aerobic
metabolic conditions
To confirm the involvement of mitochondria in
a-bisabolol toxicity, we pushed cells towards more
aerobic metabolism by growing them in Dulbecco’s
modified Eagle’s medium (DMEM) containing galac-
tose instead of glucose. Galactose is a carbon source
known to favor the oxidative phosphorylation path-
way [16]. T67 cells cultured in DMEM–galactose
showed measurably greater oxygen consumption in
state 3 and were also more sensitive to a-bisabololtoxicity (Fig. 6A).
Even fibroblasts became more sensitive to a-bisabololtoxicity when pushed to a more aerobic state. In
Fig. 6B, we show that the normal respiration rate is
maintained for only 20 min. Under these culture condi-
tions, 40 min of incubation with 5 lm a-bisabololresulted in a 50% loss of viability in normal fibroblasts,
whereas T67 cells show 100% mortality (Table S1). The
increase in a-bisabolol toxicity in cells forced to an
aerobic metabolism strengthens the hypothesis of a
mitochondrial involvement in a-bisabolol toxicity.
a-Bisabolol affects mitochondrial membrane
potential in intact cells
The effect of a-bisabolol on mitochondrial membrane
potential in digitonin-permeabilized cells was evaluated
using the fluorescent cationic probe tetramethylrod-
amine methyl ester (TMRM). T67 cells and fibroblasts,
preincubated for 30 min with or without CsA, were
treated for 40 min with a-bisabolol.In T67 cells, a-bisabolol treatment results in a
decreased membrane potential and this effect was
counteracted by CsA (Fig. 7A). As shown in Fig. 7B,
a-bisabolol did not affect fibroblasts membrane poten-
tial within 40 min of treatment.
Spectrofluorimetric data are confirmed by fluores-
cence microscopy imaging, using the same fluorogenic
probe (TMRM). T67 images collected after 20 and
40 min of a-bisabolol incubation showed a progressive
loss of mitochondrial membrane potential (Fig. 7C).
Preincubation with CsA protected T67 cells from
depolarization. Treated fibroblasts images revealed
only a slight effect of a-bisabolol on mitochondrial
membrane potential.
Discussion
The different effect of a-bisabolol on tumor versus nor-
mal cells is unusual and knowledge of the molecular
basis for this difference should be very important for the
research into and development of targeted antitumor
Table 2. Effect of 5 lM a-bisabolol on oxygen consumption in coupled rat liver mitochondria energized with glutamate–malate. State 3 of
respiration was measured in the presence of two different ADP concentrations (180 and 500 lM). 2,4-Dinitrophenol (60 lM) was added to
achieve the maximal respiration rate.
Substrate
State 4 State 3(ADP 180 lM) State 3(ADP 500 lM) 2,4-Dinitrophenol
ng AoxygenÆmin)1Æmg)1 ng AoxygenÆmin)1Æmg)1 ng AoxygenÆmin)1Æmg)1 ng AoxygenÆmin)1Æmg)1
Glutamate ⁄ malate 19.4 ± 1.7 93.9 ± 10.9 118.8 ± 5.3
Glutamate ⁄ malate + a-bisabolol 5 lM 23.5 ± 2.7 59.8 ± 7.2* 72.05 ± 6.5* 105.2 ± 12
*P < 0.05.
a-Bisabolol, an efficient apoptosis inducer E. Cavalieri et al.
3994 FEBS Journal 276 (2009) 3990–4000 ª 2009 The Authors Journal compilation ª 2009 FEBS
drugs. One possible reason for this selectivity was
exploited in a recent study by Darra et al. [13] in which
they studied the uptake of a-bisabolol in lipid rafts from
tumor and normal cells. Usually tumor plasma mem-
branes contain more lipid rafts than the normal plasma
membranes and this may explain the higher toxicity
exerted by a-bisabolol [17,18]. Moreover, a-bisabololtrafficking between the plasma membrane and the
mitochondria may be mediated by Bid, as suggested by
the binding experiments described by Darra et al. [13].
Furthermore, once absorbed in the plasma membrane,
a-bisabolol is rapidly distributed in the cytosol
and nuclei. We confirm the different accumulation of
a-bisabolol inside cells using T67 and fibroblasts.
