
In vivo grapevine anthocyanin transport involves vesicle-mediated trafficking and the contribution...
11
In vivo grapevine anthocyanin transport involves vesicle- mediated trafficking and the contribution of anthoMATE transporters and GST Camila Gomez 1 , Genevie ` ve Conejero 2 , Laurent Torregrosa 3 , Ve ´ ronique Cheynier 1 , Nancy Terrier 1 and Agne ` s Ageorges 1,* 1 INRA, UMR1083 Sciences pour l’œnologie, F-34060 Montpellier, France, 2 INRA, UMR5004 Biochimie et Physiologie Mole ´ culaire des Plantes, Plateforme Histocytologie et Imagerie Cellulaire Ve ´ge ´ tale, F-34398 Montpellier, France, and 3 INRA SupAgro, UMR1097 DIA PC, F-34060 Montpellier, France Received 22 April 2011; accepted 19 May 2011; published online 18 July 2011. * For correspondence (fax +33 (0) 499 612 857; e-mail [email protected]). SUMMARY In cells, anthocyanin pigments are synthesized at the cytoplasmic surface of the endoplasmic reticulum, and are then transported and finally accumulated inside the vacuole. In Vitis vinifera (grapevine), two kinds of molecular actors are putatively associated with the vacuolar sequestration of anthocyanins: a glutathione-S- transferase (GST) and two MATE-type transporters, named anthoMATEs. However, the sequence of events by which anthocyanins are imported into the vacuole remains unclear. We used MYBA1 transformed hairy roots as a grapevine model tissue producing anthocyanins, and took advantage of the unique autofluorescence of anthocyanins to study their cellular trafficking. In these tissues, anthocyanins were not only visible in the largest vacuoles, but were also present at higher concentrations in several vesicles of different sizes. In the cell, small vesicles actively moved alongside the tonoplast, suggesting a vesicular trafficking to the vacuole. Subcellular localization assays revealed that anthoMATE transporters were closely related with these small vesicles, whereas GST was localized in the cytoplasm around the nucleus, suggesting an association with the endoplasmic reticulum. Furthermore, cells in hairy roots expressing anthoMATE antisense did not display small vesicles filled with anthocyanins, whereas in hairy roots expressing GST antisense, anthocyanins were accumulated in vesicles but not in the vacuole. This suggests that in grapevine, anthoMATE transporters and GST are involved in different anthocyanin transport mechanisms. Keywords: anthocyanin, compartmentation, grapevine, transport, autofluorescence. INTRODUCTION The anthocyanins constitute a major flavonoid group that is widespread in plants, and in the foods made from them. They are pigments responsible for the characteristic red, blue and purple colors of many plant tissues (Harborne and Williams, 2000), which play a role in plants as visual attracters for pollinators and seed disseminators, and may be involved in protection against biotic and non-biotic stress (Harborne and Williams, 2000; Winkel-Shirley, 2001). Anthocyanins are synthesized by multi-enzyme complexes that are localized at the cytoplasmic face of the endoplasmic reticulum (ER) (Winkel-Shirley, 1999; Winkel, 2004), and then transported to the large vacuole (Grotewold, 2004). In some plants, anthocyanins are found in highly pig- mented bodies located inside the vacuole (Markham et al., 2000; Lin et al., 2003; Zhang et al., 2006; Poustka et al., 2007). These anthocyanin aggregates, referred to as anthocyaninc vacuolar inclusions (AVIs), would allow color intensification by concentrating anthocyanins above the levels that are possible in vacuolar solution (Markham et al., 2000). In Vitis vinifera (grapevine) suspension cultures, AVIs selectively accumulate acylated anthocyanins (Conn et al., 2003, 2010), suggesting that AVIs serve a specific role in aggregating or sequestering anthocyanins with particular modifications. The mechanism of anthocyanin transport to the vacuole has long been debated, with numerous models proposed such as membrane vesicle-mediated or membrane trans- porter-mediated transport (Kitamura, 2006; Grotewold and Davies, 2008; Zhao and Dixon, 2009). To date, several 960 ª 2011 The Authors The Plant Journal ª 2011 Blackwell Publishing Ltd The Plant Journal (2011) 67, 960–970 doi: 10.1111/j.1365-313X.2011.04648.x
Transcript of In vivo grapevine anthocyanin transport involves vesicle-mediated trafficking and the contribution...

In vivo grapevine anthocyanin transport involves vesicle-mediated trafficking and the contribution of anthoMATEtransporters and GST
Camila Gomez1, Genevieve Conejero2, Laurent Torregrosa3, Veronique Cheynier1, Nancy Terrier1 and Agnes Ageorges1,*
1INRA, UMR1083 Sciences pour l’œnologie, F-34060 Montpellier, France,2INRA, UMR5004 Biochimie et Physiologie Moleculaire des Plantes, Plateforme Histocytologie et Imagerie
Cellulaire Vegetale, F-34398 Montpellier, France, and3INRA SupAgro, UMR1097 DIA PC, F-34060 Montpellier, France
Received 22 April 2011; accepted 19 May 2011; published online 18 July 2011.*For correspondence (fax +33 (0) 499 612 857; e-mail [email protected]).
SUMMARY
In cells, anthocyanin pigments are synthesized at the cytoplasmic surface of the endoplasmic reticulum, and
are then transported and finally accumulated inside the vacuole. In Vitis vinifera (grapevine), two kinds of
molecular actors are putatively associated with the vacuolar sequestration of anthocyanins: a glutathione-S-
transferase (GST) and two MATE-type transporters, named anthoMATEs. However, the sequence of events by
which anthocyanins are imported into the vacuole remains unclear. We used MYBA1 transformed hairy roots
as a grapevine model tissue producing anthocyanins, and took advantage of the unique autofluorescence of
anthocyanins to study their cellular trafficking. In these tissues, anthocyanins were not only visible in the
largest vacuoles, but were also present at higher concentrations in several vesicles of different sizes. In the cell,
small vesicles actively moved alongside the tonoplast, suggesting a vesicular trafficking to the vacuole.
Subcellular localization assays revealed that anthoMATE transporters were closely related with these small
vesicles, whereas GST was localized in the cytoplasm around the nucleus, suggesting an association with the
endoplasmic reticulum. Furthermore, cells in hairy roots expressing anthoMATE antisense did not display
small vesicles filled with anthocyanins, whereas in hairy roots expressing GST antisense, anthocyanins were
accumulated in vesicles but not in the vacuole. This suggests that in grapevine, anthoMATE transporters and
GST are involved in different anthocyanin transport mechanisms.
Keywords: anthocyanin, compartmentation, grapevine, transport, autofluorescence.
INTRODUCTION
The anthocyanins constitute a major flavonoid group that is
widespread in plants, and in the foods made from them.
They are pigments responsible for the characteristic red,
blue and purple colors of many plant tissues (Harborne
and Williams, 2000), which play a role in plants as visual
attracters for pollinators and seed disseminators, and may
be involved in protection against biotic and non-biotic
stress (Harborne and Williams, 2000; Winkel-Shirley, 2001).
Anthocyanins are synthesized by multi-enzyme complexes
that are localized at the cytoplasmic face of the endoplasmic
reticulum (ER) (Winkel-Shirley, 1999; Winkel, 2004), and then
transported to the large vacuole (Grotewold, 2004).
In some plants, anthocyanins are found in highly pig-
mented bodies located inside the vacuole (Markham et al.,
2000; Lin et al., 2003; Zhang et al., 2006; Poustka et al., 2007).
These anthocyanin aggregates, referred to as anthocyaninc
vacuolar inclusions (AVIs), would allow color intensification
by concentrating anthocyanins above the levels that are
possible in vacuolar solution (Markham et al., 2000). In Vitis
vinifera (grapevine) suspension cultures, AVIs selectively
accumulate acylated anthocyanins (Conn et al., 2003, 2010),
suggesting that AVIs serve a specific role in aggregating or
sequestering anthocyanins with particular modifications.
The mechanism of anthocyanin transport to the vacuole
has long been debated, with numerous models proposed
such as membrane vesicle-mediated or membrane trans-
porter-mediated transport (Kitamura, 2006; Grotewold and
Davies, 2008; Zhao and Dixon, 2009). To date, several
960 ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd
The Plant Journal (2011) 67, 960–970 doi: 10.1111/j.1365-313X.2011.04648.x

molecular players involved in the sequestration of anthocy-
anins have been identified. The importance of glutathione-
S-transferases (GSTs) for anthocyanin transport has been
demonstrated in Zea mays (maize) BZ2 (Marrs et al., 1995),
petunia AN9 (Alfenito et al., 1998) and Arabidopsis TT19
(Kitamura et al., 2004), where mutations in the encoding
genes led to a reduction in anthocyanin accumulation and
a pigment mislocalization. Although a mechanism similar
to detoxification processes was proposed for anthocyanin
(Marrs et al., 1995), no anthocyanin-glutathione conjugate
has been observed in vitro in the presence of GST. This
protein is therefore purported to act as a cytoplasmic
anthocyanin carrier protein in vivo (Mueller et al., 2000;
Mueller and Walbot, 2001). Two major transporter families,
the ABC-C and MATE families would be also involved in
anthocyanin transport in maize (Goodman et al., 2004),
Arabidopsis (Marinova et al., 2007) and grape (Gomez et al.,
2009). However, the involvement of these different proteins
in anthocyanin transport to the vacuole is still unclear.
