
Changes in spatial point patterns of pioneer woody plants across a large tropical landslide
Upload
khangminh22Category
view
0download
0
‘OLUTION AND DIVERSITY OF VASCULAR PLANTS
. 2005. Phylogeny of cryptogrannnoid ferns and related taxa based on rbcL sequences Nordic Journal of
‘tp://amerfernsoc.org lists many resources, including pubiications, references, local, national, and internaites, commerjcal fern sites, and fern databases.
LIGNOPHYTIES—WOODY PLANTS
The lignophytes, or woody plants (also called Lignophyta),
are a monophyletic lineage of euphyllous vascular plants that
share the derived features of a vascular cambium, which
gives rise to wood, and a cork cambium, which produces
cork (Figures 5.1, 5.2). Growth of the vascular and cork
cambia is called secondary growth because it initiates after
the vertical extension of stems and roots due to cell expansion
(primary growth). A vascular cambium is a sheath, or hollow
cylinder, of cells that develops within the stems and roots as a
continuous layer, between the xylem and phloem in extant,
eustelic spermatophytes (see later discussion). The cells of
the vascular cambium divide mostly tangentially (parallel to
a tangential plane), resulting initially in two concentric layers
of cells (Figure 5.3A). One of these layers remains as the vas
cular cambium and continues to divide indefinitely; the other
layer eventually differentiates into either secondary xylem
wood, if produced to the inside of the cambium, or secondary
phloem, if produced to the outside (Figure 5.3A,B). Because
129
Ginkgophyta 144
Ginkgoaceae 145
Coniferae—Conifers 145
Pinopsida 148
Pinaceae 148
Cupressopsida 151
Araucariaceae 151
Cupressaceae 151
Podocarpaceae 154
Taxaceae 154
Gnetales 156
Ephedraceae 157
REVIEW QJESTIONS 160
EXERCISES 160
REFERENCES FOR FURTHER STUDY 161
WEB SITES 162
layers of cells are produced both to the inside and outside of
a continuously generated cambium, this type of growth is
termed bifacial. Generally, much more secondary xylem is
produced than secondary phloem. [Note that a secondary
cambium independently evolved in fossil lineages within the
lycophytes (e.g., Lepidodendron) and equisetophytes (e.g.,
Calamites), but this cambium was unifacial, producing sec
ondary xylem (wood) to the inside but no outer secondary
phloem, likely limiting in terms of an adaptive feature.]
Secondary growth results in an increase of the width or girth
of stems and roots (Figures 5.3B, 5.4). This occurs both by
expansion of the new cells generated by the cambium and by
accompanying radial divisions, increasing the number of cells
within a given growth ring. Many woody plants have regular
growth periods, e.g., forming annual rings of wood (Figure 5.4).
A cork cambium is similar to a vascular cambium, only it
differentiates near the periphery of the stem or root axis. The
cork cambium and its derivatives constitute the periderm
(referred to as the outer bark). The outermost layer of the
periderm is cork (Figure 5.3B). Cork cells contain a waxy
I 5EVOLUTION AND DIVERSITY OF
WOODY AND SEED PLANTSLlGNOPHYTESWOODY PLANTS 129
SPERMATOPHYTES—SEED PLANTS 131
Seed Evolution 131
Pollination Droplet 135
Pollen Grains 135
Pollen Tube 136
Ovule and Seed Development 136
Seed Adaptations 139
Eustele 139
DIVERSITY OF WOODYAND SEED PLANTS 139
Archeopteris 139
“Pteridosperms”—”Seed Ferns” 139
Gymnospermae—Gymnosperms 140
Cycadophyta—Cycads 140
Cycadaceae 141
Zamiaceae 142
© 2010 Elsevier Inc. All rights reserved.
doi: 10.101 61B978-0- 12-374380-000005.2

UNIT II EVOLUTION AND DIVERSITY OF PLANTS 131130 CHAPTER 5 EVOLUTION AND DIVERSITY OF WOODY AND SEED PLANTS
t
Ca a
Ia)
C C
is.sC) ç
o a) S—o o0 0 a)t a):t S a)
Ct S rjCto 0
— •0-5c .
- 0 5(/D
- Lignophyta (Woody Plants)
Spermatophyta (Seed Plants)
Gymnospermae (Gymnosperms)
Coniferae (Conifers)
Cupressopsida
a)4.,
a)
0-I.
a)oO— (/,
r Gnetales—,
4.
a)a)
a) a)S
a)C: S
S Sa) a)
CS
C
porose
epimatium +receptacle
iiiiifiiiii 1 ovule/scale
iiiiiiiifiiii leaves simple
t =extincttaxon
= extinct lineage
pollen tube—sperm nonmotile (siphonogamy)
‘1F
__
-
____I
/- I 1g., -,
W
V c •4L %H
I-
I’
:14a,, •‘
lIst. pa-. ,41 t
!%: r
-‘- . -
.—
______
-a -
-
Et :rLct
classification and identification of woody plants. Wood ana
tomical features may also be used to study the past, a spe
cialty known as dendrochronolOgy (see Chapter 10).
Another feature of lignophytes is that they possess ances
trally monopodial growth, in which a single main shoot
develops branches from lateral (usually axillary) buds (see
Chapters 4, 9). Although monopodial growth is presumed to
have arisen prior to the monilophyte—lignophyte split, it
enabled woody plants in particular the capability of forming
extensive (sometimes massive) woody branching systems,
permitting them to survive and reproduce more effectively.
SPERMATOPHYTESSEED PLANTS —
The spermatophyta, commonly called spermatophytes or
seed plants, are a monophyletic lineage within the lignophytes
(Figure 5.1). The major evolutionary novelty that unites this
group is the seed. A seed is defined as an embryo, which is an
immature diploid sporophyte developing from the zygote, sur
rounded by nutritive tissue and enveloped by a seed coat
(Figure 5.5). The embryo generally consists of an immature
root called the radicle, a shoot apical meristem called the
epicotyl, and one or more young seed leaves, the cotyledons;
the transition region between root and stem is called the
hypocotyl (Figures 5.5, 5.10). An immature seed, prior to
fertilization, is known as an ovule.
