
Individual variation in the advertisement calls of a Neotropical treefrog (Scinax constrictus)
11
Amphibia-Reptilia (2014) DOI:10.1163/15685381-00002949 Individual variation in the advertisement calls of a Neotropical treefrog (Scinax constrictus) Priscilla Guedes Gambale 1,∗ , Luciana Signorelli 2 , Rogério Pereira Bastos 2 Abstract. Studies of the variability of signals at different levels are important to resolve issues related to the evolution of a species’ recognition system. We analysed the variation within males, among individuals, and among three breeding seasons in the advertisement calls of Scinax constrictus, a Neotropical hylid frog endemic to the Cerrado of Brazil. We assessed the influence of different temperature ranges and different body condition ranges over a three-year period of breeding season on the acoustical features of the advertisement calls of 62 individuals. Air temperature had negative relationship with call duration and note number. Body condition had a negative relationship with the dominant frequency and positive effects on pulse number and note duration. The acoustic parameters of S. constrictus were stable across breeding seasons. Temporal parameters were highly variable within individuals, whereas the dominant frequency was the most stereotyped property of advertisement calls. Individuals of S. constrictus have sufficient among-male variability, especially for temporal parameters (call duration, number of notes, and note duration), to permit discrimination between conspecific calling males at a reproductive site by statistical analysis. The results highlight the informativeness of non-invasive bioacoustic features for population-level studies and biological conservation. Keywords: acoustic behaviour, acoustic discrimination, Central Brazil, intraspecific variation, temporal variability. Introduction The vocalisation behaviour of animals exhibits a continuum of variation, which ensures the recognition of conspecific calls and discrimi- nation among individuals in fishes (McKibben and Bass, 1998), birds (Gentner et al., 2000; Seddon, 2005), and mammals (McComb et al., 2000; Sousa-Lima et al., 2002; Sèbe et al., 2010). This acoustic variation has also been well documented in anurans (Gerhardt, 1991; Bee and Gerhardt, 2002; Briggs, 2010; Ro- dríguez et al., 2010), but several features are still unclear. Three levels of variation in acoustic signals are possible: within a single individual, among individuals of the same species, and among in- 1 - Departamento de Biologia, Universidade Estadual de Maringá, Programa de Pós-Graduação em Ecologia de Ambientes Aquáticos Continentais, Nupélia – Núcleo de Pesquisas em Limnologia, Ictiologia e Aqüicultura, Av. Colombo 5790, 87020-900 Maringá, Paraná, Brasil 2 - Departamento de Ecologia/Instituto de Ciências Biológ- icas, Universidade Federal de Goiás, Caixa Postal 131, 74001-970 Goiânia, Goiás, Brazil ∗ Corresponding author; e-mail: [email protected] dividuals of different species. For individual variation in acoustic signals, it is important to investigate changes in the same male with re- gard to choruses from an evolutionary perspec- tive, whereas variation among individuals may provide cues for the females’ choice of conspe- cific males (Searcy and Nowicki, 2005). How- ever, for individual signal distinctiveness by conspecific males, it is necessary that acoustic variability among individuals of a species ex- ceed that within individuals of the same species (Beecher, 1989; Gerhardt, 1991). Spectral pa- rameters (e.g., the dominant frequency) of the call are under the control of morphological traits and are more stereotyped (classified as ‘static’ by Gerhardt, 1991, 2002); in contrast, tempo- ral parameters (e.g., call duration, note number) are influenced by environmental and/or social characteristics (Tárano, 2001; Castellano et al., 2002) and are more ‘dynamic’ (Gerhardt, 1991, 2002). Nevertheless, variation in call proper- ties is a continuum between static and dynamic properties (Gerhardt, 1991) due to different se- lection pressures on the vocalisations (Witte et al., 2001; Smith and Hunter, 2005; Reinhold, 2009; Rodríguez et al., 2010). © Koninklijke Brill NV, Leiden, 2014. DOI:10.1163/15685381-00002949
Transcript of Individual variation in the advertisement calls of a Neotropical treefrog (Scinax constrictus)

Amphibia-Reptilia (2014) DOI:10.1163/15685381-00002949
Individual variation in the advertisement calls ofa Neotropical treefrog (Scinax constrictus)
Priscilla Guedes Gambale1,∗, Luciana Signorelli2, Rogério Pereira Bastos2
Abstract. Studies of the variability of signals at different levels are important to resolve issues related to the evolution of aspecies’ recognition system. We analysed the variation within males, among individuals, and among three breeding seasonsin the advertisement calls of Scinax constrictus, a Neotropical hylid frog endemic to the Cerrado of Brazil. We assessedthe influence of different temperature ranges and different body condition ranges over a three-year period of breedingseason on the acoustical features of the advertisement calls of 62 individuals. Air temperature had negative relationshipwith call duration and note number. Body condition had a negative relationship with the dominant frequency and positiveeffects on pulse number and note duration. The acoustic parameters of S. constrictus were stable across breeding seasons.Temporal parameters were highly variable within individuals, whereas the dominant frequency was the most stereotypedproperty of advertisement calls. Individuals of S. constrictus have sufficient among-male variability, especially for temporalparameters (call duration, number of notes, and note duration), to permit discrimination between conspecific calling males ata reproductive site by statistical analysis. The results highlight the informativeness of non-invasive bioacoustic features forpopulation-level studies and biological conservation.
Keywords: acoustic behaviour, acoustic discrimination, Central Brazil, intraspecific variation, temporal variability.
Introduction
The vocalisation behaviour of animals exhibitsa continuum of variation, which ensures therecognition of conspecific calls and discrimi-nation among individuals in fishes (McKibbenand Bass, 1998), birds (Gentner et al., 2000;Seddon, 2005), and mammals (McComb et al.,2000; Sousa-Lima et al., 2002; Sèbe et al.,2010). This acoustic variation has also beenwell documented in anurans (Gerhardt, 1991;Bee and Gerhardt, 2002; Briggs, 2010; Ro-dríguez et al., 2010), but several features are stillunclear.
