
Identifying advanced glycation end products as a major source of oxidants in aging: implications for...
10
Identifying Advanced Glycation End Products as a Major Source of Oxidants in Aging: Implications for the Management and/or Prevention of Reduced Renal Function in Elderly Persons Helen Vlassara, MD,* Jaime Uribarri, MD, ‡ Luigi Ferrucci, MD, § Weijing Cai, MD,* Massimo Torreggiani, MD,* , James B. Post, MD, ‡ Feng Zheng, MD, PhD,* and Gary E. Striker, MD* ,‡ Summary: Aging is characterized by increasing inflammation and oxidant stress (OS). Reduced renal function was present in more than 20% of normal-aged individuals sampled in the National Health and Nutrition Examination Survey (NHANES) cross-sectional study of the US population. Longitudinal studies in the United States and Italy showed that renal function does not decline in some individuals, suggesting that a search for causes of the loss of renal function in some persons might be indicated and interventions to reduce this outcome should be sought. Because advanced glycation end products (AGEs) induce both inflammation and OS, accumulate with age, and primarily are excreted by the kidney, one outcome of reduced renal function in aging could be decreased AGE disposal. The build-up of AGEs with reduced renal function could contribute to inflammation, increased oxidant stress, and accumulation of AGEs in aging. In fact, results from a longitudinal study of normal aging adults in Italy showed that the most significant correlation with mortality was the level of renal function. A clear link between inflammation, OS, AGEs, and chronic disease was shown in studies of mice that showed that reduction of AGE levels by drugs or decreased intake of AGEs reduces chronic kidney disease (CKD) and cardiovascular disease of aging. The data support a role for AGEs in the development of renal lesions in aging mice and reveal that AGEs in the diet are very important contributors to renal and cardiovascular lesions. AGEs signal through two receptors, one of which is anti-inflammatory (AGER1) and the other is proinflammatory (RAGE). Overexpression of AGER1 protects against OS and acute vascular injury. The reduction of AGEs in the diet is as efficient in preventing aging-related cardiovascular and renal lesions in mice as that seen with calorie restriction. Studies in normal adults of all ages and those with CKD suggest that the findings in mice may be directly applicable to both aging and CKD. Namely, the dietary content of AGEs determines the serum levels of AGEs and inflammatory mediators and urine AGE levels in both normal subjects and CKD patients. Importantly, reduction of AGEs controls these changes in both normal subjects and CKD patients, and the phenotypic changes in AGER1 are reduced in CKD patients by decreasing the amount of AGEs consumed with the diet. These data suggest that the changes in renal function in normal aging may be subject to control and this subject deserves renewed attention. Semin Nephrol 29:594-603 © 2009 Published by Elsevier Inc. Keywords: AGEs, aging, diet, CKD T wenty million adults in the United States (age, 20 y) have an increased serum cre- atinine level and the incidence increases with age according to a large cross-sectional study (National Health and Nutrition Examination Sur- vey [NHANES III]). 1 More than 20% of those age 65 years or older have decreased renal function, even when corrected for obvious causes of renal diseases such as hypertension and diabetes melli- *Division of Diabetes and Aging, Department of Geriatrics, Mount Sinai School of Medicine, New York, NY. ‡Division of Nephrology, Department of Medicine, Mount Sinai School of Medicine, New York, NY. §Longitudinal Studies Section, Clinical Research Branch, National Institute on Aging, National Institutes of Health, Baltimore, MD. Unit of Nephrology, Dialysis and Transplantation, University of Pavia, Pavia, Italy. Address reprint requests to Helen Vlassara, MD, Mount Sinai School of Medicine, Box 1640, One Gustave Levy Pl, New York, NY 10029. E-mail: [email protected] 0270-9295/09/$ - see front matter © 2009 Published by Elsevier Inc. doi:10.1016/j.semnephrol.2009.07.013 Seminars in Nephrology, Vol 29, No 6, November 2009, pp 594-603 594
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Identifying advanced glycation end products as a major source of oxidants in aging: implications for...

*
‡
§
�A
0©
5
Identifying Advanced Glycation EndProducts as a Major Source of Oxidants inAging: Implications for the Management
and/or Prevention of Reduced RenalFunction in Elderly Persons
Helen Vlassara, MD,* Jaime Uribarri, MD,‡ Luigi Ferrucci, MD,§ Weijing Cai, MD,*Massimo Torreggiani, MD,*,� James B. Post, MD,‡ Feng Zheng, MD, PhD,* and
Gary E. Striker, MD*,‡
Summary: Aging is characterized by increasing inflammation and oxidant stress (OS). Reduced renalfunction was present in more than 20% of normal-aged individuals sampled in the National Health andNutrition Examination Survey (NHANES) cross-sectional study of the US population. Longitudinalstudies in the United States and Italy showed that renal function does not decline in some individuals,suggesting that a search for causes of the loss of renal function in some persons might be indicated andinterventions to reduce this outcome should be sought. Because advanced glycation end products(AGEs) induce both inflammation and OS, accumulate with age, and primarily are excreted by thekidney, one outcome of reduced renal function in aging could be decreased AGE disposal. Thebuild-up of AGEs with reduced renal function could contribute to inflammation, increased oxidantstress, and accumulation of AGEs in aging. In fact, results from a longitudinal study of normal agingadults in Italy showed that the most significant correlation with mortality was the level of renalfunction. A clear link between inflammation, OS, AGEs, and chronic disease was shown in studies ofmice that showed that reduction of AGE levels by drugs or decreased intake of AGEs reduces chronickidney disease (CKD) and cardiovascular disease of aging. The data support a role for AGEs in thedevelopment of renal lesions in aging mice and reveal that AGEs in the diet are very importantcontributors to renal and cardiovascular lesions. AGEs signal through two receptors, one of which isanti-inflammatory (AGER1) and the other is proinflammatory (RAGE). Overexpression of AGER1protects against OS and acute vascular injury. The reduction of AGEs in the diet is as efficient inpreventing aging-related cardiovascular and renal lesions in mice as that seen with calorie restriction.Studies in normal adults of all ages and those with CKD suggest that the findings in mice may bedirectly applicable to both aging and CKD. Namely, the dietary content of AGEs determines the serumlevels of AGEs and inflammatory mediators and urine AGE levels in both normal subjects and CKDpatients. Importantly, reduction of AGEs controls these changes in both normal subjects and CKDpatients, and the phenotypic changes in AGER1 are reduced in CKD patients by decreasing the amountof AGEs consumed with the diet. These data suggest that the changes in renal function in normal agingmay be subject to control and this subject deserves renewed attention.Semin Nephrol 29:594-603 © 2009 Published by Elsevier Inc.