SEED EVOI.,UTION
The evolution of the seed involved several steps. The
exact sequence of these is not certain, and two or more
“steps” in seed evolution may have occurred concomitantly
and be functionally correlated. The probable steps in seed
evolution are as follows (Figure 5.6):
1. HeterospOry. Heterospory is the formation of two types
of haploid spores within two types of sporangia: large,
fewer-numbered megaspores, which develop via meiosis
in the megasporangium, and small, more numerous
microsporeS, the products of meiosis in the microsporafl
gium (Figures 5.6, 5.7). The ancestral condition, in which a
single spore type forms, is called “homosporyY Each
megaspore develops into a female gametophyte that bears
only archegonia; a microspore develops into a male game
tophyte, bearing only antheridia. Although heterospory
was prerequisite to seed evolution, there are fossil plants
that were heterosporous but had not evolved seeds, among
these being species of Archeopteris (Figures 5.1, 5.1 3A;
see later discussion). Note that heterospory has evolved
independently in other, nonseed plants, e.g., in the extant
lycophytes Selaginella and Isoetes and in the water ferns
(Chapter 4).
— s eustele
— — pollen tube—sperm motile (zooidogamy)
— — endosporic, male gametophyte = pollen grain
— — pollination droplet
— — integument with micropyle— — retention of megaspore within megasporangium
— — reduction to 1 megaspore per megasporangium SEED— — endosporic female gametophyte (embryo -
+ nutntive tissue— — heterospory + integuments)— — cork cambium (periderm)— vascular cambium (secondary vascular tissue, md. wood)
FIGURE 5.1 Cladogram of the woody and seed plants. Major apomorphies are indicated beside a thick hash mark. Families in bold aredescribed in detail. Modified from Bowe et al. (2000); Chaw et al. (2000); Frohlich et al. (2000); and Samigullin et al. (1999).
polymer called suberin (similar to cutin) that is quite resist- shrubs or trees with tall overstory canopies (e.g., Figure 5.2),ant to water loss (see Chapter 10). a significant ecological adaptation. Cork produced by the cork
The vascular cambium and cork cambium constituted major cambium functions as a thick layer of cells that protectsevolutionary novelties. Secondary xylem, or wood, functions the delicate vascular cambium and secondary phloem fromin structural support, enabling the plant to grow tall and acquire mechanical damage, predation, and desiccation.massive systems of lateral branches. Thus, the vascular cam- Wood anatomy can be quite complex. The details ofbium was a precursor to the formation of intricately branched cellular structure are important characters used in the
FIGURE 5.2 Composite
giant sequoia, a woody conifer that is the most massive, nonclonal
organism on Earth, and among the tallest of trees.

AND DIVERSITY OF WOODY AND SEED PLANTS UNIT II EVOLUTION AND DIVERSITY OF PLANTS 131
a.)—a.)
a.) a.) a:
a a:a:
C C— -a:
C.) a:a.,) c_)
Ieustele
— —
—pollen tube—sperm motile (zooidogamy)
endosporic, male gametophyte pollen grainpollination droplet
integument with micropyle
retention of megaspore within megasporangjumreduction to 1 megaspore per megasporangjum SEEDendosporic female gametophyte (embryo
+ nutritive tissueheterospory+ integuments)
cork cambjum (periderm)vascular cambjum (secondary vascular tissue, mci. wood)
plants. Major apomorphies are indicated beside a thick hash mark. Families in bold are10); Chaw et al. (2000); Frohljch et al. (2000); and Samigullin et al. (1999).
shrubs or trees with tall overstory canopies (e.g., Figure 5.2),a significant ecological adaptation. Cork produced by the corkcambium functions as a thick layer of cells that protectsthe delicate vascular cambium and secondary phloem frommechanical damage, predation, and desiccation.
Wood anatomy can be quite complex. The details ofcellular structure are important characters used in the
FIGURI 5.2 Composite photograph ofSequoiadendrongiganteum,
giant sequoia, a woody conifer that is the most massive, nonclonal
organism on Earth, and among the tallest of trees.
classification and identification of woody plants. Wood ana
tomical features may also be used to study the past, a spe
cialty known as dendrochronology (see Chapter 10).
Another feature of lignophytes is that they possess ances
trally monopodial growth, in which a single main shoot
develops branches from lateral (usually axillary) buds (see
Chapters 4, 9). Although monopodial growth is presumed to
have arisen prior to the monilophyte—lignophyte split, it
enabled woody plants in particular the capability of forming
extensive (sometimes massive) woody branching systems,
permitting them to survive and reproduce more effectively.
SPERMATOPHYTES—SEED PLANTS
The Spermatophyta, commonly called spermatophytes or
seed plants, are a monophyletic lineage within the lignophytes
(Figure 5.1). The major evolutionary novelty that unites this
group is the seed. A seed is defined as an embryo, which is an
immature diploid sporophyte developing from the zygote, sur
rounded by nutritive tissue and enveloped by a seed coat
(Figure 5.5). The embryo generally consists of an immature
root called the radicle, a shoot apical meristem called the
epicotyl, and one or more young seed leaves, the cotyledons;
the transition region between root and stem is called the
hypocotyl (Figures 5.5, 5.10). An immature seed, prior to
fertilization, is known as an ovule.
SEED EVOLUTION
The evolution of the seed involved several steps. The
exact sequence of these is not certain, and two or more
“steps” in seed evolution may have occurred concomitantly
and be functionally correlated. The probable steps in seed
evolution are as follows (Figure 5.6):
1. Heterospory. Heterospory is the formation of two types
of haploid spores within two types of sporangia: large,
fewer-numbered megaspores, which develop via meiosis
in the megasporangium, and small, more numerous
microspores, the products of meiosis in the microsporan
glum (Figures 5.6, 5.7). The ancestral condition, in which a
single spore type forms, is called “homospory.” Each
megaspore develops into a female gametophyte that bears
only archegonia; a microspore develops into a male game
tophyte, bearing only antheridia. Although heterospory
was prerequisite to seed evolution, there are fossil plants
that were heterosporous but had not evolved seeds, among
these being species of Archeopteris (Figures 5.1, 5.13A;
see later discussion). Note that heterospory has evolved
independently in other, nonseed plants, e.g., in the extant
lycophytes Selaginella and Isoetes and in the water ferns
(Chapter 4).
