
Evolution of the Sele River coastal plain (southern Italy) during the Late Quaternary by inland and...
24
1 23 Rendiconti Lincei SCIENZE FISICHE E NATURALI ISSN 2037-4631 Rend. Fis. Acc. Lincei DOI 10.1007/s12210-012-0165-5 Evolution of the Sele River coastal plain (southern Italy) during the Late Quaternary by inland and offshore stratigraphical analyses Pietro P. C. Aucelli, Vincenzo Amato, Francesca Budillon, Maria Rosaria Senatore, Sabrina Amodio, Carmine D’Amico, Simone Da Prato, et al.
Transcript of Evolution of the Sele River coastal plain (southern Italy) during the Late Quaternary by inland and...

1 23
Rendiconti LinceiSCIENZE FISICHE E NATURALI ISSN 2037-4631 Rend. Fis. Acc. LinceiDOI 10.1007/s12210-012-0165-5
Evolution of the Sele River coastalplain (southern Italy) during the LateQuaternary by inland and offshorestratigraphical analyses
Pietro P. C. Aucelli, Vincenzo Amato,Francesca Budillon, Maria RosariaSenatore, Sabrina Amodio, CarmineD’Amico, Simone Da Prato, et al.

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag. This e-offprint is for personal use only
and shall not be self-archived in electronic
repositories. If you wish to self-archive your
work, please use the accepted author’s
version for posting to your own website or
your institution’s repository. You may further
deposit the accepted author’s version on a
funder’s repository at a funder’s request,
provided it is not made publicly available until
12 months after publication.

LAND SEA INTERACTION IN CAMPANIA (ITALY)
Evolution of the Sele River coastal plain (southern Italy)during the Late Quaternary by inland and offshorestratigraphical analyses
Pietro P. C. Aucelli • Vincenzo Amato • Francesca Budillon • Maria Rosaria Senatore •
Sabrina Amodio • Carmine D’Amico • Simone Da Prato • Luciana Ferraro •
Gerardo Pappone • Elda Russo Ermolli
Received: 19 November 2011 / Accepted: 11 January 2012
� Springer-Verlag 2012
Abstract The late Quaternary evolution of the Sele River
coastal plain (Salerno Gulf, southern Italy) was investi-
gated through integrated stratigraphical, chronological and
palaeoecological analyses. The main environmental chan-
ges were ascribed to glacio-eustatic variations leading to
rapid ingressions alternating with coastal progradations.
The marked marine ingression of MIS 5.5 is testified by
palaeoridges now cropping out 4 km inland at 11/13 m
a.s.l. (Gromola palaeoridge). The eustatic minimum of MIS
2 is testified by lower shoreface deposits in the offshore
core record and in the seismic profiles at 120/130 m below
sea level. This prolonged sea-level fall was interrupted by
at least three rapid sea-level rises, probably related to MIS
5.3, 5.1 and 3. The evidence of the first two sea level rises
are represented by shoreface deposits in the inland S1 core
(30 m thick, 3 m a.s.l., 1.5 km inland). The highstand of
MIS 3 was identified by seismic profiles as onlapping
marine deposits. The shore deposits at 100 m b.s.l were
tentatively attributed to the lowstand of MIS 4. After the
lowstand of MIS 2, the Sele Plain was newly flooded due to
the rapid Post Glacial sea-level rise. This ingression caused
the inland migration of a barrier-lagoon system and stop-
ped at approximately 5.5 ky BP. From that moment the
shoreline started prograding up to the present position
probably due to the decrease in the sea-level rise rates and
This paper is an outcome of the FISR project VECTOR (Vulnerability
of the Italian Coastal Area and Marine Ecosystem to Climate changes
and their role in the Mediterranean carbon cycles), subproject
VULCOST (VULnerability of COaSTal environments to climate
changes) on: ‘‘Land–sea interaction and costal changes in the Sele
River plain, Campania’’.
P. P. C. Aucelli � S. Amodio � G. Pappone
Dipartimento di Scienze per l’Ambiente, Universita di Napoli
Parthenope, Centro direzionale, isola c4, 80143 Naples, Italy
e-mail: [email protected]
S. Amodio
e-mail: [email protected]
G. Pappone
e-mail: [email protected]
V. Amato (&) � C. D’Amico
Dipartimento di Scienze e Tecnologie per l’Ambiente e il
Territorio, Universita del Molise, C.da Fonte Lappone,
86090 Pesche (IS), Italy
e-mail: [email protected]
C. D’Amico
e-mail: [email protected]
F. Budillon � L. Ferraro
Istituto per l’Ambiente Marino Costiero, CNR, Calata Porta di
Massa, 80133 Naples, Italy
e-mail: [email protected]
L. Ferraro
e-mail: [email protected]
M. R. Senatore
Dipartimento di Scienze per la Biologia, la Geologia e
l’Ambiente, Universita degli Studi del Sannio,
Via Dei Mulini 59A, 82100 Benevento, Italy
e-mail: [email protected]
S. Da Prato
Istituto di Geoscienze e Georisorse, CNR, Via Moruzzi 1,
56124 Pisa, Italy
e-mail: [email protected]
E. Russo Ermolli
Dipartimento ARBOPAVE, Universita degli Studi ‘‘Federico II’’
di Napoli, Via Universita 100, 80055 Portici, Italy
e-mail: [email protected]
123
Rend. Fis. Acc. Lincei
DOI 10.1007/s12210-012-0165-5
Author's personal copy

to the volcaniclastic supplies from the Neapolitan volca-
noes, especially from the AD 79 Vesuvius eruption, also
recorded in the subbottom chirp profiles.
Keywords Late Quaternary � Sele Plain � Sea-level
changes � Facies analysis � Seismostratigraphy �Land–sea correlation
1 Introduction
Sea-level changes represent a significant factor in con-
trolling the evolution of coastal environments over geo-
logical times. Reconstructing palaeo-sea levels has been
attracting increasing interest in the last years due to the
deep impact of global warming on people living along the
coasts (Lambeck et al. 2011; IPCC 2007; Rahmstorf
2007; Siddall et al. 2006; Pirazzoli 1996). For this reason,
relatively tectonically stable areas are the best places for
this kind of research. In these areas, biological and sed-
imentological markers of the Last Interglacial (Marine
Isotopic Stage 5.5, hereinafter MIS 5.5) and Holocene
(MIS 1) sea level highstand phases are clearly recogni-
zable inland (Orru et al. 2011; Coltorti et al. 2010;
Ferranti et al. 2006; Antonioli et al. 2004; Lambeck et al.
2004; Antonioli et al. 1999), while markers of the low-
stand phases (MIS 2 and 4) can be recognized both off-
shore and inland at a considerable depth from the present
ground level (Spampinato et al. 2011; Caruso et al. 2011;
Wheatcroft and Drake 2003; Waelbroeck et al. 2002; Plint
and Nummedal 2000).
The Sele Plain/Salerno Gulf half-graben represents an
excellent area for integrated studies on the Quaternary
stratigraphic records. In fact the studies on the late Qua-
ternary morpho-stratigraphy allowed the palaeobeach
deposits of the Last Interglacial period to be recognized at
ca. 4 km inland, ca. 11/13 m a.s.l. (Brancaccio et al. 1987;
Russo and Belluomini 1992), covered by dune sands of the
Gromola-S.Cecilia-Arenosola-Aversana palaeoridges
(hereinafter GP). Moreover, the beach deposits of the
Holocene highstand are found at ca. 1.5 km inland, covered
by dune sands of the Laura palaeoridge (Brancaccio et al.
1988; ISPRA 2009; Amato et al. 2011). Other beach
deposits were recognized between 0.8 km inland and the
present shoreline, covered by dune sands of the Sterpina
palaeoridges (Brancaccio et al. 1988; ISPRA 2009; Amato
et al. 2011). The offshore sector of the Salerno Gulf shows
a wide continental shelf extending up to 15–25 km from
the present coast. Here shoreface deposits, recording the
eustatic minimum of the Last Glacial Maximum, were
found offshore in seismic and core data at ca. 8–10 km
from the present-day shoreline and at ca 110/120 m below
s.l. (Budillon et al. 1994).
In order to reconstruct the variations of the coastal and
marine palaeoenvironments due to the palaeoclimatic
changes between MIS 5 and MIS 1, a land–sea integrated
stratigraphic study was carried out, combining information
from three seismic profiles interpreted on the base of two
gravity cores (C1213 and C101), with a 30-m deep core
(S1) drilled inland at 1.5 km from the present coast. Facies
analysis was applied to all cores, also supported by pal-
aeoecological data, sequence-stratigraphic interpretation
and tephro-chronologic and 14C age constraints.
2 The inland and offshore geologic and geomorphologic
setting
The alluvial-coastal plain of the Sele River displays up to
2,400 m of post-orogenic sediment infill, accumulated in a
coastal half-graben, extending offshore in the deep Salerno
Gulf (ISPRA 2009 with references). This tectonic depres-
sion is bounded to N-NE by the horsts of the Lattari and
Picentini mountains and to the S-SE by the Cilento
mountains (Fig. 1). The submerged topography of the
Salerno Gulf displays an asymmetrical shape: a narrow
shelf domain develops down to 120–130 m around the
large submarine Salerno Valley, whereas a shelf 25 km
large and 180 m deep, surrounds the Cilento shore south of
the Sele River mouth, controlled by shallow rocky out-
crops. This physiography largely reflects the structural
configuration, mainly controlled by the Capri Master Fault
to the north (CMF in Fig. 1) and the Sele line to the south.
The Sele Plain–Salerno Gulf half-graben was charac-
terized by extensional tectonics since the Late Miocene–
Lower Pliocene (Brancaccio et al. 1987; Ortolani et al.
1979), along NW–SE and NE–SW faults. The collapse is
evidenced by the deposition of the so-called Conglomerati
di Eboli, a thick and widespread clastic unit (Early Pleis-
tocene in age). The latter is in part deeply buried in the
external sector of the plain and in the offshore sector, and
in part is largely exposed inland, along the NW margin of
the plain (Fig. 1). At the beginning of Middle Pleistocene,
the geomorphology of the plain was re-shaped by normal
and transtensional faults (Fig. 1). Part of the area carrying
Early Pleistocene fanglomerates was uplifted, while a large
part of the Plain continued to subside, accommodating the
so-called Battipaglia-Persano Supersynthem (BP in Fig. 1;
ISPRA 2009). This unit, which is up to some hundred
meters thick, covers a large part of the most internal sector
of the plain forming wide depositional terraces lying at
16–18 m a.s.l., close to the modern coast, and at 100 m
a.s.l. or more close to the mountain foot. Thickness and
facies distribution of the supersynthem’s lower part suggest
that its deposition was accompanied by subsidence and by
NW-ward tilting (ISPRA 2009).
Rend. Fis. Acc. Lincei
123
Author's personal copy
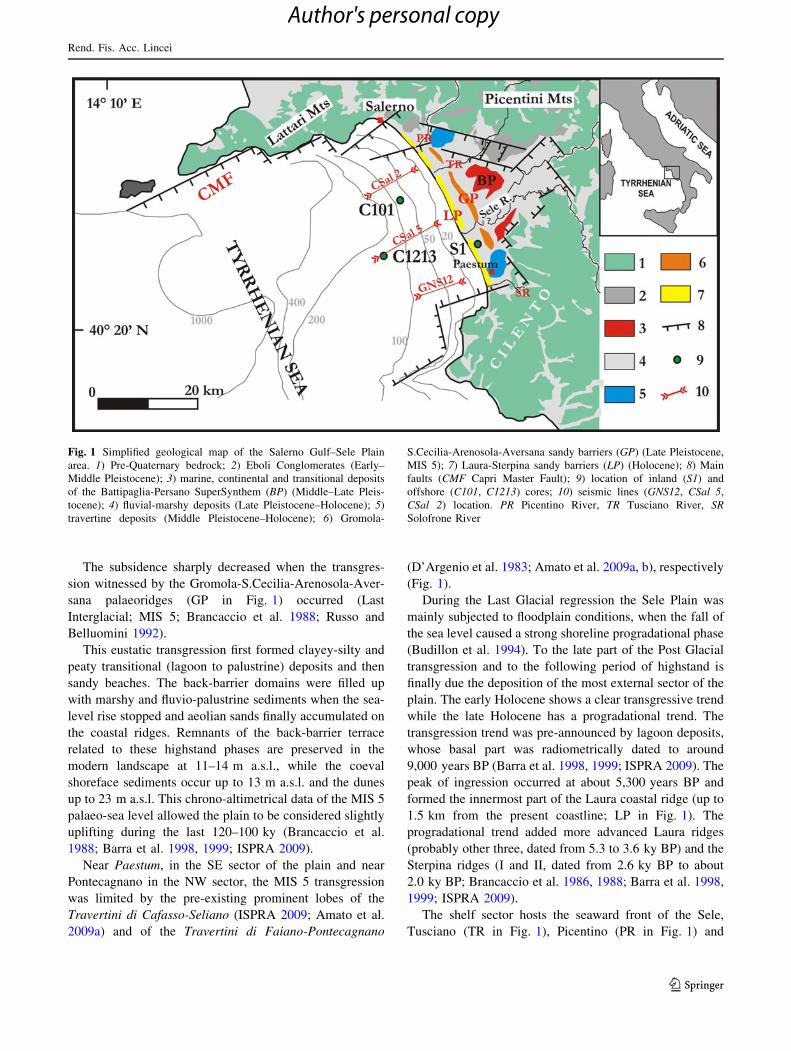
The subsidence sharply decreased when the transgres-
sion witnessed by the Gromola-S.Cecilia-Arenosola-Aver-
sana palaeoridges (GP in Fig. 1) occurred (Last
Interglacial; MIS 5; Brancaccio et al. 1988; Russo and
Belluomini 1992).
This eustatic transgression first formed clayey-silty and
peaty transitional (lagoon to palustrine) deposits and then
sandy beaches. The back-barrier domains were filled up
with marshy and fluvio-palustrine sediments when the sea-
level rise stopped and aeolian sands finally accumulated on
the coastal ridges. Remnants of the back-barrier terrace
related to these highstand phases are preserved in the
modern landscape at 11–14 m a.s.l., while the coeval
shoreface sediments occur up to 13 m a.s.l. and the dunes
up to 23 m a.s.l. This chrono-altimetrical data of the MIS 5
palaeo-sea level allowed the plain to be considered slightly
uplifting during the last 120–100 ky (Brancaccio et al.
1988; Barra et al. 1998, 1999; ISPRA 2009).
Near Paestum, in the SE sector of the plain and near
Pontecagnano in the NW sector, the MIS 5 transgression
was limited by the pre-existing prominent lobes of the
Travertini di Cafasso-Seliano (ISPRA 2009; Amato et al.
2009a) and of the Travertini di Faiano-Pontecagnano
(D’Argenio et al. 1983; Amato et al. 2009a, b), respectively
(Fig. 1).
During the Last Glacial regression the Sele Plain was
mainly subjected to floodplain conditions, when the fall of
the sea level caused a strong shoreline progradational phase
(Budillon et al. 1994). To the late part of the Post Glacial
transgression and to the following period of highstand is
finally due the deposition of the most external sector of the
plain. The early Holocene shows a clear transgressive trend
while the late Holocene has a progradational trend. The
transgression trend was pre-announced by lagoon deposits,
whose basal part was radiometrically dated to around
9,000 years BP (Barra et al. 1998, 1999; ISPRA 2009). The
peak of ingression occurred at about 5,300 years BP and
formed the innermost part of the Laura coastal ridge (up to
1.5 km from the present coastline; LP in Fig. 1). The
progradational trend added more advanced Laura ridges
(probably other three, dated from 5.3 to 3.6 ky BP) and the
Sterpina ridges (I and II, dated from 2.6 ky BP to about
2.0 ky BP; Brancaccio et al. 1986, 1988; Barra et al. 1998,
1999; ISPRA 2009).
The shelf sector hosts the seaward front of the Sele,
Tusciano (TR in Fig. 1), Picentino (PR in Fig. 1) and
Fig. 1 Simplified geological map of the Salerno Gulf–Sele Plain
area. 1) Pre-Quaternary bedrock; 2) Eboli Conglomerates (Early–
Middle Pleistocene); 3) marine, continental and transitional deposits
of the Battipaglia-Persano SuperSynthem (BP) (Middle–Late Pleis-
tocene); 4) fluvial-marshy deposits (Late Pleistocene–Holocene); 5)
travertine deposits (Middle Pleistocene–Holocene); 6) Gromola-
S.Cecilia-Arenosola-Aversana sandy barriers (GP) (Late Pleistocene,
MIS 5); 7) Laura-Sterpina sandy barriers (LP) (Holocene); 8) Main
faults (CMF Capri Master Fault); 9) location of inland (S1) and
offshore (C101, C1213) cores; 10) seismic lines (GNS12, CSal 5,
CSal 2) location. PR Picentino River, TR Tusciano River, SRSolofrone River
Rend. Fis. Acc. Lincei
123
Author's personal copy