Evidence has emerged in favor of vesicle-mediated traf-
ficking, where vesicles filled with anthocyanins or other
flavonoids travel from the ER to the tonoplast (Grotewold
and Davies, 2008). In lisianthus and Arabidopsis, anthocya-
nins are localized in vesicles that resemble ER bodies, which
may play a role in transport to the vacuole (Zhang et al.,
2006; Poustka et al., 2007). In Brassica tapetum cells, flavo-
noids have been localized first in the ER network and then
in ER-derived tapetosomes for their delivery to the pollen
surface, suggesting the existence of flavonoid vesicle traf-
ficking (Hsieh and Huang, 2007).
In grapevine, a GST-like gene was found to be upregulat-
ed in red berries (Ageorges et al., 2006). It complemented
the maize mutant BZ2, suggesting its involvement in antho-
cyanin transport (Conn et al., 2008). Recently Gomez et al.
(2009) identified two MATE proteins, anthoMATE1 (AM1)
and anthoMATE3 (AM3), which specifically mediated acyl-
ated anthocyanin transport in vitro. AM1 and AM3 were
localized mainly at the tonoplast and in membrane vesicles
attached to the nucleus (Gomez et al., 2009), suggesting that
they participate in the transport of cytoplasmic-synthesized
anthocyanins across the tonoplast. However, these obser-
vations were performed in grapevine hairy roots not accu-
mulating anthocyanins, and drawing definitive conclusions
requires further investigation in tissues accumulating
anthocyanins. The ectopic expression of anthocyanin tran-
scription factor MYBA1 in grapevine plants and hairy roots
led to the production and vacuolar storage of anthocyanins,
associated with the overexpression of GST and AM3
(Cutanda-Perez et al., 2009). The presence of AVIs was
observed, along with intense coloration and a high accu-
mulation of anthocyanins in the transformed tissues.
Recently, in V. vinifera cell suspension cultures treated with
sucrose and jasmonic acid, white-light microscopy studies
showed that small vesicles (termed AVIs, despite not
definitively being vacuolar vesicles) could proceed from
the cytoplasm across the tonoplast, and were able to fuse
intravacuolarly to give fewer, larger AVIs as the cells mature
(Conn et al., 2010). Overall, the mechanisms by which
anthocyanins and other flavonoids are imported from the
cytoplasmic surface of the ER to the vacuole are not yet
clearly identified (Grotewold, 2004; Zhao and Dixon, 2009).
In this study, we investigated the relationship between the
anthocyanin compartmentation and the subcellular locali-
zation of anthocyanin transporters to gain a further under-
standing of the relationship between transport and the
sequestration of these pigments. We identified vesicle-like
bodies of different sizes filled with anthocyanins. Inside the
cell, the small ones, localized in the cytoplasm, moved
actively alongside the tonoplast, suggesting a vesicular
trafficking. We also investigated the subcellular localization
of GST and anthoMATE transporters (AMs), and the impact
of antisense construction for these two genes in relation to
anthocyanin compartmentation. These observations sug-
gest that in grapevine, anthoMATE transporters and GST are
involved in different anthocyanin transport mechanisms.
RESULTS
Anthocyanins compartmentation in hairy root cells
derived from grapevine tissues producing anthocyanins
In order to characterize the anthocyanin compartmentation,
we generated grapevine tissues producing anthocyanins.
Transformed plantlets regenerated after the expression of
MYBA1 in embryogenic calluses (Cutanda-Perez et al., 2009)
displayed red coloration and accumulated anthocyanins.
From theses plantlets, we generated transgenic hairy roots
(HRs) expressing, under the cauliflower mosaic virus 35S
promoter (p35S), either GFP alone (control HR) or GFP fused
with HDEL (HR-GFPHDEL). HDEL corresponds to an ER
retention signal sequence that can be used as an ER marker
(Haseloff et al., 1997; Batoko et al., 2000). These transgenic
HRs harbored a violet-brown coloration that became
noticeable as soon as the root length reached a few mm.
They presented a thin diameter (1–2 mm) and displayed
reduced growth and secondary branching compared with
HRs generated from wild-type plantlets (Cutanda-Perez
et al., 2009). Transverse sections of these transgenic HRs
revealed that the epidermis and the vascular cylinder were
the most intensely colored tissues, whereas the cortex
parenchyma appeared colorless, with only a few randomly
located red cells (Figure 1a).
Wide-field microscopic observations in the pigmented
epidermal cells showed that the coloration was essentially
the result of an accumulation of anthocyanins in the large
vacuoles, which tended to occupy most of the cellular
volume (Figure 1b,c). In these pigmented cells, we observed
a high density of spherical structures, which appeared much
more intensely colored than the whole vacuole (Figure 1b,c).
Anthocyanin trafficking in grapevine 961
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), 67, 960–970

These colored structures look like vesicles and can be
classified into two groups according to their size: group I
(G.I) corresponds to very small structures (0.2–0.5 lm in
diameter), and group II (G.II) corresponds to spherical
structures larger than 1 lm in diameter. Most of the
pigmented cells accumulated anthocyanins simultaneously
in vacuoles, and in G.I and G.II structures (Table 1). One-
third of pigmented cells contained anthocyanins only in
vacuoles and in large G.II structures. By contrast, almost no
cell accumulating anthocyanins only in vesicle-like struc-
tures or exclusively in vacuoles was found (Table 1). Obser-
vations of a pigmented cell in different z-sections confirmed
the presence of several G.I structures that actively moved
inside the cytoplasm (Video Clip S1). The G.II structures,
which are very heterogeneous in size (ranging from 1 to over
15 lm), were motionless, and their localization was less
clear.
In order to determine the subcellular localization of G.I
and G.II anthocyanin-filled structures, these tissues were
examined using confocal microscopy. Anthocyanins exhibit
autofluorescent properties when excited at 543 nm (Poustka
et al., 2007). Using this autofluorescence, we observed a
similar organization to that observed in Figure 1b,c (Fig-
ure 1d). We detected numerous small vesicle-like structures
(G.I) around what we supposed to be the vacuole, and a few
large structures (G.II) localized against a boundary, assumed
to be the tonoplast, and close to G.I (Figure 1d). However, in
our experimental conditions, the subcellular localization of
(a)
(b)
(h)
(c) (d)
(g)(f)(e)
Figure 1. Wide-field and confocal microscopy images of grapevine hairy roots (HRs) producing anthocyanins. Wide-field microscopy images of control HR
transverse section (a) and pigmented epidermal cell (b) showing colored vacuoles and spherical vesicles filled with anthocyanins. Light transmitted images of
control HR longitudinal section (c) and image of vesicles in the same cell (d) showing anthocyanin autofluorescence. Visualization of specific signal of HR-GFPHDEL
in a pigmented cell by confocal microscopy (e, red channel for anthocyanins; f, green channel for GFP; g, overlay of both channels). Visualization of lipids by Nile Red
staining in a pigmented cell with epifluorescence microscope (h). Anthocyanins appear inside vesicles of different size (G.I < 1 lm in diameter; G.II > 1 lm in
diameter). G.I vesicles are visible outside the vacuole delimited by the tonoplast (b, c). In HR-GFPHDEL, anthocyanin autofluorescence appears in G.I vesicles
surrounding the nucleus, and is co-localized with the GFP-HDEL fluorescence (g). DNA is stained in blue with DAPI, red autofluorescence is generated by
anthocyanins and green fluorescence by GFP from the GFP-HDEL fusion protein. Scale bars: 0.5 mm (a); 5 lm (e, f and G); 10 lm (c, d and h). Cp, cytoplasm;
ep, epidermis; ER, endoplasmic reticulum; N, nucleus; V, vacuole; vc, vascular cylinder.
Table 1 Distribution of pigmented cells in control (i.e. MybA1-transformed) hairy roots according to the type of subcellularcompartment in which the anthocyanins were accumulated. Cellswere sorted based on the presence of anthocyanins in vacuoles (CV)and/or small (G.I) and/or large (G.II) vesicles
Compartments
% of cellsaccumulatinganthocyanins
CV + G.I + G.II 68.4 � 10.5a
CV only 1.3 � 1.5b
G.II only 0CV + G.II 30.2 � 7.8a
G.I + G.II 0
Results are expressed as a percentage of the total number of cellsobserved containing anthocyanins. Values with different letters (a, b)indicate significance at P < 0.05. Values are means � SDs from thepercentage of n epidermal cells analysed from the independent hairyroot lines (n1 = 297, n2 = 244, n3 = 237).
