
Serine threonine protein kinases of mycobacterial genus: phylogeny to function

1Department of Zoology, Swedish Museum of Natural History, Stockholm Sweden; 2Temple University, Philadelphia PA, USA;3Museum of New Zealand Te Papa Tongarewa, Wellington New Zealand; 4Institute of Marine Biology, National Sun-Yat-SenUniversity, Kaohsiung Taiwan; 5Department of Aquatic Bioscience, National Chiayi University, Chiayi Taiwan
Hagfish phylogeny and taxonomy, with description of the new genus Rubicundus(Craniata, Myxinidae)
BO FERNHOLM1, MICHAEL NOR�EN
1, SVEN O. KULLANDER1, ANDREA M. QUATTRINI
2, VINCENT ZINTZEN3, CLIVE D. ROBERTS
3,HIN-KIU MOK
4 and CHIEN-HSIEN KUO5
AbstractA recent phylogenetic analysis of the Myxinidae based on the 16S rRNA gene resulted in synonymization of Paramyxine with Eptatretus. This createdhomonymy of Paramyxine fernholmi with Eptatretus fernholmi and Paramyxine wisneri with Eptatretus wisneri. In order to resolve this nomenclaturaldilemma, we made a more extensive phylogenetic assessment of the Myxinidae and examined the nomenclature of the family. We used 75 sequences(37 of which new for this study) of a 561 bp fragment of the 16S rRNA gene, representing 33 species, and 72 sequences (37 of which new for thisstudy) of a 687 bp fragment of the cytochrome c oxidase subunit I (COI) gene, representing 23 species, to reconstruct the phylogeny of Myxinidae.The monophyly of the subfamily Myxininae, traditionally characterized by having a single pair of external gill openings, was rejected (0.50 Bayesianposterior probability) by the 16S analysis, but supported by the COI and combined COI+16S analyses (0.99 and 0.81 Bpp, respectively). The mono-phyly of the subfamily Eptatretinae, characterized by having several pairs of external gill openings, was not supported by the 16S analysis and rejectedby the COI and combined COI+16S analysis due to the placement of Eptatretus lopheliae as the earliest branch of Myxinidae (0.71 and 0.57 Bpp,respectively). Eptatretus lopheliae and Eptatretus rubicundus formed a monophyletic group and were allocated to a new genus, Rubicundus, character-ized by the presence of an elongated tubular nostril and reddish coloration. A new monotypic subfamily, Rubicundinae, was proposed for Rubicundus.The synonymy of the genera Paramyxine and Quadratus with Eptatretus was confirmed. E. fernholmi is renamed Eptatretus luzonicus. Eptatretus wis-neri was renamed Eptatretus bobwisneri. Petromyzon cirrhatus Forster, 1801, Homea banksii Fleming, 1822, and Bdellostoma forsteri M€uller, 1836are synonyms, but no type specimens are known to exist. Petromyzon cirrhatus was designated as type species of Eptatretus, conserving presentusage. Gastrobranchus dombeyi Shaw, 1804 has priority over other names for Chilean myxinids. Bdellostoma stoutii was designated as type speciesof Polistotrema Gill. The validity of the Western Atlantic Myxine limosa as distinct from the Eastern Atlantic Myxine glutinosa was confirmed.
Key words: Myxinidae – Bayesian – classification – 16S rRNA – cytochrome c oxidase subunit I
Introduction
The Myxinidae are a family of basal marine craniates distributedin all oceans except the Arctic and Southern Ocean and the RedSea (Froese and Pauly 2012). 78 species are currently recognizedas valid (Froese and Pauly 2012). Myxinids are eel-like benthicscavengers and predators on small invertebrates and fish (Zintzenet al. 2011) with reduced eyes, no paired fins, a terminal com-plex jaw apparatus and slime pores along the lower side of theirbody. The taxonomy of the Myxinidae has been based mainly onvariation in the teeth and gill structures, as well as a limited setof meristic, morphological and colour characteristics, with thelatest review of the family by Fernholm (1998). Two subfamiliesare recognized. The Myxininae Linnæus (1758) are characterizedby having one pair of external gill openings while the Eptatreti-nae Bonaparte, 1850, have several (5–16) pairs of external gillopenings (Nelson 2006).
Up to seven genera are currently recognized (Nelson 2006),but a number of them have been disputed (Strahan 1975; Fern-holm 1998; Møller and Jones 2007; Kuo et al. 2010). TheMyxininae include Myxine Linnæus, 1758, with 23 species, Nem-amyxine Richardson, 1958, with two species, and the monotypicgenera Notomyxine Nani & Gneri, 1951, and NeomyxineRichardson, 1953. A second, undescribed species of Neomyxine
was reported by Zintzen et al. (2011). The Eptatretinae, with 51species, include the genera Eptatretus Cloquet, 1819; Paramyx-ine Dean, 1904 and Quadratus Wisner, 1999.
Until recently, myxinids have been caught mostly over softbottoms. However, recent opportunities to collect and observe inthe deep-sea using submersibles, deep potting and submergedcameras, have enabled sampling and observations in a morediverse range of habitats. Eptatretus lakeside Mincarone andMcCosker (2004) from the Gal�apagos Islands was collected froma submersible in 711 m depth. Eptatretus goliath Mincarone andStewart (2006) from northern New Zealand, the largest(1275 mm long) and heaviest (6.2 kg) hagfish, was collectedfrom 811 m depth using baited traps. Eptatretus strickrottiMøller and Jones (2007) was collected with a submersible froma hydrothermal vent habitat at 2211 m depth. An undescribedspecies of Eptatretus was observed on video and collected froma deep coral sponge garden at 300–700 m depth off northernNew Zealand (Zintzen et al. 2011). Eptatretus lopheliae Fern-holm and Quattrini (2008), collected with submersible from cold-water coral habitat in 430–442 m depth, and Eptatretus rubicun-dus Kuo et al. (2010), trawled in 800 m depth north-east ofTaiwan, have been found in remarkable new biotopes for hag-fishes and also show some interesting and novel morphologies.
The taxonomy of the Myxinidae currently suffers from severalunresolved issues. The validity of the genera Paramyxine andQuadratus is uncertain. Synonymization of Paramyxine withEptatretus was suggested already by Strahan (1975) andFernholm (1998). Møller and Jones (2007), in an analysis of a16S rDNA nucleotide data set from 17 species of the Myxinidae,provided molecular support for the synonymization of Paramyx-ine and Quadratus with Eptatretus. As a consequence of synony-
Corresponding author: Michael Nor�en ([email protected])Contributing authors: Bo Fernholm ([email protected]), Sven O. Kullander([email protected]), AndreaM.Quattrini ([email protected]), VincentZintzen ([email protected]), Clive D. Roberts ([email protected]),Hin-Kiu Mok ([email protected]), Chien-Hsien Kuo ([email protected])
Accepted on 1 June 2013© 2013 Blackwell Verlag GmbH J Zoolog Syst Evol Res doi: 10.1111/jzs.12035

mizing Paramyxine with Eptatretus resulting junior homonymsrequire new names. Eptatretus wisneri McMillan (1999) is pre-dated by Paramyxine wisneri Kuo et al. (1994), and Eptatretusfernholmi McMillan and Wisner (2004) by Paramyxine fernholmiKuo et al. (1994). In addition, the taxonomy and nomenclatureof Eptatretus is complicated by the dispute around the type spe-cies of Eptatretus (Gill 1901; Eschmeyer 2012), and also by thesynonymization of Quadratus and Paramyxine by Møller andJones (2007) which remains tentative because the type species ofthese genera, as well as the putative type species of Eptatretus,were not included in their analysis.
The present study aims first to provide an improved phyloge-netic hypothesis and classification of the Myxinidae, then solvethe resulting homonymy problems, and also resolve nomencla-tural problems associated with Eptatretus.
Material and Methods
Selection of gene fragment for molecular phylogeneticanalysis
This study uses mitochondrial 16S rDNA (16S, also known as the smallsubunit, ssu) and cytochrome oxidase subunit I (COI, also known asmt-CO1 and COX1) DNA fragments because these are the only geneswith a significant number of published sequences, permitting a bettersampling of myxinid diversity, and with variation and rate of evolutionsuitable for study of myxinid interrelationships. The 33 published 16Ssequences mostly come from earlier phylogenetic studies (Kuo et al.2003, 2010; Chen et al. 2005; Møller and Jones 2007), whereas the 30published COI sequences are mostly products of the Barcode of Life con-sortium. Preliminary analysis showed that four published COI sequences(JF493943 and JF493945–493947) were obtained from misidentifiedspecimens, and GenBank was notified.
DNA extraction and PCRMitochondrial DNA was extracted using a GeneMole (Mole Genetics)fully automated liquid-handling instrument, with the MoleStrips (MoleGenetics) kit and recommended protocol. The 16S fragment wasamplified using the primers 16SARL and 16SBRH (Palumbi et al.1991), the COI fragment was amplified using the standard barcodingprimers Fish-F1 and Fish-R1 (Ward et al. 2005). PCRs (PCR cycling:94°C 4′; 35* (94°C 30″; 52°C 30″; 72°C 30″); 72°C 8′) were per-formed with the puReTaq Ready-To-Go PCR kit (Amersham Bios-ciences AB, Uppsala, Sweden). PCR products were checked onminigel and purified using the FastAP Thermosensitive Alkaline Phos-phatase (Thermo Fischer Scientific, G€oteborg, Sweden) purification kit.
SequencingSequencing of both strands of all fragments was carried out by MacrogenEurope (Amstelveen, Holland). All sequences were proofread and assem-bled using the software Geneious v. 6.1.2 (Drummond et al. 2012).
Voucher specimensVoucher specimens were deposited at the Swedish Museum of NaturalHistory, NRM (Stockholm, Sweden), Scripps Institute of Oceanography,SIO (San Diego, USA), Icelandic Museum of Natural History, NMSI(Reykjavik, Iceland) and Museum of New Zealand Te Papa Tongarewa,NMNZ (Wellington, New Zealand). Data on voucher specimens areavailable from the Global Biodiversity Information Facility (GBIF) portal(data.gbif.org).