Solofrone-Capo di Fiume (SR in Fig. 1) rivers’ alluvial
deposits. Coastal and alluvial domains prograded 15 km
seawards starting from MIS 5, in step with the general
retreat of the sea level during the Last Glacial Maximum
(MIS 2). Successively the shelf underwent drowning,
following the Late Pleistocene–Holocene transgression
(Budillon et al. 1994; Buccheri et al. 2002; Iorio et al.
2009; Sacchi et al. 2009). Consequently, the stratigraphic
pattern of the Sele River plain consists of a thick Late
Pleistocene prograding wedge topped by a marked
erosional unconformity, which, in turn, underlies the
lithosomes that were formed between 18 and 5 ky. The late
Holocene depositional unit consists of a wedge whose
thickness decreases toward the shelf edge. The most
impressive late Holocene event was caused by the AD 79
Vesuvius eruption (Lirer et al. 1973; Buccheri et al. 1994;
Sacchi et al. 2005; Insinga et al. 2008), that settled a thick
pumiceous lapilli layer, which now provides a well rec-
ognizable stratigraphic marker within the shelf and the
upper slope sediments.
3 Materials and methods
3.1 Inland core
A new deep core (30 m thick) was drilled in the inner part
of the Laura palaeoridge (40�27.3230N, 14�38.1710E,
2.85 m a.s.l., S1 in Fig. 1), using a dry-continuous
mechanic coring that allowed a 10 cm diameter core to be
extracted. The core was preserved in six coring-box, now
kept at IAMC-CNR Naples warehouses. Here, the core was
analyzed to define color, texture, grains size, shape and
composition, fossil content, sedimentary and diagenetic
structures. All these features were taken into account to
define lithofacies and their association, according to the
Unconformity Boundary Stratigraphic Unit method
(UBSU, after Salvador 1994). The most significant layers
were sampled for laboratory analyses, such as palaeoe-
cology (mollusk, ostracoda and benthic foraminifera
assemblages) palynology and 14C AMS datings, in order to
interpret at the best, the lithofacies organizations and their
chronology, and finally, the original depositional environ-
ments (Fig. 2).
3.2 Mollusks
Qualitative and quantitative malacological analyses were
carried out on 58 samples of the S1 core. Shells of each
sample were identified and counted. Taxonomy of the
species follows both the World Register of Marine Species
(WoRMS) database for marine and brackish taxa (accessed
at http://www.marinespecies.org on 2011-06-27) and the
Italian Ministry of Environment’s Checklist of the Italian
Fauna (accessed at http://www.faunaitalia.it/checklist/ on
2011-06-27) for non-marine taxa. Ecology of marine and
brackish species was defined after Peres and Picard (1964)
and Peres (1982); while for non-marine species we follow
Kerney (1999). A malacological diagram (Fig. 3) based on
numerical data was essayed; the diagram deals with the
number of shells related to species recovered from each
sample. As the different samples recorded a great vari-
ability in the total number of shells, ranging from more
than 1,400 to less than 5, percentages of species are not
considered for the construction of the diagram, that allows
to recognize several mollusk zones (related to different
environments), in accordance with the major faunal chan-
ges through the sequence.
3.3 Benthic foraminifera
The foraminiferal analysis was carried out on a total of 20
samples collected at approximately 100 cm intervals of the
S1 core (Fig. 2). Almost 300 g of sediment was wet-sieved
through 125 lm, dried at 60�C and then weighed. In the
present study data from [125 lm size fraction were ana-
lyzed. When abundant, the sediments were split by a
microsplitter in small portions for counting foraminifera.
Species determination was mainly based on studies con-
cerning the Mediterranean benthic fauna (Sgarrella and
Moncharmont Zei 1993; Fiorini and Vaiani 2001). For the
genus Ammonia, we referred to Carboni and Di Bella
(1996). Because of the general scarcity of foraminifera,
only qualitative analysis was performed on benthic species.
3.4 Ostracods
Thirty-seven samples were prepared for ostracod analysis
of the S1 core (Fig. 2). Samples were disaggregated in
warm water (95�C) adding hydrogen peroxide, washed
through a 63-lm sieve, and finally dried at 110�C. In order
to facilitate palaeoecologic interpretation, semi-quantita-
tive analyses were performed, including both juvenile and
adult specimens. Ostracod frequencies were calculated for
200 g of dried sediment. The analysis of the population
structure of each species was performed in order to
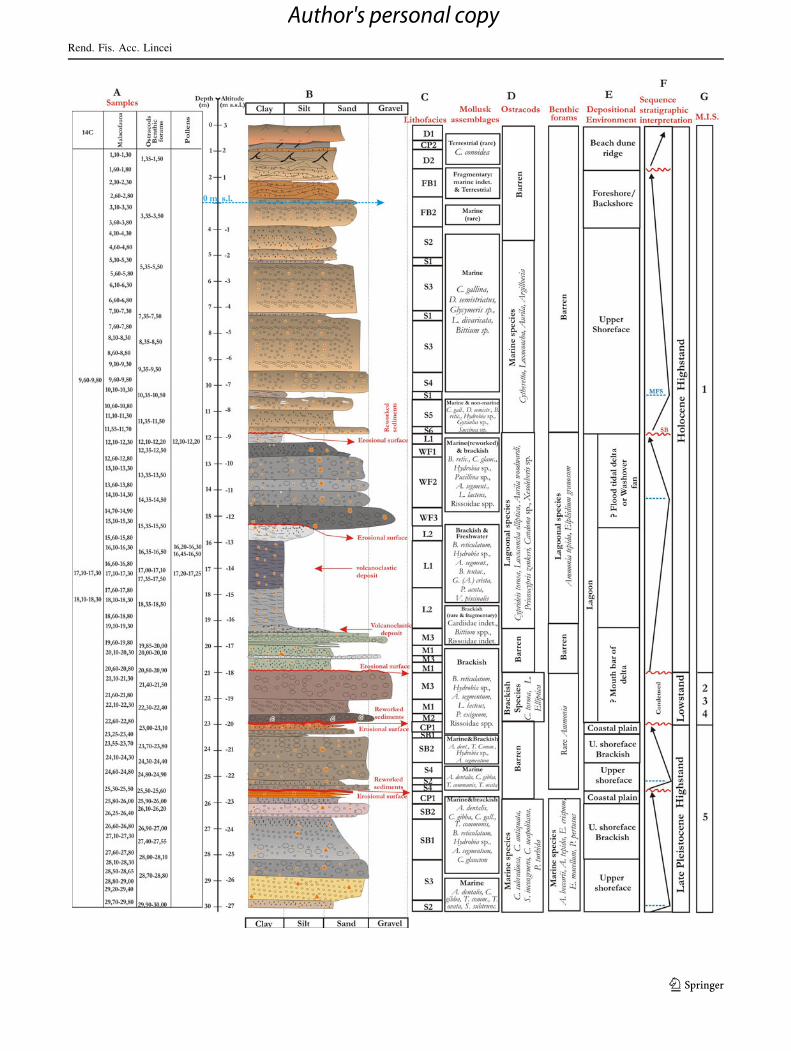
Fig. 2 Sedimentological log of the inland core S1. Columns show
from the left to the right: (a) selected samples used for 14C datings and
palaeoecological analyses (mollusks, benthic foraminifera, ostracods,
pollens); (b) grain size log; (c) lithofacies, as indicated in Table 1,
where their detailed descriptions are shown; (d) vertical distribution
of the main mollusk, ostracod and benthic foraminifera assemblages;
(e) depositional environments, also suggested by vertical evolution of
lithofacies; (f) systems-tract interpretation as suggested by retrogra-
dational and progradational facies trends (SB sequence boundary,
MFS maximum flooding surface); (g) marine isotope stages (MIS)
interpretation. See details in the text of the inland data
c
Rend. Fis. Acc. Lincei
123
Author's personal copy
Rend. Fis. Acc. Lincei
123
Author's personal copy
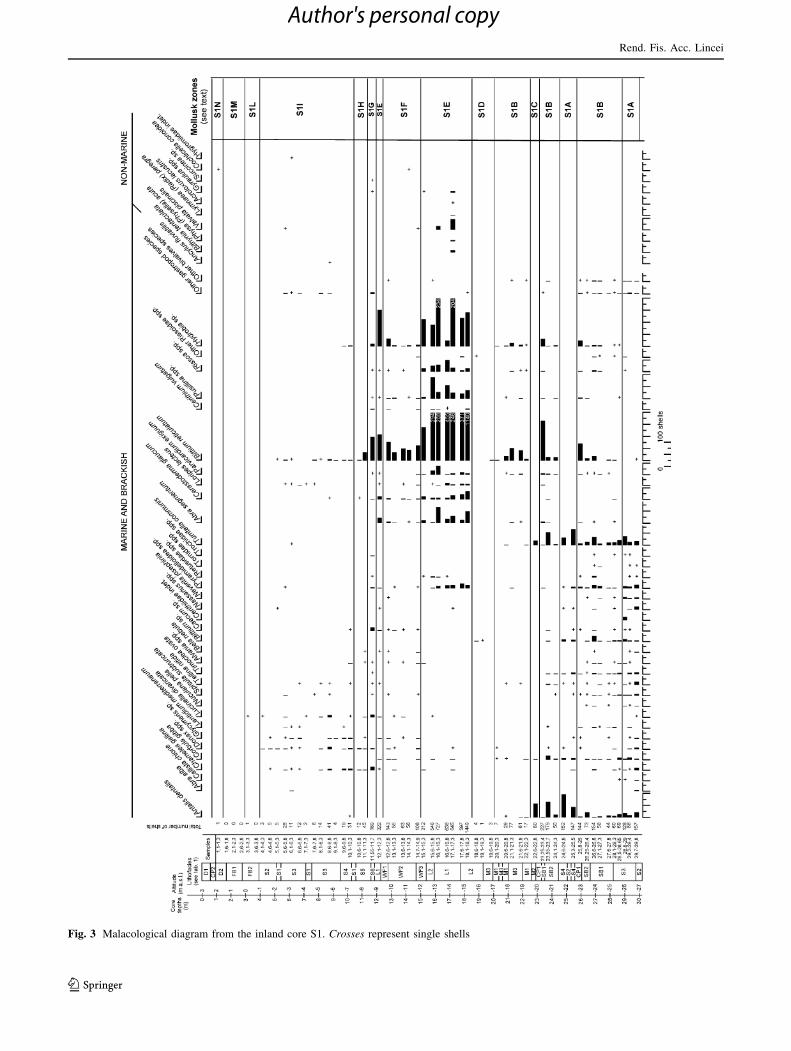
Fig. 3 Malacological diagram from the inland core S1. Crosses represent single shells
Rend. Fis. Acc. Lincei
123
Author's personal copy