The increase in cytochrome c release [7] and the
resistance to cell death conferred by CsA strongly sug-
gest mitochondrial involvement in a-bisabolol-inducedapoptosis; moreover our results show that neither
Bax ⁄Bcl-2 nor ROS production are involved. In this
scenario, ROS production observed in the T67 cell line
is not the direct cause of apoptosis, but could be a
secondary effect because of the loss of mitochondrial
integrity. In fact, using NAC as a radical scavenger,
we observed a decrease in ROS production, but we did
not prevent cell death. In T67 cell lines, death seems to
be caused by a massive mPTP opening with a conse-
quent release of mitochondrial constituents and a loss
A
B
C
Fig. 5. CsA protects T67 cells from apoptotic death induced by a-
bisabolol. (A) Pretreatment with 10 lM CsA prevented a decrease
in the of oxygen consumption rate induced by 30 min of incubation
with 5 lM a-bisabolol in both coupled and uncoupled respiration.
Uncoupling was achieved by adding FCCP to a final concentration
of 600 nM. (B) CsA prevented PARP cleavage and DNA ladder for-
mation induced by a-bisabolol. (C) FK506 did not prevent a-bisabo-
lol-induced PARP cleavage and DNA ladder formation. Cells were
treated with 10 lM CsA (B) and 1 lM FK506 (C) for 30 min and then
with 5 lM a-bisabolol or vehicle solvent. Figures show representa-
tive results from four independent experiments.
A
B
Fig. 6. Oxygen consumption rate of the T67 cell line and human
fibroblasts in the presence of galactose. Effect of a-bisabolol treat-
ment on coupled and uncoupled respiration in the T67 cell line (A)
and human fibroblasts (B). Uncoupling was achieved by adding
FCCP to a final concentration of 600 nM. Data are the mean of
three different determinations ± SD.
E. Cavalieri et al. a-Bisabolol, an efficient apoptosis inducer
FEBS Journal 276 (2009) 3990–4000 ª 2009 The Authors Journal compilation ª 2009 FEBS 3995

of respiratory activity. The inference that mPTP is a
target in the mechanism of action of this small terpene
is supported by the prevention of cell death by CsA, a
specific inhibitor of mPTP opening [19]. CsA preincu-
bation prevents a-bisabolol-induced mitochondrial
membrane depolarization both in permeabilized and
adherent T67 cells. In normal fibroblasts, 40 min of
a-bisabolol incubation did not exert any significant
effect on mitochondrial potential.
The results obtained on isolated rat liver mitochon-
dria suggest that the damage is related to a loss of mem-
brane integrity. Nevertheless, in rat liver mitochondria,
the effect of a-bisabolol on mPTP opening was slight; at
first glance, this behavior seems to be in disagreement
with the strong cell death induced (Fig. S2).
The reasons for this apparent discrepancy may be
related to the different experimental models (cell-free
vs. cell models). In a cell model, even a partial loss of
mitochondrial membrane integrity may trigger an
amplified response (e.g. cytochrome c release and
apoptosis). Loss of mitochondrial integrity may be
more dangerous for cells that depend mainly on aero-
bic rather than glycolytic metabolism. In the latter
case, the energy requirement is less dependent on mito-
chondrial activity, and so cells can survive even with
mitochondrial impairment.
Because a-bisabolol seems to be preferentially
adsorbed in lipid rafts, we speculate that its effect
may be mediated by perturbation of the lipid bilayer.
In pilot experiments, we prepared dipalmitoylphos-
phatidylcholine vesicles in the presence of 5 lm
a-bisabolol and examined them using differential scan-
ning calorimetry. Analysis showed that a-bisabololinduced a decrease in the transition temperature of the
lipid phase without broadening the differential scan-
ning calorimetry spectra (Fig. S3). This is indicative of
a change in the more external part of the lipid vesicles
and suggests that a-bisabolol is able to induce disorga-
nization on the membrane surface. In mitochondria,
such a change may be involved in the PTP regulation,
as shown by Ricchelli et al. [20].