— Lignophyta (Woody Plants) — —
Spermatophyta (Seed Plants)
- Gymnospermae (Gymnosperms) —
— Conjferae (Conifers) — —
1 — Cupressopsida
1
1
r Gnetales-,
.
a: a:a.) a.)C.) C.)a: a:
a: a:C.?
aril
Spollen tube—sperm nonmotjie (siphonogamy)
is quite resist
flstituted major‘ood, functionstall and acquirevascular cam
ately branched

2. Endospory. Endospory is the complete development of,
in this case, the female gametophyte within the original
spore wall (Figure 5.6). The ancestral condition, in which
the spore germinates and grows as an external gameto
phyte, is called exospory. The evolution of endosporic
female gametophytes was correlated with that of
endosporic male gametophytes (pollen grains); see later
discussion.
3. Reduction of megaspore number to one. Reduction of
megaspore number occurred in two ways. First, the number
of cells within the megasporangium that undergo meiosis
(each termed a megasporocyte or megaspore mother
cell) was reduced, from several to one (Figure 5.6). This
single diploid megasporocyte gives rise to four haploid
megaspores. Second, of the four haploid megaspores pro
duced by meiosis, three consistently abort, leaving only
one functional megaspore. This single megaspore also
undergoes a great increase in size, correlated with the
megasporangium.
4. Retention of the megaspore. Instead of the megaspore
being released from the sporangium (the ancestral condi
tion, as occurs in all homosporous nonseed plants),
in seed plants it is retained within the megasporangium
(Figure 5.6). This was accompanied by a reduction in
thickness of the megaspore wall.
5. Evolution of the integument & micropyle. The final
event in seed evolution was the envelopment of the
megasporangium by a layer of tissue, called the integu
ment (Figure 5.6). The integument grows from the base of
the megasporangium (which is often called a nucellus
when surrounded by an integument) and envelopes it,
except at the distal end. Fossil evidence suggests that the
integument likely evolved from separate lobes derived
from telomes (ancestral branches) that surrounded the
megasporangium. These “preovules”, i.e., ovules prior to
the evolution of integuments, possessed a rim or ring of
tissue at the apex of the megasporangium, the lagenos
tome, which functioned to funnel pollen grains to a pol
lination chamber. (See, e.g., Stewart and Rothwell 1993
for details.) The epitome of seed evolution occurred with
the evolutionary “fusion” of the telomes to form the
integument, a continuous sheath that completely sur
rounds the nucellus. The integument of all extant seed
plants has a small pore at the distal end called the micro-
pyle. The micropyle replaced the ancestral lagenostome
as the site of entry of pollen grains (or in angiosperms, of
pollen tubes). The micropyle also functions in the
mechanics of pollination droplet formation and resorp
tion (see below). Note that a single integument represents
the ancestral condition of spermatophytes; in angiosperms
a second integument layer evolved later (Chapter 6).
132 CHAPTER 5 EVOLUTION AND DIVERSITY OF WOODY AND SEED PLANTS
I
UNIT II EVOLUTION AND DIVERSITY OF PLANTS 133
I
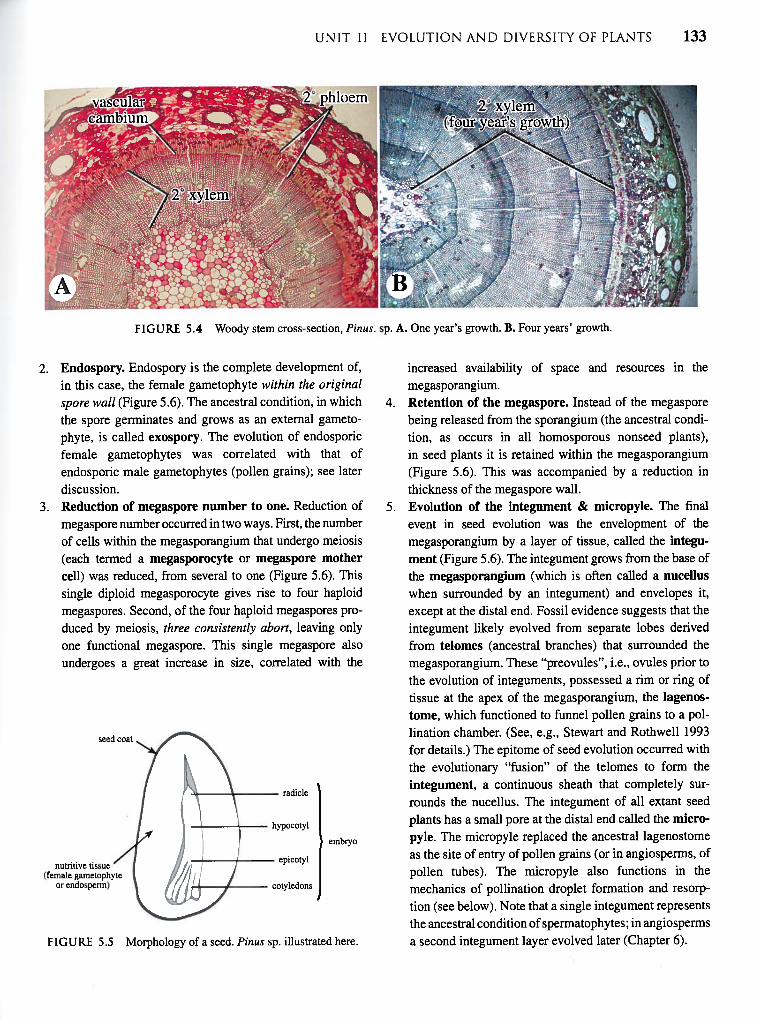
I FIGURE 5.4 Woody stem cross-section, Pinus. sp. A. One year’s growth. B. Four years’ growth.
vascularcambjum
increased availability of space and resources in the
A
B
FIGURE 5.3 A. Development of the vascular cambium. B. Development of secondary vascular tissue in the stem, illustrated here for aeusteljc stem.