separate autochthonous from displaced species (Gliozzi
2000). The autoecological data of each species and their
interpretation were performed following different authors
(e.g. Colalongo 1969; Athersuch et al. 1989; Montenegro
and Pugliese 1995; Gliozzi and Mazzini 1998; Meisch
2000).
3.5 Pollen
Only four samples were collected for pollen analysis of the
S1 core, due to unsuitable lithology (Figs. 2, 4). For each
sample, 10 g of sediment were treated with HCl and HF for
mineral dissolution. Physical enrichment procedures, such
as ZnCl2 separation and ultrasound sieving, were realized
in order to concentrate the pollen grains in the residue. The
most peaty samples were boiled in KOH 10% before being
processed. Quantitative pollen analysis was only possible
on three core samples, since the sample at -12.20 m was
barren. Ca. 300 pollen grains were counted in each sample
and 42 taxa were identified. The main sum used for the
calculation of arboreal (AP) and nonarboreal pollen (NAP)
percentages excluded marsh and water plants (including
Typha and Potamogeton), spores, algae and indeterminate
grains.
3.6 Offshore cores
The gravity core C1213 was collected in 2003 at a depth of
152 m (40�26.1330N, 14�45.9500E) (Fig. 1) and about
4.7 m of marine sediment were retrieved. The core was cut
in 1 m-long sections and split in two halves in order to
allow visual description and sediment physical measure-
ments at IAMC-CNR laboratories.
The gravity core C101 was collected in 1984 at a depth
of 87 m (40�33.0670N; 14�47.1660E) (Fig. 1) and entered
the marine sediment within the shelf down to 4.30 m bsf.
3.7 Seismic data
The subbottom profiles were acquired in 2002 using a
Chirp Cap II profiler on board of the R/V Urania, during
the acquisition of data for a geologic cartographic project
by IAMC-CNR (Fig. 1). Chirp seismic system in based on
a frequency-modulated source (FM), pinging within the
2–7 kHz band. The single channel Uniboom profiles were
shot by a EG&G mod. 230 (PSU230) Power Supply, each
150 ms at 300 J, on board of R/V Bannock during a cruise
carried out by the Earth Science Department of Naples
University in 1984.
The conversion of two-way travel time to real depth was
obtained assuming an average velocity of about
1,550 m s-1 below the sea floor (Carlson et al. 1986). The
vessel positioning was achieved by a 12-channel DGPS, a
motion sensor and a gyrocompass in order to perform a
real-time correction during the 2002 survey while a Loran
C positioning was used during the acquisition of Uniboom
profiles in 1984. Subbottom profiles investigated the shelf
record by about 60 ms two-way travel time, c. 50 m
beneath the sea floor. Maximum vertical resolution does
not exceed 30–40 cm. The Uniboom signal penetrated the
Fig. 4 Pollen diagram from core S1. Taxa percentages are plotted
against depth
Rend. Fis. Acc. Lincei
123
Author's personal copy

first 80 ms two-way travel time, c. 60 m bsf, and achieved
a vertical resolution of about 70 cm.
Seismic lines were interpreted considering reflection
geometries, relative lateral terminations among reflectors
and seismic facies analysis, according to standard seismo-
stratigraphic, echo facies interpretation (Damuth 1980;
Vail 1987) and sequence-stratigraphic criteria on conti-
nental margins (Posamentier and Allen 1999; Plint and
Nummedal 2000).
3.8 Radiometric datings
An age-depth model of the S1 drilled succession was built
thanks to radiometric dating of three biological sea-level
markers: Cerithium vulgatum, Cerastoderma glaucum and
Donax trunculus. The shells were analyzed at the CIRCE-
DSA-SUN laboratory (Center for Isotopic Research on
Cultural and Environmental heritage, Environmental Sci-
ence Department, Second University of Naples). 14C ages
were calibrated using the Calibration data set, intcal09.14c
of Reimer et al. (2009).
The age model of the C101 core is based on AMS 14C
dating performed at CIRCE-DSA-SUN Laboratory on
mixed planktonic foraminifera shells from four collected
samples. Radiocarbon ages were converted into calendar
ages through intracal09.14c (Reimer et al. 2009). The fall
deposits of the 79 AD Vesuvius eruption were considered
as chronological reference.
4 Results
4.1 The S1 core
4.1.1 Mollusks
A total of 101 mollusk taxa were recovered, including 33
bivalves, 66 gastropods and 2 scaphopods. All the identi-
fied species were previously reported for modern non-
marine, marine and lagoonal settings of the Mediterranean
area. Figure 3 provides a malacological diagram divided in
the following mollusk zones.
• Zone S1A (from -29.7/-29.8 to -28.8/-29 m; from
-25.3/-25.5 to -24.6/-24.8 m): polytypic marine
assemblages consisting mainly of Antalis dentalis,
Chamelea gallina, Corbula gibba, Donax semistriatus,
Tellina nitida, Timoclea ovata, Bittium sp., Pyramidel-
loidea spp. and Turritella communis. Shell fragments
are abundant and often rounded. Taxa of the unstable
mud biocoenosis (MI; infralittoral and circalittoral) of
Peres and Picard (1964) dominate; infralittoral elements
of the upper clean-sand (SFS; bathymetry: 0–2.5 m),
of the fine, well-sorted sand (SFBC; bathymetry:
2.5–25 m) and of the Posidonia meadows biocoenoses
(HP; bathymetry: 0.5–40 m) are also present.
• Zone S1B (from -28.5/-28.65 to -25.8/-26 m; from
-24.1/-24.3 to -23.25/-23.4 m; from -22.1/-22.3
to -20.1/-20.3 m): mixed polytypic marine and
oligotypic brackish assemblages. Besides the interval
from -22.1/-22.3 to -20.1/-20.3 m, both marine
shells and their fragments, altered and dissolved, are
frequent. Species of the marine assemblages are the
same of the zone S1A. Generally low diversity and low
abundance characterize the brackish assemblages;
shells are well preserved. Bittium reticulatum, an
euryhaline component of HP biocoenosis, also spread-
ing over brackish environments, and Hydrobia sp.,
typical of the lagoonal eurythermal and euryhaline
biocoenosis (LEE), prevail. Abra segmentum (LEE),
Parvicardium exiguum (LEE) and Rissoidae spp. (HP)
are present as associated elements.
• Zone S1C (sample -22.6/-22.8 m): abundant frag-
ments and reworked marine shells. Species of the MI
biocoenosis (A. dentalis, C. gibba, T. communis)
prevail; other species such as Glycymeris sp. (SFBC),
Nuculana pella, T. ovata and Ringicula sp. are rare and
represented only by fragments.
• Zone S1D (from -19.6/-19.8 to -18.6/-18.8 m): rare
fragments and broken specimens (Cardiidae indet.,
B. reticulatum, Rissoidae indet.).
• Zone S1E (from -18.1/-18.3 to -15.1/-15.3; sample
-12.1/-12.3 m): rich oligotypic brackish assemblages
dominated by B. reticulatum (HP) and Hydrobia sp.
(LEE). Also represented are A. segmentum (LEE),
Cerastoderma glaucum (LEE), Loripes lacteus (LEE),
P. exiguum (LEE), and some taxa requiring higher
salinity conditions, such as Cerithium vulgatum, Pyra-
midelloidea spp., Pusillina spp. (HP) and Rissoa spp.
(HP). Specimens of freshwater gastropods (Acroloxus
lacustris, Bithynia tentaculata, Gyraulus spp., Lymnaea
(Radix) peregra, Physa (Physella) acuta and Valvata
piscinalis), typical of environments with still or slow-
moving waters, and a fair number of marine infralittoral
species locally occur.
• Zone S1F (from -14.7/-14.9 to -12.6/-12.8 m):
mixed marine and oligotypic brackish assemblages.
Marine elements are reworked (e.g. C. vulgatum,
C. gallina, C. gibba, Donax spp., Glycymeris sp.);
and shells of the brackish assemblages (A. segmentum,
B. reticulatum, C. glaucum, Hydrobia sp., Pusillina
spp.) are both preserved and reworked. Rare Cochli-
cella conoidea, a terrestrial species typical of dune
environments, and V. piscinalis, a freshwater gastropod,
also occur. Shell oxidation may be locally observed.
• Zone S1G (sample -11.55/-11.70 m): mixed mollusk
assemblage with marine and brackish species.
Rend. Fis. Acc. Lincei
123
Author's personal copy

B. reticulatum (HP) prevails, and is followed by
C. gallina (SFS and SFBC), Caecum sp., Lucinella
divaricata (SRPV = upper muddy sands in sheltered
areas biocoenosis; bathymetry: 0–1 m), Hydrobia sp.
(LEE), D. semistriatus (SFS) and C. vulgatum. Shell
fragments are abundant; individuals of some species are
strongly reworked (B. reticulatum, C. vulgatum); others
are in a good state of preservation (Caecum sp.,
C. gallina, Hydrobia sp., L. divaricata). Rare Gyraulus
sp. and Succinea sp. testify some freshwater input.
• Zone S1H (from -11.1/-11.3 to -10.6/-10.8 m):
fragments and specimens of marine species (B. reticul-
atum, C. glaucum, C. gallina, Donax spp., L. divari-
cata), are generally poor represented and reworked.
• Zone S1I (from -10.1/-10.3 to -4.1/-4.3 m): marine
infralittoral assemblages dominated by infaunal and
filter feeders bivalves. Shell fragments are common;
specimens are not abundant. C. gallina (SFS and
SFBC), D. semistriatus (SFS), Glycymeris sp. (SFBC)
and L. divaricata (SRPV) prevail. Rare individuals and
fragments of non-marine species (Ancylus fluviatilis,
V. piscinalis and Hygromiidae indet.) locally occur.
• Zone S1L (from -3.6/-3.8 to -3.1/-3.3 m): few and
rounded marine shell fragments, and one specimen of
L. divaricata (SRPV).
• Zone S1M (from -2.6/-2.8 to -1.6/-1.8 m): rare
marine and terrestrial shell fragments.
• Zone S1N (sample -1.10/-1.30 m): few shell frag-
ments of terrestrial gastropods and one specimen of
C. conoidea.
4.1.2 Benthic foraminifera
From the bottom to the top of the core two associations
have been individuated (Fig. 2)
• Association A1: (from -29.90/-30.00 to -21.40/
-21.50 m): the benthic association is characterized
by the prevalence of Ammonia beccarii (Linn.) and
subordinately by the species A. tepida (Linn.), Elphi-
dium crispum (Linn.), E. macellum (Fichtel and Moll),
Peneroplis pertusus (Forskal) and Quinqueloculina spp.
All taxa are present with low percentages except in
sample -28.70/-28.80 m in which A. beccarii and
A. parkinsoniana are more abundant. A. beccarii, is
present in all samples and it is indicative of a shallow-
marine environment characterized by sandy bottoms
(Sgarrella and Moncharmont Zei 1993). Jorissen (1988)
found that this species is very abundant in the Adriatic
Sea along a belt parallel to the Italian coast at a water
depth of less than 20 m, where the highest abundance is
found between 15 and 20 m in the samples with
intermediate percentages of organic matter. The species
is totally absent in the area in front of the main Po
outlets. E. crispum, E. macellum and P. pertusus are
shallow-marine species that are commonly found as
epiphytic (Blanc-Vernet 1969; Langer 1993) and do not
tolerate high concentrations of organic matter. The
assemblage found in this interval suggests an area not
directly influenced by the river run-off and mainly
characterized by coarse substrate.
• Association A2: (from -17.35/-17.50 to -11.35/
-11.50 m): taxa are dominated by very few specimens
of the genus Ammonia. While the sample at -16.35/
-16.50 m is characterized by the highest values of
A. tepida (Cushman) and E. granosum (d’Orbigny).
E. granosum is considered by Jorissen (1988) as
inhabitant of a near-shore zone (7.5–25 m water depth),
with coarse substrata poor in organic matter; it is also
reported from lagoon and shallow-marine settings
(Zampi and D’Onofrio 1987; Albani and Serandrei
Barbero 1990; Bellotti et al. 1994). A. tepida is common
in shallow-marine environments, lagoons and deltas
(Almogi-Labin et al. 1992; Favry et al. 1998; Abu-Zied
et al. 2007); it can also be indicative of moderately
restricted conditions (Debenay et al. 2005). A. tepida
(referred to as A. parkinsoniana forma tepida by Jorissen
1988) has a very close relation to run-off systems and has a
strong correlation with high percentages of organic matter.
All taxa present in this interval are common in shallow-
marine environments subjected to fluvial influence.
4.1.3 Ostracoda
Basing on ostracod assemblages, the S1 core may be
divided from the bottom to the top in seven intervals
(Fig. 2). In the first interval the ostracods are abundant both
in species diversity and in number of specimens with an
association typical of shallow-marine environments. The
third interval is characterized by an oligotypic association,
indicative of brackish water, with a very low number of
specimens per sample. The ostracod assemblage of the fifth
interval is characterized by a relatively high species
diversity and abundant valves, typical of oligo to mesoh-
aline shallow-water environments (lagoon). The changes in
the ostracod associations are due to salinity variations. On
the contrary, the following interval is characterized by
relatively low species diversity and low number of speci-
mens; these taxa are typical of shallow-marine environ-
ments. The other intervals are barren.
4.1.4 Pollen
Pollen analysis results are presented in a detailed pollen
diagram (Fig. 4) where all the recognized taxa show their
Rend. Fis. Acc. Lincei
123
Author's personal copy