AnalysisSequences used in this study are summarized in (Table 1). All availablemyxinid 16S sequences from GenBank corresponding to the studied frag-ment and longer than 530 base pairs (bp) were included. All myxiniformCOI sequences available from GenBank on the 10th of March 2013 corre-sponding to the studied fragment and longer than 610 bp have beenincluded, except JF493943 which was excluded after analysis showed thatthe sequence is not of a myxiniform, but identical to published COI
sequences of the congrid eel Bassanago bulbiceps. The sequences werealigned using the MUSCLE (Edgar 2004) plug-in for Geneious. Align-ment of the 16S sequences was problematic, and alternative alignmentswere constructed using Geneious Alignment and the MAFFT (Katoh et al.2002, 2005) plug-in for Geneious in order to test sensitivity to alignment.
The combined data set was produced with all taxa for which both 16Sand COI data were available. The 16S alignment comprises 75 sequencesrepresenting 33 species and is 561 bp long, after insertions unique to out-groups had been deleted. The COI alignment comprises 72 sequencesrepresenting 23 species and is 687 bp long. The combined alignmentcomprises 42 sequences representing 18 species and is 1240 bp long. Allalignments are available from the authors.
Phylogenetic analysis was performed using the software MrBayes v.3.2 (Huelsenbeck and Ronquist 2001; Ronquist et al. 2012). COI datawas partitioned according to codon position (first, second, third), andparameters estimated separately for each partition. In the combined analy-sis, the 16S data comprised a fourth partition. The GTR + Γ + I modelwas used as suggested by ModelTest (Posada and Crandall 1998). Theanalysis was run for two million generations, at which time averagestandard deviation of split frequencies reported by MrBayes was ≤0.01.Samples were taken every 1000 generations, and the first 25% of sampleswere discarded as ‘burn-in’.
Parsimony bootstrap estimates indicate the amount of conflicting signalsin a data set. Parsimony bootstrap analysis was performed using the soft-ware TNT v1.1 (Goloboff et al. 2008), with 1000 replicates, 10 randomadditions per replicate, and using ‘classic search’ and TBR branch swap-ping. End gaps were coded as missing data, other gaps as a fifth state.
In all analyses, the trees were rooted with four non-myxinid chordates:Branchiostoma lanceolatum (Cephalochordata: Branchiostomatidae),Protopterus dolloi (Sarcopterygii: Lepidosirenidae), Petromyzon marinus(Petromyzontomorphi: Petromyzontidae) and Lampetra fluviatilis(Petromyzontomorphi: Petromyzontidae).
In all presented trees, (Figs 1–3) species represented by severalsequences and recovered in our analyses as monophyletic have been reducedto a single terminal. This is because intraspecific population structure is irrel-evant to the questions this study seeks to answer, and smaller trees are easierto read. However, the full, unreduced, versions of the presented trees areinstead provided as supplementary information (Figures S4–S6).
Taxonomic revision
The taxonomic and nomenclatural analysis was performed studying publi-cations and comparing with the International Code of Zoological Nomen-clature (International Commission on Zoological Nomenclature 1999),abbreviated ‘Code’ below. Entries in Eschmeyer (2012) were used asseed for collecting names and relevant taxonomic literature.
Results
Molecular phylogenetic analysis
16S data analysisThe Bayesian phylogenetic analysis of the 16S data is summarizedin Fig. 1. The base of the tree separated into two sister groups, thefirst one composed of Neomyxine sp. and Neomyxine biniplicata(Bayesian posterior probability 1.0), the second composed of allother species of Myxinidae (Bpp 0.50). This second group is fur-ther divided into three groups corresponding to Rubicundus (Bpp1.0), Myxininae excluding Neomyxine (Bpp 0.98), and Eptatretinaeexcluding Rubicundus (Bpp 1.0). Within the Myxininae, Myxinesp. ‘1-CHK-2001’, Myxine formosana, and one of the two Myxinecircifrons formed a cohesive group (Bpp 0.76), whereas remainingMyxininae formed another supported (Bpp 0.99) group with littleinternal structure. The monophyly of N. sp., N. biniplicata, M. sp.‘1-CHK-2001’, Myxine limosa and Myxine jespersenae wassupported, (Bpp ≥ 0.99), but the monophyly of Myxine glutinosawas not supported (Bpp < 0.5), and the monophyly of M. circi-frons was rejected.
In the Eptatretinae, Eptatretus cheni branched first. Eptatretusminor, E. cirrhatus and a group comprising the remaining
doi: 10.1111/jzs.12035© 2013 Blackwell Verlag GmbH
2 FERNHOLM, NOR�EN, KULLANDER, QUATTRINI, ZINTZEN, ROBERTS, MOK and KUO

Table 1. List of species and sequences used in this study. Taxa marked with an asterisk were designated out-group. Minus-sign in the voucher columnindicates absence of information. JX442444 to JX442480 and KC807319 to KC807355 are new for this study
Species GenBank# 16S GenBank# COI Voucher Origin
Eptatretus burgeri – AJ278504 – JapanEptatretus burgeri AF364616 – – TaiwanEptatretus burgeri AF364617 – – TaiwanEptatretus burgeri AY619579 – – ‘China’ (Taiwan)Eptatretus burgeri JX442457 KC807331 NRM 50265-T3326 JapanEptatretus burgeri JX442458 KC807331 NRM 50265-T3325 JapanEptatretus burgeri JX442459 KC807321 NRM 50265-T3327 JapanEptatretus burgeri JX442460 KC807323 NRM 50265-T3329 JapanEptatretus burgeri JX442461 KC807322 NRM 50265-T3328 JapanEptatretus burgeri NC_002807 NC_002807 – JapanEptatretus cf fernholmi JX442463 KC807333 – TaiwanEptatretus cheni AF364620 – – TaiwanEptatretus cheni AF364621 – – TaiwanEptatretus chinensis AY619580 – – ‘China’ (Taiwan)Eptatretus cirrhatus AF364619 – – New ZealandEptatretus cirrhatus JX442450 KC807340 NMNZ P.037107 New ZealandEptatretus cirrhatus JX442451 KC807345 NMNZ P.049408 New ZealandEptatretus cirrhatus JX442452 KC807341 NMNZ P.044178 New ZealandEptatretus cirrhatus JX442453 – NMNZ P.037133 New ZealandEptatretus deani – FJ164594 TZ05-FROSTI-325 Canada, British ColumbiaEptatretus deani – FJ164595 TZ05-FROSTI-330 Canada, British ColumbiaEptatretus deani – FJ164596 TZ05-FROSTI-331 Canada, British ColumbiaEptatretus deani – FJ164597 TZ05-FROSTI-302 Canada, British ColumbiaEptatretus deani – FJ164598 TZ05-FROSTI-215 Canada, British ColumbiaEptatretus deani – GU440316 MFC317 USAEptatretus deani EF014477 – KU 28249 USA, CaliforniaEptatretus deani EU099514 – SIO 05-95 USA, CaliforniaEptatretus longipinnis EF014476 – SAMA F07540 Australia, SouthEptatretus minor JX442454 KC807329 – USA, Gulf of MexicoEptatretus minor JX442455 KC807331 – USA, Gulf of MexicoEptatretus minor JX442456 KC807331 – USA, Gulf of MexicoEptatretus nelsoni AF364607 – – Taiwan?Eptatretus nelsoni AF364608 – – TaiwanEptatretus nelsoni AF364609 – – Taiwan?Eptatretus sheni AF364610 – – TaiwanEptatretus sp. ‘Korea’ JX442462 KC807324 NRM 50590 South KoreaEptatretus stoutii – FJ164599 NEOCAL07-0004 Canada, British ColumbiaEptatretus stoutii – FJ164600 NEOCAL07-0003 Canada, British ColumbiaEptatretus stoutii – FJ164601 NEOCAL07-0002 Canada, British ColumbiaEptatretus stoutii – FJ164602 NEOCAL07-0001 Canada, British ColumbiaEptatretus stoutii – FJ164603 TZ-06-RICKER-573 Canada, British ColumbiaEptatretus stoutii – FJ164604 TZ-06-RICKER-574 Canada, British ColumbiaEptatretus stoutii – FJ164605 TZ-06-RICKER-607 Canada, British ColumbiaEptatretus stoutii – FJ164606 TZ-06-RICKER-608 Canada, British ColumbiaEptatretus stoutii – FJ164607 TZ-06-RICKER-490 Canada, British ColumbiaEptatretus stoutii – GU440317 MFC124 USA, San DiegoEptatretus stoutii AF364618 – – USA, OregonEptatretus stoutii EU099456 – SIO 95-2 USA, CaliforniaEptatretus strickrotti EF014478 – CAS 223480 East Pacific RiseEptatretus taiwanae AF364611 – – TaiwanEptatretus yangi AF364612 – – TaiwanEptatretus yangi AF364613 – – TaiwanEptatretus yangi AF364614 – – TaiwanEptatretus yangi AF364615 – – TaiwanMyxine affinis JX442466 KC807353 UNMDP-T 0919 Argentina, Plataforma PatagoniaMyxine affinis JX442467 KC807352 UNMDP-T 0041 Argentina, Plataforma PatagoniaMyxine affinis JX442471 KC807349 UNMDP-T 0025 Argentina, Plataforma PatagoniaMyxine australis – EU074490 INIDEP-T 0132 ArgentinaMyxine australis JX442468 KC807354 UNMDP-T 0935 Argentina, Ushuaia, Mar del PlataMyxine australis JX442469 KC807351 UNMDP-T 0035 Argentina, Plataforma PatagoniaMyxine australis JX442470 KC807355 UNMDP-T 0936 Argentina, Ushuaia, Mar del PlataMyxine capensis – JF493944 ADC09_1.4#4 South Africa, Agulhas BankMyxine capensis – JF493945 ADC09_1.4 #3 South Africa, Agulhas BankMyxine capensis – JF493946 ADC09_1.4#2 South Africa, Agulhas BankMyxine capensis – JF493947 ACD07_1.4 #1 South AfricaMyxine circifrons AF364628 – – USAMyxine circifrons EU099504 – SIO 05-104 USA, CaliforniaMyxine fernholmi JX442465 KC807350 UNMDP-T 0026 Argentina, Plataforma PatagoniaMyxine formosana – JN027323 NAFF 6195 Taiwan?
doi: 10.1111/jzs.12035© 2013 Blackwell Verlag GmbH
Hagfish phylogeny and taxonomy 3

Eptatretinae formed an unresolved polytomy (Bpp 0.97). Withinthis group of Eptatretinae, E. strickrotti, E. longipinnis and agroup comprising the remaining Eptatretinae formed anotherunresolved polytomy (Bpp 0.93). Within this last group, E. de-ani + E. stoutii (Bpp 0.98) was the sister group of a clade com-prising all remaining Eptatretinae (Bpp 0.84). The Bayesiananalysis supported the monophyly of E. cheni, E. cirrhatus,E. minor, E. deani, E. stoutii and Eptatretus nelsoni (Bpp 1.0),and the monophyly of E. burgeri (0.69) and E. yangi (Bpp0.77). All included members of the proposed genus Paramyxineexcept E. cheni formed a largely unresolved group (Bpp 0.72).