In conclusion, our results confirm that a-bisabolol isable to induce apoptotic cell death preferentially in
tumor cells and suggest that its toxicity is strictly
related to its intracellular concentration. Because of its
high lipophilicity, a-bisabolol uptake should be medi-
ated by membrane lipid rafts and its intracellular traf-
ficking may involve Bid, a protein of the Bcl-2 family.
This would explain the increased uptake in cancer
cells, whereas the results obtained using CsA confirm
the involvement of mPTP opening in the cell death
mechanism.
Materials and methods
Reagents
All chemicals used were of the highest analytical grade and
purchased from Sigma Chemical Company (Milan, Italy),
unless otherwise specified.
Fibroblasts αα-bisabolol α-bisabolol
T67
0 min
20 min
40 min
α-bisabolol+ CsA
α-bisabolol+ CsA
A
B
C
Fig. 7. a-Bisabolol reduces the mitochondrial DW in T67 cells, but
not in fibroblasts. Effects of 5 lM a-bisabolol (40 min of incubation)
on mitochondrial membrane potential in T67 cells (A) and fibro-
blasts (B) in the presence or absence of 10 lM CsA. Data are
expressed as fluorescence arbitrary unitsÆ106 cells)1 and are the
mean of at least three different determinations (± SD). (C) Fluores-
cence microscopy images of fibroblasts and T67 cells in the pres-
ence of 5 lM a-bisabolol and 5 lM a-bisabolol + 10 lM CsA. Images
were collected at 0, 20 and 40 min of incubation, using 100 nM
TMRM as the mitochondrial membrane potential probe.
a-Bisabolol, an efficient apoptosis inducer E. Cavalieri et al.
3996 FEBS Journal 276 (2009) 3990–4000 ª 2009 The Authors Journal compilation ª 2009 FEBS

Cell culture
The T67 human glioma cell line was derived by Lauro et al.
[21] from a World Health Organization (WHO) Grade III
gemistocytic astrocytoma. The primary culture of human
fibroblasts was a generous gift from U. Armato (Verona
University, Italy). T67 cells and normal human fibroblasts
were cultured in DMEM (BioWhittaker, Cambrex BioSci-
ence, Verviers, Belgium), supplemented with 10% fetal
bovine serum (BioWhittaker, Cambrex BioScience),
100 UIÆmL)1 penicillin, 100 lgÆmL)1 streptomycin and
40 lgÆmL)1 gentamycin, in a 5% CO2 atmosphere at 37 �C,with saturating humidity. Cell stocks were cryopreserved
using standard methods, and stored in liquid nitrogen. Cell
viability was measured by Trypan blue exclusion [22].
a-Bisabolol treatment
Cells were treated with a-bisabolol (purum, ‡ 95%, Fluka
and Riedel-de Haen, Sigma Chemical Co.) from a freshly
prepared stock solution in absolute ethanol (1 : 8 v ⁄ v). Thea-bisabolol concentration indicated in cell experiments rep-
resents its soluble fraction in culture medium, measured as
described previously [7].
Measurement of a-bisabolol in membrane,
cytosol and nuclear fraction
T67 cells and fibroblasts treated for 15 min with a-bisabololwere resuspended and homogenized using a hand-held glass
homogenizer in 300 lL of ice-cold buffer A (20 mm
Tris ⁄HCl, pH 7.5; 2 mm EDTA; 0.5 mm EGTA; complete
EDTA-free protease inhibitor cocktail; Roche Applied
Science, Mannheim, Germany). Lysates were centrifuged at
800 g for 5 min at 4 �C in order to remove nuclei and super-
natants were centrifuged at 1 000 000 g for 20 min at 4 �C.The obtained supernatants were collected and used as the
cytosolic fraction. The membrane pellets were solubilized in
200 lL buffer A plus 1% Triton X-100 by bath sonication
and centrifuged at 12 000 g for 20 min at 4 �C in a micro-
centrifuge. The obtained supernatants were used as the
membrane fraction. a-Bisabolol concentration was analyzed
in each fraction by the gas chromatography method [23].