percentage values plotted against depth. The arboreal taxa
percentage (AP/Tot curve in Fig. 4) oscillates between 80
and 90% giving the image of a very dense forested land-
scape (intra alias Heim 1970) in which the main arboreal
elements were deciduous Quercus, Alnus, Carpinus and
Corylus. These trees do not show important variations
along the diagram, they probably represent the main ele-
ments of a forest association established on humid soils.
Vegetation associations dominated by Alnus, Corylus,
Carpinus and Ulmus now characterize high humidity
environments linked to microclimatic conditions, well
represented all over Italy (Pedrotti and Gafta 1996). These
elements are also commonly present in alluvial environ-
ments of temperate regions in central Europe (Polunin and
Walters 1987) where they form plain-wood strips, by now
relicts in Italy and almost disappeared from southern Italy,
a part from the hygrophilous woods of Mount Circeo
(Stanisci et al. 1998).
It is very interesting from a phytogeographic and bio-
climatic point of view the constant presence of Vitis sp.
The species vinifera, as its wild representative, are now
associated with the above-mentioned arboreal elements in
thermophilous humid forests from central to southern Italy
(Pignatti 1982) even if the wild representative is in constant
regression all over Europe (Arnold et al. 1998).
Small amounts of Abies, Fagus and Betula are the
only representatives of the mountain vegetation belt.
Mediterranean elements are mainly represented by
Quercus ilex and by minor percentages of Olea, Phillyrea
and Pistacia.
Poaceae are the main representatives of the herbaceous
elements; their percentages are always below 10%. The
constant presence of Cyperaceae, water plants and spores
indicates the persistence of humid environments around the
site. In sample -17.20 the presence of a dinoflagellate cyst
suggests the possible connection to sea water.
A very similar vegetation association, dominated by a
deciduous forest on humid soils with Vitis as brushwood
element, characterized the Holocene climatic optimum
(8,354–8,524 cal yr BP) in the Vendicio plain, near For-
mia, about 100 km north of Naples (Aiello et al. 2008).
Here, the dated level was sampled at a core depth corre-
sponding to ca. 16 m below the present sea level. In the S1
core the levels sampled for pollen analysis correspond to
ca. 13–14 m b.s.l. and thus their stratigraphical position is
consistent with an early–middle Holocene age.
4.1.5 Facies analysis
On the whole, 23 lithofacies, organized in 8 lithofacies
associations, were identified and their detailed descriptions
are summarized in Table 1. The vertical stratigraphic
evolution of the S1 core (Fig. 2) is here briefly described.
The lowermost interval (-30.00/-28.30 m) is charac-
terized by bioclastic sands where marine fossil fauna are
rich and diversified, indicating a high-energy shallow-
marine environment, corresponding to the infralittoral zone
(S2 and S3 lithofacies).
From -28.30 to -25.80 m depth, coarse to fine muddy
sands with bioclasts, locally intercalated to gravels, occur
(SB1 and SB2 lithofacies); marine fauna is dominant, but
associated with brackish species indicating, on the whole, a
shallow-marine environment with freshwater input.
After a thin interval made of very altered and oxidized
reddish-brown silty sands showing diagenetic structures
related to an emersion phase (CP2 lithofacies, ephemeral
coastal plain), the shallow-marine conditions are re-estab-
lished as shown by coarse sands and gravels with typical
shallow-marine fauna (S2 and S4 lithofacies), which in turn
pass upward to sands and gravels with marine and subor-
dinately brackish fauna (SB lithofacies association, up to
-23.10 m depth). Then, a second interval (0.15 m thick)
characterized by the CP1 lithofacies, testifies to a new
emersion surface.
From -23.00 to -21.00 m depth, medium sands to gravels
with rounded and discoidal pebbles and cobbles are docu-
mented; a dominant brackish fauna or barren sediments occur
(M lithofacies association, mouth bar of estuarine/delta). The
upper interval up to about -15.30 m depth is dominated by
muddy sediments with organic matter and brackish fossil
fauna, living in still waters with soft bottoms (L1 and L2
lithofacies, open to sheltered lagoon environment).
The following interval (from -15.30 to -11.75 m),
limited by two erosional surfaces, is characterized by
gravels passing gradually to muds; here brackish fauna is
associated to freshwater (characeans) and terrestrial (gas-
tropods) species. On these bases, the sediments could be
interpreted as a flooded tidal delta or as a washover fan
formed on the landward margin of a barrier island (WF
lithofacies association). After open marine conditions came
back by a coarse reworked horizon with polygenic grains
(mollusks and travertine fragments, S6 and S5 lithofacies,
from -11.75 to -10.40 m depth), followed by a thick
interval (from -10.40 to -3.90 m) with abundant coarse to
medium sands locally intercalated with silty horizons (S
lithofacies association, upper shoreface environment).
Here, the marine species are more frequent than the
brackish ones but, on the whole, the fossil fauna is less rich
and diversified than in the lower part of S1 core.
From -3.90 to -1.60 m depth, planar and cross-lami-
nated silty sands, locally oxidized, with rare fragments of
marine bivalves, together with terrestrial gastropods, indi-
cate a foreshore/backshore deposit (FB1 and FB2 lithofa-
cies), followed by fossilized and pedogenized beach dune
ridge deposits and recent aeolian sands of coastal plain
(D and CP lithofacies associations, from -1.60 to 0 m).
Rend. Fis. Acc. Lincei
123
Author's personal copy

Table 1 S1 land core: lithofacies, lithofacies association and their environmental interpretation
Lithof.
assoc.
Lithofacies Environ. interpretation
CP CP1 Reddish-brown medium-fine and coarse silty sands with abundant altered shell fragments.
Note the strong oxidation of sediments
Coastal plain
CP2 Paleosol: brownish sandy mud with organic matter and plant material
CP3 Volcaniclastic deposits formed by greenish altered cinerite with rare whitish-grey pumice. The
thickness is about 5 cm
D D1 Dark-brown medium-fine silty sands with rounded and subrounded lithic fragments topped by
roots and humified material
Beach dune ridge
D2 Greyish muddy medium sands with pedoconcretions and burrows filled by dark muddy sandy
matrix. Rare Cochlicella conoidea and fragments of indeterminate terrestrial mollusks are
present
FB FB1 Whitish-grey medium-fine silty sands and rare muddy levels; rounded and discoidal lithic and
calcareous granules locally occur; planar and cross laminations are present. Fragments of
indeterminate terrestrial mollusks and rare fragments of marine bivalves have been
recognized
Foreshore/backshore
FB2 Gravelly coarse-medium sands with rounded and discoidal lithic and biogenic (bivalves)
fragments
S S1 Planar laminated centimeter levels of alternating grey sandy silts, muds and medium-fine
sands. Detrital mineral grains are recognized: quartz, mica and others. Shallow-marine fossil
associations are poor and oligotypic with rare gastropods, benthic foraminifera (rare
Ammonia) and ostracods (rare Aurila, Loxoconcha, Argilloecia sp.). This lithofacies only
occurs in the holocenic interval
Upper shoreface
S2 Grey medium-fine silty sands with abundant bioclasts (mollusks, anellids, echinoids, corals);
nodules of organic matter locally occur. Sometimes planar laminations are recognized.
Mollusks fauna (Antalis dentalis, Turritella communis, subordinated Chamelea gallina,Corbula gibba, Donax semistriatus, Bittium sp. etc.), also as rounded bioclasts, indicate
marine environment with high-energy. Ostracofauna is rich and diversified: Cytherettasubradiosa, Carinocythereis antiquata, Semicytherura incongruens, Cytheridea neapolitana,Pontocythere turbida, with rare Hiltermannicythere turbida, Cytheretta adriatica, Costa spp.
Benthic foraminifera association is dominated by Ammonia beccarii, subordinately Ammoniatepida, Elphidium crispum, Elphidium macellum, Peneroplis pertusus and Quinqueloculinaspp. are present. All fossil associations are poor and oligothipic in the holocenic interval
S3 Dark-grey gravelly muddy sands (medium-coarse sand grains) with abundant bioclasts
(mollusks, anellids, echinoids, bryozoans and rare corals); granules and pebbles are
subrounded and discoidal. Organic matter and altered volcanic material locally occur. Fossil
associations are the same as that in the S2 lithofacies, but some brackish mollusks (Bittiumreticulatum and Hydrobia sp.) are associated. On the whole, fossil fauna are scarce and less
diversified in the holocenic interval
S4 Coarse sands and gravels with scarce silty matrix. Clasts are represented by granules, pebbles
and cobbles (max. 3 cm in size). Mollusk association is represented by Chamelea gallina,Donax semistriatus, Glycymeris sp. and Bittium sp. with rare form of Hydrobia sp. Benthic
foraminifera are rare or absent; the genera Ammonia is dominant. Ostracod association is
poor and oligotypic (rare Aurila, Loxoconcha, Argilloecia sp.).
S5 Coarse-medium reworked sands and polygenic gravels with abundant whole and fragmented
mollusks (Chamelea gallina, Donax semistriatus, very abundant Bittium sp.), rare ?travertine
fragments; very rare Ammonia occur; ostracod association is poor and oligotypic (rare Aurila,
Loxoconcha, Argilloecia sp.). This lithofacies is only documented in the holocenic interval
S6 Coarse reworked sands and polygenic gravels with abundant whole and fragmented mollusks
(Chamelea gallina, Donax semistriatus, Lucinella divaricata, Bittium sp., Caecum sp., and
Hydrobia sp.), echinoid spines. Some characeans (oogons) are locally documented.
Ostracods association is composed of Cyprideis torosa and Loxoconcha elliptica, also with
Candona neglecta, C. angolata, Aurila woodwardi, Prionocypris zenkeri and Xestoleberis sp.
Very rare Ammonia occur. This lithofacies is only documented in the holocenic interval
Rend. Fis. Acc. Lincei
123
Author's personal copy

4.1.6 Chronology
The C1 sample (Donax trunculus, 9.70 m from the top,
-6.85 m below s.l.) gave a 14C age of 4,582 ± 50 years
BP for the S4 lithofacies (calibration: 95% r2 at
5,346 ± 33 years BP). The C2 sample (Cerastoderma
glaucum, 17.20 m, 14.35 m below s.l.) gave a 14C age of
10.073 ± 49 years BP for the L1 lithofacies (calibration:
95% r2 at 11,660 ± 218 years BP), while the 14C age of
the C3 sample (Cerithium vulgatum, 18.20, 15.35 m below
Table 1 continued
Lithof.
assoc.
Lithofacies Environ. interpretation
SB SB1 Whitish-grey coarse to fine muddy sands with bioclasts and organic matter. More altered
volcanoclastic material also occurs. Mollusk associations are marine (Antalis dentalis,Turritella communis, Chamelea gallina, Lentidium mediterraneum, Bittium sp.) and brackish
(Cerastoderma glaucum, Hydrobia sp. Abra segmentum); the marine shells are more altered
and dissolved while the brackish ones are well preserved. Ostracods and benthic foraminifera
associations are the same as that in the S2 lithofacies
Upper shoreface/brackish
SB2 Whitish-grey coarse-medium sands and gravels with scarce silty sand matrix; bioclasts are
common. Also here the marine mollusk shells are partly altered and dissolved. More altered
volcanoclastic material occurs. Fossil associations are the same as that in the SB1 lithofacies
WF WF1 Dark-grey medium-fine muddy sands with volcanoclastic material and oligotyphic brackish
(Abra segmentum, Cerastoderma glaucum, Bittium reticulatum and Hydrobia sp.) and
reworked marine molluscks. Some oogones of characeans occur. Ostracod associations are
dominated by Cyprideis torosa and Loxoconcha elliptica associated with Candona neglecta,
C. angolata, Aurila woodwardi, Prionocypris zenkeri e Xestoleberis sp. Rare or totally absent
Ammonia.
?Flood tidal delta or
washover fan in brackish
lagoon
WF2 Grey medium-coarse muddy sands with pebbles; integral and fragmented reworked shells of
marine and brackish mollusks (Bittium sp. Pusillina lineolata, and Hydrobia sp.) occur. Few
oogons of characeans and rare terrestrial gastropods (Cochlicella conoidea) are present;
oxidation locally occur. Ostracod and benthic foraminifera associations are the same as that
in the WF1 lithofacies
WF3 Grey muddy gravels with subrounded and discoidal polygenic pebbles and abundant
fragmented shells of brackish (Abra segmentum, Cerastoderma glaucum, Bittium sp.
Pusillina lineolata and Hydrobia sp.) and subordinately reworked marine mollusks. Few
oogons of characeans occur. Ostracod and benthic foraminifera associations are the same of
the WF1 lithofacies
L L1 Dark-grey laminated mud with rare polygenic coarse sands; abundant brackish (Bittiumreticulatum and Hydrobia sp.) and subordinate freshwater (Armiger crista, Bityniatentaculata, Valvata piscinalis) mollusks, living in still waters with soft bottom, are
documented. An oxidated and altered greenish pumice horizon (10 cm thick) occur. Ostracod
fauna is dominated by Cyprideis torosa and Loxoconcha elliptica and associated with
Candona neglecta, C. angolata, Aurila woodwardi, Prionocypris zenkeri and Xestoleberis sp.
Benthic foraminifera association is characterized by Ammonia tepida, Elphidium granosumand deformed Ammonia tepida. Pollen analysis shows deciduous main arboreal elements
(Quercus, Alnus, Carpinus, Corilus) representing a near forest association on humid soils;
herbaceous elements are Poaceae (\10%), Cyperaceae, water plant, spores, rare
dinoflagellate cysts, indicating humid environment connected to sea water
Brackish lagoon
L2 Dark-grey silty clay with abundant coarse to fine sand intercalations; mollusks are not
abundant or rare (Bittium sp., Hydrobia sp. Cardiidae indeter., Rissoa sp.), sometimes
oxidated. Ostracod fauna is the same as that in the L1 lithofacies. Rare Ammonia or totally
absence of benthic foraminifera occur
M M1 Greenish grey medium-fine silty sands alternated with rounded and discoidal gravels or sandy
silts. Mollusks association is dominated by brackish forms (Bittium reticulatum, Hydrobiasp., Abra segmentum, Cerastoderma glaucum, Loripes lacteus, Pusillina lineolata, Rissoaspp.); the ostracods are Cyprideis torosa and Loxoconcha elliptica, subordinately Aurilawoodwardi. Very rare Ammonia beccarii
?Mouth bar of estuary/delta
M2 Muddy sandy gravels, locally oxidated, and medium-coarse sands with abundant reworked and
dissolved marine shells. Fossil associations are the same as that in the M1 lithofacies
M3 Dark-grey muddy gravels with subrounded and rounded pebbles and cobbles (until to 8 cm in
size), locally rich in organic matter. Fossil associations are the same as that in the M1
lithofacies
Rend. Fis. Acc. Lincei
123
Author's personal copy