The majority-rule parsimony bootstrap tree inferred by TNT(not shown; provided as supporting information, Figure S1) wascompatible with the majority-rule Bayesian tree inferred usingMrBayes (Fig. 1). The parsimony bootstrap tree differs byhaving lower resolution in Eptatretinae and Myxininae.
The 16S tree was sensitive to alignment, with different meth-ods and parameters of alignment resulting in different relativebranching order of Myxininae, Eptatretinae, Rubicundus andNeomyxine. However, the tree was robust to the method ofanalysis, with parsimony bootstrap and Bayesian analysis pro-ducing compatible trees when analysing the same alignment.
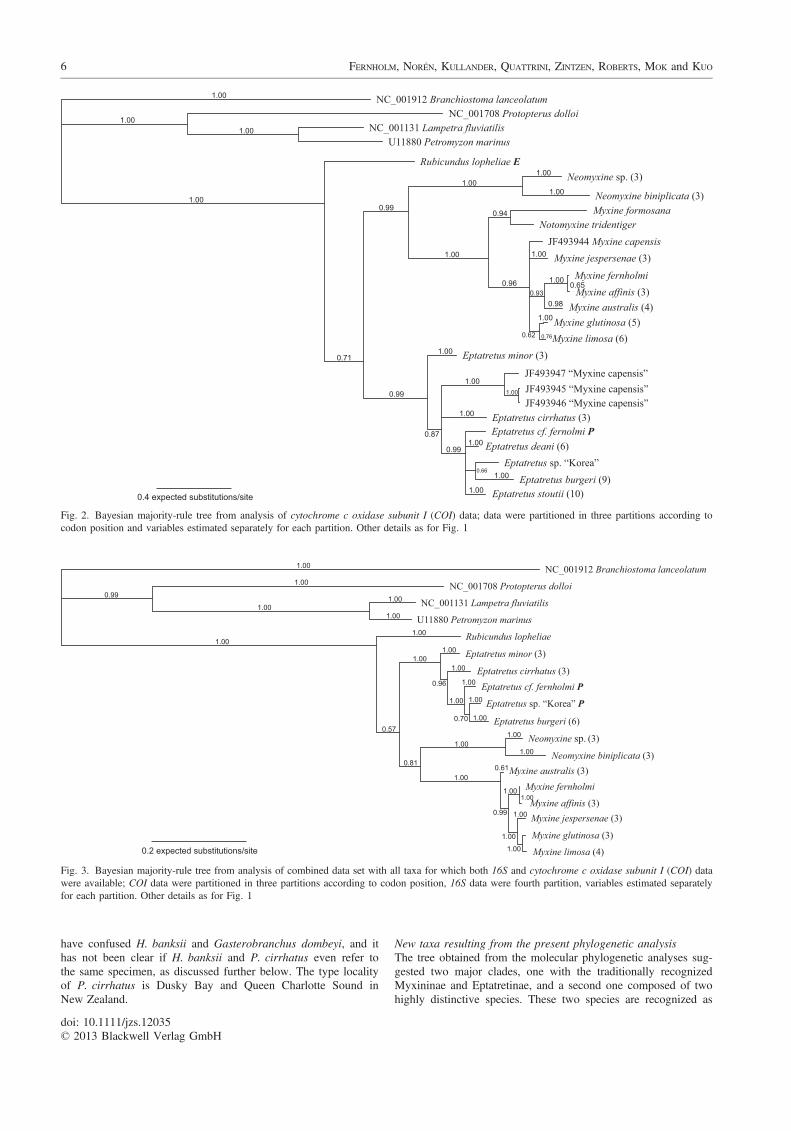
COI data analysisThe Bayesian phylogenetic analysis of the COI data is summa-rized in Fig. 2. The base of the tree separated into two sistergroups, the first one composed of Rubicundus lopheliae, thesecond one with all other species of Myxinidae (Bpp 0.71). Thissecond group was further divided into two groups corresponding
to Myxininae (Bpp 0.99) and Eptatretinae excluding Rubicundus(Bpp 0.99). Within the Myxininae, the earliest branch is Neomyx-ine sp. + Neomyxine biniplicata (Bpp 1.0). The next group tobranch comprised Myxine formosana + Notomyxine tridentiger(Bpp 0.94), while the remaining Myxininae formed a polytomy inwhich Myxine fernholmi + Myxine affinis + Myxine australisformed a cohesive group (Bpp 0.93) and Myxine glutinosa + Myx-ine limosa another (Bpp 0.62). The monophyly of Neomyxine sp.,N. biniplicata, M. affinis, M. australis, M. glutinosa, M. limosaand M. jespersenae was supported (Bpp 1.0 for all except limosa,Bpp 0.76) but the monophyly of M. capensis was rejected.
In the Eptatretinae, E. minor branched first. ‘M. capensis’,E. cirrhatus, and a group comprising the remaining Eptatretinaeformed an unresolved polytomy (Bpp 0.99). Within this group ofEptatretinae, E. sp. ‘Korea’ formed a monophyletic group withE. burgeri (Bpp 0.66). The monophyly of E. minor, ‘M. capensis’,E. cirrhatus, E. deani, E. burgeri and E. stoutii was supported(Bpp 1.0 for all).
The majority-rule parsimony bootstrap tree inferred by TNT(not shown; provided as supporting information, Figure S2) wassimilar, but conflicted with the majority-rule Bayesian tree(Fig. 1) by reconstructing Neomyxine as the earliest branchwithin Myxinidae, while Rubicundus, Eptatretinae and Myxininaeformed an unresolved trichotomy, and by having M. formosanaas the earliest branch within Myxininae.
Combined 16S+COI data analysisThe Bayesian phylogenetic analysis of all myxinids for whichboth 16S and COI data were available is summarized in Fig. 3.
Table 1. (continued)
Species GenBank# 16S GenBank# COI Voucher Origin
Myxine formosana AF364625 – – TaiwanMyxine glutinosa – KC807339 NRM 47489Myxine glutinosa – Y15182 – Sweden?Myxine glutinosa AJ404477 AJ404477 – Sweden, ‘Baltic Sea’Myxine glutinosa JX442475 KC807338 NRM 47485 Sweden, SkagerrakMyxine glutinosa JX442476 KC807337 NRM 47487 Sweden, SkagerrakMyxine jespersenae JX442472 KC807336 NMSI 760 IcelandMyxine jespersenae JX442473 KC807334 NMSI 761 IcelandMyxine jespersenae JX442474 KC807335 NMSI 759 IcelandMyxine limosa – KC015742 SIO 06-781 Canada, St. LawrenceMyxine limosa – KC015744 SIO 06-913 Canada, St. LawrenceMyxine limosa JX442477 KC807327 – USA, North CarolinaMyxine limosa JX442478 KC807326 – USA, North CarolinaMyxine limosa JX442479 KC807332 – USA, North CarolinaMyxine limosa JX442480 KC807328 –Myxine sp. ‘1-CHK-2001’ AF364622 – – TaiwanMyxine sp. ‘1-CHK-2001’ AF364623 – – TaiwanMyxine sp. ‘1-CHK-2001’ AF364624 – – TaiwanMyxine sp. ‘2-CHK-2001’ AF364626 – – TaiwanMyxine sp. ‘3-CHK-2001’ AF364627 – – TaiwanNeomyxine biniplicata JX442447 KC807347 NMNZ P.052442/TS2 New ZealandNeomyxine biniplicata JX442448 KC807348 NMNZ P.052443/TS2 New ZealandNeomyxine biniplicata JX442449 KC807346 NMNZ P.052441/TS2 New ZealandNeomyxine sp. JX442444 KC807343 NMNZ P.047438 New ZealandNeomyxine sp. JX442445 KC807342 NMNZ P.044215 New ZealandNeomyxine sp. JX442446 KC807344 NMNZ P.047862 New ZealandNotomyxine tridentiger – EU074500 INIDEP-T 0131 ArgentinaRubicundus lopheliae JX442464 KC807325 – USA, North
CarolinaRubicundus rubicundus AY033088 – ASIZP 60660 Taiwan* Branchiostomalanceolatum
NC_001912 NC_001912
* Lampetra fluviatilis NC_001131 NC_001131* Petromyzon marinus U11880 U11880* Protopterus dolloi NC_001708 NC_001708
doi: 10.1111/jzs.12035© 2013 Blackwell Verlag GmbH
4 FERNHOLM, NOR�EN, KULLANDER, QUATTRINI, ZINTZEN, ROBERTS, MOK and KUO

The tree topology is compatible with the COI tree, differing onlyin support values. Excluding intraspecific nodes, all nodes haveBpp ≥ 0.99, except for the placement of Rubicundus lopheliae asfirst branch in Myxinidae (Bpp 0.57), the placement of Neomyx-ine as the sister group of all other Myxininae (0.81) and the sis-ter group relationship of Eptatretus sp. and E. burgeri (Bpp0.70). The monophyly of all included species is recovered (Bpp1.00; M. australis Bpp 0.61).