Western blot analysis
Bax analysis
Cell pellets were suspended in 1 mL of solution containing
10 mm NaCl, 1.5 mm MgCl2, 10 mm Tris ⁄HCl, pH 7.5,
1 mm sodium orthovanadate and complete EDTA-free pro-
tease inhibitor cocktail (Boehringer-Mannheim). Cells were
then chilled on ice for 10 min and gently lysed by adding
0.3% (v ⁄ v) Nonidet-P40. In order to restore an isotonic
environment, a solution containing 525 mm mannitol,
175 mm sucrose, 12.5 mm Tris ⁄HCl, 2.5 mm EDTA,
pH 7.5 and protease inhibitor cocktail was added. Lysates
were centrifuged at 17 000 g for 30 min at 4 �C. The cyto-
solic and mitochondrial fractions so obtained were sepa-
rated on a 15% SDS polyacrylamide gels.
Bcl-2 and PARP analysis
Cells were homogenized at 4 �C in 50 mm Tris ⁄HCl, pH 8,
containing 0.1% Nonidet-P40, 200 mm KCl, 2 mm MgCl2,
50 lm ZnCl2, 2 mm dithiothreitol and EDTA-free protease
inhibitor cocktail (Boehringer-Mannheim). Aliquots of the
homogenates were loaded on 15% (for Bcl-2) or 7.5% (for
PARP) SDS polyacrylamide gels.
Electrophoresis was performed at 100 V with a running
buffer containing 0.25 m Tris ⁄HCl, pH 8.3, 1.92 m glycine
and 1% SDS. The resolved proteins were electroblotted onto
a poly(vinylidene difluoride) membrane (Immobilon P; Milli-
pore, Bedford MA, USA). Membranes were incubated with
a rabbit polyclonal IgG antibody to Bax (Upstate, Lake
Placid, NY, USA), a mouse monoclonal IgG antibody to Bcl-
2 (Upstate) and a mouse monoclonal IgG antibody to PARP
(Zymed, South San Francisco, CA, USA) and incubated with
the appropriate secondary antibody IgG–peroxidase conju-
gate (Amersham Biosciences, Little Chalfont, UK). The blots
were successively incubated with enhanced chemiluminescent
detection reagents (ECL; Amersham Biosciences) according
to the manufacturer’s instructions and proteins were detected
by exposing the blots to Kodak X-AR film.
DNA ladder
For internucleosomal DNA laddering, 3 · 106 cells, resus-
pended in 0.3 mL of culture medium containing 10% fetal
bovine serum, were incubated for 45 min at 65 �C, and then
overnight at 37 �C in the presence of 0.4 m NaCl, 5 mm
Tris ⁄HCl, pH 8, 2 mm EDTA, 4% SDS and 2 mgÆmL)1 pro-
teinase K. The lysates, brought to a final concentration of
1.58 m NaCl, were centrifuged twice for 10 min at 6000 g to
separate the DNA fragments from intact DNA. The superna-
tants were recovered and DNA was precipitated by addition
of three volumes of absolute ethanol at )80 �C for 1 h. DNA
pellets were recovered by microcentrifugation (10 min at
12 000 g), and resuspended in a minimal volume of 40 lL of
10 mm Tris ⁄HCl, pH 7.4, 1 mm EDTA, 1 mgÆmL)1 DNase-
free ribonuclease A. Aliquots of 5 lg of DNA were then
loaded onto a 1% agarose gel containing 0.25 lgÆmL)1
ethidium bromide. After electrophoresis, the DNA was
visualized by UV light and recorded photographically.
Measurement of ROS
Confluent T67 cells and fibroblasts washed with fresh
DMEM were incubated for 2 h in DMEM premixed with
E. Cavalieri et al. a-Bisabolol, an efficient apoptosis inducer
FEBS Journal 276 (2009) 3990–4000 ª 2009 The Authors Journal compilation ª 2009 FEBS 3997

10 lm dichlorodihydrofluorescein diacetate (DCF-DA).
Cells loaded with DCF-DA were subsequently washed with
NaCl ⁄Pi and incubated for 40 min in DMEM mixed with
5 lm a-bisabolol. Fluorescence was recorded in a fluores-
cence plate reader using a six-well microtiter plate after
excitation at 485 nm and emission at 535 nm at 30 �C.Cells were treated with 10 lm CsA and 10 mm NAC
during DCF-DA incubation to test their effect on ROS
production. To normalize the fluorescence detection each
well was carefully seeded with the same number of cells
[24–26].