s.l.) was 9,568 ± 39 years BP for the L2 lithofacies (cal-
ibration: 95% r2 at 10,968 ± 180 years BP).
The ages obtained from the C2 and C3 samples are not
in agreement with the stratigraphy. The bias could be due
to reworking of this part of the core during the drilling, to
reworking of the dated mollusk shells into the sediments
or to a dating error. Nevertheless, these chronologic-data
allowed the lagoon environments (L1 and L2 lithofacies)
to be referred to as the beginning of the Holocene, while
the transition to the upper shoreface was established about
5,300 years ago. In this way, if there are no dating errors,
it is possible to constraint the upper part of the S1 suc-
cession (from L1 and L2 lithofacies to the top) into the
Holocene.
4.1.7 Environmental model and sequence stratigraphy
According to the above integrated approach based mostly
on sedimentological and palaeoecological data, it has been
essayed the reconstruction of the palaeoenvironmental
evolution of the southern sector of the Sele Plain during the
Late Pleistocene–Holocene interval. To this purpose we
have taken into account also the literature of the last
10 years (cfr. Barra et al. 1999; ISPRA 2009). The envi-
ronmental model, that we propose, corresponds to a coastal
plain with a fluvial mouth passing to the open sea through a
lagoon-barrier island system, sometimes crossed by was-
hover fans and/or channels.
In addition, the detailed sedimentological and strati-
graphic analyses allowed us to individuate the retrograda-
tional and progradational coastal systems by applying the
sequence stratigraphy approach to the S1 core (Fig. 2),
even though radiometric datings are not available for the
lower part of the core. We propose that the base of the S1
reaches the Late Pleistocene Highstand (MIS 5), where the
more open marine lithofacies occur. In this interval, about
7 m thick, two main sequences were individuated, with the
lower not complete downwards. The following estuarine/
delta deposits (about 3 m thick) correspond to a lowstand
system, formed during the prolonged sea-level fall (MIS4–
MIS2 interval) that reached -120 m during the Last Gla-
cial Maximum, c. 20 ky BP (Siddall et al. 2006; Lambeck
et al. 2011; Waelbroeck et al. 2002; Caruso et al. 2011).
This is a very condensed interval characterized at the base
by erosion, meteoric diagenesis and reworking of coastal
sediments. The subsequent Holocene marine transgression
is testified by lagoon deposits (transgressive system), fol-
lowed by a very short progradation phase associated to
flood tidal delta/washover fan sediments (highstand sys-
tem); the latter are topped by a new sequence boundary
corresponding to an erosional surface. In the upper 12 m, a
fourth sequence was individuated corresponding to a beach
dune prograding system, still active at present. On the
whole, the third and fourth sequences clearly record the
Holocene sea-level rise (MIS1).
4.2 The offshore data
4.2.1 The C101 and C1213 cores
The C1213 core (Fig. 5) entered a marine succession
whose textures and biological assemblage give indication
of a progressively shallowing upward environment in the
lower 170 cm (A) and of an increasingly deep environment
in the uppermost 320 cm (C). Between them a 15 cm thick,
well-sorted, fine sand deposit with shell fragments (B) is
included, being underlain by a sharp irregular surface.
The A unit is composed of two fining upward horizons:
the lower one (sub-unit A1) includes at the base a poorly
sorted, mud-sustained deposit with bryozoans, sponges and
mollusk shells; the upper one (sub-unit A2) shows at the
base a mud-sustained, poorly sorted bed made by rhodolit-
bearing pebbles, bryozoans and mollusk shells.
The C unit consists, from the base to the top, of a 60-cm-
thick, mud-sustained, poorly sorted heterometric bioclastic
sand, whose grains are fining upwards, and of a homoge-
neous marine mud which holds two volcaniclastic deposits:
the deepest one (sub-unit C1) is a 15-cm-thick, dark-grey
ash deposit dispersed within an olive-grey muddy matrix,
slightly bioturbated and bounded by blunt surfaces; the
shallowest (sub-unit C2) is 45 cm thick and is made up of
coarse white and grey pumiceous lapilli, above a sharp and
regular surface; these distinctive lithofacies match well
with analogous tephra found in several cores in the Salerno
Gulf and respectively correlated to the AD 79 Vesuvius
Plinian eruption and to the 2.7–3.3 ky BP Somma-Vesu-
vius activity (AP eruptions; Insinga et al. 2008).
The B unit displays textures and bioclastic content
typical of a lower shoreface deposit; therefore, it registers
the shallowest marine environment within the core sedi-
ment record. Based on its stratigraphic position and litho-
logic features it can be related to the LGM lowstand, i.e. to
the stage of maximum retreat of the shoreline. On the basis
of lithofacies analysis and stratigraphic markers it is pos-
sible to infer to the base of the core a relative age as old as
MIS 3, since it passes through the lower shoreface deposit
(B) relative to the maximum glacial lowstand condition
(MIS 2) and reaches a former deposit that stands for a mid-
shelf environment (A2) and inner-shelf environment (A1).
The C101 core (Fig. 5) entered a marine succession
mostly consisting of fine siliciclastic deposits. The basal
unit (A) is constituted of mud deposits that include several
thin layers of well-sorted silt and lenses of fragmented
shells (mainly mollusks and echinoids). A 17-cm-thick
volcaniclastic deposit (sub-unit A1) occurs between 281 and
302 cm, composed of normally graded, fine to very fine
Rend. Fis. Acc. Lincei
123
Author's personal copy
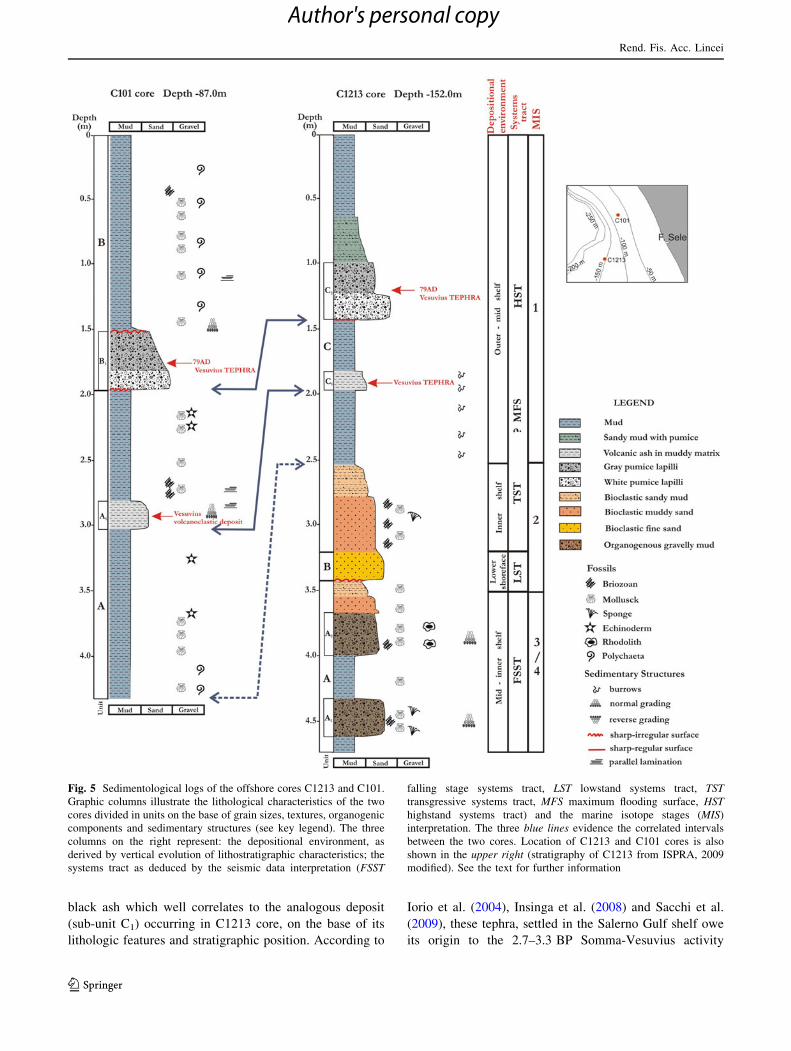
black ash which well correlates to the analogous deposit
(sub-unit C1) occurring in C1213 core, on the base of its
lithologic features and stratigraphic position. According to
Iorio et al. (2004), Insinga et al. (2008) and Sacchi et al.
(2009), these tephra, settled in the Salerno Gulf shelf owe
its origin to the 2.7–3.3 BP Somma-Vesuvius activity
Fig. 5 Sedimentological logs of the offshore cores C1213 and C101.
Graphic columns illustrate the lithological characteristics of the two
cores divided in units on the base of grain sizes, textures, organogenic
components and sedimentary structures (see key legend). The three
columns on the right represent: the depositional environment, as
derived by vertical evolution of lithostratigraphic characteristics; the
systems tract as deduced by the seismic data interpretation (FSST
falling stage systems tract, LST lowstand systems tract, TSTtransgressive systems tract, MFS maximum flooding surface, HSThighstand systems tract) and the marine isotope stages (MIS)
interpretation. The three blue lines evidence the correlated intervals
between the two cores. Location of C1213 and C101 cores is also
shown in the upper right (stratigraphy of C1213 from ISPRA, 2009
modified). See the text for further information
Rend. Fis. Acc. Lincei
123
Author's personal copy