Keywords: AGEs, aging, diet, CKDTw(v6e
Division of Diabetes and Aging, Department of Geriatrics, Mount SinaiSchool of Medicine, New York, NY.
Division of Nephrology, Department of Medicine, Mount Sinai School ofMedicine, New York, NY.
Longitudinal Studies Section, Clinical Research Branch, National Institute onAging, National Institutes of Health, Baltimore, MD.
Unit of Nephrology, Dialysis and Transplantation, University of Pavia, Pavia, Italy.ddress reprint requests to Helen Vlassara, MD, Mount Sinai School ofMedicine, Box 1640, One Gustave Levy Pl, New York, NY 10029. E-mail:[email protected]
270-9295/09/$ - see front matter
d2009 Published by Elsevier Inc. doi:10.1016/j.semnephrol.2009.07.013Seminars in N94
wenty million adults in the United States(age, �20 y) have an increased serum cre-atinine level and the incidence increases
ith age according to a large cross-sectional studyNational Health and Nutrition Examination Sur-ey [NHANES III]).1 More than 20% of those age5 years or older have decreased renal function,ven when corrected for obvious causes of renal
iseases such as hypertension and diabetes melli-ephrology, Vol 29, No 6, November 2009, pp 594-603

toHwsOfmrbwtobtAvdaaitttermstt
A
TwbtiooApnaaptryci
sgw(a
npsgimGfqzDbtagmmiciaamdwtcrgdCrAfica
A
Aawb
AGEs as a source of oxidants in aging 595
us. Thus, de novo chronic kidney disease (CKD)f aging represents an important health problem.ypertension and diabetes mellitus, both ofhich are associated with increased oxidative
tress (OS), and intercurrent events that increaseS carry a further increase in the risk of renal
ailure.1 This suggests that the aging kidney isore susceptible to injury and/or has an altered
epair response.2 There is a strong associationetween CKD and cardiovascular disease, both ofhich have been associated with an inflamma-
ory state.3-11 These data are consistent with ourbservations in experimental animals and humaneings, that the presence of adequate renal func-ion is critical to maintaining the body load ofGEs, at nontoxic levels.12 Namely, serum ad-anced glycation end products (AGEs) correlateirectly with the levels of inflammatory markersnd OS, and inversely with the clearance of cre-tinine.13 CKD in aging also is associated withncreased levels of interleukin-6, C-reactive pro-ein, and other markers of inflammation.14 Thus,he CKD of aging is associated with an inflamma-ory state, which may be associated with the gen-ral increase in OS noted with aging. CKD thatesembles that seen in human beings occurs inultiple mouse strains and in rats and con-
ists of glomerulosclerosis and hypertrophy,ubulointerstitial changes, chronic inflamma-ion, and vascular lesions.15,16
GEs
hese compounds are generated endogenouslyhen reducing sugars, ascorbate, and other car-ohydrates spontaneously react with amino-pep-ides, nucleic acids, and amine-containing lip-ds.17,18 The diet is an underappreciated sourcef AGEs.19,20 This exogenous source consistsf foods cooked at increased temperatures.21
mine-containing lipids form advanced li-oxidation end products such as 4-hydroxy-onenal, or �N-carboxymethyllysine (CML)nd their analogues, deriving from both lipidnd protein oxidation.22,23 Glycoxidation ofroteins and lipids (AGEs/advanced lipoxida-ion end products) involves the formation ofeactive carbonyls that generate reactive ox-gen species (ROS) and increased OS, termedarbonyl stress.24,25 Autoxidation of glucose
s accompanied by the generation of ROS auch as superoxide radicals.24 ROS enhancelycation, and both mechanisms can interfereith the normal clearance of macromolecules
ie, low-density lipoprotein), from circulationnd promote atherogenesis.22,23,26
The term AGEs, although often referring toonreactive terminal products, such as CML orentosidine, also includes reactive precursors,uch as methylglyoxal, glyoxal, and 1- or 3-deoxy-lucosone. Methylglyoxal constantly is producedn vivo by glucose auto-oxidation and inter-
ediates of protein and lipid metabolism.lyoxalase I is a major methylglyoxal detoxi-
ying enzyme. The action of glyoxalase I re-uires glutathione. Glutathione reacts nonen-ymatically with methylglyoxal to form S--lactoylglutathione hemithioacetal. Glyoxalase Ireaks hemithioacetal adduct to S-D-lactoylgluta-hioneion. Methylglyoxal-derived AGEs (MG)re one of the major groups of AGEs. MG isenerated primarily from the interaction ofethylglyoxal with arginine residues. MG isore active because of its dynamic reversibil-
ty. The levels of MG decrease when the con-entration of methylglyoxal decreases. AGEsncluding MG levels are increased in diabetesnd aging.27,28 The cause of increased AGEs inging is not clear. Because the kidney is theajor site for the excretion of AGEs, a mild
ecrease in renal function may be associatedith increased levels of AGEs. The accumula-
ion of AGEs with the slowly progressive de-rease in renal function in aging may induce aelease of inflammatory mediators and theeneration of ROS before a clinically evidentecrease in kidney function becomes evident.12,29
hronic inflammation and oxidative stress areisk factors for kidney aging. The significance ofGE accumulation in aging is established by thending that AGE inhibitors conferred signifi-ant protection against cardiovascular diseasend CKD in aging.30-32
GE LEVELS AND RENAL FUNCTION
GEs are excreted by the kidney, and their levelsre increased in patients with renal insufficiency,ith or without diabetes.33,34 Although it has longeen known that the levels of circulating AGEs
re increased in patients with already developed
rcdpntttbpdctkoiTpr
A
AfltSmosispatAhaRRdsasrbodtad
S
CK
G2tdbfaAtffcflpaeramktrtttcct
DA
Wcbtescrtibtt
596 H. Vlassara et al
enal failure, it now is clear that AGEs are in-reased before there is clinical evidence of re-uced renal function13,19,29 in both the normalopulation and in diabetic patients.12,20 The kid-ey is both the only major site for excretion oferminal end products of AGE metabolism and aarget for reactive early AGE intermediates. Reac-ive intermediates attached to small peptides maye filtered by the glomerulus and taken up in theroximal tubules from the filtrate or they may beirectly taken out of the blood in the peritubularapillaries. The proximal tubule may further me-abolize AGEs to nonreactive products. Thus, theidney is one of the key regulators of AGE metab-lism in aging. However, the ingested AGEs may
nduce an inflammatory response in the tubules.his being the case, it is critical to understand theathways by which AGEs mediate inflammatoryesponses in the kidney.