962 Camila Gomez et al.
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), 67, 960–970
G.II was difficult to establish. Using HR-GFPHDEL, in which
the GFP-HDEL marker is retained in the ER, we confirmed the
cytoplasmic localization of G.I (Figure 1e,f,g). The specific
green fluorescence from the GFP-HDEL marker was detected
in a network surrounding the nucleus, and we also observed
red autofluorescence provided by G.I filled with anthocya-
nins surrounding the nucleus (Figure 1e,f). Observations
under both the red and green channels with confocal
microscopy showed the co-localization of some G.I with
the ER marker, suggesting that G.Is can be considered as
ER-derived bodies (Figure 1g). Subsequently, colored cells
of control HRs were stained by Nile red dye (Greenspan
et al., 1985). We clearly observed a yellow signal associated
with G.I (Figure 1h). By contrast, no signal was detected
around large G.II, suggesting that G.I and G.II do not have
the same lipidic composition.
In order to confirm the motion of G.I and G.II structures,
we used the spinning-disc confocal microscopy system,
which enables confocal observations of very fast processes
in living cells over a long time period (Graf et al., 2005).
Video Clip S2 presents a 4D reconstruction across the cell,
moving along the optical z-axis for 45 min. We observed G.II
structures localized in the middle of the cell that remained
motionless over the 45-min period. During the same period,
small G.Is moved actively and remained localized at the
edge of the cell (Video Clip S2). Crossing the cell, we
observed small vesicle-like structures moving around and
against large G.II structures. Overall, anthocyanins are
compartmentalized in vesicle-like structures of different
sizes: (i) small G.I structures surrounding the vacuole and
exhibiting active movement inside the cytoplasm; and (ii)
large G.II structures (up to 15 lm in diameter) that do not
move, the localization of which is unclear.
Localization of AMs and GST in epidermal cells
from HRs producing anthocyanins
To elucidate the involvement of anthoMATE transporters and
GST in anthocyanin transport, we used MYBA1 transformed
plantlets to generate transgenic HRs expressing under p35S
genes encoding GFP protein fusions of AM1 (HR-AM1GFP),
AM3 (HR-AM3GFP) and GST (HR-GSTGFP). In parallel, the
subcellular localization of these proteins was performed by
immunofluorescent labeling using anti-AM and anti-GST
antibodies in HRs producing anthocyanins (HRMYBA1).
From control sample observations, we confirmed that the
green fluorescence detected in these experiments was highly
specific to the GFP fusion protein or antibodies (Figure S1).
Moreover, immunolabeling with anti-AM and anti-GST anti-
bodies in HRs containing no anthocyanin revealed only a
very weak fluorescent signal (Figure S2). In some cases,
minimal autofluorescence was also seen at the cell wall,
possibly representing other autofluorescent compounds.
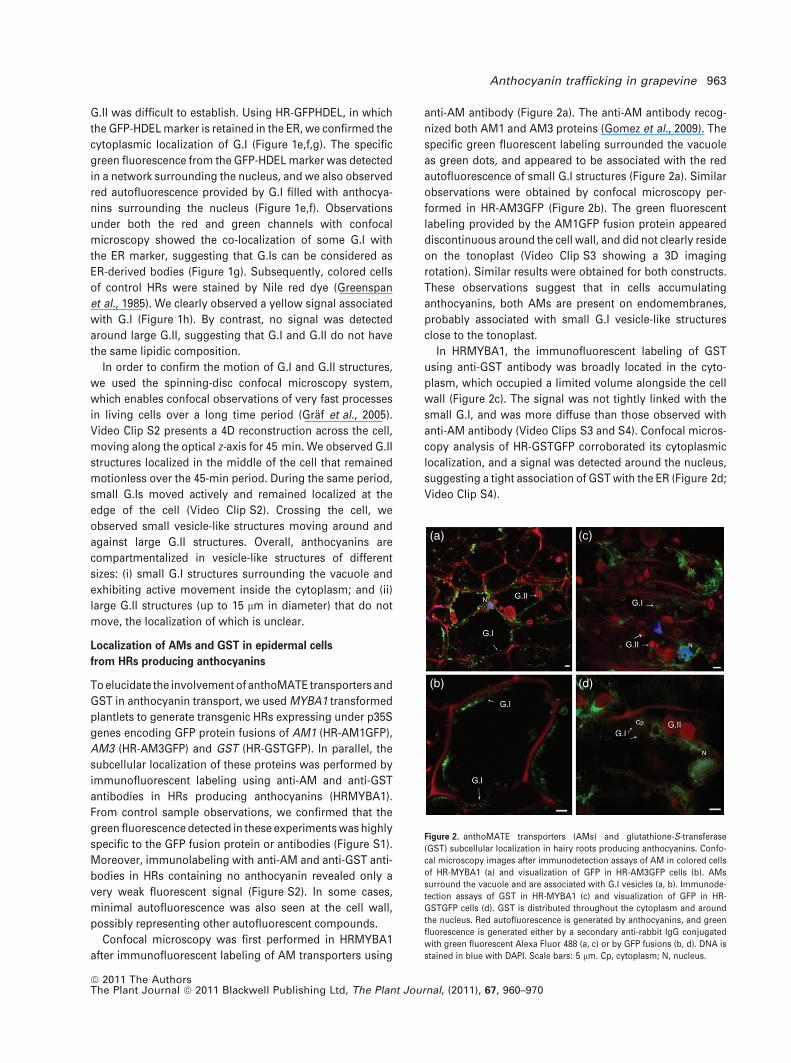
Confocal microscopy was first performed in HRMYBA1
after immunofluorescent labeling of AM transporters using
anti-AM antibody (Figure 2a). The anti-AM antibody recog-
nized both AM1 and AM3 proteins (Gomez et al., 2009). The
specific green fluorescent labeling surrounded the vacuole
as green dots, and appeared to be associated with the red
autofluorescence of small G.I structures (Figure 2a). Similar
observations were obtained by confocal microscopy per-
formed in HR-AM3GFP (Figure 2b). The green fluorescent
labeling provided by the AM1GFP fusion protein appeared
discontinuous around the cell wall, and did not clearly reside
on the tonoplast (Video Clip S3 showing a 3D imaging
rotation). Similar results were obtained for both constructs.
These observations suggest that in cells accumulating
anthocyanins, both AMs are present on endomembranes,
probably associated with small G.I vesicle-like structures
close to the tonoplast.
In HRMYBA1, the immunofluorescent labeling of GST
using anti-GST antibody was broadly located in the cyto-
plasm, which occupied a limited volume alongside the cell
wall (Figure 2c). The signal was not tightly linked with the
small G.I, and was more diffuse than those observed with
anti-AM antibody (Video Clips S3 and S4). Confocal micros-
copy analysis of HR-GSTGFP corroborated its cytoplasmic
localization, and a signal was detected around the nucleus,
suggesting a tight association of GST with the ER (Figure 2d;
Video Clip S4).
(a)
(b)
(c)
(d)
Figure 2. anthoMATE transporters (AMs) and glutathione-S-transferase
(GST) subcellular localization in hairy roots producing anthocyanins. Confo-
cal microscopy images after immunodetection assays of AM in colored cells
of HR-MYBA1 (a) and visualization of GFP in HR-AM3GFP cells (b). AMs
surround the vacuole and are associated with G.I vesicles (a, b). Immunode-
tection assays of GST in HR-MYBA1 (c) and visualization of GFP in HR-
GSTGFP cells (d). GST is distributed throughout the cytoplasm and around
the nucleus. Red autofluorescence is generated by anthocyanins, and green
fluorescence is generated either by a secondary anti-rabbit IgG conjugated
with green fluorescent Alexa Fluor 488 (a, c) or by GFP fusions (b, d). DNA is
stained in blue with DAPI. Scale bars: 5 lm. Cp, cytoplasm; N, nucleus.
Anthocyanin trafficking in grapevine 963
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), 67, 960–970

Immunodetection of AMs and GST in epicarp cells
of grape berries
To validate the results obtained in transgenic HRs in berries,
we performed immunofluorescent labeling using anti-AM
and anti-GST in fixed sections of Syrah mature grape ber-
ries. Immunofluorescent labeling was never detected when
secondary antibody was used alone (Figure S1). Under
wide-field microscopic observations, the cells of berry skin
displayed numerous colored vesicle-like structures of dif-
ferent sizes (Figure 3a). Several small anthocyanin-filled
structures were observed in the small cytoplasmic space
bordering the large vacuole, suggesting that epicarp cells
have an anthocyanin compartmentation similar to that ob-
served in HRs producing anthocyanins (Figure 3a). Confocal
microscopy of sections of mature red berries revealed that
the AM signal surrounded the vacuole (Figure 3b). More-
over, we detected a high signal associated with small
spherical structures similar to G.I in HRs (Figure 3b). Finally,
immunodetection of GST in the epicarp cells of red berries
showed that this protein was distributed throughout the
cytoplasm and around the nucleus (Figure 3c). Together,
these results obtained in mature berries corroborate the
spatial distribution of AMs and GST in transgenic HRs pro-
ducing anthocyanins.