(AP eruptions). Nevertheless, the occurrence of parallel
lamination toward the top deposit, the concentration of shell
fragments and the dubious age dating at 4.4 ky cal BP at
293 cm bsf (Table 2), let us to infer that this volcaniclastic
deposit underwent reworking processes in this sector of the
shelf.
The upper unit (B) starts from 195 to 164 cm bsf with
the tephra layer (sub-unit B1) correlated to the AD 79
Vesuvius eruption composed of angular and sub-angular
white to grey pumice lapilli laying on a basal erosive
contact. This unit is overlain by a normally graded and
reworked deposit consisting of pumice lapilli and ash, mud
supported, with fragmented organism shells up to 149 cm
bsf, thus evidencing synsedimentary reworking processes
(Buccheri et al. 2002; Sacchi et al. 2005) soon after the AD
79 eruption. Between 149 cm and the core top, the unit B is
mainly composed of mud deposits with some thin well-
sorted silty layers and widespread presence of shells,
mainly mollusks, sometimes fragmented.
According to the recognized lithofacies, the depositional
conditions have changed at the core site from a mid to an
outer shelf environment characterized by fine-grained
sedimentation with current influxes able to selected the silt
sediment and to break and drag the organism shells. On the
basis of the 14C datings (Table 2) the sediment-core is
representative of the whole Holocene marine sedimenta-
tion, since about 11.2 ky cal BP.
4.2.2 Seismic data
The very high resolution (VHR) seismic acoustic profile
CSal 5 was shot off the Sele River mouth, up to 40 m of
depth, 2.9 km off the shoreline; the CSal 2 VHR profile
was shot off in between the Tusciano and Picentino river
mouths, up to 20 m of depth, 1.9 km from the coastline.
The GNS12 uniboom profile was run off the Capo di
Fiume-Solofrone streams at a depth of 20 m, 3.6 km
distant from the Paestum coast (Figs. 1, 6).
The seismic lines go far off the present-day sand bars
that lie within 10 m below s.l. (ISPRA 2009). Some evi-
dent stratigraphic horizons can be traced beneath the sea-
floor, across the plain margins and bear a meaningful
stratigraphic reference over the Salerno Gulf and the
surrounding shelf margin. From the seafloor downwards,
they are as follows:
• a shallow unit beneath the seabed, 8–10 m thick, with
typical discontinuous reflectors possibly due to fluid
escape and plastic deformation features or sediment
undulations (Urgeles et al. 2011). Between 40 and 70 m
below s.l., in the central and northern sector of the Bay.
The unit is bounded at the base by a regular and
conformable marine reflector that lies halfway between
the AD 79 Vesuvius tephra and the present-day seabed,
thus not older than about 1 ky (ISPRA 2009);
• the conformable, isochronous, marine reflector related
to the Vesuvius AD 79 Plinian eruption, traceable from
about 22 m of depth, far beyond the shelf break and
largely reported in recent literature on the offshore;
• the diachronic unconformity (LPu) associated to the
increasing land exposure, following the sea-level retreat
since MIS 5.1; it truncates the top of the Late
Pleistocene prograding units, and marks a progressively
longer hiatus landwards. It matches locally with the
ravinement surface, that records the landward shift of
the wave base erosion during the following phase of
sea-level rise. In this case the paralic lithosomes are
missing (Cattaneo and Steel 2003);
• a set of gentle-dipping prograding lithosomes with
sigmoidal reflections above the LPu (4 in the CSal 2; at
least 1 in the CSal 5, Fig. 6), stacking in a retrogressive
arrangement towards the most recent one. The four
prograding set off the TP coast lie 70, 45, 30 and 23 m
deep b.s.l and are, respectively, 6.8, 4.3, 3.1 and 2.5 km
far from the present-day shoreline;
• a laterally continuous prograding unit (Fig. 6), trun-
cated at the top by the LPu, and mostly consisting of a
Late Pleistocene Falling Stage Systems Tract (FSST) in
the mid-shelf and of a lowstand prograding wedge
(LST), seaward (Plint and Nummedal 2000).
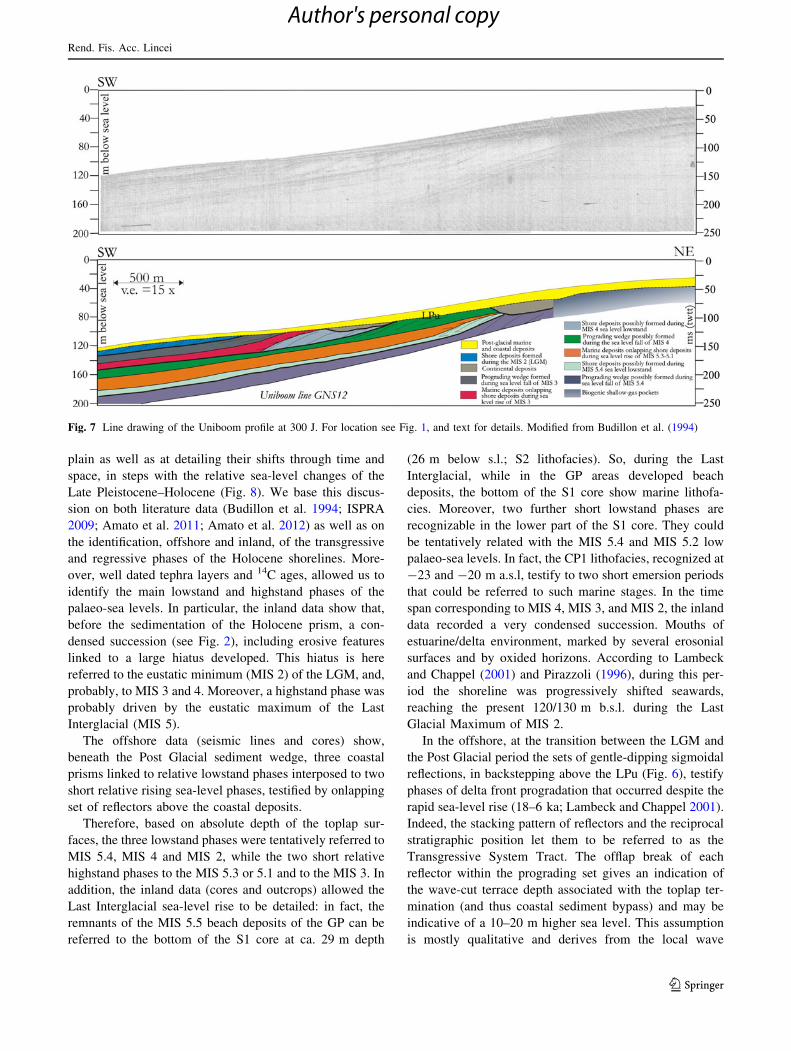
The Uniboom profile shows three prograding units
alternating with onlapping units below the LPu (Fig. 7).
This stratigraphic pattern was formerly interpreted as
due to a forced regression of the coastal units driven by the
Late Pleistocene, fourth order sea-level drop following
the Tyrrhenian highstand stage (Budillon et al. 1994).
Table 2 14C dating results of the analyzed samples in C101 core
Sample depth (cm) Lab. DSA Radiocarbon age
(years BP)
Calibrated
age (1 r)
Calibrated age
BP (years BP)
d13C (%)
C101 (-45) -811 860 ± 31 1158–1219 AD 792–631 4 ± 1
C101 (-293) -802 3,911 ± 38 2469–2396 BC 4,419–4,346 0 ± 1
C101 (-358) -804 5,520 ± 45 4445–4420 BC 6,395–6,370 1 ± 1
Mixed planktonic foraminifera shells were used as material to be dated
Rend. Fis. Acc. Lincei
123
Author's personal copy
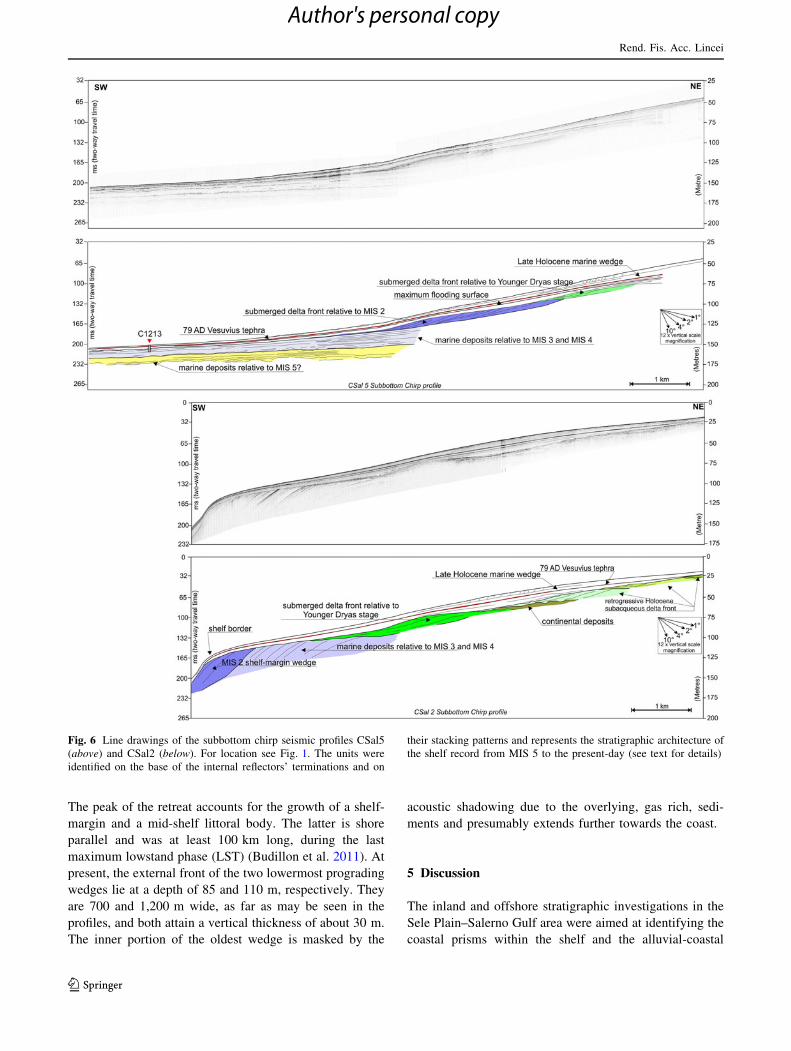
The peak of the retreat accounts for the growth of a shelf-
margin and a mid-shelf littoral body. The latter is shore
parallel and was at least 100 km long, during the last
maximum lowstand phase (LST) (Budillon et al. 2011). At
present, the external front of the two lowermost prograding
wedges lie at a depth of 85 and 110 m, respectively. They
are 700 and 1,200 m wide, as far as may be seen in the
profiles, and both attain a vertical thickness of about 30 m.
The inner portion of the oldest wedge is masked by the
acoustic shadowing due to the overlying, gas rich, sedi-
ments and presumably extends further towards the coast.
5 Discussion
The inland and offshore stratigraphic investigations in the
Sele Plain–Salerno Gulf area were aimed at identifying the
coastal prisms within the shelf and the alluvial-coastal
Fig. 6 Line drawings of the subbottom chirp seismic profiles CSal5
(above) and CSal2 (below). For location see Fig. 1. The units were
identified on the base of the internal reflectors’ terminations and on
their stacking patterns and represents the stratigraphic architecture of
the shelf record from MIS 5 to the present-day (see text for details)
Rend. Fis. Acc. Lincei
123
Author's personal copy
plain as well as at detailing their shifts through time and
space, in steps with the relative sea-level changes of the
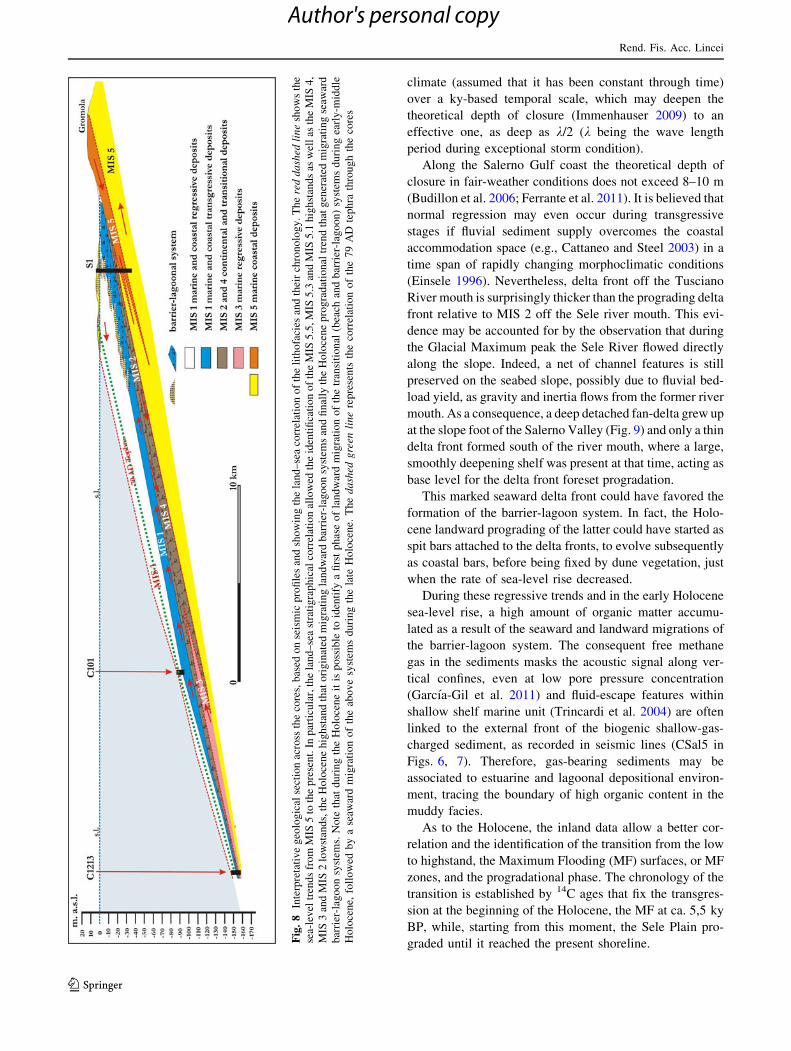
Late Pleistocene–Holocene (Fig. 8). We base this discus-
sion on both literature data (Budillon et al. 1994; ISPRA
2009; Amato et al. 2011; Amato et al. 2012) as well as on
the identification, offshore and inland, of the transgressive
and regressive phases of the Holocene shorelines. More-
over, well dated tephra layers and 14C ages, allowed us to
identify the main lowstand and highstand phases of the
palaeo-sea levels. In particular, the inland data show that,
before the sedimentation of the Holocene prism, a con-
densed succession (see Fig. 2), including erosive features
linked to a large hiatus developed. This hiatus is here
referred to the eustatic minimum (MIS 2) of the LGM, and,
probably, to MIS 3 and 4. Moreover, a highstand phase was
probably driven by the eustatic maximum of the Last
Interglacial (MIS 5).
The offshore data (seismic lines and cores) show,
beneath the Post Glacial sediment wedge, three coastal
prisms linked to relative lowstand phases interposed to two
short relative rising sea-level phases, testified by onlapping
set of reflectors above the coastal deposits.
Therefore, based on absolute depth of the toplap sur-
faces, the three lowstand phases were tentatively referred to
MIS 5.4, MIS 4 and MIS 2, while the two short relative
highstand phases to the MIS 5.3 or 5.1 and to the MIS 3. In
addition, the inland data (cores and outcrops) allowed the
Last Interglacial sea-level rise to be detailed: in fact, the
remnants of the MIS 5.5 beach deposits of the GP can be
referred to the bottom of the S1 core at ca. 29 m depth
(26 m below s.l.; S2 lithofacies). So, during the Last
Interglacial, while in the GP areas developed beach
deposits, the bottom of the S1 core show marine lithofa-
cies. Moreover, two further short lowstand phases are
recognizable in the lower part of the S1 core. They could
be tentatively related with the MIS 5.4 and MIS 5.2 low
palaeo-sea levels. In fact, the CP1 lithofacies, recognized at
-23 and -20 m a.s.l, testify to two short emersion periods
that could be referred to such marine stages. In the time
span corresponding to MIS 4, MIS 3, and MIS 2, the inland
data recorded a very condensed succession. Mouths of
estuarine/delta environment, marked by several erosonial
surfaces and by oxided horizons. According to Lambeck
and Chappel (2001) and Pirazzoli (1996), during this per-
iod the shoreline was progressively shifted seawards,
reaching the present 120/130 m b.s.l. during the Last
Glacial Maximum of MIS 2.
In the offshore, at the transition between the LGM and
the Post Glacial period the sets of gentle-dipping sigmoidal
reflections, in backstepping above the LPu (Fig. 6), testify
phases of delta front progradation that occurred despite the
rapid sea-level rise (18–6 ka; Lambeck and Chappel 2001).
Indeed, the stacking pattern of reflectors and the reciprocal
stratigraphic position let them to be referred to as the
Transgressive System Tract. The offlap break of each
reflector within the prograding set gives an indication of
the wave-cut terrace depth associated with the toplap ter-
mination (and thus coastal sediment bypass) and may be
indicative of a 10–20 m higher sea level. This assumption
is mostly qualitative and derives from the local wave
Fig. 7 Line drawing of the Uniboom profile at 300 J. For location see Fig. 1, and text for details. Modified from Budillon et al. (1994)
Rend. Fis. Acc. Lincei
123
Author's personal copy
climate (assumed that it has been constant through time)
over a ky-based temporal scale, which may deepen the
theoretical depth of closure (Immenhauser 2009) to an
effective one, as deep as k/2 (k being the wave length
period during exceptional storm condition).
Along the Salerno Gulf coast the theoretical depth of
closure in fair-weather conditions does not exceed 8–10 m
(Budillon et al. 2006; Ferrante et al. 2011). It is believed that
normal regression may even occur during transgressive
stages if fluvial sediment supply overcomes the coastal
accommodation space (e.g., Cattaneo and Steel 2003) in a
time span of rapidly changing morphoclimatic conditions
(Einsele 1996). Nevertheless, delta front off the Tusciano
River mouth is surprisingly thicker than the prograding delta
front relative to MIS 2 off the Sele river mouth. This evi-
dence may be accounted for by the observation that during
the Glacial Maximum peak the Sele River flowed directly
along the slope. Indeed, a net of channel features is still
preserved on the seabed slope, possibly due to fluvial bed-
load yield, as gravity and inertia flows from the former river
mouth. As a consequence, a deep detached fan-delta grew up
at the slope foot of the Salerno Valley (Fig. 9) and only a thin
delta front formed south of the river mouth, where a large,
smoothly deepening shelf was present at that time, acting as
base level for the delta front foreset progradation.
This marked seaward delta front could have favored the
formation of the barrier-lagoon system. In fact, the Holo-
cene landward prograding of the latter could have started as
spit bars attached to the delta fronts, to evolve subsequently
as coastal bars, before being fixed by dune vegetation, just
when the rate of sea-level rise decreased.
During these regressive trends and in the early Holocene
sea-level rise, a high amount of organic matter accumu-
lated as a result of the seaward and landward migrations of
the barrier-lagoon system. The consequent free methane
gas in the sediments masks the acoustic signal along ver-
tical confines, even at low pore pressure concentration
(Garcıa-Gil et al. 2011) and fluid-escape features within
shallow shelf marine unit (Trincardi et al. 2004) are often
linked to the external front of the biogenic shallow-gas-
charged sediment, as recorded in seismic lines (CSal5 in
Figs. 6, 7). Therefore, gas-bearing sediments may be
associated to estuarine and lagoonal depositional environ-
ment, tracing the boundary of high organic content in the
muddy facies.
As to the Holocene, the inland data allow a better cor-
relation and the identification of the transition from the low
to highstand, the Maximum Flooding (MF) surfaces, or MF
zones, and the progradational phase. The chronology of the
transition is established by 14C ages that fix the transgres-
sion at the beginning of the Holocene, the MF at ca. 5,5 ky
BP, while, starting from this moment, the Sele Plain pro-
graded until it reached the present shoreline.Fig
.8
Inte
rpre
tati
ve
geo
log
ical
sect
ion
acro
ssth
eco
res,
bas
edo
nse
ism
icp
rofi
les
and
sho
win
gth
ela
nd
–se
aco
rrel
atio
no
fth
eli
tho
faci
esan
dth
eir
chro
no
log
y.
Th
ere
dd
ash
edli
ne
sho
ws
the
sea-
lev
eltr
end
sfr
om
MIS
5to
the
pre
sen
t.In
par
ticu
lar,
the
lan
d–
sea
stra
tig
rap
hic
alco
rrel
atio
nal
low
edth
eid
enti
fica
tio
no
fth
eM
IS5
.5,M
IS5
.3an
dM
IS5
.1h
igh
stan
ds
asw
ell
asth
eM
IS4
,
MIS
3an
dM
IS2
low
stan
ds,
the
Ho
loce
ne
hig
hst
and
that
ori
gin
ated
mig
rati
ng
lan
dw
ard
bar
rier
-lag
oo
nsy
stem
san
dfi
nal
lyth
eH
olo
cen
ep
rog
rad
atio
nal
tren
dth
atg
ener
ated
mig
rati
ng
seaw
ard
bar
rier
-lag
oo
nsy
stem
s.N
ote
that
du
rin
gth
eH
olo
cen
eit
isp
oss
ible
toid
enti
fya
firs
tp
has
eo
fla
nd
war
dm
igra
tio
no
fth
etr
ansi
tio
nal
(bea
chan
db
arri
er-l
ago
on
)sy
stem
sd
uri
ng
earl
y-m
idd
le
Ho
loce
ne,
foll
ow
edb
ya
seaw
ard
mig
rati
on
of
the
abo
ve
syst
ems
du
rin
gth
ela
teH
olo
cen
e.T
he
da
shed
gre
enli
ne
rep
rese
nts
the
corr
elat
ion
of
the
79
AD
tep
hra
thro
ug
hth
eco
res
Rend. Fis. Acc. Lincei
123
Author's personal copy