The majority-rule parsimony bootstrap tree inferred by TNT(not shown; provided as supporting information, Figure S3) con-flicted with the majority-rule Bayesian tree (Fig. 3) by recon-structing Neomyxine as the earliest branch within Myxinidae andRubicundus as the sister group to Eptatretinae.
Taxonomy
Type species and validity of EptatretusThe original description of Eptatretus (Cloquet, 1819) wasbased on two species, both identified as Eptatretus Dombeii
(= Gasterobranchus dombeyi Shaw, 1804). With reference toCode article 70.3, we designate here as type species of Eptatr-etus the species corresponding to the diagnostic charactersreported by Cloquet, viz., Homea banksii Fleming (1822).Homea banksii is a junior synonym of Petromyzon cirrhatusForster (in Schneider 1801). Bdellostoma heptatrema M€uller(1836) is an unjustified replacement name for P. cirrhatus andthus an objective junior synonym. Gasterobranchus dombeyiShaw, 1804, described from Chile, is an available name for adifferent species, most likely a senior synonym of Eptatretuspolytrema (Girard, 1855a), E. nanii Wisner and McMillan(1988), or E. bischhoffi (Schneider, 1880), but the type speci-men was lost.
No types are known to exist of H. banksii or P. cirrhatus. Aneotype from the type locality area of P. cirrhatus has beenselected and is being formally designated by Zintzen et al. (inprep.) covering H. banksii, B. heptatrema and P. cirrhatus. Thisneotype designation is necessary because of the prevailing uncer-tainty concerning the type species of Eptatretus. Previous authors
1.00
0.57
0.50
1.00
1.00
1.00
0.98
1.00
1.00
0.97
1.00
0.76
0.99
0.72
1.00
1.00
1.00
0.76
0.98
0.93
1.00
0.90
0.99
1.00
0.99
0.84
0.98
0.98
0.92
0.69
1.00
1.00
0.92
0.81
1.00
0.77
NC_001912 Branchiostoma lanceolatum NC_001708 Protopterus dolloi
NC_001131 Lampetra fluviatilis U11880 Petromyzon marinus
Rubicundus rubicundus ERubicundus lopheliae E
Neomyxine sp. (3)
Neomyxine biniplicata (3)
Eptatretus cheni (2) P
EU099504 Myxine circifrons
Myxine fernholmi
Eptatretus strickrottiEptatretus longipinnis
Eptatretus minor (3)
Myxine formosana
AJ404477 Myxine glutinosa JX442475 Myxine glutinosa JX442476 Myxine glutinosa
Myxine affinis (3)
Eptatretus cirrhatus (5)
Myxine sp. “1 CHK 2001” (3)
AF364629 Myxine circifrons
Myxine jespersenae (3)
Myxine limosa (4)
Myxine australis (3)
Eptatretus sp. “Korea”
Myxine sp. “2 CHK 2001” Myxine sp. “3 CHK 2001”
AF364628 Myxine circifrons
Eptatretus sheni PEptatretus burgeri (9)
Eptatretus deani (2)
Eptatretus stoutii (2)
Eptatretus taiwanae Q, PEptatretus chinensis Q, P
Eptatretus cf. fernholmi P Eptatretus nelsoni (3) P
Eptatretus yangi (4) Q, P0.2 expected substitutions per site
1.00
Fig. 1. Bayesian majority-rule tree from analysis of mitochondrial 16S rDNA data. Values at nodes indicate Bayesian posterior probability estimates.Branch lengths are proportional to number of expected substitutions per site. Monophyletic species have been reduced to a single terminal; number ofincluded sequences is indicated in parenthesis. Sequences of species recovered as non-monophyletic are indicated by having GenBank accession num-ber in front of the species name. Species transferred to a new genus in this study have their old genus indicated with an E for Eptatretus, P for Para-myxine, Q for Quadratus
doi: 10.1111/jzs.12035© 2013 Blackwell Verlag GmbH
Hagfish phylogeny and taxonomy 5
have confused H. banksii and Gasterobranchus dombeyi, and ithas not been clear if H. banksii and P. cirrhatus even refer tothe same specimen, as discussed further below. The type localityof P. cirrhatus is Dusky Bay and Queen Charlotte Sound inNew Zealand.
New taxa resulting from the present phylogenetic analysisThe tree obtained from the molecular phylogenetic analyses sug-gested two major clades, one with the traditionally recognizedMyxininae and Eptatretinae, and a second one composed of twohighly distinctive species. These two species are recognized as
1.00
1.00
1.00
0.71
1.00
0.99
0.99
1.00
1.00
0.87
1.00
1.00
1.00
0.94
0.96
1.00
1.00
0.99
1.00
0.62
0.93
1.00
0.66
1.00
1.00
1.00
0.76
1.00
0.98
1.00
0.65
NC_001912 Branchiostoma lanceolatum
Rubicundus lopheliae E
NC_001708 Protopterus dolloi NC_001131 Lampetra fluviatilis
U11880 Petromyzon marinus
Eptatretus minor (3)
Neomyxine sp. (3)
Neomyxine biniplicata (3) Myxine formosana
Notomyxine tridentiger JF493944 Myxine capensis
JF493947 “Myxine capensis”
Eptatretus cirrhatus (3) Eptatretus cf. fernolmi P
Myxine jespersenae (3)
JF493945 “Myxine capensis” JF493946 “Myxine capensis”
Eptatretus sp. “Korea”
Eptatretus stoutii (10)
Eptatretus deani (6)
Myxine glutinosa (5) Myxine limosa (6)
Myxine fernholmi
Myxine australis (4)
Eptatretus burgeri (9)
Myxine affinis (3)
0.4 expected substitutions/site
Fig. 2. Bayesian majority-rule tree from analysis of cytochrome c oxidase subunit I (COI) data; data were partitioned in three partitions according tocodon position and variables estimated separately for each partition. Other details as for Fig. 1
1.00
1.00
0.99
0.57
1.00
1.00
1.00
1.00
0.81
1.00
1.00
0.96
1.00
1.00
1.00
1.00
1.00
0.99
0.61
1.00
1.00
0.70
1.00
1.00
1.00
1.00
1.00
1.00
1.00
1.00
NC_001912 Branchiostoma lanceolatum
Rubicundus lopheliae
NC_001708 Protopterus dolloi
NC_001131 Lampetra fluviatilis
U11880 Petromyzon marinus
Eptatretus minor (3)
Eptatretus cf. fernholmi PEptatretus cirrhatus (3)
Myxine australis (3)
Neomyxine sp. (3)
Neomyxine biniplicata (3)
Eptatretus sp. “Korea” P
Myxine fernholmi
Eptatretus burgeri (6)
Myxine jespersenae (3) Myxine affinis (3)
Myxine limosa (4)
Myxine glutinosa (3) 0.2 expected substitutions/site
Fig. 3. Bayesian majority-rule tree from analysis of combined data set with all taxa for which both 16S and cytochrome c oxidase subunit I (COI) datawere available; COI data were partitioned in three partitions according to codon position, 16S data were fourth partition, variables estimated separatelyfor each partition. Other details as for Fig. 1
doi: 10.1111/jzs.12035© 2013 Blackwell Verlag GmbH
6 FERNHOLM, NOR�EN, KULLANDER, QUATTRINI, ZINTZEN, ROBERTS, MOK and KUO

belonging to a new genus. In order to preserve the recognition ofthe subfamilies Myxininae and Eptatretinae, a new subfamily fortheir sister clade is proposed.
Rubicundus, new genus.Type species: Eptatretus rubicundus Kuo et al. (2010).Referred species: Eptatretus lopheliae, Eptatretus eos Fernholm
(1991), Eptatretus lakeside Mincarone and McCosker (2004).Etymology: Rubicundus repeats the specific epithet of the type
species, emphasizing the reddish coloration of the included spe-cies. It is a Latin adjective meaning red, here treated as a mascu-line noun in the nominative singular.
Diagnosis: Myxinids with five gill openings, uniquely distin-guished by elongated, tubular nostril (versus short in all otherMyxinidae; Fig. 4) and reddish colour (versus grey or brown,never reddish, in all other Myxinidae; Fig. 4).
Rubicundinae, new subfamily of Myxinidae.Type-genus: Rubicundus, present paper, only included genus.Diagnosis: Same as for the genus Rubicundus.
Replacement of scientific names resulting from the present phylo-genetic analysisSynonymization of species from Paramyxine with Eptatretusresulted in two cases of homonymy that requires replacementnames. Eptatretus wisneri McMillan (1999) from the Gal�apagosArchipelago becomes a junior homonym of Paramyxine wisneriKuo et al. (1994) from Taiwan. Eptatretus wisneri McMillan isherewith given the new name Eptatretus bobwisneri to maintainits affiliation to Robert L. Wisner (1921–2005) of the ScrippsInstitute of Oceanography. Eptatretus fernholmi McMillan andWisner (2004) from the Philippines becomes a junior homonymof Paramyxine fernholmi Kuo et al. (1994) from Taiwan and isherewith renamed E. luzonicus. This specific name was men-tioned by McMillan and Wisner (2004) in the description as thename initially intended for the species. The single known speci-men was caught east of the southern end of the Luzon Island inthe Philippines. The species epithet luzonicus, alluding to the typelocality, is an adjective with alternative endings -a, and -um.
Discussion
Molecular phylogenetic analysis
The COI results were robust to methods of alignment, but in the16S analysis, the position of Neomyxine, and to a lesser degree
Rubicundus, was not. The tree depicted in (Fig. 1) was based onthe alignment obtained from MUSCLE, but analysing alignmentsproduced by Geneious and MAFFT variously recovered Neomyx-ine as either (1) the earliest branch within the Myxinidae (inwhich case Rubicundus either forms an unresolved trichotomywith the Eptatretinae and Myxininae, or is the sister taxon of theMyxininae), (2) the sister taxon of the Eptatretinae or (3) as amember of an unresolved polytomy encompassing Myxininae,Eptatretinae and Rubicundus. From this, we infer that there isinsufficient information regarding the relative branching order ofNeomyxine, Rubicundus, Eptatretinae and Myxininae in our 16Sdata and that resolution at that level in the tree is due to differ-ences in alignment.