2 xylem
2 phloem
periderrn
cork(epidermis sloughed off to outside)
radicle
fluthtive tissue(female gametophyle
or endosperm)
embryo
cotyledons
FIGURE 5.5 Morphology of a seed. Pinus sp. illustrated here.
AND DIVERSITY OF WOODYAND SEED PLANTS
2. Endospory. Endospory is the complete development of,
in this case, the female gametophyte within the original
spore wall (Figure 5.6). The ancestral condition, in which
the spore germinates and grows as an external gameto
phyte, is called exospory. The evolution of endosporicfemale gametophytes was correlated with that of
endosporic male gametophytes (pollen grains); see later
discussion.3. Reduction of megaspore number to one. Reduction of
megaspore number occurred in two ways. First, the number
of cells within the megasporangium that undergo meiosis
(each termed a megasporocyte or megaspore mother
cell) was reduced, from several to one (Figure 5.6). This
single diploid megasporocyte gives rise to four haploidmegaspores. Second, of the four haploid megaspores pro
duced by meiosis, three consistently abort, leaving only
one functional megaspore. This single megaspore also
undergoes a great increase in size, correlated with the
UNIT II EVOLUTION AND DIVERSITY OF PLANTS 133
increased availability of space and resources in themegasporangium.
4. Retention of the megaspore. Instead of the megaspore
being released from the sporangium (the ancestral condi
tion, as occurs in all homosporous nonseed plants),
in seed plants it is retained within the megasporangium(Figure 5.6). This was accompanied by a reduction inthickness of the megaspore wall.
5. Evolution of the integument & micropyle. The final
event in seed evolution was the envelopment of themegasporangium by a layer of tissue, called the integu
ment (Figure 5.6). The integument grows from the base of
the megasporangium (which is often called a nucellus
when surrounded by an integument) and envelopes it,
except at the distal end. Fossil evidence suggests that theintegument likely evolved from separate lobes derived
from telomes (ancestral branches) that surrounded the
megasporangium. These “preovules”, i.e., ovules prior to
the evolution of integuments, possessed a rim or ring of
tissue at the apex of the megasporangium, the lagenos
tome, which functioned to funnel pollen grains to a pol
lination chamber. (See, e.g., Stewart and Rothwell 1993
for details.) The epitome of seed evolution occurred with
the evolutionary “fusion” of the telomes to form the
integument, a continuous sheath that completely sur
rounds the nucellus. The integument of all extant seed
plants has a small pore at the distal end called the micro-
pyle. The micropyle replaced the ancestral lagenostome
as the site of entry of pollen grains (or in angiosperms, of
pollen tubes). The micropyle also functions in the
mechanics of pollination droplet formation and resorp
tion (see below). Note that a single integument represents
the ancestral condition of spermatophytes; in angiosperms
a second integument layer evolved later (Chapter 6).
1
FIGURE 5.4 Woody stem cross-section, Pinus. sp. A. One year’s growth. B. Four years’ growth.
vascularcambium
2’ xylem
vascularcambium 2 phloem
l’phloem
1’ xylem2’ xylem
2’ phloem
periderm
cortex
seed coat
cork(epidermis sloughed off to outside)
radicle
nutritive tissue(female gametophyte
or endosperm)
embryo
epicotyl
cotyledons
ambium. B. Development of second vascular tissue in the stem, illustrated here for a FIGURE 5.5 Morphology of a seed. Pinus sp. illustrated here.

134 CHAPTER 5 EVOLUTION AND DIVERSITY OF WOODY AND SEED PLANTS UNIT II EVOLUTION AND DIVERSITY OF PLANTS 135
POLLINATION DROPLET
One possible evolutionary novelty associated with seed evo
lution is the pollination droplet. This is a droplet of liquid
that is secreted by the young ovule through the micropyle
(Figures 5.1OA, 5.171). This droplet is mostly water plus
some sugars or amino acids and is formed by the breakdown
of cells at the distal end of the megasporangium (nucellus).
The cavity formed by this breakdown of cells is called the
pollination chamber (Figure 5. bA). The pollination drop
let functions in transporting pollen grains through the micro-
pyle. This occurs by resorption of the droplet, which “pulls”
pollen grains that have contacted the droplet into the pollina
tion chamber. It is unknown whether a pollination dropletwas present in the earliest seed plants. However, the presenceof a pollination droplet in many nonflowering seed plants
suggests that its occurrence may be apomorphic for at least
the extant seed plant lineages. Note that the ovules of
angiosperms lack pollination droplets or pollination cham
bers, as flowering plants have evolved a different mechanism
of pollen grain transfer (see Chapter 6).
POLLEN GRAINSConcomitant with the evolution of the seed was the evolution of
pollen grains (Figure 5.8). A pollen grain is, technically, an
immature, endosporic male gametophyte. Endospory in pollen
grain evolution was similar to the same process in seed evolu
tion, involving the development of the male gametophyte within
the original spore wall. Pollen grains of seed plants are extremely
reduced male gametophytes, consisting of only a few cells.
They are termed “immature” male gametophytes because, at the
time of their release, they have not fully differentiated.
After being released from the microsporangium, pollen must
be transported to the micropyle of the ovule (or, in angio
sperms, to the stigmatic tissue of the carpel; see Chapter 6) in
order to ultimately effect fertilization. Wind dispersal, in com
bination with an ovule pollination droplet (see later discus
sion), was probably the ancestral means of pollen transport.