Subcellular structures of epicarp cells of grape berries
containing anthocyanin
To elucidate the anatomy of these anthocyanin-filled struc-
tures, microscopic observations were completed by trans-
mission electron microscopy (TEM) observations carried out
on transverse microsections of epidermal red cells of ber-
ries. Cells contained one or just a few large vacuolar com-
partments (Figure 3d,h). The cytoplasm of these cells had
large numbers of mitochondria, chloroplasts and vesicles
(a)
(d)
(e) (f)
(g)
(h)
(b) (c)
Figure 3. Immunolocalization of anthoMATE transporters (AMs) and glutathione-S-transferase (GST) in mature red berries, and ultrastructure of epicarp cells.
Wide-field microscopy images of epicarp from mature Syrah berries embedded in paraffin showing various red vesicles G.I and G.II in cells without staining (a).
Confocal microscopy images of epicarp cells after immunodetection assays of AM (b) and GST (c) in red Syrah berry. The green specific signal of AMs are associated
with G.I structures, surrounding the vacuole, and G.II structures (b). The green specific signal of GST is detected throughout the cytoplasm and around the nucleus
(c). DNA is stained in blue with DAPI. Red autofluorescence is generated by anthocyanins, and green fluorescence by a secondary anti-rabbit IgG conjugated with
green fluorescent Alexa Fluor 488. Ultrastructural evaluation by TEM (d–h): general view of a long cell from the epicarp showing various vesicles in the cytoplasm
(d, h); close-up image of a cytoplasmic region closed to the tonoplast showing G.I vesicles (e). Visualization of G.I under high magnification showing a membrane
around these structures (f). Part of an epicarp cell showing G.I structures penetrating the vacuole (g, h). Scale bars: 20 lm (a); 5 lm (b and c); 1 lm (d and h); 0.5 lm
(d insert); 100 nm (e and f); 500 nm (g). Chl, chloroplast; Cy, cytoplasm; CW, cell wall; m, mitochondria; N, nucleus; V, vacuole.
964 Camila Gomez et al.
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), 67, 960–970

alongside the vacuolar membrane (Figure 3d,e). The cyto-
plasmic vesicle size was around 200–300 nm (Figure 3e).
The shape, the size and the location of these cytoplasmic
vesicles were similar to the anthocyanin-filled structures
observed under wide-field microscopy (Figure 3a). These
cytoplasmic vesicles were clearly surrounded by an elec-
tronic-dense membrane (Figure 3e,f). As anthocyanins were
more or less solubilized during the dehydratation step, with
ethanol required for the sample preparation of TEM, the
anthocyanin-filled bodies appeared in some cases to be non-
osmiophilic. At higher magnification, we observed small
cytoplasmic vesicles bridging the tonoplast, penetrating
by endocytosis into the large vacuole (Figure 3g,h). In our
conditions and in the observed cells, no intravacuolar
membranous fragment was observed.
Hairy roots producing anthocyanins and expressing
antisense AMs and GST
Transgenic HRs expressing, under p35S, genes encoding
antisense AM3 (HR-AM3AS) and GST (HR-GSTAS) were
generated from MYBA1-transformed plantlets. The expres-
sion of AM1, AM3 and GST was evaluated to confirm the
repression of these genes in HR-AM3AS and HR-GSTAS
when compared with control HR (Figure 4). HR-AM3AS lines
exhibited 8.3-fold less AM1 expression and 5.6-fold less AM3
expression than control HR lines (Figure 4). HR-GSTAS lines
exhibited four-fold less GST expression than control HR
lines (Figure 4). Wide-field and confocal microscopy obser-
vations performed in HR-AM3AS, HR-GSTAS and control
HR, showed altered anthocyanin compartmentation in
HR-AM3AS and HR-GSTAS compared with control HR
(Figure 5). In cells of control HR, anthocyanins accumulated
in large vacuoles and in small G.I and large G.II structures
(Figures 5 and 6a). In HR-AM3AS, anthocyanins were mainly
accumulated in the large vacuole, and in few G.II structures
(Figures 5 and 6b,c). Moreover, no G.I structures were
observed in HR-AM3AS, suggesting that the G.II structures
are not dependent on the presence of G.I structures. In
contrast, in HR-GSTAS, virtually no colored vacuoles were
detected in epidermal cells, and anthocyanins were mostly
found in G.I and G.II structures (Figures 5 and 6d,e). These
results indicate that AMs and GST are involved in different
anthocyanin transport mechanisms.
DISCUSSION
To elucidate the anthocyanin compartmentation and the
relationship between transport and sequestration of these
Figure 4. Quantitative real-time PCR expression of AM1, AM3 and GST.
Transcript levels of AM1 (a) AM3 (b) in HR-AM3AS and control HR, and GST (c)
in HR-GSTAS and control HR. Gene expression was normalized with the
expression of VvEF1-a. Data are means of results obtained from three
independent transformed lines (two for the control) with error bars indicating
SD.
Figure 5. Effect of antisense expression of AM3 and GST on the compart-
mentation of anthocyanins in epidermal cells. Percentage of cells accumulat-
ing anthocyanins in colored vacuoles (black), G.I vesicles (white) and G.II
vesicles (grey) from control hairy root (HR), HR-AM3AS and HR-GSTAS. Data
are mean values � SDs from the percentage of n epidermal cells analysed
from three independent HR lines (control HR, n1 = 297, n2 = 244, n3 = 237; HR-
AM3AS, n1 = 42, n2 = 51, n3 = 45; HR-GSTAS, n1 = 40, n2 = 36, n3 = 31).
*Significant difference between control and transformed hairy roots
(P < 0.01).
(b)(a)
(c) (e)
(d)
Figure 6. Grapevine hairy roots (HRs) expressing antisense AM3 and GST.
Wide-field microscopy images of control HR (a), HR-AM3AS (b) and HR-
GSTAS (d). Confocal microscopy images of HR-AM3AS (c) and HR-GSTAS (e).
Epidermal cells in control HR exhibited anthocyanin accumulation in colored
vacuoles, in G.I and G.II vesicles (a). Epidermal cells in HR-AM3AS displayed
anthocyanin accumulation in large dark vacuoles; no G.I vesicles were
detected (b and c). Epidermal cells in HR-GSTAS displayed anthocyanins
compartmentalized in G.I and G.II vesicles; no colored vacuole was observed
(d and e). Red autofluorescence is generated by anthocyanins. Scale bar:
5 lm. V, vacuole.
Anthocyanin trafficking in grapevine 965
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), 67, 960–970

pigments, we developed a strategy using in vivo imaging
microscopy in grape berries naturally accumulating antho-
cyanins, and in transgenic grapevine HRs producing antho-
cyanins. Both tissues accumulate anthocyanins at high
levels, and the same anthocyanins are present in both tissue
types, although in different proportions (Cutanda-Perez
et al., 2009). Wide-field microscopy and confocal micros-
copy analysis of these tissues showed identical patterns of
anthocyanin compartmentation, indicating that transgenic
HRs accumulating anthocyanins are an appropriate model to
study mechanisms involved in anthocyanin accumulation in
grape berry.
Tissues producing anthocyanins often have unique sub-
cellular structures called ‘anthocyanoplasts’ or AVIs (Pecket
and Small, 1980). In grapevine tissues, anthocyanins are
present in two types of structures, differing mainly by their
diameter and their location inside the cell. This is in accor-
dance with observations of Conn et al. (2010), who observed
small to large red bodies in grape suspension cells. However,
both were called AVIs by the authors, despite the fact that
some of the vesicles were observed outside of the vacuole. In
lisianthus flowers, Zhang et al. (2006) reported anthocyanin
accumulating as vesicle-like bodies in the cytoplasm, which
were considered as pre-vacuolar compartments. In Arabid-
opsis and maize, cytoplasmic vesicles were also identified in
cells accumulating high levels of anthocyanins (Grotewold
et al., 1998; Irani and Grotewold, 2005; Poustka et al., 2007).
In the literature, there are some contradictory microscopic
observations showing the presence (Small and Pecket, 1982;
Jasik and Vancova, 1992; Poustka et al., 2007) or absence
(Nozue et al., 1993; Markham et al., 2000) of membranes
surrounding the anthocyanin-filled vesicles.