The normal regression trend of the shoreline was more
accentuated during historical times (last 2,500 years)
because it was supported by the emplacement of the distal
fall deposits related to Neapolitan volcanoes eruptions,
mainly the AD 79 event, and by the increasing man-
induced impact on vegetation and on alluvial-coastal
sectors.
Data correlation allows some palaeogeographical and
palaeoenvironmental aspects to be detailed for the Holo-
cene transgression and progradation phases. In particular,
at the beginning of the Holocene, the transgression was fast
and favored the landward migration of the barrier-lagoon
system, up to the maximum ingression that occurred at ca.
5,5 ky BP and reached the position of the Laura palaeo-
ridges. Then, the barrier-lagoon system migrated mainly
seaward. The presence of lagoons during the early Holo-
cene is confirmed by pollen data from the Sele Plain as well
as from very similar vegetation associations characterizing
the Holocene climatic optimum of other Tyrrhenian allu-
vial-coastal plains (Aiello et al. 2008). A different age of
the passage from a retrogradational to aprogradational
trend can be inferred for the sector of the plain between the
Tusciano and Picentino rivers, on the base of the CSal2
seismic profile. Here, the backstepping of the shore per-
sisted until approximately 3.0 ky BP, probably due to the
different vertical tectonic behaviour of the plain, during
Holocene times. In fact,according to Pappone et al. (2012)
and Vilardo et al. (2009), the dx sector of the Sele plain
is undergoing subsidence, while the sx sector should be
tectonically stable or slightly uplifting.
6 Final remarks
To summarize the above data presentation and interpreta-
tion we have assembled in Fig. 9 the main late Quaternary
Fig. 9 Salerno Gulf–Sele Plain palaeogeographical sketches from
MIS 5.5 to the present-day, showing the supposed shoreline position
and some palaeoenvironmental interpretations, derived by inland and
offshore stratigraphical data. 1) lagoon deposits, 2) sandy beach
deposits, 3) MIS 5 beach and dune deposits, 4) travertine deposits, 5)
pro-delta deposits, 6) black dashed lines indicate presumed palaeo-
channels of the Sele and Tusciano rivers, created during the sea-level
falls, 7) cores cited in the text
Rend. Fis. Acc. Lincei
123
Author's personal copy

paleogeographical features of the Sele plain/Salerno valley,
tentatively, reconstructed through the correlation of inland
and offshore data.
Starting from MIS 5.5, when the shoreline had reached
the GP, at ca 4 km inland with a palaeo-sea level of
11/13 m above s.l., the shoreline strongly prograded, until
reaching 120/130 m below s.l. (Lambeck and Chappel
2001) during the eustatic minimum of MIS 2. This long-
time lasting of sea level fall was interrupted by three short
rise periods which could be tentatively referred to MIS 5.3,
5.1 and MIS 3. In fact, if the palaeo-shorelines of MIS 5.3
and 5.1 can be located some km seawards from the S1 core,
as suggested by seismic data, two sea-level falls were
recognized in the lower part of the S1 core, at ca 20 m
below s.l. (CP lithofacies layers). These layers, testifying a
short emersion, could have formed during the MIS 5
lowstand phases, as supposed by seismic offshore data, that
show shore deposits at ca. 80 m below s.l. As to the
Holocene, the palaeo-shorelines of the maximum trans-
gressive phase can be located ca. 1.5 km inland, in proxi-
mity of the Laura palaeoridge, when the barrier-lagoon
system was at its maximum retrogradation stage, approxi-
matively at 5.5 ky BP. Then, the shoreline prograded and
the barrier-lagoon system migrated seaward, reaching the
present position. The Sterpina dune system can be related
to this moment when the stasis of the progradation allowed
the formation of the pre 2.5 ky Sterpina I ridge and then the
sin AD 79 Sterpina II ridge (Amato et al. 2012; Alberico
et al. 2011).
References
Abu-Zied RH, Keatings KW, Flower RJ (2007) Environmental
controls on foraminifera in Lake Qarun. Egypt J Foram Res
37:136–149
Aiello G, Barra D, De Pippo T, Donadio C, Miele P, Russo Ermolli E
(2008) Morphological and paleoenvironmental evolution of the
Vendicio coastal plain in the Holocene (Latium, central Italy). Il
Quaternario. Ital J Quat Sci 20:185–194
Albani AD, Serandrei Barbero R (1990) I foraminiferi della Laguna e
del Golfo di Venezia. Mem Sci Geol Padova 42:271–341
Alberico I, Amato V, Aucelli PPC, D’Argenio B, Di Paola G,
Pappone G (2011) Historical shoreline change of the Sele Plain
(southern Italy): the 1870–2009 Time Window. J Coast Res. doi:
10.2112/JCOASTRES-D-10-00197.1
Almogi-Labin A, Perelis-Grossovicz L, Raab M (1992) Living
Ammonia from a hypersaline inland pool, Dead Sea area. Isr J
Foram Res 22:257–266
Amato V, Avagliano G, Cinque A, Cipriani M, Di Paola G,
Pontrandolfo A, Rosskopf CM, Santoriello A (2009a) Geomor-
phology and geoarchaeology of the Paestum area: modification
of the physical environment in historical times. Mediterranee
112:129–135
Amato V, Aiello G, Barra D, Di Pasquale G, Di Pasquale L, Russo
Ermolli E, Aurino P, Bailo Modesti G, Basile F, Cerchiai L,
D’Andrea M, Mancusi M, Negro D, Pellegrino C, Rossi A,
Schiano Di Cola C, Viscione M (2009b) Holocene climate
forcing and man-induced environmental changes of Abitato
Antico di Pontecagnano area (Salerno, southern Italy). Mediter-
ranee 112:119–128
Amato V, Aucelli PPC, Cinque A, D’Argenio B, Di Donato V, Russo
Ermolli E, Pappone G, Petrosino P, Rosskopf CM (2011)
Holocene paleo-geographical evolution of the Sele River coastal
plain (southern Italy): new morpho-sedimentary data from the
Paestum area. Il Quaternario. Ital J Quat Sci 24:5–7
Amato V, Aucelli PPC, D’Argenio B, Da Prato S, Ferraro L, Pappone
G, Petrosino P, Rosskopf CM, Russo Ermolli E (2012)
Holocene environmental evolution of the costal sector before
the Poseidonia-Paestum archaeological area (Sele plain, southern
Italy). Rend Fis Acc Lincei. doi:10.1007/s12210-011-0161-1 (in
press)
Antonioli F, Silenzi S, Vittori E, Villani C (1999) Sea level changes
and tectonic mobility: precise measurements in three coastlines
of Italy considered stable during the last 125 ky. Phys Chem
Earth (A) 24(4):337–342
Antonioli F, Bard E, Silenzi S, Improta S (2004) 215 kyr history of
sea level oscillation from marine and continental layer of
Argentarola Cave’s speleothems (Italy). Glob Planet Change
21:66–77
Arnold C, Gillet F, Gobat JM (1998) Situation de la vigne sauvage
Vitis vinifera ssp. sylvestris en Europe. Vitis 37:159–170
Athersuch J, Horne DJ, Whittaker JE (1989) Marine and brackish
water ostracods. In: Kermack DM, Barnes RSK (eds) Synopses
of the British Fauna (New Series), vol. 43
Barra D, Calderoni G, Cinque A, De Vita P, Rosskopf CM, Russo
Ermolli E (1998) New data on the evolution of the Sele River
coastal plain (southern Italy) during the Holocene. Il Quaternario
(Ital J Quat Sci) 11:287–299
Barra D, Calderoni G, Cipriani M, De La Geniere’ J, Fiorillo L, Greco
G, Mariotti Lippi M, Mori Secci M, Pescatore T, Russo B,
Senatore MR, Tocco Sciarelli G, Thorez J (1999) Depositional
history and paleogeographic reconstruction of Sele coastal plain
during Magna Grecia settlement of Hera Argiva (southern Italy).
Geologica Romana 35:151–166
Bellotti P, Carboni MG, Di Bella L, Palagi I, Valeri P (1994) Benthic
foraminiferal assemblages in the depositional sequence of the
Tiber Delta. In: Matteucci R, Carboni MG, Pignatti JS (eds)
Studies on ecology and paleoecology of benthic communities.
Boll Soc Paleontol Ital Spec vol 2:29–40
Blanc-Vernet L (1969) Contribution a l’etude des foraminiferes de
Mediterranee. These de doctorat d’etat, travaux de la station
marine d’Endoume, Marseille
Brancaccio L, Cinque A, Belluomini G, Branca M, Delitala L (1986)
Isoleucine epimerization dating and tectonic significance of
upper Pleistocene sea level features of the Sele Plain (southern
Italy). Zeit Geomorph NF, Suppl Bd 62:159–166
Brancaccio L, Cinque A, D’Angelo G, Russo F, Santangelo N,
Sgrosso I (1987) Evoluzione tettonica e geomorfologica della
Piana del Sele (Campania, Appennino meridionale). Geogr Fis
Din Quat 10:47–55
Brancaccio L, Cinque A, Russo F, Santangelo N, Alessio M, Allegri
L, Improta S, Belluomini G, Branca M Delitala L (1988) Nuovi
dati cronologici sui depositi marini e continentali della Piana del
F. Sele e della costa settentrionale del Cilento (Campania,
Appennino meridionale). Atti del 74� Congresso Nazionale della
Soc Geol Ital A:55–62
Buccheri G, Russo B, Bertolodo M (1994) Stratigrafia e paleocli-
matologia della carota GN82-C41 (Mar Tirreno): foraminiferi e
pteropodi. Boll Soc Geol Ital 113:120–155
Buccheri G, Capretto G, Di Donato V, Esposito P, Ferruzza G,
Pescatore T, Russo Ermolli E, Senatore MR, Sprovieri M,
Bertoldo M, Carella D, Madonna G (2002) A high resolution
Rend. Fis. Acc. Lincei
123
Author's personal copy