Given alignment, the Bayesian and the parsimony bootstrapmajority-rule trees were largely compatible, differing mostly inresolution, with the exception of the placement of Neomyxine.In all parsimony bootstrap analyses, Neomyxine was the earliestbranch in Myxinidae, whereas in Bayesian analyses, Neomyxinewas the earliest branch in Myxinidae only in the 16S analysis,while Rubicundus was the earliest branch in the COI and com-bined analyses, as well as with some alternative alignments of16S. This discrepancy may be a result of signal erosion(dubbed a Class II Long Branch artefact by W€agele and Mayer2007), as all out-groups are distant, with branches roughly oneorder of magnitude longer than branches inside Myxinidae,except the long branches of Rubicundus and Neomyxine andBayesian analysis is more resistant to long branch artefacts thanparsimony analysis.
There exists no more closely related out-group taxa that couldbe added to the analysis, and performing the parsimony bootstrapanalysis without out-groups results in Neomyxine, Rubincundi-nae, Myxininae and Eptatretinae forming a polytomy. The rela-tive branching order of these taxa remains uncertain.
Our reconstructions of the phylogeny of the Myxinidae(Figs 1–3) are similar to what Kuo et al. (2010) proposed basedon an analysis of 16S data, differing mainly in the placement ofRubicundus rubicundus. In their tree, R. rubicundus was the sis-ter group to all other Eptatretinae, but with weak support (0.63Bpp), and as previously discussed, the position of Rubicundus inanalyses of 16S data is sensitive to alignment. Møller and Jones(2007) presented a tree with a topology similar to ours for theMyxininae, but different from ours for the Eptatretinae. In theirtree, E. strickrotti was the first branch within the Eptatretinae.To test if this difference was due to the increased number of
Fig. 4. Comparison of the head of Eptatretus burgeri (NRM 57630), total length 320 mm and the holotype of Rubicundus eos (NMNZ P.24262), totallength 665 mm, illustrating the diagnostic reddish colour and tubular nostril of the genus Rubicundus. Preserved specimens, colours somewhat fadedcompared to life. Left: ventral view of the head of E. burgeri. Right: ventral view of the head of R. eos. The images have been scaled and cropped,levels and sharpness have been adjusted and noise reduced
doi: 10.1111/jzs.12035© 2013 Blackwell Verlag GmbH
Hagfish phylogeny and taxonomy 7

specimens and species included in our study, we performed ananalysis with just the 27 sequences used in Møller and Jones(2007). We could not reproduce their results, and the tree obtainedwas similar to the results of the 16S analysis of this study (Fig. 1).The discrepancy may be explained by differences in method ofanalysis: Møller and Jones (2007) did not provide sufficient infor-mation on the parameters used in their analysis, such as methodof alignment, data partitioning or model used in their Bayesiananalysis, preventing us from repeating their results.
The rejected monophyly of Myxine circifrons suggests thatone or several of the sequences EU099504, AF364628 andAF364629 deposited in GenBank were obtained from misidenti-fied specimens. Voucher specimens do not exist for thesesequences, leaving their identity unresolved. The long branch ofAF364628 could be an artefact caused by an apparent sequenc-ing error at positions 72–83.
The rejected monophyly of M. capensis also seems to becaused by misidentifications. One of the four M. capensissequences (JF493944) groups inside Myxininae and is presum-ably correct, but the three remaining ‘M. capensis’ (JF493945–JF493947) group inside Eptatretinae, indicating that thesesequences represent species of Eptatretus. The sequencesJF493944–JF493947 were also included in an analysis byKuraku (2013). Their analysis (their fig. 5) also recoveredE. capensis as non-monophyletic. GenBank has been notifiedof the error. The ‘M. capensis’ sequences originate from theBarcode of Life initiative, preserved voucher specimens exist,and the sequence identity will hopefully be corrected.
The sequences KC015742 and KC015744 are listed in Gen-Bank as M. glutinosa from Canada, but in this study representM. limosa. This is not a result of misidentification: the north-west Atlantic M. limosa is often considered a synonym of thenorth-east Atlantic M. glutinosa.
Taxonomy
Twenty nominal genera have been recognized in the Myxinidae:Bdellostoma M€uller, 1836; Dodecatrema Fowler, 1947,Heptatrema Dum�eril, 1832, Heptatremus Swainson, 1839, Hep-tatretus Regan, 1912, Hexabranchus Schultze, 1835, HomeaFleming, 1822; Polistotrema Gill, 1881 and Polytrema Girard,1855b; are generally considered as junior synonyms of EptatretusCloquet, 1819; when not simply unjustified emendations. Para-myxine Dean (1904) and Quadratus Wisner (1999) are recog-nized as valid in recent publications (Nelson 2006), but also
often considered synonyms of Eptatretus (Møller and Jones2007). Myxine (Linnæus 1758) has well established juniorsynonyms Anopsis Agassiz, 1846, Anopsus Rafinesque, 1815,Gastrobranchus Bloch, 1791 and Muraenoblenna La Cep�ede,1803. Neomyxine Richardson, 1953 and Nemamyxine Richardson,1958, both with two species, and the monotypic NotomyxineNani & Gneri, 1951, are three little studied genera, universallyconsidered valid.
Traditionally, the Myxinidae has been organized in two sub-families, Eptatretinae for the hagfishes with several pairs of gillopenings and Myxininae for species with only one pair of gillopenings (Fernholm 1998; Nelson 2006). Our molecular resultsare not compatible with that of dichotomous division. The newgenus Rubicundus has several pairs of gill openings, but doesnot belong to the traditional Eptatretinae. Clearly, a more compli-cated taxonomy of the Myxinidae is emerging. We believe thathaving several pairs of external gill openings is the plesiomor-phic character state within Myxinidae, while having one pair ofexternal gill openings is a synapomorphy of the Myxininae. Neo-myxine, with a single pair of gill openings, has traditionally beenincluded in the Myxininae, a position challenged by the 16S phy-logeny (Fig. 1) and parsimony bootstrap analysis. However, theplacement of Neomyxine as the first branch in Myxinidae isdependent on alignment, and in the Bayesian COI (Fig. 2) andcombined analyses (Fig. 3), Myxininae is monophyletic. Also,Nemamyxine and Notomyxine have one pair of gill openings andare consequently traditionally assigned to the Myxininae.Pending further studies, we choose to continue the prevailingpraxis of assigning these three genera to the Myxininae.
EptatretusAny classification of myxinid species is problematic because ofuncertainty about the type species of Eptatretus, a genus tradi-tionally diagnosed by the multiple gill openings. The problemwas already recognized by Gill (1901) who presented a detailedanalysis of the status of Eptatretus and its synonym Bdellostoma.The first description of a hagfish with multiple gill openings mayhave been by Home (1815), who described it as an animal ‘inter-mediate between the lamprey and the myxine’, and considered itto be a new genus. Dum�eril (1812) established the family of‘Cyclostomes’ for Myxine, lampreys and ammocetes (not knownat the time to be larvae of lampreys). He described the anatomyof the cyclostomes in some detail, but only mentioning a singlepair of gill openings in Myxine. Cloquet (1818) reviewed thecyclostomes, basing his work mainly on Dum�eril (1812).
Fig. 5. Capture localities of Rubicundus. * Rubicundus lopheliae, ▲ Rubicundus rubicundus, ♦ Rubicundus lakeside, ● Rubicundus eos
doi: 10.1111/jzs.12035© 2013 Blackwell Verlag GmbH
8 FERNHOLM, NOR�EN, KULLANDER, QUATTRINI, ZINTZEN, ROBERTS, MOK and KUO

However, at the end of his text, Cloquet referred to Home’s(1815) paper as having about the same content as that ofDum�eril (1812). Cloquet also provided a key to cyclostomes,which distinguishes between Myxine with two ventral gill open-ings, and Eptatr�eme with seven lateral gill openings. No speciesname was associated with Eptatr�eme, but the name doubtlesslyrefers to the description by Home (1815) and should be regardedas a French vernacular name.
Cloquet (1819) listed three generic names that apparently allrefer to the Eptatr�eme of 1818. Eptacitr�ete is listed only witha reference to Eptatr�eme, for which the entry reads ‘Eptatr�emeou Eptatr�ete’. In the paragraph explaining the etymology,eptatr�eme is explained followed by the remark that Dum�erilhad first proposed the word eptacitr�ete. Below that are twouses of Eptatr�ete, and one of Eptatretus, and a mention thatDum�eril established his genus Eptatr�eme in Paris. The entry issigned by Cloquet, but it is evident that he considers Dum�erilas author of Eptatr�eme and the proposed Eptacitr�ete. No realdistinction is made between Eptatretus, Eptatr�ete andEptatr�eme, but only Eptatretus appears in italics together witha specific epithet (Eptatretus Dombeii). The position of laterworkers, adopted also by us, has obviously been to considerEptatr�ete and Eptatr�eme as vernacular names, and Eptatretusas the intended scientific name, although we share Gill’s(1901) resignation over Cloquet’s confused text.
Cloquet (1819) based the diagnosis of Eptatretus on informa-tion about two species, viz., Gastrobranche dombey, describedby La Cep�ede (1798), and the subsequently described H. banksiiFleming (1822). La Cep�ede’s description and figure (1798: pl.25, fig. 1) were based on a single dry skin. The charactersdescribed and the drawing are clearly of a hagfish, but the num-ber of gill openings is not given, and gill openings are notshown on the drawing. In the introductory text, La Cep�ede(1798: 526) makes clear that he did not count the number of gillopenings, stating that ‘All p�etromyzons have seven gills on eachside, the blind gastrobranche [Myxine glutinosa] only has six onthe left and six on the right, and one should assume that the Gas-trobranche dombey does not have more’ (‘Tous les p�etromyzonsont sept branchies de chaque cot�e; le gastrobranche aveugle[Myxine glutinosa] n’en a que six �a droite et six �a gauche, et ilest a pr�esumer que le gastrobranche dombey n’en a pas un plusgrand nombre.’). This description does not contain any informa-tion that may enable identification as to genus or species, andthe specimen is apparently lost (Wisner and McMillan 1988).The type locality is given as the sea near Chili [= Chile]. Thecollector, Joseph Dombey, collected plants in Chile 1782–1784(Ruiz 1940).