Three levels of variation in acoustic signalsare possible: within a single individual, amongindividuals of the same species, and among in-
1 - Departamento de Biologia, Universidade Estadual deMaringá, Programa de Pós-Graduação em Ecologia deAmbientes Aquáticos Continentais, Nupélia – Núcleode Pesquisas em Limnologia, Ictiologia e Aqüicultura,Av. Colombo 5790, 87020-900 Maringá, Paraná, Brasil
2 - Departamento de Ecologia/Instituto de Ciências Biológ-icas, Universidade Federal de Goiás, Caixa Postal 131,74001-970 Goiânia, Goiás, Brazil∗Corresponding author;e-mail: [email protected]
dividuals of different species. For individualvariation in acoustic signals, it is important toinvestigate changes in the same male with re-gard to choruses from an evolutionary perspec-tive, whereas variation among individuals mayprovide cues for the females’ choice of conspe-cific males (Searcy and Nowicki, 2005). How-ever, for individual signal distinctiveness byconspecific males, it is necessary that acousticvariability among individuals of a species ex-ceed that within individuals of the same species(Beecher, 1989; Gerhardt, 1991). Spectral pa-rameters (e.g., the dominant frequency) of thecall are under the control of morphological traitsand are more stereotyped (classified as ‘static’by Gerhardt, 1991, 2002); in contrast, tempo-ral parameters (e.g., call duration, note number)are influenced by environmental and/or socialcharacteristics (Tárano, 2001; Castellano et al.,2002) and are more ‘dynamic’ (Gerhardt, 1991,2002). Nevertheless, variation in call proper-ties is a continuum between static and dynamicproperties (Gerhardt, 1991) due to different se-lection pressures on the vocalisations (Witte etal., 2001; Smith and Hunter, 2005; Reinhold,2009; Rodríguez et al., 2010).
© Koninklijke Brill NV, Leiden, 2014. DOI:10.1163/15685381-00002949

2 P.G. Gambale, L. Signorelli, R.P. Bastos
Figure 1. Scinax constrictus male vocalising on vegetation at Sítio dos Pinheiros, municipality of Palmeiras, Goiás state,Brazil.
Studies on acoustic variability between sub-sequent breeding seasons are still scarce foranuran amphibians (e.g., Sullivan and Rinshaw,1992; Brooke et al., 2000; Smith and Hunter,2005). Individual hormonal changes betweenbreeding seasons (Ball, 1999), environmentalchanges (Brooke et al., 2000), changes in demo-graphic structure (Nottebohm and Nottebohm,1978; Howard and Young, 1998; Pröhl, 2003),and the social context in which males call(Wells, 1977; Bastos et al., 2011) may changeanuran advertisement calls over time. Neverthe-less, the hereditary basis of acoustic parametersmay limit individual variation in signals amongbreeding seasons (Smith and Hunter, 2005). Theassessment of call variability at several levels(among males, within males, and among years)is important for providing information abouthow individuals can be identified as belongingto one conspecific class (Gerhardt, 1991; Bee etal., 2001; Bee, 2004; Rodríguez et al., 2010).This species recognition system (Beer, 1970) isstill poorly understood.
Scinax constrictus Lima, Bastos and Giaretta,2004 is a species of the family Hylidae that is re-
stricted to the Cerrado of South America, Brazil(Valdujo et al., 2012) (fig. 1). The reproductiveactivity of this species is restricted to the rainyseason, occurring in ponds in open areas be-tween December and March (Lima et al., 2004).Lima et al. (2004) described the advertisementcall of S. constrictus as having a pulsed struc-ture, but published information on the variabil-ity of acoustic traits in this species is limited. Weexamined variation in the advertisement call ofS. constrictus to evaluate whether temperatureand body condition influenced acoustic parame-ters and to test the following hypotheses: (i) ad-vertisement calls of the population do not varyamong breeding seasons; (ii) male acoustic sig-nals vary among individuals at a breeding site;and (iii) acoustic variability should be greateramong than within individuals.
Materials and methods
We carried out field observations during three breeding sea-sons, from October to March (rainy period) in 2005-6, 2006-7, and 2007-8, in a permanent body of water at the Sítiodos Pinheiros, municipality of Palmeiras de Goiás, Goiás

Call variation of Scinax constrictus 3
State (16°47′22.3′′S 49°55′57.5′′W; 1300 m a.s.l.). The wa-ter body is a pond surrounded by corn crops, pastures, andinterfluvial and riparian vegetation (sensu Eiten, 1994). Inthe 2007 breeding season, an important reduction in ripar-ian vegetation was observed (Gambale, pers. obs.) and thischange in the habitat most likely explains the decrease in thenumber of active males (only nine males were recorded).
We conducted field work for 30 days in each breed-ing season from 18:00 h until 00:00 h, using white or redbeam flashlights, to reduce the stress on animals (Buchanan,1993). We made recordings with a digital recorder, MarantzPMD660 (uncompressed format, sampling rate = 44.1 kHz,16 bits) or an analog cassette recorder, Marantz PMD222with a Sennheiser ME66 directional microphone, at a dis-tance of approximately 0.5 m from the calling male. Toavoid recording the same male more than once, all maleswere collected immediately after the recording, subse-quently measured and marked by toe clipping (Martof,1953), and returned to the pond. After each recording, theair temperature was measured near the shore with a digi-tal thermohygrometer, Instrutherm HT-210 (precision 1°C).We measured individuals (snout-vent length [SVL]) with adigital caliper (precision 0.05 mm) and weighed them witha digital scale (precision 0.01 g).
Analogue recordings were digitised on a computer at22-kHz sampling frequency and 16-bit resolution. Digitalrecordings were resampled with Sound Forge 10.0 softwareat a frequency of 22 kHz to produce a similar spectrogramresolution for both the digital and analogue recordings. Weanalysed the temporal parameters of the call with Avisoft-SAS Lab Lite software and the spectral parameters withCool Edit 2000 and obtained frequency information throughthe Fast Fourier Transformation (FFT, 1024 points).
We analysed five advertisement calls for each individual,randomly sampled from approximately 2 minutes of record-ing of the same individual, according to Bastos et al. (2011)and Morais et al. (2012). The following acoustic parameterswere evaluated: call duration (ms), note number (notes/call),note duration (ms), pulse number (pulses/note), and domi-nant frequency (Hz). The terms relative to the bioacousticanalysis are in agreement with Lima et al. (2004). Somevoucher specimens are housed in the Zoological Collectionof the Universidade Federal de Goiás (ZUFG 3968, 3970).
Considering that anuran amphibians are ectothermic an-imals (Duellman and Trueb, 1994; Gerhardt and Huber,2002), multiple regression was employed to evaluate the in-fluences of temperature and body condition for all 62 malesrecorded. For this, we calculated the averages of the five pa-rameters of a given male and considered the averages as onedatum. To reduce the multicollinearity of factors, body con-dition was measured as the product of SVL and body mass.To evaluate the influence of breeding season on acousticcharacteristics, the seasons were also considered an inde-pendent variable in the multiple regression.