GEs AND AGE-RELATED RECEPTORS
GEs bind to cellular AGE-receptors inducing in-ammation and ROS.35,36 The AGE receptor sys-em is complex, and includes RAGE, AGER1-3,cR-II, CD36, and defensins.37 Some receptors re-ove excess AGE from the body (AGER1) and
thers increase OS and activate inflammatory re-ponses (eg, RAGE).38,39 Those that activate annflammatory response have been studied exten-ively, and are best represented by a multiligandrotein, RAGE, which is linked to OS and cellctivation.37,40 Although insight has been ob-ained from in vitro molecular studies on certainGE receptors, clarification of their in vivo roleas been difficult. For instance, reactive AGEsffect cells and cellular AGE receptors alike;AGE, the receptor most closely associated withOS and nuclear factor-kB activation, also is in-ucible by AGE and ROS.38 Despite high expres-ion/function of RAGE in diabetes, renal disease,nd aging, tissue AGE deposits remain increased,uggesting that it does not participate in AGEemoval. AGER1, which clears AGEs, appears toe suppressed or down-regulated in the presencef excess AGE-induced oxidant stress (ie, severeiabetic complications and aging).28,41 We foundhat AGER1 could have a protective functiongainst OS because it may inhibit AGE-depen-
ent cellular activation. gTUDIES IN MICE
haracterization of theidney Lesions in Aging C57B6 Mice
lomerular changes in C57B6 mice begin at 18 to0 months of age, before recognizable tubuloin-erstitial changes, and progressively increased toeath at 30 months.42-44 These mice develop al-uminuria and decreased renal function. We alsoound that AGEs increase in the kidney withging,28 and that a reduction of the intake ofGEs via the diet blocked the development of
he CKD of aging and prolonged survival. Weound that glomerular mesangial cells isolatedrom aging mice express stable phenotypichanges, including increased production of in-ammatory cytokines and connective tissueroteins, oxidant stress, cellular hypertrophy,nd increased P27 levels.43,44 Because a prolif-rative stimulus accentuates kidney lesions, weeasoned that uninephrectomy would acceler-te the CKD of aging. Uninephrectomy in 20-onth-old C57B6 mice resulted in accelerated
idney lesions (glomerular and tubulointersti-ial), increased albuminuria, and early onsetenal failure.42 The lesions closely resembledhose found in 30-month-old mice, suggestinghe added proliferative stimulus from a reduc-ion in nephron number accelerated the aginghanges and that phenotypically altered re-ruited progenitors could be the source ofhe inflammatory/prosclerotic changes.
etermination of thenatomic Site of the Phenotypic Change
e previously observed that the bone marrowontained mesenchymal progenitors in humaneings and glomerular progenitors in mice,45 andhat the bone marrow contains progenitors thatxpress stable phenotypic changes in glomerulo-clerosis and diabetes. Because the bone marrowontained altered progenitors in other chronicenal diseases, we postulated that the lesions inhe CKD of aging also may originate in a progen-tor population. Therefore, we asked whether theone marrow is a source of kidney progenitorshat carry the inflammatory/prosclerotic pheno-ypic changes of aging. We found the following.
The bone marrow contains glomerular pro-
enitors that express the inflammatory/sclero-
semablmstsTtta
leuataitmtsdm
pripmi((C
Tt
WhippNgg
nsvgspmbaaimg
ACPR
Mpliwrtmibrcertdmsceagemmtgigto
AGEs as a source of oxidants in aging 597
is phenotype. Bone marrow transplants fromarly and late aging mice transfer both the glo-erular and tubulointerstitial lesions found in
ging donors to young naive recipients. Thus,one marrow–derived progenitors mediate at
east some aspects of the aging lesions in C57B6ice.44 The lesions transferred were donor age-
pecific. Young recipients of bone marrowransplants from the oldest donors had moreevere lesions than if the donor was less old.his suggests that the number of progenitors in
he bone marrow, which express stable pheno-ypic changes, increase with age. These aspectspply to aging in both human beings and mice.
Bone marrow transplants from young donorsargely reversed kidney lesions in old recipi-nts. Note that there was a decrease in glomer-lar size, sclerosis, and cellularity, coupled withdecrease in tubulointerstitial lesions. Thus,
he glomerular and tubulointerstitial lesions ofging are reversible in C57B6 mice, and progen-tors derived from the bone marrow mediatehese changes. These data confirm that bonearrow–derived progenitors may play an ac-
ive role in the CKD of aging.44 The data alsouggest that delivery of normal progenitors re-uces the CKD of aging, in an aging environ-ent.Stable phenotypic changes in kidney-specific
rogenitors occur in a number of chronic scle-osing/inflammatory conditions, and are not lim-ted to a specific genetic background. Transfer ofrogressive sclerosing kidney lesions by bonearrow–derived progenitors also was found
n a radiation-induced mutant mouse strainROP�/�) with progressive glomerulosclerosisG2) and in a model of diabetic nephropathy57B6/db�/�.45,46
he Molecular Basis ofhe Stable Phenotypic Change
e found that mesangial cells from aging miceave a stably altered phenotype: mesangial cells
solated from 28-month-old mice show stablehenotypic changes.42 In addition, the in vitrohenotype and in vivo phenotype are correlated.amely, the stable phenotypic changes in mesan-ial cells in vitro reflect the in vivo sclerotic
lomerular changes.47,48 The stable aging phe- Cotype is proinflammatory and prosclerotic, ashown by gene expression analysis that re-ealed that the stable phenotypic changes inlomerular cells were proinflammatory and pro-clerotic.43 Although not applicable to aging,er se, stable epigenetic changes in the matrixetalloproteinase-9 promoter could be induced
y chronic hyperglycemia in vitro, a conditionssociated with increased formation of AGEsnd increased OS.48,49 We found that the revers-bility of the phenotypic changes in the matrix
etalloproteinase-9 promoter depended on theenetic background.49
melioration of Phenotypichanges in Mesangial Cells byeroxisome Proliferator Activatedeceptor-�, Pyridoxamine, and Estrogen
ice with type 2 diabetes and established ne-hropathy were treated with peroxisome pro-
iferator activated receptor-� agonists, whichmproves glucose use, or with pyridoxamine,
hich reduces AGE formation.50-52 Both drugseduced renal lesions and improved renal func-ion and favorably affected matrix turnover byesangial cells in vitro. These data provide ev-
dence that the stable phenotypic changes cane modulated, with resultant stabilization oreversal of established kidney lesions. Mesangialells express both the � and � subtypes of thestrogen receptor and estradiol administrationesults in up-regulation of matrix metallopro-einase-2 and -9.53 Estrogen receptors also areecreased at the transcriptional level in ROP�/�
ice, which are prone to develop glomerulo-clerosis, revealing that glomerulosclerosis inertain mouse strains may be associated withstrogen deficiency, a feature of the CKD inging.42-44,54 The induction of profound estro-en deficiency in ROP�/� mice results in accel-ration of the glomerular lesions.54 In thisodel, we found that the maintenance of nor-al glomerular function depended on the con-
inuous presence of physiologic levels of estro-en, and that prolonged estrogen deficiencynduced stable phenotypic changes in mesan-ial cells.55 Estrogen and progesterone recep-ors and responses to estrogen and progester-ne also were studied in the vascular cells of
3H/HeJ (toll-like receptor [TLR]4-deficient)
mrorlitcfidl
SI
Tdantso
DtAFR
AadaAsa
kmli
DDt
Acdty
matsltcdcdermicfttlmstahTcaipi
S
NC
WNdrgssiloywsm
598 H. Vlassara et al
ice and found that collagen synthesis and deg-adation were not responsive to added estrogenr progesterone, despite the presence of theseeceptors.56 These data also suggest that both theevels and responses to sex hormones may bemportant in the development of, and responseso, stable phenotypic inflammatory/sclerotichanges.55 They also suggest that the TLR4 de-ciency and transforming growth factor-�–me-iated responses may be involved, providing a
ink between AGEs and fibrosis.56
uppression ofnflammation Reduces CKD
he inflammatory aspects of progressive kidneyiseases were prevented by pentosan polysulfate,drug that decreases inflammation in the kid-
ey. Namely, pentosan polysulfate preventedhe development of kidney lesions after five-ixths nephrectomy, a model pertinent to CKDf aging.57
ecreasing AGE Intake Reduceshe CKD of Aging and Decreases AGEccumulation and Transforming Growthactor-�1 and Type IV Collagen MessengerNA Expression in the Aging Kidney
minoguanidine decreases the cardiovascularnd renal disease of aging in rats.30 Becauseietary AGEs are a major source of oxidants inging, we introduced a life-long restriction ofGEs in mice to determine if external AGEserved as a source of the AGE accumulation inging.