After being transported to the ovule (or stigmatic tissue), the
male gametophyte completes development by undergoing
additional mitotic divisions and differentiation. The male
gametophyte grows an exosporic pollen tube, which functions
antheridia
/gametophyte
male gametophyte
microsporangium
(n)
(n) SpOraflgium
1. Heterospory megasporangium
Sporophyte Body,.._-‘ (2n)
mitosis, growth, & differentiation mitosis, growth, & differentiation
7Embryo
(2n) Microsporangium Megasporangium(2n) (2n)
/ \ \mitosis, growth, & differentiation mitosis, growth, & d(fferentianon
‘\Zygote SPOROPHYTE GENERATION Microsporocyte Megasporocyte
(2n) (2N) (2n) (2n)
t——fertilization rneiosis —— meioszs ——
/‘ Jn‘jN I) GAMETOPHYTE GENERATION) (N)
Microspores Megaspores..)
(n) (n) j Conifers (mci. Gnetales) ©
Egg Sperm 1. (sperm nonflagellate in © ( (n) (n)
k and Angiosperms)female gametophyte { Archegonium Antheridium 1
(lost in the Angiosperms (reduced to absent in& some Gnetales) (n) (n) 3’ extant seed plants)
archegonia
/wall
female gametophyte(contained in megaspore)
2. Endospory
megasporangiurn
archegonia
0
1/mitosis, growth, & differentiation
mitosis, growth, & derentiaiionGametophyte
(n)Male
Female Gametophyte(n)
megasporangium
3. Reduction to 1 megaspore
megaspore
FIGURE 5.7 Life cycle of heterosporous seed plants.
gametophyte
megasporangium
4. Retention of megaspore
FIGURE 5.6 Ovule and seed evolution in the spermatophytes (hypothetical, for purpose of illustration).
5. Evolution of Integument & Micropyle

134 CHAPTER 5 EVOLUTION AND DIVERSITY OF WOODY AND SEED PLANTS UNIT II EVOLUTION AND DIVERSITY OF PLANTS 135
POLLINATION DROPLET
One possible evolutionary novelty associated with seed evo
lution is the pollination droplet. This is a droplet of liquid
that is secreted by the young ovule through the micropyle
(Figures 5.1OA, 5.171). This droplet is mostly water plus
some sugars or amino acids and is formed by the breakdown
of cells at the distal end of the megasporangium (nucellus).
The cavity formed by this breakdown of cells is called the
pollination chamber (Figure 5. bA). The pollination drop
let functions in transporting pollen grains through the micro-
pyle. This occurs by resorption of the droplet, which “pulls”
pollen grains that have contacted the droplet into the pollina
tion chamber. It is unknown whether a pollination dropletwas present in the earliest seed plants. However, the presenceof a pollination droplet in many nonflowering seed plants
suggests that its occurrence may be apomorphic for at least
the extant seed plant lineages. Note that the ovules of
angiosperms lack pollination droplets or pollination cham
bers, as flowering plants have evolved a different mechanism
of pollen grain transfer (see Chapter 6).
POLLEN GRAINSConcomitant with the evolution of the seed was the evolution of
pollen grains (Figure 5.8). A pollen grain is, technically, an
immature, endosporic male gametophyte. Endospory in pollen
grain evolution was similar to the same process in seed evolu
tion, involving the development of the male gametophyte within
the original spore wall. Pollen grains of seed plants are extremely
reduced male gametophytes, consisting of only a few cells.
They are termed “immature” male gametophytes because, at the
time of their release, they have not fully differentiated.
After being released from the microsporangium, pollen must
be transported to the micropyle of the ovule (or, in angio
sperms, to the stigmatic tissue of the carpel; see Chapter 6) in
order to ultimately effect fertilization. Wind dispersal, in com
bination with an ovule pollination droplet (see later discus
sion), was probably the ancestral means of pollen transport.
After being transported to the ovule (or stigmatic tissue), the
male gametophyte completes development by undergoing
additional mitotic divisions and differentiation. The male
gametophyte grows an exosporic pollen tube, which functions
antheridia
/gametophyte
male gametophyte
microsporangium
(n)
(n) SpOraflgium
1. Heterospory megasporangium
Sporophyte Body,.._-‘ (2n)
mitosis, growth, & differentiation mitosis, growth, & differentiation
7Embryo
(2n) Microsporangium Megasporangium(2n) (2n)
/ \ \mitosis, growth, & differentiation mitosis, growth, & d(fferentianon
‘\Zygote SPOROPHYTE GENERATION Microsporocyte Megasporocyte
(2n) (2N) (2n) (2n)
t——fertilization rneiosis —— meioszs ——
/‘ Jn‘jN I) GAMETOPHYTE GENERATION) (N)
Microspores Megaspores..)
(n) (n) j Conifers (mci. Gnetales) ©
Egg Sperm 1. (sperm nonflagellate in © ( (n) (n)
k and Angiosperms)female gametophyte { Archegonium Antheridium 1
(lost in the Angiosperms (reduced to absent in& some Gnetales) (n) (n) 3’ extant seed plants)
archegonia
/wall
female gametophyte(contained in megaspore)
2. Endospory
megasporangiurn
archegonia
0
1/mitosis, growth, & differentiation
mitosis, growth, & derentiaiionGametophyte
(n)Male
Female Gametophyte(n)
megasporangium
3. Reduction to 1 megaspore
megaspore
FIGURE 5.7 Life cycle of heterosporous seed plants.
gametophyte
megasporangium
4. Retention of megaspore
FIGURE 5.6 Ovule and seed evolution in the spermatophytes (hypothetical, for purpose of illustration).
5. Evolution of Integument & Micropyle

as a haustorial organ, obtaining nutrition by absorption from thesurrounding sporophytic tissue (Figure 5.9; see Pollen Tabe).