To evaluate call variability patterns within and among in-dividuals, we calculated the coefficient of variation (CV) =(SD/X) × 100 for each acoustic parameter. We estimatedwithin- and among-male coefficients of variation by breed-ing season and within- and among-male coefficients for all
62 males recorded. The within-male coefficients of varia-tion were based on the means and standard deviations ofthe recordings for each male. The calculation of the among-male coefficient of variation was performed with the meansand standard deviations of the males of each breeding sea-son. Acoustic parameters were classified into ‘static’ and‘dynamic’ according to the criteria suggested by Gerhardt(1991): parameters with a within-male coefficient of varia-tion lower than or equal to 5% are classified as ‘static’; thosehigher than or equal to 12% are ‘dynamic’. This classifica-tion has been widely adopted in studies of mate choice inanurans (Bee et al., 2013; Pettitt et al., 2013).
Following Márquez and Eekhout (2006), we calculatedthe ratio between the among-male coefficient (CVA) andwithin-male (CVW) variation to determine the variabil-ity among males. According to previous studies, whenCVA/CVW > 1, the parameter can function in individualrecognition (Jouventin et al., 1999; Bee et al., 2001). Weapplied the Kruskal-Wallis test (Zar, 1999) for each breed-ing season to test the significance of differences in among-male acoustic variation. The acoustic parameters were inde-pendent variables, and the individuals were dependent vari-ables. For each individual, we used five measured acousticvariables.
To determine whether the acoustic parameters of theadvertisement call of S. constrictus could be used to dis-tinguish individuals at the breeding site and to evaluateacoustic distinctiveness among individuals, we performeda Discriminant Function Analysis (DFA; Nelson and Mar-ler, 1990), considering the 2005-6 and 2006-7 breeding sea-sons separately. We did not perform a DFA for the 2007-8breeding season because the model constructed was simpler(n = 9) than the other years and difficult to compare withthe others. For DFA, we considered the five calls from eachindividual as independent samples.
Prior to the DFA, we adjusted each measured acousticvalue to a common air temperature for both breeding sea-sons (mean = 23.7°C) following Kaefer and Lima (2012).Because many of the call properties were correlated, we per-formed a principal components analysis (PCA) to create aset of five orthogonal predictor variables. The correspond-ing principal component scores were used as input variablesfor the DFA, which is a multivariate statistical method usedto distinguish between the acoustic parameters of differ-ent individuals (Bee et al., 2001; Bee, 2004; Gasser et al.,2009). We measured classification success using a ‘leave-one-out’ cross-validation procedure. The models were fittedwith n-1 remaining calls, and the ability to classify this da-tum to one of the groups was scored. Canonical functionswith eigenvalues higher than one were analysed, and vari-ables that best explained the discrimination among individ-uals were selected on the basis of their standardised betacoefficients. All the statistical analyses were in accordancewith Zar (1999) and Manly (2008) and performed with theaid of the software Statistica 7.1 (Statsoft, 2006) and ‘R forWindows’ (R project for statistical computing, 2012), withthe significance level set at P = 0.05.
4 P.G. Gambale, L. Signorelli, R.P. Bastos
Results
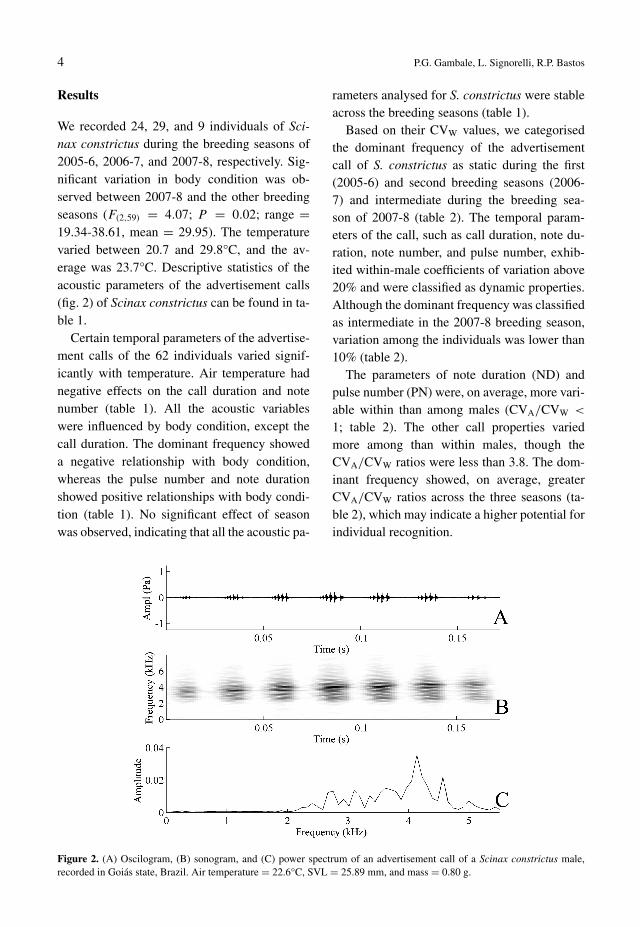
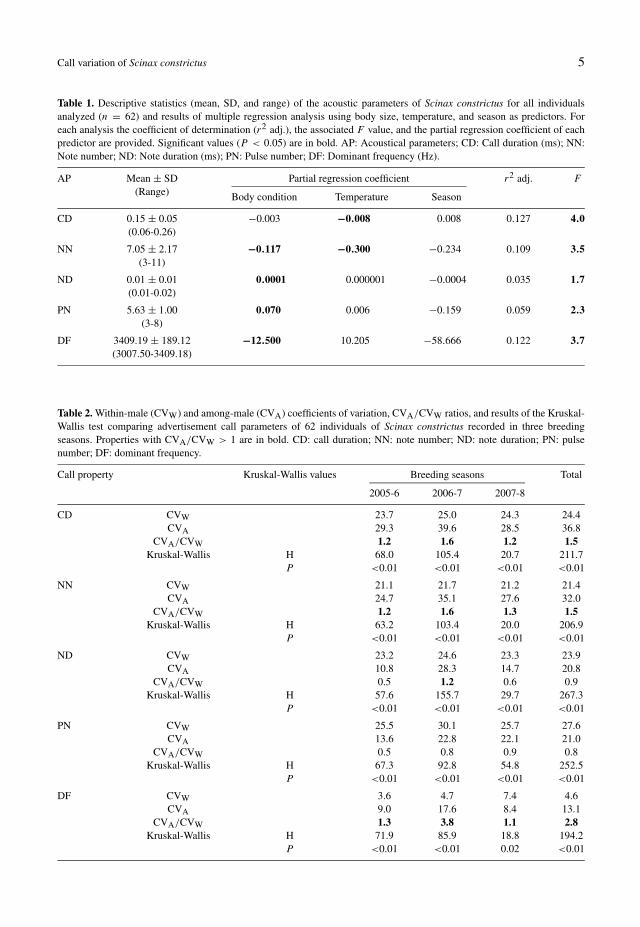
We recorded 24, 29, and 9 individuals of Sci-nax constrictus during the breeding seasons of2005-6, 2006-7, and 2007-8, respectively. Sig-nificant variation in body condition was ob-served between 2007-8 and the other breedingseasons (F(2,59) = 4.07; P = 0.02; range =19.34-38.61, mean = 29.95). The temperaturevaried between 20.7 and 29.8°C, and the av-erage was 23.7°C. Descriptive statistics of theacoustic parameters of the advertisement calls(fig. 2) of Scinax constrictus can be found in ta-ble 1.