In the present study, the color of the cytoplasmic vesicle-
like structures appeared to be red, suggesting that inside the
G.I, anthocyanins were in acidic conditions compared with
cytoplasmic neutral pH (Tanaka et al., 2008). In addition, Nile
red staining indicates the presence of lipids in G.I structures.
All these observations are in favor of the presence of a
membrane surrounding these structures in order to main-
tain the acidic conditions. Moreover, AM proteins, possess-
ing hydrophobic transmembrane domains, were closely
associated with G.I structures, and would suggest that G.Is
are membranous organelles. This corroborates the obser-
vations performed by TEM that in grapevine tissues
producing anthocyanin, we could clearly observe a mem-
brane surrounding the small cytoplasmic structures. These
cytoplasmic G.I structures could correspond to small
vesicles previously described in maize and Arabidopsis
(Grotewold et al., 1998; Irani and Grotewold, 2005; Poustka
et al., 2007).
By contrast with the cytoplasmic vesicles, and within the
set of cells observed, we did not detect any electronic-
dense membranous structures inside the vacuole. A similar
conclusion was made regarding lisianthus AVIs, for which
no membrane encompassing the AVIs inside the vacuole
could be detected, despite the complex forms of AVIs
observed (Zhang et al., 2006). In the literature, the anatom-
ical identity of these vesicles is not firmly established.
According to some authors, the ontogenesis of larger
anthocyanin-filled vesicles from smaller ones is reminiscent
of autophagy, as has already been described in maize (Irani
and Grotewold, 2005). Recently, Pourcel et al. (2010) also
suggested a role for autophagy in the formation of AVIs
in Arabidopsis. The origin of these large vesicles is thus
questionable. In grapevine tissues, the fast movements of
small G.I structures near the large G.IIs observed inside the
cell (Video Clip S2) suggest that small G.Is coalesce into
larger G.II structures, as reported in lisianthus, Arabidopsis
and grapevine suspension cells (Zhang et al., 2006; Conn
et al., 2010). However, the presence of G.IIs and the lack of
cytoplasmic G.I structures in HR-AM3AS suggest that some
G.II structures can be formed independently of the presence
of cytoplasmic G.I. The formation of large anthocyanin-
filled vesicles in grapevine tissues is thus probably the
result of different parallel mechanisms, which co-exist in
the cell.
The presence of numerous small anthocyanin-filled
structures in grapevine tissues producing anthocyanins
could be related to AM transporters. In these tissues, AM
transporters were systematically closely associated with G.I
structures. These cytoplasmic anthocyanin-filled vesicles
would then suggest the occurrence of ER-derived vesicular
transport of anthocyanins (Pecket and Small, 1980; Grote-
wold et al., 1998; Irani and Grotewold, 2005; Zhang et al.,
2006). Anthocyanin vesicular trafficking has been previ-
ously demonstrated in Arabidopsis when it accumulates
large levels of anthocyanins, as well as in lisianthus flower
petals (Zhang et al., 2006; Poustka et al., 2007). These
authors postulated that small vesicles linked to the ER
gradually developed and enlarged in the cytoplasm to
vesicles of various sizes before merging with the central
vacuole to deliver anthocyanins (Zhang et al., 2006; Poustka
et al., 2007; Conn et al., 2010). In grapevine tissues, our
in vivo images of dynamic processes also show that small
vesicles coalesce into larger vesicles. Unfortunately, the
time scale used in the video did not allow the detection of
modifications in the proportion of G.I to G.II structures, or
an increase in the volume of G.II as already observed in
previous studies (Irani and Grotewold, 2005; Conn et al.,
2010). These results raise questions about the origin of
these colored vesicles, and about the way flavonoids are
targeted to them. In vesicle-mediated transport, the traf-
ficking of proteins has been well studied and documented
in plants (Neuhaus and Paris, 2005). In contrast, the
trafficking of phytochemicals is poorly understood (Grote-
wold, 2004). In grapevine, the overexpression of flavonoid
transcription factors MYBA1, MYBPA1 and MYBPA2 in HRs
(Cutanda-Perez et al., 2009; Terrier et al., 2009) induced
966 Camila Gomez et al.
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), 67, 960–970

the expression of a set of genes associated with cellular
trafficking. Further work is clearly needed to elucidate if
anthocyanin vesicular transport takes advantage of protein
secretion machinery or develops alternative secretory
pathways.
According to previous studies (Marrs et al., 1995; Alfenito
et al., 1998; Kitamura et al., 2004), GST is one of the proteins
related to vacuolar transport of flavonoids. In grapevine
tissues accumulating anthocyanins, GST was localized in the
cytoplasm. Moreover, in HR-GSTAS, anthocyanins were
accumulated in G.I and G.II vesicles only, and not in the
vacuoles. This is in accordance with the recent results of
Kitamura et al. (2010), who also reported flavonoid accumu-
lation in small vacuolar-like vesicles in tt19 mutants. These
observations suggest that GST is not involved in the
formation of the flavonoid-filled vesicles, but is required
for the accumulation of anthocyanins into the vacuole. The
modified cellular anthocyanin compartmentation in HR-
GSTAS demonstrated the existence of anthocyanin trans-
port from the ER to the tonoplast, involving GST, in
grapevine, as had already been hypothesized in some other
plants. Nevertheless, to date the functional role of GST in the
vacuolar anthocyanin sequestration has not been clearly
described.
Two models have been classically proposed to explain
how flavonoids traffic from the surface of the ER to the
tonoplast: ligandin transport (LT) and vesicular transport
(VT) (Grotewold and Davies, 2008). According to these
models, GST would be associated with the LT mechanism,
as already described in some plants, and AMs would be
more closely related to the VT mechanism. Nevertheless,
a role of AMs in the LT mechanism cannot be excluded
because AMs were also localized to the tonoplast in cells
that do not accumulate flavonoids (Gomez et al., 2009). In
addition, HR-AM3AS still accumulate anthocyanins,
suggesting that other tonoplast transporters besides AMs
are involved, as already suggested by the specificity of AM
transporters towards acylated anthocyanins (Gomez et al.,
2009). Furthermore, the subcellular localization of GST in
the cytoplasm and around the nucleus where biosynthesis
takes place suggests that it may be involved in the escort
of anthocyanins to the tonoplast. Tissues expressing
antisense GST or AM genes still accumulate anthocyanins
not only in the cytoplasm in these cases, suggesting that
both LT and VT would coexist in HR cells. To date, GST and
transporters have been identified in the maize anthocyanin
pathway (BZ2 and MRP3) and in the Arabidopsis flavonoid
pathway (TT19 and TT12). In maize, the authors observed a
similar disruption in anthocyanin vacuolar accumulation
in both the bz2 mutant (Marrs et al., 1995) and in the
ZmMRP3 mutant (Goodman et al., 2004), suggesting that a
VT mechanism may not exist in maize, and that both GST
and MRP3 are involved in an LT mechanism. On the other
hand, in Arabidopsis seed coat cells, Kitamura et al. (2010)
observed an accumulation of PA derivatives in small
vesicles in tt19 mutants and at the cytoplasmic face of
the tonoplast in tt12 mutants, whereas flavonols were
located in the cytosol instead of inside tapetosomes in
pollen cells in both mutants (Hsieh and Huang, 2007).
Therefore, it seems that the coexistence of VT and LT
mechanisms, and the involvement of GST and transporters
in each type of mechanism, would be cell-type and/or
flavonoid-class dependant, and cannot be generalized in
the plant kingdom.
In grapevine, HRs expressing MYBA1 produce mainly
acylated anthocyanins, which are not the major form of
anthocyanins found in grape berries (Cutanda-Perez et al.,
2009). In lisianthus petals, AVIs extracted from vacuoles
selectively bind acylated diglycosides, which are the minor
form of anthocyanins in flowers (Markham et al., 2000). In
Arabidopsis, the over-accumulation of anthocyanins after
PAP1 overexpression or light stress leads to the pre-dom-
inant accumulation of acylated anthocyanins (Tohge et al.,
2005; Luo et al., 2007). In Solanum lycopersicum (tomato),
the overexpression of anthocyanin transcription factors
ANT1 and Rosea1 leads to the accumulation of acylated
anthocyanins (Mathews et al., 2003; Butelli et al., 2008).
Therefore, we cannot exclude that in our study, the over-
accumulation of anthocyanins led to a modification of a part
of the pool of anthocyanins followed by a vesicular traffick-
ing to the vacuole. This mechanism may be closely related to
the rapid removal of potentially toxic material from the
cytoplasm, and may overlap with other trafficking processes
(Snyder and Nicholson, 1990; McLusky et al., 1999; Nielsen
et al., 2004; Field et al., 2006). Therefore, the accumulation of
anthocyanins in plant tissues may be a hallmark of plant
stress, even if the role of flavonoids in the stress response is
still poorly understood (Winkel-Shirley, 2002).