Thus, restriction of AGEs in the diet reducesidney lesions in aging. We also found that theyocardial changes of aging are reduced and
ongevity is increased by the low-AGE dietaryntervention (see later).58
ecreasing AGE Intake, Withoutecreasing Calorie Intake, Reduces
he CKD of Aging and Increases Lifespan
s shown earlier the content of AGEs in the dietorrelates with serum AGE levels, OS, organysfunction, and lifespan. We also found thathe addition of a chemically defined AGE (meth-
lglyoxal–bovine serum albumin) to a low-AGE louse chow increased levels of serum AGEsnd OS (�2- to 3-fold). This provides evidencehat dietary AGEs are oxidants that can induceystemic OS, which predisposes to cardiovascu-ar disease and CKD. Calorie restriction (CR) ishe most studied means to decrease OS, in-rease longevity, and reduce OS-related organamage in mammals. We hypothesized that be-ause reduction of food intake obligatorily alsoecreases oxidant AGE intake, the beneficialffects of CR in mammals could be related toestriction of oxidants or energy. We studiedice pair-fed a CR diet or a high-AGE CR diet
n which the dietary content of AGEs was in-reased by brief heat treatment (CR-High). Weound that old mice given CR with a brief heatreatment developed high levels of 8-isopros-anes, AGEs, RAGE, and p66 shc, coupled withow AGER1, low GSH/GSSG, insulin resistance,
arked myocardial and renal fibrosis, and ahortened lifespan.58 Similar to mice main-ained on an AGE-restricted diet, old CR micelso had low OS, p66 shc, RAGE, and AGEs, butigh AGER1, coupled with a longer lifespan.hese data lead us to conclude that the benefi-ial effects of a CR diet may be related partly toreduced oxidant intake, and that a reduced
ntake of oxidant AGEs via the diet is one of therincipal determinants of oxidant status in ag-
ng mice.
TUDIES IN HUMAN BEINGS
ormal Subjects,ross-Sectional Data, New York Subjects
e focused on two distinct serum AGEs. First,�-carboxy-methyl-lysine (sCML), a terminal oxi-ant product, which is less active and likely toeflect longitudinal exposure.59 Second, methyl-lyoxal derivatives (sMG) are more active, lesstable, and readily derived from ROS of otherources. The cross-sectional analyses of approx-mately 300 healthy persons established thatevels of sCML and sMG correlate with eachther, and both are higher in older than inounger persons and in men compared withomen older than age 65.19 Also, the levels of
erum AGEs correlated with circulating inflam-atory markers, such as 8-isoprostanes, vascu-
ar cell adhesion molecule (VCAM)-1, tumor ne-

cA(satacpaTamtnccm
NL
RAysvntTeawpom
NL
TatctnSsijBc
imisohefiaeaartdtttnatiJlfrt
NL
DndecetCmwllrasbe
AGEs as a source of oxidants in aging 599
rosis factor (TNF)-�, and C-reactive protein.fter adjusting for sex, age, and renal clearancebased on estimated glomerular filtration rate),erum AGEs were independent predictors of OSmong healthy persons. A pivotal finding duringhis investigation was that dietary AGE intake,djusted for calorie and nutrient intake, signifi-antly correlated with levels of sCML, sMG, 8-iso-rostanes, VCAM-1, and mononuclear TNF-�,mong other inflammatory markers across age.hese findings suggested that persons habitu-lly consuming high-AGE diets may have abnor-ally high levels of AGEs, which may exceed
he capacity of native defenses to remove oreutralize them. Thus, independent of age,hronic consumption of AGE-rich foods mayause high OS, a common denominator forost chronic diseases.
ormal Subjects,ongitudinal Data, New York Subjects
epeated measures of serum AGEs and dietaryGEs at quarterly intervals over a period of 2ears confirmed the earlier-described relation-hips.19 Serum CML levels (as well as sMG)aried during this period and the changes sig-ificantly correlated with changes in 8-isopros-anes, peripheral mononuclear cell–derivedNF-�, and VCAM-1. Moreover, changes in di-tary AGE intake correlated with those of sCML,s well as of sMG. These findings were in lineith the baseline cross-sectional data and sup-orted the proposed link between exogenousxidant load and systemic oxidant and inflam-ation burden.