Certain temporal parameters of the advertise-ment calls of the 62 individuals varied signif-icantly with temperature. Air temperature hadnegative effects on the call duration and notenumber (table 1). All the acoustic variableswere influenced by body condition, except thecall duration. The dominant frequency showeda negative relationship with body condition,whereas the pulse number and note durationshowed positive relationships with body condi-tion (table 1). No significant effect of seasonwas observed, indicating that all the acoustic pa-
rameters analysed for S. constrictus were stableacross the breeding seasons (table 1).
Based on their CVW values, we categorisedthe dominant frequency of the advertisementcall of S. constrictus as static during the first(2005-6) and second breeding seasons (2006-7) and intermediate during the breeding sea-son of 2007-8 (table 2). The temporal param-eters of the call, such as call duration, note du-ration, note number, and pulse number, exhib-ited within-male coefficients of variation above20% and were classified as dynamic properties.Although the dominant frequency was classifiedas intermediate in the 2007-8 breeding season,variation among the individuals was lower than10% (table 2).
The parameters of note duration (ND) andpulse number (PN) were, on average, more vari-able within than among males (CVA/CVW <
1; table 2). The other call properties variedmore among than within males, though theCVA/CVW ratios were less than 3.8. The dom-inant frequency showed, on average, greaterCVA/CVW ratios across the three seasons (ta-ble 2), which may indicate a higher potential forindividual recognition.
Figure 2. (A) Oscilogram, (B) sonogram, and (C) power spectrum of an advertisement call of a Scinax constrictus male,recorded in Goiás state, Brazil. Air temperature = 22.6°C, SVL = 25.89 mm, and mass = 0.80 g.
Call variation of Scinax constrictus 5
Table 1. Descriptive statistics (mean, SD, and range) of the acoustic parameters of Scinax constrictus for all individualsanalyzed (n = 62) and results of multiple regression analysis using body size, temperature, and season as predictors. Foreach analysis the coefficient of determination (r2 adj.), the associated F value, and the partial regression coefficient of eachpredictor are provided. Significant values (P < 0.05) are in bold. AP: Acoustical parameters; CD: Call duration (ms); NN:Note number; ND: Note duration (ms); PN: Pulse number; DF: Dominant frequency (Hz).
AP Mean ± SD(Range)
Partial regression coefficient r2 adj. F
Body condition Temperature Season
CD 0.15 ± 0.05 −0.003 −0.008 0.008 0.127 4.0(0.06-0.26)
NN 7.05 ± 2.17 −0.117 −0.300 −0.234 0.109 3.5(3-11)
ND 0.01 ± 0.01 0.0001 0.000001 −0.0004 0.035 1.7(0.01-0.02)
PN 5.63 ± 1.00 0.070 0.006 −0.159 0.059 2.3(3-8)
DF 3409.19 ± 189.12 −12.500 10.205 −58.666 0.122 3.7(3007.50-3409.18)
Table 2. Within-male (CVW) and among-male (CVA) coefficients of variation, CVA/CVW ratios, and results of the Kruskal-Wallis test comparing advertisement call parameters of 62 individuals of Scinax constrictus recorded in three breedingseasons. Properties with CVA/CVW > 1 are in bold. CD: call duration; NN: note number; ND: note duration; PN: pulsenumber; DF: dominant frequency.
Call property Kruskal-Wallis values Breeding seasons Total
2005-6 2006-7 2007-8
CD CVW 23.7 25.0 24.3 24.4CVA 29.3 39.6 28.5 36.8
CVA/CVW 1.2 1.6 1.2 1.5Kruskal-Wallis H 68.0 105.4 20.7 211.7
P <0.01 <0.01 <0.01 <0.01
NN CVW 21.1 21.7 21.2 21.4CVA 24.7 35.1 27.6 32.0
CVA/CVW 1.2 1.6 1.3 1.5Kruskal-Wallis H 63.2 103.4 20.0 206.9
P <0.01 <0.01 <0.01 <0.01
ND CVW 23.2 24.6 23.3 23.9CVA 10.8 28.3 14.7 20.8
CVA/CVW 0.5 1.2 0.6 0.9Kruskal-Wallis H 57.6 155.7 29.7 267.3
P <0.01 <0.01 <0.01 <0.01
PN CVW 25.5 30.1 25.7 27.6CVA 13.6 22.8 22.1 21.0
CVA/CVW 0.5 0.8 0.9 0.8Kruskal-Wallis H 67.3 92.8 54.8 252.5
P <0.01 <0.01 <0.01 <0.01
DF CVW 3.6 4.7 7.4 4.6CVA 9.0 17.6 8.4 13.1
CVA/CVW 1.3 3.8 1.1 2.8Kruskal-Wallis H 71.9 85.9 18.8 194.2
P <0.01 <0.01 0.02 <0.01

6 P.G. Gambale, L. Signorelli, R.P. Bastos
Table 3. Results from a principal components analysis of temperature-adjusted advertisement call parameters of Scinaxconstrictus. The factor loadings for all five acoustic parameters are provided and those higher than 0.5 are in bold. CD: callduration; NN: note number; ND: note duration; PN: pulse number; DF: dominant frequency; EV: eigenvalue; V: variance(%); CV: cumulative variance (%).