record of the last deglaciation in the southern Tyrrhenian sea:
environmental and climatic evolution. Mar Geol 186:447–470
Budillon F, Pescatore T, Senatore MR (1994) Cicli deposizionali del
Pleistocene superiore-olocene sulla piattaforma continentale del
Golfo di Salerno (Tirreno meridionale). Boll Soc Geol Ital
113:303–316
Budillon F, Vicinanza D, Ferrante V, Iorio M (2006) Sediment
transport and deposition during extreme sea storm events at the
Salerno Bay (Tyrrhenian Sea): comparison of field data with
numerical model results. Nat Hazards Earth Syst Sci 6(5):839–
852
Budillon F, Aiello G, Conforti A, D’Argenio B, Ferraro L, Marsella
E, Monti L, Pelosi N, Tonielli R (2011) The coastal depositional
systems along the Campania continental margin (Italy, Southern
Tyrrhenian Sea) since the Late Pleistocene: new information
gathered in the frame of the CARG Project. In: Brugnoli et al
(eds) Marine Research @ CNR, National Research Council of
Italy, 539551. ISSN:2239-5172
Carboni MG, Di Bella L (1996) Morphological variation in the genus
Ammonia Brunnich in the Plio/Pleistocene of the Sabina
(Latium). In: Cherchi, A. (Ed.), Autoecology of selected fossil
organisms: Achievements and problems. Boll. Soc. Paleont. It.,
Special Volume 3: 63–76
Carlson RL, Gangi AF, Snow KL (1986) Empirical reflection travel
time versus depth and velocity versus depth functions for the
deep sea sediments column. J Geophys Res 91:8249–8266
Caruso A, Cosentino C, Pierre C, Sulli A (2011) Sea-level changes
during the last 41,000 years in the outer shelf of the southern
Tyrrhenian Sea: evidence from benthic foraminifera and seis-
mostratigraphic analysis. Quat Int 232:122e131
Cattaneo A, Steel RJ (2003) Transgressive deposits: a review of their
variability. Earth Sci Rev 62:187–228
Colalongo ML (1969) Ricerche sugli Ostracodi nei fondali antistanti
il delta del Po. Giornale di Geologia 36:335–362
Coltorti M, Melis E, Patta D (2010) Geomorphology, stratigraphy and
facies analysis of some Late Pleistocene and Holocene key
deposits along the coast of Sardinia (Italy). Quat Int 222:19–35
D’Argenio B, Ferreri V, Stanzione D, Brancaccio L, Ferreri M (1983)
I travertini di Pontecagnano (Campania). Geomorfologia, Sed-
imentologia, Geochimca. Boll Soc Geol Ital 102:123–136
Damuth JE (1980) Use of high-frequency (3.5–12 kHz) echograms in
the study of near-bottom sedimentation processes in the deep-
sea: a review. Mar Geol 38:51–75
Debenay JP, Millet M, Angelidis M (2005) Relationships between
foraminiferal assemblages and hydrodynamics in the Gulf of
Kalloni (Greece). J Foram Res 35:327–343
Einsele G (1996) Event deposits: the role of sediment supply and
relative sea-level changes: overview. Sediment Geol 104:11–37
Favry A, Guelorget O, Debenay JP, Perthuisot JP (1998) Distribution
des peuplements de foraminiferes actuels dans une lagune
mediterraneenne: l’etang du Prevost. Vie et milieu 48:41–53
Ferrante V, Budillon F, Iorio M, Lubritto C, Monaco M, Pugliese
Carratelli E, Vicinanza D (2011) Tracing possible offshore sand
movement during extreme sea storm events: an example from
the Salerno Bay (Southern Tyrrhenian Sea). In: Brugnoli E et al
(eds) Marine Research @ CNR, National Research Council of
Italy, 539551. ISSN:2239-5172
Ferranti L, Antonioli F, Mauz B, Amorosi A, Dai Pra G, Mastronuzzi
G, Monaco C, Orru P, Pappalardo M, Radtke U, Renda P,
Romano P, Sanso P, Verrubbi V (2006) Markers of the last
interglacial sea level high stand along the coast of Italy: tectonic
implications. Quat Int 145–146:30–54
Fiorini F, Vaiani SC (2001) Benthic foraminifers and transgressive-
regressive cycles in the Late Quaternary subsurface sediments of
the Po Plain near Ravenna (Northern Italy). Boll Soc Paleontol
Ital 40:357–403
Garcıa-Gil S, de Blas E, Martınez-Carreno N, Iglesias J, Rial-Otero R,
Simal-Gandara J, Judd AG (2011) Characterisation and pre-
liminary quantification of the methane reservoir in a coastal
sedimentary source: San Simon Bay, Rıa de Vigo, NW Spain.
Estuar Coast Shelf Sci 91:232–242
Gliozzi E (2000) Tardiglacial-Holocene freshwater ostracod assem-
blages from Ripasottile wells (Rieti basin, Latium, central Italy).
Giornale di Geologia 3(62):159–172
Gliozzi E, Mazzini I (1998) Palaeoenvironmental analysis of Early
Pleistocene brackish marshes in the Rieti and Tiberino intrapen-
ninic basins (Latium and Umbria, Italy) using ostracods.
Palaeogeogr Paleoclimatol Palaeoecol 254:235–252
Heim J (1970) Les relations entre les spectres polliniques recents et la
vegetation actuelle en Europeoccidentale. Mem Soc R Belg
4:1–181
Immenhauser A (2009) Estimating paleo-water depth from the
physical rock record. Earth Sci Rev 96:107–139
Insinga D, Molisso F, Lubritto C, Sacchi M, Passariello I, Morra V
(2008) The proximal marine record of Somma–Vesuvius volca-
nic activity in the Naples and Salerno bays, Eastern Tyrrhenian
Sea, during the last 3 kyr. J Volcanol Geotherm Res 177(1):170–
186
Iorio M, Sagnotti L, Angelino A, Budillon F (2004) High-resolution
petrophysical and paleomagnetic study of late-Holocene shelf
sediments, Salerno Gulf, Tyrrhenian Sea. Holocene 14(3):433–442
Iorio M, Liddicoat J, Budillon F, Tiano P (2009) Paleomagnetic
secular variation time constraints on Late Neogene Geological
events in slope sediment from the eastern Tyrrhenian Sea. Estuar
Coast Shelf Sci 92:233–243
IPCC (2007) Contribution of Working Group I to the Fourth
Assessment Report of the Intergovernmental Panel on Climate
Change. In: Solomon S, Qin D, Manning M, Chen Z, Marquis M,
Averyt KB, Tignor M, Miller HL (eds) Climate change 2007: the
physical science basis. Cambridge University Press, Cambridge
ISPRA (2009) Note illustrative della Carta Geologica d’Italia alla
scala 1:50.000. Foglio 486, Foce Sele. A cura di Cinque A.,
Servizio Geologico Nazionale, 1–205
Jorissen FJ (1988) Benthic foraminifera from the Adriatic Sea:
principles of phenotypic variations. Utrecht Micropaleontol Bull
37:1–176
Kerney M (1999) Atlas of the land and freshwater molluscs of Britain
and Ireland. Harley Books, Colchester
Lambeck K, Chappel J (2001) Sea level change through the last
glacial cycle. Science 292:679–686
Lambeck K, Antonioli F, Purcell A, Silenzi S (2004) Sea level change
along the Italian coast for the past 10,000 yrs. Quat Sci Rev
23:1567–1598
Lambeck K, Antonioli F, Anzidei M, Ferranti L, Leoni G (2011) Sea
level change along Italian coast during Holocene and a
prediction for the future. Quat Int 232:250–257
Langer M (1993) Epiphytic foraminifera. Mar Micropal 20:235–265
Lirer L, Pescatore TS, Booth B, Walker GPL (1973) Two plinian
pumice-fall deposits from Somma–Vesuvius. Italy Geol Soc Am
Bull 84:759–772
Meisch C (2000) Freshwater Ostracoda of Western and Central
Europe. In: Schwoerbel J, Zwick P (eds) Suesswasserfauna von
Mitteleuropa 8/3. Spektrum Akademischer Verlag, Heidelberg
Montenegro ME, Pugliese N (1995) Ostracodi della laguna di
Orbetello: tolleranza ed opportunismo. Atti Mus Geol Paleontol
Monfalcone Quad Spec 3:71–80
Orru PE, Antonioli F, Hearty PJ, Radtke U (2011) Chronostrati-
graphic confirmation of MIS 5 age of a Baymouth Bar at Is
Arenas (Cagliari, Italy). Quat Int 232:169–178
Ortolani F, Torre M, Russo B, Di Nocera S (1979) Depositi alto-
miocenici del bordo settentrionale della Piana del Sele (Camp-
ania). Boll Soc Geol Ital 98:3–14
Rend. Fis. Acc. Lincei
123
Author's personal copy

Pappone G, Aucelli PPC, Alberico I, Amato V, Antonioli F, Cesarano
M, Di Paola G, Pelosi N (2012) Relative sea-level rise and
marine erosion and inundation in the Sele river coastal plain
(Southern Italy): scenarios for the next century. Rend Fis Acc
Lincei. doi:10.1007/s12210-012-0166-4
Pedrotti F, Gafta D (1996) Ecologia delle foreste ripariali e paludose
dell’Italia. L’uomo e l’ambiente, Universita degli Studi di
Camerino, Camerino, Italy
Peres JM (1982) Major benthic assemblages. In: Kinne O (eds) Mar
Ecol 5(1):373–508
Peres JM, Picard J (1964) Nouveau manuel de Bionomie benthique de
la Mer Mediterranee. Recueil des Travaux de la Station Marine
d’Endoume Bulletin 31(47):5–137
Pignatti S (1982) Flora d’Italia. Agricole (ed), Bologna, vol. III,
pp 451–620
Pirazzoli PA (1996) Sea-level changes: the last 20,000 years. Wiley,
Chichester
Plint AG, Nummedal D (2000) The falling stage systems tract:
recognition and importance in sequence stratigraphic analysis.
In: Hunt D, Gawthorpe RL (eds) Sedimentary response to forced
regression, vol 172. Geol Soc Lond Spec Publ, pp 1–17
Polunin O., Walters M. (1987). Guida alle vegetazioni d’Europa.
Zanichelli ed., Bologna
Posamentier HW, Allen GP (1999) Siliciclastic sequence stratigra-
phy: concepts and applications. Concepts in sedimentology and
paleontology, vol 7. Society of Economic Paleontologists and
Mineralogists (SEPM)
Rahmstorf S (2007) A semi-empirical approach to projecting future
sea-level rise. Science 315:368–370
Reimer PJ, Baillie MGL, Bard E, Bayliss A, Beck JW, Blackwell PG,
Bronk Ramsey C, Buck CE, Burr GS, Edwards RL, Friedrich M,
Grootes PM, Guilderson TP, Hajdas I, Heaton TJ, Hogg AG,
Hughen KA, Kaiser KF, Kromer B, McCormac FG, Manning
SW, Reimer RW, Richards DA, Southon JR, Talamo S, Turney
CSM, van der Plicht J, Weyhenmeyer CE (2009) IntCal09 and
Marine09 radiocarbon age calibration curves, 0–50,000 years -
cal BP. Radiocarbon 51(4):1111–1150
Russo F, Belluomini G (1992) Affioramenti di depositi marini
tirreniani sulla piana in destra del Fiume Sele (Campania). Boll
Soc Geol Ital 111:25–31
Sacchi M, Insinga D, Milia A, Molisso F, Raspini A, Torrente MM,
Conforti A (2005) Stratigraphic signature of the Vesuvius 79 AD
event off the Sarno prodelta system, Naples Bay. Mar Geol
222–223:443–469
Sacchi M, Molisso F, Violante C, Esposito E (2009) Insights into flood-
dominated fan-deltas: very high-resolution seismic examples off
the Amalfi cliffed coasts, eastern Tyrrhenian Sea. Geol Soc Lond
322:33–71
Salvador A (1994) International stratigraphic guide. A guide to
stratigraphic classification, terminology, and procedure. The
International Union of Geological Sciences and The Geological
Society of America, Boulder
Sgarrella F, Moncharmont Zei M (1993) Benthic foraminifera of the
Gulf of Naples (Italy): systematics and autoecology. Boll Soc
Paleontol Ital 32:145–264
Siddall M, Chappell J, Potter EK (2006) Eustatic sea level during past
interglacials. In: Sirocko F (ed) The climate of past interglacials.
Elsevier, Amsterdam, pp 75–82
Spampinato CR, Costa B, Di Stefano A, Monaco C, Scicchitano G
(2011) The contribution of tectonics to relative sea-level change
during the Holocene in coastal south-eastern Sicily: new data
from boreholes. Quat Int 232:214–227
Stanisci A, Presti G, Blasi C (1998) I boschi igrofili del Parco
Nazionale del Circeo (Italia centrale). Ecol Mediterr 24:73–88
Trincardi F, Cattaneo A, Correggiari A, Ridente D (2004) Evidence of
soft sediment deformation, fluid escape, sediment failure and
regional weak layers within the late Quaternary mud deposits of
the Adriatic Sea. Mar Geol 213:91–119
Urgeles R, Cattaneo A, Puig P, Liquete C, De Mol B, Amblas D,
Sultan N, Trincardi F (2011) A review of undulated sediment
features on Mediterranean prodeltas: distinguishing sediment
transport structures from sediment deformation. Mar Geophys
Res 32:49–69
Vail PR (1987) Seismic stratigraphic interpretation procedure. In:
Bally AW (ed) Atlas of seismic stratigraphy. AAPG Mem
26:49–212
Vilardo G, Ventura G, Terranova C, Matano F, Nardo S (2008)
Ground deformation due to tectonic, hydrothermal, gravity,
hydrogeological, and anthropic processes in the Campania
Region (Southern Italy) from Permanent Scatterers Synthetic
Aperture Radar Interferometry. Remote Sens Environ 113:197-
212
Waelbroeck C, Labeyrie L, Michel E, Duplessy JC, Lambeck K,
McManus JF, Balbon E, Labracherie M (2002) Sea-level and
deep water temperature changes derived from benthic forami-
nifera isotopic records. Quat Sci Rev 21:295–305
Wheatcroft RA, Drake DE (2003) Post-depositional alteration and
preservation of sedimentary event layers on continental margins,
I. The role of episodic sedimentation. Mar Geol 199:123–137
Zampi M, D’Onofrio S (1987) I foraminiferi della laguna di Levante
(Orbetello, Grosseto). Atti della Soc. Sc. Nat. Mem Ser A
93:101–127
Rend. Fis. Acc. Lincei
123
Author's personal copy




