ormal Subjects,ongitudinal Data, Baltimore Subjects
he first systematic longitudinal scientific ex-mination of human aging was started in 1958:he Baltimore Longitudinal Study of Aging. Thisohort has remained as one of the most impor-ant sources of information on the aging kid-ey.60 Data from the Baltimore Longitudinaltudy of Aging confirmed that renal functionhows a progressive decline of renal functionn some normal aging adults (based on age-ad-usted standards for creatinine clearance (CrCl).61
ased on CrCl the estimated average annual
hange in CrCl equaled �0.26 mL/min/1.73 m2 in nndividuals aged 20 to 39 years, and �1.51 mL/in/1.73 m2 after age 80. These data are unique
n that the predictions were based on cross-ectional and longitudinal observations andnly reported for individuals consideredealthy based on strict standard criteria. Inter-stingly, although the longitudinal data con-rmed the cross-sectional observations, theylso identified two groups that were not appar-nt in the cross-sectional analysis, one had anccelerated decline of kidney function withge, and another that did not show any loss ofenal function with age. Surprisingly, some Bal-imore Longitudinal Study of Aging participantsid not have a significant decline in renal func-ion over 5 to 10 years. Although the number ofhese individuals was small and few were olderhan 70 years of age, these data challenge theotion that the decline of kidney function withge is unavoidable and call for increased inves-igation in this area. Similar data were reportedn a letter to the editor of the New Englandournal of Medicine by investigators examiningiving kidney donors, in which an extensiveollow-up evaluation failed to show a decline inenal function in this population selected forhe absence of kidney disease.62
ormal Subjects,ongitudinal Data, Italian Subjects
ata from another longitudinal study in Italianormal aging subjects (InCHIANTI study) ad-ressed the question of whether measured andstimated CrCl in older individuals are signifi-ant predictors of mortality.63 They used differ-nt formulae for calculating the glomerular fil-ration rate and found that after stratifying byrCl, patients with CrCl levels of less than 60L/min/1.73 m2 and 60 to 90 mL/min/1.73 m2
ere more likely to die over the 6-year fol-ow-up period compared with those with CrClevels of more than 90 mL/min/1.73 m2 (hazardatio, 1.91; 95% confidence interval, 1.11-3.29;nd hazard ratio, 1.70; 95% CI, 1.02-2.83, re-pectively). They did not find that classificationased on the Modification of Diet in Renal Dis-ase (MDRD) formula provided significant prog-
ostic information for mortality.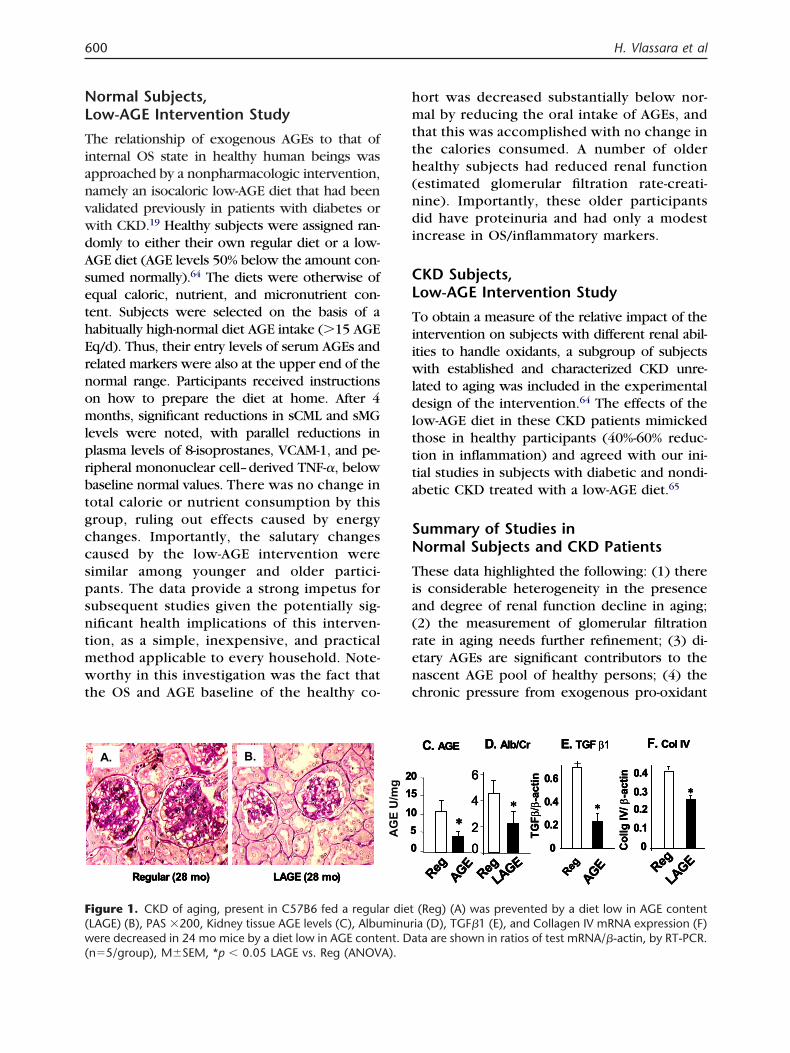
NL
TianvwdAsethErnomlprbtgccspsntmwt
hmtth(ndi
CL
Tiiwldlttta
SN
Tia(renc
F(w(
600 H. Vlassara et al
ormal Subjects,ow-AGE Intervention Study
he relationship of exogenous AGEs to that ofnternal OS state in healthy human beings waspproached by a nonpharmacologic intervention,amely an isocaloric low-AGE diet that had beenalidated previously in patients with diabetes orith CKD.19 Healthy subjects were assigned ran-omly to either their own regular diet or a low-GE diet (AGE levels 50% below the amount con-umed normally).64 The diets were otherwise ofqual caloric, nutrient, and micronutrient con-ent. Subjects were selected on the basis of aabitually high-normal diet AGE intake (�15 AGEq/d). Thus, their entry levels of serum AGEs andelated markers were also at the upper end of theormal range. Participants received instructionsn how to prepare the diet at home. After 4onths, significant reductions in sCML and sMG
evels were noted, with parallel reductions inlasma levels of 8-isoprostanes, VCAM-1, and pe-ipheral mononuclear cell–derived TNF-�, belowaseline normal values. There was no change inotal calorie or nutrient consumption by thisroup, ruling out effects caused by energyhanges. Importantly, the salutary changesaused by the low-AGE intervention wereimilar among younger and older partici-ants. The data provide a strong impetus forubsequent studies given the potentially sig-ificant health implications of this interven-ion, as a simple, inexpensive, and practicalethod applicable to every household. Note-orthy in this investigation was the fact that
he OS and AGE baseline of the healthy co-
A. B.A. B.A. B.
LAGE (28 mo)Regular (28 mo) LAGE (28 mo)Regular (28 mo) LAGE (28 mo)Regular (28 mo)
igure 1. CKD of aging, present in C57B6 fed a regulaLAGE) (B), PAS �200, Kidney tissue AGE levels (C), Albuere decreased in 24 mo mice by a diet low in AGE conte
n�5/group), M�SEM, *p � 0.05 LAGE vs. Reg (ANOVA).
ort was decreased substantially below nor-al by reducing the oral intake of AGEs, and
hat this was accomplished with no change inhe calories consumed. A number of olderealthy subjects had reduced renal functionestimated glomerular filtration rate-creati-ine). Importantly, these older participantsid have proteinuria and had only a modest
ncrease in OS/inflammatory markers.