Using non-destructive techniques, we were able to exam-
ine living cells containing anthocyanins, and obtained
in vivo images of dynamic processes. Convincing evidence
exists for at least GST-mediated transport, vesicle trafficking
and anthoMATE transporter mechanisms for the final vac-
uolar accumulation of anthocyanins in grapevine. The
existence of other putative transporters and the interaction
between GST with its ligand and/or tonoplastic transporters
remain to be elucidated in order to precisely characterize
these two mechanisms. Several questions concerning the
origin, structure, composition and behavior of anthocyanin-
containing vesicles remain. The characterization of these
vesicles and of their cargo should help, and may provide
additional insights into engineering this metabolic pathway.
EXPERIMENTAL PROCEDURES
Plasmid construction
The AM1-GFP, AM3-GFP and GST-GFP fusions were obtained byGateway cloning (Invitrogen, http://www.invitrogen.com). The stop
Anthocyanin trafficking in grapevine 967
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), 67, 960–970

codon-less AM1, AM3 and GST cDNAs were amplified using high-fidelity Taq Polymerase (Advantage-HF2 PCR kit; Clontech, http://www.clontech.com), according to the manufacturer’s instructions.The primers used for AM1 (accession no. GB FJ264202) and AM3(accession no. GB FJ264203) amplification are described in Gomezet al. (2009), and the following primers were designed for GST(accession no. GB AY971515): forward 5¢-CACCATGGTGATGAAGGTGTATGGCCCA-3¢ and R 5¢-AGCAGCGAGCTCCATGACTTTT-3¢.The resulting PCR products were transferred into pENTRTM/D-TOPO� vector (Invitrogen) and subsequently into pH7FWG2(Karimi et al., 2002) by TOPO and LR clonase reactions, according tothe manufacturer’s instructions.
The AM3 and GST cDNAs were amplified using high-fidelity TaqPolymerase (Advantage-HF2 PCR kit; Clontech), according to themanufacturer’s instructions. The following primers were used:AM3, forward 5¢-CACCATGGAGACACCGCTGCTCAAGAGC-3¢ andreverse 5¢-TCATACATCATCGGCTTCAATCC-3¢; GST forward 5¢-CAC-CATGGTGATGAAGGTGTATGGCCCA-3¢ and reverse 5¢-TCAAG-CAGCGAGCTCCATGACTTTT-3¢. The resulting PCR products weretransferred into pENTRTM/D-TOPO� vector (Invitrogen), and subse-quently into pH2WG7 (Karimi et al., 2002) by TOPO and LR clonasereactions, according to the manufacturer’s instructions.
Plant transformation
All transgenic HRs have been generated from MYBA1-transformedgrapevine plantlets (described in Cutanda-Perez et al., 2009). Theinduction and culture of transgenic HRs in grapevine were per-formed as described by Torregrosa and Bouquet (1997), withmodifications reported in Cutanda-Perez et al. (2009). For eachtransformation, four independent HR clones arising from separateinduction experiments were selected. Microscopy observationswere performed after two rounds of subculture.
DNA was extracted from 100 mg of fresh tissue using the DNAPlant Mini kit (Qiagen, http://www.qiagen.com). Transformantswere checked by PCR using the hygromycin primers, forward5¢-GCCTGAACTCACCGCGACGTC-3¢ and reverse 5¢-CAGTTTGCCAGTGATACAC-3¢.
Gene expression analysis
RNA was extracted from 150 mg of tissue using RNeasy Plant MiniKit (Qiagen), following the manufacturer’s protocol. QuantitativePCR analysis was performed as described in Cutanda-Perez et al.(2009). Gene transcripts were quantified upon normalization toVvEF1-a as the internal standard. Results are reported as 2)DCt,where DCt is the cycle threshold required to the log phase ofamplification for the experimental gene minus the same measurefor VvEF-a (Livak and Schmittgen, 2001). Primers used for amplifi-cation of GST and AM3 are described in Cutanda-Perez et al. (2009)and Gomez et al. (2009), respectively. All primer pairs were deter-mined to have equal amplification efficiency, and the PCR productswere checked by electrophoresis.
Subcellular localization
The anti-AM and anti-GST antibodies were obtained from rabbitsera following immunization with the peptides AALSIRVSNEL-GYGHPRAAK and KLEKVFDVYEQRLSKSR, respectively, preparedby Proteogenix (http://www.proteogenix.fr).
Immunocytolocalization of AM and GST in sections of grape
berries. Grape berry tissue preparation was performed asdescribed in Paciorek et al. (2006). Berries (cv. Syrah) wereharvested at 10 weeks post-flowering to obtain a ripe stage. The
procedure involved a fixation step with paraformaldehyde 4% in10 mM phosphate-buffered saline, pH 7.2 (PBS; 7 mM Na2HPO4,3 mM NaH2PO4, 120 mM NaCl, 2.7 mM KCl), dehydration steps withethanol at increasing concentrations and a 100% butanol bath(5 days). Tissue embedding was performed with successive bathsof increasing concentrations of Safesolv (Labonord, http://www.labonord.com) in butanol, then with Safesolv/Paraplast pure wax(paraffin X-TRA; McCormick Scientific, http://www.mccormicksci-entific.com) baths at increasing pure wax concentrations. Paraplastblocks were cut with a razor blade at 8-lm thickness (Leica micro-tome RM 2265; Leica, http://www.leica.com). Strips were placed ona warm water surface on a heating plate and straightened. The crosssections were transferred on silanized slides and dried completely.Samples were then dewaxed and rehydrated following the conversesteps. After a blocking incubation with BSA (2% in PBS) to reducenon-specific binding, the primary antibody solution (a rabbit anti-AM polyclonal antibody 1:200, or a rabbit anti-GST antibody 1:500)was gently poured on the microscope slides. After an overnightincubation at 4�C, the sections were washed with PBS (three timesfor 10 min each) and incubated in the secondary antibody [anti-rabbit IgG F(ab¢)2 fragment conjugated to the Alexa Fluor 488fluorochrome (Invitrogen)] for 1 h 30 min at room temperature(25�C) in the darkness. The secondary antibody was then removedand after washing in PBS, a 2-lM 4¢,6-diamidino-2-phenylindole(DAPI) solution (Sigma-Aldrich, http://www.sigmaaldrich.com) wasused to specifically stain the nuclei. The sections were mounted inMowiol anti-fading medium.
Immunocytolocalization of AM and GST in HRs. Cross sec-tions (50 lm) were obtained from fresh HRs using a vibratome(MICROM HM650V; Thermo Scientific, http://www.thermo.com),and lightly fixed for 1 h in paraformaldehyde 4% in 10 mM PBS insmall glass dishes. After washing with PBS, the sections were pre-treated with a BSA solution (3%) for h to reduce non-specific bind-ing. The sections were incubated in the polyclonal antibodiessolutions (a rabbit anti-AM polyclonal antibody 1:200 or a rabbitanti-GST antibody 1:500) overnight at 4�C. After washing with PBS,the secondary antibody Alexa488-conjugate was applied for 1 h(1:400) in the darkness. Then, after washing with PBS, sections weremounted in Mowiol antifading medium on fluoro-slides with twocircles (ESCO, http://www.escocorp.com). Controls were included inorder to show the specificity of the signal, some sections weretreated with neither anti-AM (or anti-GST) nor secondary antibody(autofluorescence), and others were treated with the secondaryantibody only.
Light microscopy and laser scanning confocal microscopy
Cross sections (50 lm) were obtained from fresh HRs using avibratome (MICROM HM650V; Thermo Scientific), dipped in PBSand directly observed on a Leica DM6000 wide-field microscopewith a PL APO objective 40 · (NA: 1.25). The pictures and videos(10 frames s)1; Video Clips S1–S4) were obtained with VOLOCITY-
ACQUISITION 5.1.0 (Perkin Elmer, http://las.perkinelmer.com).The sections were dipped in Nile red solution (2 lg ml)1) for
10 min in the dark, and were then observed after washing with LeicaDM 6000 (transmitted light and epifluorescence; filter I3 and TX,excitation 450–490 and 540–580 nm, emission 515–800 and 608–682 nm, respectively) to visualize the neutral or membrane lipids inyellow or red, respectively.
The confocal microscope imaging was performed on the Mont-pellier RIO Imaging Platform (http://www.mri.cnrs.fr), with a confo-cal laser scanning Axiovert 200M 510 META microscope (Zeiss,http://www.zeiss.com). Excitation was provided by a 405-nm laser
968 Camila Gomez et al.
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), 67, 960–970

for the DAPI stain (bandpass 422–476 nm), a helium-neon 543-nmlaser for the autofluorescence of anthocyanins (long pass 585 nm)and a 488-nm Argon laser for the Alexa 488 and GFP (bandpass 505–530 nm), using multitracking mode (fast line-wise switchingbetween excitation laser lines) to avoid the simultaneous excitationof, and hence emission from, the fluorophores (crosstalk).