Mitochondrial membrane potential in intact cells
Confluent T67 cells and fibroblasts were incubated for
40 min with 5 lm a-bisabolol, with or without 10 lm CsA
preincubation. Briefly, after trypsinization, cells were
collected and resuspended in 0.25 m sucrose, 10 mm
Hepes, 5 mm KH2PO4, 1 mm EDTA, 50 lm EGTA,
pH 7.4 with KOH. Assays were carried out in 2 mL resus-
pension medium, containing 1 · 106 cells, in the presence
of 100 nm TMRM, 1 lgÆmL)1 oligomycin and 1 lgÆmL)1
rotenone. After 10 min for stabilization, succinate ⁄Triswas added to 12 mm to energize mitochondria. Then
5 lgÆmL)1 digitonin pulses were added stepwise until
complete cell permeabilization [27]. Depolarization was
achieved by 200 nm FCCP addition. TMRM fluorescence
was measured in a Jasco FP-777 spectrofluorometer, using
558 nm as excitation wavelength and 578 nm as emission
wavelength.
Cell images were obtained using an IX50 inverted fluo-
rescence microscope (Olympus, Tokyo, Japan). Confluent
cells were washed with DMEM, incubated with 100 nm
TMRM for 20 min, then washed again with DMEM and
treated with 5 lm a-bisabolol in the presence or absence of
10 lm CsA. Images were collected at 20 and 40 min after
a-bisabolol treatment.
Oxygen consumption
Cells
Intact cells (1 · 106) were assayed for oxygen consumption
at 37 �C using a thermostatically controlled oxygraph and
Clark electrode. The respiration rate was measured in cells
treated with 5 lm a-bisabolol in DMEM. Maximal respira-
tion rate was empirically determined by stepwise addition
of the uncoupler FCCP.
For respiration assays in the presence of galactose, cells
were incubated for 24 h in DMEM without glucose, con-
taining 10% (v ⁄ v) dialyzed fetal bovine serum. This glu-
cose-free medium was supplemented with galactose (5 mm)
and pyruvate (110 mgÆL)1) as the oxidizable substrate. Res-
piration rates were measured in this medium, and expressed
as nmol O2Æmin)1Æ106 cells)1.
Rat liver mitochondria
Coupled rat liver mitochondria were prepared according to
Costantini et al. [28]. Oxygen consumption was assayed
according to Slater [29]. The maximal electron transport
capability (maximal respiration rate) was achieved by add-
ing 60 lm 2,4-dinitrophenol as the electron transfer uncou-
pler. The respiratory rates were expressed in ng atoms
oxygenÆmin)1Æmg)1 of protein referencing to 250 nmo-
l O2Æml buffer)1 as 100% at 30 �C [30].
Acknowledgements
We thank Dr Sergio Bonora for performing the differ-
ential scanning calorimetry experiments. This work
was supported financially by grant COFIN2002 of Ital-
ian Ministry of University and Scientific and research
and grant of CARIVERONA 2003 to HS.
References
1 Grassi A, Palermi G & Paradisi M (2000) Study of toler-
ance and efficacy of cosmetic preparations with lenitive
action in atopic dermatitis in children.Clin Ter 151, 77–80.
2 Jakovlev V, Isaac O, Thiemer K & Kunde R (1979)
Pharmacological investigations with compounds of
chamomile II. New investigations on the antiphlogistic
effects of (-)-alpha-bisabolol and bisabolol oxides
[author’s translation]. Planta Med 35, 125–140.
3 Szelenyi I, Isaac O & Thiemer K (1979) Pharmacological
experiments with compounds of chamomile III. Experi-
mental studies of the ulcer protective effect of chamomile
[author’s translation]. Planta Med 35, 218–227.
4 Torrado S, Torrado S, Agis A, Jimenez ME &
Cadorniga R (1995) Effect of dissolution profile and
(–)-alpha-bisabolol on the gastrotoxicity of acetylsalicylic
acid. Pharmazie 50, 141–143.