The status of Gastrobranche dombey remains in limbo. Thereis no character known by which it can be identified to genus orspecies. Putnam (1874) listed Bdellostoma polytrema as a validspecies and added in a footnote that ‘This is unquestionably thespecies described by Lacep�ede [sic], under the name of Le Gas-trobranche Dombey, and figured from a stuffed skin of a speci-men collected by Dombey in Chili’. Later, many authors havecited Gastrobranche dombey as a scientific name, although itwas consistently presented by La Cep�ede as a French vernacularname. The earliest scientific name for La Cep�ede’s speciesappeared in 1804, but priority among the two descriptions isuncertain. Gasterobranchus dombeyi Shaw (1804: 265, pl. 134)is based entirely on La Cep�ede’s description, and the figure is acopy of La Cep�ede’s illustration. Latreille (1804: 75) lists thegenus Gastrobanchus [sic] and provides a Latinized scientificname for La Cep�ede’s species as ‘gastrobanc. dombey’, withclear indication to La Cep�ede’s description. Dates of publicationfor those two works are not known to us, but we follow Esch-
meyer (2012) who gives Shaw as author. Eptatretus dombeiiDum�eril (in Cloquet 1819) is a later independent Latinization.Cuvier (1816: 121) writes Gasterobranche dombey in italics, butapparently just cites the French name used by La Cep�ede.
Several post-1899 usages of Polistotrema dombey and Bdellos-toma dombeyi as valid names were listed among unassignablerecords of Chilean hagfishes by Wisner and McMillan (1988),and therefore Gasterobranchus dombeyi is available as potentialsenior synonym for one of several species of hagfish knownfrom the Pacific coast of Chile. Girard (1855a) describedB. polytrema from Chile and reviewed La Cep�ede’s descriptionof Gastrobranche dombey, coming to the conclusion that thecharacters did not support inclusion in Bdellostoma, diagnosedby presence of eyes and multiple gill openings. Putnam (1874)nevertheless considered Gasterobranche dombey and B. poly-trema to be the same species, but dismissed the first as a vernac-ular name. This was obviously not accepted by Jordan andEvermann (1896: 6) who used Putnam’s (1874) diagnosis ofB. polytrema as a diagnosis of Polistotrema dombey, to differen-tiate between this species and E. stoutii (Lockington, 1878). Put-nam’s B. polytrema was re-identified as E. bischhoffi (Schneider,1880) by Wisner and McMillan (1988). Wisner and McMillan(1988) reported three species of Eptatretus from the Chileancoast (E. bischhoffi, E. polytrema, and E. nanii Wisner andMcMillan, 1988). One of them is likely to be identical with LaCep�ede’s species, but we leave the synonymization to a futurerevision of the concerned species.
Homea banksii was known to Cloquet (1819) from thedescription by Home (1815) based on a dissected specimen‘brought from the South Seas by Sir Joseph Banks’. The descrip-tion by Fleming (1822) is apparently based only on Home’sdescription. M€uller (1836) described the species asBdellostoma heptatrema, again based apparently only on Home’sdescription, making B. heptatrema a junior synonym ofH. banksii. There is no record of extant specimens for thedescription of Petromyzon cirrhatus Forster, 1801 (in Schneider1801: 532), which was based on a manuscript description byForster [Johann Reinhold Forster]. Schneider (1801) gave thelocality as the coastal sea in New Zealand. M€uller (1836)renamed it Bdellostoma forsteri, and gave the locality as QueenCharlotte Sound, based on Forster’s manuscript. Forster’s manu-script was edited and published by Lichtenstein (1844), includinga long, detailed description of P. cirrhatus, with the localityDusky Bay and Queen Charlotte Sound in New Zealand. Banks’sspecimen(s) may have come from the first of Cook’s journeys tothe South Sea, in which Banks participated. Reinhold and GeorgForster participated in the second of Cook’s South Sea journeys,during which the expedition anchored in Dusky Bay in Marchand April 1773 and in Queen Charlotte Sound in May andNovember 1773, and in October 1774 (Lichtenstein 1844; For-ster 2007). It remains possible, however, that there was a speci-men passed from Forster to Banks, as some objects anddrawings were sold to Banks (Harpprecht 2007:19). It seems thatthe majority or all of the animals brought back by Cook’s threejourneys were in the possession of Banks for longer or shortertime, but became widely dispersed (Whitehead 1969). The speci-men(s) examined by Home may have come from any of the jour-neys and would have been available to him at the Huntermuseum, operated by his brother-in-law, or at the Royal Collegeof Surgeons, both of which, along with other institutions in Lon-don possessed specimens from Banks (Whitehead 1969). Fishspecimens from Cook’s journey still exist in the Natural HistoryMuseum in London (Whitehead 1969), but no myxinid speci-mens coming for sure from Forster or Banks are present today inthe Natural History Museum (James Maclaine, pers. comm.,
doi: 10.1111/jzs.12035© 2013 Blackwell Verlag GmbH
Hagfish phylogeny and taxonomy 9

Sept. 2012). An unpublished drawing of a P. cirrhatus, made byGeorg Forster is, however, preserved in the Natural HistoryMuseum (Whitehead 1978; Wheeler 1981). M€uller (1836) con-sidered Forster’s and Home’s species to be different. Laterauthors, however, have synonymized H. banksii with P. cirrha-tus, and it seems not impossible that the descriptions were basedon the same specimen. If this is the case, P. cirrhatus would bean objective junior synonym of H. banksii.
There are thus no type specimens known for H. banksii orP. cirrhatus, as would be desirable for establishing the relativestatus of these names, and for a stable nomenclature of the genusEptatretus. It also turns out that two similar species of Eptatr-etus, including E. cirrhatus occur sympatrically around the SouthIsland, probably including the type locality of P. cirrhatus. Thetaxonomy of these taxa will be addressed by Zintzen et al. (inprep.) who will designate a common neotype for H. banksii,B. heptatrema and P. cirrhatus. By being a replacement name,this neotype also becomes neotype of Bdellostoma forsteri.
Cloquet’s description of Eptatretus is based on a diagnosticcharacter (number of gill openings) not known from Gastrobran-che dombey, but only from Home’s description, which he citesat length, and apparently considers to be the same species. Thereis no doubt that Gastrobranche dombey, formally established asGasterobranchus dombeyi Shaw, is the type species of Eptatr-etus by monotypy (Code articles 67.2.1 and 68.3). However, asalready put forth by Gill (1901), it is also clear that Cloquet didnot base the genus on the characters of this species, but on themisidentified specimen later named H. banksii. Consequently,Code article 70.3 applies, and we designate H. banksii as typespecies of Eptatretus. We consider H. banksii and P. cirrhatusto be synonyms. The older name, P. cirrhatus, takes priority.
Gastrobranche dombey is also the nominal type species of Polis-totrema Gill (in Jordan & Gilbert, 1880). According to Eschmeyer(2012), the genus was based on misidentified E. stoutii andB. polytrema Girard in the synonymy of Gasterobranche dombey.The original description lists ‘Polistrotrema dombey (Muller) Gill’as a species occurring on the Pacific Coast of the United States,with reference to a footnote with the two names Bdellostoma poly-trema and B. stoutii. In Jordan and Gilbert (1881), it is obvious thatthe species they call Polistotrema dombeyi is rather, by a taxo-nomic conclusion, E. stoutii. Consequently Code article 70.3applies, and we select here B. stoutii as the type species of Polisto-trema, observing Code article 70.3.2.
Paramyxine and QuadratusParamyxine was introduced by Dean (1904) to distinguish theJapanese species Paramyxine atami Dean (1904) with crowdedgill openings from species of Eptatretus, in which the gill open-ings are more spaced. Subsequently, another 13 species havebeen referred to Paramyxine and four of them eventually toQuadratus. Molecular data failed to support monophyly of eitherParamyxine or Quadratus (Møller and Jones 2007; Kuo et al.2010; this study). The type species of Quadratus is Paramyxinetaiwanae (Shen & Tao, 1975), which is included in the 16S anal-yses, and nested among species of Eptatretus. Of the seven spe-cies of Eptatretus, labelled ‘P’ in Fig. 1, which possess crowdedgill openings and are referable to Paramyxine, one species posi-tions as sister group of remaining Eptatretus and the remainderform a monophyletic group within a clade composed of specieswith spaced gill openings and mixed with species that have beenreferred to Quadratus. Although Quadratus is clearly not distinctfrom Paramyxine, and all putative Paramyxine in this analysisshould be referred to Eptatretus, P. atami was not included, andthere remains a possibility that it is genetically distinct. If thatwas the case, Paramyxine would remain a valid genus, but with
a different composition and in need of a revised diagnosis. Rec-ognizing the six-species cluster of Paramyxine as a valid genusbased on the tree topology in Fig. 1 would require recognition ofseven or eight separate genera for the other clusters as well.There is no morphological support for such extensive splitting.
Rubicundus and RubicundinaeRubicundus lopheliae formed the sister clade to the remainingMyxinidae in the COI and combined analyses, while R. lopheliaeand R. rubicundus formed a monophyletic group in a polytomywith Myxininae and Eptatretinae in the 16S analysis. Both spe-cies were placed in Eptatretus because of their multiple gillopenings, and it was expected that they would group within orwith the Eptatretinae. However, if — as has been suggested byFernholm (1998) — having multiple pairs of gill openings witha gill opening adjacent to each gill pouch is the plesiomorphicstate, then that character is not useful for informing phylogeny.