KD Subjects,ow-AGE Intervention Study
o obtain a measure of the relative impact of thentervention on subjects with different renal abil-ties to handle oxidants, a subgroup of subjects
ith established and characterized CKD unre-ated to aging was included in the experimentalesign of the intervention.64 The effects of the
ow-AGE diet in these CKD patients mimickedhose in healthy participants (40%-60% reduc-ion in inflammation) and agreed with our ini-ial studies in subjects with diabetic and nondi-betic CKD treated with a low-AGE diet.65
ummary of Studies inormal Subjects and CKD Patients
hese data highlighted the following: (1) theres considerable heterogeneity in the presencend degree of renal function decline in aging;2) the measurement of glomerular filtrationate in aging needs further refinement; (3) di-tary AGEs are significant contributors to theascent AGE pool of healthy persons; (4) thehronic pressure from exogenous pro-oxidant
RegAGE
∗
02
4
6
Reg
LAGE
∗
0
0.2
0.4
0.6
TGFβ/β
-act
in
RegAGE
∗
00.1
0.20.3
0.4
Col
lgIV
/β-a
ctin
Reg
LAGE
∗
RegAGE
∗
02
4
6
Reg
LAGE
∗
0
0.2
0.4
0.6
TGFβ/β
-act
in
RegAGE
∗
00.1
0.20.3
0.4
Col
lgIV
/β-a
ctin
Reg
LAGE
∗
RegAGE
∗
02
4
6
Reg
LAGE
∗
0
0.2
0.4
0.6
TGFβ/β
-act
in
RegAGE
∗
00.1
0.20.3
0.4
Col
lgIV
/β-a
ctin
Reg
LAGE
∗
D. Alb/Cr E. TGF β1D. Alb/Cr E. TGF β1D. Alb/Cr E. TGF β1C. AGE C. AGE C. AGE F. Col IVF. Col IVF. Col IV
(Reg) (A) was prevented by a diet low in AGE contentia (D), TGF�1 (E), and Collagen IV mRNA expression (F)ta are shown in ratios of test mRNA/�-actin, by RT-PCR.
05
10
1520
05
10
1520
05
10
1520
AG
E U
/mg
r dietminurnt. Da

ssinObccimidcaord
O
Tasrtdtacdcdlpb
R
1
1
1
1
1
1
1
AGEs as a source of oxidants in aging 601
ubstances gradually erodes native defenses,etting the stage for abnormally high OS andnflammation; (5) this erosion may be operativeot only in patients with established increasedS, but, more importantly, in healthy subjectsefore the onset of end-organ decline; (6) be-ause a physiologic intervention (dietary AGEhange) effectively ameliorated increased OS, its reasonable to conclude that increased OS
ay be reversible, whether in healthy aging orn diseases (such as CKD or metabolic syn-rome); (7) the intervention is practical be-ause both younger and older subjects wereble to prepare low-AGE diets in their homesver a 4-month period; and (8) reduction inenal function in aging is one of the best pre-ictors of mortality.
VERALL SUMMARY
he data obtained from studies in experimentalnimals, normal aging subjects, and CKD patientsuggest that oxidants contribute to changes inenal function. Both cross-sectional and longi-udinal studies of aging persons suggest that aecline in renal function with age is not inevi-able. Because reduced renal function is associ-ted with cardiovascular disease, it seems verylear that interventions should be sought. Theata in animals suggest that the observed de-line in renal function with aging may be reme-iable by control of oxidants. Importantly, the
evels of oxidants can be controlled by non-harmacologic and pharmacologic means inoth animals and human beings. (Fig. 1).
EFERENCES1. Coresh J, Astor BC, Greene T, Eknoyan G, Levey AS.
Prevalence of chronic kidney disease and decreasedkidney function in the adult US population: ThirdNational Health and Nutrition Examination Survey.Am J Kidney Dis. 2003;41:1-12.
2. Hsu CY, Ordonez JD, Chertow GM, Fan D, McCullochCE, Go AS. The risk of acute renal failure in patientswith chronic kidney disease. Kidney Int. 2008;74:101-7.
3. Finkel TH. Oxidants, oxidative stress and the biologyof ageing. Nature. 2000;408:239-47.
4. Abbatecola AM, Ferrucci L, Grella R, Bandinelli S,Bonafe M, Barbieri M, et al. Diverse effect of inflam-matory markers on insulin resistance and insulin-re-sistance syndrome in the elderly. J Am Geriatr Soc.
2004;52:399-404.5. Cappola AR, Xue QL, Ferrucci L, Guralnik JM, VolpatoS, Fried LP. Insulin-like growth factor I and interleu-kin-6 contribute synergistically to disability and mor-tality in older women. J Clin Endocrinol Metab. 2003;88:2019-25.
6. Harris TB, Ferrucci L, Tracy RP, Corti MC, WacholderS, Ettinger WH Jr, et al. Associations of elevated in-terleukin-6 and C-reactive protein levels with mortal-ity in the elderly. Am J Med. 1999;106:506-12.
7. Reuben DB, Cheh AI, Harris TB, Ferrucci L, Rowe JW,Tracy RP, et al. Peripheral blood markers of inflam-mation predict mortality and functional decline inhigh-functioning community-dwelling older persons.J Am Geriatr Soc. 2002;50:638-44.
8. Stadtman ER. Role of oxidant species in aging. CurrMed Chem. 2004;11:1105-12.
9. Foley RN, Murray AM, Li S, Herzog CA, McBean AM,Eggers PW, et al. Chronic kidney disease and the riskfor cardiovascular disease, renal replacement, anddeath in the United States Medicare population, 1998to 1999. J Am Soc Nephrol. 2005;16:489-95.
0. Kilhovd BK, Juutilainen A, Lehto S, Ronnemaa T,Torjesen PA, Birkeland KI, et al. High serum levels ofadvanced glycation end products predict increased cor-onary heart disease mortality in nondiabetic women butnot in nondiabetic men: a population-based 18-year fol-low-up study. Arterioscler Thromb Vasc Biol. 2005;25:815-20.
1. Santopinto JJ, Fox KA, Goldberg RJ, Budaj A, PineroG, Avezum A, et al. Creatinine clearance and ad-verse hospital outcomes in patients with acute cor-onary syndromes: findings from the global registryof acute coronary events (GRACE). Heart. 2003;89:1003-8.
2. Koschinsky T, He CJ, Mitsuhashi T, Bucala R, Liu C,Buenting C, et al. Orally absorbed reactive glycationproducts (glycotoxins): an environmental risk factorin diabetic nephropathy. Proc Natl Acad Sci U S A.1997;94:6474-9.