The sections were observed with Plan Neofluar 25 · /0.8 or C-Apochromat 40 · /1.2 Zeiss objectives. Pictures were processedusing the Zeiss LSM IMAGE BROWSER software and PHOTOSHOP CS3(Adobe, http://www.adobe.com). Three-dimensional pictures wereobtained with VOLOCITY VISUALIZATION 5.0.2 (Improvision; a PerkinElmer Company). The 3D video was created from Z-stack sectionsfrom the confocal microscope with the VOLOCITY VISUALIZATION usingvarious rotations of 3D rendering. The video 4D (time lapse) wasobtained with a Cell Observer SD Zeiss (multidimensional fastconfocal image acquisition or spinning disk) with a 532-nm laser.
Transmission electron microscopy (TEM)
Mature grape berry samples (epicarp) were immersed in 2.5% glu-taraldehyde in Sorensen’s buffer (0.1 M, pH 7.4) overnight at 4�C,and then rinsed in Sorensen’s buffer and post-fixed in 0.5% osmicacid for 2 h in the dark and at room temperature. After two bufferrinses, tissues were dehydrated in a graded ethanol series (30–100%). Tissues were then embedded in EmBed 812 resin using anAutomated Microwave Tissue Processor for Electronic Microscopy(Leica EM AMW). Ultra-thin sections (80 nm; Leica-Reichert Ultra-cut E) were collected at different levels of each block, and counter-stained with uranyl acetate. Observation and photography wereperformed using a Hitachi 7100 transmission electron microscope(http://www.hitachi.com) in the Centre de Ressources en ImagerieCellulaire de Montpellier, France.
ACKNOWLEDGEMENTS
The authors especially thank M. Lartaud and E. Jublanc, PHIV,Montpellier Rio Imaging, IFR 127 DAPHNE and E. Jullien from Zeissfor their helpful assistance with confocal and spinning-discmicroscopy. The authors are also grateful to C. Cazevieille andC. Sanchez for their technical assistance and interpreting dataconcerning ultrastructural evaluation. The authors thank N. Paris forproviding GFP-HDEL and C. Tesniere for providing the berries to beused in the immunocytolocalization assays, and I. Gil and G. Lopezfor their assistance in the handling of the in vitro plant material. Thiswork was supported by the European Community (STREP projectFLAVO–FOOD–CT–2004–513960) and by INRA.
SUPPORTING INFORMATION
Additional supporting information may be found in the online ver-sion of this article:Figure S1. Immunofluorescence and GFP controls by confocalmicroscopy.Figure S2. anthoMATE transporters (AMs) and glutathione-S-trans-ferase (GST) subcellular localization in hairy roots not expressingMybA1.Video Clip S1. Bright-field microscopy observation of a pigmentedcell from control hairy roots, showing small vesicles movingactively within the cytoplasm, and large vesicles located withinthe vacuole but remaining motionless. During acquisition, the focuswas successively adjusted to the cell wall, the cytoplasm and themiddle of the vacuole.Video Clip S2. Four-dimensional acquisition using spinning-discconfocal microscopy (Cell Observer SD Zeiss) of an epidermal cell ofcontrol hairy roots from Z-stack optical sections over a period of
45 min. The red autofluorescence of anthocyanins is obtained witha 532-nm laser.Video Clip S3. Three-dimensional rendering of an epidermal cell inHR-AM1GFP from Z-stack optical sections, made with VOLOCITY
VISUALIZATION software. The video shows a rotation of this 3Dreconstruction for a better visualization of the specific green signalof GFP in the cellular volume. The red autofluorescence of antho-cyanins and the green fluorescence of the AM1-GFP fusion are seenusing laser multitracking at 543 and 488 nm, respectively, on theconfocal microscope.Video Clip S4. Three-dimensional rendering of an epidermal cell inHR-GSTGFP from Z-stack optical sections, made with VOLOCITY
VISUALIZATION software. The video shows a rotation of this 3Dreconstruction for a better visualization of the specific green signalof GFP in the cellular volume. The red autofluorescence of antho-cyanins and the green fluorescence of the GST-GFP fusion are seentogether with the DAPI-stained nucleus using laser multitracking at405, 543 and 488 nm, on the confocal microscope.Please note: As a service to our authors and readers, this journalprovides supporting information supplied by the authors. Suchmaterials are peer-reviewed and may be re-organized for onlinedelivery, but are not copy-edited or typeset. Technical supportissues arising from supporting information (other than missingfiles) should be addressed to the authors.
REFERENCES
Ageorges, A., Fernandez, L., Vialet, S., Merdinoglu, D., Terrier, N. and
Romieu, C. (2006) Four specific isogenes of the anthocyanin metabolic
pathway are systematically co-expressed with the red colour of grape
berries. Plant Sci. 170, 372–383.
Alfenito, M.R., Souer, E., Goodman, C.D., Buell, R., Mol, J., Koes, R. and
Walbot, V. (1998) Functional complementation of anthocyanin sequestra-
tion in the vacuole by widely divergent glutathione S-transferases. Plant
Cell, 10, 1135–1149.
Batoko, H., Zheng, H.Q., Hawes, C. and Moore, I. (2000) A Rab1 GTPase is
required for transport between the Endoplasmic Reticulum and Golgi
apparatus and for normal Golgi movement in plants. Plant Cell, 12, 2201–
2217.
Butelli, E., Titta, L., Giorgio, M. et al. (2008) Enrichment of tomato fruit with
health-promoting anthocyanins by expression of select transcription fac-
tors. Nat. Biotechnol. 26, 1301–1308.
Conn, S., Zhang, W. and Franco, C. (2003) Anthocyanic vacuolar inclusions
(AVIs) selectively bind acylated anthocyanins in Vitis vinifera L. (grapevine)
suspension culture. Biotech. Letters, 25, 835–839.
Conn, S., Curtin, C., Bezier, A., Franco, C. and Zhang, W. (2008) Purification,
molecular cloning, and characterization of glutathione S-transferases
(GSTs) from pigmented Vitis vinifera L. cell suspension cultures as putative
anthocyanin transport proteins. J. Exp. Bot. 59, 3621–3634.
Conn, S., Franco, C. and Zhang, W. (2010) Characterization of anthocyanic
vacuolar inclusions in Vitis vinifera L. cell suspension cultures. Planta, 231,
1343–1360.
Cutanda-Perez, M.C., Ageorges, A., Gomez, C., Vialet, S., Terrier, N., Romieu,
C. and Torregrosa, L. (2009) Ectopic expression of VlmybA1 in grapevine
activates a narrow set of genes involved in anthocyanin synthesis and
transport. Plant Mol. Biol. 69, 633–648.
Field, B., Jordan, F. and Osbourn, A. (2006) First encounters – deployment of
defence-related natural products by plants. New Phytol. 172, 193–
207.
Gomez, C., Terrier, N., Torregrosa, L. et al. (2009) Grapevine MATE-Type
proteins act as vacuolar H+-dependent acylated anthocyanin transporters.
Plant Physiol. 150, 402–415.
Goodman, C.D., Casati, P. and Walbot, V. (2004) A multidrug resistance-
associated protein involved in anthocyanin transport in Zeamays. Plant
Cell, 16, 1812–1826.
Graf, R., Rietdorf, J. and Zimmermann, T. (2005) Live Cell Spinning Disk
Microscopy. In Microscopy Techniques (Rietdorf, J., ed.). Adv. Biochem.
Engin. Biotechnol. 95, 57–75.
Anthocyanin trafficking in grapevine 969
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), 67, 960–970

Greenspan, P., Mayer, E.P. and Fowler, S.D. (1985) Nile Red – A selective
fluorescent stain for inreacellular lipid droplets. J. Cell Biol. 100, 695–973.
Grotewold, E. (2004) The challenges of moving chemicals within and out of
cells: insights into the transport of plant natural products. Planta, 219, 906–
909.
Grotewold, E. and Davies, K. (2008) Trafficking and sequestration of antho-
cyanins. Nat. Prod. Comm. 3, 1251–1258.
Grotewold, E., Chamberlin, M., Snook, M., Siame, B., Butler, L., Swenson, J.,
Maddock, S., St-Clair, G. and Bowen, B. (1998) Engineering secondary
metabolism in maize cells by ectopic expression of transcription factors.
Plant Cell, 10, 721–740.
Harborne, J.B. and Williams, C.A. (2000) Advances in flavonoid research since
1992. Phytochemistry, 55, 582–596.
Haseloff, J., Siemering, K.R., Prasher, D.C. and Hodge, S. (1997) Removal of a
cryptic intron and subcellular localization of green fluorescent protein are
required to mark transgenic Arabidopsis plants brightly. Proc. Natl Acad.
Sci. USA, 94, 2122–2127.
Hsieh, K. and Huang, A.H.C. (2007) Tapetosomes in Brassica tapetum
accumulate endoplasmic reticulum-derived flavonoids and alkanes for
delivery to the pollen surface. Plant Cell, 19, 582–596.