PCA factor Acoustic parameters EV V CV
CD NN ND PN DF
Breeding season 2005-6Factor 1 0.96 0.96 −0.10 −0.31 0.78 2.57 51.48 51.48Factor 2 0.10 0.08 0.94 0.90 0.29 1.79 35.70 87.18Factor 3 −0.23 −0.23 −0.13 0.01 0.55 0.44 8.76 95.94Factor 4 0.01 0.08 −0.30 0.31 0.04 0.19 3.83 99.77Factor 5 0.07 −0.07 0.01 0.01 −0.01 0.01 0.23 100
Breeding season 2006-7Factor 1 0.93 0.95 −0.34 −0.78 0.62 2.87 57.33 57.33Factor 2 −0.17 −0.12 −0.86 −0.31 −0.44 1.08 21.69 79.02Factor 3 −0.27 −0.22 −0.26 0.04 0.65 0.62 12.43 91.48Factor 4 0.19 0.17 −0.25 0.54 0.01 0.41 8.31 99.76Factor 5 0.08 −0.08 0.01 −0.01 0.01 0.01 0.24 100
In table 3, we report the five PCA factor load-ings extracted for each breeding season (2005-6and 2006-7) from the analyses of all the acous-tic parameters of S. constrictus adjusted for tem-perature. When the raw five factor scores fromPCA were used as input in DFAs, the discrim-inant models for 2005-6 (Wilks’ λ < 0.02;F115,446 = 4.58, P < 0.001) and 2006-7(Wilks’ λ < 0.01; F140,533 = 6.36, P <
0.001) were significant. Two canonical rootsfor 2005-6 and three canonical roots for 2006-7 from DFAs had eigenvalues greater than 1and explained 75% and 88.8%, respectively, ofthe variation in the PCA factor scores for eachbreeding season (table 4). The results of theDFAs according to the cross-validation ‘leave-one-out’ process showed that 23% and 28% ofadvertisement calls were correctly classified asbelonging to the male that produced them for2005-6 (n = 24) and 2006-7 (n = 29), respec-tively.
The first and second canonical roots from theDFA for 2005-6, which explained 41.8% and33.2% of the variance, respectively, were cor-related with PCA factor 1 and PCA factor 3, re-spectively. In this case, the acoustic parametersof call duration, note number, and dominant fre-quency were the most correlated variables thatcontributed to individual distinctiveness. The
Table 4. Standardized canonical discriminant function co-efficients showing the relative importance of each PCA fac-tor scores (from the original five call properties; see ta-ble 1) in the composition of the first three discriminant func-tions in the advertisement call of Scinax constrictus duringthe breeding seasons 2005-6 and 2006-7. Coefficients withlargest values for each function are in bold.
PCA factor Discriminant functions
1 2 3
Breeding season 2005-6Factor 1 −0.92 0.40 0.30Factor 2 −0.17 0.32 −0.09Factor 3 0.18 0.91 −0.44Factor 4 0.30 0.43 0.66Factor 5 −0.77 −0.31 −0.42Eigenvalue 2.82 2.23 0.85Cumulative proportion (%) 41.84 74.99 87.61
Breeding season 2006-7Factor 1 −0.01 1.00 −0.09Factor 2 −1.17 0.07 −0.01Factor 3 −0.39 0.18 −0.90Factor 4 −0.90 −0.08 0.41Factor 5 −0.01 0.08 0.17Eigenvalue 5.70 2.47 1.17Cumulative proportion (%) 54.24 77.71 88.85
first canonical roots from the DFA for 2006-7,which explained 54.2%, were correlated withPCA factors 2 and 4, and the second and thirdcanonical roots from the DFA for 2006-7, whichexplained 23.5% and 11.1% of the variance, re-spectively, were correlated with PCA factors 1and 3, respectively (tables 3 and 4). In this case,

Call variation of Scinax constrictus 7
all the acoustic parameters were important forindividual distinctiveness. In all breeding sea-sons, the factor loadings for dominant frequencywere low compared with the temporal parame-ters.
Discussion
Temperature and body size varied among thebreeding seasons of S. constrictus. Due to theectothermic condition of anuran amphibians,acoustic parameters can be influenced by tem-perature, considering the dependence of acous-tic parameters on muscle contraction (Wells,2007). Influences of temperature on acousticcharacters can be more subtle and difficult todetect in neotropical frogs. According to Wells(2007), increased temperature promotes an in-crease in individual metabolism and, conse-quently, an increase in emission call rate inanuran amphibians, as reported by Oseen andWassersug (2002) and Guimarães and Bas-tos (2003). Nonetheless, to maintain the con-stancy of energy expenditure (Wells, 2007),some acoustic parameters may correlate nega-tively with temperature (Lingnau and Bastos,2007; Rodríguez et al., 2010).
We found evidence that the dominant fre-quency of the advertisement call of Scinax con-strictus may be an indicator of male quality,as proposed for other anurans by Smith andRoberts (2003), Byrne (2008), and Richard-son et al. (2008). Scinax constrictus males withhigher body condition values emitted lower fre-quencies; however, previous studies found thisrelationship only for SVL (e.g., Castellano etal., 1998; Guimarães and Bastos, 2003; Silva etal., 2008; Briggs, 2010). Additional experimen-tal studies designed to control other confound-ing factors, such as calling neighbours and cho-rus tenure, will be desirable to confirm the au-thenticity of the body condition signal embed-ded in the dominant frequency of S. constrictuscalls.
There are few published studies describingthe effects of body condition on temporal pa-
rameters of calls (Morais et al., 2012). It is pos-sible that larger males store larger volumes ofair in their lungs (Bernal et al., 2005), whichwould produce positive correlations with thecall rate, call duration, number of pulses, andpulse rate of advertisement calls (Friedl andKlump, 2002; Castellano and Rosso, 2006).However, individuals may change their vocal-isation in social interactions (e.g., Bernal etal., 2007); thus, negative relationships betweentemporal characteristics of the call and bodysize can also be found, especially if we considersmall males in the choruses.
Our results support the hypothesis that malesof S. constrictus vary in their calling behaviour.In the 2005-6 and 2006-7 breeding seasons,the dominant frequency of the advertisementcall had a low within-male variability and wasclassified as a static parameter following Ger-hardt’s criteria (1991). This has also been re-ported in the following other Hylidae species:Hyla cinerea, H. crucifer, and H. versicolor(Gerhardt, 1991), H. arborea (Castellano et al.,2002), Oophaga pumilio (Pröhl, 2003), Lito-ria booroolongensis (Smith and Hunter, 2005),H. intermedia (Castellano and Rosso, 2006),Agalychnis moreletii (Briggs, 2010), and Pseu-dacris maculata (Bee et al., 2010). Althoughthis property has an important role in speciesrecognition (Pettitt et al., 2013), patterns ofpreference for this parameter are variable (Ger-hardt, 1991).