POLLEN TUBEThe male gametophytes of all extant seed plants form a pollentube (Figure 5.9) soon after the pollen grains make contactwith the megasporangial (nucellar) tissue of the ovule. Inextant seed plants the ancestral type of pollen type (found incycads and ginkgophytes) was haustorial, in which the malegametophyte feeds (like a parasite) off the tissues of thenucellus. Motile sperm is delivered from this male gametophyte into a fertilization chamber, where the sperm swims tothe archegonium containing the egg, a process known aszooidogamy (zoom, animal + gamos, marriage). In the conifers (including Gnetales), pollen tubes are also haustorial, butdeliver nonmotile sperm cells to the archegonium or egg, aprocess known as siphonogamy (siphono, tube + gamos,
marriage). A type of siphonogamy evolved independently inthe angiosperms. In angiosperms, however, the pollen tubesgrow through stylar tissue prior to delivering the sperm to theegg of a female gametophyte (see Chapter 6).
OVULE AND SEED DEVELOPMENTAfter pollination, the megasporocyte develops within themegasporangium of the ovule (Figures 5.1OA, 5.11A). Themegasporocyte is a single cell that undergoes meiosis, producinga tetrad of four haploid megaspores, which in most extant seedplants are arranged in a straight line, or linearly (Figure 5.IOA).The three megaspores that are distal (away from the ovule base)abort; only the proximal megaspore (near the ovule base) continues to develop. In the pollination chamber, the resorbedpollen grains (Figures 5. iDA, 5.1 1A) develop into mature malegametophytes and form pollen tubes, which grow into the tissueof the megasporangium (Figures 5. iDA, 5.1 1B). In gymnosperms
these male gametophytes may live in the megasporangial tissue
for some time, generally several months to a year.
The functional megaspore greatly expands, accompanied
by numerous mitotic divisions, to form the endosporic
female gametophyte (Figures 5.1OA, 5.11B,C). In the seeds
of gymnosperms, archegonia differentiate at the apex of the
female gametophyte (Figure 5.11C,D). As in the nonseed
land plants, each archegonium has a large egg cell and a
short line of neck cells (plus typically a ventral canal cell or
nucleus). Eventually, the male gametophytes either release
motile sperm cells (in cycads and Ginkgo) into a cavity
between the megasporangium and female gametophyte
(known as the archegonial chamber; Figure 5.1 DA), or the
pollen tube of the male gametophyte delivers sperm cells
directly into the archegonial neck (in conifers). (Note that
micropyle
the ovules of some Gnetales and all angiosperms lack arche
gonia.) The end result is that a sperm cell from the male
gametophyte fertilizes the egg of the female gametophyte. A
long period of time (perhaps a year or more) may ensue
between pollination, which is delivery of the pollen grains
to the ovu)e, and fertilization, actual union of sperm and
egg. Note: This is not true for the flowering plants, in which
fertilization generally occurs very soon after pollination (see
Chapter 6).The resulting diploid zygote, once formed, undergoes
considerable mitotic divisions and differentiation, eventually
maturing into the embryo, the immature sporophyte (Figures
5.1DB, ShE). The tissue of the female gametophyte contin
ues to surround the embryo (Figure 5.11E) and serves as
nutritive tissue for the embryo upon seed germination (except
136 CHAPTER 5 EVOLUTION AND DIVERSITY OF WOODY AND SEED PLANTS UNIT II EVOLUTION AND DIVERSITY OF PLANTS 137
A
FIGURE 5.8 Pollen grains—immature male gametophytes of seed plants. A. Zamia sp., a cycad. B. Ginkgo biloba. C. Pinus sp., a conifer.
SICL
pollen pollengrains
pollination droplet
- integumentpollinationchamber
integument (2n)
megasporangium(nucellus)
(2n)
mitosiS and
archegonial chamber
archegonium(with egg)
differentiation
A
functionalmegaspore
(n)
(2n)
micropyle
femalegametophyte
(n)
egg)
seed coat’
mitosis and
micropyle
megaSporangiUm(nucellus)
(2n)
B
femalegametophyte
(n)
embryo(new 2n)
megasporangium
pollen tube(haustorial)
germination &
radicle
hypocotyl
epicotyl(shoot apex)
cotyledons
femalegametophyte
(n)
megaspOrangiUm(degenerate)
differentiation
FIGURE 5.10 A. Ovule development in the nonflowering spermatophytes. B. Seed development.
pollen grain(immature endosporic
male gametophyte)
mature malegametophytes, each
with pollen tube
sperm
motilesperm cell
FIGURE 5.9 Male gametophyte morphology and development in the nonflowering spermatophytes; Cycas sp., illustrated. (Reproducedand modified from Swamy, B. G. L. 1948. American Journal of Botany 35: 77—88, by permission.)
1

as a haustorial organ, obtaining nutrition by absorption from thesurrounding sporophytic tissue (Figure 5.9; see Pollen Tabe).
POLLEN TUBEThe male gametophytes of all extant seed plants form a pollentube (Figure 5.9) soon after the pollen grains make contactwith the megasporangial (nucellar) tissue of the ovule. Inextant seed plants the ancestral type of pollen type (found incycads and ginkgophytes) was haustorial, in which the malegametophyte feeds (like a parasite) off the tissues of thenucellus. Motile sperm is delivered from this male gametophyte into a fertilization chamber, where the sperm swims tothe archegonium containing the egg, a process known aszooidogamy (zoom, animal + gamos, marriage). In the conifers (including Gnetales), pollen tubes are also haustorial, butdeliver nonmotile sperm cells to the archegonium or egg, aprocess known as siphonogamy (siphono, tube + gamos,
marriage). A type of siphonogamy evolved independently inthe angiosperms. In angiosperms, however, the pollen tubesgrow through stylar tissue prior to delivering the sperm to theegg of a female gametophyte (see Chapter 6).
OVULE AND SEED DEVELOPMENTAfter pollination, the megasporocyte develops within themegasporangium of the ovule (Figures 5.1OA, 5.11A). Themegasporocyte is a single cell that undergoes meiosis, producinga tetrad of four haploid megaspores, which in most extant seedplants are arranged in a straight line, or linearly (Figure 5.IOA).The three megaspores that are distal (away from the ovule base)abort; only the proximal megaspore (near the ovule base) continues to develop. In the pollination chamber, the resorbedpollen grains (Figures 5. iDA, 5.1 1A) develop into mature malegametophytes and form pollen tubes, which grow into the tissueof the megasporangium (Figures 5. iDA, 5.1 1B). In gymnosperms
these male gametophytes may live in the megasporangial tissue
for some time, generally several months to a year.