3. Linden E, Cai W, He JC, Xue C, Li Z, Winston J, et al.Endothelial dysfunction in patients with chronic kid-ney disease results from advanced glycation end prod-ucts (AGE)-mediated inhibition of endothelial nitricoxide synthase through RAGE activation. Clin J AmSoc Nephrol. 2008;3:691-8.
4. Fried LF, Lee JS, Shlipak M, Chertow GM, Green C,Ding J, et al. Chronic kidney disease and functionallimitation in older people: health, aging and bodycomposition study. J Am Geriatr Soc. 2006;54:750-6.
5. Baylis C, Corman B. The aging kidney: insights fromexperimental studies. J Am Soc Nephrol. 1998;9:699-709.
6. Wiggins JE, Goyal M, Sanden SK, Wharram BL, Shed-den KA, Misek DE, et al. Podocyte hypertrophy, “ad-aptation,” and “decompensation” associated with glo-merular enlargement and glomerulosclerosis in theaging rat: prevention by calorie restriction. J Am Soc
Nephrol. 2005;16:2953-66.
1
1
1
2
2
2
2
2
2
2
2
2
2
3
3
3
3
3
3
3
3
3
3
4
4
4
4
602 H. Vlassara et al
7. Baynes JW, Thorpe SR. Role of oxidative stress indiabetic complications: a new perspective on an oldparadigm. Diabetes. 1999;48:1-9.
8. Hamada Y, Araki N, Koh N, Nakamura J, Horiuchi S,Hotta N. Rapid formation of advanced glycation endproducts by intermediate metabolites of glycolyticpathway and polyol pathway. Biochem Biophys ResCommun. 1996;228:539-43.
9. Uribarri J, Cai W, Peppa M, Goodman S, Ferrucci L,Striker G, et al. Circulating glycotoxins and dietaryadvanced glycation endproducts: two links to inflam-matory response, oxidative stress, and aging. J Ger-ontol A Biol Sci Med Sci. 2007;62:427-33.
0. Vlassara H, Cai W, Crandall J, Goldberg T, ObersteinR, Dardaine V, et al. Inflammatory mediators are in-duced by dietary glycotoxins, a major risk factor fordiabetic angiopathy. Proc Natl Acad Sci U S A. 2002;99:15596-601.
1. Vlassara H, Striker G. Glycotoxins in the diet promotediabetes and diabetic complications. Curr Diab Rep.2007;7:235-41.
2. Baynes JW, Thorpe SR. Glycoxidation and lipoxida-tion in atherogenesis. Free Radic Biol Med. 2000;28:1708-16.
3. Bucala R, Makita Z, Koschinsky T, Cerami A, VlassaraH. Lipid advanced glycosylation: pathway for lipidoxidation in vivo. Proc Natl Acad Sci U S A. 1993;90:6434-8.
4. Voziyan PA, Hudson BG. Pyridoxamine as a multi-functional pharmaceutical: targeting pathogenic gly-cation and oxidative damage. Cell Mol Life Sci. 2005;62:1671-81.
5. Thornalley PJ. Pharmacology of methylglyoxal: forma-tion, modification of proteins and nucleic acids, andenzymatic detoxification—a role in pathogenesis andantiproliferative chemotherapy. Gen Pharmacol. 1996;27:565-73.
6. Vlassara H. Advanced glycation in health and disease:role of the modern environment. Ann N Y Acad Sci.2005;1043:452-60.
7. Genuth S, Sun W, Cleary P, Sell DR, Dahms W, Ma-lone J, et al. Glycation and carboxymethyllysine levelsin skin collagen predict the risk of future 10-yearprogression of diabetic retinopathy and nephropathyin the diabetes control and complications trial andepidemiology of diabetes interventions and complica-tions participants with type 1 diabetes. Diabetes.2005;54:3103-11.
8. Cai W, He JC, Zhu L, Chen X, Wallenstein S, StrikerGE, et al. Reduced oxidant stress and extended life-span in mice exposed to a low glycotoxin diet: asso-ciation with increased AGER1 expression. Am JPathol. 2007;170:1893-902.
9. Oberg BP, McMenamin E, Lucas FL, McMonagle E,Morrow J, Ikizler TA, et al. Increased prevalence ofoxidant stress and inflammation in patients with mod-erate to severe chronic kidney disease. Kidney Int.2004;65:1009-16.
0. Li YM, Steffes M, Donnelly T, Liu C, Fuh H, Basgen
J, et al. Prevention of cardiovascular and renalpathology of aging by the advanced glycationinhibitor aminoguanidine. Proc Natl Acad Sci U S A.1996;93:3902-7.
1. Wolffenbuttel BH, Boulanger CM, Crijns FR, HuijbertsMS, Poitevin P, Swennen GN, et al. Breakers of ad-vanced glycation end products restore large arteryproperties in experimental diabetes. Proc Natl AcadSci U S A. 1998;95:4630-4.
2. Lin RY, Choudhury RP, Cai W, Lu M, Fallon JT, FisherEA, et al. Dietary glycotoxins promote diabetic ath-erosclerosis in apolipoprotein E-deficient mice. Ath-erosclerosis. 2003;168:213-20.
3. Himmelfarb J, McMonagle E. Manifestations of oxi-dant stress in uremia. Blood Purif. 2001;19:200-5.
4. Peppa M, Uribarri J, Cai W, Lu M, Vlassara H. Glycoxi-dation and inflammation in renal failure patients. Am JKidney Dis. 2004;43:690-5.
5. Schmidt AM, Hasu M, Popov D, Zhang JH, Chen J, YanSD, et al. Receptor for advanced glycation end prod-ucts (AGEs) has a central role in vessel wall interac-tions and gene activation in response to circulatingAGE proteins. Proc Natl Acad Sci U S A. 1994;91:8807-11.
6. Lu C, He JC, Cai W, Liu H, Zhu L, Vlassara H. Ad-vanced glycation endproduct (AGE) receptor 1 is anegative regulator of the inflammatory response toAGE in mesangial cells. Proc Natl Acad Sci U S A.2004;101:11767-72.
7. Vlassara H. The AGE-receptor in the pathogenesis ofdiabetic complications. Diabetes Metab Res Rev. 2001;17:436-43.
8. Schmidt AM, Yan SD, Yan SF, Stern DM. The multili-gand receptor RAGE as a progression factor amplify-ing immune and inflammatory responses. J Clin In-vest. 2001;108:949-55.
9. Schmidt AM, Hori O, Brett J, Yan SD, Wautier JL, SternD. Cellular receptors for advanced glycation endproducts. Implications for induction of oxidant stressand cellular dysfunction in the pathogenesis of vas-cular lesions. Arterioscler Thromb. 1994;14:1521-8.