Temporal parameters (e.g., call duration,number of notes, number of pulses, and pulseduration) were characterised as dynamic. Theyare most likely dependent on social interactionsbetween individuals within a chorus (Wells,1977; Martins and Jim, 2003; Toledo and Had-dad, 2005; Bastos et al., 2011), and differencesin the energy reserves of each male are involved(Lesbarrères and Lodé, 2002). In this way, someproperties of the call can have more variabil-ity within individuals than among males (e.g.,note duration and pulse number), which maylead to rapid changes in these characteristics.Dynamic call properties are expected to be un-

8 P.G. Gambale, L. Signorelli, R.P. Bastos
der strong directional selection; however, futurepreference tests are necessary to assess S. con-strictus female preferences for these call prop-erties.
The within-male coefficient of variability forthe dominant frequency was intermediate forthe 2007-8 season, though it was static for theother seasons (table 2), showing that a contin-uum of variability is possible (Gerhardt, 1991;Reinhold, 2009); it also may possibly havebeen an artefact of the sample sizes. Tempo-ral call parameters in other Hylidae (Gerhardt,1991; Castellano and Rosso, 2006; Friedl, 2006;Gasser et al., 2009) exhibit coefficients of in-dividual variation lower than those of S. con-strictus, and these differences can be involvedin the evolutionary history of Hylidae. The pat-tern of variation of acoustic properties mightnot be conserved in the evolution of this fam-ily. Furthermore, social interactions can favourrapid changes in temporal parameters, and con-tribute to the high individual variation of theseacoustical characteristics in S. constrictus, be-cause chorus sizes of up to ten individuals canbe observed (Lima, pers. obs.).
According to the results, the acoustic pa-rameters of S. constrictus (e.g., call dura-tion, note number, note duration, and dom-inant frequency) allow discrimination amongconspecifics at a breeding site, even with lowclassification results. Our classification successvalues were similar to those reported for Allo-bates femoralis (20-93%; n = 19; Gasser etal., 2009) for a similar number of individuals.However, it is likely that S. constrictus individu-als face acoustic discrimination tasks restrictedto a small number of calling neighbours, and,in such case, our DFAs exaggerate the acousticdiscrimination task.
The high percentages of DFA explanationwere especially related to the temporal parame-ters of the advertisement calls in the two breed-ing seasons. This pattern corroborates Gasser etal. (2009) and Pettitt et al. (2013), who reportedthe temporal parameters of the call as the ma-jor contributors for individual distinction in A.
femoralis and Anomaloglossus beebei, respec-tively. The dominant frequency also contributedto the distinctness of S. constrictus individuals,albeit less than the temporal parameters. Theseresults indicate that all the acoustic propertiesexamined for this species are potentially usefulin the recognition of individuals. In this case,not only are parameters with CVA/CVW > 1important in the recognition system, but param-eters with greater variation among males arealso important.
Acoustic parameters may have a hereditarybasis (Smith and Hunter, 2005), which shouldrestrict evolutionary change in acoustic vari-ables over time. However, temporal changes ofa call can make it more perceptible and reducethe intra- or interspecific acoustic interferencein complex choruses (Bastos and Haddad, 1995;Bee et al., 2001; Penna et al., 2005; Bastos etal., 2011). In S. constrictus, acoustic parametersdid not vary among the years, most likely due toa low amount of genetic exchange between thepopulations, which would keep acoustic param-eters similar among breeding seasons. Smithand Hunter (2005) also did not report differ-ences over years for the note duration of Litoriabooroolongensis nor did Pröhl (2003) for callduration and frequency in Oophaga pumilio.Howard and Young (1998) observed variationin the call duration of Bufo americanus duringsubsequent breeding seasons, but no variation indominant frequency was found.
In summary, this is the first study to de-scribe variation among individuals, within in-dividuals, and among different breeding sea-sons of the advertisement call in S. constrictusmales in the Cerrado domain in South America.Individual distinctness of anuran vocalisationscan be used as a non-invasive bioacoustic toolin biological conservation to census threatenedpopulations (Tripp and Otter, 2006). This tech-nique can be particularly relevant for specieswith small populations and/or low reproductionrates and could be used to monitor populations.The absence of variation between seasons sug-gests that the data from different seasons can

Call variation of Scinax constrictus 9
be pooled in population-level studies of anurancalls. Although this assumption should be testedin additional studies, our results are encourag-ing and can be especially valuable in taxonomicand evolutionary studies when data are scarceand collected in different years.
Acknowledgements. Financial support was provided byCNPq (Conselho Nacional de Desenvolvimento Científicoe Tecnológico; procs.304350/2004-4; 555286/2006-1). R.P.Bastos is grateful to CNPq and L. Signorelli and P.G. Gam-bale to Capes (Coordenação de Aperfeiçoamento de Pes-soal de Nível Superior) for fellowships. Instituto ChicoMendes de Conservação da Biobiversidade (ICMBio/RAN)provided collecting permits. We also thank the Lima familyfor granting us access to the study area and logistical sup-port, A.R. Morais, H.S. Santana and A. Rodríguez for helpin statistical analyses, V.G. Batista, R. Vieira, R. Márquez,A. Rodríguez and anonymous reviewer for critical readingof the manuscript.
References
Ball, G.F. (1999): The neuroendocrine basis of seasonalchanges in vocal behaviour among songbirds. In: TheDesign of Animal Communication, p. 213-253. Hauser,M.D., Konishi, M., Eds, MIT Press, Cambridge.
Bastos, R.P., Haddad, C.F.B. (1995): Vocalizations andacoustic interactions in Hyla elegans (Anura, Hylidae)during the reproductive activity. Naturalia São Paulo 20:165-176.
Bastos, R.P., Alcantara, M.B., Morais, A., Lignau, R., Lima,L.P., Pombal Jr., J.P., Signorelli, L. (2011): Vocal be-haviour and conspecifc call response in Scinax centralis.Herpetol. J. 21: 43-50.
Bee, M.A., Kozic, C.E., Blackwell, K.J., Gerhardt, H.C.(2001): Individual variation in advertisement calls of ter-ritorial male green frogs, Rana clamitans: Implicationsfor individual discrimination. Ethol. 107: 65-84.
Bee, M.A., Gerhardt, H.C. (2002): Individual voice recog-nition in a territorial frog (Rana catesbeiana). Proc. R.Soc. London, Ser. B 269: 1443-1448.