Rubicundus lopheliae was sampled in the western NorthAtlantic off the south-eastern United States, and R. rubicundusin the western Pacific, off the north-eastern coast of Taiwan(Fig. 3). Although they come from very distant sites, R. lophe-liae and R. rubicundus formed a monophyletic group in thisstudy. Both species were found in deep waters (400 and 800 m),and have five pairs of gills. They share two unique morphologi-cal character states, currently considered as synapomorphies: anelongated nostril (Fig. 2) and the reddish colour. Another possi-ble synapomorphy is the localization of the slime pores in thegill area in a straight line with the gill openings (Fernholm andQuattrini 2008; Kuo et al. 2010).
Based on the results of the molecular phylogenetic analysesand the presence of distinctive morphological characters, we con-sider R. lopheliae and R. rubicundus to represent a distinct line-age of the Myxinidae, and place them in a separate genus andseparate subfamily. Two more species share the same snoutshape, reddish colour and five pairs of gills, and are also referredto Rubicundus. Both are known only from a single specimeneach, and no tissue suitable for molecular phylogenetic analysisis available. Rubicundus eos (Fernholm, 1991) was describedfrom one specimen caught in trawl at about 1000 m of depth inthe southern hemisphere in the Tasman Sea, west of NewZealand. Rubicundus lakeside was based on one specimen caughtin 762 m of depth off the Gal�apagos Archipelago (Mincaroneand McCosker 2004).
Fernholm and Quattrini (2008) discussed the possibility oferecting a new genus for the long-snouted, reddish deep-waterforms but refrained mainly because of the long geographicdistances between the type localities. Kuo et al. (2010) alsodiscussed the striking similarities between these deep-living hag-fish species occurring at long distances from each other.
The extremely wide and patchy distribution of Rubicundus(Fig. 5) is likely due to insufficient sampling. Rubicundus eos,R. lakeside and R. rubicundus are all described from single speci-mens, and R. lopheliae is known from only a few individuals.The latter was collected by manned submersibles in a cold-watercoral reef, and observations of live specimens within the reef indi-cate that this is their natural habitat (Fernholm and Quattrini2008). Additional specimens have later been collected from deepcoral areas off the south-east United States (S. W. Ross, unpub-lished data). Also R. lakeside was collected using mannedsubmersibles. R. eos and R. rubicundus on the other hand werecaught by trawl, which is not the best method to collect deep-living benthic hagfishes in complex habitats. We consider it likelythat what we see here is the emerging pattern of high degree ofendemism and speciation within complex deep-sea habitats (cf.Hubbs 1959; Hart and Pearson 2011). It is expected that with the
doi: 10.1111/jzs.12035© 2013 Blackwell Verlag GmbH
10 FERNHOLM, NOR�EN, KULLANDER, QUATTRINI, ZINTZEN, ROBERTS, MOK and KUO

on-going exploration of the deep sea using appropriate new col-lecting methods, we could find more hagfish species in these habi-tats. New technology to access deep-sea habitats was identified byEschmeyer et al. (2010) as the principal trigger for the presentoverall increase in descriptions of new marine fish species.
Eptatretus strickrotti was also caught in a deep-sea habitatfrom a hydrothermal vent (Møller and Jones 2007). However,although the live specimen was described as bright pink, its mor-phology is different from the species discussed in connectionwith Rubicundus, being a very slender species with 12 pairs ofgills and lacking an elongated nostril. Our molecular phyloge-netic analysis places it in Eptatretus (Fig. 1).
MyxineThe genus Myxine was erected by Linnæus (1758) with inclusionof the single species M. glutinosa. Four syntypes (NRM 89) arepreserved in the Swedish Museum of Natural History and pro-vide a stable reference for taxonomic work on the Myxininae.
When describing M. jespersenae, Møller et al. (2005) includeda comprehensive comparison and detailed discussion of all spe-cies of Myxine. They discussed whether M. glutinosa, from thenorth-east Atlantic, and M. limosa, from the north-west Atlantic,are conspecific or distinct species. Putnam (1874) synonymizedM. limosa with M. glutinosa. Following that work, recognitionof M. limosa has varied. Jordan and Evermann (1896) followedPutnam. Regan (1913) considered M. limosa to be valid, but alsodescribed another Western Atlantic species, Myxine atlantica.Wisner and McMillan (1995) synonymized M. atlantica withM. limosa, which they considered to be distinct from M. glutin-osa based on colour characters and larger size. Møller et al.(2005) stated that the holotype of M. limosa, from Bay of Fundy,was in poor condition and could not be used to investigate usefulcharacters, concluding that input from molecular data would benecessary to assess the validity of M. limosa. In all analyses,sequences from our specimens identified as M. limosa fromNorth Carolina form a monophyletic group that is clearly sepa-rate from M. glutinosa, supporting the validity of M. limosa andthe diagnosis provided by Wisner and McMillan (1995). Ourresults supported the monophyly of M. jespersenae, but only theCOI and combined analyses supported the monophyly of M. glu-tinosa.
Acknowledgements
We thank Bodil Kajrup (Swedish Museum of Natural History) for takingphotographs of hagfishes, Georg B. Friðriksson (Icelandic Museum ofNatural History) and Juan M. D�ıaz de Astarloa (Universidad Nacional deMar del Plata) for graciously making tissue samples available to us, Nic-olas Lartillot (Universit�e de Montreal) for advice on analytical interpreta-tion and Nicolas Bailly (FishBase) for his forceful encouragement. Wealso thank Steve W. Ross (University North Carolina Wilmington) forvaluable contributions. FishBase Sweden provided financial support.
References
Chen YW, Chang HW, Mok HK (2005) Phylogenetic position ofEptatretus chinensis (Myxinidae: Myxiniformes) inferred by 16S rRNAgene sequence and morphology. Zool Stud 44:111–118.
Cloquet H (1818) Cyclostomes. In: Dictionnaire des sciences naturelles,dans lequel on traite m�ethodiquement des diff�erens etres de la nature,consid�er�es soit en eux-memes, d’apr�es l’�etat actuel de nosconnoissances, soit relativement �a l’utilit�e qu’en peuvent retirer lam�edecine, l’agriculture, le commerce et les artes. Suivi d’une biographiedes plus c�el�ebres naturalistes. Tome 12. Levrault, Strasbourg, pp 301–304.
Cloquet H (1819) Eptatr�eme ou Eptatr�ete. In: Dictionnaire des sciencesnaturelles, dans lequel on traite m�ethodiquement des diff�erens etres de
la nature, consid�er�es soit en eux-memes, d’apr�es l’�etat actuel de nosconnoissances, soit relativement �a l’utilit�e qu’en peuvent retirer lam�edecine, l’agriculture, le commerce et les artes. Suivi d’unebiographie des plus c�el�ebres naturalistes. Tome 15. Levrault,Strasbourg, pp 134–136.
Cuvier G (1816) Le r�egne animal distribu�e d’apr�es son organisation, pourservir de base a l’histoire naturelle des animaux et d’introduction al’anatomie compar�ee. Tome II, contenant les reptiles, les poissons, lesmollusques et les ann�elides. Deterville, Paris.
Dean B (1904) Notes on Japanese myxinoids. A new genus, Paramyxine,and a new species Homea okinoseana. Reference also to their eggs. JColl Sci, Imperial Univ. Tokyo, Jap. 19:1–23.
Drummond AJ, Ashton B, Buxton S, Cheung M, Cooper A, Duran C,Field M, Heled J, Kearse M, Markowitz S, Moir R, Stones-Havas S,Sturrock S, Thierer T, Wilson A (2012) Geneious v5.6, Available athttp://www.geneious.com.
Dum�eril AMC (1812) Dissertation sur la famille des poissonsCyclostomes, pour d�emontrer leurs rapports avec les animaux sansvertebr�es; suivie d’un m�emoire sur l’anatomie des Lamproies. Facult�ede M�edecine, Paris.
Edgar RC (2004) MUSCLE: multiple sequence alignment with highaccuracy and high throughput. Nucleic Acids Res 32:1792–1797.
Eschmeyer WN (Ed.) (2012) Catalog of Fishes. California Academy ofSciences, http://research.calacademy.org/research/ichthyology/catalog/fishcatmain.asp. Electronic version accessed 2012-08-26.
Eschmeyer WN, Fricke R, Fong JD, Polack DA (2010) Marine fishdiversity: history of knowledge and discovery (Pisces). Zootaxa2525:19–50.
Fernholm B (1991) Eptatretus eos: a new species of hagfish (Myxinidae)from the Tasman Sea. Japan J Ichthyol 38:115–118.
Fernholm B (1998) Hagfish systematics. In: Jørgensen JM, Lomholt JP,Weber RE, Malte H (eds), Biology of Hagfishes. Chapman & Hall,London, pp 33–44.
Fernholm B, Quattrini AM (2008) A new species of hagfish (Myxinidae:Eptatretus) associated with deep-sea coral habitat in the western NorthAtlantic. Copeia 2008:126–132.
Fleming J (1822) The philosophy of Zoology; or a general view of thestructure, functions, and classification of animals. Archibald Constable& Co., Edinburgh.
Forster G (2007) Reise um die Welt- Illustriert von eigener Hand. Miteinem biographischen Essay von Klaus Haarpprecht und einemNachwort von Frank Vorpahl. 2nd edn. Eichborn, Frankfurt am Main.Edited reprint of Forster, G., Johann Reinhold Forster’s … Reise umdie Welt …, 2nd edn. 1784.
Froese R, Pauly D (Ed.) (2012) FishBase. World Wide Web electronicpublication. www.fishbase.org, version (08/2012). [Accessed 2012-04-19].
Gill TN (1901) The proper names of Bdellostoma or Heptatrema. ProcUS Natl Mus 23:735–738.
Girard CF (1855a) Abstract of a report to Lieut. Jas. M. Gilliss, U. S. N.,upon the fishes collected during the U. S. N. Astronomical Expeditionto Chili. Proc Acad Nat Sci Philadelphia 7:197–199.