0. Schmidt AM, Yan SD, Yan SF, Stern DM. The biologyof the receptor for advanced glycation end productsand its ligands. Biochim Biophys Acta. 2000;1498:99-111.
1. He CJ, Koschinsky T, Buenting C, Vlassara H. Pres-ence of diabetic complications in type 1 diabeticpatients correlates with low expression of mononu-clear cell AGE-receptor-1 and elevated serum AGE.Mol Med. 2001;7:159-68.
2. Zheng F, Plati AR, Potier M, Schulman Y, Berho M,Banerjee A, et al. Resistance to glomerulosclerosis inB6 mice disappears after menopause. Am J Pathol.2003;162:1339-48.
3. Zheng F, Cheng QL, Plati AR, Ye SQ, Berho M, Ban-erjee A, et al. The glomerulosclerosis of aging infemales: contribution of the proinflammatory mesan-gial cell phenotype to macrophage infiltration. Am J
Pathol. 2004;165:1339-48.
4
4
4
4
4
4
5
5
5
5
5
5
5
5
5
5
6
6
6
6
6
6
AGEs as a source of oxidants in aging 603
4. Zheng F, Plati AR, Cheng Q, Bertho M, Banerjee A, PotierM, et al. Glomerular aging in females is a multi-stagereversible process mediated by phenotypic changes inprogenitors. Am J Pathol. 2005;162:355-63.
5. Cornacchia F, Fornoni A, Plati AR, Thomas A, WangY, Inverardi L, et al. Glomerulosclerosis is transmittedby bone marrow-derived mesangial cell progenitors.J Clin Invest. 2001;108:1649-56.
6. Zheng F, Cornacchia F, Schulman I, Banerjee A,Cheng QL, Potier M, et al. Development of albumin-uria and glomerular lesions in normoglycemic B6 re-cipients of db/db mice bone marrow: the role ofmesangial cell progenitors. Diabetes. 2004;53:2420-7.
7. Tack I, Elliot SJ, Potier M, Rivera A, Striker GE, StrikerLJ. Autocrine activation of the IGF-I signaling pathwayin mesangial cells isolated from diabetic NOD mice.Diabetes. 2002;51:182-8.
8. Fornoni A, Lenz O, Striker LJ, Striker GE. Glucose in-duces clonal selection and reversible dinucleotide re-peat expansion in mesangial cells isolated from glomer-ulosclerosis-prone mice. Diabetes. 2003;52:2594-602.
9. Fornoni A, Striker LJ, Zheng F, Striker GE. Reversibilityof glucose-induced changes in mesangial cell extracel-lular matrix depends on the genetic background. Dia-betes. 2002;51:499-505.
0. Zheng F, Fornoni A, Elliot SJ, Guan Y, Breyer MD,Striker LJ, et al. Upregulation of type 1 collagen byTGF-beta in mesangial cells is blocked by PPAR-gamma activation. Am J Physiol Renal Physiol. 2002;282:F639-48.
1. Zheng F, Zeng YJ, Plati AR, Elliot SJ, Berho M, PotierM, et al. Combined AGE inhibition and ACEi de-creases the progression of established diabetic ne-phropathy in B6 db/db mice. Kidney Int. 2006;70:507-14.
2. Park C, Zhang Y, Zhang X, Wu J, Chen L, Cha DR, etal. PPARalpha agonist fenofibrate improves diabeticnephropathy in db/db mice. Kidney Int. 2006;69:1511-7.
3. Potier M, Karl M, Zheng F, Elliot SJ, Striker GE, StrikerLJ. Estrogen-related abnormalities in glomerulosclero-sis-prone mice: reduced mesangial cell estrogen re-ceptor expression and prosclerotic response to estro-gens. Am J Pathol. 2002;160:1877-85.
4. Elliot SJ, Karl M, Berho M, Potier M, Zheng F, Leclercq
B, et al. Estrogen deficiency accelerates progressionof glomerulosclerosis in susceptible mice. Am JPathol. 2003;162:1441-8.
5. Karl M, Berho M, Pignac-Kobinger J, Striker GE, ElliotSJ. Differential effects of continuous and intermittent17beta-estradiol replacement and tamoxifen therapyon the prevention of glomerulosclerosis: modulationof the mesangial cell phenotype in vivo. Am J Pathol.2006;169:351-61.
6. Potier M, Karl M, Elliot SJ, Striker GE, Striker LJ.Response to sex hormones differs in atherosclerosis-susceptible and -resistant mice. Am J Physiol Endocri-nol Metab. 2003;285:E1237-45.
7. Bobadilla NA, Tack I, Tapia E, Sanchez-Lozada LG,Santamaria J, Jimenez F, et al. Pentosan polysulfateprevents glomerular hypertension and structural in-jury despite persisting hypertension in 5/6 nephrec-tomy rats. J Am Soc Nephrol. 2001;12:2080-7.
8. Cai W, He JC, Zhu L, Chen X, Zheng F, Striker GE, etal. Oral glycotoxins determine the effects of calorierestriction on oxidant stress, age-related diseases, andlifespan. Am J Pathol. 2008;173:327-36.
9. Vlassara H. Protein glycation in the kidney: role indiabetes and aging. Kidney Int. 1996;49:1795-804.
0. Ruggiero C, Metter EJ, Melenovsky V, Cherubini A,Najjar SS, Ble A, et al. High basal metabolic rate is arisk factor for mortality: the Baltimore LongitudinalStudy of Aging. J Gerontol A Biol Sci Med Sci. 2008;63:698-706.
1. Lindeman RD, Tobin J, Shock NW. Longitudinal stud-ies on the rate of decline in renal function with age.J Am Geriatr Soc. 1985;33:278-85.
2. Dols LF, Weimar W, Ijzermans JN. Long-term conse-quences of kidney donation. N Engl J Med. 2009;360:2371-2.
3. Pizzarelli F, Lauretani F, Bandinelli S, Windham GB,Corsi AM, Giannelli SV, et al. Predictivity of survivalaccording to different equations for estimating renalfunction in community-dwelling elderly subjects.Nephrol Dial Transplant. 2009;24:1197-205.
4. Vlassara H, Cai W, Goodman S, Pyzik R, Yong A, ZhuL, et al. Protection against loss of innate defenses inadulthood by low AGE intake; role of a new anti-inflammatory AGE-receptor-1. Endocrinology. 2009.In press.
5. Uribarri J, Peppa M, Cai W, Goldberg T, Lu M, He C,et al. Restriction of dietary glycotoxins reduces ex-cessive advanced glycation end products in renal fail-
ure patients. J Am Soc Nephrol. 2003;14:728-31.



