Bee, M.A. (2004): Within-individual variation in bullfrogvocalizations: implications for a vocally mediated socialrecognition system. J. Acoust. Soc. Am. 166: 3770-3781.
Bee, M.A., Cook, J.M., Love, E.K., O’Bryan, L.R., Pettitt,B.A., Schrode, K., Velez, A. (2010): Assessing acous-tic signal variability and the potential for sexual selec-tion and social recognition in Boreal chorus frogs (Pseu-dacris maculata). Ethol. 116: 564-576.
Bee, M.A., Schwartz, J.J., Summers, K. (2013): All’s wellthat begins wells: celebrating 60 years of Animal Be-haviour and 36 years of research on anuran social be-haviour. Anim. Behav. 85: 5-18.
Beecher, M.D. (1989): Signalling systems for individualrecognition: an information theory approach. Anim. Be-hav. 38: 248-261.
Beer, C.G. (1970): Individual recognition of voice in thesocial behavior of birds. Adv. Study Behav. 3: 27-74.
Bernal, X.E., Guarnizo, C., Lüddecke, H. (2005): Geo-graphic variation in advertisement call and genetic struc-ture of Colostethus palmatus (Anura, Dendrobatidae)from the Colombian Andes. Herpetologica 61: 395-408.
Bernal, X.E., Rand, A.S., Ryan, M.J. (2007): Sex differ-ences in response to nonconspecific advertisement calls:receiver permissiveness in male and female túngarafrogs. Anim. Behav. 73: 955-964.
Briggs, V.S. (2010): Call trait variation in Morelett’s treefrog, Agalychnis moreletii, of Belize. Herpetologica 66:241-249.
Brooke, P.N., Alford, R.A., Schwarzkopf, L. (2000): Envi-ronmental and social factors influence chorusing behav-ior in a tropical frog: examining various temporal andspatial scales. Behav. Ecol. Sociobiol. 49: 79-87.
Buchanan, B.W. (1993): Effects of enhanced lighting onthe behaviour of nocturnal frogs. Anim. Behav. 45: 893-899.
Byrne, P.G. (2008): Strategic male calling behavior in anaustralian terrestrial toadlet (Pseudophryne bibronii).Copeia 2008: 57-63.
Castellano, S., Cuatto, B., Rinella, R., Rosso, A., Gia-coma, C. (2002): The advertisement call of the Europeantreefrog (Hyla arborea): a multilevel study of variation.Ethol. 108: 75-89.
Castellano, S., Rosso, A. (2006): Variation in call temporalproperties and female preferences in Hyla intermedia.Behaviour 143: 405-424.
Castellano, S.B.R., Giacoma, C., Dujsebayeva, T., Odierna,G., Balletto, E. (1998): Morphometrical and acousticalcomparison between diploid and tetraploid green toads.Biol. J. Linn. Soc. 63: 257-281.
Duellman, W.E., Trueb, L. (1994): Biology of Amphibians.McGraw-Hill, New York.
Eiten, G. (1994): Vegetação do cerrado. In: Cerrado car-acterização, ocupação e perspectivas, p. 17-73. Pinto,M.N., Ed., Editora da Universidade de Brasília, Brasília.
Friedl, T.W.P., Klump, G.M. (2002): The vocal behavior ofmale European treefrogs (Hyla arborea): Implicationsfor inter- and intrasexual selection. Behaviour 139: 113-136.
Friedl, T.W.P. (2006): Individual male calling pattern andmale mating success in the European treefrog (Hylaarborea): is there evidence for directional or stabilizingselection on male calling behaviour? Ethol. 112: 116-126.
Gasser, H., Amézquita, A., Hodl, W. (2009): Who is call-ing? Intraspecific call variation in the aromobatid frogAllobates femoralis. Ethol. 115: 596-607.
Gentner, T.Q., Hulse, S.H., Bentley, G.E., Ball, G.F. (2000):Individual vocal recognition and the effect of partial le-sions to HVc on discrimination, learning, and catego-rization of conspecific song in adult songbirds. J. Neu-robiol. 42: 117-133.

10 P.G. Gambale, L. Signorelli, R.P. Bastos
Gerhardt, H.C. (1991): Female mate choice in treefrogs:static and dynamic acoustic criteria. Anim. Behav. 42:615-635.
Gerhardt, H.C. (2002): Neuroethology of sound communi-cation in anurans (frogs and toads). In: Encyclopediaof Neuroscience Oxford, p. 1904. Adelman, G., Smith,B.H., Eds, EUA, Universidade de Oxford.
Gerhardt, H.C., Huber, F. (2002): Acoustic Communicationin Insects and Anurans: Common Problems and DiverseSolutions. The University of Chicago Press, Chicago.
Guimarães, L.D., Bastos, R.P. (2003): Vocalizações e inter-ações acústicas em Hyla raniceps (Anura, Hylidae) du-rante a atividade reprodutiva. Iheringia, Sér. Zool. 93:149-158.
Howard, R.D., Young, J.R. (1998): Individual variation inmale vocal traits and female mating preferences in Bufoamericanus. Anim. Behav. 55: 1165-1179.
Jouventin, P., Aubin, T., Lengagne, T. (1999): Finding aparent in a King Penguin colony: the acoustic systemof individual recognition. Anim. Behav. 57: 1175-1183.
Kaefer, I.K., Lima, A.P. (2012): Sexual signals of the Ama-zonian frog Allobates paleovarzensis: geographic varia-tion and stereotypy of acoustic traits. Anim. Behav. 149:15-33.
Lesbarrères, D., Lodé, T. (2002): Variations in male callsand responses to an unfamiliar advertisement call aterritorial breeding anuran, Rana dalmatina: evidencefor a dear-enemy effect. Ethol. Ecol. Evol. 14: 287-295.
Lima, L.P., Bastos, R.P., Giaretta, A.A. (2004): A newScinax Wagler, 1830 of the S. rostratus group fromcentral Brazil (Amphibia, Anura, Hylidae). Arq. Mus.Nac. 62: 505-512.
Lingnau, R., Bastos, R.P. (2007): Vocalizations of theBrazilian torrent frog Hylodes heyeri (Anura: Hylodi-dae): Repertoire and influence of air temperature on ad-vertisement call variation. J. Nat. Hist. 41: 1227-1235.
Manly, B.F.J. (2008): Métodos estatísticos multivariados:Uma introdução, 3rd Edition. Editora Artmed, Brasil.