Girard CF (1855b) Fishes. In: Contributions to the Fauna of Chile.Report to Lieut. James M. Gilliss, U. S. N., upon the fishes collectedby the U. S. Naval Astronomical Expedition to the southernhemisphere during the years 1849–50–51–52. Volume 2. SmithsonianInstitution, Washington, pp 230–253.
Goloboff PA, Farris JS, Nixon KC (2008) TNT, a free program forphylogenetic analysis. Cladistics 24:774–786.
Harpprecht K (2007) Georg Forster. Das Abenteyer der Freiheit und dieLiebe zur Welt. Ein biographischer Essay. In: Forster (2007), pp 7–36.
Hart PJB, Pearson E (2011) An application of the theory of islandbiogeography to fish speciation on seamounts. Mar Ecol Prog Ser430:281–288.
Home E (1815) On the structure of the organ of respiration in animalswhich appear to hold an intermediate place between those of the classPisces and the class Vermes, and in two genera of the last mentionedclass. Philos Trans R Soc Lond 105:256–264.
Hubbs CL (1959) Initial discoveries of fish faunas on seamounts andoffshore banks in the eastern Pacific. Pac Sci 13:311–316.
Huelsenbeck JP, Ronquist F (2001) MRBAYES: Bayesian inference ofphylogeny. Bioinformatics 17:754–755.
doi: 10.1111/jzs.12035© 2013 Blackwell Verlag GmbH
Hagfish phylogeny and taxonomy 11

International Commission on Zoological Nomenclature (1999)International Code of Zoological Nomenclature, 4th edn. InternationalTrust for Zoological Nomenclature, London.
Jordan DS, Evermann BW (1896) The fishes of North and MiddleAmerica: a descriptive catalogue of the species of fish-like vertebratesfound in the waters of North America, north of the Isthmus ofPanama. Part I. Bull US Natl Mus 47:i–lx + 1–1240.
Jordan DS, Gilbert CH (1881) List of the fishes of the Pacific coast ofthe United States, with a table showing the distribution of the species.Proc US Natl Mus 3:452–458.
Katoh K, Misawa K, Kuma K, Miyata T (2002) MAFFT: a novelmethod for rapid multiple sequence alignment based on fast Fouriertransform. Nucleic Acids Res 30:3059–3066.
Katoh K, Kuma K, Toh H, Miyata T (2005) MAFFT version 5:improvement in accuracy of multiple sequence alignment. NucleicAcids Res 33:511–518.
Kuo CH, Huang KF, Mok HK (1994) Hagfishes of Taiwan (I): ataxonomic revision with description of four new Paramyxine species.Zool Stud 33:126–139.
Kuo CH, Huang S, Lee SC (2003) Phylogeny of hagfish based on themitochondrial 16S rRNA gene. Mol Phylogenet Evol 28:448–457.
Kuo CH, Lee SC, Mok HK (2010) A new species of hagfish Eptatretusrubicundus (Myxinidae: Myxiniformes) from Taiwan, with reference toits phylogenetic position based on its mitochondrial DNA sequence.Zool Stud 49:855–864.
Kuraku S (2013) Impact of asymmetric gene repertoire betweencyclostomes and gnathostomes. Semin Cell Dev Biol 24:119–127.
La Cep�ede BG (1798) Histoire naturelle des poissons. Tome premier.Plassan, Paris.
Latreille PA (1804) Tableau m�ethodique des Poissons. In: NouveauDictionnaire d’Histoire naturelle, appliqu�ee aux arts, principalement �al’Agriculture et �a l’Economie rurale et domestique, 24. Deterville,Paris, pp 71–105.
Lichtenstein H (Ed.) (1844) Descriptiones animalium quae in itinere admaris australis terras per annos 1772 1773 et 1774 suscepto collegitobservavit et delineavit Ioannes Reinoldus Forster. Acad litter RBerolini.
Linnæus C (1758) Systema naturæ per regna tria naturæ, secundumclasses, ordines, genera, species, cum characteribus, differentiis,synonymis, locis. Tomus I. Editio decima, reformata. LaurentiusSalvius, Holmiæ.
McMillan CB (1999) Three new species of hagfish (Myxinidae,Eptatretus) from the Gal�apagos Islands. Fish Bull 97:110–117.
McMillan CB, Wisner RL (2004) Review of the hagfishes (Myxinidae,Myxiniformes) of the northwestern Pacific Ocean, with descriptions ofthree new species, Eptatretus fernholmi, Paramyxine moki, and P.walkeri. Zool Stud 43:51–73.
Mincarone MM, McCosker JE (2004) Eptatretus lakeside sp. nov., a newspecies of five-gilled hagfish (Myxinidae) from the Gal�apagos Islands.Proc Calif Acad Sci 55:162–168.
Mincarone MM, Stewart AL (2006) A new species of giant seven-gilledhagfish (Myxinidae: Eptatretus) from New Zealand. Copeia 2006:225–229.
Møller PR, Jones WJ (2007) Eptatretus strickrotti n. sp. (Myxinidae):first hagfish captured from a hydrothermal vent. Biol Bull 202:55–66.
Møller PR, Feld TK, Poulsen IH, Thomsen PF, Thormar JG (2005)Myxine jespersenae, a new species of hagfish (Myxiniformes:Myxinidae) from the North Atlantic Ocean. Copeia 2005:374–385.
M€uller J (1836) Vergleichende Anatomie der Myxinoiden, derCyclostomen mit durchbohrtem Gaumen. Erster Theil. Osteologie undMyologie. Abhandl K Akad Wiss Berlin, Phys-mathem Kl 1834:65–340.
Nelson JS (2006) Fishes of the World. John Wiley and Sons, Inc.,Hoboken, New Jersey.
Palumbi SR, Martin AP, Romano S, McMillan WO (1991) The SimpleFool’s Guide to PCR. University of Hawaii, Honolulu.
Posada D, Crandall KA (1998) Modeltest: testing the model of DNAsubstitution. Bioinformatics 14:817–818.
Putnam FW (1874) Notes on Myxinidae. Notes on the genus Myxine.Proc Boston Soc Nat Hist 16:127–135.
Regan CT (1913) A revision of the myxinoids of the genus Myxine. AnnMag Nat Hist 8:395–398.
Ronquist F, Teslenko M, van der Mark P, Ayres DL, Darling A, H€ohnaS, Larget B, Liu L, Suchard MA, Huelsenbeck JP (2012) MrBayes3.2: Efficient Bayesian phylogenetic inference and model choice acrossa large model space. Syst Biol 61:539–542.
Ruiz H (1940) Travels of Ruiz, Pav�on, and Dombey in Peru and Chile(1777–1788). Bot Ser, Field Mus Nat Hist 21:1–372.
Schneider JG (1801) M. E. Blochii systema ichthyologiae. Published bythe author, Berolini.
Shaw G (1804) General Zoology or Systematic Natural History, vol. 5,part 2, Pisces: G. Kearsley, London.
Strahan R (1975) Eptatretus longipinnis, n. sp., a new hagfish (familyEptatretidae) from South Australia, with a key to the 5–7 gilledEptatretidae. Aust Zool 18:137–148.
W€agele JW, Mayer C (2007) Visualizing differences in phylogeneticinformation content of alignments and distinction of three classes oflong-branch effects. BMC Evol Biol 7:147.
Ward RD, Zemlak TS, Innes BH, Last PR, Hebert PD (2005) DNABarcoding of Australia’s fish species. Philos Trans R Soc Lond B BiolSci 360:1847–1857.
Wheeler A (1981) The Forster’s fishes. In: Cook J, The journal of H. M.S. Resolution 1772–1775. Genesis Publications, Guilford, pp 781–803.
Whitehead PJP (1969) Zoological specimens from Captain Cook’svoyages. J Soc Biblphy Nat Hist 5:161–201.
Whitehead PJP (1978) The Forster collection of zoological drawings inthe British Museum. Bull Br Mus, (Nat Hist) Hist Ser 6:25–47.
Wisner RL (1999) Descriptions of two new subfamilies and a new genusof hagfishes (Cyclostomata: Myxinidae). Zool Stud 38:307–313.
Wisner RL, McMillan CB (1988) A new species of hagfish, genusEptatretus (Cyclostomata, Myxinidae), from the Pacific Ocean nearValparaiso, Chile, with new data on E. bischoffii and E. polytrema.Trans San Diego Soc Nat Hist 21:227–244.
Wisner RL, McMillan CB (1995) Review of new world hagfishes of thegenus Myxine (Agnatha, Myxinidae) with descriptions of nine newspecies. Fish Bull 93:530–550.
Zintzen V, Roberts CD, Anderson MJ, Stewart AL, Struthers CD,Harvey ES (2011) Hagfish predatory behaviour and slime defencemechanism. Sci Rep 1:1–6.
Supporting InformationAdditional Supporting Information may be found in the onlineversion of this article:Figure S1. Majority-rule parsimony tree from analysis of 16S
data (software TNT, 1000 replicates, 10 random additions per rep-licate, using “classic search” and TBR branch swapping.Figure S2. Majority-rule parsimony tree from analysis of COI
data (software TNT, 1000 replicates, 10 random additions per rep-licate, using “classic search” and TBR branch swapping. End gapswere coded as missing data).Figure S3. Majority-rule parsimony tree from combined analy-
sis of 16S and COI data (software TNT, 1000 replicates, 10 ran-dom additions per replicate, using “classic search” and TBRbranch swapping.Figure S4. Same as Fig. 1, but monophyletic species have not
been collapsed to a single terminal.Figure S5. Same as Fig. 2, but monophyletic species have not
been collapsed to a single terminal.Figure S6. Same as Fig. 3, but monophyletic species have not
been collapsed to a single terminal.
doi: 10.1111/jzs.12035© 2013 Blackwell Verlag GmbH
12 FERNHOLM, NOR�EN, KULLANDER, QUATTRINI, ZINTZEN, ROBERTS, MOK and KUO
Copyright © 2022 FDOKUMEN




