5 Villegas LF, Marcalo A, Martin J, Fernandez ID,
Maldonado H, Vaisberg AJ & Hammond GB (2001)
(+)-epi-Alpha-bisabolol [correction of bisbolol] is
the wound-healing principle of Peperomia galioides:
investigation of the in vivo wound-healing activity of
related terpenoids. J Nat Prod 64, 1357–1359.
6 de Andrade IL, Bezerra JN, Lima MA, de Faria RA,
Lima MA, Andrade-Neto M, Cavalcanti FS & Mesqui-
ta AL (2004) Chemical composition and insecticidal
activity of essential oils from Vanillosmopsis pohlii
Baker against Bemisia argentifolii. J Agric Food Chem
52, 5879–5881.
7 Cavalieri E, Mariotto S, Fabrizi C, de Prati AC,
Gottardo R, Leone S, Berra LV, Lauro GM, Ciampa
AR & Suzuki H (2004) alpha-Bisabolol, a nontoxic
natural compound, strongly induces apoptosis in
a-Bisabolol, an efficient apoptosis inducer E. Cavalieri et al.
3998 FEBS Journal 276 (2009) 3990–4000 ª 2009 The Authors Journal compilation ª 2009 FEBS

glioma cells. Biochem Biophys Res Commun 315, 589–
594.
8 Budavari S (1996) The Merck Index, 12th edn, 208.
Merck Research Laboratories Division of Meck and
CO., Inc, Whitehouse Station, NJ.
9 Geske FJ & Gerschenson LE (2001) The biology of
apoptosis. Hum Pathol 32, 1029–1038.
10 Ghobrial IM, Witzig TE & Adjei AA (2005) Targeting
apoptosis pathways in cancer therapy. CA Cancer J
Clin 55, 178–194.
11 Lowe SW & Lin AW (2000) Apoptosis in cancer. Carci-
nogenesis 21, 485–495.
12 Kaplow R (2005) Innovations in antineoplastic therapy.
Nurs Clin N Am 40, 77–94.
13 Darra E, Abdel-Azeim S, Manara A, Shoji K, Marechal
JD, Mariotto S, Cavalieri E, Perbellini L, Pizza C, Pera-
hia D et al. (2008) Insight into the apoptosis-inducing
action of alpha-bisabolol towards malignant tumor
cells: involvement of lipid rafts and Bid. Arch Biochem
Biophys 476, 113–123.
14 Patra SK & Bettuzzi S (2007) Epigenetic DNA-methyl-
ation regulation of genes coding for lipid raft-associ-
ated components: a role for raft proteins in cell
transformation and cancer progression. Oncol Rep 17,
1279–1290.
15 Gross A, McDonnell JM & Korsmeyer SJ (1999) BCL-
2 family members and the mitochondria in apoptosis.
Genes Dev 13, 1899–1911.
16 D’Aurelio M, Pallotti F, Barrientos A, Gajewski CD,
Kwong JQ, Bruno C, Beal MF & Manfredi G (2001)
In vivo regulation of oxidative phosphorylation in cells
harboring a stop-codon mutation in mitochondrial
DNA-encoded cytochrome c oxidase subunit I. J Biol
Chem 276, 46925–46932.
17 Canuto RA, Biocca ME, Muzio G & Dianzani MU
(1989) Fatty acid composition of phospholipids in mito-
chondria and microsomes during diethylnitrosamine
carcinogenesis in rat liver. Cell Biochem Funct 7, 11–19.
18 Meng X, Riordan NH, Riordan HD, Mikirova N,
Jackson J, Gonzalez MJ, Miranda-Massari JR, Mora E
& Trinidad Castillo W (2004) Cell membrane fatty acid
composition differs between normal and malignant cell
lines. Proc R Health Sci J 23, 103–106.
19 Petronilli V, Cola C, Massari S, Colonna R & Bernardi
P (1993) Physiological effectors modify voltage sensing
by the cyclosporin A-sensitive permeability transition
pore of mitochondria. J Biol Chem 268, 21939–21945.