Márquez, R., Eekhout, X.R. (2006): Advertisement calls ofsix species of anurans from Bali, Republic of Indonesia.J. Nat. Hist. 40: 571-588.
Martins, I.A., Jim, J. (2003): Bioacoustic analysis of adver-tisement call in Hyla nana and Hyla sanborni in Botu-catu, São Paulo, Brazil. Rev. Bras. Biol. 63: 507-516.
Martof, B. (1953): Home range and movements of the greenfrog, Rana clamitans. Ecology 34: 529-543.
McComb, K., Moss, C., Sayialel, S., Baker, L. (2000):Unusually extensive networks of vocal recognition inAfrican elephants. Anim. Behav. 59: 1103-1109.
McKibben, J.R., Bass, A.H. (1998): Behavioral assessmentof acoustic parameters relevant to signal recognition andpreference in a vocal fish. J. Acoust. Soc. Am. 104:3521-3533.
Morais, A.R., Batista, V.G., Gambale, P.G., Signorelli,L., Bastos, R.P. (2012): Acoustic communication in aNeotropical frog (Dendropsophus minutus): vocal reper-toire, variability, and individual discrimination. Her-petol. J. 22: 249-257.
Nelson, D.A., Marler, P. (1990): The perception of bird-song and an ecological concept of signal space. In: Com-parative Perception, Vol. 2, p. 443-478. Stebbins, W.C.,Berkley, M.A., Eds, Wiley, New York.
Nottebohm, F., Nottebohm, M.E. (1978): Relationship be-tween song repertoire and age in the Canary, Serinus ca-naries. Z. Tierpsychol. 46: 298-305.
Oseen, K.L., Wassersug, R.J. (2002): Environmental factorsinfluencing calling in sympatric anurans. Oecologia 133:616-625.
Penna, M., Pottstock, H., Velasquez, N. (2005): Effect ofnatural and synthetic noise on evoked vocal responses ina frog of the temperate austral forest. Anim. Behav. 70:639-651.
Pettitt, B.A., Bourne, G.R., Bee, M.A. (2013): Advertise-ment call variation in the golden rocket frog (Anoma-loglossus beebei): Evidence for individual distinctive-ness. Ethol. 119: 244-256.
Pröhl, H. (2003): Variation in male calling behavior andrelation to male mating success in the strawberry poisonfrog (Dendrobates pumilio). Ethol. 109: 273-290.
R Development Core Team (2009): R: a Language andEnvironment for Statistical Computing. R Foundationfor Statistical Computing, Vienna (Austria). Availableon: http://www.R-project.org. Access on: 14/01/2013.
Reinhold, K. (2009): Variation of acoustic courtship signalsin insects and amphibians: No evidence for bimodality,but identical dependence on duration. Ethol. 115: 134-140.
Richardson, C., Lena, J.P., Joly, P., Lengagne, T. (2008): Areleaders good mates? A study of call timing and malequality in a chorus situation. Anim. Behav. 76: 1487-1495.
Rodríguez, A., De la Nuez, D., Alonso, R. (2010): In-traspecific variation in the advertisement call of thecloud-forest frog Eleutherodactylus glamyrus (Anura:Eleutherodactylidae). J. Herpetol. 44: 457-466.
Searcy, W.A., Nowicki, S. (2005): The Evolution of AnimalCommunication. Reliability and Deception in SignalingSystems. Monographs in Behavior and Ecology. Prince-ton University Press, New Jersey.
Sèbe, F., Duboscq, J., Aubin, T., Ligout, S., Poindron, P.(2010): Early vocal recognition of mother by lambs:contribution of low- and high-frequency vocalizations.Anim. Behav. 79: 1055-1066.
Seddon, N. (2005): Ecological adaptation and speciesrecognition drives vocal evolution in neotropical sub-oscine birds. Evolution 59: 200-215.
Silva, R.A., Martins, I.A., Rossa-Feres, D.C. (2008):Bioacústica e sítio de vocalização em taxocenosesde anuros de área aberta no noroeste paulista. BiotaNeotrop. 8: 123-134.
Smith, M.J., Roberts, J.D. (2003): Call structure may affectmale mating success in the quacking frog Crinia geor-giana (Anura: Myobatrachidae). Behav. Ecol. Sociobiol.53: 221-226.
Smith, M.J., Hunter, D. (2005): Temporal and geographicvariation in the advertisement call of the booroolongfrog (Litoria booroolongensis: Anura: Hylidae). Ethol.111: 1103-1115.

Call variation of Scinax constrictus 11
Sousa-Lima, R.S., Paglia, A.P., Da Fonseca, G.A.B. (2002):Signature information and individual recognition in theisolation calls of Amazonian manatees, Trichechus inun-guis (Mammalia: Sirenia). Anim. Behav. 63: 301-310.
StatSoft (2006): STATISTICA (data analysis software sys-tem), version 7.1. http://www.statsoft.com.
Sullivan, B.K., Hinshaw, S.H. (1992): Female choice andselection on male calling behaviour in the grey treefrogHyla versicolor. Anim. Behav. 44: 733-744.
Tárano, Z. (2001): Variation in male advertisement calls inthe Neotropical frog Physalaemus enesefae. Copeia 4:1064-1072.
Toledo, L.F., Haddad, C.F.B. (2005): Acoustic repertoireand calling behavior of Scinax fuscomarginatus (Anura,Hylidae). J. Herpetol. 39: 455-464.
Tripp, T.M., Otter, K.A. (2006): Vocal individuality as a po-tential long-term monitoring tool for Western Screech-owls, Megascops kennicottii. Can. J. Zool. 84: 744-753.
Valdujo, P.H., Silvano, D.L., Colli, G., Martins, M. (2012):Anuran species composition and distribution patterns inBrazilian Cerrado, a Neotropical hotspot. South Ameri-can J. Herpetol. 7: 63-78.
Wells, K.D. (1977): The social behaviour of anuran amphib-ians. Anim. Behav. 25: 666-693.
Wells, K.D. (2007): The Ecology and Behaviour of Am-phibians. University of Chicago Press, Chicago.
Witte, K., Ryan, M.J., Wilczynski, W. (2001): Changesin the frequency structure of a mating call decreaseits attractiveness to females in the cricket frog Acriscrepitans. Ethol. 107: 685-699.
Zar, J.H. (1999): Biostatistical Analysis, 4th Edition.Prentice-Hall, New Jersey.
Submitted: April 10, 2013. Final revision received: April 25,2014. Accepted: April 27, 2014.Associate Editor: Julian Glos.




