The functional megaspore greatly expands, accompanied
by numerous mitotic divisions, to form the endosporic
female gametophyte (Figures 5.1OA, 5.11B,C). In the seeds
of gymnosperms, archegonia differentiate at the apex of the
female gametophyte (Figure 5.11C,D). As in the nonseed
land plants, each archegonium has a large egg cell and a
short line of neck cells (plus typically a ventral canal cell or
nucleus). Eventually, the male gametophytes either release
motile sperm cells (in cycads and Ginkgo) into a cavity
between the megasporangium and female gametophyte
(known as the archegonial chamber; Figure 5.1 DA), or the
pollen tube of the male gametophyte delivers sperm cells
directly into the archegonial neck (in conifers). (Note that
micropyle
the ovules of some Gnetales and all angiosperms lack arche
gonia.) The end result is that a sperm cell from the male
gametophyte fertilizes the egg of the female gametophyte. A
long period of time (perhaps a year or more) may ensue
between pollination, which is delivery of the pollen grains
to the ovu)e, and fertilization, actual union of sperm and
egg. Note: This is not true for the flowering plants, in which
fertilization generally occurs very soon after pollination (see
Chapter 6).The resulting diploid zygote, once formed, undergoes
considerable mitotic divisions and differentiation, eventually
maturing into the embryo, the immature sporophyte (Figures
5.1DB, ShE). The tissue of the female gametophyte contin
ues to surround the embryo (Figure 5.11E) and serves as
nutritive tissue for the embryo upon seed germination (except
136 CHAPTER 5 EVOLUTION AND DIVERSITY OF WOODY AND SEED PLANTS UNIT II EVOLUTION AND DIVERSITY OF PLANTS 137
A
FIGURE 5.8 Pollen grains—immature male gametophytes of seed plants. A. Zamia sp., a cycad. B. Ginkgo biloba. C. Pinus sp., a conifer.
SICL
pollen pollengrains
pollination droplet
- integumentpollinationchamber
integument (2n)
megasporangium(nucellus)
(2n)
mitosiS and
archegonial chamber
archegonium(with egg)
differentiation
A
functionalmegaspore
(n)
(2n)
micropyle
femalegametophyte
(n)
egg)
seed coat’
mitosis and
micropyle
megaSporangiUm(nucellus)
(2n)
B
femalegametophyte
(n)
embryo(new 2n)
megasporangium
pollen tube(haustorial)
germination &
radicle
hypocotyl
epicotyl(shoot apex)
cotyledons
femalegametophyte
(n)
megaspOrangiUm(degenerate)
differentiation
FIGURE 5.10 A. Ovule development in the nonflowering spermatophytes. B. Seed development.
pollen grain(immature endosporic
male gametophyte)
mature malegametophytes, each
with pollen tube
sperm
motilesperm cell
FIGURE 5.9 Male gametophyte morphology and development in the nonflowering spermatophytes; Cycas sp., illustrated. (Reproducedand modified from Swamy, B. G. L. 1948. American Journal of Botany 35: 77—88, by permission.)
1

138 CHAPTER 5 EVOLUTiON AND DIVERSITY OF WOODYAND SEED PLANTS
1, phloem
1xylern
in the flowering plants; see Chapter 6). The megasporangium
(nucellus) eventually degenerates. The integument matures
into a peripheral seed coat, which may differentiate into
various hard andlor fleshy layers.
SEED ADAPTATIONS
The adaptive significance of the seed is unquestioned. First,
seeds provide protection, mostly by means of the seed coat,
from mechanical damage, desiccation, and often predation.
Second, seeds function as the dispersal unit of sexual repro
duction. In many plants the seed has become specially modi
fied for dispersal. For example, a fleshy outer seed coat layer
may function to aid in animal dispersal. In fact, in some plants
the seeds are eaten by animals, the outer fleshy layer is
digested, and the remainder of the seed (including the embryo
protected by an inner, hard seed coat layer) passes harmlessly
through the gut of the animal, ready to germinate with a built
in supply of fertilizer. In other plants, differentiation of the
seed coat into one or more wings functions in seed dispersal
by wind. Third, the seed coat may function in dormancy
mechanisms that ensure germination of the seed only under
ideal conditions of temperature, sunlight, or moisture. Fourth,
upon germination, the nutritive tissue surrounding the embryo
provides energy for the young seedling, aiding in successful
establishment.Interestingly, in seed plants the female gametophyte (which
develops within the megaspore) remains attached to and
nutritionally dependent upon the sporophyte. This is exactly
the reverse condition as is found in the liverworts, homworts,
and mosses (Chapter 3).
EUSTELEIn addition to the seed, an apomorphy for spermatophytes is
the eustele (Figure 5.12). A eustele is a primary stem vascu
lature (“primary” meaning prior to any secondary growth)
that consists of a single ring of discrete vascular bundles.
Each vascular bundle contains an internal strand of xylem
and an external strand of phloem that are radially oriented,
i.e., positioned along a radius (Figure 5.12).
The protoxylem of the vascular bundles of a eustele is
endarch in position, i.e., toward the center of the stem. This is
distinct from the exarch protoxylem of the lycophytes and the
mesarch protoxylem of most monilophytes (Chapter 4) and of
some fossil relatives that diverged prior to the seed plants.
DIVERSITY OF WOODYAND SEED PLANTS
ARCHEOPTEPJS
A well-known lignophyte that lacked seeds was the fossil plant
Archeopteris (not to be confused with the very famous fossil,
reptilian bird Archeopteryx). Archeopteris was a large tree, with
wood like a conifer but leaves like a fern (Figure 5.13A,B).
Sporangia, producing spores, were born on fertile branch
systems. Some species of Archeopteris were heterosporous.