20 Ricchelli F, Dabbeni-Sala F, Petronilli V, Bernardi P,
Hopkins B & Bova S (2005) Species-specific modulation
of the mitochondrial permeability transition by norbor-
mide. Biochim Biophys Acta 1708, 178–186.
21 Lauro GM, Di Lorenzo N, Grossi M, Maleci A &
Guidetti B (1986) Prostaglandin E2 as an immunomod-
ulating factor released in vitro by human glioma cells.
Acta Neuropathol 69, 278–282.
22 Gorman A, McCarthy J, Finucane D & Reville W
(1996) Morphological assessment of apoptosis. In
Techniques in Apoptosis, A Users Guide (Cotter TG &
Martin SJ, eds), pp 6–7. Portland Press, London.
23 Perbellini L, Gottardo R, Caprini A, Bortolotti F,
Mariotto S & Tagliaro F (2004) Determination of
alpha-bisabolol in human blood by micro-HPLC-
ion trap MS and head space-GC-MS methods.
J Chromatogr B Anal Technol Biomed Life Sci 812,
1373–377.
24 LeBel CP, Ischiropoulos H & Bondy SC (1992) Evalua-
tion of the probe 2¢,7¢-dichlorofluorescin as an indicator
of reactive oxygen species formation and oxidative
stress. Chem Res Toxicol 5, 227–231.
25 Myhre O, Andersen JM, Aarnes H & Fonnum F (2003)
Evaluation of the probes 2¢,7¢-dichlorofluorescin diace-
tate, luminol, and lucigenin as indicators of reactive
species formation. Biochem Pharmacol 65, 1575–1582.
26 Myhre O, Vestad TA, Sagstuen E, Aarnes H &
Fonnum F (2000) The effects of aliphatic (n-nonane),
naphtenic (1,2,4-trimethylcyclohexane), and aromatic
(1,2,4-trimethylbenzene) hydrocarbons on respiratory
burst in human neutrophil granulocytes. Toxicol Appl
Pharmacol 167, 222–230.
27 Floryk D & Houstek J (1999) Tetramethyl rhodamine
methyl ester (TMRM) is suitable for cytofluorometric
measurements of mitochondrial membrane potential in
cells treated with digitonin. Biosci Rep 19, 27–34.
28 Costantini P, Petronilli V, Colonna R & Bernardi P
(1995) On the effects of paraquat on isolated
mitochondria. Evidence that paraquat causes opening
of the cyclosporin A-sensitive permeability transition
pore synergistically with nitric oxide. Toxicology 99,
77–88.
29 Slater EC (1966) Our present knowledge of the mecha-
nism of energy conservation in biological oxidations.
Bull Soc Chim Biol 48, 1151–1168.
30 Estabrook RW (1967) Mitochondrial respiratory
control and the polarographic measurement of ADP:O
ratios. Methods Enzymol 10, 41–47.
Supporting information
The following supplementary material is available:
Fig. S1. Effect of different incubation times on the via-
bility of T67 cells and human fibroblasts treated with
a-bisabolol.Fig. S2. Effect of a-bisabolol on mitochondrial swelling.
Fig. S3. Effect of a-bisabolol on transition temperature
of dipalmitoylphosphatidylcholine vesicles.
Table S1. Effect of different incubation times on the
viability of T67 cells and human fibroblasts treated
with a-bisabolol, using galactose ⁄pyruvate as a respira-
tory substrate.
E. Cavalieri et al. a-Bisabolol, an efficient apoptosis inducer
FEBS Journal 276 (2009) 3990–4000 ª 2009 The Authors Journal compilation ª 2009 FEBS 3999

This supplementary material can be found in the
online article.
Please note: As a service to our authors and read-
ers, this journal provides supporting information sup-
plied by the authors. Such materials are peer-
reviewed and may be re-organized for online deliv-
ery, but are not copy-edited or typeset. Technical
support issues arising from supporting information
(other than missing files) should be addressed to the
authors.
a-Bisabolol, an efficient apoptosis inducer E. Cavalieri et al.
4000 FEBS Journal 276 (2009) 3990–4000 ª 2009 The Authors Journal compilation ª 2009 FEBS




