Irani, N.G. and Grotewold, E. (2005) Light-induced morphological alteration in
anthocyanin-accumulating vacuoles of maize cells. BMC Plant Biol. 5, 7.
Jasik, J. and Vancova, B. (1992) Cytological study of anthocyanin production
in grapevine (Vitis vinifera L.) callus cultures. Acta Bot. Hung. 37, 251–259.
Karimi, M., Inze, D. and Depicker, A. (2002) GATEWAY vectors for Agrobac-
terium-mediated plant transformation. Trends Plant Sci. 7, 193–195.
Kitamura, S. (2006) Transport of flavonoids: from cytosolic synthesis to vac-
uolar accumulation. In The Science of Flavonoids (Grotewold, E., ed.). New
York: Springer Science, LLC, pp. 123–146.
Kitamura, S., Shikazono, N. and Tanaka, A. (2004) TRANSPARENT TESTA 19
is involved in the accumulation of both anthocyanins and proanthocyani-
dins in Arabidopsis. Plant J. 37, 104–114.
Kitamura, S., Matsuda, F., Tohge, T., Yonekura-Sakakibara, K., Yamazaki, M.,
Saito, K. and Narumi, I. (2010) Metabolic profiling and cytological analysis
of proanthocyanidins in immature seeds of Arabidopsis thaliana flavonoid
accumulation mutants. Plant J. 62, 549–559.
Lin, Y., Irani, N. and Grotewold, E. (2003) Sub-cellular trafficking of phyto-
chemicals explored using auto-fluorescent compounds in maize cells. BMC
Plant Biol. 3, 10.
Livak, K.J. and Schmittgen, T.D. (2001) Analysis of relative gene expression
data using real-time quantitative PCR and the 2)DCT method. Methods, 25,
402–408.
Luo, J., Nishiyama, Y., Fuell, C. et al. (2007) Convergent evolution in the BAHD
family of acyl transferases: identification and characterization of antho-
cyanin acyl transferases from Arabidopsis thaliana. Plant J. 50, 678–695.
Marinova, K., Poucel, L., Weder, B., Schwarz, M., Barron, D., Routaboul, J.M.,
Debeaujon, I. and Klein, M. (2007) The Arabidopsis MATE transporter TT12
acts as a vacuolar flavonoid/H+-antiporter active in proanthocyanidin
accumulating cells of the seed coat. Plant Cell, 19, 2023–2038.
Markham, K.R., Gould, K.S., Winefield, C.S., Mitchell, K.A., Bloor, S.J. and
Boase, M.R. (2000) Anthocyanic vacuolar inclusions – their nature and
significance in flower colouration. Phytochemistry, 55, 327–336.
Marrs, K.A., Alfenito, M.R., Lloyd, A.M. and Walbot, V. (1995) A Glutathione-S-
Transferase Involved in Vacuolar Transfer Encoded by the Maize Gene
Bronze-2. Nature, 375, 397–400.
Mathews, H., Clendennen, S.K., Caldwell, C.G. et al. (2003) Activation tagging
in tomato identifies a transcriptional regulator of anthocyanin biosynthe-
sis, modification, and transport. Plant Cell, 15, 1689–1703.
McLusky, S.R., Bennett, M.H., Beale, M.H., Lewis, M.J., Gaskin, P. and
Mansfield, J.W. (1999) Cell wall alterations and localized accumulation of
feruloyl-3¢-methoxytyramine in onion epidermis at sites of attempted
penetration by Botrytis allii are associated with actin polarisation, peroxi-
dase activity and suppression of flavonoid biosynthesis. Plant J. 17, 523–
534.
Mueller, L.A. and Walbot, V. (2001) Models for vacuolar sequestration of
anthocyanins. In Regulation of Phytochemicals by Molecular Techniques
(Romeo, J.T., Saunders, J.A. and Matthews, J.F., eds). New York: Perg-
amon, pp. 297–312.
Mueller, L.A., Goodman, C.D., Silady, R.A. and Walbot, V. (2000) AN9, a
petunia glutathione S-transferase required for anthocyanin sequestration,
is a flavonoid-binding protein. Plant Physiol. 123, 1561–1570.
Neuhaus, J.M. and Paris, N. (2005) Plant Vacuoles: from Biogenesis to Func-
tion. In Plant Endocytosis (Samaj, J., Baluska, F. and Menzel, D., eds).
Berlin: Springer-Verlag, pp. 65–82.
Nielsen, K.A., Gotfredsen, C.H., Buch-Pedersen, M.J., Ammitzboll, H.,
Mattsson, O., Duus, J.O. and Nicholson, R.L. (2004) Inclusions of flavonoid
3-deoxyanthocyanidins in Sorghum bicolor self-organize into spherical
vesicles. Physiol. Mol. Plant Pathol. 65, 187–196.
Nozue, M., Kubo, H., Nishimura, M., Katou, A., Hattori, C., Usuda, N., Nagata,
T. and Yasuda, H. (1993) Characterization of intravacuolar pigmented
structures in anthocyanin-containing cells of sweet potato suspension
cultures. Plant Cell Physiol. 34, 803–808.
Paciorek, T., Sauer, M., Balla, J., Wisniewska, J. and Friml, J. (2006) Immu-
nocytochemical technique for protein localization in sections of plant tis-
sues. Nat. Protoc. 1, 104–107.
Pecket, R.G. and Small, C.J. (1980) Occurrence, location and development of
anthocyanoplasts. Phytochemistry, 19, 2571–2576.
Pourcel, L., Irani, N.G., Lu, Y.H., Riedl, K., Schwartz, S. and Grotewold, E.
(2010) The formation of Anthocyanic Vacuolar Inclusions in Arabidopsis
thaliana and implications for the sequestration of anthocyanin pigments.
Mol. Plant, 3, 78–90.
Poustka, F., Irani, N.G., Feller, A., Lu, Y., Pourcel, L., Frame, K. and Grotewold,
E. (2007) A trafficking pathway for anthocyanins overlaps with the endo-
plasmic reticulum-to-vacuole protein-sorting route in Arabidopsis and
contributes to the formation of vacuolar inclusions. Plant Physiol. 145,
1323–1335.
Small, C.J. and Pecket, R.G. (1982) The ultrastructure of anthocyanoplasts in
red cabagge. Planta, 154, 97–99.
Snyder, B.A. and Nicholson, R.L. (1990) Synthesis of phytoalexins in sorghum
as a site-specific response to fungal ingress. Science, 248, 1637–1639.
Tanaka, Y., Sasaki, N. and Ohmiya, A. (2008) Biosynthesis of plant pigments:
anthocyanins, betalains and carotenoids. Plant J. 54, 733–749.
Terrier, N., Torregrosa, L., Ageorges, A., Vialet, S., Verries, C., Cheynier, V.
and Romieu, C. (2009) Ectopic expression of VvMybPA2 promotes
proanthocyanidin biosynthesis in grapevine and suggests additional
targets in the pathway. Plant Physiol. 149, 1028–1041.
Tohge, T., Nishiyama, Y., Hirai, M.Y. et al. (2005) Functional genomics by
integrated analysis of metabolome and transcriptome of Arabidopsis
plants over-expressing an MYB transcription factor. Plant J. 42, 218–235.
Torregrosa, L. and Bouquet, A. (1997) Agrobacterium rhizogenes and
A. tumefaciens co-transformation to obtain grapevine hairy roots produc-
ing the coat protein of grapevine chrome mosaic nepovirus. Plant Cell
Tissue Organ Cult. 49, 53–62.
Winkel, B.S.J. (2004) Metabolic channeling in plants. Annu. Rev. Plant Biol. 55,
85–107.
Winkel-Shirley, B. (1999) Evidence for enzyme complexes in the phenyl-
propanoid and flavonoid pathways. Physiol. Plant. 107, 142–149.
Winkel-Shirley, B. (2001) Flavonoid Biosynthesis. A colorful model for
genetics, biochemistry, cell biology, and biotechnology. Plant Physiol. 126,
485–493.
Winkel-Shirley, B. (2002) Biosynthesis of flavonoids and effects of stress. Curr.
Opin. Plant Biol. 5, 218–223.
Zhang, H., Wang, L., Deroles, S., Bennett, R. and Davies, K. (2006) New insight
into the vesicles and formation of anthocyanic vacuolar inclusions in flower
petals. BMC Plant Biol. 6, 29.
Zhao, J. and Dixon, R. (2009) The ‘ins’ and ‘outs’ of flavonoid transport. Trends
Plant Sci. 15, 72–80.
970 Camila Gomez et al.
ª 2011 The AuthorsThe Plant Journal ª 2011 Blackwell Publishing Ltd, The Plant Journal, (2011), 67, 960–970




