“PTERIDOSPERMS”—”SEED FERNS”
The “pteridosperms,” or “seed ferns:’ are almost certainly
a paraphyletic group of fossil plants that had femlike foliage, yet
bore seeds. Medullosa is a well-known example of a seed fern
integument
I’,
Iintegument
femalegametophyte
Vgametophyte
.
rchegonia
/
UNIT II EVOLUTION AND DIVERSITY OF PLANTS 139
4female
gametophyte
cortex
a7pith
/--megasporocyte
A
r.
U-.
-
B C
11female
gametophyte
FIGURE 5.12 Eustele. A. Diagram of eustele. Note single ring of vascular bundles, with xylem inside, phloem outside. B. Helianthus stem
cross-section, an example of a eustele. C. Close-up of vascular bundle, showing xylem, phloem, and associated fibers.
vascularbundle
embryo
/
nuêleus
-
. ‘\sIrile••• ‘
‘.:;. ... • cells
-1
EFIGURE 5.11 Ovule and seed development, illustrated by Pinus sp. A. Young ovule, longitudinal-section, at time of pollination. Pollengrains are pulled into micropyle by resorption of pollination droplet. Meiosis of the megasporocyte has yet to occur. B. Post-pollination,showing development of the female gametophyte and haustorial pollen tube growth of the male gametophytes within tissue of megasporangium(nucellus). C. Mature ovule, showing two functional archegonia within female gametophyte. D. Close-up of archegonia, each containinga large egg cell with a surrounding layer of sterile cells and apical neck. E. Seed longitudinal-section, seed coat removed, showing embryoand surrounding nutritive layer of female gametophytic tissue.

138 CHAPTER 5 EVOLUTiON AND DIVERSITY OF WOODYAND SEED PLANTS
1, phloem
1xylern
in the flowering plants; see Chapter 6). The megasporangium
(nucellus) eventually degenerates. The integument matures
into a peripheral seed coat, which may differentiate into
various hard andlor fleshy layers.
SEED ADAPTATIONS
The adaptive significance of the seed is unquestioned. First,
seeds provide protection, mostly by means of the seed coat,
from mechanical damage, desiccation, and often predation.
Second, seeds function as the dispersal unit of sexual repro
duction. In many plants the seed has become specially modi
fied for dispersal. For example, a fleshy outer seed coat layer
may function to aid in animal dispersal. In fact, in some plants
the seeds are eaten by animals, the outer fleshy layer is
digested, and the remainder of the seed (including the embryo
protected by an inner, hard seed coat layer) passes harmlessly
through the gut of the animal, ready to germinate with a built
in supply of fertilizer. In other plants, differentiation of the
seed coat into one or more wings functions in seed dispersal
by wind. Third, the seed coat may function in dormancy
mechanisms that ensure germination of the seed only under
ideal conditions of temperature, sunlight, or moisture. Fourth,
upon germination, the nutritive tissue surrounding the embryo
provides energy for the young seedling, aiding in successful
establishment.Interestingly, in seed plants the female gametophyte (which
develops within the megaspore) remains attached to and
nutritionally dependent upon the sporophyte. This is exactly
the reverse condition as is found in the liverworts, homworts,
and mosses (Chapter 3).
EUSTELEIn addition to the seed, an apomorphy for spermatophytes is
the eustele (Figure 5.12). A eustele is a primary stem vascu
lature (“primary” meaning prior to any secondary growth)
that consists of a single ring of discrete vascular bundles.
Each vascular bundle contains an internal strand of xylem
and an external strand of phloem that are radially oriented,
i.e., positioned along a radius (Figure 5.12).
The protoxylem of the vascular bundles of a eustele is
endarch in position, i.e., toward the center of the stem. This is
distinct from the exarch protoxylem of the lycophytes and the
mesarch protoxylem of most monilophytes (Chapter 4) and of
some fossil relatives that diverged prior to the seed plants.
DIVERSITY OF WOODYAND SEED PLANTS
ARCHEOPTEPJS
A well-known lignophyte that lacked seeds was the fossil plant
Archeopteris (not to be confused with the very famous fossil,
reptilian bird Archeopteryx). Archeopteris was a large tree, with
wood like a conifer but leaves like a fern (Figure 5.13A,B).
Sporangia, producing spores, were born on fertile branch
systems. Some species of Archeopteris were heterosporous.
“PTERIDOSPERMS”—”SEED FERNS”
The “pteridosperms,” or “seed ferns:’ are almost certainly
a paraphyletic group of fossil plants that had femlike foliage, yet
bore seeds. Medullosa is a well-known example of a seed fern
integument
I’,
Iintegument
femalegametophyte
Vgametophyte
.
rchegonia
/
UNIT II EVOLUTION AND DIVERSITY OF PLANTS 139
4female
gametophyte
cortex
a7pith
/--megasporocyte
A
r.
U-.
-
B C
11female
gametophyte
FIGURE 5.12 Eustele. A. Diagram of eustele. Note single ring of vascular bundles, with xylem inside, phloem outside. B. Helianthus stem
cross-section, an example of a eustele. C. Close-up of vascular bundle, showing xylem, phloem, and associated fibers.
vascularbundle
embryo
/
nuêleus
-
. ‘\sIrile••• ‘
‘.:;. ... • cells
-1
EFIGURE 5.11 Ovule and seed development, illustrated by Pinus sp. A. Young ovule, longitudinal-section, at time of pollination. Pollengrains are pulled into micropyle by resorption of pollination droplet. Meiosis of the megasporocyte has yet to occur. B. Post-pollination,showing development of the female gametophyte and haustorial pollen tube growth of the male gametophytes within tissue of megasporangium(nucellus). C. Mature ovule, showing two functional archegonia within female gametophyte. D. Close-up of archegonia, each containinga large egg cell with a surrounding layer of sterile cells and apical neck. E. Seed longitudinal-section, seed coat removed, showing embryoand surrounding nutritive layer of female gametophytic tissue.
Copyright © 2022 FDOKUMEN




















